森林是陆地生态系统的重要组成部分, 其植被碳(C)库约占全球植被C库的80% (Köster et al ., 2015 )。倒木是森林生态系统的重要组成部分, 对于维持生物多样性以及林木的更新具有重要作用; 同时, 倒木也是重要的养分库和C库, 其养分的缓慢释放, 对于提高林地的生产力有重要意义(Harmon et al ., 1986 ; Laiho & Prescott, 2004 )。倒木在物理、化学和生物(特别是微生物、昆虫)等作用下进行的分解过程不仅可为分解者(如微生物、无脊椎动物、鸟类等)提供生存环境和必需的营养元素(Sollins, 1982 ), 而且能够释放大量的化学元素到土壤中, 丰富土壤有机质含量和种类, 促进土壤结构和功能的发育(Spears et al ., 2003 )。影响倒木分解的因素有基质质量、分解者活性以及环境因素等(如温度、水分、气体)(Harmon et al ., 1986)。而在同一环境下, 不同树种间倒木分解速率也有很大差异, 这与倒木自身的物理和化学特性有关(van Geffen et al ., 2010 )。含水率和木材密度是影响倒木分解的主要物理因素, 因而受到越来越多的关注。倒木含水率是影响倒木呼吸的重要因子, 当它超过一定的阈值后, 就会影响微生物的活动, 从而抑制倒木的分解(Bond-Lamberty et al ., 2002 )。木材分解是一个复杂的过程, 通常根据单位时间内木材质量损失或木材密度的变化, 可以定量地算出其分解速率; 木材密度与微生物侵入倒木进行分解活动的程度有密切关系, 较低密度的木质残体更容易被分解和具有更高的呼吸速率(Johnston & Crossley, 1996 ; Chamber et al ., 2001 )。影响倒木分解的化学因素主要是其易分解成分和难分解有机成分(如木质素等)的含量, 其中氮(N)、磷(P)、木质素、纤维素、C:N、C:P等是常见的衡量倒木质量的指标(Berg et al ., 2000 ), 它们不仅可以表征倒木分解过程中基质的质量变化和养分限制状况(Wassen et al ., 2005 ), 而且可以指示物质循环和能量流动的方向(Zhou et al ., 2007 )。
近年来, 有关森林倒木物理化学性质的研究引起了广泛的关注。大部分研究结果表明, 随着腐烂等级增加, 倒木C含量呈下降趋势, N、P含量则与之相反(Palviainen et al ., 2008 ; 常晨晖等, 2015 )。很多研究认为物理化学性质的变化还与树种有关。如Köster等(2015)对同一地区主要倒木树种进行C、N含量和密度的测定发现, 欧洲云杉(Picea abies )倒木的C含量随着分解程度加深而增加, 但是欧洲山杨(Populus tremula )倒木的C含量随着分解程度加深呈下降趋势, Alnus glutinosa 倒木的C含量则变化不大; 随着分解程度加深, 不同树种的木材密度均下降, 但欧洲云杉、Alnus glutinosa 以及欧洲赤松(Pinus sylvestris )比其他3种阔叶树种下降程度小。以往对倒木物理化学性质的研究大多集中在不同腐烂等级和树种对倒木物理化学性质的影响, 而径级对倒木物理化学性质影响的研究尚处于初步探索阶段。通常情况下, 小径级倒木(胸径(DBH ) < 10 cm)在森林生态系统中的数量巨大且分解较快, 有利于有机物质矿化到土壤中被土壤吸收和利用(Moroni & Ryan, 2010 )。相对于小径级倒木, 大径级倒木心材比例较大, 由于心材储存了大量能源物质和难分解组分(Schwarze, 2007 ), 可能有较高的C含量和相对较低的养分含量, 但其结构疏松, 易于分解者活动和居住, 随着分解程度加深, 物理化学性质特征更加复杂(常晨晖等, 2015 )。Harmon等(1986)提出倒木中物理化学性质的变化可能与倒木的径级有关。吕明和等(2006)在对鼎湖山锥(Castanopsis chin- ensis )的粗木质残体的分解和元素动态分析中也指出, 在分解过程中直径5-10 cm的倒木中的N、P含量持续升高, 而直径10-20 cm的倒木中的N、P含量呈下降趋势。Sakai等(2012)在对日本扁柏(Chamae- cyparis obtusa )和日本柳杉(Cryptomeria japonica )的倒木密度以及C、N含量测定的研究中发现, 随着径级增加, 倒木密度随之下降, 且径级对处于腐烂等级I和II的两种倒木的密度影响显著。由此可见, 倒木中物理化学性质的变化不仅受到腐烂等级和径级的影响, 而且可能还受两者产生的交互作用的影响。
目前, 我国部分学者对长白山红松(Pinus koraiensis )暗针叶林(杨丽韫和代力民, 2002 )、大兴安岭落叶松(Larix gmelinii )林(辛魏巍等, 2012 )、鼎湖山季风常绿阔叶林(吕明和等, 2006 )以及亚高山针叶林(常晨晖等, 2015 )等森林类型中主要组成树种的倒木在分解过程的物理化学性质变化趋势进行了研究和评述, 而缺乏径级对倒木物理化学性质影响的研究。对不同径级和腐烂等级的倒木进行物理化学性质分析才能准确地量化和预测倒木C储量和养分含量, 同时能够更深入地了解倒木物理性质与化学性质之间的关系。阔叶红松林是我国东北东部山区的地带性顶极植被, 对其倒木物种组成、数量(刘妍妍和金光泽, 2010a )、贮量特征(代力民等, 2000 )以及分布格局(刘妍妍和金光泽, 2010b )曾进行过系统研究, 然而还缺乏倒木分解动态的详细研究。本研究以典型阔叶红松林的建群种红松倒木为研究对象, 探讨其不同径级与不同腐烂等级倒木心材和边材物理化学性质变化特征, 为未来预测倒木C贮量和养分含量以及生产力变化进行合理的评估提供重要的科学依据和基础数据。
1 材料和方法
1.1 研究区概况
试验地设在黑龙江省凉水国家级自然保护区(128.89° E, 47.18° N)。该保护区位于黑龙江省伊春市带岭区境内, 属小兴安岭南部达里带岭支脉的东坡, 海拔高度280-707 m, 为典型的低山丘陵地貌。境内森林覆盖率96%, 现有原始成过熟林面积4100 hm2 , 其中原始阔叶红松林面积占80%, 蓄积量180万m3 。该地区属温带大陆性夏雨季风气候, 春季迟缓, 多大风, 降水少; 夏季短, 多雨水, 降水占全年降水量的60%以上; 秋季降温快, 多出现早霜; 冬季长且寒冷干燥。年平均气温-0.3 ℃, 年平均最高气温7.5 ℃, 年平均最低气温-6.6 ℃, 年降水量676 mm, 年蒸发量805 mm。地带性土壤为暗棕壤, 非地带性土壤为草甸土、沼泽土和泥炭土。本研究所选的样地位于保护区中段, 从凉水沟谷地到海拔600 m的中山山脊, 该群落类型以红松为优势种, 伴生的温性阔叶树种由水曲柳(Fraxinus mandschurica )、黄檗(Phellodendron amurense )、紫椴(Tilia amur- ensis )、色木槭(Acer mono )、硕桦(Betula costata )等树种组成, 寒温性树种以臭冷杉(Abies nephrol- epis )、红皮云杉(Picea koraiensis )、鱼鳞云杉(Picea jezoensis )为主, 灌木有毛榛(Corylus mandshurica )、东北山梅花(Philadelphus schrenkii )等。
该地区典型阔叶红松林的木质残体生物量为27.4 t·hm-2 , 碳贮量13.49 t·hm-2 ; 花楷槭(Acer ukur- unduense )和硕桦是阔叶木质残体的主要组成树种, 红松和臭冷杉是针叶木质残体的主要组成树种; 各树种木质残体的腐烂等级大致呈正态分布, 主要集中在II和III腐烂等级上; 除针叶树的V腐烂等级外, 各树种不同腐烂等级的木质残体数量都随着径级的增加呈减少的趋势(刘妍妍和金光泽, 2010a )。
1.2 实验方法
1.2.1 木质残体的基础特征调查
本研究在凉水自然保护区9 hm2 (300 m × 300 m)的阔叶红松林永久性固定样地上进行(Jin et al ., 2006 ), 将样地设成900个10 m × 10 m的小样方, 采用相邻网格法进行调查。对样地内所有胸径≥2 cm的木质残体进行调查, 确定木质残体的存在形式(干基折断、干中折断、枯立木、拔根倒、根桩和树段); 测量立木形式的胸径, 对倒伏的木质残体, 测量其中央直径和胸径; 采用Vertex III超声波测高仪(Haglöf Sweden AB, Långsele, Sweden)进行长度和高度的测定, 识别树种, 确定坐标位置, 并钉铝制树牌进行编号。采用5级分类系统进行腐烂等级的划分(闫恩荣等, 2005 )。
1.2.2 样品采集
倒木分4个径级, 分别为径级i (≤ 10.0 cm)、径级ii (10.1-30.0 cm)、径级iii (30.1-50.0 cm)、径级iv (>50.0 cm); 每个径级下5个腐烂等级各取3株倒木, 共计60株。为了使每个径级的样品具有代表性, 对于径级ii、iii的倒木, 分别在15 cm左右、20 cm左右、25 cm左右、35 cm左右、40 cm左右以及45 cm左右各取一株。基于前期调查, 2015年6月在样地对红松倒木进行随机取样, 采集的红松以拔根倒、干基折断(根桩<1 m)的倒木为主(目的是保证胸径位置在倒地部分), 且倒地部分的长度>1.3 m。测量的倒木直径大约位于离地面1.3 m处, 测量直径并确定测量处的腐烂等级后取样, 每株只取一次。对I-III腐烂等级的倒木, 截取5 cm厚的圆盘; 对于高度腐烂的倒木(IV和V), 用已知容积的铝盒采样。采用de Aza等(2011) 的方法将各腐烂等级的倒木样品分为心材、边材, 分别采集各个腐烂等级的倒木样品, 去除泥土、石块、根系、苔藓等附生物质, 做好标记后, 装于密封袋内保存, 带回实验室分析,共计120个样品。
倒木腐烂等级的划分按照闫恩荣等(2005)的粗木质残体(coarse woody debris, CWD) 5级分类系统(I级: 新鲜, 树木死亡不足1年; II级: 开始腐解, 小刀可刺进几mm; III级: 小刀可刺进2 cm; IV级: 小刀可刺进2-5 cm; V级: 小刀可任意刺穿木质体)。对于腐烂等级低(I-III)的倒木用胸径尺测量其直径, 对于腐烂等级高(IV和V)的倒木直径进行目测(Sefidi & Etemad, 2015 )。
1.2.3 室内分析测定
将采集的样品粉碎, 过60目筛, 于65 ℃条件下烘干至恒质量, 存放于干燥环境中待测。采用燃烧法通过multi N/C 3000分析仪(Analytik Jena AG, Jena, Germany)测定倒木全C含量; 采用消煮法通过Kjeltec TM 2300凯氏定氮仪(Foss Teactor AB, Hoganas, Sweden)测定倒木全N含量; 采用钼锑钪比色法通过722N分光光度计(上海精密科学仪器有限公司, 上海)测定其全P含量; 采用改进的Van Soest酸性洗涤木质素方法(He et al ., 2016 )测定其木质素和纤维素含量。
倒木的密度: (1)对于腐烂等级I-III的倒木, 采用排水法测定倒木样品的体积V, 即在量筒中倒入一定量的水(保证能完全淹没木块), 读出水面刻度体积V 1 , 然后将浸泡过的木块用薄片刀(体积可忽略)完全压入盛有水的量筒中且保持水不溢出, 读出刻度体积V 2 , V 2 -V 1 为木块体积V; 对于腐烂等级高(IV和V级)的倒木, 利用环刀进行体积测量, 即用腐烂的倒木将环刀填满(切忌压实), 环刀的体积即为样本倒木的体积V 。(2)将样品放在70 ℃的烘箱内烘干至恒质量G 。(3)倒木密度ρ (g·cm-3 ) = G /V 。
倒木的含水率: 用烘箱烘干倒木样品至恒质量, 记为G 1 , 根据鲜质量G 0 和干质量G 1 得出其相对含水率(%) = (G 0 - G 1 )/G 0 × 100%。
1.3 数据处理与统计方法
采用单因素方差分析分别检验倒木腐烂等级间、径级间以及心材和边材之间物理和化学性质的差异性, 并采用Duncan法进行多重比较, 显著性水平设定为p < 0.05; 采用双因素方差分析检验径级和腐烂等级对C、N、P、木质素、纤维素含量的影响。用Pearson相关分析法分析倒木心材和边材的含水率、密度与化学成分的相关关系。以上统计分析均使用SPSS 19.0统计软件完成, 使用SigmaPlot 12.5 软件进行图形绘制。
2 结果
2.1 腐烂等级、径级对倒木含水率、木材密度的影响
腐烂等级对倒木心材和边材的含水率和密度产生极显著影响(p < 0.001)(表1 )。随着腐烂等级增加, 倒木心材和边材的含水率呈上升趋势, 密度呈下降趋势。除腐烂等级III倒木心材的密度显著大于边材的密度外, 心材与边材的含水率以及密度在其余各腐烂等级、总体上均没有显著差异(p < 0.05)(图1A 、图1B )。
倒木心材和边材的密度在不同径级间具有显著差异(p < 0.05), 含水率没有显著差异(表1 )。心材和边材的密度均表现为: 径级i >径级ii >径级iv >径级iii, 且径级i倒木心材和边材的密度显著高于径级iii的密度(p < 0.05)。各径级的心材与边材的含水率以及密度均无显著差异(图1C 、图1D )。
图1 不同腐烂等级和径级下红松倒木的含水率和木材密度(平均值±标准误差, n = 3)。Total表示不同腐烂等级和径级下倒木心材和边材含水率、木材密度的总体平均值。不同小写字母表示倒木心材及边材不同腐烂等级间和径级间差异显著(p < 0.05); 不同大写字母表示倒木心材和边材间差异显著(p < 0.05)。
Fig. 1 Moisture content and wood density of logs of Pinus koraiensis at different decay classes and diameter classes (mean ± SE, n = 3). Total indicates total average value of moisture content and wood density of both heartwood and sapwood at different decay classes and diameter classes. Different lowercase letters indicate significant differences among different decay classes and diameter classes in the heartwood and sapwood (p < 0.05). Different uppercase letters indicate significant differences between heartwood and sapwood (p < 0.05).
2.2 腐烂等级、径级对倒木C、N、P、木质素、纤维素含量的影响
除心材C含量外, 腐烂等级显著影响倒木心材和边材C、N、P、纤维素和木质素的含量(p < 0.05)(表1 )。倒木边材C含量以及心材和边材N、P、木质素含量随着腐烂等级增加呈上升趋势, 而心材和边材纤维素含量具有相反的变化规律。边材腐烂等级V的C含量显著高于腐烂等级I-III (p < 0.05); 心材和边材N含量分别在腐烂等级III、IV达到最大值, 分别为: 2.41和3.19 mg.g-1 ; 心材腐烂等级III-V的P含量显著高于腐烂等级II, 边材腐烂等级IV、V的P含量显著高于其他腐烂等级; 心材腐烂等级I-III和边材腐烂等级I、II的纤维素含量显著高于其他腐烂等级; 心材和边材腐烂等级V木质素含量分别显著高于腐烂等级I-IV和I-III (p < 0.05)(图2 )。
腐烂等级V的N、P含量和腐烂等级IV的P、纤维素和木质素含量均表现为边材显著高于心材(p < 0.05), 总体平均N含量边材显著高于心材(p < 0.05), 而心材和边材的C、N、P以及纤维素、木质素的含量在其余各腐烂等级及总体平均含量上均没有显著差异(图2 )。
图2 不同腐烂等级下红松倒木的C、N、P、纤维素、木质素含量(平均值±标准误差, n = 3)。Total表示不同腐烂等级下倒木心材和边材C、N、P、纤维素和木质素的总体平均含量。不同小写字母表示倒木心材和边材在不同腐烂等级间差异显著(p < 0.05); 不同大写字母表示倒木心材和边材间差异显著(p < 0.05)。
Fig. 2 Contents of C, N, P, cellulose, lignin of logs of Pinus koraiensis at different decay classes (mean ± SE, n = 3). Total indicate total average content of C, N, P, cellulose and lignin of heartwood and sapwood at different decay classes. Different lowercase letters indicate significant differences among different decay classes in the heartwood and sapwood (p < 0.05). Different uppercase letters indicate significant differences between heartwood and sapwood (p < 0.05).
倒木心材N和P的含量在径级间差异显著(p < 0.05), 且倒木心材和边材木质素和纤维素含量受腐烂等级和径级交互作用影响显著(表1 )。心材N和P含量随着径级增加呈先升高后降低的趋势, 径级iii倒木N、P含量最高, 心材径级iii的N含量显著高于径级i, P含量显著高于其他径级(p < 0.05)。倒木心材和边材C含量、木质素、纤维素含量在不同径级间均无显著差异(图3 )。
径级i的N含量和径级ii的P含量表现为边材显著高于心材(p < 0.05), 总体平均N含量边材显著高于心材(p < 0.05), 心材和边材的C、N、P以及纤维素、木质素含量在其余各径级和总体平均含量上均没有显著差异。
图3 在不同径级下红松倒木C、N、P、纤维素和木质素含量(平均值±标准误差, n = 3)。Total表示不同径级下倒木心材和边材C、N、P、纤维素和木质素的总体平均含量。不同小写字母表示倒木心材和边材在不同径级间差异显著(p < 0.05); 不同大写字母表示倒木心材和边材间差异显著(p < 0.05)。
Fig. 3 Contents of C, N, P, cellulose, lignin of logs of Pinus koraiensis at different diameter classes (mean ± SE, n = 3). Total indicate total average content of C, N, P, cellulose, lignin of heartwood and sapwood at different diameter classes. Different lowercase letters indicate significant differences among different diameter classes in the heartwood and sapwood (p < 0.05). Different uppercase letters indicate significant differences between heartwood and sapwood (p < 0.05).
2.3 腐烂等级、径级对倒木C:N、C:P、N:P以及木质素:纤维素的影响
除心材C:P和N:P外, 倒木C:N、C:P、N:P以及木质素:纤维素在腐烂等级间存在显著差异(p < 0.05)。随着腐烂等级增加, 心材和边材的C:N值呈下降趋势, 其中心材和边材腐烂等级I的C:N值分别显著高于腐烂等级IV、V和III-V的C:N值(p < 0.05); 边材C:P值呈先升高后降低的趋势, 其中边材腐烂等级II的C:P值显著高于其他腐烂等级C:P值(p < 0.05); 边材N:P值呈先增加后减少的趋势, 在腐烂等级II时最高, 且腐烂等级II边材N:P值显著高于其他腐烂等级(p < 0.05); 心材和边材的木质素:纤维素值呈上升趋势, 心材和边材腐烂等级V的木质素:纤维素值均显著高于其他腐烂等级(p < 0.05)。腐烂等级IV和V的C:P值心材显著高于边材(p < 0.05)(表2 )。
倒木心材和边材C:N值、C:P值以及边材的N:P值在径级间存在显著差异(p < 0.05), 木质素:纤维素以及心材N:P值在径级间没有显著差异。心材和边材C:N值的动态变化基本呈“V”形,即径级i开始下降,径级iii的C:N值最低, 径级iv上升。心材和边材径级i的C:N值均分别显著高于径级ii、iii和径级iii (p < 0.05)。C:P值随着径级增加呈下降趋势, 径级i处C:P最高, 径级i心材C:P值显著大于径级iii、iv (p < 0.05), 径级i边材C:P值显著高于其他径级(p < 0.05)。随着径级增加, 心材和边材N:P值呈下降趋势, 径级i边材N:P值显著高于其他径级N:P值(p < 0.05)。径级ii心材C:P值显著高于边材C:P值(p < 0.05) (表3 )。
2.4 倒木物理性质与化学成分之间的关系
倒木物理性质与倒木化学成分表现出显著的相关性(表4 )。含水率与倒木心材和边材C、N、木质素含量以及边材P含量呈显著或极显著正相关关系,与心材和边材纤维素含量呈极显著负相关关系(p < 0.001)。倒木密度与倒木心材和边材的C、N、P以及木质素含量呈显著或极显著负相关关系, 与纤维素含量则表现为极显著正相关关系(p < 0.001)。这充分说明倒木物理性质与化学成分有密切联系。
3 讨论
本研究发现, 腐烂等级对倒木含水率和密度产生极显著影响(p < 0.001)(表1 )。随着腐烂等级增加, 倒木含水率升高, 密度下降(图1A 、1B)。这与其他地区对倒木含水率和密度的研究结果一致(吕明和等, 2006 ; Seedre et al ., 2013 ; Petrillo et al ., 2015 )。其原因主要是由于随着分解程度加深, 倒木受到淋溶、破碎化以及呼吸作用的影响, 木质变得疏松, 从而导致倒木密度下降; 随着腐烂等级增加, 其木质越疏松, 越容易吸收降水(袁杰和张硕新, 2012 ); 其次, 腐烂程度越高, 倒木与地面接触面积越大, 影响倒木含水率变化的因素增加, 例如土壤的含水率、地表水和微地貌等(Bütler et al ., 2007 )。本研究显示, 径级i-iii倒木密度呈下降趋势, 径级iv略有上升, 但与径级iii差异不显著(图1D ), 这与Sakai等(2012)的研究结果基本一致。倒木密度受许多因素的影响, 例如树种、地理环境、树龄、增长速率以及遗传因素等(Harmon et al ., 1986 ; Repola, 2006 )。Spurr和Hsuing (1954)发现, 从心材到边材, 幼龄树的密度比老龄树的大。本实验所测红松倒木的直径位置为胸径处, 可以间接代表龄级。相比大径级倒木, 小径级的倒木树龄小, 密度相对较大, 因此树龄是造成随着径级增加密度减小的主要原因。
C是构成植物体干物质的最主要元素, N和P是生态系统最主要的营养元素。N和P养分的有效性是调节植物残体分解速率和生态系统碳平衡的一个主要因素, 同时, 植物残体中N、P的矿化为土壤提供了重要的无机营养元素。本研究发现, 除心材C含量外, 腐烂等级显著地影响倒木C、N、P的含量(p < 0.05)(表1 )。随着腐烂等级增加, 边材C含量呈上升趋势, 且腐烂等级V显著高于腐烂等级I-III的C含量(图2 ), 这与Sandström等(2007)和Harmon等(2013)对裸子植物倒木C含量的研究结果一致, 而与Lombardi等(2013)和常晨晖等(2015)对倒木C含量的研究结果相反; Bütler等(2007)则发现欧洲云杉倒木随着分解程度加深C含量保持稳定。这些研究表明腐烂等级对倒木C含量的影响因环境条件和树种不同而存在差异。杨方方等(2009)认为, 倒木C含量随腐烂等级增加而增加的原因可能是木质残体分解周期较长, C损失速度与木质体腐解速度不同步, 从而在高腐烂等级C含量升高, 但导致C含量升高的深层原因有待进一步研究。由于分解初期倒木心材和边材N含量比较低, 随着腐烂等级增加, 边材与环境接触面积增大, 水热条件及通气量得到改善, 促进了微生物对边材的入侵及繁殖。同时, 寄生性微生物通过无脊椎动物侵入倒木心材, 心材结构疏松, 有利于微生物的繁殖。随着微生物增多, 一些固氮细菌数量不断增多, 对环境中N固定能力增强, N含量升高。到分解后期, 微生物竞争加剧, 呼吸作用产生的CO2 对微生物的毒害抑制了微生物的繁殖, 使得固氮细菌数量减少(常晨晖等, 2014 ), 固N能力减弱, N含量下降, 从而导致随着腐烂等级增加倒木N含量呈先升高后降低的变化趋势。虽然在分解后期略有下降, 但差异不显著, 这与其他相关研究结果基本一致(Palviainen et al ., 2008 ; 张修玉等, 2009 ; Köster et al ., 2015 )。本研究中, 倒木腐烂等级I和II的P含量相对较少, 腐烂等级III开始逐渐增加, 与杨丽韫和代力民(2002)、Palviainen等(2010)和Lombardi等(2013)的研究结果一致。这一方面可能是由于一些木材腐朽菌的菌丝把营养从土壤转移到倒木中(Clinton et al ., 2009 ), 另一方面由于有机物质的分解导致P的相对富集(Manzoni et al ., 2010 )。本研究结果发现边材的总体平均N含量显著高于心材(图2 ), 与Cowling和Merrill (1966)对裸子植物的研究结果基本一致。纤维素和木质素是倒木基质的主要组成成分, 其降解是维持生态系统碳素平衡的不可缺少的过程(Fioretto et al ., 2005 )。本研究中腐烂等级显著地影响倒木纤维素和木质素的含量(p < 0.001)(表1 ), 随着腐烂等级增加, 纤维素含量呈下降趋势, 木质素含量表现为上升趋势(图2 ), 这与以往对其他地区针叶树的研究结果相似(Ganjegunte et al ., 2004 ; Bütler et al ., 2007 ; 常晨晖等, 2015 )。这是因为纤维素是由纤维二糖单体组成的碳水化合物, 其结构相对简单, 而且有相对较多种类的微生物能够分解, 因此更容易被接触并分解(Austin & Ballaré, 2010 ); 而木质素具有独特的复杂结构, 它被认为是倒木中最难降解的组分。在分解初期随着易分解组分(可溶性C、N、P等)的不断降解, 木质素等难分解组分所占比例有所提高, 随着分解程度加深, 木质素含量升高。另外, 有关研究发现, 在分解过程中, 微生物新陈代谢能形成木质素类似物(Brandt et al ., 2010 ), 从而使倒木中木质素含量显著增加。
心材N、P含量在不同径级间差异显著(p < 0.05), 心材和边材的纤维素和木质素含量受腐烂等级和径级交互作用影响显著(p < 0.05)(表1 )。随着径级增加, 倒木心材和边材N、P含量呈先增加后减少的趋势, 径级iii倒木N、P含量最高, 且径级iii的心材P含量显著高于其他径级(p < 0.05)(图3 ), 这表明N、P含量在不同径级间存在差异, 且中等径级的倒木能够为生态系统提供更重要的养分来源。造成差异的原因主要是不同径级下倒木自身特性、物理结构以及分解者组成的影响。Stokland和Kauserud (2004)曾提出小倒木可能没有充足的养分供应分解者生长。小径级的倒木, 由于生长期相对比较短, 其本身所含养分量较大径级倒木少。其次, Harmon等(1986)发现随着径级增大, 倒木比表面积减小。这使得淋溶作用对P含量损失影响减少, 从而导致P含量不断积累。本研究中腐烂等级i至iii的倒木P含量呈上升趋势, 在腐烂等级iv倒木P含量下降, 这表明可能存在一个P含量积累的比表面积临界值, 超过临界值, 则淋溶损失速度大于积累速度导致P含量下降。分解者是倒木分解的主要影响因素, 它包括真菌、细菌、无脊椎动物等(Harmon et al ., 1986 )。径级的大小影响分解者的种类和数量(Küffer & Senn-Irlet, 2005 ; Juutilainen et al ., 2011 )。Brin等(2011)对不同径级倒木中寄生于枯木的无脊椎动物甲虫的研究发现, 大径级倒木中有更多的甲虫物种, 且不同径级倒木中的甲虫物种组成不同。Nordén等(2004)在瑞典南部阔叶林进行粗木质残体(CWD)和细木质残体(FWD)对真菌重要性的研究中发现FWD有更多子囊菌种类, CWD倒木上出现更多担子菌类, 而分解者对化学成分的不同利用影响不同径级倒木养分含量的分布。
生态化学计量学理论认为大多数情况下C:N > 27或C:P > 186时, 微生物生长受限制(Sinsabaugh et al ., 2009 )。本研究中, 心材和边材在整个分解阶段均易受到N、P限制。Laiho和Prescott (2004)认为初始N:P值也能很好地预测N、P动态, 他们在针叶树种分解过程中养分释放的研究中发现临界N:P值约为20。本研究中, 腐烂等级I-III心材和边材N:P值均大于20 (表2 ), 表现为N相对含量的富集模式, 随着腐烂等级增加, 腐烂等级IV和V边材和心材N:P值均小于20, 表现出养分含量的释放模式。通常C在大多数植物体内含量高且变化较小(Reich & Oleksyn, 2004 ), 本研究也证实了这一点, 不同径级间C含量差异不显著(表3 )。影响C与N、P比值的主要因素是N、P含量的变化(Reich & Oleksyn, 2004 )。N、P含量随着径级先增后减, 而C含量在不同径级中比较稳定, 导致了随着径级增加, C:N、C:P先降后升的趋势(表3 )。
倒木的含水率、密度与纤维素含量和木质素含量具有显著的相关性, 这可能与倒木的呼吸作用有关。含水率和密度是影响倒木呼吸的主要物理因素, 倒木的呼吸作用主要是通过微生物的分解作用来进行的(Herrmann & Bauhus, 2013 )。微生物分解对水分的增加非常敏感, 在一定范围内, 较小的水分增加会导致微生物活性的急剧增大(Progar et al ., 2000 )。由于单糖等非结构性碳水化合物易被微生物分解且含量较低, 到分解中后期, 相比木质素, 纤维素由于其结构相对简单, 成为微生物分解的主要物质。随着倒木含水率的增加, 微生物对纤维素的分解加快, 纤维素含量下降, 而木质素在倒木中的含量相对升高, 从而导致含水率与纤维素含量的负相关, 与木质素含量正相关(表4 )。本文中木材密度对倒木纤维素和木质素的影响与含水率相反。密度低, 则微生物易于侵入进行分解活动, 且其持水能力强, 更易促进微生物对纤维素的分解(Chamber et al ., 2001 )。因此密度与木质素、纤维素的关系与含水率相反(表4 )。而倒木中C、N、P含量的变化与倒木含水率和密度之间的相关性实质上是腐烂等级分别对倒木物理化学性质的影响所导致的。C、N、P与含水率均表现为随着腐烂等级增加而升高,密度则下降。因此, 含水率与C、N、P含量呈正相关关系; 密度则与倒木C、N、P含量呈负相关关系。
综上所述, 红松的倒木除心材C含量外, 倒木物理化学性质在腐烂等级间差异显著(p < 0.05), 径级对倒木心材N、P含量以及心材和边材密度影响显著(p < 0.05), 且倒木心材和边材径级iii的N、P含量最高; 纤维素和木质素的含量受腐烂等级和径级的交互作用影响显著。以上结果表明倒木心材和边材的物理化学性质在不同腐烂等级和不同径级中质量特征存在差异。对不同腐烂等级和径级的倒木进行物理化学研究, 能够为更准确地预测倒木C储量和养分含量以及生产力变化提供科学依据。受篇幅的影响, 本文只探讨了腐烂等级和径级对阔叶红松林的建群种红松倒木理化性质的影响, 在今后的研究中, 需要进一步研究对其他主要组成树种以及功能群的影响规律。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Austin AT Ballaré CL 2010 ). Dual role of lignin in plant litter decomposition in terrestrial ecosystems
.Proceedings of the National Academy of Sciences of the United States of America 107 , 4618 -4622 .
[本文引用: 1]
[2]
Berg B Johansson MB Meentemeyer V 2000 ). Litter decomposition in a transect of Norway spruce forests: Substrate quality and climate control
.Canadian Journal of Forest Research 30 , 1136 -1147 .
[本文引用: 1]
[3]
Bond-Lamberty B Wang CK Gower ST 2002 ). Annual carbon flux from woody debris for a boreal black spruce fire chronosequence
.Journal of Geophysical Research 108 , 121 -129 .
[本文引用: 1]
[4]
Brandt LA King JY Hobbie SE Milchunas DG Sinsabaugh RL 2010 ). The role of photo degradation in surface litter decomposition across a grassland ecosystem precipitation gradient
.Ecosystems 13 , 765 -781 .
[本文引用: 1]
[5]
Brin A Bouget C Brustel H Jactel H 2011 ). Diameter of downed woody debris does matter for saproxylic beetle assemblages in temperate oak and pine forests
.Journal of Insect Conservation 15 , 653 -669 .
[6]
Bütler R Patty L Bayon RCL Guenat C Schlaepfer R 2007 ). Log decay of
Picea abies in the Swiss Jura Mountains of central Europe. Forest Ecology and Management 242 , 791 -799 .
[本文引用: 2]
[7]
Chamber JQ Schime JP Nobre AD 2001 ). Respiration from coarse wood litter in centra Amazon forests
.Biogeochemistry 52 , 115 -131 .
[本文引用: 2]
[8]
Chang CH Wu FZ Yang WQ Tan B Li H Xiao S Gou XL He LN 2014 ). The dynamics of microbial community structure at different stages of log decay in an alpine forest of western Sichuan
.Chinese Journal Applied and Environmental Biology 20 , 978 -985 .(in Chinese with English abstract) [常晨晖 , 吴福忠 , 杨万勤 , 谭波 , 李晗 , 肖洒 , 苟小林 , 何丽娜 (2014 ). 川西高山森林倒木不同分解阶段的微生物群落变化特征. 应用与环境生物学报,
20 , 978 -985 .]
[本文引用: 1]
[9]
Chang CH Wu FZ Yang WQ Tan B Xiao S Li J Gou XL 2015 ). Changes in log quality at different decay stages in an alpine forest
.Chinese Journal of Plant Ecology 39 , 14 -22 .(in Chinese with English abstract) [常晨晖 , 吴福忠 , 杨万勤 , 谭波 , 肖洒 , 李俊 , 苟小林 (2015 ). 高寒森林倒木在不同分解阶段的质量变化. 植物生态学报,
39 , 14 -22 .]
[本文引用: 4]
[10]
Clinton PW Buchanan PK Wilkie JP Smaill SJ Kimberley MO 2009 ). Decomposition of
Nothofagus wood in vitro and nutrient mobilization by fungi. Canadian Journal of Forest Research 39 , 2193 -2202 .
[本文引用: 1]
[11]
Cowling EB Merrill W 1966 ). Nitrogen in wood and its role in wood deterioration
.Canadian Journal of Botany 44 , 1539 -1554 .
[12]
Dai LM Xu ZB Chen HS 2000 ). Storage dynamics of fallen trees in the broad-leaved and Korean pine mixed forest
. Acta Ecologica Sinica 20 , 412 -416 .(in Chinese with English abstract) [代力民 , 徐振邦 , 陈华 (2000 ). 阔叶红松林倒木贮量的变化规律. 生态学报,
20 , 412 -416 .]
[本文引用: 1]
[13]
de Aza CH Turrión MB Pando V Bravo F 2011 ). Carbon in heartwood, sapwood and bark along the stem profile in three Mediterranean
Pinus species. Annals of Forest Science 68 , 1067 -1076 .
[14]
Fioretto A Di Nardo C Papa S Fuggi A 2005 ). Lignin and cellulose degradation and nitrogen dynamics during de- composition of three leaf litter species in a Mediterranean ecosystem
.Soil Biology & Biochemistry 37 , 1083 -1091 .
[本文引用: 1]
[15]
Ganjegunte GK Condron LM Clinton PW Davis MR Mahieu N 2004 ). Decomposition and nutrient release from radiata pine (
Pinus radiata ) coarse woody debris. Forest Ecology and Management 187 , 197 -211 .
[本文引用: 1]
[16]
Harmon ME Fasth B Woodall CW Sexton J 2013 ). Carbon concentration of standing and downed woody detritus: Effects of tree taxa, decay class, position, and tissue type
.Forest Ecology and Management 291 , 259 -267 .
[17]
Harmon ME Franklin JF Swanson FJ Sollins P Gregory SV Lattin JD Anderson NH Cline SP Aumen NG Sedell JR Lienkaemper GW Cromack KJ Cummins KW 1986 ). Ecology of coarse woody debris in temperate ecosystems
.Advances in Ecological Research 15 , 133 -302 .
[本文引用: 3]
[18]
He W Wu F Yang W Tan B Zhao Y Wu Q He M 2016 ). Lignin degradation in foliar litter of two shrub species from the gap center to the closed canopy in an alpine fir forest
.Ecosystems 19 , 115 -128 .
[本文引用: 1]
[19]
Herrmann S Bauhus J 2013 ). Effects of moisture, temperature and decomposition stage on respirational carbon loss from coarse woody debris (CWD) of important European tree species
.Scandinavian Journal of Forest Research 28 , 346 -357 .
[本文引用: 1]
[20]
Jin GZ Xie XC Tian YY Kim JH 2006 ). The pattern of seed rain in the broadleaved-Korean pine mixed forest of Xiaoxing’an Mountains, China
.Journal of Korean Forest Society 95 , 621 -627 .
[本文引用: 1]
[21]
Johnston JM Crossley Jr DA 1996 ). The significance of coarse woody debris for the diversity of soil mites
. Forest Service 94 , 82 -87 .
[本文引用: 1]
[22]
Juutilainen K Halme P Kotiranta H Mönkkönen M 2011 ). Diameter matters in studies of dead wood and wood-inhabiting fungi
.Fungal Ecology 4 , 342 -349 .
[本文引用: 1]
[23]
Köster K Metslaid M Engelhart J Köster E 2015 ). Dead wood basic density, and the concentration of carbon and nitrogen for main tree species in managed hemiboreal forests
.Forest Ecology and Management 354 , 35 -42 .
[本文引用: 2]
[24]
Küffer N Senn-Irlet B 2005 ). Influence of forest management on the species richness and composition of wood- inhabiting basidiomycetes in Swiss forests
.Biodiversity and Conservation 14 , 2419 -2435 .
[本文引用: 1]
[25]
Laiho R Prescott CE 2004 ). Decay and nutrient dynamics of coarse woody debris in northern coniferous forests: A synthesis
.Canadian Journal of Forest Research 34 , 763 -777 .
[本文引用: 1]
[26]
Liu YY Jin GZ 2010 a). Character of coarse woody debris in a mixed broadleaved-Korean pine forest in Xiaoxing’an Mountains, China
. Scientia Silvae Sinicae 46 (4 ), 8 -14 .(in Chinese with English abstract) [刘妍妍 , 金光泽 (2010 a). 小兴安岭阔叶红松林粗木质残体基础特征. 林业科学,
46(4) , 8 -14 .]
[27]
Liu YY Jin GZ 2010 b). Spatial point pattern analysis for coarse woody debris in a mixed broad-leaved Korean pine forest in Xiaoxing’an Mountains, China
.Acta Ecologica Sinica 22 , 6072 -6081 .(in Chinese with English abstract) [刘妍妍 , 金光泽 (2010 b). 小兴安岭阔叶红松林粗木质残体空间分布的点格局分析. 生态学报,
22 , 6072 -6081 .]
[28]
Lombardi F Cherubini P Tognetti R Cocozza C Lasserre B Marchetti M 2013 ). Investigating biochemical processes to assess deadwood decay of beech and silver fir in Mediterranean Mountain forests
.Annals of Forest Science 70 , 101 -111 .
[29]
Lü MH Zhou GY Zhang DQ Guan LL 2006 ). Decomposi- tion and nutrient release from coarse woody debris of
Castanopsis chinensis in Dinghushan forest ecosystem. Journal of Tropical and Subtropical Botany 14 , 107 -112 .(in Chinese with English abstract) [吕明和 , 周国逸 , 张德强 , 官丽莉 (2006 ). 鼎湖山锥栗粗木质残体的分解和元素动态. 热带亚热带植物学报,
14 , 107 -112 .]
[本文引用: 2]
[30]
Manzoni S Trofymow JA Jackson RB Porporato A 2010 ). Stoichiometric controls on carbon, nitrogen, and phosph- orus dynamics in decomposing litter
.Ecological Monographs 80 , 89 -106 .
[本文引用: 1]
[31]
Moroni MT Ryan DAJ 2010 ). Deadwood abundance in recently harvested and old Nova Scotia hardwood forests
.Forestry 83 , 219 -227 .
[本文引用: 1]
[32]
Nordén B Ryberg M Götmark F Olausson B 2004 ). Relative importance of coarse and fine woody debris for the diversity of wood-inhabiting fungi in temperate broadleaf forests
.Biological Conservation 117 , 1 -10 .
[33]
Palviainen M Finér L Laiho R Shorohova E Kapitsa E Vanha-Majamaa I 2010 ). Phosphorus and base cation accumulation and release patterns in decomposing Scots pine, Norway spruce and silver birch stumps
.Forest Ecology and Management 260 , 1478 -1489 .
[34]
Palviainen M Laiho R Mäkinen H Finer L 2008 ). Do decomposing Scots pine, Norway spruce, and silver birch stems retain nitrogen?
Canadian Journal of Forest Research 38 , 3047 -3055 .
[本文引用: 2]
[35]
Petrillo M Cherubini P Sartori G Abiven S Ascher J Bertoldi D Egli M 2015 ). Decomposition of Norway spruce and European larch coarse woody debris (CWD) in relation to different elevation and exposure in an Alpine setting
.iForest-Biogeosciences and Forestry 9 , 154 -164 .
[本文引用: 1]
[36]
Progar RA Schowalter TD Freitag CM Morrell JJ 2000 ). Respiration from coarse woody debris as affected by moisture and saprotroph functional diversity in Western Oregon
.Oecologia 124 , 426 -431 .
[本文引用: 1]
[37]
Reich PB Oleksyn J 2004 ). Global patterns of plant leaf N and P in relation to temperature and latitude
.Proceedings of the National Academy of Sciences of the United States of America 101 , 11001 -11006 .
[本文引用: 2]
[38]
Repola J 2006 ). Models for vertical wood density of Scots pine, Norway spruce and birch stems, and their application to determine average wood density
.Silva Fennica 40 , 673 -685 .
[本文引用: 1]
[39]
Sakai Y Ugawa S Ishizuka S Takahashi M Takenaka C 2012 ). Wood density and carbon and nitrogen concentrations in deadwood of
Chamaecyparis obtusa and Cryptomeria japonica . Soil Science and Plant Nutrition 58 , 526 -537 .
[40]
Sandström F Petersson H Kruys N Ståhl G 2007 ). Biomass conversion factors (density and carbon concentration) by decay classes for dead wood of
Pinus sylvestris, Picea abies and Betula spp. in boreal forests of Sweden. Forest Ecology and Management 243 , 19 -27 .
[41]
Schwarze FW 2007 ). Wood decay under the microscope
.Fungal Biology Reviews 21 , 133 -170 .
[本文引用: 1]
[42]
Seedre M Taylor A Chen H Jõgiste K 2013 ). Deadwood density of five boreal tree species in relation to field-assigned decay class
.Forest Science 59 , 261 -266 .
[本文引用: 1]
[43]
Sefidi K Etemad V 2015 ). Dead wood characteristics influ- enceing macrofungi species abundance and diversity in Caspian natural beech (Fagus orientalis Lipsky) forests
. Forest Systems, 24, eSC03.
[本文引用: 1]
[44]
Sinsabaugh RL Hill BH Follstad Shah JJ 2009 ). Ecoenzy- matic stoichiometry of microbial organic nutrient acquisition in soil and sediment
. Nature 462 , 795 -799 .
[本文引用: 1]
[45]
Sollins P 1982 ). Input and decay of coarse woody debris in coniferous stands in western Oregon and Washington
.Canadian Journal of Forest Research 12 , 18 -28 .
[本文引用: 1]
[46]
Spears JD Holub SM Harmon ME Lajtha K 2003 ). The influence of decomposing logs on soil biology and nutrient cycling in an old-growth mixed coniferous forest in Oregon, USA
. Canadian Journal of Forest Research 33 , 2193 -2201 .
[本文引用: 1]
[47]
Spurr SH Hsiung WY 1954 ). Growth rate and specific gravity in conifers
.Journal of Forestry 52 , 191 -200 .
[48]
Stokland J Kauserud H 2004 ). Phellinus nigrolimitatus —A wood-decomposing fungus highly influenced by forestry
.Forest Ecology and Management 187 , 333 -343 .
[49]
van Geffen KG Poorter L Sass-Klaassen U van Logtestijn RS Cornelissen JH 2010 ). The trait contribution to wood decomposition rates of 15 neotropical tree species
.Ecology 91 , 3686 -3697 .
[本文引用: 1]
[50]
Wassen MJ Venterink HO Lapshina ED Tanneberger F 2005 ). Endangered plants persist under phosphorus limitation
.Nature 437 , 547 -550 .
[本文引用: 1]
[51]
Xin WW Tie N Li ZF Zhang DM Zhang ZZ 2012 ). Da- xing’anling
Larix regeneration of fallen tree nutrient con- tent research. Forest Resources Management 3 ), 94 -99 .(in Chinese with English abstract) [辛魏巍 , 铁牛 , 李卓凡 , 张冬梅 , 张最最 (2012 ). 大兴安岭兴安落叶松原始林倒木养分含量研究
. 林业资源管理, (3), 94-99].
[本文引用: 1]
[52]
Yan ER Wang XH Huang JJ 2005 ). Concept and classifica- tion of coarse woody debris in forest ecosystems
.Acta Ecologica Sinica 25 , 158 -167 .(in Chinese with English abstract) [闫恩荣 , 王希华 , 黄建军 (2005 ). 森林粗死木质残体的概念及其分类. 生态学报,
25 , 158 -167 .]
[本文引用: 1]
[53]
Yang FF Li YL Liu XZ 2009 ). Decomposition of coarse woody debris (CWD) of dominate species
Schima superba in monsoonal evergreen broadleaved forest of Dinghushan Nature Reserve. Journal of Mountains Science 27 , 442 -448 .(in Chinese with English abstract) [杨方方 , 李跃林 , 刘兴诏 (2009 ). 鼎湖山木荷粗死木质残体的分解研究. 山地学报,
27 , 442 -448 .]
[54]
Yang LY Dai LM 2002 ). The decomposition and nutrient content of fallen woods in the moss-
Pinus koraiensis dark-conifer forest at north slope of Changbai Mountain. Acta Ecologica Sinica 22 , 185 -189 .(in Chinese with English abstract) [杨丽韫 , 代力民 (2002 ). 长白山北坡苔藓红松暗针叶林倒木分解及其养分含量. 生态学报,
22 , 185 -189 .]
[本文引用: 1]
[55]
Yuan J Zhang SX 2012 ). Characteristics of fallen wood den- sity and water content dynamics of
Pinus tabulaeformis natural secondary forests in Huoditang region in Qinling Mountains. Journal of Central South University of Forestry & Technology 32 (11 ), 105 -109 .(in Chinese with English abstract) [袁杰 , 张硕新 (2012 ). 秦岭火地塘天然次生油松林倒木密度与含水量变化特征研究. 中南林业科技大学学报,
32(11) , 105 -109 .]
[本文引用: 1]
[56]
Zhang XY Guan DS Zhang HD 2009 ). Characteristics of storage and decomposition of coarse woody debris under three forests in Guangzhou
.Acta Ecologica Sinica 29 , 5227 -5236 .(in Chinese with English abstract) [张修玉 , 管东生 , 张海东 (2009 ). 三种森林粗死木质残体 (CWD)的储量与分解特征. 生态学报,
29 , 5227 -5236 .]
[本文引用: 1]
[57]
Zhou L Dai LM Gu HY Zhong L 2007 ). Review on the decomposition and influence factors of coarse woody debris in forest ecosystem
. Journal of Forestry Research 18 , 48 -54 .
[本文引用: 1]
Dual role of lignin in plant litter decomposition in terrestrial ecosystems
1
2010
... C是构成植物体干物质的最主要元素, N和P是生态系统最主要的营养元素.N和P养分的有效性是调节植物残体分解速率和生态系统碳平衡的一个主要因素, 同时, 植物残体中N、P的矿化为土壤提供了重要的无机营养元素.本研究发现, 除心材C含量外, 腐烂等级显著地影响倒木C、N、P的含量(p < 0.05)(表1 ).随着腐烂等级增加, 边材C含量呈上升趋势, 且腐烂等级V显著高于腐烂等级I-III的C含量(图2 ), 这与Sandström等(2007)和Harmon等(2013)对裸子植物倒木C含量的研究结果一致, 而与Lombardi等(2013)和常晨晖等(2015)对倒木C含量的研究结果相反; Bütler等(2007)则发现欧洲云杉倒木随着分解程度加深C含量保持稳定.这些研究表明腐烂等级对倒木C含量的影响因环境条件和树种不同而存在差异.杨方方等(2009)认为, 倒木C含量随腐烂等级增加而增加的原因可能是木质残体分解周期较长, C损失速度与木质体腐解速度不同步, 从而在高腐烂等级C含量升高, 但导致C含量升高的深层原因有待进一步研究.由于分解初期倒木心材和边材N含量比较低, 随着腐烂等级增加, 边材与环境接触面积增大, 水热条件及通气量得到改善, 促进了微生物对边材的入侵及繁殖.同时, 寄生性微生物通过无脊椎动物侵入倒木心材, 心材结构疏松, 有利于微生物的繁殖.随着微生物增多, 一些固氮细菌数量不断增多, 对环境中N固定能力增强, N含量升高.到分解后期, 微生物竞争加剧, 呼吸作用产生的CO2 对微生物的毒害抑制了微生物的繁殖, 使得固氮细菌数量减少(常晨晖等, 2014 ), 固N能力减弱, N含量下降, 从而导致随着腐烂等级增加倒木N含量呈先升高后降低的变化趋势.虽然在分解后期略有下降, 但差异不显著, 这与其他相关研究结果基本一致(Palviainen et al ., 2008 ; 张修玉等, 2009 ; Köster et al ., 2015 ).本研究中, 倒木腐烂等级I和II的P含量相对较少, 腐烂等级III开始逐渐增加, 与杨丽韫和代力民(2002)、Palviainen等(2010)和Lombardi等(2013)的研究结果一致.这一方面可能是由于一些木材腐朽菌的菌丝把营养从土壤转移到倒木中(Clinton et al ., 2009 ), 另一方面由于有机物质的分解导致P的相对富集(Manzoni et al ., 2010 ).本研究结果发现边材的总体平均N含量显著高于心材(图2 ), 与Cowling和Merrill (1966)对裸子植物的研究结果基本一致.纤维素和木质素是倒木基质的主要组成成分, 其降解是维持生态系统碳素平衡的不可缺少的过程(Fioretto et al ., 2005 ).本研究中腐烂等级显著地影响倒木纤维素和木质素的含量(p < 0.001)(表1 ), 随着腐烂等级增加, 纤维素含量呈下降趋势, 木质素含量表现为上升趋势(图2 ), 这与以往对其他地区针叶树的研究结果相似(Ganjegunte et al ., 2004 ; Bütler et al ., 2007 ; 常晨晖等, 2015 ).这是因为纤维素是由纤维二糖单体组成的碳水化合物, 其结构相对简单, 而且有相对较多种类的微生物能够分解, 因此更容易被接触并分解(Austin & Ballaré, 2010 ); 而木质素具有独特的复杂结构, 它被认为是倒木中最难降解的组分.在分解初期随着易分解组分(可溶性C、N、P等)的不断降解, 木质素等难分解组分所占比例有所提高, 随着分解程度加深, 木质素含量升高.另外, 有关研究发现, 在分解过程中, 微生物新陈代谢能形成木质素类似物(Brandt et al ., 2010 ), 从而使倒木中木质素含量显著增加. ...
Litter decomposition in a transect of Norway spruce forests: Substrate quality and climate control
1
2000
... 森林是陆地生态系统的重要组成部分, 其植被碳(C)库约占全球植被C库的80% (Köster et al ., 2015 ).倒木是森林生态系统的重要组成部分, 对于维持生物多样性以及林木的更新具有重要作用; 同时, 倒木也是重要的养分库和C库, 其养分的缓慢释放, 对于提高林地的生产力有重要意义(Harmon et al ., 1986 ; Laiho & Prescott, 2004 ).倒木在物理、化学和生物(特别是微生物、昆虫)等作用下进行的分解过程不仅可为分解者(如微生物、无脊椎动物、鸟类等)提供生存环境和必需的营养元素(Sollins, 1982 ), 而且能够释放大量的化学元素到土壤中, 丰富土壤有机质含量和种类, 促进土壤结构和功能的发育(Spears et al ., 2003 ).影响倒木分解的因素有基质质量、分解者活性以及环境因素等(如温度、水分、气体)(Harmon et al ., 1986).而在同一环境下, 不同树种间倒木分解速率也有很大差异, 这与倒木自身的物理和化学特性有关(van Geffen et al ., 2010 ).含水率和木材密度是影响倒木分解的主要物理因素, 因而受到越来越多的关注.倒木含水率是影响倒木呼吸的重要因子, 当它超过一定的阈值后, 就会影响微生物的活动, 从而抑制倒木的分解(Bond-Lamberty et al ., 2002 ).木材分解是一个复杂的过程, 通常根据单位时间内木材质量损失或木材密度的变化, 可以定量地算出其分解速率; 木材密度与微生物侵入倒木进行分解活动的程度有密切关系, 较低密度的木质残体更容易被分解和具有更高的呼吸速率(Johnston & Crossley, 1996 ; Chamber et al ., 2001 ).影响倒木分解的化学因素主要是其易分解成分和难分解有机成分(如木质素等)的含量, 其中氮(N)、磷(P)、木质素、纤维素、C:N、C:P等是常见的衡量倒木质量的指标(Berg et al ., 2000 ), 它们不仅可以表征倒木分解过程中基质的质量变化和养分限制状况(Wassen et al ., 2005 ), 而且可以指示物质循环和能量流动的方向(Zhou et al ., 2007 ). ...
Annual carbon flux from woody debris for a boreal black spruce fire chronosequence
1
2002
... 森林是陆地生态系统的重要组成部分, 其植被碳(C)库约占全球植被C库的80% (Köster et al ., 2015 ).倒木是森林生态系统的重要组成部分, 对于维持生物多样性以及林木的更新具有重要作用; 同时, 倒木也是重要的养分库和C库, 其养分的缓慢释放, 对于提高林地的生产力有重要意义(Harmon et al ., 1986 ; Laiho & Prescott, 2004 ).倒木在物理、化学和生物(特别是微生物、昆虫)等作用下进行的分解过程不仅可为分解者(如微生物、无脊椎动物、鸟类等)提供生存环境和必需的营养元素(Sollins, 1982 ), 而且能够释放大量的化学元素到土壤中, 丰富土壤有机质含量和种类, 促进土壤结构和功能的发育(Spears et al ., 2003 ).影响倒木分解的因素有基质质量、分解者活性以及环境因素等(如温度、水分、气体)(Harmon et al ., 1986).而在同一环境下, 不同树种间倒木分解速率也有很大差异, 这与倒木自身的物理和化学特性有关(van Geffen et al ., 2010 ).含水率和木材密度是影响倒木分解的主要物理因素, 因而受到越来越多的关注.倒木含水率是影响倒木呼吸的重要因子, 当它超过一定的阈值后, 就会影响微生物的活动, 从而抑制倒木的分解(Bond-Lamberty et al ., 2002 ).木材分解是一个复杂的过程, 通常根据单位时间内木材质量损失或木材密度的变化, 可以定量地算出其分解速率; 木材密度与微生物侵入倒木进行分解活动的程度有密切关系, 较低密度的木质残体更容易被分解和具有更高的呼吸速率(Johnston & Crossley, 1996 ; Chamber et al ., 2001 ).影响倒木分解的化学因素主要是其易分解成分和难分解有机成分(如木质素等)的含量, 其中氮(N)、磷(P)、木质素、纤维素、C:N、C:P等是常见的衡量倒木质量的指标(Berg et al ., 2000 ), 它们不仅可以表征倒木分解过程中基质的质量变化和养分限制状况(Wassen et al ., 2005 ), 而且可以指示物质循环和能量流动的方向(Zhou et al ., 2007 ). ...
The role of photo degradation in surface litter decomposition across a grassland ecosystem precipitation gradient
1
2010
... C是构成植物体干物质的最主要元素, N和P是生态系统最主要的营养元素.N和P养分的有效性是调节植物残体分解速率和生态系统碳平衡的一个主要因素, 同时, 植物残体中N、P的矿化为土壤提供了重要的无机营养元素.本研究发现, 除心材C含量外, 腐烂等级显著地影响倒木C、N、P的含量(p < 0.05)(表1 ).随着腐烂等级增加, 边材C含量呈上升趋势, 且腐烂等级V显著高于腐烂等级I-III的C含量(图2 ), 这与Sandström等(2007)和Harmon等(2013)对裸子植物倒木C含量的研究结果一致, 而与Lombardi等(2013)和常晨晖等(2015)对倒木C含量的研究结果相反; Bütler等(2007)则发现欧洲云杉倒木随着分解程度加深C含量保持稳定.这些研究表明腐烂等级对倒木C含量的影响因环境条件和树种不同而存在差异.杨方方等(2009)认为, 倒木C含量随腐烂等级增加而增加的原因可能是木质残体分解周期较长, C损失速度与木质体腐解速度不同步, 从而在高腐烂等级C含量升高, 但导致C含量升高的深层原因有待进一步研究.由于分解初期倒木心材和边材N含量比较低, 随着腐烂等级增加, 边材与环境接触面积增大, 水热条件及通气量得到改善, 促进了微生物对边材的入侵及繁殖.同时, 寄生性微生物通过无脊椎动物侵入倒木心材, 心材结构疏松, 有利于微生物的繁殖.随着微生物增多, 一些固氮细菌数量不断增多, 对环境中N固定能力增强, N含量升高.到分解后期, 微生物竞争加剧, 呼吸作用产生的CO2 对微生物的毒害抑制了微生物的繁殖, 使得固氮细菌数量减少(常晨晖等, 2014 ), 固N能力减弱, N含量下降, 从而导致随着腐烂等级增加倒木N含量呈先升高后降低的变化趋势.虽然在分解后期略有下降, 但差异不显著, 这与其他相关研究结果基本一致(Palviainen et al ., 2008 ; 张修玉等, 2009 ; Köster et al ., 2015 ).本研究中, 倒木腐烂等级I和II的P含量相对较少, 腐烂等级III开始逐渐增加, 与杨丽韫和代力民(2002)、Palviainen等(2010)和Lombardi等(2013)的研究结果一致.这一方面可能是由于一些木材腐朽菌的菌丝把营养从土壤转移到倒木中(Clinton et al ., 2009 ), 另一方面由于有机物质的分解导致P的相对富集(Manzoni et al ., 2010 ).本研究结果发现边材的总体平均N含量显著高于心材(图2 ), 与Cowling和Merrill (1966)对裸子植物的研究结果基本一致.纤维素和木质素是倒木基质的主要组成成分, 其降解是维持生态系统碳素平衡的不可缺少的过程(Fioretto et al ., 2005 ).本研究中腐烂等级显著地影响倒木纤维素和木质素的含量(p < 0.001)(表1 ), 随着腐烂等级增加, 纤维素含量呈下降趋势, 木质素含量表现为上升趋势(图2 ), 这与以往对其他地区针叶树的研究结果相似(Ganjegunte et al ., 2004 ; Bütler et al ., 2007 ; 常晨晖等, 2015 ).这是因为纤维素是由纤维二糖单体组成的碳水化合物, 其结构相对简单, 而且有相对较多种类的微生物能够分解, 因此更容易被接触并分解(Austin & Ballaré, 2010 ); 而木质素具有独特的复杂结构, 它被认为是倒木中最难降解的组分.在分解初期随着易分解组分(可溶性C、N、P等)的不断降解, 木质素等难分解组分所占比例有所提高, 随着分解程度加深, 木质素含量升高.另外, 有关研究发现, 在分解过程中, 微生物新陈代谢能形成木质素类似物(Brandt et al ., 2010 ), 从而使倒木中木质素含量显著增加. ...
Diameter of downed woody debris does matter for saproxylic beetle assemblages in temperate oak and pine forests
2011
Log decay of
2
2007
... 本研究发现, 腐烂等级对倒木含水率和密度产生极显著影响(p < 0.001)(表1 ).随着腐烂等级增加, 倒木含水率升高, 密度下降(图1A 、1B).这与其他地区对倒木含水率和密度的研究结果一致(吕明和等, 2006 ; Seedre et al ., 2013 ; Petrillo et al ., 2015 ).其原因主要是由于随着分解程度加深, 倒木受到淋溶、破碎化以及呼吸作用的影响, 木质变得疏松, 从而导致倒木密度下降; 随着腐烂等级增加, 其木质越疏松, 越容易吸收降水(袁杰和张硕新, 2012 ); 其次, 腐烂程度越高, 倒木与地面接触面积越大, 影响倒木含水率变化的因素增加, 例如土壤的含水率、地表水和微地貌等(Bütler et al ., 2007 ).本研究显示, 径级i-iii倒木密度呈下降趋势, 径级iv略有上升, 但与径级iii差异不显著(图1D ), 这与Sakai等(2012)的研究结果基本一致.倒木密度受许多因素的影响, 例如树种、地理环境、树龄、增长速率以及遗传因素等(Harmon et al ., 1986 ; Repola, 2006 ).Spurr和Hsuing (1954)发现, 从心材到边材, 幼龄树的密度比老龄树的大.本实验所测红松倒木的直径位置为胸径处, 可以间接代表龄级.相比大径级倒木, 小径级的倒木树龄小, 密度相对较大, 因此树龄是造成随着径级增加密度减小的主要原因. ...
... C是构成植物体干物质的最主要元素, N和P是生态系统最主要的营养元素.N和P养分的有效性是调节植物残体分解速率和生态系统碳平衡的一个主要因素, 同时, 植物残体中N、P的矿化为土壤提供了重要的无机营养元素.本研究发现, 除心材C含量外, 腐烂等级显著地影响倒木C、N、P的含量(p < 0.05)(表1 ).随着腐烂等级增加, 边材C含量呈上升趋势, 且腐烂等级V显著高于腐烂等级I-III的C含量(图2 ), 这与Sandström等(2007)和Harmon等(2013)对裸子植物倒木C含量的研究结果一致, 而与Lombardi等(2013)和常晨晖等(2015)对倒木C含量的研究结果相反; Bütler等(2007)则发现欧洲云杉倒木随着分解程度加深C含量保持稳定.这些研究表明腐烂等级对倒木C含量的影响因环境条件和树种不同而存在差异.杨方方等(2009)认为, 倒木C含量随腐烂等级增加而增加的原因可能是木质残体分解周期较长, C损失速度与木质体腐解速度不同步, 从而在高腐烂等级C含量升高, 但导致C含量升高的深层原因有待进一步研究.由于分解初期倒木心材和边材N含量比较低, 随着腐烂等级增加, 边材与环境接触面积增大, 水热条件及通气量得到改善, 促进了微生物对边材的入侵及繁殖.同时, 寄生性微生物通过无脊椎动物侵入倒木心材, 心材结构疏松, 有利于微生物的繁殖.随着微生物增多, 一些固氮细菌数量不断增多, 对环境中N固定能力增强, N含量升高.到分解后期, 微生物竞争加剧, 呼吸作用产生的CO2 对微生物的毒害抑制了微生物的繁殖, 使得固氮细菌数量减少(常晨晖等, 2014 ), 固N能力减弱, N含量下降, 从而导致随着腐烂等级增加倒木N含量呈先升高后降低的变化趋势.虽然在分解后期略有下降, 但差异不显著, 这与其他相关研究结果基本一致(Palviainen et al ., 2008 ; 张修玉等, 2009 ; Köster et al ., 2015 ).本研究中, 倒木腐烂等级I和II的P含量相对较少, 腐烂等级III开始逐渐增加, 与杨丽韫和代力民(2002)、Palviainen等(2010)和Lombardi等(2013)的研究结果一致.这一方面可能是由于一些木材腐朽菌的菌丝把营养从土壤转移到倒木中(Clinton et al ., 2009 ), 另一方面由于有机物质的分解导致P的相对富集(Manzoni et al ., 2010 ).本研究结果发现边材的总体平均N含量显著高于心材(图2 ), 与Cowling和Merrill (1966)对裸子植物的研究结果基本一致.纤维素和木质素是倒木基质的主要组成成分, 其降解是维持生态系统碳素平衡的不可缺少的过程(Fioretto et al ., 2005 ).本研究中腐烂等级显著地影响倒木纤维素和木质素的含量(p < 0.001)(表1 ), 随着腐烂等级增加, 纤维素含量呈下降趋势, 木质素含量表现为上升趋势(图2 ), 这与以往对其他地区针叶树的研究结果相似(Ganjegunte et al ., 2004 ; Bütler et al ., 2007 ; 常晨晖等, 2015 ).这是因为纤维素是由纤维二糖单体组成的碳水化合物, 其结构相对简单, 而且有相对较多种类的微生物能够分解, 因此更容易被接触并分解(Austin & Ballaré, 2010 ); 而木质素具有独特的复杂结构, 它被认为是倒木中最难降解的组分.在分解初期随着易分解组分(可溶性C、N、P等)的不断降解, 木质素等难分解组分所占比例有所提高, 随着分解程度加深, 木质素含量升高.另外, 有关研究发现, 在分解过程中, 微生物新陈代谢能形成木质素类似物(Brandt et al ., 2010 ), 从而使倒木中木质素含量显著增加. ...
Respiration from coarse wood litter in centra Amazon forests
2
2001
... 森林是陆地生态系统的重要组成部分, 其植被碳(C)库约占全球植被C库的80% (Köster et al ., 2015 ).倒木是森林生态系统的重要组成部分, 对于维持生物多样性以及林木的更新具有重要作用; 同时, 倒木也是重要的养分库和C库, 其养分的缓慢释放, 对于提高林地的生产力有重要意义(Harmon et al ., 1986 ; Laiho & Prescott, 2004 ).倒木在物理、化学和生物(特别是微生物、昆虫)等作用下进行的分解过程不仅可为分解者(如微生物、无脊椎动物、鸟类等)提供生存环境和必需的营养元素(Sollins, 1982 ), 而且能够释放大量的化学元素到土壤中, 丰富土壤有机质含量和种类, 促进土壤结构和功能的发育(Spears et al ., 2003 ).影响倒木分解的因素有基质质量、分解者活性以及环境因素等(如温度、水分、气体)(Harmon et al ., 1986).而在同一环境下, 不同树种间倒木分解速率也有很大差异, 这与倒木自身的物理和化学特性有关(van Geffen et al ., 2010 ).含水率和木材密度是影响倒木分解的主要物理因素, 因而受到越来越多的关注.倒木含水率是影响倒木呼吸的重要因子, 当它超过一定的阈值后, 就会影响微生物的活动, 从而抑制倒木的分解(Bond-Lamberty et al ., 2002 ).木材分解是一个复杂的过程, 通常根据单位时间内木材质量损失或木材密度的变化, 可以定量地算出其分解速率; 木材密度与微生物侵入倒木进行分解活动的程度有密切关系, 较低密度的木质残体更容易被分解和具有更高的呼吸速率(Johnston & Crossley, 1996 ; Chamber et al ., 2001 ).影响倒木分解的化学因素主要是其易分解成分和难分解有机成分(如木质素等)的含量, 其中氮(N)、磷(P)、木质素、纤维素、C:N、C:P等是常见的衡量倒木质量的指标(Berg et al ., 2000 ), 它们不仅可以表征倒木分解过程中基质的质量变化和养分限制状况(Wassen et al ., 2005 ), 而且可以指示物质循环和能量流动的方向(Zhou et al ., 2007 ). ...
... 倒木的含水率、密度与纤维素含量和木质素含量具有显著的相关性, 这可能与倒木的呼吸作用有关.含水率和密度是影响倒木呼吸的主要物理因素, 倒木的呼吸作用主要是通过微生物的分解作用来进行的(Herrmann & Bauhus, 2013 ).微生物分解对水分的增加非常敏感, 在一定范围内, 较小的水分增加会导致微生物活性的急剧增大(Progar et al ., 2000 ).由于单糖等非结构性碳水化合物易被微生物分解且含量较低, 到分解中后期, 相比木质素, 纤维素由于其结构相对简单, 成为微生物分解的主要物质.随着倒木含水率的增加, 微生物对纤维素的分解加快, 纤维素含量下降, 而木质素在倒木中的含量相对升高, 从而导致含水率与纤维素含量的负相关, 与木质素含量正相关(表4 ).本文中木材密度对倒木纤维素和木质素的影响与含水率相反.密度低, 则微生物易于侵入进行分解活动, 且其持水能力强, 更易促进微生物对纤维素的分解(Chamber et al ., 2001 ).因此密度与木质素、纤维素的关系与含水率相反(表4 ).而倒木中C、N、P含量的变化与倒木含水率和密度之间的相关性实质上是腐烂等级分别对倒木物理化学性质的影响所导致的.C、N、P与含水率均表现为随着腐烂等级增加而升高,密度则下降.因此, 含水率与C、N、P含量呈正相关关系; 密度则与倒木C、N、P含量呈负相关关系. ...
川西高山森林倒木不同分解阶段的微生物群落变化特征. 应用与环境生物学报,
1
2014
... C是构成植物体干物质的最主要元素, N和P是生态系统最主要的营养元素.N和P养分的有效性是调节植物残体分解速率和生态系统碳平衡的一个主要因素, 同时, 植物残体中N、P的矿化为土壤提供了重要的无机营养元素.本研究发现, 除心材C含量外, 腐烂等级显著地影响倒木C、N、P的含量(p < 0.05)(表1 ).随着腐烂等级增加, 边材C含量呈上升趋势, 且腐烂等级V显著高于腐烂等级I-III的C含量(图2 ), 这与Sandström等(2007)和Harmon等(2013)对裸子植物倒木C含量的研究结果一致, 而与Lombardi等(2013)和常晨晖等(2015)对倒木C含量的研究结果相反; Bütler等(2007)则发现欧洲云杉倒木随着分解程度加深C含量保持稳定.这些研究表明腐烂等级对倒木C含量的影响因环境条件和树种不同而存在差异.杨方方等(2009)认为, 倒木C含量随腐烂等级增加而增加的原因可能是木质残体分解周期较长, C损失速度与木质体腐解速度不同步, 从而在高腐烂等级C含量升高, 但导致C含量升高的深层原因有待进一步研究.由于分解初期倒木心材和边材N含量比较低, 随着腐烂等级增加, 边材与环境接触面积增大, 水热条件及通气量得到改善, 促进了微生物对边材的入侵及繁殖.同时, 寄生性微生物通过无脊椎动物侵入倒木心材, 心材结构疏松, 有利于微生物的繁殖.随着微生物增多, 一些固氮细菌数量不断增多, 对环境中N固定能力增强, N含量升高.到分解后期, 微生物竞争加剧, 呼吸作用产生的CO2 对微生物的毒害抑制了微生物的繁殖, 使得固氮细菌数量减少(常晨晖等, 2014 ), 固N能力减弱, N含量下降, 从而导致随着腐烂等级增加倒木N含量呈先升高后降低的变化趋势.虽然在分解后期略有下降, 但差异不显著, 这与其他相关研究结果基本一致(Palviainen et al ., 2008 ; 张修玉等, 2009 ; Köster et al ., 2015 ).本研究中, 倒木腐烂等级I和II的P含量相对较少, 腐烂等级III开始逐渐增加, 与杨丽韫和代力民(2002)、Palviainen等(2010)和Lombardi等(2013)的研究结果一致.这一方面可能是由于一些木材腐朽菌的菌丝把营养从土壤转移到倒木中(Clinton et al ., 2009 ), 另一方面由于有机物质的分解导致P的相对富集(Manzoni et al ., 2010 ).本研究结果发现边材的总体平均N含量显著高于心材(图2 ), 与Cowling和Merrill (1966)对裸子植物的研究结果基本一致.纤维素和木质素是倒木基质的主要组成成分, 其降解是维持生态系统碳素平衡的不可缺少的过程(Fioretto et al ., 2005 ).本研究中腐烂等级显著地影响倒木纤维素和木质素的含量(p < 0.001)(表1 ), 随着腐烂等级增加, 纤维素含量呈下降趋势, 木质素含量表现为上升趋势(图2 ), 这与以往对其他地区针叶树的研究结果相似(Ganjegunte et al ., 2004 ; Bütler et al ., 2007 ; 常晨晖等, 2015 ).这是因为纤维素是由纤维二糖单体组成的碳水化合物, 其结构相对简单, 而且有相对较多种类的微生物能够分解, 因此更容易被接触并分解(Austin & Ballaré, 2010 ); 而木质素具有独特的复杂结构, 它被认为是倒木中最难降解的组分.在分解初期随着易分解组分(可溶性C、N、P等)的不断降解, 木质素等难分解组分所占比例有所提高, 随着分解程度加深, 木质素含量升高.另外, 有关研究发现, 在分解过程中, 微生物新陈代谢能形成木质素类似物(Brandt et al ., 2010 ), 从而使倒木中木质素含量显著增加. ...
高寒森林倒木在不同分解阶段的质量变化. 植物生态学报,
4
2015
... 近年来, 有关森林倒木物理化学性质的研究引起了广泛的关注.大部分研究结果表明, 随着腐烂等级增加, 倒木C含量呈下降趋势, N、P含量则与之相反(Palviainen et al ., 2008 ; 常晨晖等, 2015 ).很多研究认为物理化学性质的变化还与树种有关.如Köster等(2015)对同一地区主要倒木树种进行C、N含量和密度的测定发现, 欧洲云杉(Picea abies )倒木的C含量随着分解程度加深而增加, 但是欧洲山杨(Populus tremula )倒木的C含量随着分解程度加深呈下降趋势, Alnus glutinosa 倒木的C含量则变化不大; 随着分解程度加深, 不同树种的木材密度均下降, 但欧洲云杉、Alnus glutinosa 以及欧洲赤松(Pinus sylvestris )比其他3种阔叶树种下降程度小.以往对倒木物理化学性质的研究大多集中在不同腐烂等级和树种对倒木物理化学性质的影响, 而径级对倒木物理化学性质影响的研究尚处于初步探索阶段.通常情况下, 小径级倒木(胸径(DBH ) < 10 cm)在森林生态系统中的数量巨大且分解较快, 有利于有机物质矿化到土壤中被土壤吸收和利用(Moroni & Ryan, 2010 ).相对于小径级倒木, 大径级倒木心材比例较大, 由于心材储存了大量能源物质和难分解组分(Schwarze, 2007 ), 可能有较高的C含量和相对较低的养分含量, 但其结构疏松, 易于分解者活动和居住, 随着分解程度加深, 物理化学性质特征更加复杂(常晨晖等, 2015 ).Harmon等(1986)提出倒木中物理化学性质的变化可能与倒木的径级有关.吕明和等(2006)在对鼎湖山锥(Castanopsis chin- ensis )的粗木质残体的分解和元素动态分析中也指出, 在分解过程中直径5-10 cm的倒木中的N、P含量持续升高, 而直径10-20 cm的倒木中的N、P含量呈下降趋势.Sakai等(2012)在对日本扁柏(Chamae- cyparis obtusa )和日本柳杉(Cryptomeria japonica )的倒木密度以及C、N含量测定的研究中发现, 随着径级增加, 倒木密度随之下降, 且径级对处于腐烂等级I和II的两种倒木的密度影响显著.由此可见, 倒木中物理化学性质的变化不仅受到腐烂等级和径级的影响, 而且可能还受两者产生的交互作用的影响. ...
... ), 可能有较高的C含量和相对较低的养分含量, 但其结构疏松, 易于分解者活动和居住, 随着分解程度加深, 物理化学性质特征更加复杂(常晨晖等, 2015 ).Harmon等(1986)提出倒木中物理化学性质的变化可能与倒木的径级有关.吕明和等(2006)在对鼎湖山锥(Castanopsis chin- ensis )的粗木质残体的分解和元素动态分析中也指出, 在分解过程中直径5-10 cm的倒木中的N、P含量持续升高, 而直径10-20 cm的倒木中的N、P含量呈下降趋势.Sakai等(2012)在对日本扁柏(Chamae- cyparis obtusa )和日本柳杉(Cryptomeria japonica )的倒木密度以及C、N含量测定的研究中发现, 随着径级增加, 倒木密度随之下降, 且径级对处于腐烂等级I和II的两种倒木的密度影响显著.由此可见, 倒木中物理化学性质的变化不仅受到腐烂等级和径级的影响, 而且可能还受两者产生的交互作用的影响. ...
... 目前, 我国部分学者对长白山红松(Pinus koraiensis )暗针叶林(杨丽韫和代力民, 2002 )、大兴安岭落叶松(Larix gmelinii )林(辛魏巍等, 2012 )、鼎湖山季风常绿阔叶林(吕明和等, 2006 )以及亚高山针叶林(常晨晖等, 2015 )等森林类型中主要组成树种的倒木在分解过程的物理化学性质变化趋势进行了研究和评述, 而缺乏径级对倒木物理化学性质影响的研究.对不同径级和腐烂等级的倒木进行物理化学性质分析才能准确地量化和预测倒木C储量和养分含量, 同时能够更深入地了解倒木物理性质与化学性质之间的关系.阔叶红松林是我国东北东部山区的地带性顶极植被, 对其倒木物种组成、数量(刘妍妍和金光泽, 2010a )、贮量特征(代力民等, 2000 )以及分布格局(刘妍妍和金光泽, 2010b )曾进行过系统研究, 然而还缺乏倒木分解动态的详细研究.本研究以典型阔叶红松林的建群种红松倒木为研究对象, 探讨其不同径级与不同腐烂等级倒木心材和边材物理化学性质变化特征, 为未来预测倒木C贮量和养分含量以及生产力变化进行合理的评估提供重要的科学依据和基础数据. ...
... C是构成植物体干物质的最主要元素, N和P是生态系统最主要的营养元素.N和P养分的有效性是调节植物残体分解速率和生态系统碳平衡的一个主要因素, 同时, 植物残体中N、P的矿化为土壤提供了重要的无机营养元素.本研究发现, 除心材C含量外, 腐烂等级显著地影响倒木C、N、P的含量(p < 0.05)(表1 ).随着腐烂等级增加, 边材C含量呈上升趋势, 且腐烂等级V显著高于腐烂等级I-III的C含量(图2 ), 这与Sandström等(2007)和Harmon等(2013)对裸子植物倒木C含量的研究结果一致, 而与Lombardi等(2013)和常晨晖等(2015)对倒木C含量的研究结果相反; Bütler等(2007)则发现欧洲云杉倒木随着分解程度加深C含量保持稳定.这些研究表明腐烂等级对倒木C含量的影响因环境条件和树种不同而存在差异.杨方方等(2009)认为, 倒木C含量随腐烂等级增加而增加的原因可能是木质残体分解周期较长, C损失速度与木质体腐解速度不同步, 从而在高腐烂等级C含量升高, 但导致C含量升高的深层原因有待进一步研究.由于分解初期倒木心材和边材N含量比较低, 随着腐烂等级增加, 边材与环境接触面积增大, 水热条件及通气量得到改善, 促进了微生物对边材的入侵及繁殖.同时, 寄生性微生物通过无脊椎动物侵入倒木心材, 心材结构疏松, 有利于微生物的繁殖.随着微生物增多, 一些固氮细菌数量不断增多, 对环境中N固定能力增强, N含量升高.到分解后期, 微生物竞争加剧, 呼吸作用产生的CO2 对微生物的毒害抑制了微生物的繁殖, 使得固氮细菌数量减少(常晨晖等, 2014 ), 固N能力减弱, N含量下降, 从而导致随着腐烂等级增加倒木N含量呈先升高后降低的变化趋势.虽然在分解后期略有下降, 但差异不显著, 这与其他相关研究结果基本一致(Palviainen et al ., 2008 ; 张修玉等, 2009 ; Köster et al ., 2015 ).本研究中, 倒木腐烂等级I和II的P含量相对较少, 腐烂等级III开始逐渐增加, 与杨丽韫和代力民(2002)、Palviainen等(2010)和Lombardi等(2013)的研究结果一致.这一方面可能是由于一些木材腐朽菌的菌丝把营养从土壤转移到倒木中(Clinton et al ., 2009 ), 另一方面由于有机物质的分解导致P的相对富集(Manzoni et al ., 2010 ).本研究结果发现边材的总体平均N含量显著高于心材(图2 ), 与Cowling和Merrill (1966)对裸子植物的研究结果基本一致.纤维素和木质素是倒木基质的主要组成成分, 其降解是维持生态系统碳素平衡的不可缺少的过程(Fioretto et al ., 2005 ).本研究中腐烂等级显著地影响倒木纤维素和木质素的含量(p < 0.001)(表1 ), 随着腐烂等级增加, 纤维素含量呈下降趋势, 木质素含量表现为上升趋势(图2 ), 这与以往对其他地区针叶树的研究结果相似(Ganjegunte et al ., 2004 ; Bütler et al ., 2007 ; 常晨晖等, 2015 ).这是因为纤维素是由纤维二糖单体组成的碳水化合物, 其结构相对简单, 而且有相对较多种类的微生物能够分解, 因此更容易被接触并分解(Austin & Ballaré, 2010 ); 而木质素具有独特的复杂结构, 它被认为是倒木中最难降解的组分.在分解初期随着易分解组分(可溶性C、N、P等)的不断降解, 木质素等难分解组分所占比例有所提高, 随着分解程度加深, 木质素含量升高.另外, 有关研究发现, 在分解过程中, 微生物新陈代谢能形成木质素类似物(Brandt et al ., 2010 ), 从而使倒木中木质素含量显著增加. ...
Decomposition of
1
2009
... C是构成植物体干物质的最主要元素, N和P是生态系统最主要的营养元素.N和P养分的有效性是调节植物残体分解速率和生态系统碳平衡的一个主要因素, 同时, 植物残体中N、P的矿化为土壤提供了重要的无机营养元素.本研究发现, 除心材C含量外, 腐烂等级显著地影响倒木C、N、P的含量(p < 0.05)(表1 ).随着腐烂等级增加, 边材C含量呈上升趋势, 且腐烂等级V显著高于腐烂等级I-III的C含量(图2 ), 这与Sandström等(2007)和Harmon等(2013)对裸子植物倒木C含量的研究结果一致, 而与Lombardi等(2013)和常晨晖等(2015)对倒木C含量的研究结果相反; Bütler等(2007)则发现欧洲云杉倒木随着分解程度加深C含量保持稳定.这些研究表明腐烂等级对倒木C含量的影响因环境条件和树种不同而存在差异.杨方方等(2009)认为, 倒木C含量随腐烂等级增加而增加的原因可能是木质残体分解周期较长, C损失速度与木质体腐解速度不同步, 从而在高腐烂等级C含量升高, 但导致C含量升高的深层原因有待进一步研究.由于分解初期倒木心材和边材N含量比较低, 随着腐烂等级增加, 边材与环境接触面积增大, 水热条件及通气量得到改善, 促进了微生物对边材的入侵及繁殖.同时, 寄生性微生物通过无脊椎动物侵入倒木心材, 心材结构疏松, 有利于微生物的繁殖.随着微生物增多, 一些固氮细菌数量不断增多, 对环境中N固定能力增强, N含量升高.到分解后期, 微生物竞争加剧, 呼吸作用产生的CO2 对微生物的毒害抑制了微生物的繁殖, 使得固氮细菌数量减少(常晨晖等, 2014 ), 固N能力减弱, N含量下降, 从而导致随着腐烂等级增加倒木N含量呈先升高后降低的变化趋势.虽然在分解后期略有下降, 但差异不显著, 这与其他相关研究结果基本一致(Palviainen et al ., 2008 ; 张修玉等, 2009 ; Köster et al ., 2015 ).本研究中, 倒木腐烂等级I和II的P含量相对较少, 腐烂等级III开始逐渐增加, 与杨丽韫和代力民(2002)、Palviainen等(2010)和Lombardi等(2013)的研究结果一致.这一方面可能是由于一些木材腐朽菌的菌丝把营养从土壤转移到倒木中(Clinton et al ., 2009 ), 另一方面由于有机物质的分解导致P的相对富集(Manzoni et al ., 2010 ).本研究结果发现边材的总体平均N含量显著高于心材(图2 ), 与Cowling和Merrill (1966)对裸子植物的研究结果基本一致.纤维素和木质素是倒木基质的主要组成成分, 其降解是维持生态系统碳素平衡的不可缺少的过程(Fioretto et al ., 2005 ).本研究中腐烂等级显著地影响倒木纤维素和木质素的含量(p < 0.001)(表1 ), 随着腐烂等级增加, 纤维素含量呈下降趋势, 木质素含量表现为上升趋势(图2 ), 这与以往对其他地区针叶树的研究结果相似(Ganjegunte et al ., 2004 ; Bütler et al ., 2007 ; 常晨晖等, 2015 ).这是因为纤维素是由纤维二糖单体组成的碳水化合物, 其结构相对简单, 而且有相对较多种类的微生物能够分解, 因此更容易被接触并分解(Austin & Ballaré, 2010 ); 而木质素具有独特的复杂结构, 它被认为是倒木中最难降解的组分.在分解初期随着易分解组分(可溶性C、N、P等)的不断降解, 木质素等难分解组分所占比例有所提高, 随着分解程度加深, 木质素含量升高.另外, 有关研究发现, 在分解过程中, 微生物新陈代谢能形成木质素类似物(Brandt et al ., 2010 ), 从而使倒木中木质素含量显著增加. ...
Nitrogen in wood and its role in wood deterioration
1966
阔叶红松林倒木贮量的变化规律. 生态学报,
1
2000
... 目前, 我国部分学者对长白山红松(Pinus koraiensis )暗针叶林(杨丽韫和代力民, 2002 )、大兴安岭落叶松(Larix gmelinii )林(辛魏巍等, 2012 )、鼎湖山季风常绿阔叶林(吕明和等, 2006 )以及亚高山针叶林(常晨晖等, 2015 )等森林类型中主要组成树种的倒木在分解过程的物理化学性质变化趋势进行了研究和评述, 而缺乏径级对倒木物理化学性质影响的研究.对不同径级和腐烂等级的倒木进行物理化学性质分析才能准确地量化和预测倒木C储量和养分含量, 同时能够更深入地了解倒木物理性质与化学性质之间的关系.阔叶红松林是我国东北东部山区的地带性顶极植被, 对其倒木物种组成、数量(刘妍妍和金光泽, 2010a )、贮量特征(代力民等, 2000 )以及分布格局(刘妍妍和金光泽, 2010b )曾进行过系统研究, 然而还缺乏倒木分解动态的详细研究.本研究以典型阔叶红松林的建群种红松倒木为研究对象, 探讨其不同径级与不同腐烂等级倒木心材和边材物理化学性质变化特征, 为未来预测倒木C贮量和养分含量以及生产力变化进行合理的评估提供重要的科学依据和基础数据. ...
Carbon in heartwood, sapwood and bark along the stem profile in three Mediterranean
2011
Lignin and cellulose degradation and nitrogen dynamics during de- composition of three leaf litter species in a Mediterranean ecosystem
1
2005
... C是构成植物体干物质的最主要元素, N和P是生态系统最主要的营养元素.N和P养分的有效性是调节植物残体分解速率和生态系统碳平衡的一个主要因素, 同时, 植物残体中N、P的矿化为土壤提供了重要的无机营养元素.本研究发现, 除心材C含量外, 腐烂等级显著地影响倒木C、N、P的含量(p < 0.05)(表1 ).随着腐烂等级增加, 边材C含量呈上升趋势, 且腐烂等级V显著高于腐烂等级I-III的C含量(图2 ), 这与Sandström等(2007)和Harmon等(2013)对裸子植物倒木C含量的研究结果一致, 而与Lombardi等(2013)和常晨晖等(2015)对倒木C含量的研究结果相反; Bütler等(2007)则发现欧洲云杉倒木随着分解程度加深C含量保持稳定.这些研究表明腐烂等级对倒木C含量的影响因环境条件和树种不同而存在差异.杨方方等(2009)认为, 倒木C含量随腐烂等级增加而增加的原因可能是木质残体分解周期较长, C损失速度与木质体腐解速度不同步, 从而在高腐烂等级C含量升高, 但导致C含量升高的深层原因有待进一步研究.由于分解初期倒木心材和边材N含量比较低, 随着腐烂等级增加, 边材与环境接触面积增大, 水热条件及通气量得到改善, 促进了微生物对边材的入侵及繁殖.同时, 寄生性微生物通过无脊椎动物侵入倒木心材, 心材结构疏松, 有利于微生物的繁殖.随着微生物增多, 一些固氮细菌数量不断增多, 对环境中N固定能力增强, N含量升高.到分解后期, 微生物竞争加剧, 呼吸作用产生的CO2 对微生物的毒害抑制了微生物的繁殖, 使得固氮细菌数量减少(常晨晖等, 2014 ), 固N能力减弱, N含量下降, 从而导致随着腐烂等级增加倒木N含量呈先升高后降低的变化趋势.虽然在分解后期略有下降, 但差异不显著, 这与其他相关研究结果基本一致(Palviainen et al ., 2008 ; 张修玉等, 2009 ; Köster et al ., 2015 ).本研究中, 倒木腐烂等级I和II的P含量相对较少, 腐烂等级III开始逐渐增加, 与杨丽韫和代力民(2002)、Palviainen等(2010)和Lombardi等(2013)的研究结果一致.这一方面可能是由于一些木材腐朽菌的菌丝把营养从土壤转移到倒木中(Clinton et al ., 2009 ), 另一方面由于有机物质的分解导致P的相对富集(Manzoni et al ., 2010 ).本研究结果发现边材的总体平均N含量显著高于心材(图2 ), 与Cowling和Merrill (1966)对裸子植物的研究结果基本一致.纤维素和木质素是倒木基质的主要组成成分, 其降解是维持生态系统碳素平衡的不可缺少的过程(Fioretto et al ., 2005 ).本研究中腐烂等级显著地影响倒木纤维素和木质素的含量(p < 0.001)(表1 ), 随着腐烂等级增加, 纤维素含量呈下降趋势, 木质素含量表现为上升趋势(图2 ), 这与以往对其他地区针叶树的研究结果相似(Ganjegunte et al ., 2004 ; Bütler et al ., 2007 ; 常晨晖等, 2015 ).这是因为纤维素是由纤维二糖单体组成的碳水化合物, 其结构相对简单, 而且有相对较多种类的微生物能够分解, 因此更容易被接触并分解(Austin & Ballaré, 2010 ); 而木质素具有独特的复杂结构, 它被认为是倒木中最难降解的组分.在分解初期随着易分解组分(可溶性C、N、P等)的不断降解, 木质素等难分解组分所占比例有所提高, 随着分解程度加深, 木质素含量升高.另外, 有关研究发现, 在分解过程中, 微生物新陈代谢能形成木质素类似物(Brandt et al ., 2010 ), 从而使倒木中木质素含量显著增加. ...
Decomposition and nutrient release from radiata pine (
1
2004
... C是构成植物体干物质的最主要元素, N和P是生态系统最主要的营养元素.N和P养分的有效性是调节植物残体分解速率和生态系统碳平衡的一个主要因素, 同时, 植物残体中N、P的矿化为土壤提供了重要的无机营养元素.本研究发现, 除心材C含量外, 腐烂等级显著地影响倒木C、N、P的含量(p < 0.05)(表1 ).随着腐烂等级增加, 边材C含量呈上升趋势, 且腐烂等级V显著高于腐烂等级I-III的C含量(图2 ), 这与Sandström等(2007)和Harmon等(2013)对裸子植物倒木C含量的研究结果一致, 而与Lombardi等(2013)和常晨晖等(2015)对倒木C含量的研究结果相反; Bütler等(2007)则发现欧洲云杉倒木随着分解程度加深C含量保持稳定.这些研究表明腐烂等级对倒木C含量的影响因环境条件和树种不同而存在差异.杨方方等(2009)认为, 倒木C含量随腐烂等级增加而增加的原因可能是木质残体分解周期较长, C损失速度与木质体腐解速度不同步, 从而在高腐烂等级C含量升高, 但导致C含量升高的深层原因有待进一步研究.由于分解初期倒木心材和边材N含量比较低, 随着腐烂等级增加, 边材与环境接触面积增大, 水热条件及通气量得到改善, 促进了微生物对边材的入侵及繁殖.同时, 寄生性微生物通过无脊椎动物侵入倒木心材, 心材结构疏松, 有利于微生物的繁殖.随着微生物增多, 一些固氮细菌数量不断增多, 对环境中N固定能力增强, N含量升高.到分解后期, 微生物竞争加剧, 呼吸作用产生的CO2 对微生物的毒害抑制了微生物的繁殖, 使得固氮细菌数量减少(常晨晖等, 2014 ), 固N能力减弱, N含量下降, 从而导致随着腐烂等级增加倒木N含量呈先升高后降低的变化趋势.虽然在分解后期略有下降, 但差异不显著, 这与其他相关研究结果基本一致(Palviainen et al ., 2008 ; 张修玉等, 2009 ; Köster et al ., 2015 ).本研究中, 倒木腐烂等级I和II的P含量相对较少, 腐烂等级III开始逐渐增加, 与杨丽韫和代力民(2002)、Palviainen等(2010)和Lombardi等(2013)的研究结果一致.这一方面可能是由于一些木材腐朽菌的菌丝把营养从土壤转移到倒木中(Clinton et al ., 2009 ), 另一方面由于有机物质的分解导致P的相对富集(Manzoni et al ., 2010 ).本研究结果发现边材的总体平均N含量显著高于心材(图2 ), 与Cowling和Merrill (1966)对裸子植物的研究结果基本一致.纤维素和木质素是倒木基质的主要组成成分, 其降解是维持生态系统碳素平衡的不可缺少的过程(Fioretto et al ., 2005 ).本研究中腐烂等级显著地影响倒木纤维素和木质素的含量(p < 0.001)(表1 ), 随着腐烂等级增加, 纤维素含量呈下降趋势, 木质素含量表现为上升趋势(图2 ), 这与以往对其他地区针叶树的研究结果相似(Ganjegunte et al ., 2004 ; Bütler et al ., 2007 ; 常晨晖等, 2015 ).这是因为纤维素是由纤维二糖单体组成的碳水化合物, 其结构相对简单, 而且有相对较多种类的微生物能够分解, 因此更容易被接触并分解(Austin & Ballaré, 2010 ); 而木质素具有独特的复杂结构, 它被认为是倒木中最难降解的组分.在分解初期随着易分解组分(可溶性C、N、P等)的不断降解, 木质素等难分解组分所占比例有所提高, 随着分解程度加深, 木质素含量升高.另外, 有关研究发现, 在分解过程中, 微生物新陈代谢能形成木质素类似物(Brandt et al ., 2010 ), 从而使倒木中木质素含量显著增加. ...
Carbon concentration of standing and downed woody detritus: Effects of tree taxa, decay class, position, and tissue type
2013
Ecology of coarse woody debris in temperate ecosystems
3
1986
... 森林是陆地生态系统的重要组成部分, 其植被碳(C)库约占全球植被C库的80% (Köster et al ., 2015 ).倒木是森林生态系统的重要组成部分, 对于维持生物多样性以及林木的更新具有重要作用; 同时, 倒木也是重要的养分库和C库, 其养分的缓慢释放, 对于提高林地的生产力有重要意义(Harmon et al ., 1986 ; Laiho & Prescott, 2004 ).倒木在物理、化学和生物(特别是微生物、昆虫)等作用下进行的分解过程不仅可为分解者(如微生物、无脊椎动物、鸟类等)提供生存环境和必需的营养元素(Sollins, 1982 ), 而且能够释放大量的化学元素到土壤中, 丰富土壤有机质含量和种类, 促进土壤结构和功能的发育(Spears et al ., 2003 ).影响倒木分解的因素有基质质量、分解者活性以及环境因素等(如温度、水分、气体)(Harmon et al ., 1986).而在同一环境下, 不同树种间倒木分解速率也有很大差异, 这与倒木自身的物理和化学特性有关(van Geffen et al ., 2010 ).含水率和木材密度是影响倒木分解的主要物理因素, 因而受到越来越多的关注.倒木含水率是影响倒木呼吸的重要因子, 当它超过一定的阈值后, 就会影响微生物的活动, 从而抑制倒木的分解(Bond-Lamberty et al ., 2002 ).木材分解是一个复杂的过程, 通常根据单位时间内木材质量损失或木材密度的变化, 可以定量地算出其分解速率; 木材密度与微生物侵入倒木进行分解活动的程度有密切关系, 较低密度的木质残体更容易被分解和具有更高的呼吸速率(Johnston & Crossley, 1996 ; Chamber et al ., 2001 ).影响倒木分解的化学因素主要是其易分解成分和难分解有机成分(如木质素等)的含量, 其中氮(N)、磷(P)、木质素、纤维素、C:N、C:P等是常见的衡量倒木质量的指标(Berg et al ., 2000 ), 它们不仅可以表征倒木分解过程中基质的质量变化和养分限制状况(Wassen et al ., 2005 ), 而且可以指示物质循环和能量流动的方向(Zhou et al ., 2007 ). ...
... 本研究发现, 腐烂等级对倒木含水率和密度产生极显著影响(p < 0.001)(表1 ).随着腐烂等级增加, 倒木含水率升高, 密度下降(图1A 、1B).这与其他地区对倒木含水率和密度的研究结果一致(吕明和等, 2006 ; Seedre et al ., 2013 ; Petrillo et al ., 2015 ).其原因主要是由于随着分解程度加深, 倒木受到淋溶、破碎化以及呼吸作用的影响, 木质变得疏松, 从而导致倒木密度下降; 随着腐烂等级增加, 其木质越疏松, 越容易吸收降水(袁杰和张硕新, 2012 ); 其次, 腐烂程度越高, 倒木与地面接触面积越大, 影响倒木含水率变化的因素增加, 例如土壤的含水率、地表水和微地貌等(Bütler et al ., 2007 ).本研究显示, 径级i-iii倒木密度呈下降趋势, 径级iv略有上升, 但与径级iii差异不显著(图1D ), 这与Sakai等(2012)的研究结果基本一致.倒木密度受许多因素的影响, 例如树种、地理环境、树龄、增长速率以及遗传因素等(Harmon et al ., 1986 ; Repola, 2006 ).Spurr和Hsuing (1954)发现, 从心材到边材, 幼龄树的密度比老龄树的大.本实验所测红松倒木的直径位置为胸径处, 可以间接代表龄级.相比大径级倒木, 小径级的倒木树龄小, 密度相对较大, 因此树龄是造成随着径级增加密度减小的主要原因. ...
... 心材N、P含量在不同径级间差异显著(p < 0.05), 心材和边材的纤维素和木质素含量受腐烂等级和径级交互作用影响显著(p < 0.05)(表1 ).随着径级增加, 倒木心材和边材N、P含量呈先增加后减少的趋势, 径级iii倒木N、P含量最高, 且径级iii的心材P含量显著高于其他径级(p < 0.05)(图3 ), 这表明N、P含量在不同径级间存在差异, 且中等径级的倒木能够为生态系统提供更重要的养分来源.造成差异的原因主要是不同径级下倒木自身特性、物理结构以及分解者组成的影响.Stokland和Kauserud (2004)曾提出小倒木可能没有充足的养分供应分解者生长.小径级的倒木, 由于生长期相对比较短, 其本身所含养分量较大径级倒木少.其次, Harmon等(1986)发现随着径级增大, 倒木比表面积减小.这使得淋溶作用对P含量损失影响减少, 从而导致P含量不断积累.本研究中腐烂等级i至iii的倒木P含量呈上升趋势, 在腐烂等级iv倒木P含量下降, 这表明可能存在一个P含量积累的比表面积临界值, 超过临界值, 则淋溶损失速度大于积累速度导致P含量下降.分解者是倒木分解的主要影响因素, 它包括真菌、细菌、无脊椎动物等(Harmon et al ., 1986 ).径级的大小影响分解者的种类和数量(Küffer & Senn-Irlet, 2005 ; Juutilainen et al ., 2011 ).Brin等(2011)对不同径级倒木中寄生于枯木的无脊椎动物甲虫的研究发现, 大径级倒木中有更多的甲虫物种, 且不同径级倒木中的甲虫物种组成不同.Nordén等(2004)在瑞典南部阔叶林进行粗木质残体(CWD)和细木质残体(FWD)对真菌重要性的研究中发现FWD有更多子囊菌种类, CWD倒木上出现更多担子菌类, 而分解者对化学成分的不同利用影响不同径级倒木养分含量的分布. ...
Lignin degradation in foliar litter of two shrub species from the gap center to the closed canopy in an alpine fir forest
1
2016
... 将采集的样品粉碎, 过60目筛, 于65 ℃条件下烘干至恒质量, 存放于干燥环境中待测.采用燃烧法通过multi N/C 3000分析仪(Analytik Jena AG, Jena, Germany)测定倒木全C含量; 采用消煮法通过Kjeltec TM 2300凯氏定氮仪(Foss Teactor AB, Hoganas, Sweden)测定倒木全N含量; 采用钼锑钪比色法通过722N分光光度计(上海精密科学仪器有限公司, 上海)测定其全P含量; 采用改进的Van Soest酸性洗涤木质素方法(He et al ., 2016 )测定其木质素和纤维素含量. ...
Effects of moisture, temperature and decomposition stage on respirational carbon loss from coarse woody debris (CWD) of important European tree species
1
2013
... 倒木的含水率、密度与纤维素含量和木质素含量具有显著的相关性, 这可能与倒木的呼吸作用有关.含水率和密度是影响倒木呼吸的主要物理因素, 倒木的呼吸作用主要是通过微生物的分解作用来进行的(Herrmann & Bauhus, 2013 ).微生物分解对水分的增加非常敏感, 在一定范围内, 较小的水分增加会导致微生物活性的急剧增大(Progar et al ., 2000 ).由于单糖等非结构性碳水化合物易被微生物分解且含量较低, 到分解中后期, 相比木质素, 纤维素由于其结构相对简单, 成为微生物分解的主要物质.随着倒木含水率的增加, 微生物对纤维素的分解加快, 纤维素含量下降, 而木质素在倒木中的含量相对升高, 从而导致含水率与纤维素含量的负相关, 与木质素含量正相关(表4 ).本文中木材密度对倒木纤维素和木质素的影响与含水率相反.密度低, 则微生物易于侵入进行分解活动, 且其持水能力强, 更易促进微生物对纤维素的分解(Chamber et al ., 2001 ).因此密度与木质素、纤维素的关系与含水率相反(表4 ).而倒木中C、N、P含量的变化与倒木含水率和密度之间的相关性实质上是腐烂等级分别对倒木物理化学性质的影响所导致的.C、N、P与含水率均表现为随着腐烂等级增加而升高,密度则下降.因此, 含水率与C、N、P含量呈正相关关系; 密度则与倒木C、N、P含量呈负相关关系. ...
The pattern of seed rain in the broadleaved-Korean pine mixed forest of Xiaoxing’an Mountains, China
1
2006
... 本研究在凉水自然保护区9 hm2 (300 m × 300 m)的阔叶红松林永久性固定样地上进行(Jin et al ., 2006 ), 将样地设成900个10 m × 10 m的小样方, 采用相邻网格法进行调查.对样地内所有胸径≥2 cm的木质残体进行调查, 确定木质残体的存在形式(干基折断、干中折断、枯立木、拔根倒、根桩和树段); 测量立木形式的胸径, 对倒伏的木质残体, 测量其中央直径和胸径; 采用Vertex III超声波测高仪(Haglöf Sweden AB, Långsele, Sweden)进行长度和高度的测定, 识别树种, 确定坐标位置, 并钉铝制树牌进行编号.采用5级分类系统进行腐烂等级的划分(闫恩荣等, 2005 ). ...
The significance of coarse woody debris for the diversity of soil mites
1
1996
... 森林是陆地生态系统的重要组成部分, 其植被碳(C)库约占全球植被C库的80% (Köster et al ., 2015 ).倒木是森林生态系统的重要组成部分, 对于维持生物多样性以及林木的更新具有重要作用; 同时, 倒木也是重要的养分库和C库, 其养分的缓慢释放, 对于提高林地的生产力有重要意义(Harmon et al ., 1986 ; Laiho & Prescott, 2004 ).倒木在物理、化学和生物(特别是微生物、昆虫)等作用下进行的分解过程不仅可为分解者(如微生物、无脊椎动物、鸟类等)提供生存环境和必需的营养元素(Sollins, 1982 ), 而且能够释放大量的化学元素到土壤中, 丰富土壤有机质含量和种类, 促进土壤结构和功能的发育(Spears et al ., 2003 ).影响倒木分解的因素有基质质量、分解者活性以及环境因素等(如温度、水分、气体)(Harmon et al ., 1986).而在同一环境下, 不同树种间倒木分解速率也有很大差异, 这与倒木自身的物理和化学特性有关(van Geffen et al ., 2010 ).含水率和木材密度是影响倒木分解的主要物理因素, 因而受到越来越多的关注.倒木含水率是影响倒木呼吸的重要因子, 当它超过一定的阈值后, 就会影响微生物的活动, 从而抑制倒木的分解(Bond-Lamberty et al ., 2002 ).木材分解是一个复杂的过程, 通常根据单位时间内木材质量损失或木材密度的变化, 可以定量地算出其分解速率; 木材密度与微生物侵入倒木进行分解活动的程度有密切关系, 较低密度的木质残体更容易被分解和具有更高的呼吸速率(Johnston & Crossley, 1996 ; Chamber et al ., 2001 ).影响倒木分解的化学因素主要是其易分解成分和难分解有机成分(如木质素等)的含量, 其中氮(N)、磷(P)、木质素、纤维素、C:N、C:P等是常见的衡量倒木质量的指标(Berg et al ., 2000 ), 它们不仅可以表征倒木分解过程中基质的质量变化和养分限制状况(Wassen et al ., 2005 ), 而且可以指示物质循环和能量流动的方向(Zhou et al ., 2007 ). ...
Diameter matters in studies of dead wood and wood-inhabiting fungi
1
2011
... 心材N、P含量在不同径级间差异显著(p < 0.05), 心材和边材的纤维素和木质素含量受腐烂等级和径级交互作用影响显著(p < 0.05)(表1 ).随着径级增加, 倒木心材和边材N、P含量呈先增加后减少的趋势, 径级iii倒木N、P含量最高, 且径级iii的心材P含量显著高于其他径级(p < 0.05)(图3 ), 这表明N、P含量在不同径级间存在差异, 且中等径级的倒木能够为生态系统提供更重要的养分来源.造成差异的原因主要是不同径级下倒木自身特性、物理结构以及分解者组成的影响.Stokland和Kauserud (2004)曾提出小倒木可能没有充足的养分供应分解者生长.小径级的倒木, 由于生长期相对比较短, 其本身所含养分量较大径级倒木少.其次, Harmon等(1986)发现随着径级增大, 倒木比表面积减小.这使得淋溶作用对P含量损失影响减少, 从而导致P含量不断积累.本研究中腐烂等级i至iii的倒木P含量呈上升趋势, 在腐烂等级iv倒木P含量下降, 这表明可能存在一个P含量积累的比表面积临界值, 超过临界值, 则淋溶损失速度大于积累速度导致P含量下降.分解者是倒木分解的主要影响因素, 它包括真菌、细菌、无脊椎动物等(Harmon et al ., 1986 ).径级的大小影响分解者的种类和数量(Küffer & Senn-Irlet, 2005 ; Juutilainen et al ., 2011 ).Brin等(2011)对不同径级倒木中寄生于枯木的无脊椎动物甲虫的研究发现, 大径级倒木中有更多的甲虫物种, 且不同径级倒木中的甲虫物种组成不同.Nordén等(2004)在瑞典南部阔叶林进行粗木质残体(CWD)和细木质残体(FWD)对真菌重要性的研究中发现FWD有更多子囊菌种类, CWD倒木上出现更多担子菌类, 而分解者对化学成分的不同利用影响不同径级倒木养分含量的分布. ...
Dead wood basic density, and the concentration of carbon and nitrogen for main tree species in managed hemiboreal forests
2
2015
... 森林是陆地生态系统的重要组成部分, 其植被碳(C)库约占全球植被C库的80% (Köster et al ., 2015 ).倒木是森林生态系统的重要组成部分, 对于维持生物多样性以及林木的更新具有重要作用; 同时, 倒木也是重要的养分库和C库, 其养分的缓慢释放, 对于提高林地的生产力有重要意义(Harmon et al ., 1986 ; Laiho & Prescott, 2004 ).倒木在物理、化学和生物(特别是微生物、昆虫)等作用下进行的分解过程不仅可为分解者(如微生物、无脊椎动物、鸟类等)提供生存环境和必需的营养元素(Sollins, 1982 ), 而且能够释放大量的化学元素到土壤中, 丰富土壤有机质含量和种类, 促进土壤结构和功能的发育(Spears et al ., 2003 ).影响倒木分解的因素有基质质量、分解者活性以及环境因素等(如温度、水分、气体)(Harmon et al ., 1986).而在同一环境下, 不同树种间倒木分解速率也有很大差异, 这与倒木自身的物理和化学特性有关(van Geffen et al ., 2010 ).含水率和木材密度是影响倒木分解的主要物理因素, 因而受到越来越多的关注.倒木含水率是影响倒木呼吸的重要因子, 当它超过一定的阈值后, 就会影响微生物的活动, 从而抑制倒木的分解(Bond-Lamberty et al ., 2002 ).木材分解是一个复杂的过程, 通常根据单位时间内木材质量损失或木材密度的变化, 可以定量地算出其分解速率; 木材密度与微生物侵入倒木进行分解活动的程度有密切关系, 较低密度的木质残体更容易被分解和具有更高的呼吸速率(Johnston & Crossley, 1996 ; Chamber et al ., 2001 ).影响倒木分解的化学因素主要是其易分解成分和难分解有机成分(如木质素等)的含量, 其中氮(N)、磷(P)、木质素、纤维素、C:N、C:P等是常见的衡量倒木质量的指标(Berg et al ., 2000 ), 它们不仅可以表征倒木分解过程中基质的质量变化和养分限制状况(Wassen et al ., 2005 ), 而且可以指示物质循环和能量流动的方向(Zhou et al ., 2007 ). ...
... C是构成植物体干物质的最主要元素, N和P是生态系统最主要的营养元素.N和P养分的有效性是调节植物残体分解速率和生态系统碳平衡的一个主要因素, 同时, 植物残体中N、P的矿化为土壤提供了重要的无机营养元素.本研究发现, 除心材C含量外, 腐烂等级显著地影响倒木C、N、P的含量(p < 0.05)(表1 ).随着腐烂等级增加, 边材C含量呈上升趋势, 且腐烂等级V显著高于腐烂等级I-III的C含量(图2 ), 这与Sandström等(2007)和Harmon等(2013)对裸子植物倒木C含量的研究结果一致, 而与Lombardi等(2013)和常晨晖等(2015)对倒木C含量的研究结果相反; Bütler等(2007)则发现欧洲云杉倒木随着分解程度加深C含量保持稳定.这些研究表明腐烂等级对倒木C含量的影响因环境条件和树种不同而存在差异.杨方方等(2009)认为, 倒木C含量随腐烂等级增加而增加的原因可能是木质残体分解周期较长, C损失速度与木质体腐解速度不同步, 从而在高腐烂等级C含量升高, 但导致C含量升高的深层原因有待进一步研究.由于分解初期倒木心材和边材N含量比较低, 随着腐烂等级增加, 边材与环境接触面积增大, 水热条件及通气量得到改善, 促进了微生物对边材的入侵及繁殖.同时, 寄生性微生物通过无脊椎动物侵入倒木心材, 心材结构疏松, 有利于微生物的繁殖.随着微生物增多, 一些固氮细菌数量不断增多, 对环境中N固定能力增强, N含量升高.到分解后期, 微生物竞争加剧, 呼吸作用产生的CO2 对微生物的毒害抑制了微生物的繁殖, 使得固氮细菌数量减少(常晨晖等, 2014 ), 固N能力减弱, N含量下降, 从而导致随着腐烂等级增加倒木N含量呈先升高后降低的变化趋势.虽然在分解后期略有下降, 但差异不显著, 这与其他相关研究结果基本一致(Palviainen et al ., 2008 ; 张修玉等, 2009 ; Köster et al ., 2015 ).本研究中, 倒木腐烂等级I和II的P含量相对较少, 腐烂等级III开始逐渐增加, 与杨丽韫和代力民(2002)、Palviainen等(2010)和Lombardi等(2013)的研究结果一致.这一方面可能是由于一些木材腐朽菌的菌丝把营养从土壤转移到倒木中(Clinton et al ., 2009 ), 另一方面由于有机物质的分解导致P的相对富集(Manzoni et al ., 2010 ).本研究结果发现边材的总体平均N含量显著高于心材(图2 ), 与Cowling和Merrill (1966)对裸子植物的研究结果基本一致.纤维素和木质素是倒木基质的主要组成成分, 其降解是维持生态系统碳素平衡的不可缺少的过程(Fioretto et al ., 2005 ).本研究中腐烂等级显著地影响倒木纤维素和木质素的含量(p < 0.001)(表1 ), 随着腐烂等级增加, 纤维素含量呈下降趋势, 木质素含量表现为上升趋势(图2 ), 这与以往对其他地区针叶树的研究结果相似(Ganjegunte et al ., 2004 ; Bütler et al ., 2007 ; 常晨晖等, 2015 ).这是因为纤维素是由纤维二糖单体组成的碳水化合物, 其结构相对简单, 而且有相对较多种类的微生物能够分解, 因此更容易被接触并分解(Austin & Ballaré, 2010 ); 而木质素具有独特的复杂结构, 它被认为是倒木中最难降解的组分.在分解初期随着易分解组分(可溶性C、N、P等)的不断降解, 木质素等难分解组分所占比例有所提高, 随着分解程度加深, 木质素含量升高.另外, 有关研究发现, 在分解过程中, 微生物新陈代谢能形成木质素类似物(Brandt et al ., 2010 ), 从而使倒木中木质素含量显著增加. ...
Influence of forest management on the species richness and composition of wood- inhabiting basidiomycetes in Swiss forests
1
2005
... 心材N、P含量在不同径级间差异显著(p < 0.05), 心材和边材的纤维素和木质素含量受腐烂等级和径级交互作用影响显著(p < 0.05)(表1 ).随着径级增加, 倒木心材和边材N、P含量呈先增加后减少的趋势, 径级iii倒木N、P含量最高, 且径级iii的心材P含量显著高于其他径级(p < 0.05)(图3 ), 这表明N、P含量在不同径级间存在差异, 且中等径级的倒木能够为生态系统提供更重要的养分来源.造成差异的原因主要是不同径级下倒木自身特性、物理结构以及分解者组成的影响.Stokland和Kauserud (2004)曾提出小倒木可能没有充足的养分供应分解者生长.小径级的倒木, 由于生长期相对比较短, 其本身所含养分量较大径级倒木少.其次, Harmon等(1986)发现随着径级增大, 倒木比表面积减小.这使得淋溶作用对P含量损失影响减少, 从而导致P含量不断积累.本研究中腐烂等级i至iii的倒木P含量呈上升趋势, 在腐烂等级iv倒木P含量下降, 这表明可能存在一个P含量积累的比表面积临界值, 超过临界值, 则淋溶损失速度大于积累速度导致P含量下降.分解者是倒木分解的主要影响因素, 它包括真菌、细菌、无脊椎动物等(Harmon et al ., 1986 ).径级的大小影响分解者的种类和数量(Küffer & Senn-Irlet, 2005 ; Juutilainen et al ., 2011 ).Brin等(2011)对不同径级倒木中寄生于枯木的无脊椎动物甲虫的研究发现, 大径级倒木中有更多的甲虫物种, 且不同径级倒木中的甲虫物种组成不同.Nordén等(2004)在瑞典南部阔叶林进行粗木质残体(CWD)和细木质残体(FWD)对真菌重要性的研究中发现FWD有更多子囊菌种类, CWD倒木上出现更多担子菌类, 而分解者对化学成分的不同利用影响不同径级倒木养分含量的分布. ...
Decay and nutrient dynamics of coarse woody debris in northern coniferous forests: A synthesis
1
2004
... 森林是陆地生态系统的重要组成部分, 其植被碳(C)库约占全球植被C库的80% (Köster et al ., 2015 ).倒木是森林生态系统的重要组成部分, 对于维持生物多样性以及林木的更新具有重要作用; 同时, 倒木也是重要的养分库和C库, 其养分的缓慢释放, 对于提高林地的生产力有重要意义(Harmon et al ., 1986 ; Laiho & Prescott, 2004 ).倒木在物理、化学和生物(特别是微生物、昆虫)等作用下进行的分解过程不仅可为分解者(如微生物、无脊椎动物、鸟类等)提供生存环境和必需的营养元素(Sollins, 1982 ), 而且能够释放大量的化学元素到土壤中, 丰富土壤有机质含量和种类, 促进土壤结构和功能的发育(Spears et al ., 2003 ).影响倒木分解的因素有基质质量、分解者活性以及环境因素等(如温度、水分、气体)(Harmon et al ., 1986).而在同一环境下, 不同树种间倒木分解速率也有很大差异, 这与倒木自身的物理和化学特性有关(van Geffen et al ., 2010 ).含水率和木材密度是影响倒木分解的主要物理因素, 因而受到越来越多的关注.倒木含水率是影响倒木呼吸的重要因子, 当它超过一定的阈值后, 就会影响微生物的活动, 从而抑制倒木的分解(Bond-Lamberty et al ., 2002 ).木材分解是一个复杂的过程, 通常根据单位时间内木材质量损失或木材密度的变化, 可以定量地算出其分解速率; 木材密度与微生物侵入倒木进行分解活动的程度有密切关系, 较低密度的木质残体更容易被分解和具有更高的呼吸速率(Johnston & Crossley, 1996 ; Chamber et al ., 2001 ).影响倒木分解的化学因素主要是其易分解成分和难分解有机成分(如木质素等)的含量, 其中氮(N)、磷(P)、木质素、纤维素、C:N、C:P等是常见的衡量倒木质量的指标(Berg et al ., 2000 ), 它们不仅可以表征倒木分解过程中基质的质量变化和养分限制状况(Wassen et al ., 2005 ), 而且可以指示物质循环和能量流动的方向(Zhou et al ., 2007 ). ...
a). 小兴安岭阔叶红松林粗木质残体基础特征. 林业科学,
2010
b). 小兴安岭阔叶红松林粗木质残体空间分布的点格局分析. 生态学报,
2010
Investigating biochemical processes to assess deadwood decay of beech and silver fir in Mediterranean Mountain forests
2013
鼎湖山锥栗粗木质残体的分解和元素动态. 热带亚热带植物学报,
2
2006
... 目前, 我国部分学者对长白山红松(Pinus koraiensis )暗针叶林(杨丽韫和代力民, 2002 )、大兴安岭落叶松(Larix gmelinii )林(辛魏巍等, 2012 )、鼎湖山季风常绿阔叶林(吕明和等, 2006 )以及亚高山针叶林(常晨晖等, 2015 )等森林类型中主要组成树种的倒木在分解过程的物理化学性质变化趋势进行了研究和评述, 而缺乏径级对倒木物理化学性质影响的研究.对不同径级和腐烂等级的倒木进行物理化学性质分析才能准确地量化和预测倒木C储量和养分含量, 同时能够更深入地了解倒木物理性质与化学性质之间的关系.阔叶红松林是我国东北东部山区的地带性顶极植被, 对其倒木物种组成、数量(刘妍妍和金光泽, 2010a )、贮量特征(代力民等, 2000 )以及分布格局(刘妍妍和金光泽, 2010b )曾进行过系统研究, 然而还缺乏倒木分解动态的详细研究.本研究以典型阔叶红松林的建群种红松倒木为研究对象, 探讨其不同径级与不同腐烂等级倒木心材和边材物理化学性质变化特征, 为未来预测倒木C贮量和养分含量以及生产力变化进行合理的评估提供重要的科学依据和基础数据. ...
... 本研究发现, 腐烂等级对倒木含水率和密度产生极显著影响(p < 0.001)(表1 ).随着腐烂等级增加, 倒木含水率升高, 密度下降(图1A 、1B).这与其他地区对倒木含水率和密度的研究结果一致(吕明和等, 2006 ; Seedre et al ., 2013 ; Petrillo et al ., 2015 ).其原因主要是由于随着分解程度加深, 倒木受到淋溶、破碎化以及呼吸作用的影响, 木质变得疏松, 从而导致倒木密度下降; 随着腐烂等级增加, 其木质越疏松, 越容易吸收降水(袁杰和张硕新, 2012 ); 其次, 腐烂程度越高, 倒木与地面接触面积越大, 影响倒木含水率变化的因素增加, 例如土壤的含水率、地表水和微地貌等(Bütler et al ., 2007 ).本研究显示, 径级i-iii倒木密度呈下降趋势, 径级iv略有上升, 但与径级iii差异不显著(图1D ), 这与Sakai等(2012)的研究结果基本一致.倒木密度受许多因素的影响, 例如树种、地理环境、树龄、增长速率以及遗传因素等(Harmon et al ., 1986 ; Repola, 2006 ).Spurr和Hsuing (1954)发现, 从心材到边材, 幼龄树的密度比老龄树的大.本实验所测红松倒木的直径位置为胸径处, 可以间接代表龄级.相比大径级倒木, 小径级的倒木树龄小, 密度相对较大, 因此树龄是造成随着径级增加密度减小的主要原因. ...
Stoichiometric controls on carbon, nitrogen, and phosph- orus dynamics in decomposing litter
1
2010
... C是构成植物体干物质的最主要元素, N和P是生态系统最主要的营养元素.N和P养分的有效性是调节植物残体分解速率和生态系统碳平衡的一个主要因素, 同时, 植物残体中N、P的矿化为土壤提供了重要的无机营养元素.本研究发现, 除心材C含量外, 腐烂等级显著地影响倒木C、N、P的含量(p < 0.05)(表1 ).随着腐烂等级增加, 边材C含量呈上升趋势, 且腐烂等级V显著高于腐烂等级I-III的C含量(图2 ), 这与Sandström等(2007)和Harmon等(2013)对裸子植物倒木C含量的研究结果一致, 而与Lombardi等(2013)和常晨晖等(2015)对倒木C含量的研究结果相反; Bütler等(2007)则发现欧洲云杉倒木随着分解程度加深C含量保持稳定.这些研究表明腐烂等级对倒木C含量的影响因环境条件和树种不同而存在差异.杨方方等(2009)认为, 倒木C含量随腐烂等级增加而增加的原因可能是木质残体分解周期较长, C损失速度与木质体腐解速度不同步, 从而在高腐烂等级C含量升高, 但导致C含量升高的深层原因有待进一步研究.由于分解初期倒木心材和边材N含量比较低, 随着腐烂等级增加, 边材与环境接触面积增大, 水热条件及通气量得到改善, 促进了微生物对边材的入侵及繁殖.同时, 寄生性微生物通过无脊椎动物侵入倒木心材, 心材结构疏松, 有利于微生物的繁殖.随着微生物增多, 一些固氮细菌数量不断增多, 对环境中N固定能力增强, N含量升高.到分解后期, 微生物竞争加剧, 呼吸作用产生的CO2 对微生物的毒害抑制了微生物的繁殖, 使得固氮细菌数量减少(常晨晖等, 2014 ), 固N能力减弱, N含量下降, 从而导致随着腐烂等级增加倒木N含量呈先升高后降低的变化趋势.虽然在分解后期略有下降, 但差异不显著, 这与其他相关研究结果基本一致(Palviainen et al ., 2008 ; 张修玉等, 2009 ; Köster et al ., 2015 ).本研究中, 倒木腐烂等级I和II的P含量相对较少, 腐烂等级III开始逐渐增加, 与杨丽韫和代力民(2002)、Palviainen等(2010)和Lombardi等(2013)的研究结果一致.这一方面可能是由于一些木材腐朽菌的菌丝把营养从土壤转移到倒木中(Clinton et al ., 2009 ), 另一方面由于有机物质的分解导致P的相对富集(Manzoni et al ., 2010 ).本研究结果发现边材的总体平均N含量显著高于心材(图2 ), 与Cowling和Merrill (1966)对裸子植物的研究结果基本一致.纤维素和木质素是倒木基质的主要组成成分, 其降解是维持生态系统碳素平衡的不可缺少的过程(Fioretto et al ., 2005 ).本研究中腐烂等级显著地影响倒木纤维素和木质素的含量(p < 0.001)(表1 ), 随着腐烂等级增加, 纤维素含量呈下降趋势, 木质素含量表现为上升趋势(图2 ), 这与以往对其他地区针叶树的研究结果相似(Ganjegunte et al ., 2004 ; Bütler et al ., 2007 ; 常晨晖等, 2015 ).这是因为纤维素是由纤维二糖单体组成的碳水化合物, 其结构相对简单, 而且有相对较多种类的微生物能够分解, 因此更容易被接触并分解(Austin & Ballaré, 2010 ); 而木质素具有独特的复杂结构, 它被认为是倒木中最难降解的组分.在分解初期随着易分解组分(可溶性C、N、P等)的不断降解, 木质素等难分解组分所占比例有所提高, 随着分解程度加深, 木质素含量升高.另外, 有关研究发现, 在分解过程中, 微生物新陈代谢能形成木质素类似物(Brandt et al ., 2010 ), 从而使倒木中木质素含量显著增加. ...
Deadwood abundance in recently harvested and old Nova Scotia hardwood forests
1
2010
... 近年来, 有关森林倒木物理化学性质的研究引起了广泛的关注.大部分研究结果表明, 随着腐烂等级增加, 倒木C含量呈下降趋势, N、P含量则与之相反(Palviainen et al ., 2008 ; 常晨晖等, 2015 ).很多研究认为物理化学性质的变化还与树种有关.如Köster等(2015)对同一地区主要倒木树种进行C、N含量和密度的测定发现, 欧洲云杉(Picea abies )倒木的C含量随着分解程度加深而增加, 但是欧洲山杨(Populus tremula )倒木的C含量随着分解程度加深呈下降趋势, Alnus glutinosa 倒木的C含量则变化不大; 随着分解程度加深, 不同树种的木材密度均下降, 但欧洲云杉、Alnus glutinosa 以及欧洲赤松(Pinus sylvestris )比其他3种阔叶树种下降程度小.以往对倒木物理化学性质的研究大多集中在不同腐烂等级和树种对倒木物理化学性质的影响, 而径级对倒木物理化学性质影响的研究尚处于初步探索阶段.通常情况下, 小径级倒木(胸径(DBH ) < 10 cm)在森林生态系统中的数量巨大且分解较快, 有利于有机物质矿化到土壤中被土壤吸收和利用(Moroni & Ryan, 2010 ).相对于小径级倒木, 大径级倒木心材比例较大, 由于心材储存了大量能源物质和难分解组分(Schwarze, 2007 ), 可能有较高的C含量和相对较低的养分含量, 但其结构疏松, 易于分解者活动和居住, 随着分解程度加深, 物理化学性质特征更加复杂(常晨晖等, 2015 ).Harmon等(1986)提出倒木中物理化学性质的变化可能与倒木的径级有关.吕明和等(2006)在对鼎湖山锥(Castanopsis chin- ensis )的粗木质残体的分解和元素动态分析中也指出, 在分解过程中直径5-10 cm的倒木中的N、P含量持续升高, 而直径10-20 cm的倒木中的N、P含量呈下降趋势.Sakai等(2012)在对日本扁柏(Chamae- cyparis obtusa )和日本柳杉(Cryptomeria japonica )的倒木密度以及C、N含量测定的研究中发现, 随着径级增加, 倒木密度随之下降, 且径级对处于腐烂等级I和II的两种倒木的密度影响显著.由此可见, 倒木中物理化学性质的变化不仅受到腐烂等级和径级的影响, 而且可能还受两者产生的交互作用的影响. ...
Relative importance of coarse and fine woody debris for the diversity of wood-inhabiting fungi in temperate broadleaf forests
2004
Phosphorus and base cation accumulation and release patterns in decomposing Scots pine, Norway spruce and silver birch stumps
2010
Do decomposing Scots pine, Norway spruce, and silver birch stems retain nitrogen?
2
2008
... 近年来, 有关森林倒木物理化学性质的研究引起了广泛的关注.大部分研究结果表明, 随着腐烂等级增加, 倒木C含量呈下降趋势, N、P含量则与之相反(Palviainen et al ., 2008 ; 常晨晖等, 2015 ).很多研究认为物理化学性质的变化还与树种有关.如Köster等(2015)对同一地区主要倒木树种进行C、N含量和密度的测定发现, 欧洲云杉(Picea abies )倒木的C含量随着分解程度加深而增加, 但是欧洲山杨(Populus tremula )倒木的C含量随着分解程度加深呈下降趋势, Alnus glutinosa 倒木的C含量则变化不大; 随着分解程度加深, 不同树种的木材密度均下降, 但欧洲云杉、Alnus glutinosa 以及欧洲赤松(Pinus sylvestris )比其他3种阔叶树种下降程度小.以往对倒木物理化学性质的研究大多集中在不同腐烂等级和树种对倒木物理化学性质的影响, 而径级对倒木物理化学性质影响的研究尚处于初步探索阶段.通常情况下, 小径级倒木(胸径(DBH ) < 10 cm)在森林生态系统中的数量巨大且分解较快, 有利于有机物质矿化到土壤中被土壤吸收和利用(Moroni & Ryan, 2010 ).相对于小径级倒木, 大径级倒木心材比例较大, 由于心材储存了大量能源物质和难分解组分(Schwarze, 2007 ), 可能有较高的C含量和相对较低的养分含量, 但其结构疏松, 易于分解者活动和居住, 随着分解程度加深, 物理化学性质特征更加复杂(常晨晖等, 2015 ).Harmon等(1986)提出倒木中物理化学性质的变化可能与倒木的径级有关.吕明和等(2006)在对鼎湖山锥(Castanopsis chin- ensis )的粗木质残体的分解和元素动态分析中也指出, 在分解过程中直径5-10 cm的倒木中的N、P含量持续升高, 而直径10-20 cm的倒木中的N、P含量呈下降趋势.Sakai等(2012)在对日本扁柏(Chamae- cyparis obtusa )和日本柳杉(Cryptomeria japonica )的倒木密度以及C、N含量测定的研究中发现, 随着径级增加, 倒木密度随之下降, 且径级对处于腐烂等级I和II的两种倒木的密度影响显著.由此可见, 倒木中物理化学性质的变化不仅受到腐烂等级和径级的影响, 而且可能还受两者产生的交互作用的影响. ...
... C是构成植物体干物质的最主要元素, N和P是生态系统最主要的营养元素.N和P养分的有效性是调节植物残体分解速率和生态系统碳平衡的一个主要因素, 同时, 植物残体中N、P的矿化为土壤提供了重要的无机营养元素.本研究发现, 除心材C含量外, 腐烂等级显著地影响倒木C、N、P的含量(p < 0.05)(表1 ).随着腐烂等级增加, 边材C含量呈上升趋势, 且腐烂等级V显著高于腐烂等级I-III的C含量(图2 ), 这与Sandström等(2007)和Harmon等(2013)对裸子植物倒木C含量的研究结果一致, 而与Lombardi等(2013)和常晨晖等(2015)对倒木C含量的研究结果相反; Bütler等(2007)则发现欧洲云杉倒木随着分解程度加深C含量保持稳定.这些研究表明腐烂等级对倒木C含量的影响因环境条件和树种不同而存在差异.杨方方等(2009)认为, 倒木C含量随腐烂等级增加而增加的原因可能是木质残体分解周期较长, C损失速度与木质体腐解速度不同步, 从而在高腐烂等级C含量升高, 但导致C含量升高的深层原因有待进一步研究.由于分解初期倒木心材和边材N含量比较低, 随着腐烂等级增加, 边材与环境接触面积增大, 水热条件及通气量得到改善, 促进了微生物对边材的入侵及繁殖.同时, 寄生性微生物通过无脊椎动物侵入倒木心材, 心材结构疏松, 有利于微生物的繁殖.随着微生物增多, 一些固氮细菌数量不断增多, 对环境中N固定能力增强, N含量升高.到分解后期, 微生物竞争加剧, 呼吸作用产生的CO2 对微生物的毒害抑制了微生物的繁殖, 使得固氮细菌数量减少(常晨晖等, 2014 ), 固N能力减弱, N含量下降, 从而导致随着腐烂等级增加倒木N含量呈先升高后降低的变化趋势.虽然在分解后期略有下降, 但差异不显著, 这与其他相关研究结果基本一致(Palviainen et al ., 2008 ; 张修玉等, 2009 ; Köster et al ., 2015 ).本研究中, 倒木腐烂等级I和II的P含量相对较少, 腐烂等级III开始逐渐增加, 与杨丽韫和代力民(2002)、Palviainen等(2010)和Lombardi等(2013)的研究结果一致.这一方面可能是由于一些木材腐朽菌的菌丝把营养从土壤转移到倒木中(Clinton et al ., 2009 ), 另一方面由于有机物质的分解导致P的相对富集(Manzoni et al ., 2010 ).本研究结果发现边材的总体平均N含量显著高于心材(图2 ), 与Cowling和Merrill (1966)对裸子植物的研究结果基本一致.纤维素和木质素是倒木基质的主要组成成分, 其降解是维持生态系统碳素平衡的不可缺少的过程(Fioretto et al ., 2005 ).本研究中腐烂等级显著地影响倒木纤维素和木质素的含量(p < 0.001)(表1 ), 随着腐烂等级增加, 纤维素含量呈下降趋势, 木质素含量表现为上升趋势(图2 ), 这与以往对其他地区针叶树的研究结果相似(Ganjegunte et al ., 2004 ; Bütler et al ., 2007 ; 常晨晖等, 2015 ).这是因为纤维素是由纤维二糖单体组成的碳水化合物, 其结构相对简单, 而且有相对较多种类的微生物能够分解, 因此更容易被接触并分解(Austin & Ballaré, 2010 ); 而木质素具有独特的复杂结构, 它被认为是倒木中最难降解的组分.在分解初期随着易分解组分(可溶性C、N、P等)的不断降解, 木质素等难分解组分所占比例有所提高, 随着分解程度加深, 木质素含量升高.另外, 有关研究发现, 在分解过程中, 微生物新陈代谢能形成木质素类似物(Brandt et al ., 2010 ), 从而使倒木中木质素含量显著增加. ...
Decomposition of Norway spruce and European larch coarse woody debris (CWD) in relation to different elevation and exposure in an Alpine setting
1
2015
... 本研究发现, 腐烂等级对倒木含水率和密度产生极显著影响(p < 0.001)(表1 ).随着腐烂等级增加, 倒木含水率升高, 密度下降(图1A 、1B).这与其他地区对倒木含水率和密度的研究结果一致(吕明和等, 2006 ; Seedre et al ., 2013 ; Petrillo et al ., 2015 ).其原因主要是由于随着分解程度加深, 倒木受到淋溶、破碎化以及呼吸作用的影响, 木质变得疏松, 从而导致倒木密度下降; 随着腐烂等级增加, 其木质越疏松, 越容易吸收降水(袁杰和张硕新, 2012 ); 其次, 腐烂程度越高, 倒木与地面接触面积越大, 影响倒木含水率变化的因素增加, 例如土壤的含水率、地表水和微地貌等(Bütler et al ., 2007 ).本研究显示, 径级i-iii倒木密度呈下降趋势, 径级iv略有上升, 但与径级iii差异不显著(图1D ), 这与Sakai等(2012)的研究结果基本一致.倒木密度受许多因素的影响, 例如树种、地理环境、树龄、增长速率以及遗传因素等(Harmon et al ., 1986 ; Repola, 2006 ).Spurr和Hsuing (1954)发现, 从心材到边材, 幼龄树的密度比老龄树的大.本实验所测红松倒木的直径位置为胸径处, 可以间接代表龄级.相比大径级倒木, 小径级的倒木树龄小, 密度相对较大, 因此树龄是造成随着径级增加密度减小的主要原因. ...
Respiration from coarse woody debris as affected by moisture and saprotroph functional diversity in Western Oregon
1
2000
... 倒木的含水率、密度与纤维素含量和木质素含量具有显著的相关性, 这可能与倒木的呼吸作用有关.含水率和密度是影响倒木呼吸的主要物理因素, 倒木的呼吸作用主要是通过微生物的分解作用来进行的(Herrmann & Bauhus, 2013 ).微生物分解对水分的增加非常敏感, 在一定范围内, 较小的水分增加会导致微生物活性的急剧增大(Progar et al ., 2000 ).由于单糖等非结构性碳水化合物易被微生物分解且含量较低, 到分解中后期, 相比木质素, 纤维素由于其结构相对简单, 成为微生物分解的主要物质.随着倒木含水率的增加, 微生物对纤维素的分解加快, 纤维素含量下降, 而木质素在倒木中的含量相对升高, 从而导致含水率与纤维素含量的负相关, 与木质素含量正相关(表4 ).本文中木材密度对倒木纤维素和木质素的影响与含水率相反.密度低, 则微生物易于侵入进行分解活动, 且其持水能力强, 更易促进微生物对纤维素的分解(Chamber et al ., 2001 ).因此密度与木质素、纤维素的关系与含水率相反(表4 ).而倒木中C、N、P含量的变化与倒木含水率和密度之间的相关性实质上是腐烂等级分别对倒木物理化学性质的影响所导致的.C、N、P与含水率均表现为随着腐烂等级增加而升高,密度则下降.因此, 含水率与C、N、P含量呈正相关关系; 密度则与倒木C、N、P含量呈负相关关系. ...
Global patterns of plant leaf N and P in relation to temperature and latitude
2
2004
... 生态化学计量学理论认为大多数情况下C:N > 27或C:P > 186时, 微生物生长受限制(Sinsabaugh et al ., 2009 ).本研究中, 心材和边材在整个分解阶段均易受到N、P限制.Laiho和Prescott (2004)认为初始N:P值也能很好地预测N、P动态, 他们在针叶树种分解过程中养分释放的研究中发现临界N:P值约为20.本研究中, 腐烂等级I-III心材和边材N:P值均大于20 (表2 ), 表现为N相对含量的富集模式, 随着腐烂等级增加, 腐烂等级IV和V边材和心材N:P值均小于20, 表现出养分含量的释放模式.通常C在大多数植物体内含量高且变化较小(Reich & Oleksyn, 2004 ), 本研究也证实了这一点, 不同径级间C含量差异不显著(表3 ).影响C与N、P比值的主要因素是N、P含量的变化(Reich & Oleksyn, 2004 ).N、P含量随着径级先增后减, 而C含量在不同径级中比较稳定, 导致了随着径级增加, C:N、C:P先降后升的趋势(表3 ). ...
... ).影响C与N、P比值的主要因素是N、P含量的变化(Reich & Oleksyn, 2004 ).N、P含量随着径级先增后减, 而C含量在不同径级中比较稳定, 导致了随着径级增加, C:N、C:P先降后升的趋势(表3 ). ...
Models for vertical wood density of Scots pine, Norway spruce and birch stems, and their application to determine average wood density
1
2006
... 本研究发现, 腐烂等级对倒木含水率和密度产生极显著影响(p < 0.001)(表1 ).随着腐烂等级增加, 倒木含水率升高, 密度下降(图1A 、1B).这与其他地区对倒木含水率和密度的研究结果一致(吕明和等, 2006 ; Seedre et al ., 2013 ; Petrillo et al ., 2015 ).其原因主要是由于随着分解程度加深, 倒木受到淋溶、破碎化以及呼吸作用的影响, 木质变得疏松, 从而导致倒木密度下降; 随着腐烂等级增加, 其木质越疏松, 越容易吸收降水(袁杰和张硕新, 2012 ); 其次, 腐烂程度越高, 倒木与地面接触面积越大, 影响倒木含水率变化的因素增加, 例如土壤的含水率、地表水和微地貌等(Bütler et al ., 2007 ).本研究显示, 径级i-iii倒木密度呈下降趋势, 径级iv略有上升, 但与径级iii差异不显著(图1D ), 这与Sakai等(2012)的研究结果基本一致.倒木密度受许多因素的影响, 例如树种、地理环境、树龄、增长速率以及遗传因素等(Harmon et al ., 1986 ; Repola, 2006 ).Spurr和Hsuing (1954)发现, 从心材到边材, 幼龄树的密度比老龄树的大.本实验所测红松倒木的直径位置为胸径处, 可以间接代表龄级.相比大径级倒木, 小径级的倒木树龄小, 密度相对较大, 因此树龄是造成随着径级增加密度减小的主要原因. ...
Wood density and carbon and nitrogen concentrations in deadwood of
2012
Biomass conversion factors (density and carbon concentration) by decay classes for dead wood of
2007
Wood decay under the microscope
1
2007
... 近年来, 有关森林倒木物理化学性质的研究引起了广泛的关注.大部分研究结果表明, 随着腐烂等级增加, 倒木C含量呈下降趋势, N、P含量则与之相反(Palviainen et al ., 2008 ; 常晨晖等, 2015 ).很多研究认为物理化学性质的变化还与树种有关.如Köster等(2015)对同一地区主要倒木树种进行C、N含量和密度的测定发现, 欧洲云杉(Picea abies )倒木的C含量随着分解程度加深而增加, 但是欧洲山杨(Populus tremula )倒木的C含量随着分解程度加深呈下降趋势, Alnus glutinosa 倒木的C含量则变化不大; 随着分解程度加深, 不同树种的木材密度均下降, 但欧洲云杉、Alnus glutinosa 以及欧洲赤松(Pinus sylvestris )比其他3种阔叶树种下降程度小.以往对倒木物理化学性质的研究大多集中在不同腐烂等级和树种对倒木物理化学性质的影响, 而径级对倒木物理化学性质影响的研究尚处于初步探索阶段.通常情况下, 小径级倒木(胸径(DBH ) < 10 cm)在森林生态系统中的数量巨大且分解较快, 有利于有机物质矿化到土壤中被土壤吸收和利用(Moroni & Ryan, 2010 ).相对于小径级倒木, 大径级倒木心材比例较大, 由于心材储存了大量能源物质和难分解组分(Schwarze, 2007 ), 可能有较高的C含量和相对较低的养分含量, 但其结构疏松, 易于分解者活动和居住, 随着分解程度加深, 物理化学性质特征更加复杂(常晨晖等, 2015 ).Harmon等(1986)提出倒木中物理化学性质的变化可能与倒木的径级有关.吕明和等(2006)在对鼎湖山锥(Castanopsis chin- ensis )的粗木质残体的分解和元素动态分析中也指出, 在分解过程中直径5-10 cm的倒木中的N、P含量持续升高, 而直径10-20 cm的倒木中的N、P含量呈下降趋势.Sakai等(2012)在对日本扁柏(Chamae- cyparis obtusa )和日本柳杉(Cryptomeria japonica )的倒木密度以及C、N含量测定的研究中发现, 随着径级增加, 倒木密度随之下降, 且径级对处于腐烂等级I和II的两种倒木的密度影响显著.由此可见, 倒木中物理化学性质的变化不仅受到腐烂等级和径级的影响, 而且可能还受两者产生的交互作用的影响. ...
Deadwood density of five boreal tree species in relation to field-assigned decay class
1
2013
... 本研究发现, 腐烂等级对倒木含水率和密度产生极显著影响(p < 0.001)(表1 ).随着腐烂等级增加, 倒木含水率升高, 密度下降(图1A 、1B).这与其他地区对倒木含水率和密度的研究结果一致(吕明和等, 2006 ; Seedre et al ., 2013 ; Petrillo et al ., 2015 ).其原因主要是由于随着分解程度加深, 倒木受到淋溶、破碎化以及呼吸作用的影响, 木质变得疏松, 从而导致倒木密度下降; 随着腐烂等级增加, 其木质越疏松, 越容易吸收降水(袁杰和张硕新, 2012 ); 其次, 腐烂程度越高, 倒木与地面接触面积越大, 影响倒木含水率变化的因素增加, 例如土壤的含水率、地表水和微地貌等(Bütler et al ., 2007 ).本研究显示, 径级i-iii倒木密度呈下降趋势, 径级iv略有上升, 但与径级iii差异不显著(图1D ), 这与Sakai等(2012)的研究结果基本一致.倒木密度受许多因素的影响, 例如树种、地理环境、树龄、增长速率以及遗传因素等(Harmon et al ., 1986 ; Repola, 2006 ).Spurr和Hsuing (1954)发现, 从心材到边材, 幼龄树的密度比老龄树的大.本实验所测红松倒木的直径位置为胸径处, 可以间接代表龄级.相比大径级倒木, 小径级的倒木树龄小, 密度相对较大, 因此树龄是造成随着径级增加密度减小的主要原因. ...
Dead wood characteristics influ- enceing macrofungi species abundance and diversity in Caspian natural beech (Fagus orientalis Lipsky) forests
1
2015
... 倒木腐烂等级的划分按照闫恩荣等(2005)的粗木质残体(coarse woody debris, CWD) 5级分类系统(I级: 新鲜, 树木死亡不足1年; II级: 开始腐解, 小刀可刺进几mm; III级: 小刀可刺进2 cm; IV级: 小刀可刺进2-5 cm; V级: 小刀可任意刺穿木质体).对于腐烂等级低(I-III)的倒木用胸径尺测量其直径, 对于腐烂等级高(IV和V)的倒木直径进行目测(Sefidi & Etemad, 2015 ). ...
Ecoenzy- matic stoichiometry of microbial organic nutrient acquisition in soil and sediment
1
2009
... 生态化学计量学理论认为大多数情况下C:N > 27或C:P > 186时, 微生物生长受限制(Sinsabaugh et al ., 2009 ).本研究中, 心材和边材在整个分解阶段均易受到N、P限制.Laiho和Prescott (2004)认为初始N:P值也能很好地预测N、P动态, 他们在针叶树种分解过程中养分释放的研究中发现临界N:P值约为20.本研究中, 腐烂等级I-III心材和边材N:P值均大于20 (表2 ), 表现为N相对含量的富集模式, 随着腐烂等级增加, 腐烂等级IV和V边材和心材N:P值均小于20, 表现出养分含量的释放模式.通常C在大多数植物体内含量高且变化较小(Reich & Oleksyn, 2004 ), 本研究也证实了这一点, 不同径级间C含量差异不显著(表3 ).影响C与N、P比值的主要因素是N、P含量的变化(Reich & Oleksyn, 2004 ).N、P含量随着径级先增后减, 而C含量在不同径级中比较稳定, 导致了随着径级增加, C:N、C:P先降后升的趋势(表3 ). ...
Input and decay of coarse woody debris in coniferous stands in western Oregon and Washington
1
1982
... 森林是陆地生态系统的重要组成部分, 其植被碳(C)库约占全球植被C库的80% (Köster et al ., 2015 ).倒木是森林生态系统的重要组成部分, 对于维持生物多样性以及林木的更新具有重要作用; 同时, 倒木也是重要的养分库和C库, 其养分的缓慢释放, 对于提高林地的生产力有重要意义(Harmon et al ., 1986 ; Laiho & Prescott, 2004 ).倒木在物理、化学和生物(特别是微生物、昆虫)等作用下进行的分解过程不仅可为分解者(如微生物、无脊椎动物、鸟类等)提供生存环境和必需的营养元素(Sollins, 1982 ), 而且能够释放大量的化学元素到土壤中, 丰富土壤有机质含量和种类, 促进土壤结构和功能的发育(Spears et al ., 2003 ).影响倒木分解的因素有基质质量、分解者活性以及环境因素等(如温度、水分、气体)(Harmon et al ., 1986).而在同一环境下, 不同树种间倒木分解速率也有很大差异, 这与倒木自身的物理和化学特性有关(van Geffen et al ., 2010 ).含水率和木材密度是影响倒木分解的主要物理因素, 因而受到越来越多的关注.倒木含水率是影响倒木呼吸的重要因子, 当它超过一定的阈值后, 就会影响微生物的活动, 从而抑制倒木的分解(Bond-Lamberty et al ., 2002 ).木材分解是一个复杂的过程, 通常根据单位时间内木材质量损失或木材密度的变化, 可以定量地算出其分解速率; 木材密度与微生物侵入倒木进行分解活动的程度有密切关系, 较低密度的木质残体更容易被分解和具有更高的呼吸速率(Johnston & Crossley, 1996 ; Chamber et al ., 2001 ).影响倒木分解的化学因素主要是其易分解成分和难分解有机成分(如木质素等)的含量, 其中氮(N)、磷(P)、木质素、纤维素、C:N、C:P等是常见的衡量倒木质量的指标(Berg et al ., 2000 ), 它们不仅可以表征倒木分解过程中基质的质量变化和养分限制状况(Wassen et al ., 2005 ), 而且可以指示物质循环和能量流动的方向(Zhou et al ., 2007 ). ...
The influence of decomposing logs on soil biology and nutrient cycling in an old-growth mixed coniferous forest in Oregon, USA
1
2003
... 森林是陆地生态系统的重要组成部分, 其植被碳(C)库约占全球植被C库的80% (Köster et al ., 2015 ).倒木是森林生态系统的重要组成部分, 对于维持生物多样性以及林木的更新具有重要作用; 同时, 倒木也是重要的养分库和C库, 其养分的缓慢释放, 对于提高林地的生产力有重要意义(Harmon et al ., 1986 ; Laiho & Prescott, 2004 ).倒木在物理、化学和生物(特别是微生物、昆虫)等作用下进行的分解过程不仅可为分解者(如微生物、无脊椎动物、鸟类等)提供生存环境和必需的营养元素(Sollins, 1982 ), 而且能够释放大量的化学元素到土壤中, 丰富土壤有机质含量和种类, 促进土壤结构和功能的发育(Spears et al ., 2003 ).影响倒木分解的因素有基质质量、分解者活性以及环境因素等(如温度、水分、气体)(Harmon et al ., 1986).而在同一环境下, 不同树种间倒木分解速率也有很大差异, 这与倒木自身的物理和化学特性有关(van Geffen et al ., 2010 ).含水率和木材密度是影响倒木分解的主要物理因素, 因而受到越来越多的关注.倒木含水率是影响倒木呼吸的重要因子, 当它超过一定的阈值后, 就会影响微生物的活动, 从而抑制倒木的分解(Bond-Lamberty et al ., 2002 ).木材分解是一个复杂的过程, 通常根据单位时间内木材质量损失或木材密度的变化, 可以定量地算出其分解速率; 木材密度与微生物侵入倒木进行分解活动的程度有密切关系, 较低密度的木质残体更容易被分解和具有更高的呼吸速率(Johnston & Crossley, 1996 ; Chamber et al ., 2001 ).影响倒木分解的化学因素主要是其易分解成分和难分解有机成分(如木质素等)的含量, 其中氮(N)、磷(P)、木质素、纤维素、C:N、C:P等是常见的衡量倒木质量的指标(Berg et al ., 2000 ), 它们不仅可以表征倒木分解过程中基质的质量变化和养分限制状况(Wassen et al ., 2005 ), 而且可以指示物质循环和能量流动的方向(Zhou et al ., 2007 ). ...
Growth rate and specific gravity in conifers
1954
Phellinus nigrolimitatus —A wood-decomposing fungus highly influenced by forestry
2004
The trait contribution to wood decomposition rates of 15 neotropical tree species
1
2010
... 森林是陆地生态系统的重要组成部分, 其植被碳(C)库约占全球植被C库的80% (Köster et al ., 2015 ).倒木是森林生态系统的重要组成部分, 对于维持生物多样性以及林木的更新具有重要作用; 同时, 倒木也是重要的养分库和C库, 其养分的缓慢释放, 对于提高林地的生产力有重要意义(Harmon et al ., 1986 ; Laiho & Prescott, 2004 ).倒木在物理、化学和生物(特别是微生物、昆虫)等作用下进行的分解过程不仅可为分解者(如微生物、无脊椎动物、鸟类等)提供生存环境和必需的营养元素(Sollins, 1982 ), 而且能够释放大量的化学元素到土壤中, 丰富土壤有机质含量和种类, 促进土壤结构和功能的发育(Spears et al ., 2003 ).影响倒木分解的因素有基质质量、分解者活性以及环境因素等(如温度、水分、气体)(Harmon et al ., 1986).而在同一环境下, 不同树种间倒木分解速率也有很大差异, 这与倒木自身的物理和化学特性有关(van Geffen et al ., 2010 ).含水率和木材密度是影响倒木分解的主要物理因素, 因而受到越来越多的关注.倒木含水率是影响倒木呼吸的重要因子, 当它超过一定的阈值后, 就会影响微生物的活动, 从而抑制倒木的分解(Bond-Lamberty et al ., 2002 ).木材分解是一个复杂的过程, 通常根据单位时间内木材质量损失或木材密度的变化, 可以定量地算出其分解速率; 木材密度与微生物侵入倒木进行分解活动的程度有密切关系, 较低密度的木质残体更容易被分解和具有更高的呼吸速率(Johnston & Crossley, 1996 ; Chamber et al ., 2001 ).影响倒木分解的化学因素主要是其易分解成分和难分解有机成分(如木质素等)的含量, 其中氮(N)、磷(P)、木质素、纤维素、C:N、C:P等是常见的衡量倒木质量的指标(Berg et al ., 2000 ), 它们不仅可以表征倒木分解过程中基质的质量变化和养分限制状况(Wassen et al ., 2005 ), 而且可以指示物质循环和能量流动的方向(Zhou et al ., 2007 ). ...
Endangered plants persist under phosphorus limitation
1
2005
... 森林是陆地生态系统的重要组成部分, 其植被碳(C)库约占全球植被C库的80% (Köster et al ., 2015 ).倒木是森林生态系统的重要组成部分, 对于维持生物多样性以及林木的更新具有重要作用; 同时, 倒木也是重要的养分库和C库, 其养分的缓慢释放, 对于提高林地的生产力有重要意义(Harmon et al ., 1986 ; Laiho & Prescott, 2004 ).倒木在物理、化学和生物(特别是微生物、昆虫)等作用下进行的分解过程不仅可为分解者(如微生物、无脊椎动物、鸟类等)提供生存环境和必需的营养元素(Sollins, 1982 ), 而且能够释放大量的化学元素到土壤中, 丰富土壤有机质含量和种类, 促进土壤结构和功能的发育(Spears et al ., 2003 ).影响倒木分解的因素有基质质量、分解者活性以及环境因素等(如温度、水分、气体)(Harmon et al ., 1986).而在同一环境下, 不同树种间倒木分解速率也有很大差异, 这与倒木自身的物理和化学特性有关(van Geffen et al ., 2010 ).含水率和木材密度是影响倒木分解的主要物理因素, 因而受到越来越多的关注.倒木含水率是影响倒木呼吸的重要因子, 当它超过一定的阈值后, 就会影响微生物的活动, 从而抑制倒木的分解(Bond-Lamberty et al ., 2002 ).木材分解是一个复杂的过程, 通常根据单位时间内木材质量损失或木材密度的变化, 可以定量地算出其分解速率; 木材密度与微生物侵入倒木进行分解活动的程度有密切关系, 较低密度的木质残体更容易被分解和具有更高的呼吸速率(Johnston & Crossley, 1996 ; Chamber et al ., 2001 ).影响倒木分解的化学因素主要是其易分解成分和难分解有机成分(如木质素等)的含量, 其中氮(N)、磷(P)、木质素、纤维素、C:N、C:P等是常见的衡量倒木质量的指标(Berg et al ., 2000 ), 它们不仅可以表征倒木分解过程中基质的质量变化和养分限制状况(Wassen et al ., 2005 ), 而且可以指示物质循环和能量流动的方向(Zhou et al ., 2007 ). ...
大兴安岭兴安落叶松原始林倒木养分含量研究
1
2012
... 目前, 我国部分学者对长白山红松(Pinus koraiensis )暗针叶林(杨丽韫和代力民, 2002 )、大兴安岭落叶松(Larix gmelinii )林(辛魏巍等, 2012 )、鼎湖山季风常绿阔叶林(吕明和等, 2006 )以及亚高山针叶林(常晨晖等, 2015 )等森林类型中主要组成树种的倒木在分解过程的物理化学性质变化趋势进行了研究和评述, 而缺乏径级对倒木物理化学性质影响的研究.对不同径级和腐烂等级的倒木进行物理化学性质分析才能准确地量化和预测倒木C储量和养分含量, 同时能够更深入地了解倒木物理性质与化学性质之间的关系.阔叶红松林是我国东北东部山区的地带性顶极植被, 对其倒木物种组成、数量(刘妍妍和金光泽, 2010a )、贮量特征(代力民等, 2000 )以及分布格局(刘妍妍和金光泽, 2010b )曾进行过系统研究, 然而还缺乏倒木分解动态的详细研究.本研究以典型阔叶红松林的建群种红松倒木为研究对象, 探讨其不同径级与不同腐烂等级倒木心材和边材物理化学性质变化特征, 为未来预测倒木C贮量和养分含量以及生产力变化进行合理的评估提供重要的科学依据和基础数据. ...
森林粗死木质残体的概念及其分类. 生态学报,
1
2005
... 本研究在凉水自然保护区9 hm2 (300 m × 300 m)的阔叶红松林永久性固定样地上进行(Jin et al ., 2006 ), 将样地设成900个10 m × 10 m的小样方, 采用相邻网格法进行调查.对样地内所有胸径≥2 cm的木质残体进行调查, 确定木质残体的存在形式(干基折断、干中折断、枯立木、拔根倒、根桩和树段); 测量立木形式的胸径, 对倒伏的木质残体, 测量其中央直径和胸径; 采用Vertex III超声波测高仪(Haglöf Sweden AB, Långsele, Sweden)进行长度和高度的测定, 识别树种, 确定坐标位置, 并钉铝制树牌进行编号.采用5级分类系统进行腐烂等级的划分(闫恩荣等, 2005 ). ...
鼎湖山木荷粗死木质残体的分解研究. 山地学报,
2009
长白山北坡苔藓红松暗针叶林倒木分解及其养分含量. 生态学报,
1
2002
... 目前, 我国部分学者对长白山红松(Pinus koraiensis )暗针叶林(杨丽韫和代力民, 2002 )、大兴安岭落叶松(Larix gmelinii )林(辛魏巍等, 2012 )、鼎湖山季风常绿阔叶林(吕明和等, 2006 )以及亚高山针叶林(常晨晖等, 2015 )等森林类型中主要组成树种的倒木在分解过程的物理化学性质变化趋势进行了研究和评述, 而缺乏径级对倒木物理化学性质影响的研究.对不同径级和腐烂等级的倒木进行物理化学性质分析才能准确地量化和预测倒木C储量和养分含量, 同时能够更深入地了解倒木物理性质与化学性质之间的关系.阔叶红松林是我国东北东部山区的地带性顶极植被, 对其倒木物种组成、数量(刘妍妍和金光泽, 2010a )、贮量特征(代力民等, 2000 )以及分布格局(刘妍妍和金光泽, 2010b )曾进行过系统研究, 然而还缺乏倒木分解动态的详细研究.本研究以典型阔叶红松林的建群种红松倒木为研究对象, 探讨其不同径级与不同腐烂等级倒木心材和边材物理化学性质变化特征, 为未来预测倒木C贮量和养分含量以及生产力变化进行合理的评估提供重要的科学依据和基础数据. ...
秦岭火地塘天然次生油松林倒木密度与含水量变化特征研究. 中南林业科技大学学报,
1
2012
... 本研究发现, 腐烂等级对倒木含水率和密度产生极显著影响(p < 0.001)(表1 ).随着腐烂等级增加, 倒木含水率升高, 密度下降(图1A 、1B).这与其他地区对倒木含水率和密度的研究结果一致(吕明和等, 2006 ; Seedre et al ., 2013 ; Petrillo et al ., 2015 ).其原因主要是由于随着分解程度加深, 倒木受到淋溶、破碎化以及呼吸作用的影响, 木质变得疏松, 从而导致倒木密度下降; 随着腐烂等级增加, 其木质越疏松, 越容易吸收降水(袁杰和张硕新, 2012 ); 其次, 腐烂程度越高, 倒木与地面接触面积越大, 影响倒木含水率变化的因素增加, 例如土壤的含水率、地表水和微地貌等(Bütler et al ., 2007 ).本研究显示, 径级i-iii倒木密度呈下降趋势, 径级iv略有上升, 但与径级iii差异不显著(图1D ), 这与Sakai等(2012)的研究结果基本一致.倒木密度受许多因素的影响, 例如树种、地理环境、树龄、增长速率以及遗传因素等(Harmon et al ., 1986 ; Repola, 2006 ).Spurr和Hsuing (1954)发现, 从心材到边材, 幼龄树的密度比老龄树的大.本实验所测红松倒木的直径位置为胸径处, 可以间接代表龄级.相比大径级倒木, 小径级的倒木树龄小, 密度相对较大, 因此树龄是造成随着径级增加密度减小的主要原因. ...
三种森林粗死木质残体 (CWD)的储量与分解特征. 生态学报,
1
2009
... C是构成植物体干物质的最主要元素, N和P是生态系统最主要的营养元素.N和P养分的有效性是调节植物残体分解速率和生态系统碳平衡的一个主要因素, 同时, 植物残体中N、P的矿化为土壤提供了重要的无机营养元素.本研究发现, 除心材C含量外, 腐烂等级显著地影响倒木C、N、P的含量(p < 0.05)(表1 ).随着腐烂等级增加, 边材C含量呈上升趋势, 且腐烂等级V显著高于腐烂等级I-III的C含量(图2 ), 这与Sandström等(2007)和Harmon等(2013)对裸子植物倒木C含量的研究结果一致, 而与Lombardi等(2013)和常晨晖等(2015)对倒木C含量的研究结果相反; Bütler等(2007)则发现欧洲云杉倒木随着分解程度加深C含量保持稳定.这些研究表明腐烂等级对倒木C含量的影响因环境条件和树种不同而存在差异.杨方方等(2009)认为, 倒木C含量随腐烂等级增加而增加的原因可能是木质残体分解周期较长, C损失速度与木质体腐解速度不同步, 从而在高腐烂等级C含量升高, 但导致C含量升高的深层原因有待进一步研究.由于分解初期倒木心材和边材N含量比较低, 随着腐烂等级增加, 边材与环境接触面积增大, 水热条件及通气量得到改善, 促进了微生物对边材的入侵及繁殖.同时, 寄生性微生物通过无脊椎动物侵入倒木心材, 心材结构疏松, 有利于微生物的繁殖.随着微生物增多, 一些固氮细菌数量不断增多, 对环境中N固定能力增强, N含量升高.到分解后期, 微生物竞争加剧, 呼吸作用产生的CO2 对微生物的毒害抑制了微生物的繁殖, 使得固氮细菌数量减少(常晨晖等, 2014 ), 固N能力减弱, N含量下降, 从而导致随着腐烂等级增加倒木N含量呈先升高后降低的变化趋势.虽然在分解后期略有下降, 但差异不显著, 这与其他相关研究结果基本一致(Palviainen et al ., 2008 ; 张修玉等, 2009 ; Köster et al ., 2015 ).本研究中, 倒木腐烂等级I和II的P含量相对较少, 腐烂等级III开始逐渐增加, 与杨丽韫和代力民(2002)、Palviainen等(2010)和Lombardi等(2013)的研究结果一致.这一方面可能是由于一些木材腐朽菌的菌丝把营养从土壤转移到倒木中(Clinton et al ., 2009 ), 另一方面由于有机物质的分解导致P的相对富集(Manzoni et al ., 2010 ).本研究结果发现边材的总体平均N含量显著高于心材(图2 ), 与Cowling和Merrill (1966)对裸子植物的研究结果基本一致.纤维素和木质素是倒木基质的主要组成成分, 其降解是维持生态系统碳素平衡的不可缺少的过程(Fioretto et al ., 2005 ).本研究中腐烂等级显著地影响倒木纤维素和木质素的含量(p < 0.001)(表1 ), 随着腐烂等级增加, 纤维素含量呈下降趋势, 木质素含量表现为上升趋势(图2 ), 这与以往对其他地区针叶树的研究结果相似(Ganjegunte et al ., 2004 ; Bütler et al ., 2007 ; 常晨晖等, 2015 ).这是因为纤维素是由纤维二糖单体组成的碳水化合物, 其结构相对简单, 而且有相对较多种类的微生物能够分解, 因此更容易被接触并分解(Austin & Ballaré, 2010 ); 而木质素具有独特的复杂结构, 它被认为是倒木中最难降解的组分.在分解初期随着易分解组分(可溶性C、N、P等)的不断降解, 木质素等难分解组分所占比例有所提高, 随着分解程度加深, 木质素含量升高.另外, 有关研究发现, 在分解过程中, 微生物新陈代谢能形成木质素类似物(Brandt et al ., 2010 ), 从而使倒木中木质素含量显著增加. ...
Review on the decomposition and influence factors of coarse woody debris in forest ecosystem
1
2007
... 森林是陆地生态系统的重要组成部分, 其植被碳(C)库约占全球植被C库的80% (Köster et al ., 2015 ).倒木是森林生态系统的重要组成部分, 对于维持生物多样性以及林木的更新具有重要作用; 同时, 倒木也是重要的养分库和C库, 其养分的缓慢释放, 对于提高林地的生产力有重要意义(Harmon et al ., 1986 ; Laiho & Prescott, 2004 ).倒木在物理、化学和生物(特别是微生物、昆虫)等作用下进行的分解过程不仅可为分解者(如微生物、无脊椎动物、鸟类等)提供生存环境和必需的营养元素(Sollins, 1982 ), 而且能够释放大量的化学元素到土壤中, 丰富土壤有机质含量和种类, 促进土壤结构和功能的发育(Spears et al ., 2003 ).影响倒木分解的因素有基质质量、分解者活性以及环境因素等(如温度、水分、气体)(Harmon et al ., 1986).而在同一环境下, 不同树种间倒木分解速率也有很大差异, 这与倒木自身的物理和化学特性有关(van Geffen et al ., 2010 ).含水率和木材密度是影响倒木分解的主要物理因素, 因而受到越来越多的关注.倒木含水率是影响倒木呼吸的重要因子, 当它超过一定的阈值后, 就会影响微生物的活动, 从而抑制倒木的分解(Bond-Lamberty et al ., 2002 ).木材分解是一个复杂的过程, 通常根据单位时间内木材质量损失或木材密度的变化, 可以定量地算出其分解速率; 木材密度与微生物侵入倒木进行分解活动的程度有密切关系, 较低密度的木质残体更容易被分解和具有更高的呼吸速率(Johnston & Crossley, 1996 ; Chamber et al ., 2001 ).影响倒木分解的化学因素主要是其易分解成分和难分解有机成分(如木质素等)的含量, 其中氮(N)、磷(P)、木质素、纤维素、C:N、C:P等是常见的衡量倒木质量的指标(Berg et al ., 2000 ), 它们不仅可以表征倒木分解过程中基质的质量变化和养分限制状况(Wassen et al ., 2005 ), 而且可以指示物质循环和能量流动的方向(Zhou et al ., 2007 ). ...


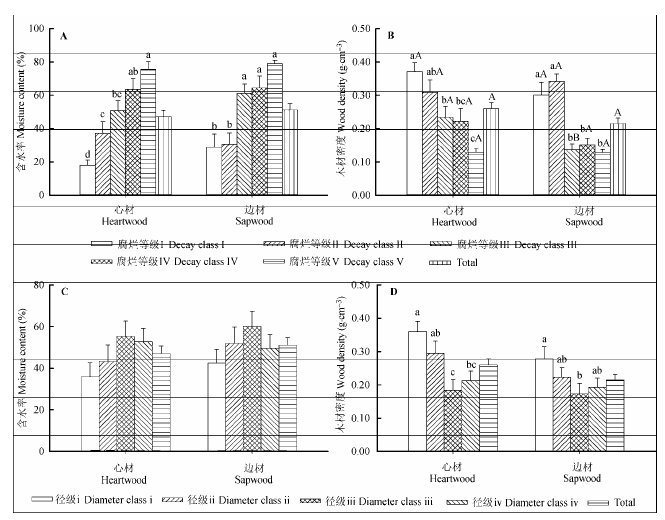
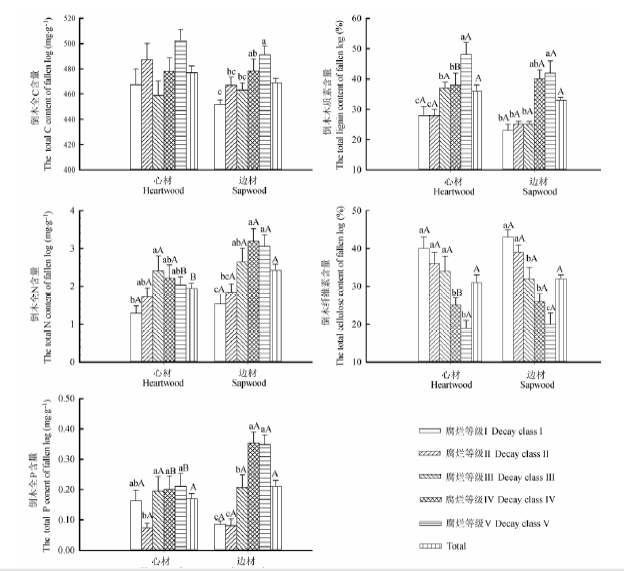
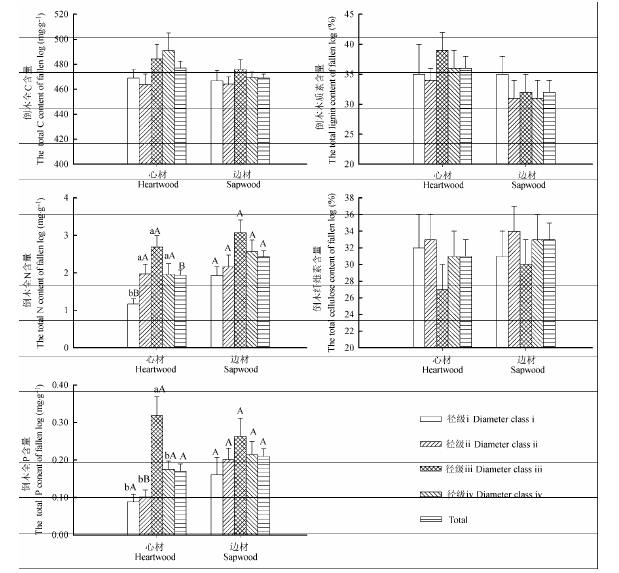
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}