陈云
CHEN Yun
通讯作者:
责任编辑:
收稿日期: 2016-06-6
接受日期: 2016-09-21
网络出版日期: 2016-11-10
版权声明: 2016 植物生态学报编辑部 本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。
基金资助:
作者简介:
* 共同第一作者 Co-first author (E-mail: 464272387@qq.com; tingwang01@126.com)
展开
摘要
以木札岭世界地质公园3 hm2的温带落叶阔叶林为研究对象, 分析了群落的物种组成、胸径结构、群落分类和主要物种的空间分布格局。结果显示: 样地内共有木本植物31科52属85种, 以蔷薇科、桦木科、卫矛科、忍冬科、杨柳科和槭树科为主, 锐齿槲栎(Quercus aliena var. acuteserrata)和华山松(Pinus armandii)为群落内优势种, 稀有种和偶见种分别占总物种数的20.0%和28.24%。物种径级分布遵从典型的倒“J”型, 植物群落更新状况良好。多元回归树经过交叉验证认为可将该样地划分为4个群落类型: 1)锐齿槲栎+华山松+秦岭木姜子(Litsea tsinlingensis) +微毛樱桃(Cerasus clarofolia) +三桠乌药(Lindera obtusiloba)群落; 2)锐齿槲栎+腺柳(Salix chaenomeloides) +湖北花楸(Sorbus hupehensis)群落; 3)锐齿槲栎+臭椿(Ailanthus altissima) +微毛樱桃+秦岭木姜子群落; 4)锐齿槲栎+白蜡(Fraxinus chinensis) +秦岭木姜子+山梅花(Philadelphus incanus)群落。在完全随机分布模型下, 样地中主要物种在整个研究尺度中全部呈聚集分布, 并且这些物种主要聚集分布在不同的生境中, 具有明显的地形生境偏好;在异质性泊松分布模型下, 这些物种在不同尺度上主要呈随机分布或规则分布。该研究结果表明地形生境异质性是影响木札岭样地物种空间分布的重要因素。通过该研究可增加对木札岭世界地质公园植物组成、群落结构和群落分布的认识, 同时可为该区域生物多样性保护和森林管理提供参考。
关键词:
Abstract
Aims The objective of this paper is to quantify the species composition and spatial distribution pattern in a deciduous broad-leaved forest in temperate to subtropical ecological transition zone.
Methods In this study, a 3-hm2 forest was selected in the temperate to subtropical ecological transition zone to analyze the community species composition, structure of diameter at breast height, community classification and spatial distribution pattern of dominant tree species.
Important findings Our results showed that in the plot there were 85 species, 52 genera and 31 families, mainly composed of Betulaceae, Celastraceae, Caprifoliaceae, Salicaceae and Aceraceae. Quercus aliena var. acuteserrata and Pinus armandii are dominant species of the community tree layer. The rare species and occasional species accounted for 20.0% and 28.24% of total species respectively. Size distribution of all species showed an invert J-shape, which indicates that the community is in a stable and normal growth status. Using multiple regression trees, the community in this plot can be divided into four categories: 1) Quercus aliena var. acuteserrata + Pinus armandii + Litsea tsinlingensis + Cerasus clarofolia + Lindera obtusiloba; 2) Quercus aliena var. acuteserrata + Salix chaenomeloides + Sorbus hupehensis; 3) Quercus aliena var. acuteserrata + Ailanthus altissima + Cerasus clarofolia + Litsea tsinlingensis; 4) Quercus aliena var. acuteserrata + Fraxinus chinensis + Litsea tsinlingensis + Philadelphus incanus. Under the completely random distribution model, the main species in the plot display clustered distributions, with the different species occurring in different habitat types, showing obvious terrain habitat preferences. However, under the heterogeneous Poisson distribution model, these species at different scales are distributed randomly or regularly. This study helps to understand the plant community species composition of the Muzhaling World Geopark, community structure and community distribution. The results show that the terrain habitat heterogeneity is an important factor influencing the spatial distribution of the species. The present work improves the understanding of plant community in Muzhaling World Geopark, and provides technical reference for biodiversity conservation and forest management of this area.
Keywords:
植物群落结构不仅能反映群落中物种之间的关系, 也能反映环境对物种生存的影响, 它是群落生态学研究的基础(李博, 2000)。而不同区域的植物群落在物种组成、结构功能和分布格局等方面存在很大差异。生态过渡带是两个或者多个生态系统之 间的过渡区域, 其植物群落有自身的结构特征,同时还兼具周围生态系统的特点, 更为重要的是, 生态过渡带对环境变化的反应更为敏感(袁志良等, 2013)。
物种的空间分布格局是物种自身特性、种间关系和环境条件综合作用的结果(Sterner et al., 1986)。已有很多研究表明扩散限制、密度制约和生境异质性是影响物种空间分布格局的3个重要因素(梁爽等, 2014; 王慧杰等, 2016)。扩散限制主要强调了机遇因素在物种分布中的作用(Hubbell, 2001); 密度制约认为物种种群扩张会受到负反馈的调节作用, 可避免单优群落的出现(祝燕, 2009); 生境异质性认为环境的筛选作用使物种聚集分布在适宜其生长的环境中, 生态位分化促使物种共存(Harms et al., 2001)。众多的解释机制也说明了物种空间分布格局的形成机制是复杂的。分析物种的空间分布格局特征将有助于揭示群落结构的形成机理和潜在的生态学过程。
锐齿槲栎(Quercus aliena var. acuteserrata)林是河南省落叶阔林区内较为稳定的森林植被类型, 对该区域生态系统结构和功能的维持起着重要的作用(王亚平等, 2014)。本研究区域选择在河南省木札岭世界地质公园, 地处典型的暖温带和北亚热带生态过渡区。木札岭锐齿槲栎林群落保护较好, 近代没有受到大面积砍伐, 人为干扰较小。未查阅到该区域植物群落的相关研究文献。
本研究选择木札岭核心区域, 设置了3 hm2长期动态监测样地。基于第一次调查数据, 初步探讨以下问题: (1)木札岭森林由哪些树种组成?优势种、伴生种、稀有种、优势科有哪些?胸径结构有什么特点?(2)基于多元回归树(multivariate regression tree, MRT)对该群落进行分类。(3)采用不同模型分析主要物种的空间分布格局, 初步探讨该区域植物空间分布格局的形成机理。希望通过本研究增加人们对该区域植物群落的认识, 为生物多样性保护和管理措施的制定提供参考。
木札岭位于河南洛阳市嵩县东南部, 属于伏牛山国家级自然保护区, 是世界地质公园。总面积125 km2, 森林覆盖率达98.5%, 属大陆性季风气候区。木札岭地处暖温带和北亚热带过渡区, 位于秦岭与淮河重要地理分界线上。特殊的地理区位, 孕育了其独特、原始和丰富多彩的生物资源。通过2015年对该区域生物资源的全面普查, 保护区内有国家级保护植物领春木(Eupulea pleiosperma)、水曲柳(Fraxinus mandshurica)、暖木(Meliosma veitchiorum)、毛蒟(Piper puberulum)在内的植物2000多种, 包括金钱豹(Panthera pardus)、香獐(Moschus moschiferus)等在内的高等动物260多种。
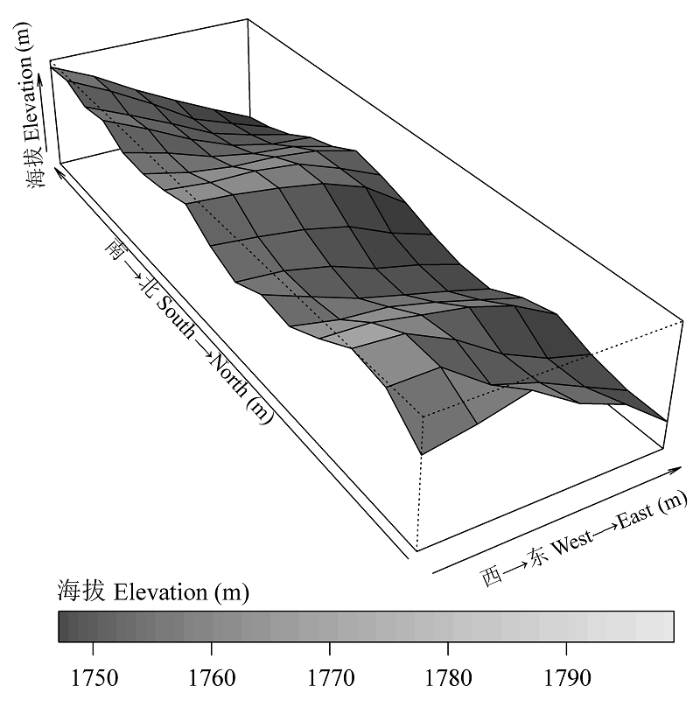
2015年, 经过实地踏查, 在木札岭核心地带选择保存较好的森林群落, 设置东西100 m、南北300 m、面积为3 hm2的固定监测样地(平均海拔1776.65 m)(图1)。样地地理坐标为33.75° N, 112.25° E。用全站仪将样地划分成75个20 m × 20 m的样方, 再把每个样方划分为16个5 m × 5 m的小样方, 每个20 m × 20 m样方的4个角用水泥桩作永久标记。调查样地内所有胸径(DBH) ≥1.0 cm的木本植物, 并挂铝牌标记, 每个需要测量的植株在1.3 m处涂上油漆, 记录种名、胸径、树高、坐标等。
图1 木札岭3 hm2永久监测样地地形图。
Fig. 1 The topography map of the Muzhaling plot with the size of 3 hm2.
根据Hubbell和Foster (1986)的定义, 把≤1株·hm-2的物种定义为稀有种, 2-10株·hm-2的物种定义为偶见种。根据物种的重要值确定样地内的优势物种, 重要值的计算公式为: 重要值= (相对多度(%) +相对频度(%) +相对胸高断面积(%))/3, 其中相对多度为某个种的株数与所有种的总株数的比, 相对频度为某个种基于20 m × 20 m样方出现的次数与所有种出现的总次数的比, 相对胸高断面积为 某个种的胸高断面积与所有种的胸高断面积之和 的比。
采用MRT进行群落分类, 自变量为75个样方的海拔、坡向、坡度和凹凸度, 因变量为物种在75个样方内的重要值, 此处重要值的计算公式为: 重要值= (相对多度(%) +相对胸高断面积(%))/2。MRT基于R语言中mvpart程序包进行。海拔采用的是样方44个顶点海拔的平均值; 样方4个顶点任取3个顶点可以组成4个不同的平面, 这4个平面与样方投影水平面夹角的平均值为样方的坡度; 这4个平面与正北方向偏差角度的平均值为坡向值; 凹凸度是样方的海拔减去相邻的8个样方海拔的平均值, 处于样方边缘样方的凹凸度为样方中心的海拔减去4个顶点海拔的平均值(Yamakura et al., 1995; Valencia et al., 2004; Nee, 2005)。其中坡向进行了转换, 使正北方向为-1, 正南方向为1, 转换采用的公式为: 坡向= -cos (坡向×π/180)。
双关联函数g(r)函数是由Ripley’s K函数推演而来, g函数能更加敏感地判别某一尺度上点的实际分布偏离期望值的程度。因此本文采用g(r)函数分析方法分析优势种的空间分布格局(Stoyan & Penttinen, 2000)。选择的模型有完全空间随机模型和异质泊松模型。完全空间随机模型又称均质性泊松过程, 是空间点格局中使用最广泛的一种, 该模型假设任何一个个体在研究区域内任何一个位置上出现的几率相等, 同时点与点之间是彼此独立的, 在生态学中常用来假设物种的空间分布不受任何生物或非生物过程影响。异质泊松模型依据密度函数决定个体的分布, 可以排除大尺度环境异质性的影响, 与完全随机模型相比, 它主要用来模拟物种与其生境之间的相互关系(郭屹立等, 2015)。本研究基于R语言中spatstat程序包分析了这两种模型下物种的空间分布格局。
样地内共有木本植物31科52属85种, 其中华山松(Pinus armandii)和油松(Pinus tabuliformis)为裸子植物, 其他都为被子植物。属和种占优势的科主要有蔷薇科(10属16种)、桦木科(4属6种)、卫矛科(2属6种)、忍冬科(3属5种)、杨柳科(2属5种)和槭树科(1属5种)。样地内独立个体数为8487株(包括分枝819个分枝), 单位面积个体数为2829株·hm-2。本文所有统计分析数据均基于独立个体。
样地内优势种明显, 从重要值(表1)来看, 锐齿槲栎、秦岭木姜子(Litsea tsinlingensis)、华山松、微毛樱桃(Cerasus clarofolia)、三桠乌药(Lindera obtusiloba)、湖北花楸(Sorbus hupehensis)、油松、华东椴(Tilia japonica)、漆树(Toxicodendron vernicifluum)和腺柳(Salix chaenomeloides)的重要值分别是30.4%、4.8%、4.6%、4.3%、2.3%、2.2%、2.1%、2.0%、1.9%、1.7%, 这些物种数占总物种数的11.8%, 重要值占56.3%。从物种的多度来看, 样地内个体数最多的是锐齿槲栎, 共2237株, 占总植株数的26.2%。从胸高截面积来看, 样地内胸高断面积主要集中在锐齿槲栎(65.1 m2), 其次是华山松(7.7 m2)。样地中稀有种较多, 根据统计, 样地内有稀有种17种, 偶见种24种, 分别占总物种数的20.0%和28.24%。
表1 木札岭森林样地重要值前10位的物种
Table 1 Top 10 species based on important value in the Muzhaling forest plot
| 物种 Species | 多度 Abundance | 平均胸径 Mean diameter at breast height (cm) | 胸高断面积 Basal area (m2) | 重要值 Important value (%) |
|---|---|---|---|---|
| 锐齿槲栎 Quercus aliena var. acuteserrata | 2 237 | 16.5 | 65.1 | 30.4 |
| 秦岭木姜子 Litsea tsinlingensis | 931 | 4.1 | 2.2 | 4.8 |
| 华山松 Pinus armandii | 429 | 12.5 | 7.7 | 4.6 |
| 微毛樱桃 Cerasus clarofolia | 727 | 6.0 | 3.3 | 4.3 |
| 三桠乌药 Lindera obtusiloba | 382 | 5.7 | 1.4 | 2.3 |
| 湖北花楸 Sorbus hupehensis | 433 | 2.9 | 0.5 | 2.2 |
| 油松 Pinus tabulaeformis | 145 | 14.7 | 3.5 | 2.1 |
| 华东椴 Sorbus hupehensis | 267 | 7.1 | 1.7 | 2.0 |
| 漆树 Toxicodendron vernicifluum | 167 | 11.1 | 2.7 | 1.9 |
| 腺柳 Salix chaenomeloides | 165 | 11.3 | 2.1 | 1.7 |
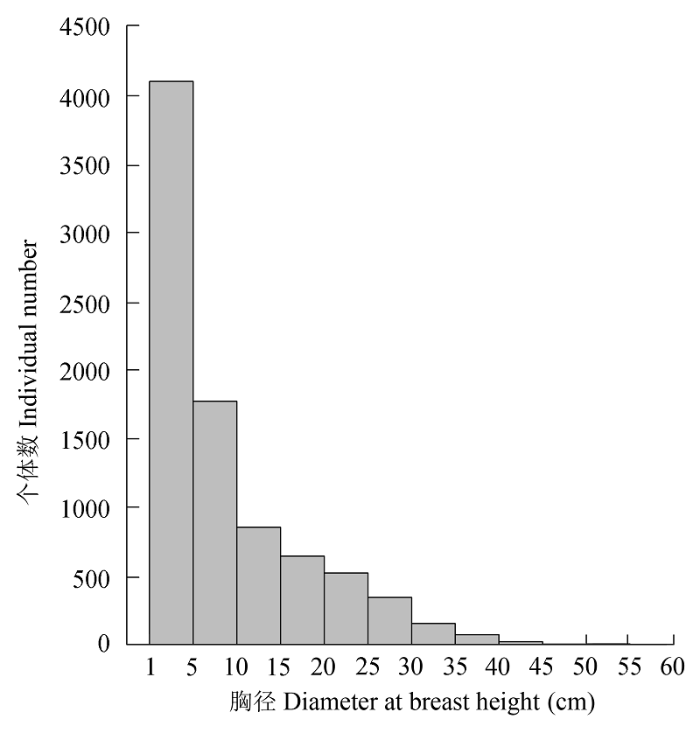
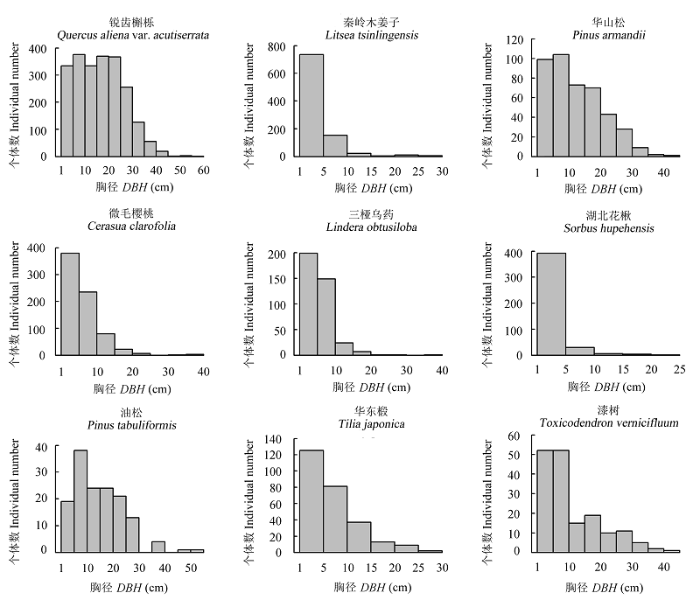
样地内植株径级分布遵从典型的倒“J”型, 随着胸径的增大, 植物株数呈迅速减少的趋势(图2)。其中, 径级1-5 cm的个体数达4100株, 占总个体数的48.31%。样地中主要物种的径级分布如图3所示, 各有不同。秦岭木姜子、微毛樱桃、三桠乌药、湖北花楸和华东椴的径级结构呈倒“J”型分布; 锐齿槲栎、油松和漆树的径级结构呈波动型分布; 华山松的径级结构呈接近正比例下降趋势。
图2 木札岭3 hm2样地物种胸径结构分布图。
Fig. 2 Distribution of diameter at breast height class in Mu- zhaling forest plot of 3 hm2.
图3 木札岭3 hm2样地9个主要物种胸径结构分布图。
Fig. 3 Size-class of diameter at breast height (DBH) distributions of nine dominant species in the Muzhaling forest plot t of 3 hm2.
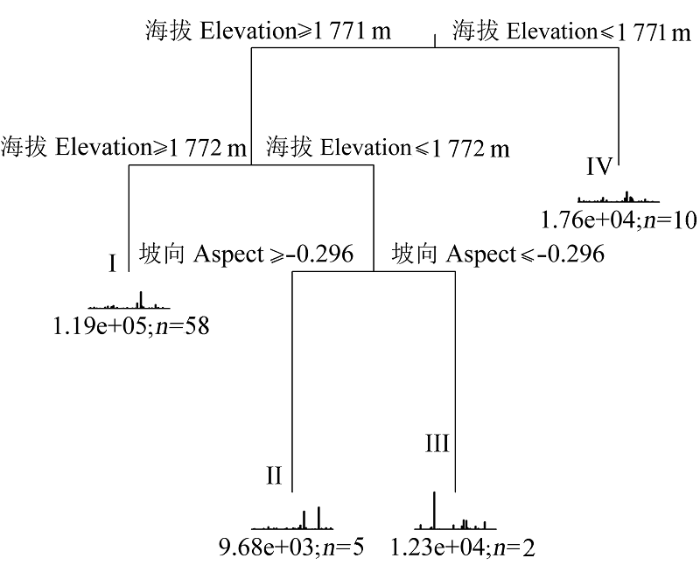
MRT以海拔和坡度将75个样方划分为4类(图4)。第一次分割以海拔1771 m为节点, 分为两组, 随后第一组又以海拔和坡度分为3组。因为植物群落空间分布存在连续性, 因此我们参照赖江山等(2010)的方法, 对分类结果进行修正, 即如果一个样方与其相邻的样方均不属于一类, 则这个样方变为周围样方中比例最大的那类群落。本分类结果中, 第410号样方本属于群落IV, 但相邻的8个样方均属于群落I, 修正后该样方归属于群落I。
图4 基于多元回归树的木札岭森林样地群落划分。I、II、III和IV为4个群落编号。n, 样方数量。
Fig. 4 Multivariate regression tree for community classification in the Muzhaling forest plot. I, II, III and IV are the number of four communities. n, number of plots.
依据植物群落分类和命名原则(赖江山等, 2010), 4类群落分别为: 1)锐齿槲栎+华山松+秦岭木姜子+微毛樱桃+三桠乌药群落; 2)锐齿槲栎+腺柳+湖北花楸群落; 3)锐齿槲栎+臭椿(Ailanthus altissima) +微毛樱桃+秦岭木姜子群落; 4)锐齿槲栎+白蜡(Fraxinus chinensis) +秦岭木姜子+山梅花(Philadelphus incanus)群落。
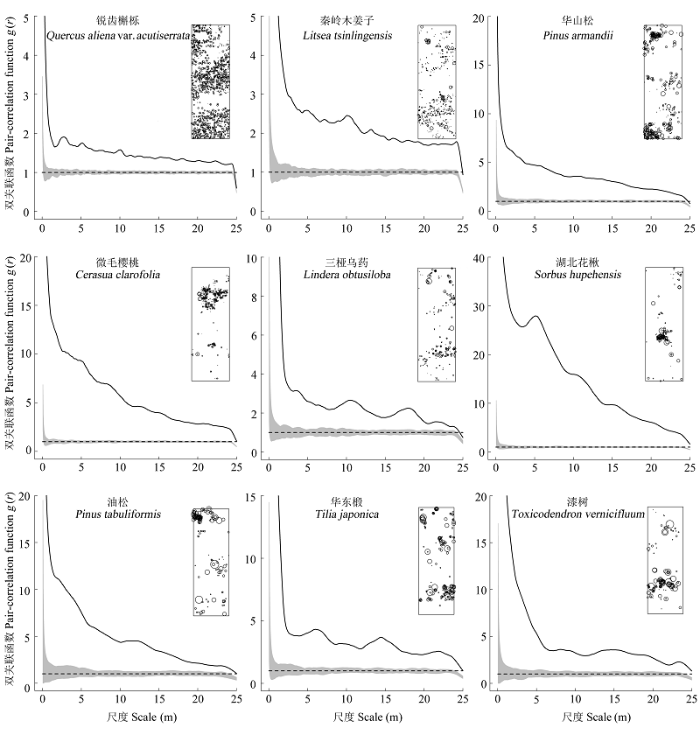
通过点格局结果可以看出, 在完全随机状态模型下, 样地重要值前9位物种的空间分布格局在整个研究尺度上都表现为明显的聚集分布格局, 随着尺度的增大空间分布格局的聚集强度减弱(图5)。
图5 木札岭样地9个主要物种完全随机状态的空间分布格局。实线为实际观测值, 虚线为理论值, 灰色区间为置信区间。图中子图为物种在样地中的空间分布图。
Fig. 5 Spatial distribution pattern under complete spatial randomness null model of nine dominant species in the Muzhaling forest plot. The solid line is the observed value, and the dotted line for the theoretical value. Grey interval is the confidence interval. The subset shows the species distribution in the plot.
对比图中物种分布的散点图, 可以发现不同物种种群聚集分布于不同的地形生境中, 如锐齿槲栎主要聚集分布在山脊和缓坡附近; 华山松主要聚集分布在采光较好的陡坡上; 微毛樱桃主要聚集分布在阳坡上; 湖北花楸主要集中分布在沟谷; 华东椴和漆树主要聚集分布在山脊上。
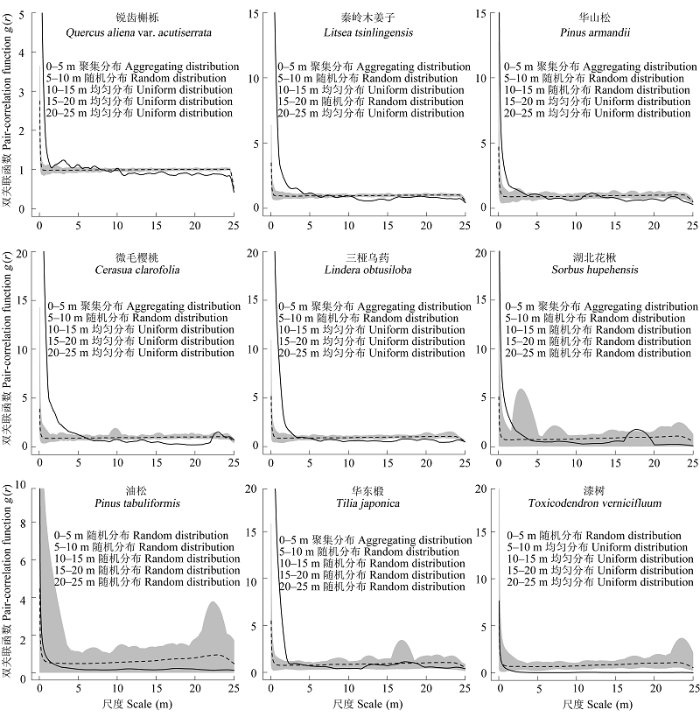
使用异质性泊松模型剔除生境异质性后发现, 这些物种的聚集程度都明显下降(图6)。油松在整个研究尺度上表现为随机分布; 漆树在0-5 m的尺度上呈随机分布, 随后呈规则分布; 其他7种物种全都在0-5 m的尺度左右呈聚集分布, 随后呈随机分布, 当尺度达15 m后, 逐渐转化为规则分布。
图6 木札岭样地主要物种异质性泊松分布下的空间分布格局。实线为实际观测值, 虚线为理论值, 灰色区间为置信区间。
Fig. 6 Spatial distribution pattern under heterogeneity Poisson null model of nine dominant species in the Muzhaling forest plot. The solid line is the observed value, and the dotted line for the theoretical value. Grey interval is the confidence interval.
样地内小径级物种占总个体数的48.31%, 所占比例较大, 这一方面说明样地中幼苗充足, 群落更新状况良好; 另一方面是因为样地内小乔木和灌木 (如秦岭木姜子和湖北花楸)数量较多, 但这些物种胸径很小。样地内优势种锐齿槲栎, 以及油松和漆树的径级结构呈波动型分布, 这可能暗示着该群落在生长过程中曾受到某种历史条件的干扰, 当然其中具体的原因还有待进一步的分析和监测。样地内胸径最大的物种为锐齿槲栎(55.2 cm)。值得注意的是, 本样地所有物种的平均胸径为8.92 cm, 这远大于亚热带的天童山样地(5.66 cm)、古田山样地(5.21 cm)和温带地区的小兴安岭样地(7.41 cm)(徐丽娜和金光泽, 2012; 宋永昌等, 2015), 这可能是因为样地选择在木札岭核心区域, 该区域植物群落在近代没有被大面积砍伐过, 群落保护较好。
很多关于植物群落分类的研究常采用聚类分析或双向指示种分析, 这些分类结果往往需要人为判别(黄永涛等, 2015)。自然分类一直是人们追求的目标。MRT是一种无需人为判别的一种较新的数量分类方法(De’Ath, 2002)。赖江山等(2010)基于古田山大样地详细介绍了MRT分类方法; 随后很多学者也尝试采用此方法在不同区域对植物群落进行划分; 张文静等(2015)比较了MRT和双向指示种分析两种方法, 认为划分大样地连续样方时, MRT更有优势。因此本研究采用MRT对木札岭样地进行群落划分, 经过交叉验证认为可将该样地划分为4类群落类型。在局部区域, 地形因子往往是形成生境的最根本因素, 特别是在生境异质性较大的地区(赖江山等, 2010)。本研究中MRT根据海拔和坡度将样方分为 4类, 海拔作为地理梯度, 包含了温度、湿度和光照等各种环境因子, 而坡度直接影响土壤水分和养分的分布(赖江山等, 2010)。因此, 本研究中MRT的分类结果基本反映了地形生境对样地中植物群落分布的影响。
已有研究表明, 在自然植物群落中, 大部分物种常常表现为聚集分布(祝燕, 2009)。本研究结果显示, 在完全随机分布模型下, 样地中主要物种在整个研究尺度中全部呈聚集分布, 这与已有的研究结果一致。Getzin等(2008)研究表明在大于10 m的尺度上, 如果呈现聚集分布, 就可以解释为是受到生境异质性的影响, 本研究中的目标物种在整个研究尺度上全部呈聚集分布。结合物种的空间分布散点图, 可以直观地发现这些物种聚集分布在不同的生境中, 具有明显的地形生境偏好。不同物种在不同的地形生境中形成不同的斑块, 导致物种之间相互隔离或者不相关联。同时, 在异质性泊松分布模型排除环境异质性的影响下, 这些物种在不同尺度上主要呈随机分布或规则分布。这就很好地证明了地形生境异质性是影响该群落物种分布的重要因素。这一结果与斯里兰卡热带森林样地(Wiegand et al., 2007)、广西弄岗北热带样地(郭屹立等, 2015)和长白山温带森林样地(Wang et al., 2010)的研究结果一致, 同时也很好地支持了生境隔离假说。
同时, 已有研究表明扩散限制是引起同种聚集的主要因素之一(黄运峰等, 2012; Jia et al., 2015)。一方面, 种子扩散大部分会分布在母株周围, 距离母株越远, 种子数量越少, 从而导致种群聚集; 另一面, 生境异质性也是种群聚集分布的重要因素(祝燕, 2009)。然而本研究中, 在较大尺度上(>20 m)所有目标物种仍表现出聚集格局, 显然这种分布格局更可能是由生境异质性引起的。
生境异质性效应可能会掩盖物种分布格局的形式和种群统计行为(祝燕, 2009), 因此有必要排除环境异质性的影响, 从而探讨植物之间纯粹的相互关系。完全随机分布模型下, 目标物种在整个研究尺度上全部呈聚集分布, 地形生境异质性显著影响着该群落物种分布。另外, 排除生境异质性的影响之后, 物种则主要呈随机分布或规则分布。这两种结果是不矛盾的, 只是生物学的解释不同。包含生境异质性描述的是研究样地内点的分布强度在大尺度上的变异; 排除生境异质性之后, 描述的是植物之间相互作用的规律(祝燕, 2009)。相关研究已表明成年树一般更倾向于随机或规则分布(祝燕, 2009)。由于种间和种内竞争引起死亡, 从而个体的空间分布越来越均匀(李立等, 2010)。木札岭森林群落没有被大面积砍伐过, 群落保存较好, 属于中老龄林, 因此种间和种内的竞争可能导致样地内主要物种呈随机分布或规则分布(李立等, 2010), 这就解释了在异质性泊松分布模型排除环境异质性的影响之后, 物种主要呈随机分布或规则分布这种现象。因此密度制约可能是影响物种随机分布或规则分布的一个原因。当然, 影响物种空间分布的原因是复杂的, 密度制约是否确切影响该区域物种分布还有待于进一步深入探索。而木札岭森林动态样地的建立, 为更好地理解森林群落的空间分布提供了很好的研究平台。
本研究结果增加了人们对木札岭世界地质公园植物组成、群落结构和群落分布的认识, 同时表明地形生境异质性是影响木札岭样地物种空间分布的重要因素, 为该区域植物群落的研究积累了基础资料, 同时为该区域生物多样性保护和森林管理提供了参考。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
| [1] |
Multivariate regression trees: A new technique for modeling species-environment relationships . |
| [2] |
Heterogeneity influences spatial patterns and demographics in forest stands .
|
| [3] |
Spatial distribution of tree species in a tropical karst seasonal rainforest in Nonggang, Guangxi, Southern China .广西弄岗北热带喀斯特季节性雨林监测样地种群空间点格局分析 . |
| [4] |
Habitat associations of trees and shrubs in a 50-ha neotropical forest plot . |
| [5] |
Spatial pattern of trees in tropical lowland rain forest in Bawangling of Hainan Island, China .海南岛霸王岭热带低地雨林树木的空间格局 . |
| [6] |
Quantitative classification of the subtropical evergreen-deciduous broadleaved mixed forest and the deciduous and evergreen species composition structure across two national nature reserves in the southwest of Hubei, China .鄂西南两个自然保护区亚热带常绿落叶阔叶混交林类型及其常绿和落叶物种组成结构分析 . |
| [7] |
The Unified Neutral Theory of Biodiversity and Biogeography . |
| [8] |
Commonness and rarity in a neotropical forest: Implications for tropical tree conservation . In: |
| [9] |
Effects of environmental and spatial heterogeneity on tree community assembly in Baotianman National Nature Reserve, Henan, China . |
| [10] |
Numerical classification of associations in subtropical evergreen broad-leaved forest based on multivariate regression trees—A case study of 24 hm2 Gutianshan forest plot in China .基于多元回归树的常绿阔叶林群丛数量分类——以古田山24公顷森林样地为例 . |
| [11] |
|
| [12] |
Spatial patterns of Castanopsis eyrei and Schima superba in mid-subtropical broadleaved evergreen forest in Gutianshan National Reserve, China .古田山常绿阔叶林优势树种甜槠和木荷的空间格局分析 . |
| [13] |
Spatial distribution pattern of the dominant speciesGironniera subaequalis in tropical montane rainforest of Jianfengling, Hainan Island, China .尖峰岭热带山地雨林优势树种白颜树空间分布格局 . |
| [14] |
The neutral theory of biodiversity: Do the numbers add up? |
| [15] |
Synthetic comparison of eight dynamics plots in evergreen broadleaf forests, China .中国常绿阔叶林8大动态监测样地植被的综合比较 . |
| [16] |
Testing for life historical changes in spatial patterns of four tropical tree species . |
| [17] |
Recent applications of point process methods in forestry statistics . |
| [18] |
Tree species distributions and local habitat variation in the Amazon: A large plot in eastern Ecuador . |
| [19] |
Density-dependent effects inPicea schrenkiana forests in Tianshan Mountains .天山雪岭云杉森林群落的密度制约效应 . |
| [20] |
Species associations in an old-growth temperate forest in Northeastern China . |
| [21] |
Species composition and spatial distribution in Baotianman National Nature Reserve shaw .宝天曼杂木林样地的物种组成及空间分布格局 . |
| [22] |
Species associations in a heterogeneous Sri Lankan dipterocarp forest . |
| [23] |
Species composition and community structure of a typical mixed broadleaved-Korean pine (Pinus koraiensis) forest plot in Liangshui Nature Reserve, Northeast China .小兴安岭凉水典型阔叶红松林动态监测样地: 物种组成与群落结构 . |
| [24] |
Topography of a large-scale research plot established within a tropical rain forest at Lambir, Sarawak . |
| [25] |
Species habitat correlation analysis in temperate- subtropical ecological transition zone .暖温带-北亚热带生态过渡区物种生境相关性分析 . |
| [26] |
A com- parison of multivariate regression tree and two-way indi- cator species analysis in plant community classification .多元回归树与双向指示种分析在群落分类中的应用比较 .
|
| [27] |
The Prevalence of Density Dependence in Gutianshan Subtropical Evergreen Broadleaved Forest, China .古田山亚热带常绿阔叶林密度制约普遍性研究 . |

Copyright © 2026 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19
![]()
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}