植物作为生态系统的第一生产者, 受环境变化的影响是直接而多方面的。叶片是植物进化过程中对环境变化最敏感且可塑性较大的器官, 在不同的选择压力下形成了各种适应类型(Hickey, 1984 ; 祁如虎等, 1998 ; Gielwanowska et al ., 2005 ), 其形态结构特征最能体现环境因子的影响或植物对环境的适应(王勋陵和王静, 1989 ; Krober et al ., 2015 )。
干旱半干旱区的荒漠环境属于一种极端环境, 降水稀少, 气候干燥, 昼夜温差大, 地面温度高, 土壤贫瘠, 风沙活动强烈……这些恶劣条件无时不在限制着荒漠植物的生存和繁衍。但即使在这种条件下, 仍有许多植物分布于干旱半干旱区, 且能很好地繁衍生息。那么, 这些植物有何特殊的形态结构来适应环境呢?如何通过植物形态结构特征判断其适应能力? 这些问题引发不少科学家对荒漠植物研究的极大关注。Schow提出“旱生植物”概念已有100多年了。 国内外不少学者研究过旱生植物的形态结构(Shield, 1950 ; 王勋陵和马骥, 1999 ; 严巧娣等, 2008 ; Liu et al ., 2012 , 2015 ; Perrone et al ., 2015 ), 但以各自有限数量的旱生植物形态结构描述为主, 至今未见对较大数量旱生植物叶片表皮微形态与结构特征进行数量化整理, 并对其类型划分、变化规律、演化趋势、适应对策等重要问题进行系统讨论的 报道。
植物叶片适应环境的形态结构变化是多方面的。在众多生态因子中, 叶片受水分、温度和光因子的影响最为显著(王勋陵和王静, 1989 ; 王勋陵, 1993 ; Aronne & de Micco 2001 ; Arena et al ., 2008 ; Jaleel et al ., 2009a , 2009b ), 主要表现为叶形的变化、叶片表皮附属物的变化、叶片的厚度及解剖结构的差异(Talukdar, 2015 )。对于荒漠植物来说, 夏季极端干旱时它们可能同时遭受以上各种环境因子的胁迫。但其最主要的限制因子为水分, 因而表现出典型的旱生植物形态结构特征。由于遗传因子与环境因子的共同作用, 在旱生植物结构的适应性上出现了两种演化趋势, 即趋同性和趋异性, 也就是说, 在相同环境下的植物演化出相似的形态结构适应性, 而不同环境下的同类植物演化出不同的形态结构(Lakusic et al ., 2010 )。因此, 要系统分析荒漠植物叶片形态结构特征的适应性变化, 需要将一个区域不同植物和不同梯度生境下的同一植物做全面分析。
植物叶片对环境的响应首先是叶片表皮对环境的感应和变化(Pal et al ., 2002 ; Bargel et al ., 2004 ; Koch et al ., 2009 ; Adebowale et al ., 2014 )。生长于干燥的气候和土壤条件下的荒漠植物, 叶表皮细胞会变小, 切向壁加厚, 具有内皮层。叶表皮角质膜厚度增加, 角质层由光滑变得粗糙, 能够反射阳光, 减少叶片表面空气的流动, 降低蒸腾作用, 防止水分过度丧失(贺金生等, 1994 ; 杨明博等, 2007 )。气孔的分布特征、密度和面积等都会受到影响, 气孔多分布于叶片下表皮, 这种分布模式既可促进植物与外界环境进行气体交换, 又能保持水分。随着环境中水分和湿度的减少, 气孔密度增加, 气孔面积则向小型化发展, 气孔多下陷形成气孔窝或其上有突出的角质膜(Sam et al ., 2000 ; Bosabalidis & Kofidis, 2002 ; 高建平等, 2003 ; Xu & Zhou, 2008 ; Gan et al ., 2010 ; de Micco & Aronne, 2012 )。叶表皮附属物(如表皮毛和表面疣状突起物)也发生变化, 它们可有效地避免叶肉组织受损(Gairola et al ., 2009 ; Koch et al ., 2009 ; Gomes et al ., 2011 ; Noman et al ., 2012 ), 表皮蜡质(waxes)晶体形成二维或三维结构, 具有降低表皮的可湿性以及反射强光等作用(Wagner et al ., 2003 ; Koch & Ensikat, 2008 )。
植物叶表皮微形态特征已用于解决物种分类问题、建立物种进化关系、阐明表皮微形态结构的生态适应性等多种研究目的(Dickison, 2000 ; Scatena et al ., 2005 ; Stenglein et al ., 2005 ; Aliero et al ., 2006 ; Chen et al ., 2010 ; Zhou & Xia, 2012 ; Adebowale et al ., 2014 ; Deng et al ., 2014 ; Zhang et al ., 2014 ; Francisco et al ., 2015 ; 朱广龙等, 2016 )。为了探讨荒漠植物叶片表皮微形态结构的适应特征及其亲缘关系, 选择分布于同一区域的不同植物和不同降水量梯度生境下的同一植物, 利用扫描电镜观察分析了117种荒漠植物叶片的表皮微形态结构, 旨在阐明荒漠植物对逆境的适应机理及其演化趋势, 并为优良固沙植物的筛选提供理论依据。
1 材料和方法
1.1 野外采样样品
荒漠植物叶片采自中国干旱半干旱荒漠区野外自然生境(包括人工固沙植被和自然演替植被), 包含28科74属117种200多个自然居群的植物叶片(或同化枝), 采集的物种及采集地地理位置见附录I。2012年和2013年的5-7月, 采集成熟的嫩叶和同化枝。长度小于1.0 cm的叶片直接放入固定液, 大于1.0 cm的叶片或同化枝取中段部分剪成0.5 cm × 0.5 cm大小或0.5 cm长度放入固定液, 同化枝选取0.5 cm中部片段放入固定液。固定液为2.5%戊二醛磷酸缓冲液(0.1 mol·L-1 , pH 7.2)。
1.2 扫描电镜观察
扫描电镜材料的制备参考Liu等(2012) 的方法, 制备过程如下: 将固定24 h后的材料经磷酸缓冲液(0.1 mol·L-1 , pH 7.2)清洗2次, 各10 min, 然后用1.0% OsO4 磷酸缓冲液进行最后固定过夜。过夜后的材料用以上磷酸缓冲液清洗后, 在不同浓度的乙醇(30%、50%、70%、96%、100%)中进行脱水, 乙醇浓度逐渐增加。然后在干燥器JFD-310中干燥后, 喷20 nm厚的胶体金, 用扫描电镜(JSM-6380, JEOL, Tokyo, Japan)观察叶上、下表皮微形态特征并进行拍照。
2 结果
2.1 荒漠植物叶片(或同化枝)表皮微形态基本特征
荒漠植物由于生境严酷, 叶片一般较小, 特别是灌木, 有些叶片缩小为细棒状或完全退化, 以同化枝来代替叶片进行光合过程而完成植物的生活史, 如麻黄属(Ephedra )、沙拐枣属(Calligonum )、梭梭属(Haloxylon )植物。因此, 对于有叶片的植物选取叶片部分, 而叶片退化的材料选取同化枝进行主要光合器官的表皮微形态结构特征分析。
通过对中国干旱半干旱荒漠区28科74属117种200多个自然居群的植物叶片表皮微形态特征的扫描, 发现荒漠植物叶片(或同化枝)表皮细微结构存在显著差异(表1 )。表皮细胞形态总体有两种: 不规则形和多边形, 但不同植物细胞形态各异, 棒状叶片和同化枝表皮细胞多为长条形, 片状叶片细胞形态一般为不规则或多边形。表皮细胞垂周壁式样有直线形、弓形或曲线形。荒漠植物叶片表皮的一个明显特征是表皮附属物相当丰富, 包括大量的表皮绒毛、角质膜蜡质片层或晶体颗粒、表面瘤状或疣状突起, 以及相对下陷且密度较低的气孔器。表皮绒毛类型多样, 从结构上可划分为具囊状(腺毛)和不具囊状(非腺毛)两类, 从形态上可划分为有囊状、柳叶状、针状、卷丝状、星状、小草状、棒状等, 且具有表皮绒毛的植物一般绒毛数量较多。表面蜡质晶体形态各异, 有颗粒状、直立鳞片状、膜片状、壳状或痂状、晶簇状、棒状以及多种形态的混合, 且大多数植物角质膜蜡质层晶体密度很高。灌木的气孔相对于草本来说, 多数为下陷型, 气孔密度为100-200个·mm-2 的植物较多, 有少数植物的气孔集中于叶面的其中一面, 而另一面没有气孔。
2.2 荒漠植物叶片(或同化枝)表皮微形态结构基本类型
按照表皮微形态结构和表皮附属物的组成, 荒漠植物表皮微形态可分为以下11个主要类型:
(1)表面粗糙 上下表皮细胞完全被表皮角质膜蜡质层覆盖, 蜡质层在表皮上如同一个罩壳阻断表皮与外界的直接接触, 表皮细胞形态需要去除蜡质层才可见, 蜡质形态有颗粒状、膜片状、晶簇状或多种形态的混合, 表面不平滑。代表植物有膜果麻黄(Ephedra przewalskii )(图1A )、霸王(Zygophyllum xanthoxylon )(图1B )灌木铁线莲(Clematis fruticosa )等。
图1 荒漠植物叶片(同化枝)表皮微形态结构特征。A, 膜果麻黄。B, 霸王。
Fig. 1 Micromorphological and structural characteristics of leaf (or assimilation branches) epidermis of desert plants. A, Ephedra przewalskii. B, Zygophyllum xanthoxylon .
(2)表面完全被毛 形态各异的表皮绒毛完全覆盖叶片上下表面(图2 ), 表皮细胞形态需要去除掉绒毛才可观察到, 绒毛的形态有柳叶状、针状、卷丝状、星状、小草状等。代表植物有沙枣(Elaeagnus angustifolia )(图2 A)、刺疙瘩(Olgaea tangutica )(图2 B)、柠条锦鸡儿(Caragana korshinskii )、卷叶锦鸡儿(Caragana ordosica )、沙冬青(Ammopiptanthus mongolicus )、唐古特白刺(Nitraria tangutorum )(图2 C)、珍珠猪毛菜(Salsola passerina )(图2 D)、驼绒藜(Ceratoides latens )(图2 E)、蓍状亚菊(Ajania achilloides )(图2 F)等。
图2 荒漠植物叶片表皮微形态结构特征。A, 沙枣。B, 刺疙瘩。C, 唐古特白刺。D, 珍珠猪毛菜。E, 驼绒藜。F, 蓍状亚菊。
Fig. 2 Micromorphological and structural characteristics of leaf epidermis of desert plants. A, Elaeagnus angustifolia . B, Olgaea tangutica . C, Nitraria tangutorum . D, Salsola passerina . E, Ceratoides latens . F, Ajania achilloides .
(3)表皮被囊状毛 上下表皮细胞形态清晰可见, 表皮绒毛附属物为囊状, 表皮有或无蜡质层覆盖(图3 )。代表植物有甘草(Glycyrrhiza uralensis )(图3 A)、百里香(Thymus mongolicus )(图3 B)、单脉大黄(Rheum uninerve )(图3 C)等。
图3 荒漠植物叶片表皮微形态结构特征。A, 甘草。B, 百里香。C, 单脉大黄。
Fig. 3 Micromorphological and structural characteristics of leaf epidermis of desert plants. A, Glycyrrhiza uralensis . B, Thymus mongolicus . C, Rheum uninerve .
(4)表皮囊状毛和其他形态绒毛混生 表皮细胞被完全覆盖或不完全覆盖, 囊状和非囊状绒毛共生在一起(图4 )。代表植物有: 蒙古莸(Caryopteris mongholica )(图4 A)、香青兰(Dracocephalum moldavica )(图4 B)、白莲蒿(Artemisia sacrorum )(图4 C)、星毛短舌菊(Brachanthemum pulvinatum )(图4 D)等。
图4 荒漠植物叶片表皮微形态结构特征。A, 蒙古莸。B, 香青兰。C, 白莲蒿。D, 星毛短舌菊。
Fig. 4 Micromorphological and structural characteristics of leaf epidermis of desert plants. A, Caryopteris mongholica . B, Dracocephalum moldavica . C, Artemisia sacrorum . D, Brachanthemum pulvinatum .
(5)上下表面完全异面 上表皮被绒毛, 下表皮主要被粗糙痂状、壳状或膜片状蜡质层。代表植物主要是柳属(Salix )植物(图5 ), 如旱柳(Salix capitata ) (图5 A、5B)、黄柳(Salix gordejevii )(图5 C、5D)、乌柳(Salix microstachya )、沙柳(Salix psammophila )等。
图5 荒漠植物叶片表皮微形态结构特征。A, 旱柳上表皮。B, 旱柳下表皮。C, 黄柳上表皮。D, 黄柳下表皮。
Fig. 5 Micromorphological and structural characteristics of leaf epidermis of desert plants. A, Adaxial surface of Salix capitata . B, Abaxial surface of Salix capitata . C, Adaxial surface of Salix gordejevii . D, Abaxial surface of Salix gordejevii .
(6)气孔器集中于下表面 上表皮光滑, 无气孔器, 下表皮有高密度气孔器, 代表植物有蒙古扁桃(Prunus mongolica )(图6A 、6B)、柄扁桃(Prunus pedunculata )、杠柳(Periploca sepium )(图6C 、6D)等。
图6 荒漠植物叶片表皮微形态结构特征。A, 蒙古扁桃上表皮。B, 蒙古扁桃下表皮。C, 杠柳上表皮。D, 杠柳下表皮。
Fig. 6 Micromorphological and structural characteristics of leaf epidermis of desert plants. A, Adaxial surface of Prunus mongolica . B, Abaxial surface of Prunus mongolica . C, Adaxial surface of Periploca sepium . D, Abaxial surface of Periploca sepium .
(7)疣状 表皮无绒毛附属物, 表皮细胞突起的宽度大于高度, 基部不收缩(图7 )。代表植物有合头草(Sympegma regelii )(图7 A)、芦苇(Phragmites com- munis )(图7 B)等。
图7 荒漠植物叶片表皮微形态结构特征。A, 合头草。B, 芦苇。
Fig. 7 Micromorphological and structural characteristics of leaf epidermis of desert plants. A, Sympegma regelii . B, Phragmites communis .
(8)瘤状 表皮无绒毛或有少量绒毛, 表皮细胞突起的高度大于或等于宽度, 顶部半圆形(图8 )。代表植物有小獐毛(Aeluropus littoralis )(图8 A)、罗布麻(Apocynum venetum )(图8 B)等。
图8 荒漠植物叶片表皮微形态结构特征。A, 小獐毛。B, 罗布麻。
Fig. 8 Micromorphological and structural characteristics of leaf epidermis of desert plants. A, Aeluropus littoralis . B, Apocynum venetum .
(9)光滑型 荒漠植物中为数不多的植物叶片表面相对比较光滑, 气孔密度较高, 气孔深陷或不下陷, 没有其他附属物。代表植物有沙葱(Allium mongolicum )(图9A )、沙木蓼(Atraphaxis bracteata )、骆驼蓬属(Peganum )植物如多裂骆驼蓬(Peganum multisectum )(图9C )、牛心朴子(Cynanchum komarovii )(图9B )、乌丹蒿(Artemisia wudanica )(图9D )、籽蒿(Artemisia sphaerocephala )、瓦松(Orostachys fimbriatus )等。
图9 荒漠植物叶片(或同化枝)表皮微形态结构特征。A, 沙葱。B, 牛心朴子。C, 多裂骆驼蓬。D, 乌丹蒿。
Fig. 9 Micromorphological and structural characteristics of leaf (or assimilation branches) epidermis of desert plants. A, Allium mongolicum . B, Cynanchum. komarovii . C, Peganum multisectum . D, Artemisia wudanica .
(10)褶皱状 叶片表面形成凸凹不平、大小不等、形状不同的皱褶, 气孔器穴居于皱褶凹陷处, 表皮无绒毛附属物。代表植物有短叶假木贼(Anabasis brevifolia )、梭梭(Haloxylon ammodendron )(图10A )、木本猪毛菜(Salsola arbuscula )、猪毛菜(Salsola collina )(图10B )、黑果枸杞(Lycium ruthenicum )、红砂属(Reaumuria )(图10C )和柽柳属(Tamarix )(图10D )的植物等。
图10 荒漠植物叶片(或同化枝)表皮微形态结构特征。A, 梭梭。B, 猪毛菜。C, 红砂。D, 柽柳。
Fig. 10 Micromorphological and structural characteristics of leaf (or assimilation branches) epidermis of desert plants. A, Haloxylon ammodendron . B, Salsola collina . C, Reaumuria soongorica . D, Tamarix chinensis .
(11)多种附属物混生状 多数片状叶片的荒漠植物叶表皮都有绒毛、蜡质晶体等多种附属物混生, 但叶片表皮细胞形态清晰可见。代表植物有雾冰藜(Bassia dasyphylla )(图11A )、沙米(Agriophyllum squ- arrosum )(图11B )、骆驼刺(Alhagi sparsifolia )(图11C )、兴安胡枝子(Lespedeza davurica )、部分锦鸡儿属(Caragana )植物如荒漠锦鸡儿(Caragana robor- ovskyi )(图11D )、岩黄芪属(Hedysarum )植物、油蒿(Artemisia ordosica )、差巴嘎蒿(Artemisia halodendron )、绵刺(Potaninia mongolica )、猫头刺(Oxytropis aciphylla )(图11E )、半日花(Helianthemum songaricum )(图11F )、花花柴(Karelinia caspia )(图11G )、中亚紫菀木(Asterothamnus centrali-asiaticus )(图11 H)等。
图11 荒漠植物叶片表皮微形态结构特征。A, 雾冰藜。B, 沙米。C, 骆驼刺。D, 荒漠锦鸡儿。E, 猫头刺。F, 半日花。G, 花花柴。H, 中亚紫菀木。
Fig. 11 Micromorphological and structural characteristics of leaf epidermis of desert plants. A, Bassia dasyphylla . B, Agriophyllum squarrosum . C, Alhagi sparsifolia . D, Caragana roborovskyi . E, Oxytropis aciphylla . F, Helianthemum songaricum . G, Karelinia caspia . H, Asterothamnus centrali-asiaticus .
2.3 荒漠植物叶片(或同化枝)表皮微形态抗逆适应主要类型
植物叶片形态结构与抗逆性关系的研究表明, 植物叶片小、表皮附属物密度大、且表皮皱缩程度高, 则抗旱性强(王勋陵和马骥, 1999 ; 周智斌和李培军, 2002 ; Bargel et al ., 2004 ; 迟丽华和宋凤斌, 2006 ; Adebowale et al ., 2014 ; 赵小仙等, 2014 ; 朱广龙等, 2016 )。通过比较荒漠植物叶片(或同化枝)的表皮微形态结构, 可以从表皮细胞形态, 表皮附属物如绒毛、蜡质、气孔等的形态、密度、结构和分布特征来进行环境适应性分析。根据表皮微形态结构和附属物与抗逆性的关系, 将荒漠植物适应环境胁迫的表皮微形态分为以下6种主要类型。
(1)完全依赖于表皮毛 形态各异的表皮绒毛完全覆盖叶片上下表面, 阻止水分蒸腾和反射强光, 同时具有吸附空气中的水汽的功能。代表性植物为表皮微形态结构基本类型(2)中的沙枣、柠条锦鸡儿、珍珠猪毛菜、蒙古莸、蓍状亚菊等。
(2)完全依赖于角质层蜡质 不同形态的蜡质层完全覆盖叶片表面, 表面粗糙, 阻止水分蒸腾和反射强光。代表性植物为表皮微形态结构基本类型(1)中的膜果麻黄、霸王、灌木铁线莲等。
(3)依赖于表皮凹凸结构 表皮细胞堆积呈凹凸状, 气孔深陷凹槽内, 气孔指数很小, 阻止蒸腾。代表性植物为表皮微形态结构基本类型(10)中的红砂(Reaumuria soongorica )、柽柳(Tamarix chinensis )、叶假木贼等。
(4)依赖于表面突起 表面有大量的形态各异的凸起, 增加叶表面积, 气孔密度相对较低, 减少蒸腾, 增强吸收水汽的功能。代表性植物为表皮微形态结构基本类型(7)、(8)中的罗布麻、小獐毛、芦苇等。
(5)依赖于表皮混生附属物 各种表皮附属物混生, 多数植物表皮绒毛不完全覆盖表面, 有明显的角质膜蜡质片层结构, 气孔密度较高, 增强了各种附属物协调抵御逆境的能力。代表性植物为表皮微形态结构基本类型(4)、(5)、(11)中的沙米、骆驼刺等。
(6)上表皮异化 上表面气孔指数为0, 减少气孔性水分散失, 并增强对强光的反射。代表性植物为表皮微形态结构基本类型(6)中的蒙古扁桃、柄扁桃、杠柳等。
这6种主要适应类型植物的抗旱性大小需根据各自植物微形态结构对抗旱性的贡献进行统计分析。
3 讨论
荒漠植物由于长期适应所生长的自然环境, 发展了一套最适宜自身生长发育的形态结构特征, 并采取各种途径来抵御或忍耐干旱胁迫对它的影响, 因而具有一定程度的耐旱性。这种耐旱性是一种复合性状, 包括植物的形态结构特征、水分生理特征、生理生化反应、组织细胞、光合器官以及原生质结构等特征的综合反映(Klich, 2000 ; Chaves et al ., 2003 ; Ogle & Reynolds, 2004 )。不同的荒漠植物对水分和不利环境的适应和耐受程度不同, 适应方式多种多样, 按照不同的分类标准, 荒漠植物有多种适应类型。通常的分类标准包括耐旱程度、耐旱方式、叶片形态、生活史、光合碳代谢途径等(赵哈林, 2012 )。各种类型的植物又有各自的适应特征。本研究是针对我国干旱半干旱区荒漠植物叶表皮微形态结构特征的综合分析, 主要集中于该区域的建群种灌木物种。根据荒漠植物叶表皮微形态基本结构特征和表皮附属物的类型与数量, 将荒漠植物表皮微形态类型直观地划分为11种, 而将抗逆适应微形态结构划分为6种类型。荒漠植物叶表皮微形态结构类型复杂, 反映了植物自身的遗传特性及其与环境的密切关系。植物叶片表皮微形态主要由遗传因素决定, 但表皮附属物绒毛数量, 蜡质形态与厚度, 气孔的形态、密度以及分布特征等与环境因子密切相关, 是植物对环境的响应(Barthlott, 1981 ; Casson & Hetherington, 2010 ; Ghahremaninejad et al ., 2012 )。因此, 基于这些表皮主要附属物特征对荒漠植物的表皮微形态类型和抗逆适应类型进行划分, 能够从微观形态结构特征进一步揭示物种的亲缘关系和表皮微形态结构的生态适应意义。
3.1 荒漠植物叶表皮微形态结构的多样性及其亲缘关系
荒漠植物叶表皮微形态结构在不同分类等级上表现出丰富的多样性, 深刻地反映了植物的亲缘关系, 不同程度地揭示了它们之间错综复杂的系统演化关系。叶片表皮微形态结构基本特征根据表皮细胞形态结构和表皮附属物类型, 分为疣状、瘤状、皱褶状、相对光滑型、表面完全被毛、表面完全被蜡质层、不同形态结构的绒毛和蜡质层混合覆盖、上下完全异面以及气孔器集中于下表面等11种类型, 有些类型还包括若干种亚型。如柳属中观察的4种植物, 尽管种及地理分布完全不同, 但它们都为典型的异面叶, 叶片上下表皮都是完全不同的形态。上表皮绒毛密度较高, 不同的种有不同形态和密度的绒毛; 表皮绒毛密度低或不具绒毛, 但有柳属植物特有的角质膜蜡质纹饰(王东超等, 2012 ; Ghahremaninejad et al ., 2012 ), 如黄柳的痂状蜡质层、旱柳和乌柳的壳状蜡质层、沙柳的膜片状蜡质晶体。麻黄属的膜果麻黄、斑子麻黄(Ephedra rhytidosperma )和草麻黄(Ephedra sinica )中, 3种同化枝的角质膜蜡质晶体数量依次减少, 斑子麻黄与草麻黄的表皮形态结构接近, 显示亲缘关系较近。膜果麻黄表皮粗糙, 完全被蜡质层覆盖。锦鸡儿属(Caragana )是豆科蝶形花亚科中较大的一个属, 叶片上下表皮微形态基本一致, 多数为等面叶。但表皮毛的形态密度各异, 柠条锦鸡儿、小叶锦鸡儿(Caragana microphlla )和中间锦鸡儿(Caragana intermedia )同属小叶系, 亲缘关系相近, 表皮毛为柳叶状, 但绒毛密度不同, 柠条锦鸡儿表皮毛完全覆盖叶面, 小叶锦鸡儿和中间锦鸡儿表皮毛密度较小, 且它们的鳞片状蜡质晶体密度相似, 显示他们之间更近的亲缘关系, 这与其他分子生物学依据得到的结果不完全一致(Guo et al ., 2008 )。究其原因, 主要是因为这3种锦鸡儿属植物之间由于分布区域重叠或交叉, 因此存在复杂的基因交流(Ma et al ., 2003 ), 不同种的亲缘关系与其分布区域的其他种之间的相互杂交有关, 甚至出现了杂交过渡态类型, 因而给分类带来了难题, 单纯某一种分类依据是不能完全决定其亲缘关系的。荒漠锦鸡儿(Caragana roborovskyi )和卷叶锦鸡儿(Caragana ordosica )绒毛为棒状, 表皮细胞和蜡质层形态亦相似。短脚锦鸡儿(Caragana brachypoda )和狭叶锦鸡儿(Caragana stenophylla )表皮毛为柳叶带乳突型, 上表皮气孔器深陷, 气孔器为不等型或不规则型, 气孔口三角形或多边形。红花锦鸡儿(Caragana rosea )为明显不同的类群, 表皮无绒毛结构, 且蜡质层形态不同于以上几种的鳞片状, 而是紧贴表皮的膜片层结构或颗粒状。
同一种植物可以是单一类型的表皮形态结构, 也可以是复合类型的, 也可能是过渡态的。在同一个属中可以有一种或数种不同的叶表皮形态结构, 它们之间的形态可能是连续的、渐变的, 也可能是间断的、区分明显的, 但无一不反映出它们之间存在的亲缘关系, 揭示它们之间极其复杂的演化关系。如猪毛菜属(Salsola )植物, 从已观测的4种植物表皮形态结构来看, 珍珠猪毛菜表皮完全被柳叶状毛, 木本猪毛菜、松叶猪毛菜(Salsola laricifolia )与猪毛菜表皮无毛, 但表皮细胞形态各异, 猪毛菜的表皮细胞皱褶状, 气孔器下陷, 松叶猪毛菜表皮细胞皱褶程度低且气孔不下陷。蒿属(Artermisia )是菊科的一个大属, 存在着不少分类上的争议, 从已观测的6种来看, 叶表皮形态结构各自不同, 同属龙蒿亚属的籽蒿、油蒿和差巴嘎蒿的叶表皮结构为渐变式的, 籽蒿表皮基本没有表皮毛, 油蒿有少量表皮毛, 差巴嘎蒿有高密度的表皮毛, 气孔器都不下陷, 但油蒿和籽蒿的表皮细胞形态和气孔器结构相同, 显示了更近的亲缘关系。乌丹蒿表皮有少数绒毛, 与油蒿和籽蒿接近, 但表皮细胞形态不同。白沙蒿(Artermisia blepharolepis )表皮细胞形态和气孔器与籽蒿相同, 但表皮有大量膜片状和颗粒状蜡质晶体, 表面不光滑。而白莲蒿表皮除了卷丝状毛外, 还有囊状毛, 为另一种形态结构类型。
另外, 按照表皮细胞形态和表皮附属物绒毛、蜡质晶体的特征进行基本形态结构的分类, 多种属于同一种类型的叶表皮微形态结构特征的植物, 它们出现在不同的属甚至亲缘关系相距甚远的科中。如蒙古扁桃和杠柳分别属于蔷薇科和萝藦科, 但两者叶表皮微形态结构属于同种类型, 上表皮无气孔器, 下表皮有高密度的气孔器; 唐古特白刺和柠条锦鸡儿的表皮完全被柳叶状绒毛等。不同科属的植物出现相近的表皮微形态结构, 与它们在相同生境中的进化过程有关, 但它们在表皮细胞形态、气孔器结构与类型、蜡质层结构等其他微形态结构特征方面还有明显差异。
植物叶片表皮微形态特征的多样性不仅在很大程度上表达了遗传的多样性和稳定性, 而且具有重要的系统学和分类学意义, 有助于一些分类问题的解决。近年来不少研究发现叶表皮性状在一定程度上能反映分类群间的系统发生, 可用于科下属间关系的探讨(Kong et al ., 2001 )。以骆驼蓬属为例, 1889年Maxim将多裂骆驼蓬定为骆驼蓬(Peganum harmala )的一个变种Peganum harmala var. multisectum , 1949年Bobr却将其定为独立的种(中国科学院中国植物志编辑委员会, 1998),《中国沙漠植物志》亦将之作为种(刘瑛心, 1987 ), 但国内外一些学者仍倾向于将其作为骆驼蓬的一个变种。马骥等(1997, 2003)结合种子微形态结构和有关化学成分、地理分布等方面的研究结果, 支持将多裂骆驼蓬作为一个独立的种处理的观点。叶片表皮微形态实验结果表明: 从粗略的叶表皮微形态基本特征统计来看, 二者基本特征相同, 但骆驼蓬表皮细胞呈褶皱状, 而多裂骆驼蓬叶表皮基本光滑, 有明显的区别, 赞成将二者作为不同的独立种的观点。
综上所述, 荒漠植物叶表皮微形态结构既连续又间断, 既有过渡又有交叉, 在不同分类等级上表现出丰富的多样性, 为植物分类科学的进展及植物进化提供了丰富的信息资料。
3.2 荒漠植物叶表皮附属物的生理功能与生态环境的联系
大多数荒漠植物叶片的一个显著特点是表皮被绒毛, 叶表皮毛的出现与长期的干旱气候有关, 是植物对干旱环境的适应特征(Retamales & Scharaschkin, 2015 )。植物叶片表皮毛分两种: 腺毛和非腺毛(Werker, 1993 ; Navarro & Oualidi, 1999 )。非腺毛的主要功能与植物的防御有关。目前确认的植物叶片表皮非腺毛具有的生理生态功能主要有: 减轻紫外辐射(Karabourniotis et al ., 1992 ; Gregoriou et al ., 2007 )、降低边界层的传导(Wuenscher, 1970 ; Kenzo et al ., 2008 )、降低叶温(Ehleringer & Mooney, 1978 ), 以及减少对太阳辐射的接收(Ehleringer et al ., 1976 )等。这些功能决定了干旱区荒漠植物抗逆能力的大小。干旱地区的叶表皮毛密度比湿润地区的高(Smith & Nobel, 1977 ; Aronne & de Micco, 2001 ), 就单个物种而言, 表皮毛密度与温度和辐射密切相关(Perez-Estrada et al ., 2000 )。在热带雨林树种中, 浓密的表皮毛很少见(Turner, 2001 ), Roth (1984)调查的250种树中仅有15种有较厚的表皮毛, 且较厚的表皮毛一般在干旱、高温和强辐射条件下产生。而在干旱半干旱的荒漠区, 大多数植物都具有浓密的表皮毛。显然, 荒漠植物叶上密生的表皮毛对于植物的生活有重要的意义。从Ehleringer等 (1976)、Ehleringer和Bjorkman (1978)、Ehleringer和Mooney (1978)研究的荒漠灌木Encelia farinosa 的叶表皮毛对叶片光谱特性和光合、蒸腾等生理活动的影响来看, 沙漠毒菊在不同生长季生长的叶片具有不同数量的表皮毛, 表皮毛的变化足够使叶片对光合有效辐射的吸收从81%降到29%, 对总的太阳辐射的吸收从46%降到16%, 而亲缘关系较近却没有表皮毛的同属物种Encelia californiea 的有效光合辐射吸收率为84%。同时, 由于对太阳辐射吸收的减少, 导致叶面温度和植物蒸腾速率的降低。荒漠环境气温经常超过40 ℃, 而植物最适宜的光合温度是25 ℃, 当温度超过35 ℃时植物的光合速率会突降, 但较厚的叶表皮毛能使叶温显著低于气温而接近光合适宜温度, 同时也避免植物达到叶片致死温度。但叶表皮毛也有不利的一面, 表皮毛的覆盖降低了光合速率, 因此同一植物在不同的生境和生长季长出的叶片会出现表皮毛数量不同的特征, 这也是植物适应环境的一种权衡策略(Kenzo et al ., 2008 )。从本研究中的117种荒漠植物叶表皮特征来看, 片状叶片基本被有表皮毛, 并有不少叶片的表皮毛数量较多, 直接增加了叶片的边界层厚度, 个别植物的上下表皮毛厚度远超于叶片厚度, 如表皮微形态抗逆适应主要类型的第一类植物中的柠条锦鸡儿、卷叶锦鸡儿和沙枣等。可见荒漠植物中叶片表皮毛的作用是相当重要的, 它们不仅能调节植物的生理功能, 而且可抵御风沙等机械损伤的直接伤害。这些物种在不同的自然生境中其表皮毛数量亦不同(数据省略), 说明生态环境深刻地影响着植物形态结构的形成与变异。
而对于腺毛来说, 其主要作为一个油性物质的合成、分泌和累积的器官, 有效地防御动物的捕食、病菌入侵以及物种竞争(Werker, 1993 ; Wagner et al ., 2003 ; Kremer et al ., 2014 ; Munien et al ., 2015 )。实际上, 腺毛也是次生代谢产物的一个独立的细胞工厂(Tissier, 2012 )。腺毛的形态随植物的组织器官和种类不同而呈多样化, 其形成和分泌的主要油性物质及其产物组成受遗传和环境两方面的因素控制(Bosabalidis & Kofidis, 2002 ; Shafiee-Hajiabad et al ., 2014 ), 腺毛结构的多样性也可以为植物分类学提供依据(Gairola et al ., 2009 ; Rusydi et al ., 2013 )。对117种荒漠植物叶片表皮扫描发现: 不少旱生植物除具有非腺体表皮毛外, 还同时具有腺毛, 并且有些腺毛体积较大, 如蒙古莸、甘草、香青兰、星毛短舌菊、百里香等。显然, 对于具有腺毛的荒漠植物, 腺毛具有重要的功能。但目前关于不同植物腺毛的形态、形成以及功能的研究还很有局限性。那么, 这些腺毛在植物适应荒漠环境的过程中其形态结构与环境因子有何关系、主要分泌哪些化学物质、发挥什么样的功能?这些问题亟待下一步的深入研究。
荒漠植物叶片的另一个显著特点是表皮有较厚的角质膜蜡质层。角质膜蜡质层与表皮毛有共同的功能, 是植物表皮的保护屏障, 主要由长链碳氢化合物组成(Shepherd & Wynne, 2006 )。叶片对光的反射由叶面形貌决定, 表皮毛和蜡质层是决定荒漠植物叶表面形貌的关键。Holmes和Keiller (2002)研究了45种桉属和伽蓝菜属植物的蜡质叶, 发现蜡质层对紫外光(330 nm)和光合有效辐射(680 nm)的反射比各种形态的表皮毛更强, 去除蜡质层通常会减少对这两种光的反射, 特别是对紫外光的反射, 但对表皮无蜡质的叶片影响不大。同时不同的物种对不同波长的光的反射亦不同, 表明蜡质晶体的形态直接影响着对光的反射, 并且不同温度下的反射强度也不同, 可见蜡质层对光的反射特性变化很大(Holmes & Keiller, 2002 )。叶表皮蜡质层参与植物对水分、盐、低温、污染、机械损伤等多种胁迫响应的过程, 提高植物对胁迫的抗性(Cameron, 1970 ; Bargel et al ., 2004 ; Perrone et al ., 2013 , 2015 )。因此, 本研究中117种荒漠植物叶片表皮颗粒状、膜片状、晶簇状、鳞片状、杆状等各种不同形态的蜡质晶体既是植物长期适应环境的结果, 也对荒漠植物在严酷环境中的生长和生存至关重要。总的来说, 这些植物中灌木的蜡质层密度高于草本, 无表皮毛的叶片蜡质层密度高于表皮毛覆盖的, 特别是以同化枝为主要光合器官和棒状叶片的植物, 表皮基本没有表皮毛覆盖, 主要的附属物是蜡质层, 如麻黄属、沙拐枣属、红砂属的植物等, 弥补了表皮毛的功能。
另外, 表面乳突或表皮皱褶, 一方面增加了叶表面积, 另一方面使气孔器能够穴居于凹陷处而避免强光的直射(Adebowale et al ., 2014 )。选取的117种荒漠植物的气孔密度主要集中于100-200个·mm-2 , 个别植物的气孔密度相当低, 如红砂属、柽柳属植物、沙枣、沙棘(Hippophae rhamnoides )等, 并且多数灌木的气孔器下陷, 降低了水分的气孔性散失。同时, 从结果可以看出气孔密度的高低与表皮其他附属物也有关系, 表皮毛和蜡质少的植物, 气孔密度一般相对较高, 如表皮微形态抗逆适应主要类型的第(5)、(6)类植物中的骆驼刺、杠柳等。表皮附属物的基本功能是保护叶片免受不利环境的直接损伤, 从而调节植物的生理功能。
综上所述, 荒漠植物对环境的适应, 从表皮微形态结构基本特征来看, 以同化枝为主要光合器官的植物, 同化枝表皮主要的附属物是角质膜蜡质层以及气孔器, 基本没有表皮绒毛, 如梭梭属、沙拐枣属、麻黄属的植物等。肉质棒状叶片表皮微形态结构与同化枝一样也不具有绒毛结构, 如黑果枸杞、红砂属植物、霸王、盐爪爪、沙葱等。同化枝和棒状叶片表皮附属物蜡质层晶体形态各异, 蜡质层厚度与植物的抗旱性密切相关。不同生活型植物中, 草本的蜡质片层相对较少, 多数灌木都具有较厚的角质膜蜡质层。表皮附属物绒毛结构在多数片状叶片中基本都可以见到, 绒毛的形态结构多样化程度很高, 基本结构分为有囊和无囊两类, 形态各异。绒毛在表皮的分布密度从零星分布到完全覆盖的都有, 多数密度较高。对于大多数荒漠植物而言, 绒毛密度低, 则蜡质层晶体密度高, 绒毛和蜡质晶体相互协调覆盖叶片表皮。表皮绒毛和蜡质晶体覆盖少的植物, 则气孔密度相对较高, 气孔密度最高的荒漠植物叶片见于上表面无气孔器的, 如蒙古扁桃、杠柳等。灌木的气孔下陷较深, 以降低水分的气孔性散失, 草本植物的气孔基本不深陷。从荒漠植物叶片表皮的主要功能来看, 荒漠植物叶表皮微形态结构的适应特征是表皮附属物(主要是绒毛和角质膜蜡质层)与表皮结构(凹凸、乳突和气孔器等)相互协调, 共同抵御强光, 降低叶片的蒸腾并吸收空气中的水汽来适应干旱和其他不利的荒漠环境条件。
附录 Supplementary Material
附录I 荒漠植物叶片采集的物种种类、采集地理位置、植物生活型及采集时间
Appendix I List of the species, collection sites, life form and selected time of desert plants in this study
http://www.plant-ecology.com/fileup/PDF/cjpe.2016.0129-A1.PDF
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Adebowale A Naidoo Y Lamb J Nicholas A 2014 ). Comparative foliar epidermal micromorphology of Southern African Strychnos L.(Loganiaceae): Taxonomic, ecological and cytological considerations
. Plant Systematics and Evolution , 300 , 127 -138 .
[本文引用: 4]
[2]
Aliero AA Grierson DS Afolayan AJ 2006 ). The foliar micromorphology of Solanum pseudocapsicum
. Flora , 201 , 326 -330 .
[本文引用: 1]
[3]
Arena C Vitale L de Santo VA 2008 ). Paraheliotropism in Robinia pseudoacacia L.: An efficient strategy to optimise photosynthetic performance under natural environmental conditions
. Plant Biology , 10 , 194 -201 .
[本文引用: 1]
[4]
Aronne G de Micco V 2001 ). Seasonal dimorphism in the Mediterranean Cistus incanus L. subsp. incanus
. Annal Botany , 87 , 789 -794 .
[本文引用: 2]
[5]
Bargel H Barthlott W Koch K Schreiber L Neinhuis C 2004 ). Plant cuticles: Multifunctional interfaces between plant and environment. In: Hemsley AR, Poole I eds. Evolution of Plant Physiology
. Academic Press , London . 171 -194 .
[本文引用: 3]
[6]
Barthlott W 1981 ). Epidermal and seed surface characters of plants: Systematic applicability and some evolutionary aspects
.Nordic Journal of Botany , 1 , 345 -355 .
[本文引用: 1]
[7]
Bosabalidis AM Kofidis G 2002 ). Comparative effects of drought stress on leaf anatomy of two olive cultivars
.Plant Science , 163 , 375 -379 .
[本文引用: 2]
[8]
Cameron RJ 1970 ). Light intensity and the growth of Eucalyptus seedlings II. The effect of cuticular waxes on light absorption in leaves of Eucalyptus species
. Australia Journal of Botany , 18 , 275 -284 .
[本文引用: 1]
[9]
Casson SA Hetherington AM 2010 ). Environmental regulation of stomatal development
.Current Opinion of Plant Biology , 13 , 90 -95 .
[本文引用: 1]
[10]
Chaves MM Maroco JP Pereira JS 2003 ). Understanding plant responses to drought—From genes to the whole plant
.Functional Plant Biology , 30 , 239 -264 .
[本文引用: 1]
[11]
Chen G Sun W Sun H 2010 ). Leaf epidermal characteristics of Asiatic buddleja L. under scanning electron microscope: Insights into chromosomal and taxonomic significance
. Flora , 205 , 777 -785 .
[本文引用: 1]
[12]
Chi LH Song FB 2006 ). Analyse of leaves’ structure character and ecological adaptation of ten plants in the west alkaline district of Songnen plain
.Ecology and Environment , 15 , 1269 -1273 . (in Chinese with English abstract)[迟丽华 , 宋凤斌 (2006 ). 松嫩平原西部盐碱地区10种植物叶片结构特征及其生态适应性
. 生态环境 , 15 , 1269 -1273 .]
[本文引用: 1]
[13]
de Micco V Aronne G 2012 ). Morpho-anatomical traits for plant adaptation to drought. In: Aroca R ed. Plant Responses to Drought Stress. Springer, Berlin.
[本文引用: 1]
[14]
Deng M Hipp A Song Y Li Q Coombes A Cotton A 2014 ). Leaf epidermal features of Quercus subgenus Cyclo- balanopsis(Fagaceae) and their systematic significance
. Botanical Journal of the Linnean Society , 176 , 224 -259 .
[本文引用: 1]
[15]
Dickison WC 2000 ). Integrative Plant Anatomy . Academic Press , San Diego, USA .
[本文引用: 1]
[16]
Editorial Committee of Flora of China , Chinese Academy of Sciences 1998 ). Flora of China . Science Press , Beijing . (in Chinese)[中国科学院中国植物志编辑委员会 (1998 ). 中国植物志 . 科学出版社 , 北京 .]
[17]
Ehleringer J Bjorkman O 1978 ). Pubescence and leaf spectral characteristics in a desert shrub,Encelia farinosa
. Oecologia , 36 , 137 -148 .
[18]
Ehleringer J Bjorkman O Mooney HA 1976 ). Leaf pubescence: Effects on absorptance and photosynthesis in a desert shrub
.Science , 192 , 376 -377 .
[本文引用: 1]
[19]
Ehleringer JR Mooney HA 1978 ). Leaf hairs: Effects on physiological activity and adaptive value to a desert shrub
.Oecologia , 37 , 183 -200 .
[本文引用: 1]
[20]
Francisco A Porto M Ascensao L 2015 ). Morphological phylogenetic analysis of Ophrys(Orchidaceae): Insights from morpho-anatomical floral traits into the interspecific relationships in an unresolved clade
. Botanical Journal of the Linnean Society , 179 , 454 -476 .
[本文引用: 1]
[21]
Gairola S Naidoo Y Bhatt A Nicholas A 2009 ). An investigation of the foliar trichomes of Tetradenia riparia(Hochst.) Codd [Lamiaceae]: An important medicinal plant of Southern Africa
. Flora , 204 , 325 -330 .
[本文引用: 2]
[22]
Gan Y Zhou L Shen ZJ Shen ZX Zhang YQ Wang GX 2010 ). Stomatal clustering, a new marker for environmental perception and adaptation in terrestrial plants
.Botanical Studies , 51 , 325 -336 .
[本文引用: 1]
[23]
Gao JP Wang YH Chen DF 2003 ). Anatomical characteristics of leaf epidermis and vessel elements of Schisandra sphenanthera from different districts and their relationships to environmental factors
. Acta Botanica Boreali-Occidentalia Sinica , 23 , 715 -723 . (in Chinese with English abstract)[高建平 , 王彦涵 , 陈道峰 (2003 ). 不同产地华中五味子叶表皮结构和导管分子的解剖学特征及其与环境因子的关系
. 西北植物学报 , 23 , 715 -723 .]
[本文引用: 1]
[24]
Ghahremaninejad F Khalili Z Maassoumi AA Mirzaie- Nodoushan H Riahi M 2012 ). Leaf epidermal features ofSalix species (Salicaceae) and their systematic significance
. American Journal of Botany , 99 , 769 -777 .
[本文引用: 2]
[25]
Gielwanowska I Szczuka E Bednara J Gorecki R 2005 ). Anatomical features and ultrastructure of Deschampsia antarctica(Poaceae) leaves from different growing habitats
. Annals of Botany , 96 , 1109 -1119 .
[本文引用: 1]
[26]
Gomes MP Marques TCLLSM Nogueira MOG de Castro EM Soares AM 2011 ). Ecophysiological and anatomical changes due to uptake and accumulation of heavy metal in Brachiaria decumbens
. Scientific Agriculture , 68 , 566 -573 .
[本文引用: 1]
[27]
Gregoriou K Pontikis K Vemmos S 2007 ). Effects of reduced irradiance on leaf morphology, photosynthetic capacity, and fruit yield in olive (Olea europaea L.)
. Photosynthetica , 45 , 172 -181 .
[本文引用: 1]
[28]
Guo H Gao Y Ma C Ren A Wu J 2008 ). Genetic diversity and genetic relationship of Caragana microphylla, Caragana davazamcii and Caragana korshinskii on the Inner Mongolia Plateau, China
. Acta Ecologica Sinica , 28 , 3729 -3736 .
[本文引用: 1]
[29]
He JS Chen WL Wang XL 1994 ). Morphological and anatomical features of Quercus section Suber and its adaptation to the ecological environment
. Acta Phytoecologica Sinica , 18 , 219 -227 . (in Chinese with English abstract)[贺金生 , 陈伟烈 , 王勋陵 (1994 ). 高山栎叶的形态结构及其与生态环境的关系
. 植物生态学报 , 18 , 219 -227 .]
[本文引用: 1]
[30]
Hickey LJ 1984 ).Changes in the angiosperm flora across the cretaceus tertiary boundary
. In: Bergren WA, van Couvering JA eds. Catastrophes and Earth History. Princeton University Press, Princeton, USA.
[本文引用: 1]
[31]
Holmes MG Keiller DR 2002 ). Effects of pubescence and waxes on the reflectance of leaves in the ultraviolet and photosynthetic wavebands: A comparison of a range of species
.Plant, Cell & Environment , 25 , 85 -93 .
[本文引用: 1]
[32]
Jaleel CA Gopi R Azooz MM Panneerselvam R 2009 a). Leaf anatomical modifications in Catharanthus roseus as affected by plant growth promoters and retardants
.Global Journal of Molecular Science , 4 , 1 -5 .
[本文引用: 1]
[33]
Jaleel CA Manivannan P Wahid A Farooq M Jasim Al-Juburi H Somasundaram R Panneerselvam R 2009 b). Drought stress in plants: A review on morphological characteristics and pigments composition
.International Journal of Agriculture and Biology , 11 , 100 -105 .
[本文引用: 1]
[34]
Karabourniotis G Papadopoulos K Papamarkou M Manetas Y 1992 ). Ultraviolet-B radiation absorbing capacity of leaf hairs
.Physiologia Plantarum , 86 , 414 -418 .
[本文引用: 1]
[35]
Kenzo T Yoneda R Azani MA Majid NM 2008 ). Changes in leaf water use after removal of leaf lower surface hairs onMallotus macrostachyus(Euphorbiaceae) in a tropical secondary forest in Malaysia
. Journal of Forest Research , 13 , 137 -142 .
[本文引用: 2]
[36]
Klich MG 2000 ). Leaf variations in Elaeagnus angustifolia related to environmental heterogeneity
. Environmental and Experimental Botany , 44 , 171 -183 .
[本文引用: 1]
[37]
Koch K Bhushan B Barthlott W 2009 ). Multifunctional surface structures of plants: An inspiration for biomimetics
.Progress of Material Science , 54 , 137 -178 .
[本文引用: 2]
[38]
Koch K Ensikat HJ 2008 ). The hydrophobic coatings of plant surfaces: Epicuticular wax crystals and their morphologies, crystallinity and molecular self-assembly
.Micron , 39 , 759 -772 .
[本文引用: 1]
[39]
Kong HZ 2001 ). Comparative morphology of leaf epidermis in the Chloranthaceae
.Botanical Journal of Linnean Society , 136 , 279 -294 .
[本文引用: 1]
[40]
Kremer D Dunkic V Ruscic M Matevski V Ballian D Bogunic F Eleftheriadou E Stesevic D Kosalec I Bezic N Staben- theiner E 2014 ). Micromorphological traits and essential oil contents of Micromeria kerneri Murb. and M. juliana(L.) Benth. 98 , 128 -136 .
[本文引用: 1]
[41]
Krober W Heklau H Bruelheide H 2015 ). Leaf morphology of 40 evergreen and deciduous broad leaved subtropical tree species and relationships to functional ecophysiological traits
.Plant Biology , 17 , 373 -383 .
[本文引用: 1]
[42]
Lakusic B Stevanovic B Jancic R Lakusic D 2010 ). Habitat- related adaptations in morphology and anatomy of Teucrium(Lamiaceae) species from the Balkan Peninsula (Serbia and Montenegro)
. Flora , 205 , 633 -646 .
[本文引用: 1]
[43]
Liu Y Li X Chen G Li M Liu M Liu D 2015 ). Epidermal micromorphology and mesophyll structure of Populus euphratica heteromorphic leaves at different development stages
. PLOS ONE , 10 , e0137701 . doi:10.1371/journal.pone.0137701 .
[本文引用: 1]
[44]
Liu Y Li X Liu M Cao B Tan H Wang J Li X 2012 ). Responses of three different ecotypes of reed (Phragmites communis Trin.) to their natural habitats: Leaf surface micro-morphology, anatomy, chloroplast ultrastructure and physio-chemical characteristics
. Plant Physiology and Biochemistry , 51 , 159 -167 .
[本文引用: 2]
[45]
Liu YX 1987 ). Desert Flora of China, Vol. 2 . Science Press , Beijing . (in Chinese)[刘瑛心 (1987 ). 中国沙漠植物志, 第2卷 . 科学出版社, 北京 .]
[本文引用: 1]
[46]
Ma CC Gao YB Guo HY Wang JL 2003 ). Interspecific transition among Caragana microphylla, C. davazamcii and C. korshinskii along geographic gradient I. Ecological and RAPD evidence
. Acta Botany Sinica , 45 , 1218 -1227 .
[本文引用: 1]
[47]
Ma J Li JZ Chao Z Zhang HW Liu CM 2003 ). A study on microstructural feature of the seeds of desert plants in northwest China
.Journal of Zhejiang Normal University (Natural Science Edition) , 26 , 109 -115 . (in Chinese with English abstract)[马骥 , 李俊祯 , 晁志 , 张宏伟 , 刘传明 (2003 ). 64种荒漠植物种子微形态的研究
. 浙江师范大学学报(自然科学版) , 26 , 109 -115 .]
[48]
Ma J Wang XL Zhao SL 1997 ). A study on the seeds micro- morphological characteristics ofPeganum from China northwest and its taxonomic and ecological significance
. Journal of Wuhan Botanical Research , 4 , 323 -327 . (in Chinese with English abstract)[马骥 , 王勋陵 , 赵松岭 (1997 ). 骆驼蓬属种子微形态及其生态学与分类学意义
. 武汉植物学研究 , 4 , 323 -327 .]
[49]
Munien P Naidoo Y Naidoo G 2015 ). Micromorphology, histochemistry and ultrastructure of the foliar trichomes of Withania somnifera(L.) Dunal (Solanaceae)
. Planta , 242 , 1107 -1122 .
[本文引用: 1]
[50]
Navarro T Oualidi JE 1999 ). Trichome morphology in Teucrium L.(Labiatae). A taxonomic review
. Anales del Jardin Botanico de Madrid , 57 , 277 -297 .
[本文引用: 1]
[51]
Noman A Hameed M Ali Q Aqeel M 2012 ). Foliar tissue architectural diversity among three species of genus Hibiscus for better adaptability under industrial environment
. International Journal of Environmental Science , 2 , 2212 -2222 .
[本文引用: 1]
[52]
Ogle K Reynolds JF 2004 ). Plant responses to precipitation in desert ecosystems: Integrating functional types, pulses, thresholds, and delays
.Oecologia , 141 , 282 -294 .
[本文引用: 1]
[53]
Pal A Kulshreshtha K Ahmed KJ Behl HM 2002 ). Do leaf surface characters play a role in plant resistance to auto-exhaust pollution?
Flora , 197 , 47 -55 .
[本文引用: 1]
[54]
Perez-Estrada LB Cano-Santana Z Oyama K 2000 ). Variation in leaf trichomes of Wigandia urens: Environmental factors and physiological consequences
. Tree Physiology , 20 , 629 -632 .
[本文引用: 1]
[55]
Perrone R de Rosa P de Castro O Colombo P 2013 ). Leaf and stem anatomy in eight Hypericum species (Clusiaceae)
. Acta Botanica Croatica , 72 , 269 -286 .
[本文引用: 1]
[56]
Perrone R Salmeri C Brullo S Colombo P de Castro O 2015 ). What do leaf anatomy and micro-morphology tell us about the psammophilous Pancratium maritimum L.(Amaryllidaceae) in response to sand dune conditions?
Flora , 213 , 20 -31 .
[本文引用: 2]
[57]
Qi RH Li YZ Fan JP 1998 ). The idioblasts and their ecological significance in leaves of 16 alpine plants
.Acta Botanica Boreali-Occidentalia Sinica , 18 , 270 -276 . (in Chinese with English abstract)[祁如虎 , 李有忠 , 范建平 (1998 ). 16种高山植物叶片内的异细胞及其生态学意义
. 西北植物学报 , 18 , 270 -276 .]
[本文引用: 1]
[58]
Retamales HA Scharaschkin T 2015 ). Comparative leaf anatomy and micromorphology of the Chilean Myrtaceae: Taxonomic and ecological implications
.Flora , 217 , 138 -154 .
[本文引用: 1]
[59]
Roth I 1984 ). Stratification of Tropical Forests As Seen in Leaf Structure
. Dr W Junk Publishers, Hague.
[60]
Rusydi A Talip N Latip J Rahman RA Sharif I 2013 ). Morphology of trichomes inPogostemon cablin Benth.(Lamiaceae)
. Australian Journal of Crop Science , 7 , 744 -749 .
[本文引用: 1]
[61]
Sam O Jerez E Dell’amiico J Ruiz Sanchez MC 2000 ). Water stress induced changes in anatomy of tomato leaf epidermis
.Biologia Plantarum , 43 , 275 -277 .
[本文引用: 1]
[62]
Scatena VL Giulietti AM Borba EL van den Berg C 2005 ). Anatomy of Brazilian Eriocaulaceae: Correlation with taxonomy and habitat using multivariate analyses
.Plant Systematics and Evolution , 253 , 1 -22 .
[本文引用: 1]
[63]
Shafiee-Hajiabad M Hardt M Honermeier B 2014 ). Comparative investigation about the trichome morphology of common oregano (Origanum vulgare L. subsp. vulgare) and Greek oregano(Origanum vulgare L. subsp. hirtum)
. Journal of Applied Research on Medicinal and Aromatic Plants , 1 , 50 -58 .
[本文引用: 1]
[64]
Shepherd T Wynne GD 2006 ). The effects of stress on plant cuticular waxes
.New Phytologist , 171 , 469 -499 .
[本文引用: 1]
[65]
Shield LM 1950 ). Leaf xeromorphy as related to physiological and structural influences
.Botanical Review , 16 , 4399 -4471 .
[本文引用: 1]
[66]
Smith WK Nobel PS 1977 ). Influences of seasonal changes in leaf morphology on water-use efficiency for three desert broadleaf shrubs
. Ecology , 58 , 1033 -1043 .
[本文引用: 1]
[67]
Stenglein SA Arambarri AM Balatti PA 2005 ). Leaf epidermal characters related with plant’s passive resistance to pathogens vary among accessions of wild beans Phaseolus vulgaris var. aborigineus (Leguminosae-Phaseoleae)
. Flora , 200 , 285 -295 .
[本文引用: 1]
[68]
Talukdar T 2015 ). Micromorphomics: A morphological dissection to unveil environmental stress
. In: Barh D, Khan MS, Davies E eds. Plant Omics: The Omics of Plant Science. Springer, New Delhi , India . 615 -632 .
[本文引用: 1]
[69]
Tissier A 2012 ). Trichome specific expression: Promoters and their applications
. In: Ciftci YO ed. Transgenic Plants- Advances and Limitations. Tech Europe, Rijeka, Croatia.
[本文引用: 1]
[70]
Turner IM 2001 ). The Ecology of Trees in the Tropical Rain Forest . Cambridge University Press , Cambridge, UK .
[本文引用: 1]
[71]
Wagner P Furstner R Barthlott W Neinhuis C 2003 ). Quantitative assessment to the structural basis of water repellency in natural and technical surfaces
.Journal of Experimental Botany , 54 , 1295 -1303 .
[本文引用: 2]
[72]
Wang DC Yang YP Chen JH Li X 2012 ). Leaf epidermal microfeatures of 28 Salix species under scanning electronic microscope and their taxonomical significances
. Plant Diversity and Resources , 34 , 430 -442 . (in Chinese with English abstract)[王东超 , 杨永平 , 陈家辉 , 李馨 (2012 ). 28种柳属植物的叶表皮微形态特征及其分类学意义
. 植物分类与资源学报 , 34 , 430 -442 .]
[本文引用: 1]
[73]
Wang XL 1993 ). Advances of plant ecological anatomy
.Chinese Bulletin of Botany , 10 (Suppl. ), 1 -10 . (in Chinese with English abstract)[王勋陵 (1993 ).植物生态解剖学研究发展
.植物学通报 , 10 (Suppl. ), 1 -10 .]
[本文引用: 1]
[74]
Wang XL Ma J 1999 ). A study on leaf-structure and the diversity of xerophytes ecology adaptation
.Acta Ecologica Sinica , 19 , 787 -792 . (in Chinese with English abstract)[王勋陵 , 马骥 (1999 ). 从旱生植物叶结构探讨其生态适应的多样性
. 生态学报 , 19 , 787 -792 .]
[本文引用: 2]
[75]
Wang XL Wang J 1989 ). Plant Morphology, Structure and Environment . Lanzhou University Press , Lanzhou . (in Chinese)[王勋陵 , 王静 (1989 ). 植物形态结构与环境 .兰州大学出版社 , 兰州 .]
[本文引用: 2]
[76]
Werker E 1993 ). Function of essential oil-secreting glandular trichome in aromatic plants of the Lamiaceae—A review
.The Flavour and Fragrance Journal , 8 , 249 -255 .
[本文引用: 2]
[77]
Wuenscher JE 1970 ). The effect of leaf hairs of Verbascum thapsus on leaf energy exchange
. New Phytologist , 69 , 65 -73 .
[本文引用: 1]
[78]
Xu Z Zhou G 2008 ). Responses of leaf stomatal density to water status and its relationship with photosynthesis in a grass
.Journal of Experimental Botany , 59 , 3317 -3325 .
[本文引用: 1]
[79]
Yan QD Su PX Chen HB Zhang LM 2008 ). Comparative studies on crystal idioblasts of five C4 desert plants
.Journal of Plant Ecology (Chinese Version) , 32 , 873 -888 . (in Chinese with English abstract)[严巧娣 , 苏培玺 , 陈宏彬 , 张岭梅 (2008 ). 五种C4 荒漠植物光合器官中含晶细胞的比较分析
. 植物生态学报 , 32 , 873 -888 .]
[本文引用: 1]
[80]
Yang MB Yang JL Yang JY Liang N Qing H 2007 ). Changes on characteristics of the leaf epidermis and genetic diversity of Caragana davazamcii in different habitats in Ordos plateau, China
. Journal of Plant Ecology (Chinese Version) , 31 , 1181 -1189 . (in Chinese with English abstract)[杨明博 , 杨吉力 , 杨九艳 , 梁娜 , 清华 (2007 ). 鄂尔多斯高原不同生境条件下中间锦鸡儿植物叶片表皮特征及遗传多样性变化分析
. 植物生态学报 , 31 , 1181 -1189 .]
[本文引用: 1]
[81]
Zhang Y Zeng C Li D 2014 ). Scanning electron microscopy of the leaf epidermis in Arundinarieae(Poaceae: Bambusoideae): Evolutionary implications of selected micromorphological features
. Botanical Journal of the Linnean Society , 176 , 46 -65 .
[本文引用: 1]
[82]
Zhao HL 2012 ). Desert Ecology . Science Press , Beijing . (in Chi- nese)[赵哈林 (2012 ). 沙漠生态学 . 科学出版社 , 北京 .]
[本文引用: 1]
[83]
Zhao XX Li Y Su SP Bai X 2014 ). Drought resistance analysis based on anatomical structures of assimilating shoots of Calligonum mongolicum from three geographic populations
. Journal of Desert Research , 34 , 1293 -1300 . (in Chinese with English abstract)[赵小仙 , 李毅 , 苏世平 , 白潇 (2014 ). 3个地理种群蒙古沙拐枣同化枝解剖结构及抗旱性比较
.中国沙漠 , 34 , 1293 -1300 .]
[本文引用: 1]
[84]
Zhou W Xia NH 2012 ). Leaf epidermal features of Lithocarpus (Fagaceae) from China and their systematic significance
. Botanical Journal of Linnean Society , 168 , 216 -228 .
[本文引用: 1]
[85]
Zhou ZB Li PJ 2002 ). A review on the phytotomy research of xerophytes in China
.Arid Zone Research , 19 (1 ), 35 -40 . (in Chinese with English abstract)[周智斌 , 李培军 (2002 ). 我国旱生植物的形态解剖学研究
. 干旱区研究 , 19 (1 ), 35 -40 .]
[本文引用: 1]
[86]
Zhu GL Deng RH Wei XZ 2016 ). Leaf epidermal micromorphology of Ziziphus jujuba var. spinosa in response to a gradient of drought stress
. Acta Ecologica Sinica , 36 , 1 -11 . (in Chinese with English abstract)[朱广龙 , 邓荣华 , 魏学智 (2016 ). 酸枣叶表皮微形态对不同生态环境的适应特征
. 生态学报 , 36 , 1 -11 .]
[本文引用: 2]
Comparative foliar epidermal micromorphology of Southern African Strychnos L.(Loganiaceae): Taxonomic, ecological and cytological considerations
4
2014
... 植物叶片对环境的响应首先是叶片表皮对环境的感应和变化(Pal et al ., 2002 ; Bargel et al ., 2004 ; Koch et al ., 2009 ; Adebowale et al ., 2014 ).生长于干燥的气候和土壤条件下的荒漠植物, 叶表皮细胞会变小, 切向壁加厚, 具有内皮层.叶表皮角质膜厚度增加, 角质层由光滑变得粗糙, 能够反射阳光, 减少叶片表面空气的流动, 降低蒸腾作用, 防止水分过度丧失(贺金生等, 1994 ; 杨明博等, 2007 ).气孔的分布特征、密度和面积等都会受到影响, 气孔多分布于叶片下表皮, 这种分布模式既可促进植物与外界环境进行气体交换, 又能保持水分.随着环境中水分和湿度的减少, 气孔密度增加, 气孔面积则向小型化发展, 气孔多下陷形成气孔窝或其上有突出的角质膜(Sam et al ., 2000 ; Bosabalidis & Kofidis, 2002 ; 高建平等, 2003 ; Xu & Zhou, 2008 ; Gan et al ., 2010 ; de Micco & Aronne, 2012 ).叶表皮附属物(如表皮毛和表面疣状突起物)也发生变化, 它们可有效地避免叶肉组织受损(Gairola et al ., 2009 ; Koch et al ., 2009 ; Gomes et al ., 2011 ; Noman et al ., 2012 ), 表皮蜡质(waxes)晶体形成二维或三维结构, 具有降低表皮的可湿性以及反射强光等作用(Wagner et al ., 2003 ; Koch & Ensikat, 2008 ). ...
... 植物叶表皮微形态特征已用于解决物种分类问题、建立物种进化关系、阐明表皮微形态结构的生态适应性等多种研究目的(Dickison, 2000 ; Scatena et al ., 2005 ; Stenglein et al ., 2005 ; Aliero et al ., 2006 ; Chen et al ., 2010 ; Zhou & Xia, 2012 ; Adebowale et al ., 2014 ; Deng et al ., 2014 ; Zhang et al ., 2014 ; Francisco et al ., 2015 ; 朱广龙等, 2016 ).为了探讨荒漠植物叶片表皮微形态结构的适应特征及其亲缘关系, 选择分布于同一区域的不同植物和不同降水量梯度生境下的同一植物, 利用扫描电镜观察分析了117种荒漠植物叶片的表皮微形态结构, 旨在阐明荒漠植物对逆境的适应机理及其演化趋势, 并为优良固沙植物的筛选提供理论依据. ...
... 植物叶片形态结构与抗逆性关系的研究表明, 植物叶片小、表皮附属物密度大、且表皮皱缩程度高, 则抗旱性强(王勋陵和马骥, 1999 ; 周智斌和李培军, 2002 ; Bargel et al ., 2004 ; 迟丽华和宋凤斌, 2006 ; Adebowale et al ., 2014 ; 赵小仙等, 2014 ; 朱广龙等, 2016 ).通过比较荒漠植物叶片(或同化枝)的表皮微形态结构, 可以从表皮细胞形态, 表皮附属物如绒毛、蜡质、气孔等的形态、密度、结构和分布特征来进行环境适应性分析.根据表皮微形态结构和附属物与抗逆性的关系, 将荒漠植物适应环境胁迫的表皮微形态分为以下6种主要类型. ...
... 另外, 表面乳突或表皮皱褶, 一方面增加了叶表面积, 另一方面使气孔器能够穴居于凹陷处而避免强光的直射(Adebowale et al ., 2014 ).选取的117种荒漠植物的气孔密度主要集中于100-200个·mm-2 , 个别植物的气孔密度相当低, 如红砂属、柽柳属植物、沙枣、沙棘(Hippophae rhamnoides )等, 并且多数灌木的气孔器下陷, 降低了水分的气孔性散失.同时, 从结果可以看出气孔密度的高低与表皮其他附属物也有关系, 表皮毛和蜡质少的植物, 气孔密度一般相对较高, 如表皮微形态抗逆适应主要类型的第(5)、(6)类植物中的骆驼刺、杠柳等.表皮附属物的基本功能是保护叶片免受不利环境的直接损伤, 从而调节植物的生理功能. ...
The foliar micromorphology of Solanum pseudocapsicum
1
2006
... 植物叶表皮微形态特征已用于解决物种分类问题、建立物种进化关系、阐明表皮微形态结构的生态适应性等多种研究目的(Dickison, 2000 ; Scatena et al ., 2005 ; Stenglein et al ., 2005 ; Aliero et al ., 2006 ; Chen et al ., 2010 ; Zhou & Xia, 2012 ; Adebowale et al ., 2014 ; Deng et al ., 2014 ; Zhang et al ., 2014 ; Francisco et al ., 2015 ; 朱广龙等, 2016 ).为了探讨荒漠植物叶片表皮微形态结构的适应特征及其亲缘关系, 选择分布于同一区域的不同植物和不同降水量梯度生境下的同一植物, 利用扫描电镜观察分析了117种荒漠植物叶片的表皮微形态结构, 旨在阐明荒漠植物对逆境的适应机理及其演化趋势, 并为优良固沙植物的筛选提供理论依据. ...
Paraheliotropism in Robinia pseudoacacia L.: An efficient strategy to optimise photosynthetic performance under natural environmental conditions
1
2008
... 植物叶片适应环境的形态结构变化是多方面的.在众多生态因子中, 叶片受水分、温度和光因子的影响最为显著(王勋陵和王静, 1989 ; 王勋陵, 1993 ; Aronne & de Micco 2001 ; Arena et al ., 2008 ; Jaleel et al ., 2009a , 2009b ), 主要表现为叶形的变化、叶片表皮附属物的变化、叶片的厚度及解剖结构的差异(Talukdar, 2015 ).对于荒漠植物来说, 夏季极端干旱时它们可能同时遭受以上各种环境因子的胁迫.但其最主要的限制因子为水分, 因而表现出典型的旱生植物形态结构特征.由于遗传因子与环境因子的共同作用, 在旱生植物结构的适应性上出现了两种演化趋势, 即趋同性和趋异性, 也就是说, 在相同环境下的植物演化出相似的形态结构适应性, 而不同环境下的同类植物演化出不同的形态结构(Lakusic et al ., 2010 ).因此, 要系统分析荒漠植物叶片形态结构特征的适应性变化, 需要将一个区域不同植物和不同梯度生境下的同一植物做全面分析. ...
Seasonal dimorphism in the Mediterranean Cistus incanus L. subsp. incanus
2
2001
... 植物叶片适应环境的形态结构变化是多方面的.在众多生态因子中, 叶片受水分、温度和光因子的影响最为显著(王勋陵和王静, 1989 ; 王勋陵, 1993 ; Aronne & de Micco 2001 ; Arena et al ., 2008 ; Jaleel et al ., 2009a , 2009b ), 主要表现为叶形的变化、叶片表皮附属物的变化、叶片的厚度及解剖结构的差异(Talukdar, 2015 ).对于荒漠植物来说, 夏季极端干旱时它们可能同时遭受以上各种环境因子的胁迫.但其最主要的限制因子为水分, 因而表现出典型的旱生植物形态结构特征.由于遗传因子与环境因子的共同作用, 在旱生植物结构的适应性上出现了两种演化趋势, 即趋同性和趋异性, 也就是说, 在相同环境下的植物演化出相似的形态结构适应性, 而不同环境下的同类植物演化出不同的形态结构(Lakusic et al ., 2010 ).因此, 要系统分析荒漠植物叶片形态结构特征的适应性变化, 需要将一个区域不同植物和不同梯度生境下的同一植物做全面分析. ...
... 大多数荒漠植物叶片的一个显著特点是表皮被绒毛, 叶表皮毛的出现与长期的干旱气候有关, 是植物对干旱环境的适应特征(Retamales & Scharaschkin, 2015 ).植物叶片表皮毛分两种: 腺毛和非腺毛(Werker, 1993 ; Navarro & Oualidi, 1999 ).非腺毛的主要功能与植物的防御有关.目前确认的植物叶片表皮非腺毛具有的生理生态功能主要有: 减轻紫外辐射(Karabourniotis et al ., 1992 ; Gregoriou et al ., 2007 )、降低边界层的传导(Wuenscher, 1970 ; Kenzo et al ., 2008 )、降低叶温(Ehleringer & Mooney, 1978 ), 以及减少对太阳辐射的接收(Ehleringer et al ., 1976 )等.这些功能决定了干旱区荒漠植物抗逆能力的大小.干旱地区的叶表皮毛密度比湿润地区的高(Smith & Nobel, 1977 ; Aronne & de Micco, 2001 ), 就单个物种而言, 表皮毛密度与温度和辐射密切相关(Perez-Estrada et al ., 2000 ).在热带雨林树种中, 浓密的表皮毛很少见(Turner, 2001 ), Roth (1984)调查的250种树中仅有15种有较厚的表皮毛, 且较厚的表皮毛一般在干旱、高温和强辐射条件下产生.而在干旱半干旱的荒漠区, 大多数植物都具有浓密的表皮毛.显然, 荒漠植物叶上密生的表皮毛对于植物的生活有重要的意义.从Ehleringer等 (1976)、Ehleringer和Bjorkman (1978)、Ehleringer和Mooney (1978)研究的荒漠灌木Encelia farinosa 的叶表皮毛对叶片光谱特性和光合、蒸腾等生理活动的影响来看, 沙漠毒菊在不同生长季生长的叶片具有不同数量的表皮毛, 表皮毛的变化足够使叶片对光合有效辐射的吸收从81%降到29%, 对总的太阳辐射的吸收从46%降到16%, 而亲缘关系较近却没有表皮毛的同属物种Encelia californiea 的有效光合辐射吸收率为84%.同时, 由于对太阳辐射吸收的减少, 导致叶面温度和植物蒸腾速率的降低.荒漠环境气温经常超过40 ℃, 而植物最适宜的光合温度是25 ℃, 当温度超过35 ℃时植物的光合速率会突降, 但较厚的叶表皮毛能使叶温显著低于气温而接近光合适宜温度, 同时也避免植物达到叶片致死温度.但叶表皮毛也有不利的一面, 表皮毛的覆盖降低了光合速率, 因此同一植物在不同的生境和生长季长出的叶片会出现表皮毛数量不同的特征, 这也是植物适应环境的一种权衡策略(Kenzo et al ., 2008 ).从本研究中的117种荒漠植物叶表皮特征来看, 片状叶片基本被有表皮毛, 并有不少叶片的表皮毛数量较多, 直接增加了叶片的边界层厚度, 个别植物的上下表皮毛厚度远超于叶片厚度, 如表皮微形态抗逆适应主要类型的第一类植物中的柠条锦鸡儿、卷叶锦鸡儿和沙枣等.可见荒漠植物中叶片表皮毛的作用是相当重要的, 它们不仅能调节植物的生理功能, 而且可抵御风沙等机械损伤的直接伤害.这些物种在不同的自然生境中其表皮毛数量亦不同(数据省略), 说明生态环境深刻地影响着植物形态结构的形成与变异. ...
Plant cuticles: Multifunctional interfaces between plant and environment. In: Hemsley AR, Poole I eds. Evolution of Plant Physiology
3
2004
... 植物叶片对环境的响应首先是叶片表皮对环境的感应和变化(Pal et al ., 2002 ; Bargel et al ., 2004 ; Koch et al ., 2009 ; Adebowale et al ., 2014 ).生长于干燥的气候和土壤条件下的荒漠植物, 叶表皮细胞会变小, 切向壁加厚, 具有内皮层.叶表皮角质膜厚度增加, 角质层由光滑变得粗糙, 能够反射阳光, 减少叶片表面空气的流动, 降低蒸腾作用, 防止水分过度丧失(贺金生等, 1994 ; 杨明博等, 2007 ).气孔的分布特征、密度和面积等都会受到影响, 气孔多分布于叶片下表皮, 这种分布模式既可促进植物与外界环境进行气体交换, 又能保持水分.随着环境中水分和湿度的减少, 气孔密度增加, 气孔面积则向小型化发展, 气孔多下陷形成气孔窝或其上有突出的角质膜(Sam et al ., 2000 ; Bosabalidis & Kofidis, 2002 ; 高建平等, 2003 ; Xu & Zhou, 2008 ; Gan et al ., 2010 ; de Micco & Aronne, 2012 ).叶表皮附属物(如表皮毛和表面疣状突起物)也发生变化, 它们可有效地避免叶肉组织受损(Gairola et al ., 2009 ; Koch et al ., 2009 ; Gomes et al ., 2011 ; Noman et al ., 2012 ), 表皮蜡质(waxes)晶体形成二维或三维结构, 具有降低表皮的可湿性以及反射强光等作用(Wagner et al ., 2003 ; Koch & Ensikat, 2008 ). ...
... 植物叶片形态结构与抗逆性关系的研究表明, 植物叶片小、表皮附属物密度大、且表皮皱缩程度高, 则抗旱性强(王勋陵和马骥, 1999 ; 周智斌和李培军, 2002 ; Bargel et al ., 2004 ; 迟丽华和宋凤斌, 2006 ; Adebowale et al ., 2014 ; 赵小仙等, 2014 ; 朱广龙等, 2016 ).通过比较荒漠植物叶片(或同化枝)的表皮微形态结构, 可以从表皮细胞形态, 表皮附属物如绒毛、蜡质、气孔等的形态、密度、结构和分布特征来进行环境适应性分析.根据表皮微形态结构和附属物与抗逆性的关系, 将荒漠植物适应环境胁迫的表皮微形态分为以下6种主要类型. ...
... 荒漠植物叶片的另一个显著特点是表皮有较厚的角质膜蜡质层.角质膜蜡质层与表皮毛有共同的功能, 是植物表皮的保护屏障, 主要由长链碳氢化合物组成(Shepherd & Wynne, 2006 ).叶片对光的反射由叶面形貌决定, 表皮毛和蜡质层是决定荒漠植物叶表面形貌的关键.Holmes和Keiller (2002)研究了45种桉属和伽蓝菜属植物的蜡质叶, 发现蜡质层对紫外光(330 nm)和光合有效辐射(680 nm)的反射比各种形态的表皮毛更强, 去除蜡质层通常会减少对这两种光的反射, 特别是对紫外光的反射, 但对表皮无蜡质的叶片影响不大.同时不同的物种对不同波长的光的反射亦不同, 表明蜡质晶体的形态直接影响着对光的反射, 并且不同温度下的反射强度也不同, 可见蜡质层对光的反射特性变化很大(Holmes & Keiller, 2002 ).叶表皮蜡质层参与植物对水分、盐、低温、污染、机械损伤等多种胁迫响应的过程, 提高植物对胁迫的抗性(Cameron, 1970 ; Bargel et al ., 2004 ; Perrone et al ., 2013 , 2015 ).因此, 本研究中117种荒漠植物叶片表皮颗粒状、膜片状、晶簇状、鳞片状、杆状等各种不同形态的蜡质晶体既是植物长期适应环境的结果, 也对荒漠植物在严酷环境中的生长和生存至关重要.总的来说, 这些植物中灌木的蜡质层密度高于草本, 无表皮毛的叶片蜡质层密度高于表皮毛覆盖的, 特别是以同化枝为主要光合器官和棒状叶片的植物, 表皮基本没有表皮毛覆盖, 主要的附属物是蜡质层, 如麻黄属、沙拐枣属、红砂属的植物等, 弥补了表皮毛的功能. ...
Epidermal and seed surface characters of plants: Systematic applicability and some evolutionary aspects
1
1981
... 荒漠植物由于长期适应所生长的自然环境, 发展了一套最适宜自身生长发育的形态结构特征, 并采取各种途径来抵御或忍耐干旱胁迫对它的影响, 因而具有一定程度的耐旱性.这种耐旱性是一种复合性状, 包括植物的形态结构特征、水分生理特征、生理生化反应、组织细胞、光合器官以及原生质结构等特征的综合反映(Klich, 2000 ; Chaves et al ., 2003 ; Ogle & Reynolds, 2004 ).不同的荒漠植物对水分和不利环境的适应和耐受程度不同, 适应方式多种多样, 按照不同的分类标准, 荒漠植物有多种适应类型.通常的分类标准包括耐旱程度、耐旱方式、叶片形态、生活史、光合碳代谢途径等(赵哈林, 2012 ).各种类型的植物又有各自的适应特征.本研究是针对我国干旱半干旱区荒漠植物叶表皮微形态结构特征的综合分析, 主要集中于该区域的建群种灌木物种.根据荒漠植物叶表皮微形态基本结构特征和表皮附属物的类型与数量, 将荒漠植物表皮微形态类型直观地划分为11种, 而将抗逆适应微形态结构划分为6种类型.荒漠植物叶表皮微形态结构类型复杂, 反映了植物自身的遗传特性及其与环境的密切关系.植物叶片表皮微形态主要由遗传因素决定, 但表皮附属物绒毛数量, 蜡质形态与厚度, 气孔的形态、密度以及分布特征等与环境因子密切相关, 是植物对环境的响应(Barthlott, 1981 ; Casson & Hetherington, 2010 ; Ghahremaninejad et al ., 2012 ).因此, 基于这些表皮主要附属物特征对荒漠植物的表皮微形态类型和抗逆适应类型进行划分, 能够从微观形态结构特征进一步揭示物种的亲缘关系和表皮微形态结构的生态适应意义. ...
Comparative effects of drought stress on leaf anatomy of two olive cultivars
2
2002
... 植物叶片对环境的响应首先是叶片表皮对环境的感应和变化(Pal et al ., 2002 ; Bargel et al ., 2004 ; Koch et al ., 2009 ; Adebowale et al ., 2014 ).生长于干燥的气候和土壤条件下的荒漠植物, 叶表皮细胞会变小, 切向壁加厚, 具有内皮层.叶表皮角质膜厚度增加, 角质层由光滑变得粗糙, 能够反射阳光, 减少叶片表面空气的流动, 降低蒸腾作用, 防止水分过度丧失(贺金生等, 1994 ; 杨明博等, 2007 ).气孔的分布特征、密度和面积等都会受到影响, 气孔多分布于叶片下表皮, 这种分布模式既可促进植物与外界环境进行气体交换, 又能保持水分.随着环境中水分和湿度的减少, 气孔密度增加, 气孔面积则向小型化发展, 气孔多下陷形成气孔窝或其上有突出的角质膜(Sam et al ., 2000 ; Bosabalidis & Kofidis, 2002 ; 高建平等, 2003 ; Xu & Zhou, 2008 ; Gan et al ., 2010 ; de Micco & Aronne, 2012 ).叶表皮附属物(如表皮毛和表面疣状突起物)也发生变化, 它们可有效地避免叶肉组织受损(Gairola et al ., 2009 ; Koch et al ., 2009 ; Gomes et al ., 2011 ; Noman et al ., 2012 ), 表皮蜡质(waxes)晶体形成二维或三维结构, 具有降低表皮的可湿性以及反射强光等作用(Wagner et al ., 2003 ; Koch & Ensikat, 2008 ). ...
... 而对于腺毛来说, 其主要作为一个油性物质的合成、分泌和累积的器官, 有效地防御动物的捕食、病菌入侵以及物种竞争(Werker, 1993 ; Wagner et al ., 2003 ; Kremer et al ., 2014 ; Munien et al ., 2015 ).实际上, 腺毛也是次生代谢产物的一个独立的细胞工厂(Tissier, 2012 ).腺毛的形态随植物的组织器官和种类不同而呈多样化, 其形成和分泌的主要油性物质及其产物组成受遗传和环境两方面的因素控制(Bosabalidis & Kofidis, 2002 ; Shafiee-Hajiabad et al ., 2014 ), 腺毛结构的多样性也可以为植物分类学提供依据(Gairola et al ., 2009 ; Rusydi et al ., 2013 ).对117种荒漠植物叶片表皮扫描发现: 不少旱生植物除具有非腺体表皮毛外, 还同时具有腺毛, 并且有些腺毛体积较大, 如蒙古莸、甘草、香青兰、星毛短舌菊、百里香等.显然, 对于具有腺毛的荒漠植物, 腺毛具有重要的功能.但目前关于不同植物腺毛的形态、形成以及功能的研究还很有局限性.那么, 这些腺毛在植物适应荒漠环境的过程中其形态结构与环境因子有何关系、主要分泌哪些化学物质、发挥什么样的功能?这些问题亟待下一步的深入研究. ...
Light intensity and the growth of Eucalyptus seedlings II. The effect of cuticular waxes on light absorption in leaves of Eucalyptus species
1
1970
... 荒漠植物叶片的另一个显著特点是表皮有较厚的角质膜蜡质层.角质膜蜡质层与表皮毛有共同的功能, 是植物表皮的保护屏障, 主要由长链碳氢化合物组成(Shepherd & Wynne, 2006 ).叶片对光的反射由叶面形貌决定, 表皮毛和蜡质层是决定荒漠植物叶表面形貌的关键.Holmes和Keiller (2002)研究了45种桉属和伽蓝菜属植物的蜡质叶, 发现蜡质层对紫外光(330 nm)和光合有效辐射(680 nm)的反射比各种形态的表皮毛更强, 去除蜡质层通常会减少对这两种光的反射, 特别是对紫外光的反射, 但对表皮无蜡质的叶片影响不大.同时不同的物种对不同波长的光的反射亦不同, 表明蜡质晶体的形态直接影响着对光的反射, 并且不同温度下的反射强度也不同, 可见蜡质层对光的反射特性变化很大(Holmes & Keiller, 2002 ).叶表皮蜡质层参与植物对水分、盐、低温、污染、机械损伤等多种胁迫响应的过程, 提高植物对胁迫的抗性(Cameron, 1970 ; Bargel et al ., 2004 ; Perrone et al ., 2013 , 2015 ).因此, 本研究中117种荒漠植物叶片表皮颗粒状、膜片状、晶簇状、鳞片状、杆状等各种不同形态的蜡质晶体既是植物长期适应环境的结果, 也对荒漠植物在严酷环境中的生长和生存至关重要.总的来说, 这些植物中灌木的蜡质层密度高于草本, 无表皮毛的叶片蜡质层密度高于表皮毛覆盖的, 特别是以同化枝为主要光合器官和棒状叶片的植物, 表皮基本没有表皮毛覆盖, 主要的附属物是蜡质层, 如麻黄属、沙拐枣属、红砂属的植物等, 弥补了表皮毛的功能. ...
Environmental regulation of stomatal development
1
2010
... 荒漠植物由于长期适应所生长的自然环境, 发展了一套最适宜自身生长发育的形态结构特征, 并采取各种途径来抵御或忍耐干旱胁迫对它的影响, 因而具有一定程度的耐旱性.这种耐旱性是一种复合性状, 包括植物的形态结构特征、水分生理特征、生理生化反应、组织细胞、光合器官以及原生质结构等特征的综合反映(Klich, 2000 ; Chaves et al ., 2003 ; Ogle & Reynolds, 2004 ).不同的荒漠植物对水分和不利环境的适应和耐受程度不同, 适应方式多种多样, 按照不同的分类标准, 荒漠植物有多种适应类型.通常的分类标准包括耐旱程度、耐旱方式、叶片形态、生活史、光合碳代谢途径等(赵哈林, 2012 ).各种类型的植物又有各自的适应特征.本研究是针对我国干旱半干旱区荒漠植物叶表皮微形态结构特征的综合分析, 主要集中于该区域的建群种灌木物种.根据荒漠植物叶表皮微形态基本结构特征和表皮附属物的类型与数量, 将荒漠植物表皮微形态类型直观地划分为11种, 而将抗逆适应微形态结构划分为6种类型.荒漠植物叶表皮微形态结构类型复杂, 反映了植物自身的遗传特性及其与环境的密切关系.植物叶片表皮微形态主要由遗传因素决定, 但表皮附属物绒毛数量, 蜡质形态与厚度, 气孔的形态、密度以及分布特征等与环境因子密切相关, 是植物对环境的响应(Barthlott, 1981 ; Casson & Hetherington, 2010 ; Ghahremaninejad et al ., 2012 ).因此, 基于这些表皮主要附属物特征对荒漠植物的表皮微形态类型和抗逆适应类型进行划分, 能够从微观形态结构特征进一步揭示物种的亲缘关系和表皮微形态结构的生态适应意义. ...
Understanding plant responses to drought—From genes to the whole plant
1
2003
... 荒漠植物由于长期适应所生长的自然环境, 发展了一套最适宜自身生长发育的形态结构特征, 并采取各种途径来抵御或忍耐干旱胁迫对它的影响, 因而具有一定程度的耐旱性.这种耐旱性是一种复合性状, 包括植物的形态结构特征、水分生理特征、生理生化反应、组织细胞、光合器官以及原生质结构等特征的综合反映(Klich, 2000 ; Chaves et al ., 2003 ; Ogle & Reynolds, 2004 ).不同的荒漠植物对水分和不利环境的适应和耐受程度不同, 适应方式多种多样, 按照不同的分类标准, 荒漠植物有多种适应类型.通常的分类标准包括耐旱程度、耐旱方式、叶片形态、生活史、光合碳代谢途径等(赵哈林, 2012 ).各种类型的植物又有各自的适应特征.本研究是针对我国干旱半干旱区荒漠植物叶表皮微形态结构特征的综合分析, 主要集中于该区域的建群种灌木物种.根据荒漠植物叶表皮微形态基本结构特征和表皮附属物的类型与数量, 将荒漠植物表皮微形态类型直观地划分为11种, 而将抗逆适应微形态结构划分为6种类型.荒漠植物叶表皮微形态结构类型复杂, 反映了植物自身的遗传特性及其与环境的密切关系.植物叶片表皮微形态主要由遗传因素决定, 但表皮附属物绒毛数量, 蜡质形态与厚度, 气孔的形态、密度以及分布特征等与环境因子密切相关, 是植物对环境的响应(Barthlott, 1981 ; Casson & Hetherington, 2010 ; Ghahremaninejad et al ., 2012 ).因此, 基于这些表皮主要附属物特征对荒漠植物的表皮微形态类型和抗逆适应类型进行划分, 能够从微观形态结构特征进一步揭示物种的亲缘关系和表皮微形态结构的生态适应意义. ...
Leaf epidermal characteristics of Asiatic buddleja L. under scanning electron microscope: Insights into chromosomal and taxonomic significance
1
2010
... 植物叶表皮微形态特征已用于解决物种分类问题、建立物种进化关系、阐明表皮微形态结构的生态适应性等多种研究目的(Dickison, 2000 ; Scatena et al ., 2005 ; Stenglein et al ., 2005 ; Aliero et al ., 2006 ; Chen et al ., 2010 ; Zhou & Xia, 2012 ; Adebowale et al ., 2014 ; Deng et al ., 2014 ; Zhang et al ., 2014 ; Francisco et al ., 2015 ; 朱广龙等, 2016 ).为了探讨荒漠植物叶片表皮微形态结构的适应特征及其亲缘关系, 选择分布于同一区域的不同植物和不同降水量梯度生境下的同一植物, 利用扫描电镜观察分析了117种荒漠植物叶片的表皮微形态结构, 旨在阐明荒漠植物对逆境的适应机理及其演化趋势, 并为优良固沙植物的筛选提供理论依据. ...
松嫩平原西部盐碱地区10种植物叶片结构特征及其生态适应性
1
2006
... 植物叶片形态结构与抗逆性关系的研究表明, 植物叶片小、表皮附属物密度大、且表皮皱缩程度高, 则抗旱性强(王勋陵和马骥, 1999 ; 周智斌和李培军, 2002 ; Bargel et al ., 2004 ; 迟丽华和宋凤斌, 2006 ; Adebowale et al ., 2014 ; 赵小仙等, 2014 ; 朱广龙等, 2016 ).通过比较荒漠植物叶片(或同化枝)的表皮微形态结构, 可以从表皮细胞形态, 表皮附属物如绒毛、蜡质、气孔等的形态、密度、结构和分布特征来进行环境适应性分析.根据表皮微形态结构和附属物与抗逆性的关系, 将荒漠植物适应环境胁迫的表皮微形态分为以下6种主要类型. ...
1
2012
... 植物叶片对环境的响应首先是叶片表皮对环境的感应和变化(Pal et al ., 2002 ; Bargel et al ., 2004 ; Koch et al ., 2009 ; Adebowale et al ., 2014 ).生长于干燥的气候和土壤条件下的荒漠植物, 叶表皮细胞会变小, 切向壁加厚, 具有内皮层.叶表皮角质膜厚度增加, 角质层由光滑变得粗糙, 能够反射阳光, 减少叶片表面空气的流动, 降低蒸腾作用, 防止水分过度丧失(贺金生等, 1994 ; 杨明博等, 2007 ).气孔的分布特征、密度和面积等都会受到影响, 气孔多分布于叶片下表皮, 这种分布模式既可促进植物与外界环境进行气体交换, 又能保持水分.随着环境中水分和湿度的减少, 气孔密度增加, 气孔面积则向小型化发展, 气孔多下陷形成气孔窝或其上有突出的角质膜(Sam et al ., 2000 ; Bosabalidis & Kofidis, 2002 ; 高建平等, 2003 ; Xu & Zhou, 2008 ; Gan et al ., 2010 ; de Micco & Aronne, 2012 ).叶表皮附属物(如表皮毛和表面疣状突起物)也发生变化, 它们可有效地避免叶肉组织受损(Gairola et al ., 2009 ; Koch et al ., 2009 ; Gomes et al ., 2011 ; Noman et al ., 2012 ), 表皮蜡质(waxes)晶体形成二维或三维结构, 具有降低表皮的可湿性以及反射强光等作用(Wagner et al ., 2003 ; Koch & Ensikat, 2008 ). ...
Leaf epidermal features of Quercus subgenus Cyclo- balanopsis(Fagaceae) and their systematic significance
1
2014
... 植物叶表皮微形态特征已用于解决物种分类问题、建立物种进化关系、阐明表皮微形态结构的生态适应性等多种研究目的(Dickison, 2000 ; Scatena et al ., 2005 ; Stenglein et al ., 2005 ; Aliero et al ., 2006 ; Chen et al ., 2010 ; Zhou & Xia, 2012 ; Adebowale et al ., 2014 ; Deng et al ., 2014 ; Zhang et al ., 2014 ; Francisco et al ., 2015 ; 朱广龙等, 2016 ).为了探讨荒漠植物叶片表皮微形态结构的适应特征及其亲缘关系, 选择分布于同一区域的不同植物和不同降水量梯度生境下的同一植物, 利用扫描电镜观察分析了117种荒漠植物叶片的表皮微形态结构, 旨在阐明荒漠植物对逆境的适应机理及其演化趋势, 并为优良固沙植物的筛选提供理论依据. ...
1
2000
... 植物叶表皮微形态特征已用于解决物种分类问题、建立物种进化关系、阐明表皮微形态结构的生态适应性等多种研究目的(Dickison, 2000 ; Scatena et al ., 2005 ; Stenglein et al ., 2005 ; Aliero et al ., 2006 ; Chen et al ., 2010 ; Zhou & Xia, 2012 ; Adebowale et al ., 2014 ; Deng et al ., 2014 ; Zhang et al ., 2014 ; Francisco et al ., 2015 ; 朱广龙等, 2016 ).为了探讨荒漠植物叶片表皮微形态结构的适应特征及其亲缘关系, 选择分布于同一区域的不同植物和不同降水量梯度生境下的同一植物, 利用扫描电镜观察分析了117种荒漠植物叶片的表皮微形态结构, 旨在阐明荒漠植物对逆境的适应机理及其演化趋势, 并为优良固沙植物的筛选提供理论依据. ...
Pubescence and leaf spectral characteristics in a desert shrub,Encelia farinosa
1978
Leaf pubescence: Effects on absorptance and photosynthesis in a desert shrub
1
1976
... 大多数荒漠植物叶片的一个显著特点是表皮被绒毛, 叶表皮毛的出现与长期的干旱气候有关, 是植物对干旱环境的适应特征(Retamales & Scharaschkin, 2015 ).植物叶片表皮毛分两种: 腺毛和非腺毛(Werker, 1993 ; Navarro & Oualidi, 1999 ).非腺毛的主要功能与植物的防御有关.目前确认的植物叶片表皮非腺毛具有的生理生态功能主要有: 减轻紫外辐射(Karabourniotis et al ., 1992 ; Gregoriou et al ., 2007 )、降低边界层的传导(Wuenscher, 1970 ; Kenzo et al ., 2008 )、降低叶温(Ehleringer & Mooney, 1978 ), 以及减少对太阳辐射的接收(Ehleringer et al ., 1976 )等.这些功能决定了干旱区荒漠植物抗逆能力的大小.干旱地区的叶表皮毛密度比湿润地区的高(Smith & Nobel, 1977 ; Aronne & de Micco, 2001 ), 就单个物种而言, 表皮毛密度与温度和辐射密切相关(Perez-Estrada et al ., 2000 ).在热带雨林树种中, 浓密的表皮毛很少见(Turner, 2001 ), Roth (1984)调查的250种树中仅有15种有较厚的表皮毛, 且较厚的表皮毛一般在干旱、高温和强辐射条件下产生.而在干旱半干旱的荒漠区, 大多数植物都具有浓密的表皮毛.显然, 荒漠植物叶上密生的表皮毛对于植物的生活有重要的意义.从Ehleringer等 (1976)、Ehleringer和Bjorkman (1978)、Ehleringer和Mooney (1978)研究的荒漠灌木Encelia farinosa 的叶表皮毛对叶片光谱特性和光合、蒸腾等生理活动的影响来看, 沙漠毒菊在不同生长季生长的叶片具有不同数量的表皮毛, 表皮毛的变化足够使叶片对光合有效辐射的吸收从81%降到29%, 对总的太阳辐射的吸收从46%降到16%, 而亲缘关系较近却没有表皮毛的同属物种Encelia californiea 的有效光合辐射吸收率为84%.同时, 由于对太阳辐射吸收的减少, 导致叶面温度和植物蒸腾速率的降低.荒漠环境气温经常超过40 ℃, 而植物最适宜的光合温度是25 ℃, 当温度超过35 ℃时植物的光合速率会突降, 但较厚的叶表皮毛能使叶温显著低于气温而接近光合适宜温度, 同时也避免植物达到叶片致死温度.但叶表皮毛也有不利的一面, 表皮毛的覆盖降低了光合速率, 因此同一植物在不同的生境和生长季长出的叶片会出现表皮毛数量不同的特征, 这也是植物适应环境的一种权衡策略(Kenzo et al ., 2008 ).从本研究中的117种荒漠植物叶表皮特征来看, 片状叶片基本被有表皮毛, 并有不少叶片的表皮毛数量较多, 直接增加了叶片的边界层厚度, 个别植物的上下表皮毛厚度远超于叶片厚度, 如表皮微形态抗逆适应主要类型的第一类植物中的柠条锦鸡儿、卷叶锦鸡儿和沙枣等.可见荒漠植物中叶片表皮毛的作用是相当重要的, 它们不仅能调节植物的生理功能, 而且可抵御风沙等机械损伤的直接伤害.这些物种在不同的自然生境中其表皮毛数量亦不同(数据省略), 说明生态环境深刻地影响着植物形态结构的形成与变异. ...
Leaf hairs: Effects on physiological activity and adaptive value to a desert shrub
1
1978
... 大多数荒漠植物叶片的一个显著特点是表皮被绒毛, 叶表皮毛的出现与长期的干旱气候有关, 是植物对干旱环境的适应特征(Retamales & Scharaschkin, 2015 ).植物叶片表皮毛分两种: 腺毛和非腺毛(Werker, 1993 ; Navarro & Oualidi, 1999 ).非腺毛的主要功能与植物的防御有关.目前确认的植物叶片表皮非腺毛具有的生理生态功能主要有: 减轻紫外辐射(Karabourniotis et al ., 1992 ; Gregoriou et al ., 2007 )、降低边界层的传导(Wuenscher, 1970 ; Kenzo et al ., 2008 )、降低叶温(Ehleringer & Mooney, 1978 ), 以及减少对太阳辐射的接收(Ehleringer et al ., 1976 )等.这些功能决定了干旱区荒漠植物抗逆能力的大小.干旱地区的叶表皮毛密度比湿润地区的高(Smith & Nobel, 1977 ; Aronne & de Micco, 2001 ), 就单个物种而言, 表皮毛密度与温度和辐射密切相关(Perez-Estrada et al ., 2000 ).在热带雨林树种中, 浓密的表皮毛很少见(Turner, 2001 ), Roth (1984)调查的250种树中仅有15种有较厚的表皮毛, 且较厚的表皮毛一般在干旱、高温和强辐射条件下产生.而在干旱半干旱的荒漠区, 大多数植物都具有浓密的表皮毛.显然, 荒漠植物叶上密生的表皮毛对于植物的生活有重要的意义.从Ehleringer等 (1976)、Ehleringer和Bjorkman (1978)、Ehleringer和Mooney (1978)研究的荒漠灌木Encelia farinosa 的叶表皮毛对叶片光谱特性和光合、蒸腾等生理活动的影响来看, 沙漠毒菊在不同生长季生长的叶片具有不同数量的表皮毛, 表皮毛的变化足够使叶片对光合有效辐射的吸收从81%降到29%, 对总的太阳辐射的吸收从46%降到16%, 而亲缘关系较近却没有表皮毛的同属物种Encelia californiea 的有效光合辐射吸收率为84%.同时, 由于对太阳辐射吸收的减少, 导致叶面温度和植物蒸腾速率的降低.荒漠环境气温经常超过40 ℃, 而植物最适宜的光合温度是25 ℃, 当温度超过35 ℃时植物的光合速率会突降, 但较厚的叶表皮毛能使叶温显著低于气温而接近光合适宜温度, 同时也避免植物达到叶片致死温度.但叶表皮毛也有不利的一面, 表皮毛的覆盖降低了光合速率, 因此同一植物在不同的生境和生长季长出的叶片会出现表皮毛数量不同的特征, 这也是植物适应环境的一种权衡策略(Kenzo et al ., 2008 ).从本研究中的117种荒漠植物叶表皮特征来看, 片状叶片基本被有表皮毛, 并有不少叶片的表皮毛数量较多, 直接增加了叶片的边界层厚度, 个别植物的上下表皮毛厚度远超于叶片厚度, 如表皮微形态抗逆适应主要类型的第一类植物中的柠条锦鸡儿、卷叶锦鸡儿和沙枣等.可见荒漠植物中叶片表皮毛的作用是相当重要的, 它们不仅能调节植物的生理功能, 而且可抵御风沙等机械损伤的直接伤害.这些物种在不同的自然生境中其表皮毛数量亦不同(数据省略), 说明生态环境深刻地影响着植物形态结构的形成与变异. ...
Morphological phylogenetic analysis of Ophrys(Orchidaceae): Insights from morpho-anatomical floral traits into the interspecific relationships in an unresolved clade
1
2015
... 植物叶表皮微形态特征已用于解决物种分类问题、建立物种进化关系、阐明表皮微形态结构的生态适应性等多种研究目的(Dickison, 2000 ; Scatena et al ., 2005 ; Stenglein et al ., 2005 ; Aliero et al ., 2006 ; Chen et al ., 2010 ; Zhou & Xia, 2012 ; Adebowale et al ., 2014 ; Deng et al ., 2014 ; Zhang et al ., 2014 ; Francisco et al ., 2015 ; 朱广龙等, 2016 ).为了探讨荒漠植物叶片表皮微形态结构的适应特征及其亲缘关系, 选择分布于同一区域的不同植物和不同降水量梯度生境下的同一植物, 利用扫描电镜观察分析了117种荒漠植物叶片的表皮微形态结构, 旨在阐明荒漠植物对逆境的适应机理及其演化趋势, 并为优良固沙植物的筛选提供理论依据. ...
An investigation of the foliar trichomes of Tetradenia riparia(Hochst.) Codd [Lamiaceae]: An important medicinal plant of Southern Africa
2
2009
... 植物叶片对环境的响应首先是叶片表皮对环境的感应和变化(Pal et al ., 2002 ; Bargel et al ., 2004 ; Koch et al ., 2009 ; Adebowale et al ., 2014 ).生长于干燥的气候和土壤条件下的荒漠植物, 叶表皮细胞会变小, 切向壁加厚, 具有内皮层.叶表皮角质膜厚度增加, 角质层由光滑变得粗糙, 能够反射阳光, 减少叶片表面空气的流动, 降低蒸腾作用, 防止水分过度丧失(贺金生等, 1994 ; 杨明博等, 2007 ).气孔的分布特征、密度和面积等都会受到影响, 气孔多分布于叶片下表皮, 这种分布模式既可促进植物与外界环境进行气体交换, 又能保持水分.随着环境中水分和湿度的减少, 气孔密度增加, 气孔面积则向小型化发展, 气孔多下陷形成气孔窝或其上有突出的角质膜(Sam et al ., 2000 ; Bosabalidis & Kofidis, 2002 ; 高建平等, 2003 ; Xu & Zhou, 2008 ; Gan et al ., 2010 ; de Micco & Aronne, 2012 ).叶表皮附属物(如表皮毛和表面疣状突起物)也发生变化, 它们可有效地避免叶肉组织受损(Gairola et al ., 2009 ; Koch et al ., 2009 ; Gomes et al ., 2011 ; Noman et al ., 2012 ), 表皮蜡质(waxes)晶体形成二维或三维结构, 具有降低表皮的可湿性以及反射强光等作用(Wagner et al ., 2003 ; Koch & Ensikat, 2008 ). ...
... 而对于腺毛来说, 其主要作为一个油性物质的合成、分泌和累积的器官, 有效地防御动物的捕食、病菌入侵以及物种竞争(Werker, 1993 ; Wagner et al ., 2003 ; Kremer et al ., 2014 ; Munien et al ., 2015 ).实际上, 腺毛也是次生代谢产物的一个独立的细胞工厂(Tissier, 2012 ).腺毛的形态随植物的组织器官和种类不同而呈多样化, 其形成和分泌的主要油性物质及其产物组成受遗传和环境两方面的因素控制(Bosabalidis & Kofidis, 2002 ; Shafiee-Hajiabad et al ., 2014 ), 腺毛结构的多样性也可以为植物分类学提供依据(Gairola et al ., 2009 ; Rusydi et al ., 2013 ).对117种荒漠植物叶片表皮扫描发现: 不少旱生植物除具有非腺体表皮毛外, 还同时具有腺毛, 并且有些腺毛体积较大, 如蒙古莸、甘草、香青兰、星毛短舌菊、百里香等.显然, 对于具有腺毛的荒漠植物, 腺毛具有重要的功能.但目前关于不同植物腺毛的形态、形成以及功能的研究还很有局限性.那么, 这些腺毛在植物适应荒漠环境的过程中其形态结构与环境因子有何关系、主要分泌哪些化学物质、发挥什么样的功能?这些问题亟待下一步的深入研究. ...
Stomatal clustering, a new marker for environmental perception and adaptation in terrestrial plants
1
2010
... 植物叶片对环境的响应首先是叶片表皮对环境的感应和变化(Pal et al ., 2002 ; Bargel et al ., 2004 ; Koch et al ., 2009 ; Adebowale et al ., 2014 ).生长于干燥的气候和土壤条件下的荒漠植物, 叶表皮细胞会变小, 切向壁加厚, 具有内皮层.叶表皮角质膜厚度增加, 角质层由光滑变得粗糙, 能够反射阳光, 减少叶片表面空气的流动, 降低蒸腾作用, 防止水分过度丧失(贺金生等, 1994 ; 杨明博等, 2007 ).气孔的分布特征、密度和面积等都会受到影响, 气孔多分布于叶片下表皮, 这种分布模式既可促进植物与外界环境进行气体交换, 又能保持水分.随着环境中水分和湿度的减少, 气孔密度增加, 气孔面积则向小型化发展, 气孔多下陷形成气孔窝或其上有突出的角质膜(Sam et al ., 2000 ; Bosabalidis & Kofidis, 2002 ; 高建平等, 2003 ; Xu & Zhou, 2008 ; Gan et al ., 2010 ; de Micco & Aronne, 2012 ).叶表皮附属物(如表皮毛和表面疣状突起物)也发生变化, 它们可有效地避免叶肉组织受损(Gairola et al ., 2009 ; Koch et al ., 2009 ; Gomes et al ., 2011 ; Noman et al ., 2012 ), 表皮蜡质(waxes)晶体形成二维或三维结构, 具有降低表皮的可湿性以及反射强光等作用(Wagner et al ., 2003 ; Koch & Ensikat, 2008 ). ...
不同产地华中五味子叶表皮结构和导管分子的解剖学特征及其与环境因子的关系
1
2003
... 植物叶片对环境的响应首先是叶片表皮对环境的感应和变化(Pal et al ., 2002 ; Bargel et al ., 2004 ; Koch et al ., 2009 ; Adebowale et al ., 2014 ).生长于干燥的气候和土壤条件下的荒漠植物, 叶表皮细胞会变小, 切向壁加厚, 具有内皮层.叶表皮角质膜厚度增加, 角质层由光滑变得粗糙, 能够反射阳光, 减少叶片表面空气的流动, 降低蒸腾作用, 防止水分过度丧失(贺金生等, 1994 ; 杨明博等, 2007 ).气孔的分布特征、密度和面积等都会受到影响, 气孔多分布于叶片下表皮, 这种分布模式既可促进植物与外界环境进行气体交换, 又能保持水分.随着环境中水分和湿度的减少, 气孔密度增加, 气孔面积则向小型化发展, 气孔多下陷形成气孔窝或其上有突出的角质膜(Sam et al ., 2000 ; Bosabalidis & Kofidis, 2002 ; 高建平等, 2003 ; Xu & Zhou, 2008 ; Gan et al ., 2010 ; de Micco & Aronne, 2012 ).叶表皮附属物(如表皮毛和表面疣状突起物)也发生变化, 它们可有效地避免叶肉组织受损(Gairola et al ., 2009 ; Koch et al ., 2009 ; Gomes et al ., 2011 ; Noman et al ., 2012 ), 表皮蜡质(waxes)晶体形成二维或三维结构, 具有降低表皮的可湿性以及反射强光等作用(Wagner et al ., 2003 ; Koch & Ensikat, 2008 ). ...
Leaf epidermal features ofSalix species (Salicaceae) and their systematic significance
2
2012
... 荒漠植物由于长期适应所生长的自然环境, 发展了一套最适宜自身生长发育的形态结构特征, 并采取各种途径来抵御或忍耐干旱胁迫对它的影响, 因而具有一定程度的耐旱性.这种耐旱性是一种复合性状, 包括植物的形态结构特征、水分生理特征、生理生化反应、组织细胞、光合器官以及原生质结构等特征的综合反映(Klich, 2000 ; Chaves et al ., 2003 ; Ogle & Reynolds, 2004 ).不同的荒漠植物对水分和不利环境的适应和耐受程度不同, 适应方式多种多样, 按照不同的分类标准, 荒漠植物有多种适应类型.通常的分类标准包括耐旱程度、耐旱方式、叶片形态、生活史、光合碳代谢途径等(赵哈林, 2012 ).各种类型的植物又有各自的适应特征.本研究是针对我国干旱半干旱区荒漠植物叶表皮微形态结构特征的综合分析, 主要集中于该区域的建群种灌木物种.根据荒漠植物叶表皮微形态基本结构特征和表皮附属物的类型与数量, 将荒漠植物表皮微形态类型直观地划分为11种, 而将抗逆适应微形态结构划分为6种类型.荒漠植物叶表皮微形态结构类型复杂, 反映了植物自身的遗传特性及其与环境的密切关系.植物叶片表皮微形态主要由遗传因素决定, 但表皮附属物绒毛数量, 蜡质形态与厚度, 气孔的形态、密度以及分布特征等与环境因子密切相关, 是植物对环境的响应(Barthlott, 1981 ; Casson & Hetherington, 2010 ; Ghahremaninejad et al ., 2012 ).因此, 基于这些表皮主要附属物特征对荒漠植物的表皮微形态类型和抗逆适应类型进行划分, 能够从微观形态结构特征进一步揭示物种的亲缘关系和表皮微形态结构的生态适应意义. ...
... 荒漠植物叶表皮微形态结构在不同分类等级上表现出丰富的多样性, 深刻地反映了植物的亲缘关系, 不同程度地揭示了它们之间错综复杂的系统演化关系.叶片表皮微形态结构基本特征根据表皮细胞形态结构和表皮附属物类型, 分为疣状、瘤状、皱褶状、相对光滑型、表面完全被毛、表面完全被蜡质层、不同形态结构的绒毛和蜡质层混合覆盖、上下完全异面以及气孔器集中于下表面等11种类型, 有些类型还包括若干种亚型.如柳属中观察的4种植物, 尽管种及地理分布完全不同, 但它们都为典型的异面叶, 叶片上下表皮都是完全不同的形态.上表皮绒毛密度较高, 不同的种有不同形态和密度的绒毛; 表皮绒毛密度低或不具绒毛, 但有柳属植物特有的角质膜蜡质纹饰(王东超等, 2012 ; Ghahremaninejad et al ., 2012 ), 如黄柳的痂状蜡质层、旱柳和乌柳的壳状蜡质层、沙柳的膜片状蜡质晶体.麻黄属的膜果麻黄、斑子麻黄(Ephedra rhytidosperma )和草麻黄(Ephedra sinica )中, 3种同化枝的角质膜蜡质晶体数量依次减少, 斑子麻黄与草麻黄的表皮形态结构接近, 显示亲缘关系较近.膜果麻黄表皮粗糙, 完全被蜡质层覆盖.锦鸡儿属(Caragana )是豆科蝶形花亚科中较大的一个属, 叶片上下表皮微形态基本一致, 多数为等面叶.但表皮毛的形态密度各异, 柠条锦鸡儿、小叶锦鸡儿(Caragana microphlla )和中间锦鸡儿(Caragana intermedia )同属小叶系, 亲缘关系相近, 表皮毛为柳叶状, 但绒毛密度不同, 柠条锦鸡儿表皮毛完全覆盖叶面, 小叶锦鸡儿和中间锦鸡儿表皮毛密度较小, 且它们的鳞片状蜡质晶体密度相似, 显示他们之间更近的亲缘关系, 这与其他分子生物学依据得到的结果不完全一致(Guo et al ., 2008 ).究其原因, 主要是因为这3种锦鸡儿属植物之间由于分布区域重叠或交叉, 因此存在复杂的基因交流(Ma et al ., 2003 ), 不同种的亲缘关系与其分布区域的其他种之间的相互杂交有关, 甚至出现了杂交过渡态类型, 因而给分类带来了难题, 单纯某一种分类依据是不能完全决定其亲缘关系的.荒漠锦鸡儿(Caragana roborovskyi )和卷叶锦鸡儿(Caragana ordosica )绒毛为棒状, 表皮细胞和蜡质层形态亦相似.短脚锦鸡儿(Caragana brachypoda )和狭叶锦鸡儿(Caragana stenophylla )表皮毛为柳叶带乳突型, 上表皮气孔器深陷, 气孔器为不等型或不规则型, 气孔口三角形或多边形.红花锦鸡儿(Caragana rosea )为明显不同的类群, 表皮无绒毛结构, 且蜡质层形态不同于以上几种的鳞片状, 而是紧贴表皮的膜片层结构或颗粒状. ...
Anatomical features and ultrastructure of Deschampsia antarctica(Poaceae) leaves from different growing habitats
1
2005
... 植物作为生态系统的第一生产者, 受环境变化的影响是直接而多方面的.叶片是植物进化过程中对环境变化最敏感且可塑性较大的器官, 在不同的选择压力下形成了各种适应类型(Hickey, 1984 ; 祁如虎等, 1998 ; Gielwanowska et al ., 2005 ), 其形态结构特征最能体现环境因子的影响或植物对环境的适应(王勋陵和王静, 1989 ; Krober et al ., 2015 ). ...
Ecophysiological and anatomical changes due to uptake and accumulation of heavy metal in Brachiaria decumbens
1
2011
... 植物叶片对环境的响应首先是叶片表皮对环境的感应和变化(Pal et al ., 2002 ; Bargel et al ., 2004 ; Koch et al ., 2009 ; Adebowale et al ., 2014 ).生长于干燥的气候和土壤条件下的荒漠植物, 叶表皮细胞会变小, 切向壁加厚, 具有内皮层.叶表皮角质膜厚度增加, 角质层由光滑变得粗糙, 能够反射阳光, 减少叶片表面空气的流动, 降低蒸腾作用, 防止水分过度丧失(贺金生等, 1994 ; 杨明博等, 2007 ).气孔的分布特征、密度和面积等都会受到影响, 气孔多分布于叶片下表皮, 这种分布模式既可促进植物与外界环境进行气体交换, 又能保持水分.随着环境中水分和湿度的减少, 气孔密度增加, 气孔面积则向小型化发展, 气孔多下陷形成气孔窝或其上有突出的角质膜(Sam et al ., 2000 ; Bosabalidis & Kofidis, 2002 ; 高建平等, 2003 ; Xu & Zhou, 2008 ; Gan et al ., 2010 ; de Micco & Aronne, 2012 ).叶表皮附属物(如表皮毛和表面疣状突起物)也发生变化, 它们可有效地避免叶肉组织受损(Gairola et al ., 2009 ; Koch et al ., 2009 ; Gomes et al ., 2011 ; Noman et al ., 2012 ), 表皮蜡质(waxes)晶体形成二维或三维结构, 具有降低表皮的可湿性以及反射强光等作用(Wagner et al ., 2003 ; Koch & Ensikat, 2008 ). ...
Effects of reduced irradiance on leaf morphology, photosynthetic capacity, and fruit yield in olive (Olea europaea L.)
1
2007
... 大多数荒漠植物叶片的一个显著特点是表皮被绒毛, 叶表皮毛的出现与长期的干旱气候有关, 是植物对干旱环境的适应特征(Retamales & Scharaschkin, 2015 ).植物叶片表皮毛分两种: 腺毛和非腺毛(Werker, 1993 ; Navarro & Oualidi, 1999 ).非腺毛的主要功能与植物的防御有关.目前确认的植物叶片表皮非腺毛具有的生理生态功能主要有: 减轻紫外辐射(Karabourniotis et al ., 1992 ; Gregoriou et al ., 2007 )、降低边界层的传导(Wuenscher, 1970 ; Kenzo et al ., 2008 )、降低叶温(Ehleringer & Mooney, 1978 ), 以及减少对太阳辐射的接收(Ehleringer et al ., 1976 )等.这些功能决定了干旱区荒漠植物抗逆能力的大小.干旱地区的叶表皮毛密度比湿润地区的高(Smith & Nobel, 1977 ; Aronne & de Micco, 2001 ), 就单个物种而言, 表皮毛密度与温度和辐射密切相关(Perez-Estrada et al ., 2000 ).在热带雨林树种中, 浓密的表皮毛很少见(Turner, 2001 ), Roth (1984)调查的250种树中仅有15种有较厚的表皮毛, 且较厚的表皮毛一般在干旱、高温和强辐射条件下产生.而在干旱半干旱的荒漠区, 大多数植物都具有浓密的表皮毛.显然, 荒漠植物叶上密生的表皮毛对于植物的生活有重要的意义.从Ehleringer等 (1976)、Ehleringer和Bjorkman (1978)、Ehleringer和Mooney (1978)研究的荒漠灌木Encelia farinosa 的叶表皮毛对叶片光谱特性和光合、蒸腾等生理活动的影响来看, 沙漠毒菊在不同生长季生长的叶片具有不同数量的表皮毛, 表皮毛的变化足够使叶片对光合有效辐射的吸收从81%降到29%, 对总的太阳辐射的吸收从46%降到16%, 而亲缘关系较近却没有表皮毛的同属物种Encelia californiea 的有效光合辐射吸收率为84%.同时, 由于对太阳辐射吸收的减少, 导致叶面温度和植物蒸腾速率的降低.荒漠环境气温经常超过40 ℃, 而植物最适宜的光合温度是25 ℃, 当温度超过35 ℃时植物的光合速率会突降, 但较厚的叶表皮毛能使叶温显著低于气温而接近光合适宜温度, 同时也避免植物达到叶片致死温度.但叶表皮毛也有不利的一面, 表皮毛的覆盖降低了光合速率, 因此同一植物在不同的生境和生长季长出的叶片会出现表皮毛数量不同的特征, 这也是植物适应环境的一种权衡策略(Kenzo et al ., 2008 ).从本研究中的117种荒漠植物叶表皮特征来看, 片状叶片基本被有表皮毛, 并有不少叶片的表皮毛数量较多, 直接增加了叶片的边界层厚度, 个别植物的上下表皮毛厚度远超于叶片厚度, 如表皮微形态抗逆适应主要类型的第一类植物中的柠条锦鸡儿、卷叶锦鸡儿和沙枣等.可见荒漠植物中叶片表皮毛的作用是相当重要的, 它们不仅能调节植物的生理功能, 而且可抵御风沙等机械损伤的直接伤害.这些物种在不同的自然生境中其表皮毛数量亦不同(数据省略), 说明生态环境深刻地影响着植物形态结构的形成与变异. ...
Genetic diversity and genetic relationship of Caragana microphylla, Caragana davazamcii and Caragana korshinskii on the Inner Mongolia Plateau, China
1
2008
... 荒漠植物叶表皮微形态结构在不同分类等级上表现出丰富的多样性, 深刻地反映了植物的亲缘关系, 不同程度地揭示了它们之间错综复杂的系统演化关系.叶片表皮微形态结构基本特征根据表皮细胞形态结构和表皮附属物类型, 分为疣状、瘤状、皱褶状、相对光滑型、表面完全被毛、表面完全被蜡质层、不同形态结构的绒毛和蜡质层混合覆盖、上下完全异面以及气孔器集中于下表面等11种类型, 有些类型还包括若干种亚型.如柳属中观察的4种植物, 尽管种及地理分布完全不同, 但它们都为典型的异面叶, 叶片上下表皮都是完全不同的形态.上表皮绒毛密度较高, 不同的种有不同形态和密度的绒毛; 表皮绒毛密度低或不具绒毛, 但有柳属植物特有的角质膜蜡质纹饰(王东超等, 2012 ; Ghahremaninejad et al ., 2012 ), 如黄柳的痂状蜡质层、旱柳和乌柳的壳状蜡质层、沙柳的膜片状蜡质晶体.麻黄属的膜果麻黄、斑子麻黄(Ephedra rhytidosperma )和草麻黄(Ephedra sinica )中, 3种同化枝的角质膜蜡质晶体数量依次减少, 斑子麻黄与草麻黄的表皮形态结构接近, 显示亲缘关系较近.膜果麻黄表皮粗糙, 完全被蜡质层覆盖.锦鸡儿属(Caragana )是豆科蝶形花亚科中较大的一个属, 叶片上下表皮微形态基本一致, 多数为等面叶.但表皮毛的形态密度各异, 柠条锦鸡儿、小叶锦鸡儿(Caragana microphlla )和中间锦鸡儿(Caragana intermedia )同属小叶系, 亲缘关系相近, 表皮毛为柳叶状, 但绒毛密度不同, 柠条锦鸡儿表皮毛完全覆盖叶面, 小叶锦鸡儿和中间锦鸡儿表皮毛密度较小, 且它们的鳞片状蜡质晶体密度相似, 显示他们之间更近的亲缘关系, 这与其他分子生物学依据得到的结果不完全一致(Guo et al ., 2008 ).究其原因, 主要是因为这3种锦鸡儿属植物之间由于分布区域重叠或交叉, 因此存在复杂的基因交流(Ma et al ., 2003 ), 不同种的亲缘关系与其分布区域的其他种之间的相互杂交有关, 甚至出现了杂交过渡态类型, 因而给分类带来了难题, 单纯某一种分类依据是不能完全决定其亲缘关系的.荒漠锦鸡儿(Caragana roborovskyi )和卷叶锦鸡儿(Caragana ordosica )绒毛为棒状, 表皮细胞和蜡质层形态亦相似.短脚锦鸡儿(Caragana brachypoda )和狭叶锦鸡儿(Caragana stenophylla )表皮毛为柳叶带乳突型, 上表皮气孔器深陷, 气孔器为不等型或不规则型, 气孔口三角形或多边形.红花锦鸡儿(Caragana rosea )为明显不同的类群, 表皮无绒毛结构, 且蜡质层形态不同于以上几种的鳞片状, 而是紧贴表皮的膜片层结构或颗粒状. ...
高山栎叶的形态结构及其与生态环境的关系
1
1994
... 植物叶片对环境的响应首先是叶片表皮对环境的感应和变化(Pal et al ., 2002 ; Bargel et al ., 2004 ; Koch et al ., 2009 ; Adebowale et al ., 2014 ).生长于干燥的气候和土壤条件下的荒漠植物, 叶表皮细胞会变小, 切向壁加厚, 具有内皮层.叶表皮角质膜厚度增加, 角质层由光滑变得粗糙, 能够反射阳光, 减少叶片表面空气的流动, 降低蒸腾作用, 防止水分过度丧失(贺金生等, 1994 ; 杨明博等, 2007 ).气孔的分布特征、密度和面积等都会受到影响, 气孔多分布于叶片下表皮, 这种分布模式既可促进植物与外界环境进行气体交换, 又能保持水分.随着环境中水分和湿度的减少, 气孔密度增加, 气孔面积则向小型化发展, 气孔多下陷形成气孔窝或其上有突出的角质膜(Sam et al ., 2000 ; Bosabalidis & Kofidis, 2002 ; 高建平等, 2003 ; Xu & Zhou, 2008 ; Gan et al ., 2010 ; de Micco & Aronne, 2012 ).叶表皮附属物(如表皮毛和表面疣状突起物)也发生变化, 它们可有效地避免叶肉组织受损(Gairola et al ., 2009 ; Koch et al ., 2009 ; Gomes et al ., 2011 ; Noman et al ., 2012 ), 表皮蜡质(waxes)晶体形成二维或三维结构, 具有降低表皮的可湿性以及反射强光等作用(Wagner et al ., 2003 ; Koch & Ensikat, 2008 ). ...
Changes in the angiosperm flora across the cretaceus tertiary boundary
1
1984
... 植物作为生态系统的第一生产者, 受环境变化的影响是直接而多方面的.叶片是植物进化过程中对环境变化最敏感且可塑性较大的器官, 在不同的选择压力下形成了各种适应类型(Hickey, 1984 ; 祁如虎等, 1998 ; Gielwanowska et al ., 2005 ), 其形态结构特征最能体现环境因子的影响或植物对环境的适应(王勋陵和王静, 1989 ; Krober et al ., 2015 ). ...
Effects of pubescence and waxes on the reflectance of leaves in the ultraviolet and photosynthetic wavebands: A comparison of a range of species
1
2002
... 荒漠植物叶片的另一个显著特点是表皮有较厚的角质膜蜡质层.角质膜蜡质层与表皮毛有共同的功能, 是植物表皮的保护屏障, 主要由长链碳氢化合物组成(Shepherd & Wynne, 2006 ).叶片对光的反射由叶面形貌决定, 表皮毛和蜡质层是决定荒漠植物叶表面形貌的关键.Holmes和Keiller (2002)研究了45种桉属和伽蓝菜属植物的蜡质叶, 发现蜡质层对紫外光(330 nm)和光合有效辐射(680 nm)的反射比各种形态的表皮毛更强, 去除蜡质层通常会减少对这两种光的反射, 特别是对紫外光的反射, 但对表皮无蜡质的叶片影响不大.同时不同的物种对不同波长的光的反射亦不同, 表明蜡质晶体的形态直接影响着对光的反射, 并且不同温度下的反射强度也不同, 可见蜡质层对光的反射特性变化很大(Holmes & Keiller, 2002 ).叶表皮蜡质层参与植物对水分、盐、低温、污染、机械损伤等多种胁迫响应的过程, 提高植物对胁迫的抗性(Cameron, 1970 ; Bargel et al ., 2004 ; Perrone et al ., 2013 , 2015 ).因此, 本研究中117种荒漠植物叶片表皮颗粒状、膜片状、晶簇状、鳞片状、杆状等各种不同形态的蜡质晶体既是植物长期适应环境的结果, 也对荒漠植物在严酷环境中的生长和生存至关重要.总的来说, 这些植物中灌木的蜡质层密度高于草本, 无表皮毛的叶片蜡质层密度高于表皮毛覆盖的, 特别是以同化枝为主要光合器官和棒状叶片的植物, 表皮基本没有表皮毛覆盖, 主要的附属物是蜡质层, 如麻黄属、沙拐枣属、红砂属的植物等, 弥补了表皮毛的功能. ...
Leaf anatomical modifications in Catharanthus roseus as affected by plant growth promoters and retardants
1
2009
... 植物叶片适应环境的形态结构变化是多方面的.在众多生态因子中, 叶片受水分、温度和光因子的影响最为显著(王勋陵和王静, 1989 ; 王勋陵, 1993 ; Aronne & de Micco 2001 ; Arena et al ., 2008 ; Jaleel et al ., 2009a , 2009b ), 主要表现为叶形的变化、叶片表皮附属物的变化、叶片的厚度及解剖结构的差异(Talukdar, 2015 ).对于荒漠植物来说, 夏季极端干旱时它们可能同时遭受以上各种环境因子的胁迫.但其最主要的限制因子为水分, 因而表现出典型的旱生植物形态结构特征.由于遗传因子与环境因子的共同作用, 在旱生植物结构的适应性上出现了两种演化趋势, 即趋同性和趋异性, 也就是说, 在相同环境下的植物演化出相似的形态结构适应性, 而不同环境下的同类植物演化出不同的形态结构(Lakusic et al ., 2010 ).因此, 要系统分析荒漠植物叶片形态结构特征的适应性变化, 需要将一个区域不同植物和不同梯度生境下的同一植物做全面分析. ...
Drought stress in plants: A review on morphological characteristics and pigments composition
1
2009
... 植物叶片适应环境的形态结构变化是多方面的.在众多生态因子中, 叶片受水分、温度和光因子的影响最为显著(王勋陵和王静, 1989 ; 王勋陵, 1993 ; Aronne & de Micco 2001 ; Arena et al ., 2008 ; Jaleel et al ., 2009a , 2009b ), 主要表现为叶形的变化、叶片表皮附属物的变化、叶片的厚度及解剖结构的差异(Talukdar, 2015 ).对于荒漠植物来说, 夏季极端干旱时它们可能同时遭受以上各种环境因子的胁迫.但其最主要的限制因子为水分, 因而表现出典型的旱生植物形态结构特征.由于遗传因子与环境因子的共同作用, 在旱生植物结构的适应性上出现了两种演化趋势, 即趋同性和趋异性, 也就是说, 在相同环境下的植物演化出相似的形态结构适应性, 而不同环境下的同类植物演化出不同的形态结构(Lakusic et al ., 2010 ).因此, 要系统分析荒漠植物叶片形态结构特征的适应性变化, 需要将一个区域不同植物和不同梯度生境下的同一植物做全面分析. ...
Ultraviolet-B radiation absorbing capacity of leaf hairs
1
1992
... 大多数荒漠植物叶片的一个显著特点是表皮被绒毛, 叶表皮毛的出现与长期的干旱气候有关, 是植物对干旱环境的适应特征(Retamales & Scharaschkin, 2015 ).植物叶片表皮毛分两种: 腺毛和非腺毛(Werker, 1993 ; Navarro & Oualidi, 1999 ).非腺毛的主要功能与植物的防御有关.目前确认的植物叶片表皮非腺毛具有的生理生态功能主要有: 减轻紫外辐射(Karabourniotis et al ., 1992 ; Gregoriou et al ., 2007 )、降低边界层的传导(Wuenscher, 1970 ; Kenzo et al ., 2008 )、降低叶温(Ehleringer & Mooney, 1978 ), 以及减少对太阳辐射的接收(Ehleringer et al ., 1976 )等.这些功能决定了干旱区荒漠植物抗逆能力的大小.干旱地区的叶表皮毛密度比湿润地区的高(Smith & Nobel, 1977 ; Aronne & de Micco, 2001 ), 就单个物种而言, 表皮毛密度与温度和辐射密切相关(Perez-Estrada et al ., 2000 ).在热带雨林树种中, 浓密的表皮毛很少见(Turner, 2001 ), Roth (1984)调查的250种树中仅有15种有较厚的表皮毛, 且较厚的表皮毛一般在干旱、高温和强辐射条件下产生.而在干旱半干旱的荒漠区, 大多数植物都具有浓密的表皮毛.显然, 荒漠植物叶上密生的表皮毛对于植物的生活有重要的意义.从Ehleringer等 (1976)、Ehleringer和Bjorkman (1978)、Ehleringer和Mooney (1978)研究的荒漠灌木Encelia farinosa 的叶表皮毛对叶片光谱特性和光合、蒸腾等生理活动的影响来看, 沙漠毒菊在不同生长季生长的叶片具有不同数量的表皮毛, 表皮毛的变化足够使叶片对光合有效辐射的吸收从81%降到29%, 对总的太阳辐射的吸收从46%降到16%, 而亲缘关系较近却没有表皮毛的同属物种Encelia californiea 的有效光合辐射吸收率为84%.同时, 由于对太阳辐射吸收的减少, 导致叶面温度和植物蒸腾速率的降低.荒漠环境气温经常超过40 ℃, 而植物最适宜的光合温度是25 ℃, 当温度超过35 ℃时植物的光合速率会突降, 但较厚的叶表皮毛能使叶温显著低于气温而接近光合适宜温度, 同时也避免植物达到叶片致死温度.但叶表皮毛也有不利的一面, 表皮毛的覆盖降低了光合速率, 因此同一植物在不同的生境和生长季长出的叶片会出现表皮毛数量不同的特征, 这也是植物适应环境的一种权衡策略(Kenzo et al ., 2008 ).从本研究中的117种荒漠植物叶表皮特征来看, 片状叶片基本被有表皮毛, 并有不少叶片的表皮毛数量较多, 直接增加了叶片的边界层厚度, 个别植物的上下表皮毛厚度远超于叶片厚度, 如表皮微形态抗逆适应主要类型的第一类植物中的柠条锦鸡儿、卷叶锦鸡儿和沙枣等.可见荒漠植物中叶片表皮毛的作用是相当重要的, 它们不仅能调节植物的生理功能, 而且可抵御风沙等机械损伤的直接伤害.这些物种在不同的自然生境中其表皮毛数量亦不同(数据省略), 说明生态环境深刻地影响着植物形态结构的形成与变异. ...
Changes in leaf water use after removal of leaf lower surface hairs onMallotus macrostachyus(Euphorbiaceae) in a tropical secondary forest in Malaysia
2
2008
... 大多数荒漠植物叶片的一个显著特点是表皮被绒毛, 叶表皮毛的出现与长期的干旱气候有关, 是植物对干旱环境的适应特征(Retamales & Scharaschkin, 2015 ).植物叶片表皮毛分两种: 腺毛和非腺毛(Werker, 1993 ; Navarro & Oualidi, 1999 ).非腺毛的主要功能与植物的防御有关.目前确认的植物叶片表皮非腺毛具有的生理生态功能主要有: 减轻紫外辐射(Karabourniotis et al ., 1992 ; Gregoriou et al ., 2007 )、降低边界层的传导(Wuenscher, 1970 ; Kenzo et al ., 2008 )、降低叶温(Ehleringer & Mooney, 1978 ), 以及减少对太阳辐射的接收(Ehleringer et al ., 1976 )等.这些功能决定了干旱区荒漠植物抗逆能力的大小.干旱地区的叶表皮毛密度比湿润地区的高(Smith & Nobel, 1977 ; Aronne & de Micco, 2001 ), 就单个物种而言, 表皮毛密度与温度和辐射密切相关(Perez-Estrada et al ., 2000 ).在热带雨林树种中, 浓密的表皮毛很少见(Turner, 2001 ), Roth (1984)调查的250种树中仅有15种有较厚的表皮毛, 且较厚的表皮毛一般在干旱、高温和强辐射条件下产生.而在干旱半干旱的荒漠区, 大多数植物都具有浓密的表皮毛.显然, 荒漠植物叶上密生的表皮毛对于植物的生活有重要的意义.从Ehleringer等 (1976)、Ehleringer和Bjorkman (1978)、Ehleringer和Mooney (1978)研究的荒漠灌木Encelia farinosa 的叶表皮毛对叶片光谱特性和光合、蒸腾等生理活动的影响来看, 沙漠毒菊在不同生长季生长的叶片具有不同数量的表皮毛, 表皮毛的变化足够使叶片对光合有效辐射的吸收从81%降到29%, 对总的太阳辐射的吸收从46%降到16%, 而亲缘关系较近却没有表皮毛的同属物种Encelia californiea 的有效光合辐射吸收率为84%.同时, 由于对太阳辐射吸收的减少, 导致叶面温度和植物蒸腾速率的降低.荒漠环境气温经常超过40 ℃, 而植物最适宜的光合温度是25 ℃, 当温度超过35 ℃时植物的光合速率会突降, 但较厚的叶表皮毛能使叶温显著低于气温而接近光合适宜温度, 同时也避免植物达到叶片致死温度.但叶表皮毛也有不利的一面, 表皮毛的覆盖降低了光合速率, 因此同一植物在不同的生境和生长季长出的叶片会出现表皮毛数量不同的特征, 这也是植物适应环境的一种权衡策略(Kenzo et al ., 2008 ).从本研究中的117种荒漠植物叶表皮特征来看, 片状叶片基本被有表皮毛, 并有不少叶片的表皮毛数量较多, 直接增加了叶片的边界层厚度, 个别植物的上下表皮毛厚度远超于叶片厚度, 如表皮微形态抗逆适应主要类型的第一类植物中的柠条锦鸡儿、卷叶锦鸡儿和沙枣等.可见荒漠植物中叶片表皮毛的作用是相当重要的, 它们不仅能调节植物的生理功能, 而且可抵御风沙等机械损伤的直接伤害.这些物种在不同的自然生境中其表皮毛数量亦不同(数据省略), 说明生态环境深刻地影响着植物形态结构的形成与变异. ...
... 的有效光合辐射吸收率为84%.同时, 由于对太阳辐射吸收的减少, 导致叶面温度和植物蒸腾速率的降低.荒漠环境气温经常超过40 ℃, 而植物最适宜的光合温度是25 ℃, 当温度超过35 ℃时植物的光合速率会突降, 但较厚的叶表皮毛能使叶温显著低于气温而接近光合适宜温度, 同时也避免植物达到叶片致死温度.但叶表皮毛也有不利的一面, 表皮毛的覆盖降低了光合速率, 因此同一植物在不同的生境和生长季长出的叶片会出现表皮毛数量不同的特征, 这也是植物适应环境的一种权衡策略(Kenzo et al ., 2008 ).从本研究中的117种荒漠植物叶表皮特征来看, 片状叶片基本被有表皮毛, 并有不少叶片的表皮毛数量较多, 直接增加了叶片的边界层厚度, 个别植物的上下表皮毛厚度远超于叶片厚度, 如表皮微形态抗逆适应主要类型的第一类植物中的柠条锦鸡儿、卷叶锦鸡儿和沙枣等.可见荒漠植物中叶片表皮毛的作用是相当重要的, 它们不仅能调节植物的生理功能, 而且可抵御风沙等机械损伤的直接伤害.这些物种在不同的自然生境中其表皮毛数量亦不同(数据省略), 说明生态环境深刻地影响着植物形态结构的形成与变异. ...
Leaf variations in Elaeagnus angustifolia related to environmental heterogeneity
1
2000
... 荒漠植物由于长期适应所生长的自然环境, 发展了一套最适宜自身生长发育的形态结构特征, 并采取各种途径来抵御或忍耐干旱胁迫对它的影响, 因而具有一定程度的耐旱性.这种耐旱性是一种复合性状, 包括植物的形态结构特征、水分生理特征、生理生化反应、组织细胞、光合器官以及原生质结构等特征的综合反映(Klich, 2000 ; Chaves et al ., 2003 ; Ogle & Reynolds, 2004 ).不同的荒漠植物对水分和不利环境的适应和耐受程度不同, 适应方式多种多样, 按照不同的分类标准, 荒漠植物有多种适应类型.通常的分类标准包括耐旱程度、耐旱方式、叶片形态、生活史、光合碳代谢途径等(赵哈林, 2012 ).各种类型的植物又有各自的适应特征.本研究是针对我国干旱半干旱区荒漠植物叶表皮微形态结构特征的综合分析, 主要集中于该区域的建群种灌木物种.根据荒漠植物叶表皮微形态基本结构特征和表皮附属物的类型与数量, 将荒漠植物表皮微形态类型直观地划分为11种, 而将抗逆适应微形态结构划分为6种类型.荒漠植物叶表皮微形态结构类型复杂, 反映了植物自身的遗传特性及其与环境的密切关系.植物叶片表皮微形态主要由遗传因素决定, 但表皮附属物绒毛数量, 蜡质形态与厚度, 气孔的形态、密度以及分布特征等与环境因子密切相关, 是植物对环境的响应(Barthlott, 1981 ; Casson & Hetherington, 2010 ; Ghahremaninejad et al ., 2012 ).因此, 基于这些表皮主要附属物特征对荒漠植物的表皮微形态类型和抗逆适应类型进行划分, 能够从微观形态结构特征进一步揭示物种的亲缘关系和表皮微形态结构的生态适应意义. ...
Multifunctional surface structures of plants: An inspiration for biomimetics
2
2009
... 植物叶片对环境的响应首先是叶片表皮对环境的感应和变化(Pal et al ., 2002 ; Bargel et al ., 2004 ; Koch et al ., 2009 ; Adebowale et al ., 2014 ).生长于干燥的气候和土壤条件下的荒漠植物, 叶表皮细胞会变小, 切向壁加厚, 具有内皮层.叶表皮角质膜厚度增加, 角质层由光滑变得粗糙, 能够反射阳光, 减少叶片表面空气的流动, 降低蒸腾作用, 防止水分过度丧失(贺金生等, 1994 ; 杨明博等, 2007 ).气孔的分布特征、密度和面积等都会受到影响, 气孔多分布于叶片下表皮, 这种分布模式既可促进植物与外界环境进行气体交换, 又能保持水分.随着环境中水分和湿度的减少, 气孔密度增加, 气孔面积则向小型化发展, 气孔多下陷形成气孔窝或其上有突出的角质膜(Sam et al ., 2000 ; Bosabalidis & Kofidis, 2002 ; 高建平等, 2003 ; Xu & Zhou, 2008 ; Gan et al ., 2010 ; de Micco & Aronne, 2012 ).叶表皮附属物(如表皮毛和表面疣状突起物)也发生变化, 它们可有效地避免叶肉组织受损(Gairola et al ., 2009 ; Koch et al ., 2009 ; Gomes et al ., 2011 ; Noman et al ., 2012 ), 表皮蜡质(waxes)晶体形成二维或三维结构, 具有降低表皮的可湿性以及反射强光等作用(Wagner et al ., 2003 ; Koch & Ensikat, 2008 ). ...
... ; Koch et al ., 2009 ; Gomes et al ., 2011 ; Noman et al ., 2012 ), 表皮蜡质(waxes)晶体形成二维或三维结构, 具有降低表皮的可湿性以及反射强光等作用(Wagner et al ., 2003 ; Koch & Ensikat, 2008 ). ...
The hydrophobic coatings of plant surfaces: Epicuticular wax crystals and their morphologies, crystallinity and molecular self-assembly
1
2008
... 植物叶片对环境的响应首先是叶片表皮对环境的感应和变化(Pal et al ., 2002 ; Bargel et al ., 2004 ; Koch et al ., 2009 ; Adebowale et al ., 2014 ).生长于干燥的气候和土壤条件下的荒漠植物, 叶表皮细胞会变小, 切向壁加厚, 具有内皮层.叶表皮角质膜厚度增加, 角质层由光滑变得粗糙, 能够反射阳光, 减少叶片表面空气的流动, 降低蒸腾作用, 防止水分过度丧失(贺金生等, 1994 ; 杨明博等, 2007 ).气孔的分布特征、密度和面积等都会受到影响, 气孔多分布于叶片下表皮, 这种分布模式既可促进植物与外界环境进行气体交换, 又能保持水分.随着环境中水分和湿度的减少, 气孔密度增加, 气孔面积则向小型化发展, 气孔多下陷形成气孔窝或其上有突出的角质膜(Sam et al ., 2000 ; Bosabalidis & Kofidis, 2002 ; 高建平等, 2003 ; Xu & Zhou, 2008 ; Gan et al ., 2010 ; de Micco & Aronne, 2012 ).叶表皮附属物(如表皮毛和表面疣状突起物)也发生变化, 它们可有效地避免叶肉组织受损(Gairola et al ., 2009 ; Koch et al ., 2009 ; Gomes et al ., 2011 ; Noman et al ., 2012 ), 表皮蜡质(waxes)晶体形成二维或三维结构, 具有降低表皮的可湿性以及反射强光等作用(Wagner et al ., 2003 ; Koch & Ensikat, 2008 ). ...
Comparative morphology of leaf epidermis in the Chloranthaceae
1
2001
... 植物叶片表皮微形态特征的多样性不仅在很大程度上表达了遗传的多样性和稳定性, 而且具有重要的系统学和分类学意义, 有助于一些分类问题的解决.近年来不少研究发现叶表皮性状在一定程度上能反映分类群间的系统发生, 可用于科下属间关系的探讨(Kong et al ., 2001 ).以骆驼蓬属为例, 1889年Maxim将多裂骆驼蓬定为骆驼蓬(Peganum harmala )的一个变种Peganum harmala var. multisectum , 1949年Bobr却将其定为独立的种(中国科学院中国植物志编辑委员会, 1998),《中国沙漠植物志》亦将之作为种(刘瑛心, 1987 ), 但国内外一些学者仍倾向于将其作为骆驼蓬的一个变种.马骥等(1997, 2003)结合种子微形态结构和有关化学成分、地理分布等方面的研究结果, 支持将多裂骆驼蓬作为一个独立的种处理的观点.叶片表皮微形态实验结果表明: 从粗略的叶表皮微形态基本特征统计来看, 二者基本特征相同, 但骆驼蓬表皮细胞呈褶皱状, 而多裂骆驼蓬叶表皮基本光滑, 有明显的区别, 赞成将二者作为不同的独立种的观点. ...
1
2014
... 而对于腺毛来说, 其主要作为一个油性物质的合成、分泌和累积的器官, 有效地防御动物的捕食、病菌入侵以及物种竞争(Werker, 1993 ; Wagner et al ., 2003 ; Kremer et al ., 2014 ; Munien et al ., 2015 ).实际上, 腺毛也是次生代谢产物的一个独立的细胞工厂(Tissier, 2012 ).腺毛的形态随植物的组织器官和种类不同而呈多样化, 其形成和分泌的主要油性物质及其产物组成受遗传和环境两方面的因素控制(Bosabalidis & Kofidis, 2002 ; Shafiee-Hajiabad et al ., 2014 ), 腺毛结构的多样性也可以为植物分类学提供依据(Gairola et al ., 2009 ; Rusydi et al ., 2013 ).对117种荒漠植物叶片表皮扫描发现: 不少旱生植物除具有非腺体表皮毛外, 还同时具有腺毛, 并且有些腺毛体积较大, 如蒙古莸、甘草、香青兰、星毛短舌菊、百里香等.显然, 对于具有腺毛的荒漠植物, 腺毛具有重要的功能.但目前关于不同植物腺毛的形态、形成以及功能的研究还很有局限性.那么, 这些腺毛在植物适应荒漠环境的过程中其形态结构与环境因子有何关系、主要分泌哪些化学物质、发挥什么样的功能?这些问题亟待下一步的深入研究. ...
Leaf morphology of 40 evergreen and deciduous broad leaved subtropical tree species and relationships to functional ecophysiological traits
1
2015
... 植物作为生态系统的第一生产者, 受环境变化的影响是直接而多方面的.叶片是植物进化过程中对环境变化最敏感且可塑性较大的器官, 在不同的选择压力下形成了各种适应类型(Hickey, 1984 ; 祁如虎等, 1998 ; Gielwanowska et al ., 2005 ), 其形态结构特征最能体现环境因子的影响或植物对环境的适应(王勋陵和王静, 1989 ; Krober et al ., 2015 ). ...
Habitat- related adaptations in morphology and anatomy of Teucrium(Lamiaceae) species from the Balkan Peninsula (Serbia and Montenegro)
1
2010
... 植物叶片适应环境的形态结构变化是多方面的.在众多生态因子中, 叶片受水分、温度和光因子的影响最为显著(王勋陵和王静, 1989 ; 王勋陵, 1993 ; Aronne & de Micco 2001 ; Arena et al ., 2008 ; Jaleel et al ., 2009a , 2009b ), 主要表现为叶形的变化、叶片表皮附属物的变化、叶片的厚度及解剖结构的差异(Talukdar, 2015 ).对于荒漠植物来说, 夏季极端干旱时它们可能同时遭受以上各种环境因子的胁迫.但其最主要的限制因子为水分, 因而表现出典型的旱生植物形态结构特征.由于遗传因子与环境因子的共同作用, 在旱生植物结构的适应性上出现了两种演化趋势, 即趋同性和趋异性, 也就是说, 在相同环境下的植物演化出相似的形态结构适应性, 而不同环境下的同类植物演化出不同的形态结构(Lakusic et al ., 2010 ).因此, 要系统分析荒漠植物叶片形态结构特征的适应性变化, 需要将一个区域不同植物和不同梯度生境下的同一植物做全面分析. ...
Epidermal micromorphology and mesophyll structure of Populus euphratica heteromorphic leaves at different development stages
1
2015
... 干旱半干旱区的荒漠环境属于一种极端环境, 降水稀少, 气候干燥, 昼夜温差大, 地面温度高, 土壤贫瘠, 风沙活动强烈……这些恶劣条件无时不在限制着荒漠植物的生存和繁衍.但即使在这种条件下, 仍有许多植物分布于干旱半干旱区, 且能很好地繁衍生息.那么, 这些植物有何特殊的形态结构来适应环境呢?如何通过植物形态结构特征判断其适应能力? 这些问题引发不少科学家对荒漠植物研究的极大关注.Schow提出“旱生植物”概念已有100多年了. 国内外不少学者研究过旱生植物的形态结构(Shield, 1950 ; 王勋陵和马骥, 1999 ; 严巧娣等, 2008 ; Liu et al ., 2012 , 2015 ; Perrone et al ., 2015 ), 但以各自有限数量的旱生植物形态结构描述为主, 至今未见对较大数量旱生植物叶片表皮微形态与结构特征进行数量化整理, 并对其类型划分、变化规律、演化趋势、适应对策等重要问题进行系统讨论的 报道. ...
Responses of three different ecotypes of reed (Phragmites communis Trin.) to their natural habitats: Leaf surface micro-morphology, anatomy, chloroplast ultrastructure and physio-chemical characteristics
2
2012
... 干旱半干旱区的荒漠环境属于一种极端环境, 降水稀少, 气候干燥, 昼夜温差大, 地面温度高, 土壤贫瘠, 风沙活动强烈……这些恶劣条件无时不在限制着荒漠植物的生存和繁衍.但即使在这种条件下, 仍有许多植物分布于干旱半干旱区, 且能很好地繁衍生息.那么, 这些植物有何特殊的形态结构来适应环境呢?如何通过植物形态结构特征判断其适应能力? 这些问题引发不少科学家对荒漠植物研究的极大关注.Schow提出“旱生植物”概念已有100多年了. 国内外不少学者研究过旱生植物的形态结构(Shield, 1950 ; 王勋陵和马骥, 1999 ; 严巧娣等, 2008 ; Liu et al ., 2012 , 2015 ; Perrone et al ., 2015 ), 但以各自有限数量的旱生植物形态结构描述为主, 至今未见对较大数量旱生植物叶片表皮微形态与结构特征进行数量化整理, 并对其类型划分、变化规律、演化趋势、适应对策等重要问题进行系统讨论的 报道. ...
... 扫描电镜材料的制备参考Liu等(2012) 的方法, 制备过程如下: 将固定24 h后的材料经磷酸缓冲液(0.1 mol·L-1 , pH 7.2)清洗2次, 各10 min, 然后用1.0% OsO4 磷酸缓冲液进行最后固定过夜.过夜后的材料用以上磷酸缓冲液清洗后, 在不同浓度的乙醇(30%、50%、70%、96%、100%)中进行脱水, 乙醇浓度逐渐增加.然后在干燥器JFD-310中干燥后, 喷20 nm厚的胶体金, 用扫描电镜(JSM-6380, JEOL, Tokyo, Japan)观察叶上、下表皮微形态特征并进行拍照. ...
1
1987
... 植物叶片表皮微形态特征的多样性不仅在很大程度上表达了遗传的多样性和稳定性, 而且具有重要的系统学和分类学意义, 有助于一些分类问题的解决.近年来不少研究发现叶表皮性状在一定程度上能反映分类群间的系统发生, 可用于科下属间关系的探讨(Kong et al ., 2001 ).以骆驼蓬属为例, 1889年Maxim将多裂骆驼蓬定为骆驼蓬(Peganum harmala )的一个变种Peganum harmala var. multisectum , 1949年Bobr却将其定为独立的种(中国科学院中国植物志编辑委员会, 1998),《中国沙漠植物志》亦将之作为种(刘瑛心, 1987 ), 但国内外一些学者仍倾向于将其作为骆驼蓬的一个变种.马骥等(1997, 2003)结合种子微形态结构和有关化学成分、地理分布等方面的研究结果, 支持将多裂骆驼蓬作为一个独立的种处理的观点.叶片表皮微形态实验结果表明: 从粗略的叶表皮微形态基本特征统计来看, 二者基本特征相同, 但骆驼蓬表皮细胞呈褶皱状, 而多裂骆驼蓬叶表皮基本光滑, 有明显的区别, 赞成将二者作为不同的独立种的观点. ...
Interspecific transition among Caragana microphylla, C. davazamcii and C. korshinskii along geographic gradient I. Ecological and RAPD evidence
1
2003
... 荒漠植物叶表皮微形态结构在不同分类等级上表现出丰富的多样性, 深刻地反映了植物的亲缘关系, 不同程度地揭示了它们之间错综复杂的系统演化关系.叶片表皮微形态结构基本特征根据表皮细胞形态结构和表皮附属物类型, 分为疣状、瘤状、皱褶状、相对光滑型、表面完全被毛、表面完全被蜡质层、不同形态结构的绒毛和蜡质层混合覆盖、上下完全异面以及气孔器集中于下表面等11种类型, 有些类型还包括若干种亚型.如柳属中观察的4种植物, 尽管种及地理分布完全不同, 但它们都为典型的异面叶, 叶片上下表皮都是完全不同的形态.上表皮绒毛密度较高, 不同的种有不同形态和密度的绒毛; 表皮绒毛密度低或不具绒毛, 但有柳属植物特有的角质膜蜡质纹饰(王东超等, 2012 ; Ghahremaninejad et al ., 2012 ), 如黄柳的痂状蜡质层、旱柳和乌柳的壳状蜡质层、沙柳的膜片状蜡质晶体.麻黄属的膜果麻黄、斑子麻黄(Ephedra rhytidosperma )和草麻黄(Ephedra sinica )中, 3种同化枝的角质膜蜡质晶体数量依次减少, 斑子麻黄与草麻黄的表皮形态结构接近, 显示亲缘关系较近.膜果麻黄表皮粗糙, 完全被蜡质层覆盖.锦鸡儿属(Caragana )是豆科蝶形花亚科中较大的一个属, 叶片上下表皮微形态基本一致, 多数为等面叶.但表皮毛的形态密度各异, 柠条锦鸡儿、小叶锦鸡儿(Caragana microphlla )和中间锦鸡儿(Caragana intermedia )同属小叶系, 亲缘关系相近, 表皮毛为柳叶状, 但绒毛密度不同, 柠条锦鸡儿表皮毛完全覆盖叶面, 小叶锦鸡儿和中间锦鸡儿表皮毛密度较小, 且它们的鳞片状蜡质晶体密度相似, 显示他们之间更近的亲缘关系, 这与其他分子生物学依据得到的结果不完全一致(Guo et al ., 2008 ).究其原因, 主要是因为这3种锦鸡儿属植物之间由于分布区域重叠或交叉, 因此存在复杂的基因交流(Ma et al ., 2003 ), 不同种的亲缘关系与其分布区域的其他种之间的相互杂交有关, 甚至出现了杂交过渡态类型, 因而给分类带来了难题, 单纯某一种分类依据是不能完全决定其亲缘关系的.荒漠锦鸡儿(Caragana roborovskyi )和卷叶锦鸡儿(Caragana ordosica )绒毛为棒状, 表皮细胞和蜡质层形态亦相似.短脚锦鸡儿(Caragana brachypoda )和狭叶锦鸡儿(Caragana stenophylla )表皮毛为柳叶带乳突型, 上表皮气孔器深陷, 气孔器为不等型或不规则型, 气孔口三角形或多边形.红花锦鸡儿(Caragana rosea )为明显不同的类群, 表皮无绒毛结构, 且蜡质层形态不同于以上几种的鳞片状, 而是紧贴表皮的膜片层结构或颗粒状. ...
骆驼蓬属种子微形态及其生态学与分类学意义
1997
Micromorphology, histochemistry and ultrastructure of the foliar trichomes of Withania somnifera(L.) Dunal (Solanaceae)
1
2015
... 而对于腺毛来说, 其主要作为一个油性物质的合成、分泌和累积的器官, 有效地防御动物的捕食、病菌入侵以及物种竞争(Werker, 1993 ; Wagner et al ., 2003 ; Kremer et al ., 2014 ; Munien et al ., 2015 ).实际上, 腺毛也是次生代谢产物的一个独立的细胞工厂(Tissier, 2012 ).腺毛的形态随植物的组织器官和种类不同而呈多样化, 其形成和分泌的主要油性物质及其产物组成受遗传和环境两方面的因素控制(Bosabalidis & Kofidis, 2002 ; Shafiee-Hajiabad et al ., 2014 ), 腺毛结构的多样性也可以为植物分类学提供依据(Gairola et al ., 2009 ; Rusydi et al ., 2013 ).对117种荒漠植物叶片表皮扫描发现: 不少旱生植物除具有非腺体表皮毛外, 还同时具有腺毛, 并且有些腺毛体积较大, 如蒙古莸、甘草、香青兰、星毛短舌菊、百里香等.显然, 对于具有腺毛的荒漠植物, 腺毛具有重要的功能.但目前关于不同植物腺毛的形态、形成以及功能的研究还很有局限性.那么, 这些腺毛在植物适应荒漠环境的过程中其形态结构与环境因子有何关系、主要分泌哪些化学物质、发挥什么样的功能?这些问题亟待下一步的深入研究. ...
Trichome morphology in Teucrium L.(Labiatae). A taxonomic review
1
1999
... 大多数荒漠植物叶片的一个显著特点是表皮被绒毛, 叶表皮毛的出现与长期的干旱气候有关, 是植物对干旱环境的适应特征(Retamales & Scharaschkin, 2015 ).植物叶片表皮毛分两种: 腺毛和非腺毛(Werker, 1993 ; Navarro & Oualidi, 1999 ).非腺毛的主要功能与植物的防御有关.目前确认的植物叶片表皮非腺毛具有的生理生态功能主要有: 减轻紫外辐射(Karabourniotis et al ., 1992 ; Gregoriou et al ., 2007 )、降低边界层的传导(Wuenscher, 1970 ; Kenzo et al ., 2008 )、降低叶温(Ehleringer & Mooney, 1978 ), 以及减少对太阳辐射的接收(Ehleringer et al ., 1976 )等.这些功能决定了干旱区荒漠植物抗逆能力的大小.干旱地区的叶表皮毛密度比湿润地区的高(Smith & Nobel, 1977 ; Aronne & de Micco, 2001 ), 就单个物种而言, 表皮毛密度与温度和辐射密切相关(Perez-Estrada et al ., 2000 ).在热带雨林树种中, 浓密的表皮毛很少见(Turner, 2001 ), Roth (1984)调查的250种树中仅有15种有较厚的表皮毛, 且较厚的表皮毛一般在干旱、高温和强辐射条件下产生.而在干旱半干旱的荒漠区, 大多数植物都具有浓密的表皮毛.显然, 荒漠植物叶上密生的表皮毛对于植物的生活有重要的意义.从Ehleringer等 (1976)、Ehleringer和Bjorkman (1978)、Ehleringer和Mooney (1978)研究的荒漠灌木Encelia farinosa 的叶表皮毛对叶片光谱特性和光合、蒸腾等生理活动的影响来看, 沙漠毒菊在不同生长季生长的叶片具有不同数量的表皮毛, 表皮毛的变化足够使叶片对光合有效辐射的吸收从81%降到29%, 对总的太阳辐射的吸收从46%降到16%, 而亲缘关系较近却没有表皮毛的同属物种Encelia californiea 的有效光合辐射吸收率为84%.同时, 由于对太阳辐射吸收的减少, 导致叶面温度和植物蒸腾速率的降低.荒漠环境气温经常超过40 ℃, 而植物最适宜的光合温度是25 ℃, 当温度超过35 ℃时植物的光合速率会突降, 但较厚的叶表皮毛能使叶温显著低于气温而接近光合适宜温度, 同时也避免植物达到叶片致死温度.但叶表皮毛也有不利的一面, 表皮毛的覆盖降低了光合速率, 因此同一植物在不同的生境和生长季长出的叶片会出现表皮毛数量不同的特征, 这也是植物适应环境的一种权衡策略(Kenzo et al ., 2008 ).从本研究中的117种荒漠植物叶表皮特征来看, 片状叶片基本被有表皮毛, 并有不少叶片的表皮毛数量较多, 直接增加了叶片的边界层厚度, 个别植物的上下表皮毛厚度远超于叶片厚度, 如表皮微形态抗逆适应主要类型的第一类植物中的柠条锦鸡儿、卷叶锦鸡儿和沙枣等.可见荒漠植物中叶片表皮毛的作用是相当重要的, 它们不仅能调节植物的生理功能, 而且可抵御风沙等机械损伤的直接伤害.这些物种在不同的自然生境中其表皮毛数量亦不同(数据省略), 说明生态环境深刻地影响着植物形态结构的形成与变异. ...
Foliar tissue architectural diversity among three species of genus Hibiscus for better adaptability under industrial environment
1
2012
... 植物叶片对环境的响应首先是叶片表皮对环境的感应和变化(Pal et al ., 2002 ; Bargel et al ., 2004 ; Koch et al ., 2009 ; Adebowale et al ., 2014 ).生长于干燥的气候和土壤条件下的荒漠植物, 叶表皮细胞会变小, 切向壁加厚, 具有内皮层.叶表皮角质膜厚度增加, 角质层由光滑变得粗糙, 能够反射阳光, 减少叶片表面空气的流动, 降低蒸腾作用, 防止水分过度丧失(贺金生等, 1994 ; 杨明博等, 2007 ).气孔的分布特征、密度和面积等都会受到影响, 气孔多分布于叶片下表皮, 这种分布模式既可促进植物与外界环境进行气体交换, 又能保持水分.随着环境中水分和湿度的减少, 气孔密度增加, 气孔面积则向小型化发展, 气孔多下陷形成气孔窝或其上有突出的角质膜(Sam et al ., 2000 ; Bosabalidis & Kofidis, 2002 ; 高建平等, 2003 ; Xu & Zhou, 2008 ; Gan et al ., 2010 ; de Micco & Aronne, 2012 ).叶表皮附属物(如表皮毛和表面疣状突起物)也发生变化, 它们可有效地避免叶肉组织受损(Gairola et al ., 2009 ; Koch et al ., 2009 ; Gomes et al ., 2011 ; Noman et al ., 2012 ), 表皮蜡质(waxes)晶体形成二维或三维结构, 具有降低表皮的可湿性以及反射强光等作用(Wagner et al ., 2003 ; Koch & Ensikat, 2008 ). ...
Plant responses to precipitation in desert ecosystems: Integrating functional types, pulses, thresholds, and delays
1
2004
... 荒漠植物由于长期适应所生长的自然环境, 发展了一套最适宜自身生长发育的形态结构特征, 并采取各种途径来抵御或忍耐干旱胁迫对它的影响, 因而具有一定程度的耐旱性.这种耐旱性是一种复合性状, 包括植物的形态结构特征、水分生理特征、生理生化反应、组织细胞、光合器官以及原生质结构等特征的综合反映(Klich, 2000 ; Chaves et al ., 2003 ; Ogle & Reynolds, 2004 ).不同的荒漠植物对水分和不利环境的适应和耐受程度不同, 适应方式多种多样, 按照不同的分类标准, 荒漠植物有多种适应类型.通常的分类标准包括耐旱程度、耐旱方式、叶片形态、生活史、光合碳代谢途径等(赵哈林, 2012 ).各种类型的植物又有各自的适应特征.本研究是针对我国干旱半干旱区荒漠植物叶表皮微形态结构特征的综合分析, 主要集中于该区域的建群种灌木物种.根据荒漠植物叶表皮微形态基本结构特征和表皮附属物的类型与数量, 将荒漠植物表皮微形态类型直观地划分为11种, 而将抗逆适应微形态结构划分为6种类型.荒漠植物叶表皮微形态结构类型复杂, 反映了植物自身的遗传特性及其与环境的密切关系.植物叶片表皮微形态主要由遗传因素决定, 但表皮附属物绒毛数量, 蜡质形态与厚度, 气孔的形态、密度以及分布特征等与环境因子密切相关, 是植物对环境的响应(Barthlott, 1981 ; Casson & Hetherington, 2010 ; Ghahremaninejad et al ., 2012 ).因此, 基于这些表皮主要附属物特征对荒漠植物的表皮微形态类型和抗逆适应类型进行划分, 能够从微观形态结构特征进一步揭示物种的亲缘关系和表皮微形态结构的生态适应意义. ...
Do leaf surface characters play a role in plant resistance to auto-exhaust pollution?
1
2002
... 植物叶片对环境的响应首先是叶片表皮对环境的感应和变化(Pal et al ., 2002 ; Bargel et al ., 2004 ; Koch et al ., 2009 ; Adebowale et al ., 2014 ).生长于干燥的气候和土壤条件下的荒漠植物, 叶表皮细胞会变小, 切向壁加厚, 具有内皮层.叶表皮角质膜厚度增加, 角质层由光滑变得粗糙, 能够反射阳光, 减少叶片表面空气的流动, 降低蒸腾作用, 防止水分过度丧失(贺金生等, 1994 ; 杨明博等, 2007 ).气孔的分布特征、密度和面积等都会受到影响, 气孔多分布于叶片下表皮, 这种分布模式既可促进植物与外界环境进行气体交换, 又能保持水分.随着环境中水分和湿度的减少, 气孔密度增加, 气孔面积则向小型化发展, 气孔多下陷形成气孔窝或其上有突出的角质膜(Sam et al ., 2000 ; Bosabalidis & Kofidis, 2002 ; 高建平等, 2003 ; Xu & Zhou, 2008 ; Gan et al ., 2010 ; de Micco & Aronne, 2012 ).叶表皮附属物(如表皮毛和表面疣状突起物)也发生变化, 它们可有效地避免叶肉组织受损(Gairola et al ., 2009 ; Koch et al ., 2009 ; Gomes et al ., 2011 ; Noman et al ., 2012 ), 表皮蜡质(waxes)晶体形成二维或三维结构, 具有降低表皮的可湿性以及反射强光等作用(Wagner et al ., 2003 ; Koch & Ensikat, 2008 ). ...
Variation in leaf trichomes of Wigandia urens: Environmental factors and physiological consequences
1
2000
... 大多数荒漠植物叶片的一个显著特点是表皮被绒毛, 叶表皮毛的出现与长期的干旱气候有关, 是植物对干旱环境的适应特征(Retamales & Scharaschkin, 2015 ).植物叶片表皮毛分两种: 腺毛和非腺毛(Werker, 1993 ; Navarro & Oualidi, 1999 ).非腺毛的主要功能与植物的防御有关.目前确认的植物叶片表皮非腺毛具有的生理生态功能主要有: 减轻紫外辐射(Karabourniotis et al ., 1992 ; Gregoriou et al ., 2007 )、降低边界层的传导(Wuenscher, 1970 ; Kenzo et al ., 2008 )、降低叶温(Ehleringer & Mooney, 1978 ), 以及减少对太阳辐射的接收(Ehleringer et al ., 1976 )等.这些功能决定了干旱区荒漠植物抗逆能力的大小.干旱地区的叶表皮毛密度比湿润地区的高(Smith & Nobel, 1977 ; Aronne & de Micco, 2001 ), 就单个物种而言, 表皮毛密度与温度和辐射密切相关(Perez-Estrada et al ., 2000 ).在热带雨林树种中, 浓密的表皮毛很少见(Turner, 2001 ), Roth (1984)调查的250种树中仅有15种有较厚的表皮毛, 且较厚的表皮毛一般在干旱、高温和强辐射条件下产生.而在干旱半干旱的荒漠区, 大多数植物都具有浓密的表皮毛.显然, 荒漠植物叶上密生的表皮毛对于植物的生活有重要的意义.从Ehleringer等 (1976)、Ehleringer和Bjorkman (1978)、Ehleringer和Mooney (1978)研究的荒漠灌木Encelia farinosa 的叶表皮毛对叶片光谱特性和光合、蒸腾等生理活动的影响来看, 沙漠毒菊在不同生长季生长的叶片具有不同数量的表皮毛, 表皮毛的变化足够使叶片对光合有效辐射的吸收从81%降到29%, 对总的太阳辐射的吸收从46%降到16%, 而亲缘关系较近却没有表皮毛的同属物种Encelia californiea 的有效光合辐射吸收率为84%.同时, 由于对太阳辐射吸收的减少, 导致叶面温度和植物蒸腾速率的降低.荒漠环境气温经常超过40 ℃, 而植物最适宜的光合温度是25 ℃, 当温度超过35 ℃时植物的光合速率会突降, 但较厚的叶表皮毛能使叶温显著低于气温而接近光合适宜温度, 同时也避免植物达到叶片致死温度.但叶表皮毛也有不利的一面, 表皮毛的覆盖降低了光合速率, 因此同一植物在不同的生境和生长季长出的叶片会出现表皮毛数量不同的特征, 这也是植物适应环境的一种权衡策略(Kenzo et al ., 2008 ).从本研究中的117种荒漠植物叶表皮特征来看, 片状叶片基本被有表皮毛, 并有不少叶片的表皮毛数量较多, 直接增加了叶片的边界层厚度, 个别植物的上下表皮毛厚度远超于叶片厚度, 如表皮微形态抗逆适应主要类型的第一类植物中的柠条锦鸡儿、卷叶锦鸡儿和沙枣等.可见荒漠植物中叶片表皮毛的作用是相当重要的, 它们不仅能调节植物的生理功能, 而且可抵御风沙等机械损伤的直接伤害.这些物种在不同的自然生境中其表皮毛数量亦不同(数据省略), 说明生态环境深刻地影响着植物形态结构的形成与变异. ...
Leaf and stem anatomy in eight Hypericum species (Clusiaceae)
1
2013
... 荒漠植物叶片的另一个显著特点是表皮有较厚的角质膜蜡质层.角质膜蜡质层与表皮毛有共同的功能, 是植物表皮的保护屏障, 主要由长链碳氢化合物组成(Shepherd & Wynne, 2006 ).叶片对光的反射由叶面形貌决定, 表皮毛和蜡质层是决定荒漠植物叶表面形貌的关键.Holmes和Keiller (2002)研究了45种桉属和伽蓝菜属植物的蜡质叶, 发现蜡质层对紫外光(330 nm)和光合有效辐射(680 nm)的反射比各种形态的表皮毛更强, 去除蜡质层通常会减少对这两种光的反射, 特别是对紫外光的反射, 但对表皮无蜡质的叶片影响不大.同时不同的物种对不同波长的光的反射亦不同, 表明蜡质晶体的形态直接影响着对光的反射, 并且不同温度下的反射强度也不同, 可见蜡质层对光的反射特性变化很大(Holmes & Keiller, 2002 ).叶表皮蜡质层参与植物对水分、盐、低温、污染、机械损伤等多种胁迫响应的过程, 提高植物对胁迫的抗性(Cameron, 1970 ; Bargel et al ., 2004 ; Perrone et al ., 2013 , 2015 ).因此, 本研究中117种荒漠植物叶片表皮颗粒状、膜片状、晶簇状、鳞片状、杆状等各种不同形态的蜡质晶体既是植物长期适应环境的结果, 也对荒漠植物在严酷环境中的生长和生存至关重要.总的来说, 这些植物中灌木的蜡质层密度高于草本, 无表皮毛的叶片蜡质层密度高于表皮毛覆盖的, 特别是以同化枝为主要光合器官和棒状叶片的植物, 表皮基本没有表皮毛覆盖, 主要的附属物是蜡质层, 如麻黄属、沙拐枣属、红砂属的植物等, 弥补了表皮毛的功能. ...
What do leaf anatomy and micro-morphology tell us about the psammophilous Pancratium maritimum L.(Amaryllidaceae) in response to sand dune conditions?
2
2015
... 干旱半干旱区的荒漠环境属于一种极端环境, 降水稀少, 气候干燥, 昼夜温差大, 地面温度高, 土壤贫瘠, 风沙活动强烈……这些恶劣条件无时不在限制着荒漠植物的生存和繁衍.但即使在这种条件下, 仍有许多植物分布于干旱半干旱区, 且能很好地繁衍生息.那么, 这些植物有何特殊的形态结构来适应环境呢?如何通过植物形态结构特征判断其适应能力? 这些问题引发不少科学家对荒漠植物研究的极大关注.Schow提出“旱生植物”概念已有100多年了. 国内外不少学者研究过旱生植物的形态结构(Shield, 1950 ; 王勋陵和马骥, 1999 ; 严巧娣等, 2008 ; Liu et al ., 2012 , 2015 ; Perrone et al ., 2015 ), 但以各自有限数量的旱生植物形态结构描述为主, 至今未见对较大数量旱生植物叶片表皮微形态与结构特征进行数量化整理, 并对其类型划分、变化规律、演化趋势、适应对策等重要问题进行系统讨论的 报道. ...
... 荒漠植物叶片的另一个显著特点是表皮有较厚的角质膜蜡质层.角质膜蜡质层与表皮毛有共同的功能, 是植物表皮的保护屏障, 主要由长链碳氢化合物组成(Shepherd & Wynne, 2006 ).叶片对光的反射由叶面形貌决定, 表皮毛和蜡质层是决定荒漠植物叶表面形貌的关键.Holmes和Keiller (2002)研究了45种桉属和伽蓝菜属植物的蜡质叶, 发现蜡质层对紫外光(330 nm)和光合有效辐射(680 nm)的反射比各种形态的表皮毛更强, 去除蜡质层通常会减少对这两种光的反射, 特别是对紫外光的反射, 但对表皮无蜡质的叶片影响不大.同时不同的物种对不同波长的光的反射亦不同, 表明蜡质晶体的形态直接影响着对光的反射, 并且不同温度下的反射强度也不同, 可见蜡质层对光的反射特性变化很大(Holmes & Keiller, 2002 ).叶表皮蜡质层参与植物对水分、盐、低温、污染、机械损伤等多种胁迫响应的过程, 提高植物对胁迫的抗性(Cameron, 1970 ; Bargel et al ., 2004 ; Perrone et al ., 2013 , 2015 ).因此, 本研究中117种荒漠植物叶片表皮颗粒状、膜片状、晶簇状、鳞片状、杆状等各种不同形态的蜡质晶体既是植物长期适应环境的结果, 也对荒漠植物在严酷环境中的生长和生存至关重要.总的来说, 这些植物中灌木的蜡质层密度高于草本, 无表皮毛的叶片蜡质层密度高于表皮毛覆盖的, 特别是以同化枝为主要光合器官和棒状叶片的植物, 表皮基本没有表皮毛覆盖, 主要的附属物是蜡质层, 如麻黄属、沙拐枣属、红砂属的植物等, 弥补了表皮毛的功能. ...
16种高山植物叶片内的异细胞及其生态学意义
1
1998
... 植物作为生态系统的第一生产者, 受环境变化的影响是直接而多方面的.叶片是植物进化过程中对环境变化最敏感且可塑性较大的器官, 在不同的选择压力下形成了各种适应类型(Hickey, 1984 ; 祁如虎等, 1998 ; Gielwanowska et al ., 2005 ), 其形态结构特征最能体现环境因子的影响或植物对环境的适应(王勋陵和王静, 1989 ; Krober et al ., 2015 ). ...
Comparative leaf anatomy and micromorphology of the Chilean Myrtaceae: Taxonomic and ecological implications
1
2015
... 大多数荒漠植物叶片的一个显著特点是表皮被绒毛, 叶表皮毛的出现与长期的干旱气候有关, 是植物对干旱环境的适应特征(Retamales & Scharaschkin, 2015 ).植物叶片表皮毛分两种: 腺毛和非腺毛(Werker, 1993 ; Navarro & Oualidi, 1999 ).非腺毛的主要功能与植物的防御有关.目前确认的植物叶片表皮非腺毛具有的生理生态功能主要有: 减轻紫外辐射(Karabourniotis et al ., 1992 ; Gregoriou et al ., 2007 )、降低边界层的传导(Wuenscher, 1970 ; Kenzo et al ., 2008 )、降低叶温(Ehleringer & Mooney, 1978 ), 以及减少对太阳辐射的接收(Ehleringer et al ., 1976 )等.这些功能决定了干旱区荒漠植物抗逆能力的大小.干旱地区的叶表皮毛密度比湿润地区的高(Smith & Nobel, 1977 ; Aronne & de Micco, 2001 ), 就单个物种而言, 表皮毛密度与温度和辐射密切相关(Perez-Estrada et al ., 2000 ).在热带雨林树种中, 浓密的表皮毛很少见(Turner, 2001 ), Roth (1984)调查的250种树中仅有15种有较厚的表皮毛, 且较厚的表皮毛一般在干旱、高温和强辐射条件下产生.而在干旱半干旱的荒漠区, 大多数植物都具有浓密的表皮毛.显然, 荒漠植物叶上密生的表皮毛对于植物的生活有重要的意义.从Ehleringer等 (1976)、Ehleringer和Bjorkman (1978)、Ehleringer和Mooney (1978)研究的荒漠灌木Encelia farinosa 的叶表皮毛对叶片光谱特性和光合、蒸腾等生理活动的影响来看, 沙漠毒菊在不同生长季生长的叶片具有不同数量的表皮毛, 表皮毛的变化足够使叶片对光合有效辐射的吸收从81%降到29%, 对总的太阳辐射的吸收从46%降到16%, 而亲缘关系较近却没有表皮毛的同属物种Encelia californiea 的有效光合辐射吸收率为84%.同时, 由于对太阳辐射吸收的减少, 导致叶面温度和植物蒸腾速率的降低.荒漠环境气温经常超过40 ℃, 而植物最适宜的光合温度是25 ℃, 当温度超过35 ℃时植物的光合速率会突降, 但较厚的叶表皮毛能使叶温显著低于气温而接近光合适宜温度, 同时也避免植物达到叶片致死温度.但叶表皮毛也有不利的一面, 表皮毛的覆盖降低了光合速率, 因此同一植物在不同的生境和生长季长出的叶片会出现表皮毛数量不同的特征, 这也是植物适应环境的一种权衡策略(Kenzo et al ., 2008 ).从本研究中的117种荒漠植物叶表皮特征来看, 片状叶片基本被有表皮毛, 并有不少叶片的表皮毛数量较多, 直接增加了叶片的边界层厚度, 个别植物的上下表皮毛厚度远超于叶片厚度, 如表皮微形态抗逆适应主要类型的第一类植物中的柠条锦鸡儿、卷叶锦鸡儿和沙枣等.可见荒漠植物中叶片表皮毛的作用是相当重要的, 它们不仅能调节植物的生理功能, 而且可抵御风沙等机械损伤的直接伤害.这些物种在不同的自然生境中其表皮毛数量亦不同(数据省略), 说明生态环境深刻地影响着植物形态结构的形成与变异. ...
Stratification of Tropical Forests As Seen in Leaf Structure
1984
Morphology of trichomes inPogostemon cablin Benth.(Lamiaceae)
1
2013
... 而对于腺毛来说, 其主要作为一个油性物质的合成、分泌和累积的器官, 有效地防御动物的捕食、病菌入侵以及物种竞争(Werker, 1993 ; Wagner et al ., 2003 ; Kremer et al ., 2014 ; Munien et al ., 2015 ).实际上, 腺毛也是次生代谢产物的一个独立的细胞工厂(Tissier, 2012 ).腺毛的形态随植物的组织器官和种类不同而呈多样化, 其形成和分泌的主要油性物质及其产物组成受遗传和环境两方面的因素控制(Bosabalidis & Kofidis, 2002 ; Shafiee-Hajiabad et al ., 2014 ), 腺毛结构的多样性也可以为植物分类学提供依据(Gairola et al ., 2009 ; Rusydi et al ., 2013 ).对117种荒漠植物叶片表皮扫描发现: 不少旱生植物除具有非腺体表皮毛外, 还同时具有腺毛, 并且有些腺毛体积较大, 如蒙古莸、甘草、香青兰、星毛短舌菊、百里香等.显然, 对于具有腺毛的荒漠植物, 腺毛具有重要的功能.但目前关于不同植物腺毛的形态、形成以及功能的研究还很有局限性.那么, 这些腺毛在植物适应荒漠环境的过程中其形态结构与环境因子有何关系、主要分泌哪些化学物质、发挥什么样的功能?这些问题亟待下一步的深入研究. ...
Water stress induced changes in anatomy of tomato leaf epidermis
1
2000
... 植物叶片对环境的响应首先是叶片表皮对环境的感应和变化(Pal et al ., 2002 ; Bargel et al ., 2004 ; Koch et al ., 2009 ; Adebowale et al ., 2014 ).生长于干燥的气候和土壤条件下的荒漠植物, 叶表皮细胞会变小, 切向壁加厚, 具有内皮层.叶表皮角质膜厚度增加, 角质层由光滑变得粗糙, 能够反射阳光, 减少叶片表面空气的流动, 降低蒸腾作用, 防止水分过度丧失(贺金生等, 1994 ; 杨明博等, 2007 ).气孔的分布特征、密度和面积等都会受到影响, 气孔多分布于叶片下表皮, 这种分布模式既可促进植物与外界环境进行气体交换, 又能保持水分.随着环境中水分和湿度的减少, 气孔密度增加, 气孔面积则向小型化发展, 气孔多下陷形成气孔窝或其上有突出的角质膜(Sam et al ., 2000 ; Bosabalidis & Kofidis, 2002 ; 高建平等, 2003 ; Xu & Zhou, 2008 ; Gan et al ., 2010 ; de Micco & Aronne, 2012 ).叶表皮附属物(如表皮毛和表面疣状突起物)也发生变化, 它们可有效地避免叶肉组织受损(Gairola et al ., 2009 ; Koch et al ., 2009 ; Gomes et al ., 2011 ; Noman et al ., 2012 ), 表皮蜡质(waxes)晶体形成二维或三维结构, 具有降低表皮的可湿性以及反射强光等作用(Wagner et al ., 2003 ; Koch & Ensikat, 2008 ). ...
Anatomy of Brazilian Eriocaulaceae: Correlation with taxonomy and habitat using multivariate analyses
1
2005
... 植物叶表皮微形态特征已用于解决物种分类问题、建立物种进化关系、阐明表皮微形态结构的生态适应性等多种研究目的(Dickison, 2000 ; Scatena et al ., 2005 ; Stenglein et al ., 2005 ; Aliero et al ., 2006 ; Chen et al ., 2010 ; Zhou & Xia, 2012 ; Adebowale et al ., 2014 ; Deng et al ., 2014 ; Zhang et al ., 2014 ; Francisco et al ., 2015 ; 朱广龙等, 2016 ).为了探讨荒漠植物叶片表皮微形态结构的适应特征及其亲缘关系, 选择分布于同一区域的不同植物和不同降水量梯度生境下的同一植物, 利用扫描电镜观察分析了117种荒漠植物叶片的表皮微形态结构, 旨在阐明荒漠植物对逆境的适应机理及其演化趋势, 并为优良固沙植物的筛选提供理论依据. ...
Comparative investigation about the trichome morphology of common oregano (Origanum vulgare L. subsp. vulgare) and Greek oregano(Origanum vulgare L. subsp. hirtum)
1
2014
... 而对于腺毛来说, 其主要作为一个油性物质的合成、分泌和累积的器官, 有效地防御动物的捕食、病菌入侵以及物种竞争(Werker, 1993 ; Wagner et al ., 2003 ; Kremer et al ., 2014 ; Munien et al ., 2015 ).实际上, 腺毛也是次生代谢产物的一个独立的细胞工厂(Tissier, 2012 ).腺毛的形态随植物的组织器官和种类不同而呈多样化, 其形成和分泌的主要油性物质及其产物组成受遗传和环境两方面的因素控制(Bosabalidis & Kofidis, 2002 ; Shafiee-Hajiabad et al ., 2014 ), 腺毛结构的多样性也可以为植物分类学提供依据(Gairola et al ., 2009 ; Rusydi et al ., 2013 ).对117种荒漠植物叶片表皮扫描发现: 不少旱生植物除具有非腺体表皮毛外, 还同时具有腺毛, 并且有些腺毛体积较大, 如蒙古莸、甘草、香青兰、星毛短舌菊、百里香等.显然, 对于具有腺毛的荒漠植物, 腺毛具有重要的功能.但目前关于不同植物腺毛的形态、形成以及功能的研究还很有局限性.那么, 这些腺毛在植物适应荒漠环境的过程中其形态结构与环境因子有何关系、主要分泌哪些化学物质、发挥什么样的功能?这些问题亟待下一步的深入研究. ...
The effects of stress on plant cuticular waxes
1
2006
... 荒漠植物叶片的另一个显著特点是表皮有较厚的角质膜蜡质层.角质膜蜡质层与表皮毛有共同的功能, 是植物表皮的保护屏障, 主要由长链碳氢化合物组成(Shepherd & Wynne, 2006 ).叶片对光的反射由叶面形貌决定, 表皮毛和蜡质层是决定荒漠植物叶表面形貌的关键.Holmes和Keiller (2002)研究了45种桉属和伽蓝菜属植物的蜡质叶, 发现蜡质层对紫外光(330 nm)和光合有效辐射(680 nm)的反射比各种形态的表皮毛更强, 去除蜡质层通常会减少对这两种光的反射, 特别是对紫外光的反射, 但对表皮无蜡质的叶片影响不大.同时不同的物种对不同波长的光的反射亦不同, 表明蜡质晶体的形态直接影响着对光的反射, 并且不同温度下的反射强度也不同, 可见蜡质层对光的反射特性变化很大(Holmes & Keiller, 2002 ).叶表皮蜡质层参与植物对水分、盐、低温、污染、机械损伤等多种胁迫响应的过程, 提高植物对胁迫的抗性(Cameron, 1970 ; Bargel et al ., 2004 ; Perrone et al ., 2013 , 2015 ).因此, 本研究中117种荒漠植物叶片表皮颗粒状、膜片状、晶簇状、鳞片状、杆状等各种不同形态的蜡质晶体既是植物长期适应环境的结果, 也对荒漠植物在严酷环境中的生长和生存至关重要.总的来说, 这些植物中灌木的蜡质层密度高于草本, 无表皮毛的叶片蜡质层密度高于表皮毛覆盖的, 特别是以同化枝为主要光合器官和棒状叶片的植物, 表皮基本没有表皮毛覆盖, 主要的附属物是蜡质层, 如麻黄属、沙拐枣属、红砂属的植物等, 弥补了表皮毛的功能. ...
Leaf xeromorphy as related to physiological and structural influences
1
1950
... 干旱半干旱区的荒漠环境属于一种极端环境, 降水稀少, 气候干燥, 昼夜温差大, 地面温度高, 土壤贫瘠, 风沙活动强烈……这些恶劣条件无时不在限制着荒漠植物的生存和繁衍.但即使在这种条件下, 仍有许多植物分布于干旱半干旱区, 且能很好地繁衍生息.那么, 这些植物有何特殊的形态结构来适应环境呢?如何通过植物形态结构特征判断其适应能力? 这些问题引发不少科学家对荒漠植物研究的极大关注.Schow提出“旱生植物”概念已有100多年了. 国内外不少学者研究过旱生植物的形态结构(Shield, 1950 ; 王勋陵和马骥, 1999 ; 严巧娣等, 2008 ; Liu et al ., 2012 , 2015 ; Perrone et al ., 2015 ), 但以各自有限数量的旱生植物形态结构描述为主, 至今未见对较大数量旱生植物叶片表皮微形态与结构特征进行数量化整理, 并对其类型划分、变化规律、演化趋势、适应对策等重要问题进行系统讨论的 报道. ...
Influences of seasonal changes in leaf morphology on water-use efficiency for three desert broadleaf shrubs
1
1977
... 大多数荒漠植物叶片的一个显著特点是表皮被绒毛, 叶表皮毛的出现与长期的干旱气候有关, 是植物对干旱环境的适应特征(Retamales & Scharaschkin, 2015 ).植物叶片表皮毛分两种: 腺毛和非腺毛(Werker, 1993 ; Navarro & Oualidi, 1999 ).非腺毛的主要功能与植物的防御有关.目前确认的植物叶片表皮非腺毛具有的生理生态功能主要有: 减轻紫外辐射(Karabourniotis et al ., 1992 ; Gregoriou et al ., 2007 )、降低边界层的传导(Wuenscher, 1970 ; Kenzo et al ., 2008 )、降低叶温(Ehleringer & Mooney, 1978 ), 以及减少对太阳辐射的接收(Ehleringer et al ., 1976 )等.这些功能决定了干旱区荒漠植物抗逆能力的大小.干旱地区的叶表皮毛密度比湿润地区的高(Smith & Nobel, 1977 ; Aronne & de Micco, 2001 ), 就单个物种而言, 表皮毛密度与温度和辐射密切相关(Perez-Estrada et al ., 2000 ).在热带雨林树种中, 浓密的表皮毛很少见(Turner, 2001 ), Roth (1984)调查的250种树中仅有15种有较厚的表皮毛, 且较厚的表皮毛一般在干旱、高温和强辐射条件下产生.而在干旱半干旱的荒漠区, 大多数植物都具有浓密的表皮毛.显然, 荒漠植物叶上密生的表皮毛对于植物的生活有重要的意义.从Ehleringer等 (1976)、Ehleringer和Bjorkman (1978)、Ehleringer和Mooney (1978)研究的荒漠灌木Encelia farinosa 的叶表皮毛对叶片光谱特性和光合、蒸腾等生理活动的影响来看, 沙漠毒菊在不同生长季生长的叶片具有不同数量的表皮毛, 表皮毛的变化足够使叶片对光合有效辐射的吸收从81%降到29%, 对总的太阳辐射的吸收从46%降到16%, 而亲缘关系较近却没有表皮毛的同属物种Encelia californiea 的有效光合辐射吸收率为84%.同时, 由于对太阳辐射吸收的减少, 导致叶面温度和植物蒸腾速率的降低.荒漠环境气温经常超过40 ℃, 而植物最适宜的光合温度是25 ℃, 当温度超过35 ℃时植物的光合速率会突降, 但较厚的叶表皮毛能使叶温显著低于气温而接近光合适宜温度, 同时也避免植物达到叶片致死温度.但叶表皮毛也有不利的一面, 表皮毛的覆盖降低了光合速率, 因此同一植物在不同的生境和生长季长出的叶片会出现表皮毛数量不同的特征, 这也是植物适应环境的一种权衡策略(Kenzo et al ., 2008 ).从本研究中的117种荒漠植物叶表皮特征来看, 片状叶片基本被有表皮毛, 并有不少叶片的表皮毛数量较多, 直接增加了叶片的边界层厚度, 个别植物的上下表皮毛厚度远超于叶片厚度, 如表皮微形态抗逆适应主要类型的第一类植物中的柠条锦鸡儿、卷叶锦鸡儿和沙枣等.可见荒漠植物中叶片表皮毛的作用是相当重要的, 它们不仅能调节植物的生理功能, 而且可抵御风沙等机械损伤的直接伤害.这些物种在不同的自然生境中其表皮毛数量亦不同(数据省略), 说明生态环境深刻地影响着植物形态结构的形成与变异. ...
Leaf epidermal characters related with plant’s passive resistance to pathogens vary among accessions of wild beans Phaseolus vulgaris var. aborigineus (Leguminosae-Phaseoleae)
1
2005
... 植物叶表皮微形态特征已用于解决物种分类问题、建立物种进化关系、阐明表皮微形态结构的生态适应性等多种研究目的(Dickison, 2000 ; Scatena et al ., 2005 ; Stenglein et al ., 2005 ; Aliero et al ., 2006 ; Chen et al ., 2010 ; Zhou & Xia, 2012 ; Adebowale et al ., 2014 ; Deng et al ., 2014 ; Zhang et al ., 2014 ; Francisco et al ., 2015 ; 朱广龙等, 2016 ).为了探讨荒漠植物叶片表皮微形态结构的适应特征及其亲缘关系, 选择分布于同一区域的不同植物和不同降水量梯度生境下的同一植物, 利用扫描电镜观察分析了117种荒漠植物叶片的表皮微形态结构, 旨在阐明荒漠植物对逆境的适应机理及其演化趋势, 并为优良固沙植物的筛选提供理论依据. ...
Micromorphomics: A morphological dissection to unveil environmental stress
1
2015
... 植物叶片适应环境的形态结构变化是多方面的.在众多生态因子中, 叶片受水分、温度和光因子的影响最为显著(王勋陵和王静, 1989 ; 王勋陵, 1993 ; Aronne & de Micco 2001 ; Arena et al ., 2008 ; Jaleel et al ., 2009a , 2009b ), 主要表现为叶形的变化、叶片表皮附属物的变化、叶片的厚度及解剖结构的差异(Talukdar, 2015 ).对于荒漠植物来说, 夏季极端干旱时它们可能同时遭受以上各种环境因子的胁迫.但其最主要的限制因子为水分, 因而表现出典型的旱生植物形态结构特征.由于遗传因子与环境因子的共同作用, 在旱生植物结构的适应性上出现了两种演化趋势, 即趋同性和趋异性, 也就是说, 在相同环境下的植物演化出相似的形态结构适应性, 而不同环境下的同类植物演化出不同的形态结构(Lakusic et al ., 2010 ).因此, 要系统分析荒漠植物叶片形态结构特征的适应性变化, 需要将一个区域不同植物和不同梯度生境下的同一植物做全面分析. ...
Trichome specific expression: Promoters and their applications
1
2012
... 而对于腺毛来说, 其主要作为一个油性物质的合成、分泌和累积的器官, 有效地防御动物的捕食、病菌入侵以及物种竞争(Werker, 1993 ; Wagner et al ., 2003 ; Kremer et al ., 2014 ; Munien et al ., 2015 ).实际上, 腺毛也是次生代谢产物的一个独立的细胞工厂(Tissier, 2012 ).腺毛的形态随植物的组织器官和种类不同而呈多样化, 其形成和分泌的主要油性物质及其产物组成受遗传和环境两方面的因素控制(Bosabalidis & Kofidis, 2002 ; Shafiee-Hajiabad et al ., 2014 ), 腺毛结构的多样性也可以为植物分类学提供依据(Gairola et al ., 2009 ; Rusydi et al ., 2013 ).对117种荒漠植物叶片表皮扫描发现: 不少旱生植物除具有非腺体表皮毛外, 还同时具有腺毛, 并且有些腺毛体积较大, 如蒙古莸、甘草、香青兰、星毛短舌菊、百里香等.显然, 对于具有腺毛的荒漠植物, 腺毛具有重要的功能.但目前关于不同植物腺毛的形态、形成以及功能的研究还很有局限性.那么, 这些腺毛在植物适应荒漠环境的过程中其形态结构与环境因子有何关系、主要分泌哪些化学物质、发挥什么样的功能?这些问题亟待下一步的深入研究. ...
1
2001
... 大多数荒漠植物叶片的一个显著特点是表皮被绒毛, 叶表皮毛的出现与长期的干旱气候有关, 是植物对干旱环境的适应特征(Retamales & Scharaschkin, 2015 ).植物叶片表皮毛分两种: 腺毛和非腺毛(Werker, 1993 ; Navarro & Oualidi, 1999 ).非腺毛的主要功能与植物的防御有关.目前确认的植物叶片表皮非腺毛具有的生理生态功能主要有: 减轻紫外辐射(Karabourniotis et al ., 1992 ; Gregoriou et al ., 2007 )、降低边界层的传导(Wuenscher, 1970 ; Kenzo et al ., 2008 )、降低叶温(Ehleringer & Mooney, 1978 ), 以及减少对太阳辐射的接收(Ehleringer et al ., 1976 )等.这些功能决定了干旱区荒漠植物抗逆能力的大小.干旱地区的叶表皮毛密度比湿润地区的高(Smith & Nobel, 1977 ; Aronne & de Micco, 2001 ), 就单个物种而言, 表皮毛密度与温度和辐射密切相关(Perez-Estrada et al ., 2000 ).在热带雨林树种中, 浓密的表皮毛很少见(Turner, 2001 ), Roth (1984)调查的250种树中仅有15种有较厚的表皮毛, 且较厚的表皮毛一般在干旱、高温和强辐射条件下产生.而在干旱半干旱的荒漠区, 大多数植物都具有浓密的表皮毛.显然, 荒漠植物叶上密生的表皮毛对于植物的生活有重要的意义.从Ehleringer等 (1976)、Ehleringer和Bjorkman (1978)、Ehleringer和Mooney (1978)研究的荒漠灌木Encelia farinosa 的叶表皮毛对叶片光谱特性和光合、蒸腾等生理活动的影响来看, 沙漠毒菊在不同生长季生长的叶片具有不同数量的表皮毛, 表皮毛的变化足够使叶片对光合有效辐射的吸收从81%降到29%, 对总的太阳辐射的吸收从46%降到16%, 而亲缘关系较近却没有表皮毛的同属物种Encelia californiea 的有效光合辐射吸收率为84%.同时, 由于对太阳辐射吸收的减少, 导致叶面温度和植物蒸腾速率的降低.荒漠环境气温经常超过40 ℃, 而植物最适宜的光合温度是25 ℃, 当温度超过35 ℃时植物的光合速率会突降, 但较厚的叶表皮毛能使叶温显著低于气温而接近光合适宜温度, 同时也避免植物达到叶片致死温度.但叶表皮毛也有不利的一面, 表皮毛的覆盖降低了光合速率, 因此同一植物在不同的生境和生长季长出的叶片会出现表皮毛数量不同的特征, 这也是植物适应环境的一种权衡策略(Kenzo et al ., 2008 ).从本研究中的117种荒漠植物叶表皮特征来看, 片状叶片基本被有表皮毛, 并有不少叶片的表皮毛数量较多, 直接增加了叶片的边界层厚度, 个别植物的上下表皮毛厚度远超于叶片厚度, 如表皮微形态抗逆适应主要类型的第一类植物中的柠条锦鸡儿、卷叶锦鸡儿和沙枣等.可见荒漠植物中叶片表皮毛的作用是相当重要的, 它们不仅能调节植物的生理功能, 而且可抵御风沙等机械损伤的直接伤害.这些物种在不同的自然生境中其表皮毛数量亦不同(数据省略), 说明生态环境深刻地影响着植物形态结构的形成与变异. ...
Quantitative assessment to the structural basis of water repellency in natural and technical surfaces
2
2003
... 植物叶片对环境的响应首先是叶片表皮对环境的感应和变化(Pal et al ., 2002 ; Bargel et al ., 2004 ; Koch et al ., 2009 ; Adebowale et al ., 2014 ).生长于干燥的气候和土壤条件下的荒漠植物, 叶表皮细胞会变小, 切向壁加厚, 具有内皮层.叶表皮角质膜厚度增加, 角质层由光滑变得粗糙, 能够反射阳光, 减少叶片表面空气的流动, 降低蒸腾作用, 防止水分过度丧失(贺金生等, 1994 ; 杨明博等, 2007 ).气孔的分布特征、密度和面积等都会受到影响, 气孔多分布于叶片下表皮, 这种分布模式既可促进植物与外界环境进行气体交换, 又能保持水分.随着环境中水分和湿度的减少, 气孔密度增加, 气孔面积则向小型化发展, 气孔多下陷形成气孔窝或其上有突出的角质膜(Sam et al ., 2000 ; Bosabalidis & Kofidis, 2002 ; 高建平等, 2003 ; Xu & Zhou, 2008 ; Gan et al ., 2010 ; de Micco & Aronne, 2012 ).叶表皮附属物(如表皮毛和表面疣状突起物)也发生变化, 它们可有效地避免叶肉组织受损(Gairola et al ., 2009 ; Koch et al ., 2009 ; Gomes et al ., 2011 ; Noman et al ., 2012 ), 表皮蜡质(waxes)晶体形成二维或三维结构, 具有降低表皮的可湿性以及反射强光等作用(Wagner et al ., 2003 ; Koch & Ensikat, 2008 ). ...
... 而对于腺毛来说, 其主要作为一个油性物质的合成、分泌和累积的器官, 有效地防御动物的捕食、病菌入侵以及物种竞争(Werker, 1993 ; Wagner et al ., 2003 ; Kremer et al ., 2014 ; Munien et al ., 2015 ).实际上, 腺毛也是次生代谢产物的一个独立的细胞工厂(Tissier, 2012 ).腺毛的形态随植物的组织器官和种类不同而呈多样化, 其形成和分泌的主要油性物质及其产物组成受遗传和环境两方面的因素控制(Bosabalidis & Kofidis, 2002 ; Shafiee-Hajiabad et al ., 2014 ), 腺毛结构的多样性也可以为植物分类学提供依据(Gairola et al ., 2009 ; Rusydi et al ., 2013 ).对117种荒漠植物叶片表皮扫描发现: 不少旱生植物除具有非腺体表皮毛外, 还同时具有腺毛, 并且有些腺毛体积较大, 如蒙古莸、甘草、香青兰、星毛短舌菊、百里香等.显然, 对于具有腺毛的荒漠植物, 腺毛具有重要的功能.但目前关于不同植物腺毛的形态、形成以及功能的研究还很有局限性.那么, 这些腺毛在植物适应荒漠环境的过程中其形态结构与环境因子有何关系、主要分泌哪些化学物质、发挥什么样的功能?这些问题亟待下一步的深入研究. ...
28种柳属植物的叶表皮微形态特征及其分类学意义
1
2012
... 荒漠植物叶表皮微形态结构在不同分类等级上表现出丰富的多样性, 深刻地反映了植物的亲缘关系, 不同程度地揭示了它们之间错综复杂的系统演化关系.叶片表皮微形态结构基本特征根据表皮细胞形态结构和表皮附属物类型, 分为疣状、瘤状、皱褶状、相对光滑型、表面完全被毛、表面完全被蜡质层、不同形态结构的绒毛和蜡质层混合覆盖、上下完全异面以及气孔器集中于下表面等11种类型, 有些类型还包括若干种亚型.如柳属中观察的4种植物, 尽管种及地理分布完全不同, 但它们都为典型的异面叶, 叶片上下表皮都是完全不同的形态.上表皮绒毛密度较高, 不同的种有不同形态和密度的绒毛; 表皮绒毛密度低或不具绒毛, 但有柳属植物特有的角质膜蜡质纹饰(王东超等, 2012 ; Ghahremaninejad et al ., 2012 ), 如黄柳的痂状蜡质层、旱柳和乌柳的壳状蜡质层、沙柳的膜片状蜡质晶体.麻黄属的膜果麻黄、斑子麻黄(Ephedra rhytidosperma )和草麻黄(Ephedra sinica )中, 3种同化枝的角质膜蜡质晶体数量依次减少, 斑子麻黄与草麻黄的表皮形态结构接近, 显示亲缘关系较近.膜果麻黄表皮粗糙, 完全被蜡质层覆盖.锦鸡儿属(Caragana )是豆科蝶形花亚科中较大的一个属, 叶片上下表皮微形态基本一致, 多数为等面叶.但表皮毛的形态密度各异, 柠条锦鸡儿、小叶锦鸡儿(Caragana microphlla )和中间锦鸡儿(Caragana intermedia )同属小叶系, 亲缘关系相近, 表皮毛为柳叶状, 但绒毛密度不同, 柠条锦鸡儿表皮毛完全覆盖叶面, 小叶锦鸡儿和中间锦鸡儿表皮毛密度较小, 且它们的鳞片状蜡质晶体密度相似, 显示他们之间更近的亲缘关系, 这与其他分子生物学依据得到的结果不完全一致(Guo et al ., 2008 ).究其原因, 主要是因为这3种锦鸡儿属植物之间由于分布区域重叠或交叉, 因此存在复杂的基因交流(Ma et al ., 2003 ), 不同种的亲缘关系与其分布区域的其他种之间的相互杂交有关, 甚至出现了杂交过渡态类型, 因而给分类带来了难题, 单纯某一种分类依据是不能完全决定其亲缘关系的.荒漠锦鸡儿(Caragana roborovskyi )和卷叶锦鸡儿(Caragana ordosica )绒毛为棒状, 表皮细胞和蜡质层形态亦相似.短脚锦鸡儿(Caragana brachypoda )和狭叶锦鸡儿(Caragana stenophylla )表皮毛为柳叶带乳突型, 上表皮气孔器深陷, 气孔器为不等型或不规则型, 气孔口三角形或多边形.红花锦鸡儿(Caragana rosea )为明显不同的类群, 表皮无绒毛结构, 且蜡质层形态不同于以上几种的鳞片状, 而是紧贴表皮的膜片层结构或颗粒状. ...
植物生态解剖学研究发展
1
1993
... 植物叶片适应环境的形态结构变化是多方面的.在众多生态因子中, 叶片受水分、温度和光因子的影响最为显著(王勋陵和王静, 1989 ; 王勋陵, 1993 ; Aronne & de Micco 2001 ; Arena et al ., 2008 ; Jaleel et al ., 2009a , 2009b ), 主要表现为叶形的变化、叶片表皮附属物的变化、叶片的厚度及解剖结构的差异(Talukdar, 2015 ).对于荒漠植物来说, 夏季极端干旱时它们可能同时遭受以上各种环境因子的胁迫.但其最主要的限制因子为水分, 因而表现出典型的旱生植物形态结构特征.由于遗传因子与环境因子的共同作用, 在旱生植物结构的适应性上出现了两种演化趋势, 即趋同性和趋异性, 也就是说, 在相同环境下的植物演化出相似的形态结构适应性, 而不同环境下的同类植物演化出不同的形态结构(Lakusic et al ., 2010 ).因此, 要系统分析荒漠植物叶片形态结构特征的适应性变化, 需要将一个区域不同植物和不同梯度生境下的同一植物做全面分析. ...
从旱生植物叶结构探讨其生态适应的多样性
2
1999
... 干旱半干旱区的荒漠环境属于一种极端环境, 降水稀少, 气候干燥, 昼夜温差大, 地面温度高, 土壤贫瘠, 风沙活动强烈……这些恶劣条件无时不在限制着荒漠植物的生存和繁衍.但即使在这种条件下, 仍有许多植物分布于干旱半干旱区, 且能很好地繁衍生息.那么, 这些植物有何特殊的形态结构来适应环境呢?如何通过植物形态结构特征判断其适应能力? 这些问题引发不少科学家对荒漠植物研究的极大关注.Schow提出“旱生植物”概念已有100多年了. 国内外不少学者研究过旱生植物的形态结构(Shield, 1950 ; 王勋陵和马骥, 1999 ; 严巧娣等, 2008 ; Liu et al ., 2012 , 2015 ; Perrone et al ., 2015 ), 但以各自有限数量的旱生植物形态结构描述为主, 至今未见对较大数量旱生植物叶片表皮微形态与结构特征进行数量化整理, 并对其类型划分、变化规律、演化趋势、适应对策等重要问题进行系统讨论的 报道. ...
... 植物叶片形态结构与抗逆性关系的研究表明, 植物叶片小、表皮附属物密度大、且表皮皱缩程度高, 则抗旱性强(王勋陵和马骥, 1999 ; 周智斌和李培军, 2002 ; Bargel et al ., 2004 ; 迟丽华和宋凤斌, 2006 ; Adebowale et al ., 2014 ; 赵小仙等, 2014 ; 朱广龙等, 2016 ).通过比较荒漠植物叶片(或同化枝)的表皮微形态结构, 可以从表皮细胞形态, 表皮附属物如绒毛、蜡质、气孔等的形态、密度、结构和分布特征来进行环境适应性分析.根据表皮微形态结构和附属物与抗逆性的关系, 将荒漠植物适应环境胁迫的表皮微形态分为以下6种主要类型. ...
2
1989
... 植物作为生态系统的第一生产者, 受环境变化的影响是直接而多方面的.叶片是植物进化过程中对环境变化最敏感且可塑性较大的器官, 在不同的选择压力下形成了各种适应类型(Hickey, 1984 ; 祁如虎等, 1998 ; Gielwanowska et al ., 2005 ), 其形态结构特征最能体现环境因子的影响或植物对环境的适应(王勋陵和王静, 1989 ; Krober et al ., 2015 ). ...
... 植物叶片适应环境的形态结构变化是多方面的.在众多生态因子中, 叶片受水分、温度和光因子的影响最为显著(王勋陵和王静, 1989 ; 王勋陵, 1993 ; Aronne & de Micco 2001 ; Arena et al ., 2008 ; Jaleel et al ., 2009a , 2009b ), 主要表现为叶形的变化、叶片表皮附属物的变化、叶片的厚度及解剖结构的差异(Talukdar, 2015 ).对于荒漠植物来说, 夏季极端干旱时它们可能同时遭受以上各种环境因子的胁迫.但其最主要的限制因子为水分, 因而表现出典型的旱生植物形态结构特征.由于遗传因子与环境因子的共同作用, 在旱生植物结构的适应性上出现了两种演化趋势, 即趋同性和趋异性, 也就是说, 在相同环境下的植物演化出相似的形态结构适应性, 而不同环境下的同类植物演化出不同的形态结构(Lakusic et al ., 2010 ).因此, 要系统分析荒漠植物叶片形态结构特征的适应性变化, 需要将一个区域不同植物和不同梯度生境下的同一植物做全面分析. ...
Function of essential oil-secreting glandular trichome in aromatic plants of the Lamiaceae—A review
2
1993
... 大多数荒漠植物叶片的一个显著特点是表皮被绒毛, 叶表皮毛的出现与长期的干旱气候有关, 是植物对干旱环境的适应特征(Retamales & Scharaschkin, 2015 ).植物叶片表皮毛分两种: 腺毛和非腺毛(Werker, 1993 ; Navarro & Oualidi, 1999 ).非腺毛的主要功能与植物的防御有关.目前确认的植物叶片表皮非腺毛具有的生理生态功能主要有: 减轻紫外辐射(Karabourniotis et al ., 1992 ; Gregoriou et al ., 2007 )、降低边界层的传导(Wuenscher, 1970 ; Kenzo et al ., 2008 )、降低叶温(Ehleringer & Mooney, 1978 ), 以及减少对太阳辐射的接收(Ehleringer et al ., 1976 )等.这些功能决定了干旱区荒漠植物抗逆能力的大小.干旱地区的叶表皮毛密度比湿润地区的高(Smith & Nobel, 1977 ; Aronne & de Micco, 2001 ), 就单个物种而言, 表皮毛密度与温度和辐射密切相关(Perez-Estrada et al ., 2000 ).在热带雨林树种中, 浓密的表皮毛很少见(Turner, 2001 ), Roth (1984)调查的250种树中仅有15种有较厚的表皮毛, 且较厚的表皮毛一般在干旱、高温和强辐射条件下产生.而在干旱半干旱的荒漠区, 大多数植物都具有浓密的表皮毛.显然, 荒漠植物叶上密生的表皮毛对于植物的生活有重要的意义.从Ehleringer等 (1976)、Ehleringer和Bjorkman (1978)、Ehleringer和Mooney (1978)研究的荒漠灌木Encelia farinosa 的叶表皮毛对叶片光谱特性和光合、蒸腾等生理活动的影响来看, 沙漠毒菊在不同生长季生长的叶片具有不同数量的表皮毛, 表皮毛的变化足够使叶片对光合有效辐射的吸收从81%降到29%, 对总的太阳辐射的吸收从46%降到16%, 而亲缘关系较近却没有表皮毛的同属物种Encelia californiea 的有效光合辐射吸收率为84%.同时, 由于对太阳辐射吸收的减少, 导致叶面温度和植物蒸腾速率的降低.荒漠环境气温经常超过40 ℃, 而植物最适宜的光合温度是25 ℃, 当温度超过35 ℃时植物的光合速率会突降, 但较厚的叶表皮毛能使叶温显著低于气温而接近光合适宜温度, 同时也避免植物达到叶片致死温度.但叶表皮毛也有不利的一面, 表皮毛的覆盖降低了光合速率, 因此同一植物在不同的生境和生长季长出的叶片会出现表皮毛数量不同的特征, 这也是植物适应环境的一种权衡策略(Kenzo et al ., 2008 ).从本研究中的117种荒漠植物叶表皮特征来看, 片状叶片基本被有表皮毛, 并有不少叶片的表皮毛数量较多, 直接增加了叶片的边界层厚度, 个别植物的上下表皮毛厚度远超于叶片厚度, 如表皮微形态抗逆适应主要类型的第一类植物中的柠条锦鸡儿、卷叶锦鸡儿和沙枣等.可见荒漠植物中叶片表皮毛的作用是相当重要的, 它们不仅能调节植物的生理功能, 而且可抵御风沙等机械损伤的直接伤害.这些物种在不同的自然生境中其表皮毛数量亦不同(数据省略), 说明生态环境深刻地影响着植物形态结构的形成与变异. ...
... 而对于腺毛来说, 其主要作为一个油性物质的合成、分泌和累积的器官, 有效地防御动物的捕食、病菌入侵以及物种竞争(Werker, 1993 ; Wagner et al ., 2003 ; Kremer et al ., 2014 ; Munien et al ., 2015 ).实际上, 腺毛也是次生代谢产物的一个独立的细胞工厂(Tissier, 2012 ).腺毛的形态随植物的组织器官和种类不同而呈多样化, 其形成和分泌的主要油性物质及其产物组成受遗传和环境两方面的因素控制(Bosabalidis & Kofidis, 2002 ; Shafiee-Hajiabad et al ., 2014 ), 腺毛结构的多样性也可以为植物分类学提供依据(Gairola et al ., 2009 ; Rusydi et al ., 2013 ).对117种荒漠植物叶片表皮扫描发现: 不少旱生植物除具有非腺体表皮毛外, 还同时具有腺毛, 并且有些腺毛体积较大, 如蒙古莸、甘草、香青兰、星毛短舌菊、百里香等.显然, 对于具有腺毛的荒漠植物, 腺毛具有重要的功能.但目前关于不同植物腺毛的形态、形成以及功能的研究还很有局限性.那么, 这些腺毛在植物适应荒漠环境的过程中其形态结构与环境因子有何关系、主要分泌哪些化学物质、发挥什么样的功能?这些问题亟待下一步的深入研究. ...
The effect of leaf hairs of Verbascum thapsus on leaf energy exchange
1
1970
... 大多数荒漠植物叶片的一个显著特点是表皮被绒毛, 叶表皮毛的出现与长期的干旱气候有关, 是植物对干旱环境的适应特征(Retamales & Scharaschkin, 2015 ).植物叶片表皮毛分两种: 腺毛和非腺毛(Werker, 1993 ; Navarro & Oualidi, 1999 ).非腺毛的主要功能与植物的防御有关.目前确认的植物叶片表皮非腺毛具有的生理生态功能主要有: 减轻紫外辐射(Karabourniotis et al ., 1992 ; Gregoriou et al ., 2007 )、降低边界层的传导(Wuenscher, 1970 ; Kenzo et al ., 2008 )、降低叶温(Ehleringer & Mooney, 1978 ), 以及减少对太阳辐射的接收(Ehleringer et al ., 1976 )等.这些功能决定了干旱区荒漠植物抗逆能力的大小.干旱地区的叶表皮毛密度比湿润地区的高(Smith & Nobel, 1977 ; Aronne & de Micco, 2001 ), 就单个物种而言, 表皮毛密度与温度和辐射密切相关(Perez-Estrada et al ., 2000 ).在热带雨林树种中, 浓密的表皮毛很少见(Turner, 2001 ), Roth (1984)调查的250种树中仅有15种有较厚的表皮毛, 且较厚的表皮毛一般在干旱、高温和强辐射条件下产生.而在干旱半干旱的荒漠区, 大多数植物都具有浓密的表皮毛.显然, 荒漠植物叶上密生的表皮毛对于植物的生活有重要的意义.从Ehleringer等 (1976)、Ehleringer和Bjorkman (1978)、Ehleringer和Mooney (1978)研究的荒漠灌木Encelia farinosa 的叶表皮毛对叶片光谱特性和光合、蒸腾等生理活动的影响来看, 沙漠毒菊在不同生长季生长的叶片具有不同数量的表皮毛, 表皮毛的变化足够使叶片对光合有效辐射的吸收从81%降到29%, 对总的太阳辐射的吸收从46%降到16%, 而亲缘关系较近却没有表皮毛的同属物种Encelia californiea 的有效光合辐射吸收率为84%.同时, 由于对太阳辐射吸收的减少, 导致叶面温度和植物蒸腾速率的降低.荒漠环境气温经常超过40 ℃, 而植物最适宜的光合温度是25 ℃, 当温度超过35 ℃时植物的光合速率会突降, 但较厚的叶表皮毛能使叶温显著低于气温而接近光合适宜温度, 同时也避免植物达到叶片致死温度.但叶表皮毛也有不利的一面, 表皮毛的覆盖降低了光合速率, 因此同一植物在不同的生境和生长季长出的叶片会出现表皮毛数量不同的特征, 这也是植物适应环境的一种权衡策略(Kenzo et al ., 2008 ).从本研究中的117种荒漠植物叶表皮特征来看, 片状叶片基本被有表皮毛, 并有不少叶片的表皮毛数量较多, 直接增加了叶片的边界层厚度, 个别植物的上下表皮毛厚度远超于叶片厚度, 如表皮微形态抗逆适应主要类型的第一类植物中的柠条锦鸡儿、卷叶锦鸡儿和沙枣等.可见荒漠植物中叶片表皮毛的作用是相当重要的, 它们不仅能调节植物的生理功能, 而且可抵御风沙等机械损伤的直接伤害.这些物种在不同的自然生境中其表皮毛数量亦不同(数据省略), 说明生态环境深刻地影响着植物形态结构的形成与变异. ...
Responses of leaf stomatal density to water status and its relationship with photosynthesis in a grass
1
2008
... 植物叶片对环境的响应首先是叶片表皮对环境的感应和变化(Pal et al ., 2002 ; Bargel et al ., 2004 ; Koch et al ., 2009 ; Adebowale et al ., 2014 ).生长于干燥的气候和土壤条件下的荒漠植物, 叶表皮细胞会变小, 切向壁加厚, 具有内皮层.叶表皮角质膜厚度增加, 角质层由光滑变得粗糙, 能够反射阳光, 减少叶片表面空气的流动, 降低蒸腾作用, 防止水分过度丧失(贺金生等, 1994 ; 杨明博等, 2007 ).气孔的分布特征、密度和面积等都会受到影响, 气孔多分布于叶片下表皮, 这种分布模式既可促进植物与外界环境进行气体交换, 又能保持水分.随着环境中水分和湿度的减少, 气孔密度增加, 气孔面积则向小型化发展, 气孔多下陷形成气孔窝或其上有突出的角质膜(Sam et al ., 2000 ; Bosabalidis & Kofidis, 2002 ; 高建平等, 2003 ; Xu & Zhou, 2008 ; Gan et al ., 2010 ; de Micco & Aronne, 2012 ).叶表皮附属物(如表皮毛和表面疣状突起物)也发生变化, 它们可有效地避免叶肉组织受损(Gairola et al ., 2009 ; Koch et al ., 2009 ; Gomes et al ., 2011 ; Noman et al ., 2012 ), 表皮蜡质(waxes)晶体形成二维或三维结构, 具有降低表皮的可湿性以及反射强光等作用(Wagner et al ., 2003 ; Koch & Ensikat, 2008 ). ...
五种C4 荒漠植物光合器官中含晶细胞的比较分析
1
2008
... 干旱半干旱区的荒漠环境属于一种极端环境, 降水稀少, 气候干燥, 昼夜温差大, 地面温度高, 土壤贫瘠, 风沙活动强烈……这些恶劣条件无时不在限制着荒漠植物的生存和繁衍.但即使在这种条件下, 仍有许多植物分布于干旱半干旱区, 且能很好地繁衍生息.那么, 这些植物有何特殊的形态结构来适应环境呢?如何通过植物形态结构特征判断其适应能力? 这些问题引发不少科学家对荒漠植物研究的极大关注.Schow提出“旱生植物”概念已有100多年了. 国内外不少学者研究过旱生植物的形态结构(Shield, 1950 ; 王勋陵和马骥, 1999 ; 严巧娣等, 2008 ; Liu et al ., 2012 , 2015 ; Perrone et al ., 2015 ), 但以各自有限数量的旱生植物形态结构描述为主, 至今未见对较大数量旱生植物叶片表皮微形态与结构特征进行数量化整理, 并对其类型划分、变化规律、演化趋势、适应对策等重要问题进行系统讨论的 报道. ...
鄂尔多斯高原不同生境条件下中间锦鸡儿植物叶片表皮特征及遗传多样性变化分析
1
2007
... 植物叶片对环境的响应首先是叶片表皮对环境的感应和变化(Pal et al ., 2002 ; Bargel et al ., 2004 ; Koch et al ., 2009 ; Adebowale et al ., 2014 ).生长于干燥的气候和土壤条件下的荒漠植物, 叶表皮细胞会变小, 切向壁加厚, 具有内皮层.叶表皮角质膜厚度增加, 角质层由光滑变得粗糙, 能够反射阳光, 减少叶片表面空气的流动, 降低蒸腾作用, 防止水分过度丧失(贺金生等, 1994 ; 杨明博等, 2007 ).气孔的分布特征、密度和面积等都会受到影响, 气孔多分布于叶片下表皮, 这种分布模式既可促进植物与外界环境进行气体交换, 又能保持水分.随着环境中水分和湿度的减少, 气孔密度增加, 气孔面积则向小型化发展, 气孔多下陷形成气孔窝或其上有突出的角质膜(Sam et al ., 2000 ; Bosabalidis & Kofidis, 2002 ; 高建平等, 2003 ; Xu & Zhou, 2008 ; Gan et al ., 2010 ; de Micco & Aronne, 2012 ).叶表皮附属物(如表皮毛和表面疣状突起物)也发生变化, 它们可有效地避免叶肉组织受损(Gairola et al ., 2009 ; Koch et al ., 2009 ; Gomes et al ., 2011 ; Noman et al ., 2012 ), 表皮蜡质(waxes)晶体形成二维或三维结构, 具有降低表皮的可湿性以及反射强光等作用(Wagner et al ., 2003 ; Koch & Ensikat, 2008 ). ...
Scanning electron microscopy of the leaf epidermis in Arundinarieae(Poaceae: Bambusoideae): Evolutionary implications of selected micromorphological features
1
2014
... 植物叶表皮微形态特征已用于解决物种分类问题、建立物种进化关系、阐明表皮微形态结构的生态适应性等多种研究目的(Dickison, 2000 ; Scatena et al ., 2005 ; Stenglein et al ., 2005 ; Aliero et al ., 2006 ; Chen et al ., 2010 ; Zhou & Xia, 2012 ; Adebowale et al ., 2014 ; Deng et al ., 2014 ; Zhang et al ., 2014 ; Francisco et al ., 2015 ; 朱广龙等, 2016 ).为了探讨荒漠植物叶片表皮微形态结构的适应特征及其亲缘关系, 选择分布于同一区域的不同植物和不同降水量梯度生境下的同一植物, 利用扫描电镜观察分析了117种荒漠植物叶片的表皮微形态结构, 旨在阐明荒漠植物对逆境的适应机理及其演化趋势, 并为优良固沙植物的筛选提供理论依据. ...
1
2012
... 荒漠植物由于长期适应所生长的自然环境, 发展了一套最适宜自身生长发育的形态结构特征, 并采取各种途径来抵御或忍耐干旱胁迫对它的影响, 因而具有一定程度的耐旱性.这种耐旱性是一种复合性状, 包括植物的形态结构特征、水分生理特征、生理生化反应、组织细胞、光合器官以及原生质结构等特征的综合反映(Klich, 2000 ; Chaves et al ., 2003 ; Ogle & Reynolds, 2004 ).不同的荒漠植物对水分和不利环境的适应和耐受程度不同, 适应方式多种多样, 按照不同的分类标准, 荒漠植物有多种适应类型.通常的分类标准包括耐旱程度、耐旱方式、叶片形态、生活史、光合碳代谢途径等(赵哈林, 2012 ).各种类型的植物又有各自的适应特征.本研究是针对我国干旱半干旱区荒漠植物叶表皮微形态结构特征的综合分析, 主要集中于该区域的建群种灌木物种.根据荒漠植物叶表皮微形态基本结构特征和表皮附属物的类型与数量, 将荒漠植物表皮微形态类型直观地划分为11种, 而将抗逆适应微形态结构划分为6种类型.荒漠植物叶表皮微形态结构类型复杂, 反映了植物自身的遗传特性及其与环境的密切关系.植物叶片表皮微形态主要由遗传因素决定, 但表皮附属物绒毛数量, 蜡质形态与厚度, 气孔的形态、密度以及分布特征等与环境因子密切相关, 是植物对环境的响应(Barthlott, 1981 ; Casson & Hetherington, 2010 ; Ghahremaninejad et al ., 2012 ).因此, 基于这些表皮主要附属物特征对荒漠植物的表皮微形态类型和抗逆适应类型进行划分, 能够从微观形态结构特征进一步揭示物种的亲缘关系和表皮微形态结构的生态适应意义. ...
3个地理种群蒙古沙拐枣同化枝解剖结构及抗旱性比较
1
2014
... 植物叶片形态结构与抗逆性关系的研究表明, 植物叶片小、表皮附属物密度大、且表皮皱缩程度高, 则抗旱性强(王勋陵和马骥, 1999 ; 周智斌和李培军, 2002 ; Bargel et al ., 2004 ; 迟丽华和宋凤斌, 2006 ; Adebowale et al ., 2014 ; 赵小仙等, 2014 ; 朱广龙等, 2016 ).通过比较荒漠植物叶片(或同化枝)的表皮微形态结构, 可以从表皮细胞形态, 表皮附属物如绒毛、蜡质、气孔等的形态、密度、结构和分布特征来进行环境适应性分析.根据表皮微形态结构和附属物与抗逆性的关系, 将荒漠植物适应环境胁迫的表皮微形态分为以下6种主要类型. ...
Leaf epidermal features of Lithocarpus (Fagaceae) from China and their systematic significance
1
2012
... 植物叶表皮微形态特征已用于解决物种分类问题、建立物种进化关系、阐明表皮微形态结构的生态适应性等多种研究目的(Dickison, 2000 ; Scatena et al ., 2005 ; Stenglein et al ., 2005 ; Aliero et al ., 2006 ; Chen et al ., 2010 ; Zhou & Xia, 2012 ; Adebowale et al ., 2014 ; Deng et al ., 2014 ; Zhang et al ., 2014 ; Francisco et al ., 2015 ; 朱广龙等, 2016 ).为了探讨荒漠植物叶片表皮微形态结构的适应特征及其亲缘关系, 选择分布于同一区域的不同植物和不同降水量梯度生境下的同一植物, 利用扫描电镜观察分析了117种荒漠植物叶片的表皮微形态结构, 旨在阐明荒漠植物对逆境的适应机理及其演化趋势, 并为优良固沙植物的筛选提供理论依据. ...
我国旱生植物的形态解剖学研究
1
2002
... 植物叶片形态结构与抗逆性关系的研究表明, 植物叶片小、表皮附属物密度大、且表皮皱缩程度高, 则抗旱性强(王勋陵和马骥, 1999 ; 周智斌和李培军, 2002 ; Bargel et al ., 2004 ; 迟丽华和宋凤斌, 2006 ; Adebowale et al ., 2014 ; 赵小仙等, 2014 ; 朱广龙等, 2016 ).通过比较荒漠植物叶片(或同化枝)的表皮微形态结构, 可以从表皮细胞形态, 表皮附属物如绒毛、蜡质、气孔等的形态、密度、结构和分布特征来进行环境适应性分析.根据表皮微形态结构和附属物与抗逆性的关系, 将荒漠植物适应环境胁迫的表皮微形态分为以下6种主要类型. ...
酸枣叶表皮微形态对不同生态环境的适应特征
2
2016
... 植物叶表皮微形态特征已用于解决物种分类问题、建立物种进化关系、阐明表皮微形态结构的生态适应性等多种研究目的(Dickison, 2000 ; Scatena et al ., 2005 ; Stenglein et al ., 2005 ; Aliero et al ., 2006 ; Chen et al ., 2010 ; Zhou & Xia, 2012 ; Adebowale et al ., 2014 ; Deng et al ., 2014 ; Zhang et al ., 2014 ; Francisco et al ., 2015 ; 朱广龙等, 2016 ).为了探讨荒漠植物叶片表皮微形态结构的适应特征及其亲缘关系, 选择分布于同一区域的不同植物和不同降水量梯度生境下的同一植物, 利用扫描电镜观察分析了117种荒漠植物叶片的表皮微形态结构, 旨在阐明荒漠植物对逆境的适应机理及其演化趋势, 并为优良固沙植物的筛选提供理论依据. ...
... 植物叶片形态结构与抗逆性关系的研究表明, 植物叶片小、表皮附属物密度大、且表皮皱缩程度高, 则抗旱性强(王勋陵和马骥, 1999 ; 周智斌和李培军, 2002 ; Bargel et al ., 2004 ; 迟丽华和宋凤斌, 2006 ; Adebowale et al ., 2014 ; 赵小仙等, 2014 ; 朱广龙等, 2016 ).通过比较荒漠植物叶片(或同化枝)的表皮微形态结构, 可以从表皮细胞形态, 表皮附属物如绒毛、蜡质、气孔等的形态、密度、结构和分布特征来进行环境适应性分析.根据表皮微形态结构和附属物与抗逆性的关系, 将荒漠植物适应环境胁迫的表皮微形态分为以下6种主要类型. ...
 , 李新荣, 李蒙蒙, 刘丹, 张雯莉
, 李新荣, 李蒙蒙, 刘丹, 张雯莉


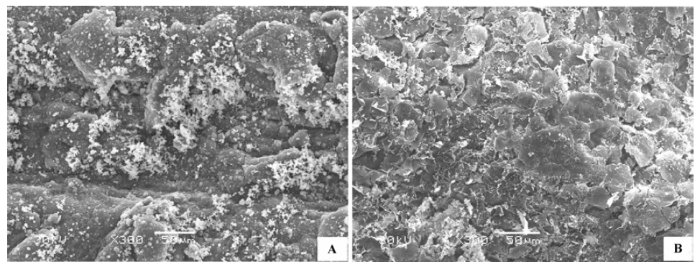
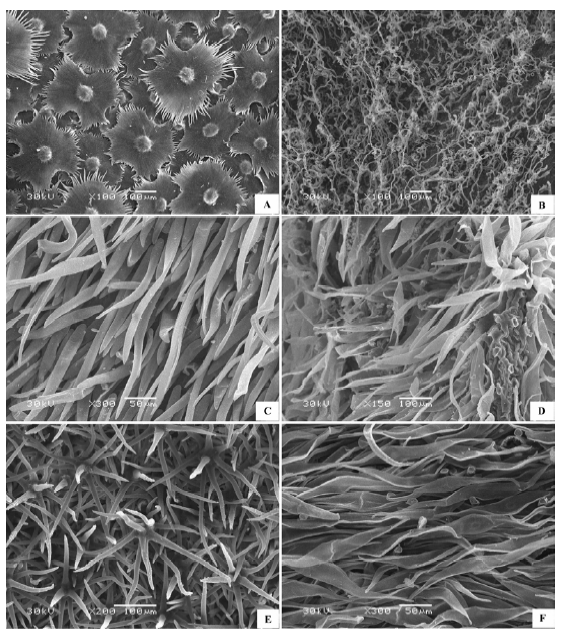
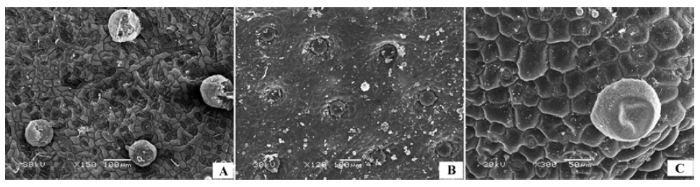
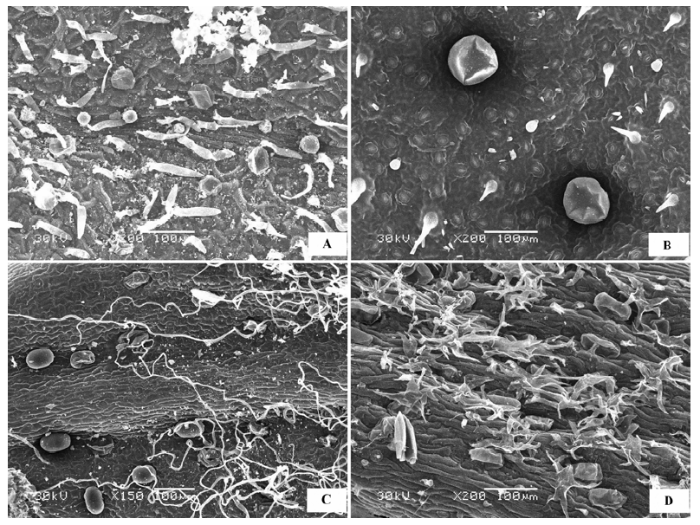
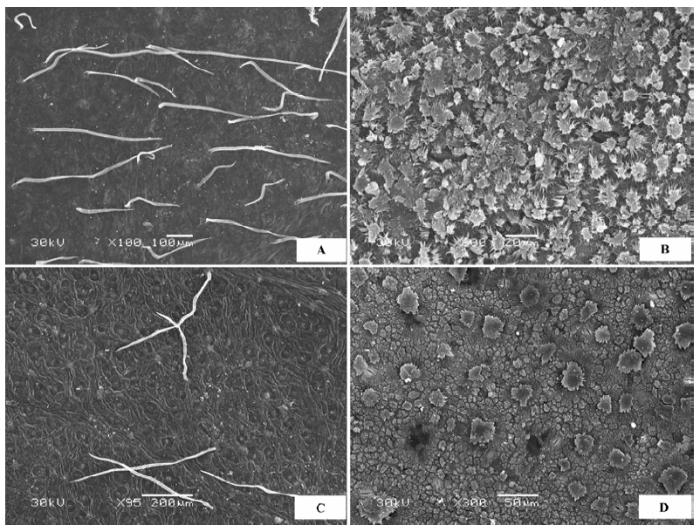
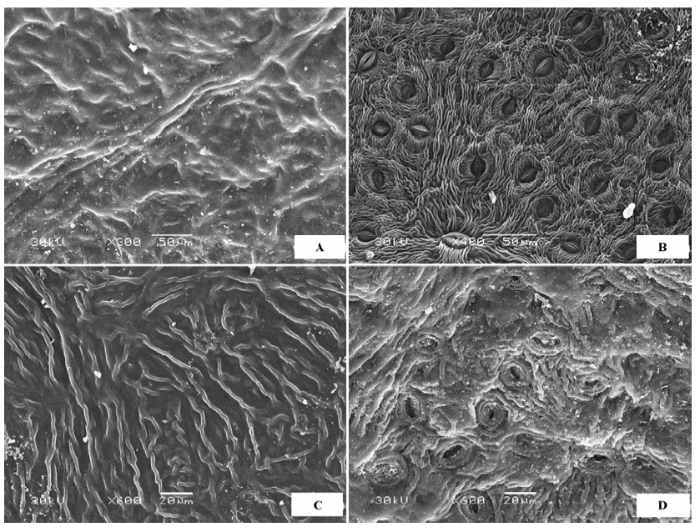
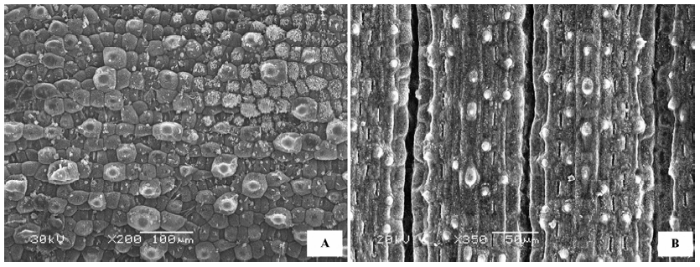
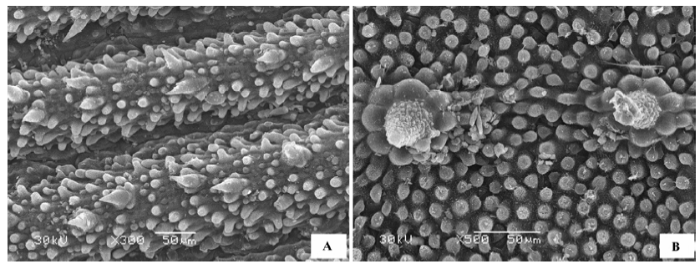
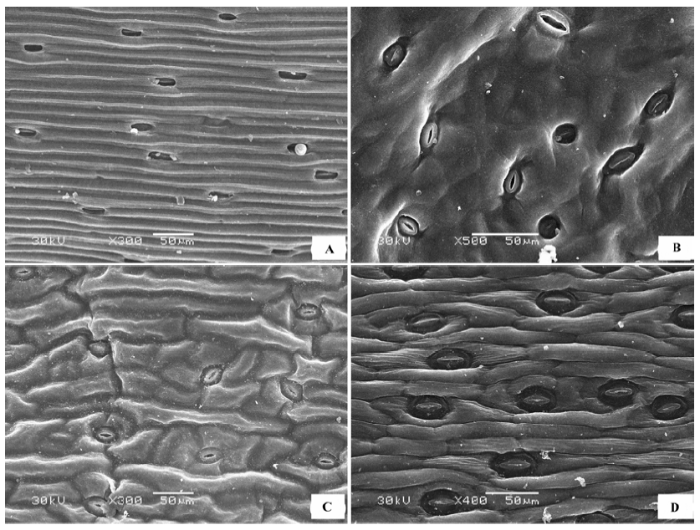
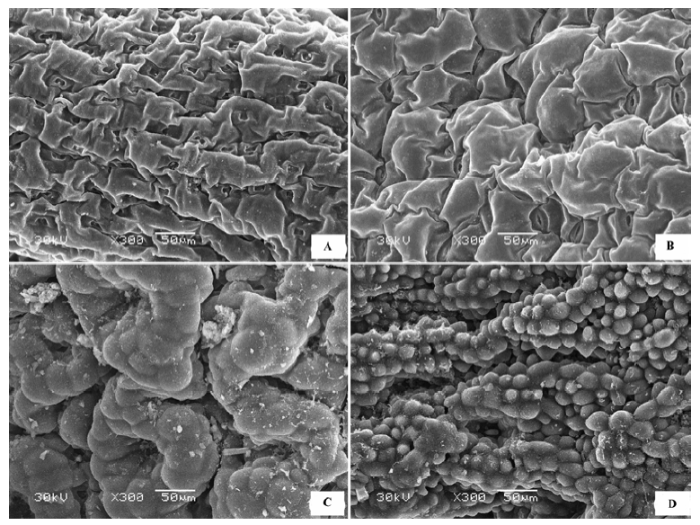
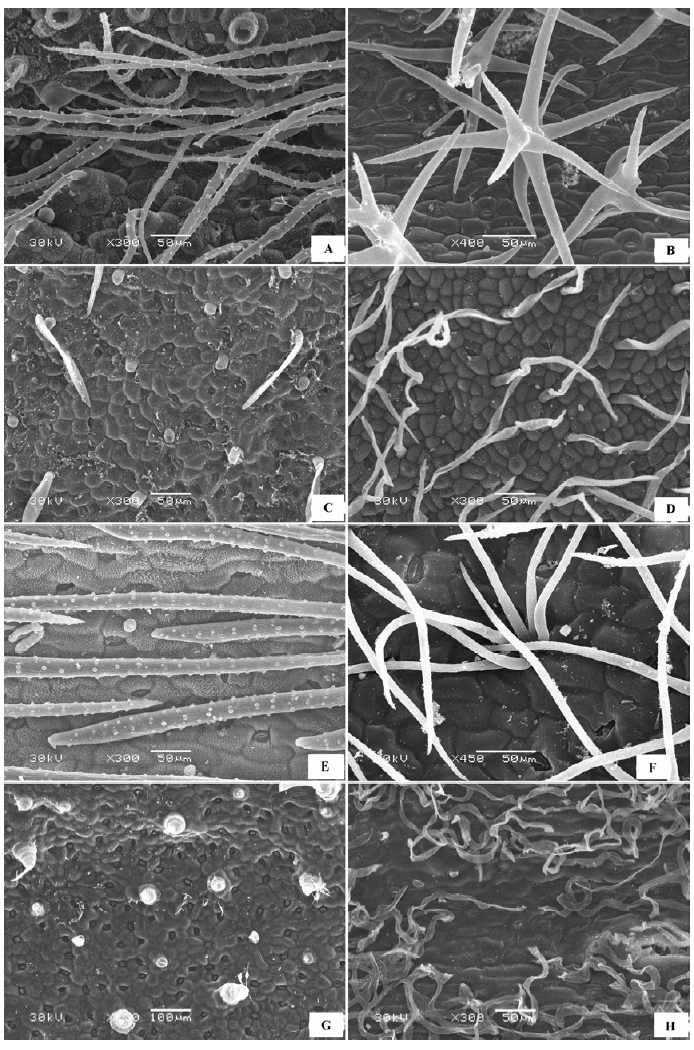
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}