植物表型多样性是基因表达和所处环境交互作用的结果, 表现为种群在不同分布区的压力选择中发生不可逆变异, 并经稳定遗传产生表型变异, 表型变异在适应与进化上具有重要意义(李斌等, 2002 )。研究表型变异格局及其与地理生态因子的关系, 不仅有助于了解植物应对环境因子变化的可塑性, 同时也可以揭示遗传因素和环境因素在植物生态适应中的相互作用(李因刚等, 2014 )。近年来, 表型变异的研究主要集中在分析变异程度进行种质资源分类上, 前人已通过表型对杏(Armeniaca vulgaris )(刘娟等, 2014 )、秋子梨(Pyrus ussuriensis )(马腾等, 2011 ; 许靖诗等, 2015 )、无患子(Sapindus mukorossi )(刁松锋等, 2014 )、柿(Diospyros kaki )(梁玉琴等, 2015 )、五角枫(Acer pictum subsp. mono )(姬志峰等, 2012 )等大量树种进行了研究, 并解释了各个树种的遗传变异规律。在环境因素方面, 毛梾(Cornus walteri )(康永祥等, 2011 )种实性状与纬度和降水量呈梯度变化规律, 川西云杉(Picea likiangensis var. rubescens )(辜云杰等, 2009 )种实性状存在经度、纬度并存的变异模式。另外, 以往形态特征的研究中, 大多以分析各种源差异程度和选择优良性状的遗传材料为重点, 很少进一步在地理生态上 开展重要性状研究, 探究物种适应性和性状分布 格局。
山杏(Armeniaca sibirica )为蔷薇科李亚科杏属植物(俞德浚, 1986 ), 主要分布在俄罗斯的西伯利亚、蒙古国和我国, 我国主要分布于内蒙古、辽宁、河北等省区, 耐干旱、瘠薄, 适应性强, 作为先锋树种应用于沙漠防护林营造, 杏仁粗脂肪含量45.64%, 粗蛋白含量28%, 苦杏仁苷含量4.5% (包文泉, 2013 ), 是重要的木本油料树种和药用资源。近几年来, 荒漠化的加重以及全球气候的变化以及农业耕地和城市发展冲突的加剧, 使得山杏在防治荒漠化上的生态功能以及作为木本油料植物节约耕地的重大意义越来越受到关注。
目前对山杏的研究多集中在主要经济性状变异(李明等, 2011 ; 包文泉等, 2014 )、成分提取(马玉花等, 2007 )、生长生殖(刘梦培等, 2014 )、抗寒性(张睿等, 2005 )和S基因型鉴定(刘梦培, 2015 )等方面。关于山杏天然生态种群的表型多样性以及与地理生态的适应关系少有研究, 这在一定程度上制约了山杏遗传多样性和良种选育工作的开展。本研究以内蒙古13个天然分布的山杏种群为研究对象, 结合地理生态因子调查分析叶、果、核、仁的23个表型性状, 旨在揭示内蒙古山杏种群间和种群内的变异规律和性状分布格局, 为山杏种质资源收集和良种选育工作提供科学依据。
1 材料和方法
1.1 材料来源和样品采集
在对内蒙古山杏文献资料搜集以及野外实地调查的基础上, 分别于2012年及2013年每年果实成熟的7-8月份完成叶和果实的野外采集工作。采样点分别位于内蒙古的扎赉特旗(P1)、科尔沁右翼前旗察尔森(P2)、科尔沁右翼中旗(P3)、扎鲁特旗(P4)、科尔沁左翼后旗(P5)、阿鲁科尔沁旗(P6)、巴林右旗(P7)、敖汉旗(P8)、克什克腾旗(P9)、卓资县(P10)、和林格尔县(P11)、土默特左旗万家沟(P12)、乌拉山(P13), 共计13个种群130个植株, 取样时株间距大于25 m, 即母树树高的5倍以上(辜云杰等, 2009 )。每一单株从东西南北4个方向随机采摘健康、无病虫害的果实、成熟叶片各10个, 低温保鲜带回实验室备用。采样点的地理位置、海拔和气候因子等见表1 。地理位置和海拔为采样时现场记录, 气候数据来源于中国气象数据网(data.cma.cn)。
1.2 表型性状的测定
参照《杏种质资源描述规范和数据标准》(刘宁和刘威生, 2006 )和《中国果树志: 杏卷》(张加延和张钊, 2003 )选择遗传上相对稳定、易于获得和测定的23个表型性状。用电子天平精确到0.01 g测单果质量、核干质量、仁干质量, 用游标卡尺精确到0.01 mm测叶长、叶宽、叶柄长、叶尖长、果纵径、果横径、果侧径、核纵径、核横径、核侧径、核壳厚、仁纵径、仁横径、仁侧径等性状。根据叶片长宽, 果实的纵径、横径, 核的纵径、横径, 仁的纵径、横径分别计算叶形指数、果形指数、核形指数和仁形指数。根据单果质量、干核质量和干仁质量计算出核率和出仁率。
1.3 数据处理和分析
采用巢式方差分析研究山杏的23个表型性状的变异, 线性模型为: yijk = μ + αi + βj(i) + εijk 。其中, yijk 为第i 个种群的第j 个个体的第k 个观测值, μ 为总体平均值; αi 为第i 个种群的效应值(固定), βj(i) 为第i 个种群内第j 个个体的效应值(随机), εijk 为第ijk 个观测值的试验误差。计算各性状平均值、标准偏差、变异系数(CV )和表型分化系数(V st ), 进行表型性状间的差异性检验和多重比较分析, 其中CV =标准偏差/ 平均值, V st = δ 2 t/s /(δ 2 t/s + δ 2 s ), 其中δ 2 t/s 为种群间方差值, δ 2 s 为种群内方差值(葛颂等, 1988 )。CV 表明了性状变异的离散s程度, V st 描述的是表型变异在种群间贡献的大小。计算表型性状与生态因子间的相关系数, 并对性状进行主成分分析。数据处理在Excel 2010和SPSS 20.0软件中完成。
2 结果
2.1 种群间和种群内的表型变异
表2 为山杏的23个表型性状在种群间和种群内层次上的变异分析。经F 值检验, 23个参试表型性状, 除单果质量和核形指数在种群内无显著差异外, 其他性状在种群间和种群内层次上都存在极显著差异(p < 0.01)。表型性状的平均值、标准偏差和多重比较结果(附录I)表明, 山杏表型性状在种群间存在显著差异。山杏叶长最长、叶尖长最长、仁形指数最大的是P8种群, 叶宽最宽、仁纵径最长、仁侧径最长的是P2种群, 叶柄长最长、核横径最长、仁横径最长的是P9种群, 叶形指数最大、果形指数最大、核壳厚最大、核形指数最大、出核率最大的是P13种群, 果纵径最大、果横径最大、果侧径最大、单果质量最大、核纵径最大、核侧径最大、核形指数最大、核干质量最大的是P11种群, 仁干质量最大、出仁率最大的是P10种群。
2.2 种群间的表型分化
通过巢式方差分析, 得到山杏23个表型性状种群内和种群间的方差分量和各性状的表型分化系数(附录II), 结果表明23个表型性状在种群间和种群内的平均方差分量百分比分别为47.15%和16.43%, 随机误差36.43%。种群间的表型分化系数的变异范围是49.27%-97.47%, 单果质量的表型分化系数最大, 在种群间分化较大, 其次是果横径和叶柄长, 而仁纵径和仁形指数较小, 在种群间分化较小, 相对稳定。山杏各表型性状平均表型分化系数为73.03%, 说明表型变异中种群间的贡献为73.03%, 种群内的贡献为26.97%, 种群间的变异程度大于种群内, 种群间的变异是表型变异的主要来源。
2.3 表型性状变异特征
变异系数反映了性状的离散程度, 变异系数越大, 则性状值离散程度越大, 表型多样性越丰富。山杏种群23个表型性状的变异系数分析(附录III)表明, 山杏叶、果、核、仁各性状的平均变异系数为14.28%, 变异幅度为7.01%-27.23%。其中单果质量的变异系数(27.23%)最大, 其次为核壳厚(25.49%)和核干质量(22.56%), 果形指数的变异系数(7.01%)最小。从不同的部位来看, 核的变异系数(15.67%)最大, 果的变异系数(12.11%)最小。13个种群表型性状的平均变异系数在11.65%-17.37%, P12种群和P6种群的表型多样性较为丰富, P2种群和P1种群的表型多样性程度较低。
2.4 表型性状与地理生态因子的相关性
山杏23个表型性状与采样点地理生态因子间的相关性分析见表3 , 综合来说, 地理生态因子与表型性状的相关性如下: 经度(8.372) >年降水量(7.927) >年日照时间(7.486) >纬度(7.157) >海拔(7.069) >年平均气温(6.700) >无霜期(4.304)。其中经度与果形指数呈极显著负相关关系(p < 0.01), 与叶形指数和核纵径呈显著负相关关系(p < 0.05), 与叶尖长和仁侧径呈显著正相关关系(p < 0.05); 年降水量与叶柄长呈极显著正相关关系(p < 0.01), 与叶长、叶宽、叶尖长呈显著正相关关系(p < 0.05), 与果形指数呈显著负相关关系(p < 0.05); 年日照时间与叶长、叶宽、叶柄长、叶尖长呈显著负相关关系(p < 0.05), 与出核率呈显著正相关关系(p < 0.05); 纬度与果形指数呈极显著负相关关系(p < 0.01), 与叶长呈显著正相关关系(p < 0.05); 海拔与叶形指数和果形指数呈极显著正相关关系(p < 0.01), 与叶宽和仁侧径呈显著负相关关系(p < 0.05); 年平均气温与仁纵径呈显著负相关关系(p < 0.05)。无霜期与各性状的相关关系并不显著。
2.5 表型性状的主成分分析和种群聚类分析
通过对山杏资源的23个表型性状进行主成分分析(表4 ), 得到5个特征值大于1的主成分, 贡献率高达90.484%。第一主成分(PC-1)主要包含叶长、叶宽、叶柄长、叶尖长、果纵径等, 主要表征叶性状; 第二主成分(PC-2)主要包含单果质量、核纵径、核横径、核侧径、核壳厚、核形指数、核干质量等, 表征核性状; 第三主成分(PC-3)主要包含出核率、仁纵径、仁横径、仁侧径、仁形指数等, 表征仁性状; 第四主成分(PC-4)主要包含仁干质量、果侧径、果形指数、核形指数等, 表征仁干质量和形状指标; 第五主成分(PC-5)主要包含出仁率。
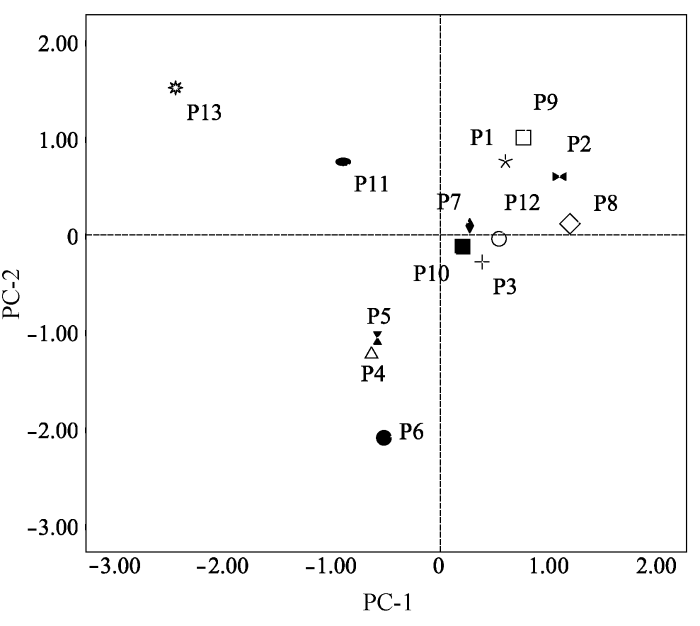
PC-1和PC-2的贡献率分别是38.557%和29.782%, 以PC-1和PC-2绘制散点图, 图1 表明地理分布对无患子表型性状的影响较大, 且对核性状(PC-2)的影响大于叶性状(PC-1)。P3、P7、P10、P12种群的表型性状受地理因子影响较小, 其他种群受地理因子影响较大, 其中P13种群的叶性状和P6种群的核性状受地理因子影响最大。
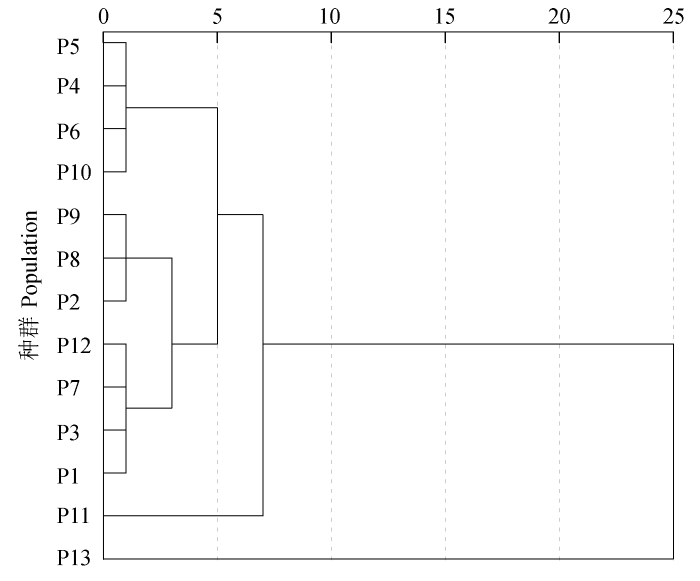
13个种群间用组间连接法进行聚类, 得到种群聚类图(图2 )。种群根据表型性状可以分为5类, 第一类为P4、P5、P6、P10; 第二类为P2、P8、P9; 第三类为P1、P3、P7、P12; 第四类为P11; 第五类为P13。
图1 基于表型性状(PC-1, PC-2)的山杏种群关系。种群代码见表1 。
Fig. 1 The relationship among different Armeniaca sibirica populations based on PC-1 and PC-2. Population code see Table 1 .
图2 基于表型性状的山杏种群聚类分析。种群同表1 。
Fig. 2 Cluster dendrogram of Armeniaca sibirica populations based on the phenotypic traits. Population see Table 1 .
3 讨论
3.1 山杏不同种群表型变异来源
内蒙古山杏主要分布于大兴安岭南麓东坡和阴山山脉中段大青山南坡的灌丛草原和低山丘陵地区。通过对该分布区的13个种群的叶、果、核、仁等23个表型性状的研究发现, 山杏的表型性状在种群间和种群内都存在极显著差异, 这与杏(刘娟等, 2014 )、浙江柿(Diospyros glaucifolia )(井振华等, 2010 )、文冠果(Xanthoceras sorbifolia )(牟洪香等, 2007 )等经济树种的研究结果一致。表型分化系数在某种程度上反映了山杏的环境适应性, 种群表型分化系数的均值为73.03%, 高于山苍子(Litsea cubeba )(66.66%)(田胜平等, 2012 )、无患子(62.21%)(刁松锋等, 2014 )、浙江楠(Phoebe chekiangensis ) (41.43%)(李因刚等, 2014 )、浙江柿(40.15%)(井振华等, 2010 ), 和滇龙胆(Gentiana rigescens ) (73.14%) (杨维泽等, 2011 )相当, 低于夏蜡梅(Sinocalycanthus chinensis )(89.30%)(金则新等, 2012 ), 表明了山杏种群表型分化程度较高, 对环境的适应性较强, 在内蒙古横跨13.71个经度(1523 km)范围, 在224-2083 m海拔范围内自然分布, 造成了不同种群表型性状的异质性。种群间的变异程度大于种群内, 种群间的变异是表型变异的主要来源, 这可能与山杏的地理分布和繁殖系统有关。种群间变异的量值取决于其地理和生态分布区(杨继, 1991 ), 内蒙古的山杏多分布于荒山丘陵和沙地边缘, 东西地理分布的间断性和海拔的差异等地理隔离促进了种群间的分化。另外, 交配系统也是影响植物种群遗传结构的重要因素(Hamrick & Godt, 1990 ), 山杏主要为虫媒异花授粉, 其花粉的传播距离远低于风媒花植物, 且杏花开放主要受温度影响, 不同地理种群花期的差异阻碍了种群间的花粉传播和基因交流, 导致不同种群间的遗传分化加剧。山杏自然生境贫瘠, 结实量少, 且种仁易被鸟兽取食, 加上海拔障碍, 使种子传播受阻导致种群间种子交流困难, 从而促进了种群间的遗传分化。分化较大的种群在各自生境中的适应和进化促成了山杏丰富的遗传多样性。
3.2 山杏种群的表型变异特征
变异系数越大, 性状观测值离散程度越大, 表型多样性程度越高。山杏叶、果、核、仁性状的平均变异系数大小顺序为: 核(15.67%) >叶(14.51%) >仁(14.13%) >果(12.11%), 核与叶的变异系数较大可能与日照及海拔有关, 采样过程中发现P13种群海拔(2083 m)和年日照时间(3205 h)都高于其他种群, 其形态表现出叶子小, 树形小, 干上多刺, 结果少, 果核大, 核壳厚等具有适应意义的自我保护特性, 类似情况见于西南桦(Betula alnoides )(曾杰等, 2005 )在低海拔、风速小地区增大种翅以促进种子散布。另外P11种群的果实大于其他种群, 可能与其在果实形态上与普通杏接近有关。山杏叶、核、果、仁的表征质量和大小的性状变异系数均值为15.25%, 而叶形指数、果形指数、核形指数、仁形指数等表征形状的性状变异系数均值仅为9.45%, 表明山杏质量和大小的变异程度较大, 而形状较为稳定, 这与对毛梾(康永祥等, 2011 )和无患子(刁松锋等, 2014 )表型性状的研究结果一致。单果质量、核干质量、仁干质量等重要经济性状的遗传变异较大, 受外界环境选择的压力较大, 在选择上更有潜力, 这为山杏的仁用良种选育提供了丰富的物质 基础。
3.3 山杏种群的表型性状与地理生态因子的相 关性
山杏13个种群的分布区均属于中温带季风气候, 具有降水量少且分布不均匀、寒暑变化剧烈的特点, 纬度范围40.38°-46.59° N, 经度范围109.04°- 122.75° E, 海拔范围245-2083 m, 广泛的经度范围和海拔差异使得表型性状存在较大的异质性。除无霜期外, 其他地理生态因子对山杏的表型性状都有显著影响。7个生态因子中, 经度对山杏表型性状的综合影响最大, 东部种群的叶形更圆, 叶尖趋于长尾尖, 果形更接近圆球形, 仁也更趋于饱满; 西部种群叶形卵圆, 叶尖趋于钝尖, 果形趋于长圆形。随着年降水量的增加, 叶子变大, 叶柄变长, 果更趋于圆球形。随着年日照时间的增加, 叶子变小, 核变大, 核壳变厚, 核形趋于长圆形, 仁变小。这可能是强光照下植物的适应机制, 叶子变小以减小蒸腾作用失水, 核壳变厚以保护种子在不利条件下的生存繁殖。而随着纬度的增加, 叶长变长, 果形指数减小。随着海拔的增加, 叶子变小, 核变大, 核壳变厚, 仁侧径变小, 果形趋于长圆, 其对性状的影响与年日照时间类似。年平均气温和无霜期对表型性状无显著影响。因此, 由于复杂的地理环境, 不同于青海云杉(Picea crassifolia )(王娅丽等, 2008 )单一的经度变化规律, 也不同于无患子(刁松锋等, 2014 )以纬度和年平均气温为主的变化规律和五角枫(张翠琴等, 2015 )以经度和纬度并存的变异趋势, 山杏不同种群表型性状的变异与地理经度、年降水量以及年日照时间3个主要地理气候因子相关。
根据主成分分析以及成分得分散点图, P4、P5、P6种群可归为一类, 其地理分布距离较近, 海拔和年日照时间基本相同, 水热条件较为接近; 而P1、P2、P8、P9种群归为一类, 其年降水量彼此接近且海拔和年平均气温相对较低; P3、P7、P10、P12种群归为一类, 其年日照时间较长, 接近3000 h; P11和P13种群本应归为一类, 但由于乌拉山的海拔(2083 m)比和林格尔县(1160 m)高将近一倍, 其较长的日照时间和较少的降水量, 使其种群表型性状产生较为独特的变异, 形成叶小, 干上生刺, 结果量少, 核壳很厚等一系列自我保护机制(李帅锋等, 2013 )。而聚类分析的结果与主成分分析结果基本一致, 不同种群间表型的差异与地理气候因子密切相关, 由此可见, 利用地理区划对山杏种群进行表型上的划分是可行的。而要进一步解读表型变异和生态因子的关系, 应该利用系统的生态学方法, 扩大样地及样本量, 以得到更加精确可信的结果。
3.4 山杏种质资源的保育和利用
山杏生境贫瘠, 适应性极强, 杏仁脂肪酸、粗蛋白和苦杏仁苷含量丰富, 具有重要的生态价值、经济价值和药用价值, 其自然分布环境复杂多样, 地理隔离严重, 在种群内和种群间层次上都存在丰富的变异, 这为良种选育提供了物质基础。近年来, 荒漠化的加重以及全球气候的变化使山杏种质资源受到严重侵蚀, 遗传多样性逐渐丧失(曹扬, 2014 )。为了充分利用现有天然资源, 选育优良资源进行山杏的遗传改良, 培育优质高产稳定的品种, 特提出如下建议: (1)协调林牧关系, 加强对野生资源的保护, 充分发挥山杏在贫瘠山地的生态价值和经济创收价值; (2)山杏表型变异主要来源于种群间, 要重视不同地理种群间目标性状的分布规律, 增加种群间的采样数, 选择特定种群后更有效地进行定向改良(如仁用、壳用、抗寒等); (3)山杏表型变异与地理经度、年降水量以及年日照时间相关, 加大这三个因子的采样范围, 以得到更多变异特征; (4)山杏的经济价值需要充分挖掘, 在丰富的表型变异基础上营建母树林或种子园, 进行良种化和产业化发展, 发挥其重要木本油料和高品质蛋白作物的作用; (5)同时需要将分子手段和表型变异以及地理生态相结合, 进行遗传多样性研究, 构建核心种质, 提取有保护意义和利用价值的资源。
附录I 山杏13个种群表型性状的平均值、标准偏差及多重比较
Supplemen I Mean value, standard deviation and multiple comparison of phenotypic traits of 13 Armeniaca sibirica populations
http://www.plant-ecology.com/fileup/PDF/cjpe.2016.0172-S1.xls
附录II 山杏表型性状的方差分量及种群间表型分化系数
Supplemen II Variance components and phenotypic differentiation coefficients among Armeniaca sibirica populations
http://www.plant-ecology.com/fileup/PDF/cjpe.2016.0172-S2.xls
附录III 山杏表型性状的变异系数
Supplemen III Variance components of phenotypic traits in Armeniaca sibirica populations
http://www.plant-ecology.com/fileup/PDF/cjpe.2016.0172-S3.xls
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Bao WQ 2013 ). Evaluation of Important Genetic Variation on Armeniaca sibirica in Inner Mongolia and Its Excellent Plant Screening. Master degree dissertation,
Inner Mongolia Agricultural University , Hohhot . 32 -40 .(in Chinese) [包文泉 (2013 ). 内蒙古地区山杏重要性状遗传变异评价及优株选择
. 硕士学位论文, 内蒙古农业大学, 呼和浩特 . 32 -40 .]
[本文引用: 1]
[2]
Bao WQ Wuyun TN Yin MY 2014 ). Genetic variation of fine individual plants’ seed characteristics of wild apricot in Inner Mongolia.
Nonwood Forest Research , 32 (4 ), 13 -17, 72 .(in Chinese with English abstract) [包文泉 , 乌云塔娜 , 尹明宇 (2014 ). 内蒙古野生山杏优良单株果核性状的遗传变异分析.
经济林研究 ,32 (4 ), 13 -17, 72 .]
[本文引用: 1]
[3]
Cao Y 2014 ). Analysis preservation of Armeniaca sibirica germplasm resources in Inner Mongolia.
Inner Mongolia Forestry Investigation and Design , 37 (3 ), 103 -105 .(in Chinese with English abstract) [曹扬 (2014 ). 内蒙古自治区山杏种质资源保存库建设分析.
内蒙古林业调查设计 ,37 (3 ), 103 -105 .
[本文引用: 1]
[4]
Diao SF Shao WH Jiang JM Dong RX Sun HG 2014 ). Phenotypic diversity in natural populations of Sapindus mukorossi based on fruit and seed traits.
Acta Ecologica Sinica , 34 , 1451 -1460 .(in Chinese with English abstract) [刁松锋 , 邵文豪 , 姜景民 , 董汝湘 , 孙洪刚 (2014 ). 基于种实性状的无患子天然群体表型多样性研究.
生态学报 ,34 , 1451 -1460 .]
[本文引用: 4]
[5]
Ge S Wang MX Chen YW 1988 ). An analysis of population genetic structure of masson pine by isozyme technique.
Scientia Silvae Sinicae , 24 , 399 -410 .(in Chinese with English abstract) [葛颂 , 王明庥 , 陈岳武 (1988 ). 用同工酶研究马尾松种群的遗传结构.
林业科学 ,24 , 399 -410 .]
[本文引用: 1]
[6]
Gu YJ Luo JX Wu YW Cao XJ 2009 ). Phenotypic diversity in natural populations of Picea balfouriana in Sichuan, China.
Chinese Journal of Plant Ecology , 33 , 291 -301 .(in Chinese with English abstract) [辜云杰 , 罗建勋 , 吴远伟 , 曾小军 (2009 ). 川西云杉天然种群表型多样性.
植物生态学报 ,33 , 291 -301 .]
[本文引用: 2]
[7]
Hamrick JL Godt NJW 1990 ). Allozyme Diversity in Plant Species. Sinauer Association,
Sunderland , USA . 43 -63 .
[本文引用: 1]
[8]
Jing ZH Li H Shao WH Yue HF Jiang JM 2010 ). Phenotypic diversity of natural populations in Diospyros glaucifolia.
Bulletin of Botanical Research , 30 , 325 -331 .(in Chinese with English abstract) [井振华 , 李皓 , 邵文豪 , 岳华峰 , 姜景民 (2010 ). 浙江柿天然群体表型多样性研究.
植物研究 ,30 , 325 -331 .]
[本文引用: 2]
[9]
Jin ZX Gu JJ Li JM 2012). Genetic variation among populations of the endangered Sinocalycanthus chinensis based on morphological traits and ISSR profiles.
Acta Ecologica Sinica , 2012 , 32 , 3849 -3858 .(in Chinese with English abstract) [金则新 , 顾婧婧 , 李钧敏 (2012 ). 基于形态及分子标记的濒危植物夏蜡梅自然居群的遗传变异研究.
生态学报 ,32 , 3849 -3858 .]
[本文引用: 1]
[10]
Ji ZF Gao YH Li L Mao SX Zhao L Geng QY Wang YL 2012 ). Phenotypic diversity of populations of Acer mono in Huoshan mountain of Shanxi at different altitude.
Acta Horticulturae Sinica , 39 , 2217 -2228 .(in Chinese with English abstract) [姬志峰 , 高亚卉 , 李乐 , 毛思雪 , 赵亮 , 耿全英 , 王祎玲 (2012 ). 山西霍山五角枫不同海拔种群的表型多样性研究.
园艺学报 ,39 , 2217 -2228 .]
[本文引用: 1]
[11]
Kang YX Zhao BX Yun YJ Chen M 2011 ). Study on phenotypic diversity of seeds and fruits’ characteristics in Cornus walteri. Journal of Northwest A & F University (
Nature Science Edition ), 39 (9 ), 107 -117 .(in Chinese with English abstract) [康永祥 , 赵宝鑫 , 贠玉洁 , 陈绵 (2011 ). 毛梾天然群体种实表型多样性研究.
西北农林科技大学学报(自然科学版) ,39 (9 ), 107 -117 .]
[本文引用: 2]
[12]
Li B Gu WC Lu BM 2002 ). A study on phenotypic diversity of seeds and cones characteristics in Pinus bungeana.
Biodiversity Science , 10 , 181 -188 .(in Chinese with English abstract) [李斌 , 顾万春 , 卢宝明 (2002 ). 白皮松天然种群种实性状表型多样性研究.
生物多样性 ,10 , 181 -188 .]
[本文引用: 1]
[13]
Li M Zhao Z Yang JA Lu B 2011 ). Genetic diversity analysis on germplasm of Armeniaca sibirica in different counties in Loess Plateau. Journal of Northwest A & F University (
Nature Science Edition ), 39 (2 ), 143 -149, 156 (in Chinese with English abstract). [李明 , 赵忠 , 杨吉安 , 卢斌 (2011 ). 黄土高原不同县域山杏种质遗传多样性研究.
西北农林科技大学学报(自然科学版) ,39 (2 ), 143 -149, 156 .]
[本文引用: 1]
[14]
Li SF Su JR Liu WD Lang XD Zhang ZJ Su L Jia CXZ Yang HJ 2013 ). Phenotypic variations in cones and seeds of natural Pinus kesiya var. langbianensis populations in Yunnan Province, China.
Chinese Journal of Plant Ecology , 37 , 998 -1009 .(in Chinese with English abstract) [李帅锋 , 苏建荣 , 刘万德 , 郎学东 , 张志钧 , 苏磊 , 贾呈鑫卓 , 杨华景 (2013 ). 思茅松天然群体种实表型变异.
植物生态学报 ,37 , 998 -1009 .]
[本文引用: 1]
[15]
Li YG Liu XH Ma JW Shi CG Zhu GQ 2014 ). Phenotypic variations in populations of Phoebe chekiangensis.
Chinese Journal of Plant Ecology , 38 , 1315 -1324 .(in Chinese with English abstract) [李因刚 , 柳新红 , 马俊伟 , 石从广 , 朱光权 (2014 ). 浙江楠种群表型变异.
植物生态学报 ,38 , 1315 -1324 .]
[本文引用: 2]
[16]
Liang YQ Han WJ Zhang JJ Sun P Liang JJ Fu JM 2015 ). Phenotypic diversity of persimmon germplasms in Henan Province.
Journal of China Agricultural University , 20 (1 ), 74 -85 .(in Chinese with English abstract) [梁玉琴 , 韩卫娟 , 张嘉嘉 , 孙鹏 , 梁晋军 , 傅建敏 (2015 ). 河南省柿种质资源表型多样性研究.
中国农业大学学报 ,20 (1 ), 74 -85 .]
[本文引用: 1]
[17]
Liu J Liao K Mansur N Zhao SR Liu H Jia Y 2014 ). Research on phenotypic diversity of apricot germplasm resources in Xinjiang.
Journal of Fruit Science , 31 , 1047 -1056 .(in Chinese with English abstract) [刘娟 , 廖康 , 曼苏尔·那斯尔 , 赵世荣 , 刘欢 , 贾杨 (2014 ). 新疆杏种质资源表型多样性研究.
果树学报 ,31 , 1047 -1056 .]
[本文引用: 2]
[18]
Liu MP Du HY Fu JM Zhu GP Zhao H Liu HM Wuyun TN 2014 ). Reproductive ecology of Armeniaca sibirica with cold resistant in in Inner Mongolia.
Acta Botanica Boreali-Occidentalia Sinica , 34 , 1143 -1151 .(in Chinese with English abstract) [刘梦培 , 杜红岩 , 傅建敏 , 朱高浦 , 赵罕 , 刘慧敏 , 乌云塔娜 (2014 ). 内蒙古居群抗寒山杏繁殖生态学研究.
西北植物学报 ,34 , 1143 -1151 .]
[本文引用: 1]
[19]
Liu MP 2015 ). Study on Pollination Pharacteristic and S Genotype Identification of Self-incompatibility of Kernel- apricot. PhD dissertation,
Chinese Academy of Forestry , Beijing . 56 -86 .(in Chinese) [刘梦培 (2015 ). 仁用杏自交不亲和的授粉特性和S基因型鉴定研究
. 博士学位论文, 中国林业科学研究院, 北京 . 56 -58 .]
[本文引用: 1]
[20]
Liu N Liu WS Descriptors and Data Standard for Apricot (Armeniaca Mill.) . China Agriculture Press , Beijing .(in Chinese) [刘宁 , 刘威生 (2006). 杏种质资源描述规范和数据标准 . 中国农业出版社 , 北京.]
[本文引用: 1]
[21]
Ma T Wuyun TN Deng WT Kang X Luo J 2011 ). Investigation on wild Pyrus ussuriensis Maxim resources of Chifeng area in Inner Mongolia.
Guangdong Agricultural Sciences , 38 (2 ), 45 -47 .(in Chinese with English abstract) [马腾 , 乌云塔娜 , 邓文韬 , 康秀 , 罗健 (2011 ). 内蒙古赤峰地区野生秋子梨调查研究.
广东农业科学 ,38 (2 ), 45 -47 .]
[本文引用: 1]
[22]
Ma YH Zhao Z Li KY Ma XH Guo CJ Shi QH Zhu HL 2007 ). Oil content and composition of almond from different producing area.
Journal of the Chinese Cereals and Oils Association , 23 (4 ), 272 -275 .(in Chinese with English abstract) [马玉花 , 赵忠 , 李科友 , 马希汉 , 郭婵娟 , 史清华 , 朱海兰 (2007 ). 超临界CO2 流体萃取杏仁油工艺研究.
农业工程学报 ,23 (4 ), 272 -275 .]
[本文引用: 1]
[23]
Mou HX Hou XC Liu QZ 2007 ). Study on the phenotype diversity of woody energy plant Xanthoceras sorbifolia.
Forest Research , 20 , 350 -355 .(in Chinese with English abstract) [牟洪香 , 侯新村 , 刘巧哲 (2007 ). 木本能源植物文冠果的表型多样性研究.
林业科学研究 ,20 , 350 -355 .]
[本文引用: 1]
[24]
Tian SP Wang YD Chen YC Han XJ 2012 ). Phenotypic diversity of natural Litsea cubeba populations leaf and fruit traits.
Chinese Journal of Ecology , 31 , 1665 -1672 .(in Chinese with English abstract) [田胜平 , 汪阳东 , 陈益存 , 韩小娇 (2012 ). 山苍子天然种群叶片和种实性状的表型多样性.
生态学杂志 ,31 , 1665 -1672 .]
[本文引用: 1]
[25]
Wang YL Li Y 2008 ). Study on phenotypic diversity of cone and seed in natural populations of Picea crassifolia in Qilian Mountain, China. Journal of Plant Ecology (
Chinese Version ), 32 , 355 -362 .(in Chinese with English abstract) [王娅丽 , 李毅 (2008 ). 祁连山青海云杉天然群体的种实性状表型多样性.
植物生态学报 ,32 , 355 -362 .]
[本文引用: 1]
[26]
Xu JS Wuyun TN Wang GJ 2015 ). Study on morphological variation of Chinese wild Pyrus ussuriensis Maxim.
Journal of Central South University of Forestry & Technology , 35 (8 ), 64 -68 .(in Chinese with English abstract) [许靖诗 , 乌云塔娜 , 王广军 (2015 ). 中国野生秋子梨果实形态变异研究.
中南林业科技大学学报 ,35 (8 ), 64 -68 .]
[本文引用: 1]
[27]
Yang J 1991 ). Infraspecific variation in plant and the exploring methods,
Journal of Wuhan Botanical Research , 9 , 185 -195 .(in Chinese with English abstract) [杨继 (1991 ). 植物种内形态变异的机制及其研究方法.
武汉植物学研究 , 9 , 185 -195 .]
[本文引用: 1]
[28]
Yang WZ Jin H Yang MQ Zhao ZL Zhang ZH Wang YZ Zhang JY 2011 ). Phenotypic diversity and environment relations in Gentiana rigescens of Yunnan. Aata Botanica Boreali-
Occidentalia Sinica , 32 , 1326 -1334 .(in Chinese with English abstract) [杨维泽 , 金航 , 杨美权 , 赵振玲 , 张智慧 , 王元忠 , 张金渝 (2011 ). 云南滇龙胆居群表型多样性及其与环境关系研究.
西北植物学报 ,32 , 1326 -1334 .]
[本文引用: 1]
[29]
Yu DX Flora of China . Vol. 38. Science Press , Beijing . 24 -31 .(in Chinese) [俞德浚 (1986). 中国植物志 . 第三十八卷. 科学出版社 , 北京 . 24 -31 .]
[本文引用: 1]
[30]
Zeng J Zheng HS Gan SM Bai JY 2005 ). Phenotypic variation in natural populations of Betula alnoides in Guangxi, China.
Scientia Silvae Sinicae , 41 (2 ), 59 -65 .(in Chinese with English abstract) [曾杰 , 郑海水 , 甘四明 , 白嘉雨 (2005 ). 广西西南桦天然居群的表型变异.
林业科学 ,41 (2 ), 59 -65 .]
[本文引用: 1]
[31]
Zhang CQ Ji ZF Lin LL Zhao RH Wang YL 2015 ). Phenotypic diversity of Acer mono Maxim population.
Acta Ecologica Sinica , 35 , 5343 -5352 .(in Chinese with English abstract) [张翠琴 , 姬志峰 , 林丽丽 , 赵瑞华 , 王祎玲 (2015 ). 五角枫种群表型多样性.
生态学报 ,35 , 5343 -5352 .]
[本文引用: 1]
[32]
Zhang JY Zhang Z Chinese Fruit Tree, Apricot . China Forestry Publishing House ,Beijing . 93 -559 .(in Chinese) [张加延 , 张钊 (2003). 中国果树志 , 杏卷. 中国林业出版社 , 北京 . 93 -559 .]
[本文引用: 1]
[33]
Zhang R Wei AZ Yang H Yang TX Sa WQ Yang XN 2005 ). Cold resistances of the young fruits of well- performing apricot varieties with sweet almonds.
Acta Botanica Boreali-Occidentalia Sinica , 12 , 2510 -2513 .(in Chinese with English abstract) [张睿 , 魏安智 , 杨恒 , 杨途熙 , 撒文清 , 杨向纳 (2005 ). 甜山杏优株幼果抗寒性研究.
西北植物学报 ,12 , 2510 -2513 .]
[本文引用: 1]
内蒙古地区山杏重要性状遗传变异评价及优株选择
1
2013
... 山杏(Armeniaca sibirica )为蔷薇科李亚科杏属植物(俞德浚, 1986 ), 主要分布在俄罗斯的西伯利亚、蒙古国和我国, 我国主要分布于内蒙古、辽宁、河北等省区, 耐干旱、瘠薄, 适应性强, 作为先锋树种应用于沙漠防护林营造, 杏仁粗脂肪含量45.64%, 粗蛋白含量28%, 苦杏仁苷含量4.5% (包文泉, 2013 ), 是重要的木本油料树种和药用资源.近几年来, 荒漠化的加重以及全球气候的变化以及农业耕地和城市发展冲突的加剧, 使得山杏在防治荒漠化上的生态功能以及作为木本油料植物节约耕地的重大意义越来越受到关注. ...
内蒙古野生山杏优良单株果核性状的遗传变异分析.
1
2014
... 目前对山杏的研究多集中在主要经济性状变异(李明等, 2011 ; 包文泉等, 2014 )、成分提取(马玉花等, 2007 )、生长生殖(刘梦培等, 2014 )、抗寒性(张睿等, 2005 )和S基因型鉴定(刘梦培, 2015 )等方面.关于山杏天然生态种群的表型多样性以及与地理生态的适应关系少有研究, 这在一定程度上制约了山杏遗传多样性和良种选育工作的开展.本研究以内蒙古13个天然分布的山杏种群为研究对象, 结合地理生态因子调查分析叶、果、核、仁的23个表型性状, 旨在揭示内蒙古山杏种群间和种群内的变异规律和性状分布格局, 为山杏种质资源收集和良种选育工作提供科学依据. ...
内蒙古自治区山杏种质资源保存库建设分析.
1
2014
... 山杏生境贫瘠, 适应性极强, 杏仁脂肪酸、粗蛋白和苦杏仁苷含量丰富, 具有重要的生态价值、经济价值和药用价值, 其自然分布环境复杂多样, 地理隔离严重, 在种群内和种群间层次上都存在丰富的变异, 这为良种选育提供了物质基础.近年来, 荒漠化的加重以及全球气候的变化使山杏种质资源受到严重侵蚀, 遗传多样性逐渐丧失(曹扬, 2014 ).为了充分利用现有天然资源, 选育优良资源进行山杏的遗传改良, 培育优质高产稳定的品种, 特提出如下建议: (1)协调林牧关系, 加强对野生资源的保护, 充分发挥山杏在贫瘠山地的生态价值和经济创收价值; (2)山杏表型变异主要来源于种群间, 要重视不同地理种群间目标性状的分布规律, 增加种群间的采样数, 选择特定种群后更有效地进行定向改良(如仁用、壳用、抗寒等); (3)山杏表型变异与地理经度、年降水量以及年日照时间相关, 加大这三个因子的采样范围, 以得到更多变异特征; (4)山杏的经济价值需要充分挖掘, 在丰富的表型变异基础上营建母树林或种子园, 进行良种化和产业化发展, 发挥其重要木本油料和高品质蛋白作物的作用; (5)同时需要将分子手段和表型变异以及地理生态相结合, 进行遗传多样性研究, 构建核心种质, 提取有保护意义和利用价值的资源. ...
基于种实性状的无患子天然群体表型多样性研究.
4
2014
... 植物表型多样性是基因表达和所处环境交互作用的结果, 表现为种群在不同分布区的压力选择中发生不可逆变异, 并经稳定遗传产生表型变异, 表型变异在适应与进化上具有重要意义(李斌等, 2002 ).研究表型变异格局及其与地理生态因子的关系, 不仅有助于了解植物应对环境因子变化的可塑性, 同时也可以揭示遗传因素和环境因素在植物生态适应中的相互作用(李因刚等, 2014 ).近年来, 表型变异的研究主要集中在分析变异程度进行种质资源分类上, 前人已通过表型对杏(Armeniaca vulgaris )(刘娟等, 2014 )、秋子梨(Pyrus ussuriensis )(马腾等, 2011 ; 许靖诗等, 2015 )、无患子(Sapindus mukorossi )(刁松锋等, 2014 )、柿(Diospyros kaki )(梁玉琴等, 2015 )、五角枫(Acer pictum subsp. mono )(姬志峰等, 2012 )等大量树种进行了研究, 并解释了各个树种的遗传变异规律.在环境因素方面, 毛梾(Cornus walteri )(康永祥等, 2011 )种实性状与纬度和降水量呈梯度变化规律, 川西云杉(Picea likiangensis var. rubescens )(辜云杰等, 2009 )种实性状存在经度、纬度并存的变异模式.另外, 以往形态特征的研究中, 大多以分析各种源差异程度和选择优良性状的遗传材料为重点, 很少进一步在地理生态上 开展重要性状研究, 探究物种适应性和性状分布 格局. ...
... 内蒙古山杏主要分布于大兴安岭南麓东坡和阴山山脉中段大青山南坡的灌丛草原和低山丘陵地区.通过对该分布区的13个种群的叶、果、核、仁等23个表型性状的研究发现, 山杏的表型性状在种群间和种群内都存在极显著差异, 这与杏(刘娟等, 2014 )、浙江柿(Diospyros glaucifolia )(井振华等, 2010 )、文冠果(Xanthoceras sorbifolia )(牟洪香等, 2007 )等经济树种的研究结果一致.表型分化系数在某种程度上反映了山杏的环境适应性, 种群表型分化系数的均值为73.03%, 高于山苍子(Litsea cubeba )(66.66%)(田胜平等, 2012 )、无患子(62.21%)(刁松锋等, 2014 )、浙江楠(Phoebe chekiangensis ) (41.43%)(李因刚等, 2014 )、浙江柿(40.15%)(井振华等, 2010 ), 和滇龙胆(Gentiana rigescens ) (73.14%) (杨维泽等, 2011 )相当, 低于夏蜡梅(Sinocalycanthus chinensis )(89.30%)(金则新等, 2012 ), 表明了山杏种群表型分化程度较高, 对环境的适应性较强, 在内蒙古横跨13.71个经度(1523 km)范围, 在224-2083 m海拔范围内自然分布, 造成了不同种群表型性状的异质性.种群间的变异程度大于种群内, 种群间的变异是表型变异的主要来源, 这可能与山杏的地理分布和繁殖系统有关.种群间变异的量值取决于其地理和生态分布区(杨继, 1991 ), 内蒙古的山杏多分布于荒山丘陵和沙地边缘, 东西地理分布的间断性和海拔的差异等地理隔离促进了种群间的分化.另外, 交配系统也是影响植物种群遗传结构的重要因素(Hamrick & Godt, 1990 ), 山杏主要为虫媒异花授粉, 其花粉的传播距离远低于风媒花植物, 且杏花开放主要受温度影响, 不同地理种群花期的差异阻碍了种群间的花粉传播和基因交流, 导致不同种群间的遗传分化加剧.山杏自然生境贫瘠, 结实量少, 且种仁易被鸟兽取食, 加上海拔障碍, 使种子传播受阻导致种群间种子交流困难, 从而促进了种群间的遗传分化.分化较大的种群在各自生境中的适应和进化促成了山杏丰富的遗传多样性. ...
... 变异系数越大, 性状观测值离散程度越大, 表型多样性程度越高.山杏叶、果、核、仁性状的平均变异系数大小顺序为: 核(15.67%) >叶(14.51%) >仁(14.13%) >果(12.11%), 核与叶的变异系数较大可能与日照及海拔有关, 采样过程中发现P13种群海拔(2083 m)和年日照时间(3205 h)都高于其他种群, 其形态表现出叶子小, 树形小, 干上多刺, 结果少, 果核大, 核壳厚等具有适应意义的自我保护特性, 类似情况见于西南桦(Betula alnoides )(曾杰等, 2005 )在低海拔、风速小地区增大种翅以促进种子散布.另外P11种群的果实大于其他种群, 可能与其在果实形态上与普通杏接近有关.山杏叶、核、果、仁的表征质量和大小的性状变异系数均值为15.25%, 而叶形指数、果形指数、核形指数、仁形指数等表征形状的性状变异系数均值仅为9.45%, 表明山杏质量和大小的变异程度较大, 而形状较为稳定, 这与对毛梾(康永祥等, 2011 )和无患子(刁松锋等, 2014 )表型性状的研究结果一致.单果质量、核干质量、仁干质量等重要经济性状的遗传变异较大, 受外界环境选择的压力较大, 在选择上更有潜力, 这为山杏的仁用良种选育提供了丰富的物质 基础. ...
... 山杏13个种群的分布区均属于中温带季风气候, 具有降水量少且分布不均匀、寒暑变化剧烈的特点, 纬度范围40.38°-46.59° N, 经度范围109.04°- 122.75° E, 海拔范围245-2083 m, 广泛的经度范围和海拔差异使得表型性状存在较大的异质性.除无霜期外, 其他地理生态因子对山杏的表型性状都有显著影响.7个生态因子中, 经度对山杏表型性状的综合影响最大, 东部种群的叶形更圆, 叶尖趋于长尾尖, 果形更接近圆球形, 仁也更趋于饱满; 西部种群叶形卵圆, 叶尖趋于钝尖, 果形趋于长圆形.随着年降水量的增加, 叶子变大, 叶柄变长, 果更趋于圆球形.随着年日照时间的增加, 叶子变小, 核变大, 核壳变厚, 核形趋于长圆形, 仁变小.这可能是强光照下植物的适应机制, 叶子变小以减小蒸腾作用失水, 核壳变厚以保护种子在不利条件下的生存繁殖.而随着纬度的增加, 叶长变长, 果形指数减小.随着海拔的增加, 叶子变小, 核变大, 核壳变厚, 仁侧径变小, 果形趋于长圆, 其对性状的影响与年日照时间类似.年平均气温和无霜期对表型性状无显著影响.因此, 由于复杂的地理环境, 不同于青海云杉(Picea crassifolia )(王娅丽等, 2008 )单一的经度变化规律, 也不同于无患子(刁松锋等, 2014 )以纬度和年平均气温为主的变化规律和五角枫(张翠琴等, 2015 )以经度和纬度并存的变异趋势, 山杏不同种群表型性状的变异与地理经度、年降水量以及年日照时间3个主要地理气候因子相关. ...
用同工酶研究马尾松种群的遗传结构.
1
1988
... 采用巢式方差分析研究山杏的23个表型性状的变异, 线性模型为: yijk = μ + αi + βj(i) + εijk .其中, yijk 为第i 个种群的第j 个个体的第k 个观测值, μ 为总体平均值; αi 为第i 个种群的效应值(固定), βj(i) 为第i 个种群内第j 个个体的效应值(随机), εijk 为第ijk 个观测值的试验误差.计算各性状平均值、标准偏差、变异系数(CV )和表型分化系数(V st ), 进行表型性状间的差异性检验和多重比较分析, 其中CV =标准偏差/ 平均值, V st = δ 2 t/s /(δ 2 t/s + δ 2 s ), 其中δ 2 t/s 为种群间方差值, δ 2 s 为种群内方差值(葛颂等, 1988 ).CV 表明了性状变异的离散s程度, V st 描述的是表型变异在种群间贡献的大小.计算表型性状与生态因子间的相关系数, 并对性状进行主成分分析.数据处理在Excel 2010和SPSS 20.0软件中完成. ...
川西云杉天然种群表型多样性.
2
2009
... 植物表型多样性是基因表达和所处环境交互作用的结果, 表现为种群在不同分布区的压力选择中发生不可逆变异, 并经稳定遗传产生表型变异, 表型变异在适应与进化上具有重要意义(李斌等, 2002 ).研究表型变异格局及其与地理生态因子的关系, 不仅有助于了解植物应对环境因子变化的可塑性, 同时也可以揭示遗传因素和环境因素在植物生态适应中的相互作用(李因刚等, 2014 ).近年来, 表型变异的研究主要集中在分析变异程度进行种质资源分类上, 前人已通过表型对杏(Armeniaca vulgaris )(刘娟等, 2014 )、秋子梨(Pyrus ussuriensis )(马腾等, 2011 ; 许靖诗等, 2015 )、无患子(Sapindus mukorossi )(刁松锋等, 2014 )、柿(Diospyros kaki )(梁玉琴等, 2015 )、五角枫(Acer pictum subsp. mono )(姬志峰等, 2012 )等大量树种进行了研究, 并解释了各个树种的遗传变异规律.在环境因素方面, 毛梾(Cornus walteri )(康永祥等, 2011 )种实性状与纬度和降水量呈梯度变化规律, 川西云杉(Picea likiangensis var. rubescens )(辜云杰等, 2009 )种实性状存在经度、纬度并存的变异模式.另外, 以往形态特征的研究中, 大多以分析各种源差异程度和选择优良性状的遗传材料为重点, 很少进一步在地理生态上 开展重要性状研究, 探究物种适应性和性状分布 格局. ...
... 在对内蒙古山杏文献资料搜集以及野外实地调查的基础上, 分别于2012年及2013年每年果实成熟的7-8月份完成叶和果实的野外采集工作.采样点分别位于内蒙古的扎赉特旗(P1)、科尔沁右翼前旗察尔森(P2)、科尔沁右翼中旗(P3)、扎鲁特旗(P4)、科尔沁左翼后旗(P5)、阿鲁科尔沁旗(P6)、巴林右旗(P7)、敖汉旗(P8)、克什克腾旗(P9)、卓资县(P10)、和林格尔县(P11)、土默特左旗万家沟(P12)、乌拉山(P13), 共计13个种群130个植株, 取样时株间距大于25 m, 即母树树高的5倍以上(辜云杰等, 2009 ).每一单株从东西南北4个方向随机采摘健康、无病虫害的果实、成熟叶片各10个, 低温保鲜带回实验室备用.采样点的地理位置、海拔和气候因子等见表1 .地理位置和海拔为采样时现场记录, 气候数据来源于中国气象数据网(data.cma.cn). ...
Allozyme Diversity in Plant Species. Sinauer Association,
1
1990
... 内蒙古山杏主要分布于大兴安岭南麓东坡和阴山山脉中段大青山南坡的灌丛草原和低山丘陵地区.通过对该分布区的13个种群的叶、果、核、仁等23个表型性状的研究发现, 山杏的表型性状在种群间和种群内都存在极显著差异, 这与杏(刘娟等, 2014 )、浙江柿(Diospyros glaucifolia )(井振华等, 2010 )、文冠果(Xanthoceras sorbifolia )(牟洪香等, 2007 )等经济树种的研究结果一致.表型分化系数在某种程度上反映了山杏的环境适应性, 种群表型分化系数的均值为73.03%, 高于山苍子(Litsea cubeba )(66.66%)(田胜平等, 2012 )、无患子(62.21%)(刁松锋等, 2014 )、浙江楠(Phoebe chekiangensis ) (41.43%)(李因刚等, 2014 )、浙江柿(40.15%)(井振华等, 2010 ), 和滇龙胆(Gentiana rigescens ) (73.14%) (杨维泽等, 2011 )相当, 低于夏蜡梅(Sinocalycanthus chinensis )(89.30%)(金则新等, 2012 ), 表明了山杏种群表型分化程度较高, 对环境的适应性较强, 在内蒙古横跨13.71个经度(1523 km)范围, 在224-2083 m海拔范围内自然分布, 造成了不同种群表型性状的异质性.种群间的变异程度大于种群内, 种群间的变异是表型变异的主要来源, 这可能与山杏的地理分布和繁殖系统有关.种群间变异的量值取决于其地理和生态分布区(杨继, 1991 ), 内蒙古的山杏多分布于荒山丘陵和沙地边缘, 东西地理分布的间断性和海拔的差异等地理隔离促进了种群间的分化.另外, 交配系统也是影响植物种群遗传结构的重要因素(Hamrick & Godt, 1990 ), 山杏主要为虫媒异花授粉, 其花粉的传播距离远低于风媒花植物, 且杏花开放主要受温度影响, 不同地理种群花期的差异阻碍了种群间的花粉传播和基因交流, 导致不同种群间的遗传分化加剧.山杏自然生境贫瘠, 结实量少, 且种仁易被鸟兽取食, 加上海拔障碍, 使种子传播受阻导致种群间种子交流困难, 从而促进了种群间的遗传分化.分化较大的种群在各自生境中的适应和进化促成了山杏丰富的遗传多样性. ...
浙江柿天然群体表型多样性研究.
2
2010
... 内蒙古山杏主要分布于大兴安岭南麓东坡和阴山山脉中段大青山南坡的灌丛草原和低山丘陵地区.通过对该分布区的13个种群的叶、果、核、仁等23个表型性状的研究发现, 山杏的表型性状在种群间和种群内都存在极显著差异, 这与杏(刘娟等, 2014 )、浙江柿(Diospyros glaucifolia )(井振华等, 2010 )、文冠果(Xanthoceras sorbifolia )(牟洪香等, 2007 )等经济树种的研究结果一致.表型分化系数在某种程度上反映了山杏的环境适应性, 种群表型分化系数的均值为73.03%, 高于山苍子(Litsea cubeba )(66.66%)(田胜平等, 2012 )、无患子(62.21%)(刁松锋等, 2014 )、浙江楠(Phoebe chekiangensis ) (41.43%)(李因刚等, 2014 )、浙江柿(40.15%)(井振华等, 2010 ), 和滇龙胆(Gentiana rigescens ) (73.14%) (杨维泽等, 2011 )相当, 低于夏蜡梅(Sinocalycanthus chinensis )(89.30%)(金则新等, 2012 ), 表明了山杏种群表型分化程度较高, 对环境的适应性较强, 在内蒙古横跨13.71个经度(1523 km)范围, 在224-2083 m海拔范围内自然分布, 造成了不同种群表型性状的异质性.种群间的变异程度大于种群内, 种群间的变异是表型变异的主要来源, 这可能与山杏的地理分布和繁殖系统有关.种群间变异的量值取决于其地理和生态分布区(杨继, 1991 ), 内蒙古的山杏多分布于荒山丘陵和沙地边缘, 东西地理分布的间断性和海拔的差异等地理隔离促进了种群间的分化.另外, 交配系统也是影响植物种群遗传结构的重要因素(Hamrick & Godt, 1990 ), 山杏主要为虫媒异花授粉, 其花粉的传播距离远低于风媒花植物, 且杏花开放主要受温度影响, 不同地理种群花期的差异阻碍了种群间的花粉传播和基因交流, 导致不同种群间的遗传分化加剧.山杏自然生境贫瘠, 结实量少, 且种仁易被鸟兽取食, 加上海拔障碍, 使种子传播受阻导致种群间种子交流困难, 从而促进了种群间的遗传分化.分化较大的种群在各自生境中的适应和进化促成了山杏丰富的遗传多样性. ...
... )、浙江柿(40.15%)(井振华等, 2010 ), 和滇龙胆(Gentiana rigescens ) (73.14%) (杨维泽等, 2011 )相当, 低于夏蜡梅(Sinocalycanthus chinensis )(89.30%)(金则新等, 2012 ), 表明了山杏种群表型分化程度较高, 对环境的适应性较强, 在内蒙古横跨13.71个经度(1523 km)范围, 在224-2083 m海拔范围内自然分布, 造成了不同种群表型性状的异质性.种群间的变异程度大于种群内, 种群间的变异是表型变异的主要来源, 这可能与山杏的地理分布和繁殖系统有关.种群间变异的量值取决于其地理和生态分布区(杨继, 1991 ), 内蒙古的山杏多分布于荒山丘陵和沙地边缘, 东西地理分布的间断性和海拔的差异等地理隔离促进了种群间的分化.另外, 交配系统也是影响植物种群遗传结构的重要因素(Hamrick & Godt, 1990 ), 山杏主要为虫媒异花授粉, 其花粉的传播距离远低于风媒花植物, 且杏花开放主要受温度影响, 不同地理种群花期的差异阻碍了种群间的花粉传播和基因交流, 导致不同种群间的遗传分化加剧.山杏自然生境贫瘠, 结实量少, 且种仁易被鸟兽取食, 加上海拔障碍, 使种子传播受阻导致种群间种子交流困难, 从而促进了种群间的遗传分化.分化较大的种群在各自生境中的适应和进化促成了山杏丰富的遗传多样性. ...
基于形态及分子标记的濒危植物夏蜡梅自然居群的遗传变异研究.
1
2012
... 内蒙古山杏主要分布于大兴安岭南麓东坡和阴山山脉中段大青山南坡的灌丛草原和低山丘陵地区.通过对该分布区的13个种群的叶、果、核、仁等23个表型性状的研究发现, 山杏的表型性状在种群间和种群内都存在极显著差异, 这与杏(刘娟等, 2014 )、浙江柿(Diospyros glaucifolia )(井振华等, 2010 )、文冠果(Xanthoceras sorbifolia )(牟洪香等, 2007 )等经济树种的研究结果一致.表型分化系数在某种程度上反映了山杏的环境适应性, 种群表型分化系数的均值为73.03%, 高于山苍子(Litsea cubeba )(66.66%)(田胜平等, 2012 )、无患子(62.21%)(刁松锋等, 2014 )、浙江楠(Phoebe chekiangensis ) (41.43%)(李因刚等, 2014 )、浙江柿(40.15%)(井振华等, 2010 ), 和滇龙胆(Gentiana rigescens ) (73.14%) (杨维泽等, 2011 )相当, 低于夏蜡梅(Sinocalycanthus chinensis )(89.30%)(金则新等, 2012 ), 表明了山杏种群表型分化程度较高, 对环境的适应性较强, 在内蒙古横跨13.71个经度(1523 km)范围, 在224-2083 m海拔范围内自然分布, 造成了不同种群表型性状的异质性.种群间的变异程度大于种群内, 种群间的变异是表型变异的主要来源, 这可能与山杏的地理分布和繁殖系统有关.种群间变异的量值取决于其地理和生态分布区(杨继, 1991 ), 内蒙古的山杏多分布于荒山丘陵和沙地边缘, 东西地理分布的间断性和海拔的差异等地理隔离促进了种群间的分化.另外, 交配系统也是影响植物种群遗传结构的重要因素(Hamrick & Godt, 1990 ), 山杏主要为虫媒异花授粉, 其花粉的传播距离远低于风媒花植物, 且杏花开放主要受温度影响, 不同地理种群花期的差异阻碍了种群间的花粉传播和基因交流, 导致不同种群间的遗传分化加剧.山杏自然生境贫瘠, 结实量少, 且种仁易被鸟兽取食, 加上海拔障碍, 使种子传播受阻导致种群间种子交流困难, 从而促进了种群间的遗传分化.分化较大的种群在各自生境中的适应和进化促成了山杏丰富的遗传多样性. ...
山西霍山五角枫不同海拔种群的表型多样性研究.
1
2012
... 植物表型多样性是基因表达和所处环境交互作用的结果, 表现为种群在不同分布区的压力选择中发生不可逆变异, 并经稳定遗传产生表型变异, 表型变异在适应与进化上具有重要意义(李斌等, 2002 ).研究表型变异格局及其与地理生态因子的关系, 不仅有助于了解植物应对环境因子变化的可塑性, 同时也可以揭示遗传因素和环境因素在植物生态适应中的相互作用(李因刚等, 2014 ).近年来, 表型变异的研究主要集中在分析变异程度进行种质资源分类上, 前人已通过表型对杏(Armeniaca vulgaris )(刘娟等, 2014 )、秋子梨(Pyrus ussuriensis )(马腾等, 2011 ; 许靖诗等, 2015 )、无患子(Sapindus mukorossi )(刁松锋等, 2014 )、柿(Diospyros kaki )(梁玉琴等, 2015 )、五角枫(Acer pictum subsp. mono )(姬志峰等, 2012 )等大量树种进行了研究, 并解释了各个树种的遗传变异规律.在环境因素方面, 毛梾(Cornus walteri )(康永祥等, 2011 )种实性状与纬度和降水量呈梯度变化规律, 川西云杉(Picea likiangensis var. rubescens )(辜云杰等, 2009 )种实性状存在经度、纬度并存的变异模式.另外, 以往形态特征的研究中, 大多以分析各种源差异程度和选择优良性状的遗传材料为重点, 很少进一步在地理生态上 开展重要性状研究, 探究物种适应性和性状分布 格局. ...
毛梾天然群体种实表型多样性研究.
2
2011
... 植物表型多样性是基因表达和所处环境交互作用的结果, 表现为种群在不同分布区的压力选择中发生不可逆变异, 并经稳定遗传产生表型变异, 表型变异在适应与进化上具有重要意义(李斌等, 2002 ).研究表型变异格局及其与地理生态因子的关系, 不仅有助于了解植物应对环境因子变化的可塑性, 同时也可以揭示遗传因素和环境因素在植物生态适应中的相互作用(李因刚等, 2014 ).近年来, 表型变异的研究主要集中在分析变异程度进行种质资源分类上, 前人已通过表型对杏(Armeniaca vulgaris )(刘娟等, 2014 )、秋子梨(Pyrus ussuriensis )(马腾等, 2011 ; 许靖诗等, 2015 )、无患子(Sapindus mukorossi )(刁松锋等, 2014 )、柿(Diospyros kaki )(梁玉琴等, 2015 )、五角枫(Acer pictum subsp. mono )(姬志峰等, 2012 )等大量树种进行了研究, 并解释了各个树种的遗传变异规律.在环境因素方面, 毛梾(Cornus walteri )(康永祥等, 2011 )种实性状与纬度和降水量呈梯度变化规律, 川西云杉(Picea likiangensis var. rubescens )(辜云杰等, 2009 )种实性状存在经度、纬度并存的变异模式.另外, 以往形态特征的研究中, 大多以分析各种源差异程度和选择优良性状的遗传材料为重点, 很少进一步在地理生态上 开展重要性状研究, 探究物种适应性和性状分布 格局. ...
... 变异系数越大, 性状观测值离散程度越大, 表型多样性程度越高.山杏叶、果、核、仁性状的平均变异系数大小顺序为: 核(15.67%) >叶(14.51%) >仁(14.13%) >果(12.11%), 核与叶的变异系数较大可能与日照及海拔有关, 采样过程中发现P13种群海拔(2083 m)和年日照时间(3205 h)都高于其他种群, 其形态表现出叶子小, 树形小, 干上多刺, 结果少, 果核大, 核壳厚等具有适应意义的自我保护特性, 类似情况见于西南桦(Betula alnoides )(曾杰等, 2005 )在低海拔、风速小地区增大种翅以促进种子散布.另外P11种群的果实大于其他种群, 可能与其在果实形态上与普通杏接近有关.山杏叶、核、果、仁的表征质量和大小的性状变异系数均值为15.25%, 而叶形指数、果形指数、核形指数、仁形指数等表征形状的性状变异系数均值仅为9.45%, 表明山杏质量和大小的变异程度较大, 而形状较为稳定, 这与对毛梾(康永祥等, 2011 )和无患子(刁松锋等, 2014 )表型性状的研究结果一致.单果质量、核干质量、仁干质量等重要经济性状的遗传变异较大, 受外界环境选择的压力较大, 在选择上更有潜力, 这为山杏的仁用良种选育提供了丰富的物质 基础. ...
白皮松天然种群种实性状表型多样性研究.
1
2002
... 植物表型多样性是基因表达和所处环境交互作用的结果, 表现为种群在不同分布区的压力选择中发生不可逆变异, 并经稳定遗传产生表型变异, 表型变异在适应与进化上具有重要意义(李斌等, 2002 ).研究表型变异格局及其与地理生态因子的关系, 不仅有助于了解植物应对环境因子变化的可塑性, 同时也可以揭示遗传因素和环境因素在植物生态适应中的相互作用(李因刚等, 2014 ).近年来, 表型变异的研究主要集中在分析变异程度进行种质资源分类上, 前人已通过表型对杏(Armeniaca vulgaris )(刘娟等, 2014 )、秋子梨(Pyrus ussuriensis )(马腾等, 2011 ; 许靖诗等, 2015 )、无患子(Sapindus mukorossi )(刁松锋等, 2014 )、柿(Diospyros kaki )(梁玉琴等, 2015 )、五角枫(Acer pictum subsp. mono )(姬志峰等, 2012 )等大量树种进行了研究, 并解释了各个树种的遗传变异规律.在环境因素方面, 毛梾(Cornus walteri )(康永祥等, 2011 )种实性状与纬度和降水量呈梯度变化规律, 川西云杉(Picea likiangensis var. rubescens )(辜云杰等, 2009 )种实性状存在经度、纬度并存的变异模式.另外, 以往形态特征的研究中, 大多以分析各种源差异程度和选择优良性状的遗传材料为重点, 很少进一步在地理生态上 开展重要性状研究, 探究物种适应性和性状分布 格局. ...
黄土高原不同县域山杏种质遗传多样性研究.
1
2011
... 目前对山杏的研究多集中在主要经济性状变异(李明等, 2011 ; 包文泉等, 2014 )、成分提取(马玉花等, 2007 )、生长生殖(刘梦培等, 2014 )、抗寒性(张睿等, 2005 )和S基因型鉴定(刘梦培, 2015 )等方面.关于山杏天然生态种群的表型多样性以及与地理生态的适应关系少有研究, 这在一定程度上制约了山杏遗传多样性和良种选育工作的开展.本研究以内蒙古13个天然分布的山杏种群为研究对象, 结合地理生态因子调查分析叶、果、核、仁的23个表型性状, 旨在揭示内蒙古山杏种群间和种群内的变异规律和性状分布格局, 为山杏种质资源收集和良种选育工作提供科学依据. ...
思茅松天然群体种实表型变异.
1
2013
... 根据主成分分析以及成分得分散点图, P4、P5、P6种群可归为一类, 其地理分布距离较近, 海拔和年日照时间基本相同, 水热条件较为接近; 而P1、P2、P8、P9种群归为一类, 其年降水量彼此接近且海拔和年平均气温相对较低; P3、P7、P10、P12种群归为一类, 其年日照时间较长, 接近3000 h; P11和P13种群本应归为一类, 但由于乌拉山的海拔(2083 m)比和林格尔县(1160 m)高将近一倍, 其较长的日照时间和较少的降水量, 使其种群表型性状产生较为独特的变异, 形成叶小, 干上生刺, 结果量少, 核壳很厚等一系列自我保护机制(李帅锋等, 2013 ).而聚类分析的结果与主成分分析结果基本一致, 不同种群间表型的差异与地理气候因子密切相关, 由此可见, 利用地理区划对山杏种群进行表型上的划分是可行的.而要进一步解读表型变异和生态因子的关系, 应该利用系统的生态学方法, 扩大样地及样本量, 以得到更加精确可信的结果. ...
浙江楠种群表型变异.
2
2014
... 植物表型多样性是基因表达和所处环境交互作用的结果, 表现为种群在不同分布区的压力选择中发生不可逆变异, 并经稳定遗传产生表型变异, 表型变异在适应与进化上具有重要意义(李斌等, 2002 ).研究表型变异格局及其与地理生态因子的关系, 不仅有助于了解植物应对环境因子变化的可塑性, 同时也可以揭示遗传因素和环境因素在植物生态适应中的相互作用(李因刚等, 2014 ).近年来, 表型变异的研究主要集中在分析变异程度进行种质资源分类上, 前人已通过表型对杏(Armeniaca vulgaris )(刘娟等, 2014 )、秋子梨(Pyrus ussuriensis )(马腾等, 2011 ; 许靖诗等, 2015 )、无患子(Sapindus mukorossi )(刁松锋等, 2014 )、柿(Diospyros kaki )(梁玉琴等, 2015 )、五角枫(Acer pictum subsp. mono )(姬志峰等, 2012 )等大量树种进行了研究, 并解释了各个树种的遗传变异规律.在环境因素方面, 毛梾(Cornus walteri )(康永祥等, 2011 )种实性状与纬度和降水量呈梯度变化规律, 川西云杉(Picea likiangensis var. rubescens )(辜云杰等, 2009 )种实性状存在经度、纬度并存的变异模式.另外, 以往形态特征的研究中, 大多以分析各种源差异程度和选择优良性状的遗传材料为重点, 很少进一步在地理生态上 开展重要性状研究, 探究物种适应性和性状分布 格局. ...
... 内蒙古山杏主要分布于大兴安岭南麓东坡和阴山山脉中段大青山南坡的灌丛草原和低山丘陵地区.通过对该分布区的13个种群的叶、果、核、仁等23个表型性状的研究发现, 山杏的表型性状在种群间和种群内都存在极显著差异, 这与杏(刘娟等, 2014 )、浙江柿(Diospyros glaucifolia )(井振华等, 2010 )、文冠果(Xanthoceras sorbifolia )(牟洪香等, 2007 )等经济树种的研究结果一致.表型分化系数在某种程度上反映了山杏的环境适应性, 种群表型分化系数的均值为73.03%, 高于山苍子(Litsea cubeba )(66.66%)(田胜平等, 2012 )、无患子(62.21%)(刁松锋等, 2014 )、浙江楠(Phoebe chekiangensis ) (41.43%)(李因刚等, 2014 )、浙江柿(40.15%)(井振华等, 2010 ), 和滇龙胆(Gentiana rigescens ) (73.14%) (杨维泽等, 2011 )相当, 低于夏蜡梅(Sinocalycanthus chinensis )(89.30%)(金则新等, 2012 ), 表明了山杏种群表型分化程度较高, 对环境的适应性较强, 在内蒙古横跨13.71个经度(1523 km)范围, 在224-2083 m海拔范围内自然分布, 造成了不同种群表型性状的异质性.种群间的变异程度大于种群内, 种群间的变异是表型变异的主要来源, 这可能与山杏的地理分布和繁殖系统有关.种群间变异的量值取决于其地理和生态分布区(杨继, 1991 ), 内蒙古的山杏多分布于荒山丘陵和沙地边缘, 东西地理分布的间断性和海拔的差异等地理隔离促进了种群间的分化.另外, 交配系统也是影响植物种群遗传结构的重要因素(Hamrick & Godt, 1990 ), 山杏主要为虫媒异花授粉, 其花粉的传播距离远低于风媒花植物, 且杏花开放主要受温度影响, 不同地理种群花期的差异阻碍了种群间的花粉传播和基因交流, 导致不同种群间的遗传分化加剧.山杏自然生境贫瘠, 结实量少, 且种仁易被鸟兽取食, 加上海拔障碍, 使种子传播受阻导致种群间种子交流困难, 从而促进了种群间的遗传分化.分化较大的种群在各自生境中的适应和进化促成了山杏丰富的遗传多样性. ...
河南省柿种质资源表型多样性研究.
1
2015
... 植物表型多样性是基因表达和所处环境交互作用的结果, 表现为种群在不同分布区的压力选择中发生不可逆变异, 并经稳定遗传产生表型变异, 表型变异在适应与进化上具有重要意义(李斌等, 2002 ).研究表型变异格局及其与地理生态因子的关系, 不仅有助于了解植物应对环境因子变化的可塑性, 同时也可以揭示遗传因素和环境因素在植物生态适应中的相互作用(李因刚等, 2014 ).近年来, 表型变异的研究主要集中在分析变异程度进行种质资源分类上, 前人已通过表型对杏(Armeniaca vulgaris )(刘娟等, 2014 )、秋子梨(Pyrus ussuriensis )(马腾等, 2011 ; 许靖诗等, 2015 )、无患子(Sapindus mukorossi )(刁松锋等, 2014 )、柿(Diospyros kaki )(梁玉琴等, 2015 )、五角枫(Acer pictum subsp. mono )(姬志峰等, 2012 )等大量树种进行了研究, 并解释了各个树种的遗传变异规律.在环境因素方面, 毛梾(Cornus walteri )(康永祥等, 2011 )种实性状与纬度和降水量呈梯度变化规律, 川西云杉(Picea likiangensis var. rubescens )(辜云杰等, 2009 )种实性状存在经度、纬度并存的变异模式.另外, 以往形态特征的研究中, 大多以分析各种源差异程度和选择优良性状的遗传材料为重点, 很少进一步在地理生态上 开展重要性状研究, 探究物种适应性和性状分布 格局. ...
新疆杏种质资源表型多样性研究.
2
2014
... 植物表型多样性是基因表达和所处环境交互作用的结果, 表现为种群在不同分布区的压力选择中发生不可逆变异, 并经稳定遗传产生表型变异, 表型变异在适应与进化上具有重要意义(李斌等, 2002 ).研究表型变异格局及其与地理生态因子的关系, 不仅有助于了解植物应对环境因子变化的可塑性, 同时也可以揭示遗传因素和环境因素在植物生态适应中的相互作用(李因刚等, 2014 ).近年来, 表型变异的研究主要集中在分析变异程度进行种质资源分类上, 前人已通过表型对杏(Armeniaca vulgaris )(刘娟等, 2014 )、秋子梨(Pyrus ussuriensis )(马腾等, 2011 ; 许靖诗等, 2015 )、无患子(Sapindus mukorossi )(刁松锋等, 2014 )、柿(Diospyros kaki )(梁玉琴等, 2015 )、五角枫(Acer pictum subsp. mono )(姬志峰等, 2012 )等大量树种进行了研究, 并解释了各个树种的遗传变异规律.在环境因素方面, 毛梾(Cornus walteri )(康永祥等, 2011 )种实性状与纬度和降水量呈梯度变化规律, 川西云杉(Picea likiangensis var. rubescens )(辜云杰等, 2009 )种实性状存在经度、纬度并存的变异模式.另外, 以往形态特征的研究中, 大多以分析各种源差异程度和选择优良性状的遗传材料为重点, 很少进一步在地理生态上 开展重要性状研究, 探究物种适应性和性状分布 格局. ...
... 内蒙古山杏主要分布于大兴安岭南麓东坡和阴山山脉中段大青山南坡的灌丛草原和低山丘陵地区.通过对该分布区的13个种群的叶、果、核、仁等23个表型性状的研究发现, 山杏的表型性状在种群间和种群内都存在极显著差异, 这与杏(刘娟等, 2014 )、浙江柿(Diospyros glaucifolia )(井振华等, 2010 )、文冠果(Xanthoceras sorbifolia )(牟洪香等, 2007 )等经济树种的研究结果一致.表型分化系数在某种程度上反映了山杏的环境适应性, 种群表型分化系数的均值为73.03%, 高于山苍子(Litsea cubeba )(66.66%)(田胜平等, 2012 )、无患子(62.21%)(刁松锋等, 2014 )、浙江楠(Phoebe chekiangensis ) (41.43%)(李因刚等, 2014 )、浙江柿(40.15%)(井振华等, 2010 ), 和滇龙胆(Gentiana rigescens ) (73.14%) (杨维泽等, 2011 )相当, 低于夏蜡梅(Sinocalycanthus chinensis )(89.30%)(金则新等, 2012 ), 表明了山杏种群表型分化程度较高, 对环境的适应性较强, 在内蒙古横跨13.71个经度(1523 km)范围, 在224-2083 m海拔范围内自然分布, 造成了不同种群表型性状的异质性.种群间的变异程度大于种群内, 种群间的变异是表型变异的主要来源, 这可能与山杏的地理分布和繁殖系统有关.种群间变异的量值取决于其地理和生态分布区(杨继, 1991 ), 内蒙古的山杏多分布于荒山丘陵和沙地边缘, 东西地理分布的间断性和海拔的差异等地理隔离促进了种群间的分化.另外, 交配系统也是影响植物种群遗传结构的重要因素(Hamrick & Godt, 1990 ), 山杏主要为虫媒异花授粉, 其花粉的传播距离远低于风媒花植物, 且杏花开放主要受温度影响, 不同地理种群花期的差异阻碍了种群间的花粉传播和基因交流, 导致不同种群间的遗传分化加剧.山杏自然生境贫瘠, 结实量少, 且种仁易被鸟兽取食, 加上海拔障碍, 使种子传播受阻导致种群间种子交流困难, 从而促进了种群间的遗传分化.分化较大的种群在各自生境中的适应和进化促成了山杏丰富的遗传多样性. ...
内蒙古居群抗寒山杏繁殖生态学研究.
1
2014
... 目前对山杏的研究多集中在主要经济性状变异(李明等, 2011 ; 包文泉等, 2014 )、成分提取(马玉花等, 2007 )、生长生殖(刘梦培等, 2014 )、抗寒性(张睿等, 2005 )和S基因型鉴定(刘梦培, 2015 )等方面.关于山杏天然生态种群的表型多样性以及与地理生态的适应关系少有研究, 这在一定程度上制约了山杏遗传多样性和良种选育工作的开展.本研究以内蒙古13个天然分布的山杏种群为研究对象, 结合地理生态因子调查分析叶、果、核、仁的23个表型性状, 旨在揭示内蒙古山杏种群间和种群内的变异规律和性状分布格局, 为山杏种质资源收集和良种选育工作提供科学依据. ...
仁用杏自交不亲和的授粉特性和S基因型鉴定研究
1
2015
... 目前对山杏的研究多集中在主要经济性状变异(李明等, 2011 ; 包文泉等, 2014 )、成分提取(马玉花等, 2007 )、生长生殖(刘梦培等, 2014 )、抗寒性(张睿等, 2005 )和S基因型鉴定(刘梦培, 2015 )等方面.关于山杏天然生态种群的表型多样性以及与地理生态的适应关系少有研究, 这在一定程度上制约了山杏遗传多样性和良种选育工作的开展.本研究以内蒙古13个天然分布的山杏种群为研究对象, 结合地理生态因子调查分析叶、果、核、仁的23个表型性状, 旨在揭示内蒙古山杏种群间和种群内的变异规律和性状分布格局, 为山杏种质资源收集和良种选育工作提供科学依据. ...
1
... 参照《杏种质资源描述规范和数据标准》(刘宁和刘威生, 2006 )和《中国果树志: 杏卷》(张加延和张钊, 2003 )选择遗传上相对稳定、易于获得和测定的23个表型性状.用电子天平精确到0.01 g测单果质量、核干质量、仁干质量, 用游标卡尺精确到0.01 mm测叶长、叶宽、叶柄长、叶尖长、果纵径、果横径、果侧径、核纵径、核横径、核侧径、核壳厚、仁纵径、仁横径、仁侧径等性状.根据叶片长宽, 果实的纵径、横径, 核的纵径、横径, 仁的纵径、横径分别计算叶形指数、果形指数、核形指数和仁形指数.根据单果质量、干核质量和干仁质量计算出核率和出仁率. ...
内蒙古赤峰地区野生秋子梨调查研究.
1
2011
... 植物表型多样性是基因表达和所处环境交互作用的结果, 表现为种群在不同分布区的压力选择中发生不可逆变异, 并经稳定遗传产生表型变异, 表型变异在适应与进化上具有重要意义(李斌等, 2002 ).研究表型变异格局及其与地理生态因子的关系, 不仅有助于了解植物应对环境因子变化的可塑性, 同时也可以揭示遗传因素和环境因素在植物生态适应中的相互作用(李因刚等, 2014 ).近年来, 表型变异的研究主要集中在分析变异程度进行种质资源分类上, 前人已通过表型对杏(Armeniaca vulgaris )(刘娟等, 2014 )、秋子梨(Pyrus ussuriensis )(马腾等, 2011 ; 许靖诗等, 2015 )、无患子(Sapindus mukorossi )(刁松锋等, 2014 )、柿(Diospyros kaki )(梁玉琴等, 2015 )、五角枫(Acer pictum subsp. mono )(姬志峰等, 2012 )等大量树种进行了研究, 并解释了各个树种的遗传变异规律.在环境因素方面, 毛梾(Cornus walteri )(康永祥等, 2011 )种实性状与纬度和降水量呈梯度变化规律, 川西云杉(Picea likiangensis var. rubescens )(辜云杰等, 2009 )种实性状存在经度、纬度并存的变异模式.另外, 以往形态特征的研究中, 大多以分析各种源差异程度和选择优良性状的遗传材料为重点, 很少进一步在地理生态上 开展重要性状研究, 探究物种适应性和性状分布 格局. ...
超临界CO2 流体萃取杏仁油工艺研究.
1
2007
... 目前对山杏的研究多集中在主要经济性状变异(李明等, 2011 ; 包文泉等, 2014 )、成分提取(马玉花等, 2007 )、生长生殖(刘梦培等, 2014 )、抗寒性(张睿等, 2005 )和S基因型鉴定(刘梦培, 2015 )等方面.关于山杏天然生态种群的表型多样性以及与地理生态的适应关系少有研究, 这在一定程度上制约了山杏遗传多样性和良种选育工作的开展.本研究以内蒙古13个天然分布的山杏种群为研究对象, 结合地理生态因子调查分析叶、果、核、仁的23个表型性状, 旨在揭示内蒙古山杏种群间和种群内的变异规律和性状分布格局, 为山杏种质资源收集和良种选育工作提供科学依据. ...
木本能源植物文冠果的表型多样性研究.
1
2007
... 内蒙古山杏主要分布于大兴安岭南麓东坡和阴山山脉中段大青山南坡的灌丛草原和低山丘陵地区.通过对该分布区的13个种群的叶、果、核、仁等23个表型性状的研究发现, 山杏的表型性状在种群间和种群内都存在极显著差异, 这与杏(刘娟等, 2014 )、浙江柿(Diospyros glaucifolia )(井振华等, 2010 )、文冠果(Xanthoceras sorbifolia )(牟洪香等, 2007 )等经济树种的研究结果一致.表型分化系数在某种程度上反映了山杏的环境适应性, 种群表型分化系数的均值为73.03%, 高于山苍子(Litsea cubeba )(66.66%)(田胜平等, 2012 )、无患子(62.21%)(刁松锋等, 2014 )、浙江楠(Phoebe chekiangensis ) (41.43%)(李因刚等, 2014 )、浙江柿(40.15%)(井振华等, 2010 ), 和滇龙胆(Gentiana rigescens ) (73.14%) (杨维泽等, 2011 )相当, 低于夏蜡梅(Sinocalycanthus chinensis )(89.30%)(金则新等, 2012 ), 表明了山杏种群表型分化程度较高, 对环境的适应性较强, 在内蒙古横跨13.71个经度(1523 km)范围, 在224-2083 m海拔范围内自然分布, 造成了不同种群表型性状的异质性.种群间的变异程度大于种群内, 种群间的变异是表型变异的主要来源, 这可能与山杏的地理分布和繁殖系统有关.种群间变异的量值取决于其地理和生态分布区(杨继, 1991 ), 内蒙古的山杏多分布于荒山丘陵和沙地边缘, 东西地理分布的间断性和海拔的差异等地理隔离促进了种群间的分化.另外, 交配系统也是影响植物种群遗传结构的重要因素(Hamrick & Godt, 1990 ), 山杏主要为虫媒异花授粉, 其花粉的传播距离远低于风媒花植物, 且杏花开放主要受温度影响, 不同地理种群花期的差异阻碍了种群间的花粉传播和基因交流, 导致不同种群间的遗传分化加剧.山杏自然生境贫瘠, 结实量少, 且种仁易被鸟兽取食, 加上海拔障碍, 使种子传播受阻导致种群间种子交流困难, 从而促进了种群间的遗传分化.分化较大的种群在各自生境中的适应和进化促成了山杏丰富的遗传多样性. ...
山苍子天然种群叶片和种实性状的表型多样性.
1
2012
... 内蒙古山杏主要分布于大兴安岭南麓东坡和阴山山脉中段大青山南坡的灌丛草原和低山丘陵地区.通过对该分布区的13个种群的叶、果、核、仁等23个表型性状的研究发现, 山杏的表型性状在种群间和种群内都存在极显著差异, 这与杏(刘娟等, 2014 )、浙江柿(Diospyros glaucifolia )(井振华等, 2010 )、文冠果(Xanthoceras sorbifolia )(牟洪香等, 2007 )等经济树种的研究结果一致.表型分化系数在某种程度上反映了山杏的环境适应性, 种群表型分化系数的均值为73.03%, 高于山苍子(Litsea cubeba )(66.66%)(田胜平等, 2012 )、无患子(62.21%)(刁松锋等, 2014 )、浙江楠(Phoebe chekiangensis ) (41.43%)(李因刚等, 2014 )、浙江柿(40.15%)(井振华等, 2010 ), 和滇龙胆(Gentiana rigescens ) (73.14%) (杨维泽等, 2011 )相当, 低于夏蜡梅(Sinocalycanthus chinensis )(89.30%)(金则新等, 2012 ), 表明了山杏种群表型分化程度较高, 对环境的适应性较强, 在内蒙古横跨13.71个经度(1523 km)范围, 在224-2083 m海拔范围内自然分布, 造成了不同种群表型性状的异质性.种群间的变异程度大于种群内, 种群间的变异是表型变异的主要来源, 这可能与山杏的地理分布和繁殖系统有关.种群间变异的量值取决于其地理和生态分布区(杨继, 1991 ), 内蒙古的山杏多分布于荒山丘陵和沙地边缘, 东西地理分布的间断性和海拔的差异等地理隔离促进了种群间的分化.另外, 交配系统也是影响植物种群遗传结构的重要因素(Hamrick & Godt, 1990 ), 山杏主要为虫媒异花授粉, 其花粉的传播距离远低于风媒花植物, 且杏花开放主要受温度影响, 不同地理种群花期的差异阻碍了种群间的花粉传播和基因交流, 导致不同种群间的遗传分化加剧.山杏自然生境贫瘠, 结实量少, 且种仁易被鸟兽取食, 加上海拔障碍, 使种子传播受阻导致种群间种子交流困难, 从而促进了种群间的遗传分化.分化较大的种群在各自生境中的适应和进化促成了山杏丰富的遗传多样性. ...
祁连山青海云杉天然群体的种实性状表型多样性.
1
2008
... 山杏13个种群的分布区均属于中温带季风气候, 具有降水量少且分布不均匀、寒暑变化剧烈的特点, 纬度范围40.38°-46.59° N, 经度范围109.04°- 122.75° E, 海拔范围245-2083 m, 广泛的经度范围和海拔差异使得表型性状存在较大的异质性.除无霜期外, 其他地理生态因子对山杏的表型性状都有显著影响.7个生态因子中, 经度对山杏表型性状的综合影响最大, 东部种群的叶形更圆, 叶尖趋于长尾尖, 果形更接近圆球形, 仁也更趋于饱满; 西部种群叶形卵圆, 叶尖趋于钝尖, 果形趋于长圆形.随着年降水量的增加, 叶子变大, 叶柄变长, 果更趋于圆球形.随着年日照时间的增加, 叶子变小, 核变大, 核壳变厚, 核形趋于长圆形, 仁变小.这可能是强光照下植物的适应机制, 叶子变小以减小蒸腾作用失水, 核壳变厚以保护种子在不利条件下的生存繁殖.而随着纬度的增加, 叶长变长, 果形指数减小.随着海拔的增加, 叶子变小, 核变大, 核壳变厚, 仁侧径变小, 果形趋于长圆, 其对性状的影响与年日照时间类似.年平均气温和无霜期对表型性状无显著影响.因此, 由于复杂的地理环境, 不同于青海云杉(Picea crassifolia )(王娅丽等, 2008 )单一的经度变化规律, 也不同于无患子(刁松锋等, 2014 )以纬度和年平均气温为主的变化规律和五角枫(张翠琴等, 2015 )以经度和纬度并存的变异趋势, 山杏不同种群表型性状的变异与地理经度、年降水量以及年日照时间3个主要地理气候因子相关. ...
中国野生秋子梨果实形态变异研究.
1
2015
... 植物表型多样性是基因表达和所处环境交互作用的结果, 表现为种群在不同分布区的压力选择中发生不可逆变异, 并经稳定遗传产生表型变异, 表型变异在适应与进化上具有重要意义(李斌等, 2002 ).研究表型变异格局及其与地理生态因子的关系, 不仅有助于了解植物应对环境因子变化的可塑性, 同时也可以揭示遗传因素和环境因素在植物生态适应中的相互作用(李因刚等, 2014 ).近年来, 表型变异的研究主要集中在分析变异程度进行种质资源分类上, 前人已通过表型对杏(Armeniaca vulgaris )(刘娟等, 2014 )、秋子梨(Pyrus ussuriensis )(马腾等, 2011 ; 许靖诗等, 2015 )、无患子(Sapindus mukorossi )(刁松锋等, 2014 )、柿(Diospyros kaki )(梁玉琴等, 2015 )、五角枫(Acer pictum subsp. mono )(姬志峰等, 2012 )等大量树种进行了研究, 并解释了各个树种的遗传变异规律.在环境因素方面, 毛梾(Cornus walteri )(康永祥等, 2011 )种实性状与纬度和降水量呈梯度变化规律, 川西云杉(Picea likiangensis var. rubescens )(辜云杰等, 2009 )种实性状存在经度、纬度并存的变异模式.另外, 以往形态特征的研究中, 大多以分析各种源差异程度和选择优良性状的遗传材料为重点, 很少进一步在地理生态上 开展重要性状研究, 探究物种适应性和性状分布 格局. ...
植物种内形态变异的机制及其研究方法.
1
1991
... 内蒙古山杏主要分布于大兴安岭南麓东坡和阴山山脉中段大青山南坡的灌丛草原和低山丘陵地区.通过对该分布区的13个种群的叶、果、核、仁等23个表型性状的研究发现, 山杏的表型性状在种群间和种群内都存在极显著差异, 这与杏(刘娟等, 2014 )、浙江柿(Diospyros glaucifolia )(井振华等, 2010 )、文冠果(Xanthoceras sorbifolia )(牟洪香等, 2007 )等经济树种的研究结果一致.表型分化系数在某种程度上反映了山杏的环境适应性, 种群表型分化系数的均值为73.03%, 高于山苍子(Litsea cubeba )(66.66%)(田胜平等, 2012 )、无患子(62.21%)(刁松锋等, 2014 )、浙江楠(Phoebe chekiangensis ) (41.43%)(李因刚等, 2014 )、浙江柿(40.15%)(井振华等, 2010 ), 和滇龙胆(Gentiana rigescens ) (73.14%) (杨维泽等, 2011 )相当, 低于夏蜡梅(Sinocalycanthus chinensis )(89.30%)(金则新等, 2012 ), 表明了山杏种群表型分化程度较高, 对环境的适应性较强, 在内蒙古横跨13.71个经度(1523 km)范围, 在224-2083 m海拔范围内自然分布, 造成了不同种群表型性状的异质性.种群间的变异程度大于种群内, 种群间的变异是表型变异的主要来源, 这可能与山杏的地理分布和繁殖系统有关.种群间变异的量值取决于其地理和生态分布区(杨继, 1991 ), 内蒙古的山杏多分布于荒山丘陵和沙地边缘, 东西地理分布的间断性和海拔的差异等地理隔离促进了种群间的分化.另外, 交配系统也是影响植物种群遗传结构的重要因素(Hamrick & Godt, 1990 ), 山杏主要为虫媒异花授粉, 其花粉的传播距离远低于风媒花植物, 且杏花开放主要受温度影响, 不同地理种群花期的差异阻碍了种群间的花粉传播和基因交流, 导致不同种群间的遗传分化加剧.山杏自然生境贫瘠, 结实量少, 且种仁易被鸟兽取食, 加上海拔障碍, 使种子传播受阻导致种群间种子交流困难, 从而促进了种群间的遗传分化.分化较大的种群在各自生境中的适应和进化促成了山杏丰富的遗传多样性. ...
云南滇龙胆居群表型多样性及其与环境关系研究.
1
2011
... 内蒙古山杏主要分布于大兴安岭南麓东坡和阴山山脉中段大青山南坡的灌丛草原和低山丘陵地区.通过对该分布区的13个种群的叶、果、核、仁等23个表型性状的研究发现, 山杏的表型性状在种群间和种群内都存在极显著差异, 这与杏(刘娟等, 2014 )、浙江柿(Diospyros glaucifolia )(井振华等, 2010 )、文冠果(Xanthoceras sorbifolia )(牟洪香等, 2007 )等经济树种的研究结果一致.表型分化系数在某种程度上反映了山杏的环境适应性, 种群表型分化系数的均值为73.03%, 高于山苍子(Litsea cubeba )(66.66%)(田胜平等, 2012 )、无患子(62.21%)(刁松锋等, 2014 )、浙江楠(Phoebe chekiangensis ) (41.43%)(李因刚等, 2014 )、浙江柿(40.15%)(井振华等, 2010 ), 和滇龙胆(Gentiana rigescens ) (73.14%) (杨维泽等, 2011 )相当, 低于夏蜡梅(Sinocalycanthus chinensis )(89.30%)(金则新等, 2012 ), 表明了山杏种群表型分化程度较高, 对环境的适应性较强, 在内蒙古横跨13.71个经度(1523 km)范围, 在224-2083 m海拔范围内自然分布, 造成了不同种群表型性状的异质性.种群间的变异程度大于种群内, 种群间的变异是表型变异的主要来源, 这可能与山杏的地理分布和繁殖系统有关.种群间变异的量值取决于其地理和生态分布区(杨继, 1991 ), 内蒙古的山杏多分布于荒山丘陵和沙地边缘, 东西地理分布的间断性和海拔的差异等地理隔离促进了种群间的分化.另外, 交配系统也是影响植物种群遗传结构的重要因素(Hamrick & Godt, 1990 ), 山杏主要为虫媒异花授粉, 其花粉的传播距离远低于风媒花植物, 且杏花开放主要受温度影响, 不同地理种群花期的差异阻碍了种群间的花粉传播和基因交流, 导致不同种群间的遗传分化加剧.山杏自然生境贫瘠, 结实量少, 且种仁易被鸟兽取食, 加上海拔障碍, 使种子传播受阻导致种群间种子交流困难, 从而促进了种群间的遗传分化.分化较大的种群在各自生境中的适应和进化促成了山杏丰富的遗传多样性. ...
1
... 山杏(Armeniaca sibirica )为蔷薇科李亚科杏属植物(俞德浚, 1986 ), 主要分布在俄罗斯的西伯利亚、蒙古国和我国, 我国主要分布于内蒙古、辽宁、河北等省区, 耐干旱、瘠薄, 适应性强, 作为先锋树种应用于沙漠防护林营造, 杏仁粗脂肪含量45.64%, 粗蛋白含量28%, 苦杏仁苷含量4.5% (包文泉, 2013 ), 是重要的木本油料树种和药用资源.近几年来, 荒漠化的加重以及全球气候的变化以及农业耕地和城市发展冲突的加剧, 使得山杏在防治荒漠化上的生态功能以及作为木本油料植物节约耕地的重大意义越来越受到关注. ...
广西西南桦天然居群的表型变异.
1
2005
... 变异系数越大, 性状观测值离散程度越大, 表型多样性程度越高.山杏叶、果、核、仁性状的平均变异系数大小顺序为: 核(15.67%) >叶(14.51%) >仁(14.13%) >果(12.11%), 核与叶的变异系数较大可能与日照及海拔有关, 采样过程中发现P13种群海拔(2083 m)和年日照时间(3205 h)都高于其他种群, 其形态表现出叶子小, 树形小, 干上多刺, 结果少, 果核大, 核壳厚等具有适应意义的自我保护特性, 类似情况见于西南桦(Betula alnoides )(曾杰等, 2005 )在低海拔、风速小地区增大种翅以促进种子散布.另外P11种群的果实大于其他种群, 可能与其在果实形态上与普通杏接近有关.山杏叶、核、果、仁的表征质量和大小的性状变异系数均值为15.25%, 而叶形指数、果形指数、核形指数、仁形指数等表征形状的性状变异系数均值仅为9.45%, 表明山杏质量和大小的变异程度较大, 而形状较为稳定, 这与对毛梾(康永祥等, 2011 )和无患子(刁松锋等, 2014 )表型性状的研究结果一致.单果质量、核干质量、仁干质量等重要经济性状的遗传变异较大, 受外界环境选择的压力较大, 在选择上更有潜力, 这为山杏的仁用良种选育提供了丰富的物质 基础. ...
五角枫种群表型多样性.
1
2015
... 山杏13个种群的分布区均属于中温带季风气候, 具有降水量少且分布不均匀、寒暑变化剧烈的特点, 纬度范围40.38°-46.59° N, 经度范围109.04°- 122.75° E, 海拔范围245-2083 m, 广泛的经度范围和海拔差异使得表型性状存在较大的异质性.除无霜期外, 其他地理生态因子对山杏的表型性状都有显著影响.7个生态因子中, 经度对山杏表型性状的综合影响最大, 东部种群的叶形更圆, 叶尖趋于长尾尖, 果形更接近圆球形, 仁也更趋于饱满; 西部种群叶形卵圆, 叶尖趋于钝尖, 果形趋于长圆形.随着年降水量的增加, 叶子变大, 叶柄变长, 果更趋于圆球形.随着年日照时间的增加, 叶子变小, 核变大, 核壳变厚, 核形趋于长圆形, 仁变小.这可能是强光照下植物的适应机制, 叶子变小以减小蒸腾作用失水, 核壳变厚以保护种子在不利条件下的生存繁殖.而随着纬度的增加, 叶长变长, 果形指数减小.随着海拔的增加, 叶子变小, 核变大, 核壳变厚, 仁侧径变小, 果形趋于长圆, 其对性状的影响与年日照时间类似.年平均气温和无霜期对表型性状无显著影响.因此, 由于复杂的地理环境, 不同于青海云杉(Picea crassifolia )(王娅丽等, 2008 )单一的经度变化规律, 也不同于无患子(刁松锋等, 2014 )以纬度和年平均气温为主的变化规律和五角枫(张翠琴等, 2015 )以经度和纬度并存的变异趋势, 山杏不同种群表型性状的变异与地理经度、年降水量以及年日照时间3个主要地理气候因子相关. ...
1
... 参照《杏种质资源描述规范和数据标准》(刘宁和刘威生, 2006 )和《中国果树志: 杏卷》(张加延和张钊, 2003 )选择遗传上相对稳定、易于获得和测定的23个表型性状.用电子天平精确到0.01 g测单果质量、核干质量、仁干质量, 用游标卡尺精确到0.01 mm测叶长、叶宽、叶柄长、叶尖长、果纵径、果横径、果侧径、核纵径、核横径、核侧径、核壳厚、仁纵径、仁横径、仁侧径等性状.根据叶片长宽, 果实的纵径、横径, 核的纵径、横径, 仁的纵径、横径分别计算叶形指数、果形指数、核形指数和仁形指数.根据单果质量、干核质量和干仁质量计算出核率和出仁率. ...
甜山杏优株幼果抗寒性研究.
1
2005
... 目前对山杏的研究多集中在主要经济性状变异(李明等, 2011 ; 包文泉等, 2014 )、成分提取(马玉花等, 2007 )、生长生殖(刘梦培等, 2014 )、抗寒性(张睿等, 2005 )和S基因型鉴定(刘梦培, 2015 )等方面.关于山杏天然生态种群的表型多样性以及与地理生态的适应关系少有研究, 这在一定程度上制约了山杏遗传多样性和良种选育工作的开展.本研究以内蒙古13个天然分布的山杏种群为研究对象, 结合地理生态因子调查分析叶、果、核、仁的23个表型性状, 旨在揭示内蒙古山杏种群间和种群内的变异规律和性状分布格局, 为山杏种质资源收集和良种选育工作提供科学依据. ...



{kind=link}
{kind=link}
{kind=link}
{kind=link}