作为植物碳水耦合权衡的重要器官, 叶片在进化过程中对环境的变化较敏感且可塑性较大(Scoffoni et al ., 2015 ), 从叶片形态解剖结构了解植物生态策略在异质生境中的变化对植物的结构和生理变化具有重要意义(Maricle et al ., 2009 )。叶脉是影响叶片水分供应和利用的重要结构, 在一定意义上反映了叶片资源流通能力(Sack & Frole, 2006 ; Sack & Scoffoni, 2013 )。叶脉密度作为叶脉网络功能性状的重要指标(Brodribb et al ., 2010 ; Blonder et al ., 2011 ), 可反映叶脉系统的水分、养分和光合产物等物质运输的效率和安全性以及抵抗外界干扰的能力, 是理解植物环境可塑性机制的重要途径(李乐等, 2013 )。气孔是植物叶片与外界进行水气交换的唯一可调节通道, 通过保卫细胞对环境和内源信号的感知, 与植物的光合作用、蒸腾作用等生理生态功能密切相关(杨惠敏和王根轩, 2001 )。叶脉与气孔是叶片水分供给和散失的重要因素, 两者的大小和数量的配置直接影响着叶片的水分利用效率, 进而影响叶片结构及生理功能的调节机制(Fiorin et al ., 2016 )。当植株处于高光强条件下时, 相对湿度下降, 叶片受到的水分胁迫增加, 叶片在获取光合收益的同时也需兼顾内部水分动态平衡(Fiorin et al ., 2016 ), 此时, 植株往往通过调整净光合速率与蒸腾速率来应对不利环境, 较高的蒸腾速率要求叶片发育较多的叶脉以提供供水系统, 在叶片的蒸腾作用中通过控制气孔密度和气孔大小决定进入大气的水分和CO2 , 降低植物的呼吸和蒸腾成本。叶脉密度与气孔性状共同作用体现了气孔蒸腾对水分的需求和叶脉系统对水分的供给相互匹配与权衡(Brodribb & Jordan, 2011 )。坡向作为重要的地形因素, 控制了太阳辐射和降水的空间再分配(Cartón et al ., 2004 ), 由坡向引起的土壤水分、光辐射、光照时长和大气温度等环境因子形成的生境梯度变化影响着群落组成(Bazzaz et al ., 1987 ; 韦莉莉等, 2005 ), 致使群落环境及其密度制约下的种群内植株的生态位存在差异, 群落内部郁闭度和干扰程度不同, 所接受的光合有效辐射也不同, 从而影响植物的空间分布格局及光照、温度等环境(党晶晶等, 2014 ; 杜晶等, 2016 ), 进而影响植物的形态。为了提高光合能力并保证水分供需间的平衡, 植株叶片会适当调整叶脉密度与气孔密度、气孔大小的生物量分配, 补充蒸腾带来的大量水分和能量的消耗(Hetherington & Woodward, 2003 ), 同时也使叶片中单位碳对光合作用的投资效益最大(Brodribb & Jordan, 2011 ), 从而提高生存适合度和竞争能力。因此, 研究微地形影响下叶脉密度与气孔性状的关系, 有助于探讨植物叶片内部水分供应与散失之间的平衡关系, 对植物生理生态功能的维持具有指导意义。
刺槐(Robinia pseudoacacia )是温带落叶乔木, 是我国华北和西北地区绿化荒山和防风固沙的主要植物, 具有固氮能力强和碳储存量大的优势, 作为黄土丘陵区的主要人工造林树种, 其根系发达、生长迅速、成活率高和环境适应性强等特点在保持水土和生态防护中发挥着重要作用。近年来, 关于刺槐在干旱半干旱地区的种植范围和生态适应性等问题引起了学界的广泛关注(于占辉等, 2009 ; 李军等, 2010 ; 申家朋和张文辉, 2014 )。学术界对气孔性状之间的关系(杨惠敏和王根轩, 2001 ; 王曙光等, 2013 )、气孔与环境因子的关系(左应梅等, 2011 ; 王娜等, 2016 )、气孔性状对水分利用的影响(高洁等, 2004 ; 杨利民等, 2007 )、叶脉网络功能性状与叶片光合作用(Brodribb et al ., 2010 ; Sack et al ., 2012 )、水分利用能力(Zhang et al ., 2012 ; Sack & Scoffoni, 2013 )等问题进行了研究, 阐述了植物叶片在适应特定生境中各种性状的塑造, 而将叶脉与气孔性状结合起来, 探讨二者在异质性生境中叶片内部水分传输及水分平衡适应策略的分析和认识目前还比较薄弱。鉴于此, 本文以兰州北山的人工刺槐林为研究对象, 试图揭示以下两个问题: (1)不同坡向上刺槐叶脉密度与气孔密度、气孔大小之间存在何种关系?(2)这种关系的形成原因有哪些?理解异质生境中植物叶片的物理构建方式, 可为提高半干旱人工林资源的利用率和绿化覆盖率提供理论依据。
1 研究地区概况和研究方法
1.1 研究区概况
试验区位于兰州市北山九州台西北师范大学绿化基地(36.17°-36.23° N, 103.21°-103.25° E), 区内海拔1536-1914 m, 温带大陆性季风气候显著, 年平均气温5-9 ℃, ≥10 ℃的活动积温3385.4 ℃, 年日照时间为2607.6 h, 年太阳总辐射529.6-614.0 kJ·cm-2 , 年降水量327.7 mm, 且主要集中在6-9月, 约占全年降水量的60%, 相反, 年蒸发量很大, 达1650 mm, 为降水量的5倍多(史元春等, 2015 )。土壤为黄土母质上发育起来的灰钙土。供试的刺槐林建植于2002年, 栽培的刺槐为同一品种的同龄幼苗, 平均造林密度为2500株·hm-2 , 每年灌溉4-6次, 灌溉量约800 m3 ·hm-2 。植被群落以刺槐和侧柏(Platycladus orientalis )人工林为主, 主要林下植物有红砂(Reaumuria songarica )、枸杞(Lycium chinense )、碱蓬(Suaeda glauca )、白茎盐生草(Halogeton arachnoideus )、柽柳(Tamarix chinensis )、柠条锦鸡儿(Caragana korshinskii )等。
1.2 实验设计
实验设计于2015年8-9月的刺槐生长旺季完成, 在地图上将研究区划分为4个坡向, 在多次现场踏勘的基础上, 各坡向沿垂直海拔梯度(1550-1750 m)间隔50 m布置长180 m、宽10 m的水平样带1条, 总计20条水平样带, 每条水平样带上间隔5 m设置12个10 m × 10 m的样方, 用GPS测量每个样点的经纬度和海拔。针对各样点内胸径≥1 cm的每个个体进行群落学特征调查, 记录其种名、株高、胸径、冠幅等指标, 并测定各样地群落的平均高度和郁闭度。
对土壤含水量进行采样, 每个样点用土钻(直径4 cm)在0-30 cm土层范围分3层间隔10 cm取土样, 重复3次, 同时用土钻(直径4 cm)在每个样点取0-30 cm混合土样, 重复3次, 所有样品均剔除明显的植物根段和枯落物等杂质, 装入编号的铝盒中。样品带回实验室后, 称量土壤样品质量, 然后在105 ℃的烘箱内烘12 h以上至恒质量, 并计算出各层及0-30 cm土层土壤质量含水量。
1.3 植物采样和测量
1.3.1 植物采样
对刺槐进行采样, 对于株高2 m以上的刺槐, 用高枝剪截取树冠中部外层不同方位的当年生小枝4个; 对于株高2 m以下的刺槐, 直接从冠层中部外层4个方位剪取4个无明显破损叶片的小枝, 在每个枝条上取4片生长良好的叶片做好标记进行以下步骤(所有工作借助人字梯完成)。
(1)植物光合参数的测定: 选择晴朗天气的9:00-12:00进行气体交换参数的测定。光合测定使用GFS-3000便携式光合测量系统(Heinz Walz GmbH, Bavaria, Germany), 测量过程中使用人工红蓝光源, 光合有效辐射(PAR )为1200 μmol·m-2 ·s-1 , CO2 浓度约为340 μmol·mol-1 , 相对湿度(RH )保持在40%-50%, 流速设定为750 μmol·s-1 , 叶室面积为8 cm2 , 对做好标记的每个叶片记录5组数据用于统计, 分析分别测定的叶片净光合速率(P n )、蒸腾速率(T r )等参数(任青吉等, 2015 )。对于不能充满叶室的叶片, 则保存于湿润的封口袋中, 带回实验室, 用便携式激光叶面积仪(CI-202, Walz, Camas, USA)测定叶面积, 进而计算P n 和T r 。
(2) PAR 的测定: 选择晴朗无云的天气, 用手持光量子计(3415F, Walz, Plainfield, USA)于9:00- 11:00对4个坡向内的刺槐进行PAR 测定, 测定位置分别在每个植株上选取的4个枝条向外5 cm处。
1.3.2 叶片性状测量
将样品带回实验室后, 摘除小枝上的叶片, 小枝用于其他分析。
(1)气孔密度和气孔大小的测定: 采用印记法, 将采集的叶片用纸巾擦净后, 选取叶片背面中部采集气孔样品(1 cm × 1 cm), 用透明指甲油涂抹在样品上, 待晾干后取出印记叶片放在载玻片上, 置于Motic SFC-18生物显微镜下观察并拍照, 每个叶片拍摄10个视野, 放大100倍观察气孔密度, 在400×放大倍数下观察气孔长度, 用图形软件ImageJ对含有气孔的照片进行编辑处理, 统计样品内的气孔个数。由于气孔的张开程度随时在变, 选用保卫细胞的长度代表气孔的最大张开程度, 即气孔的大小, 而不是测定气孔的实际张开程度(Xu & Zhou, 2008 )。计算气孔密度: 气孔密度=气孔数/面积。
(2)叶脉密度的测定: 在测完光合参数的植株上采集已标记的叶片, 用含有37%甲醛溶液、50%乙醇和13%冰醋酸溶液的福尔马林-乙酸溶液固定, 然后用含有5% NaOH的乙醇进行化学清理, 用0.1%番红-固绿染色液进行双重染色, 用蒸馏水清洗3遍, 在数码显微镜(Pro 2009a, Regent Instruments, Quebec, Canada)下观察、扫描并拍照(Berlyn & Miksche, 1976 ; 徐婷等, 2016 )。每个叶片拍摄3个视野(叶片两端及中间部位, 包含中脉), 利用CAD软件描绘叶脉长度, 计算所选图像叶脉密度的平均值, 叶脉密度=叶脉长度/叶面积。
1.4 空间数据
对研究区1:50000地形图利用ArcGIS进行数字化分析, 得到栅格大小为25 m的数字高程模型(digital elevation model, DEM)。然后在DEM基础上利用Spatial Analyst模块中的Surface Analysis命令提取坡向, 其中坡向采用3次有限差分方法计算得到(Skidmore, 1989 ), 表示地表面上一点的切平面的法线矢量n 在水平面的投影n xoy 与过该点的正北方向的夹角, 最后利用提取出的坡向指标建立每块样地的数据查找表, 其中坡向组别以国际划分法(陈瑶等, 2006 )确定(圆周角属于315°-360°和0-45°定为正北坡向)。
1.5 数据分析
将坡向划分为北坡(315°-360°和0°-45°)、东坡(45°-135°)、西坡(225°-315°)和南坡(135°-225°), 数据分析前将每个样点刺槐叶片的气孔密度、气孔大小和叶脉密度的平均值进行对数(以10为底)转换后使之符合正态分布。对叶脉密度与气孔性状关系的研究, 采用y = bxa , 线性转换成log(y ) = log(b ) + a log(x ), 式中x 和y 是两个特征参数, b 代表性状关系的截距, a 表示相关性的斜率, 即异速生长参数或相对生长的指数, 当a = 1时, 表示两者是等速关系; 当a > 1时, 表示y 的增加程度大于x 的增加程度; 当a < 1时, 表示y 的增加程度小于x 的增加程度。数据分析主要采用标准化主轴估计(SMA)的方法(Warton et al ., 2006 ), 由软件(S)MATR Version 2.0 (Falster et al ., 2012 )计算完成。每一个回归斜率的置信区间根据Pitman (1939)的方法计算, 并采用Warton和Weber (2002)的方法对每个密度的斜率进行异质性测试。不同坡向间植物功能性状平均值的差异比较采用单因素方差分析(α = 0.05), 实验所用数据采用SPSS 16.0软件进行处理分析。
2 结果和分析
2.1 不同坡向上刺槐林样地群落和土壤含水量特征
如表1 所示, 随着坡向由南坡向东坡、西坡和北坡转变, 刺槐林样地群落的郁闭度、高度和土壤含水量均呈增大趋势。从南坡到北坡, 群落郁闭度增加了29.34%, 植株高度增加了32.09%, 土壤含水量增加了28.40%, 在东坡和西坡之间无显著差异(p > 0.05), 表明植株更适合在北坡生长。
2.2 不同坡向上刺槐叶片性状和光合特性
如表2 所示, 随着坡向的转变, 刺槐叶脉密度、气孔密度、光合有效辐射、净光合速率和蒸腾速率呈减小趋势, 从南坡到北坡分别减少了23.36%、9.8%、23.82%、30.29%和33.05%; 刺槐的气孔大小与叶面积则呈增大趋势, 从南坡到北坡分别增加了22.18%和46.91%, 这些性状值在东坡和西坡之间无显著差异(p > 0.05)。
2.3 不同坡向上刺槐叶脉密度与气孔密度的关系
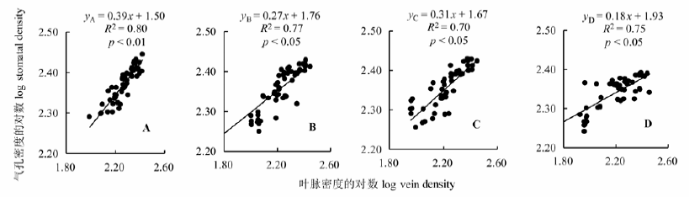
如图1 所示, 在4个坡向上, 刺槐叶脉密度和气孔密度均呈显著正相关关系(p < 0.05), 但在南坡存在极显著正相关关系(p < 0.01), 刺槐叶脉密度与气孔密度在北坡、东坡和西坡呈显著正相关关系(p < 0.05)。通过SMA分析得出, 叶脉密度与气孔密度显著的异速生长关系, 表明在各个坡向上, 气孔密度随叶脉密度的增加而增加, 气孔密度的生长速率显著小于叶脉密度的增长速度, 且在不同坡向生境两者呈协同变异关系。
图1 不同坡向上刺槐叶脉密度与气孔密度的关系。A, 南坡。B, 东坡。C, 西坡。D, 北坡。y 和x 指两个特征参数: 气孔密度对数和叶脉密度对数, y A = 0.39x + 1.50代表南坡气孔密度与叶脉密度对数间的线性关系, 以此类推。
Fig. 1 Relationship between vein density and stomatal density of Robinia pseudoacacia in different aspects. A, South slope. B, East slope. C, West slope. D, North slope. y and x is two parameter: log stomatal density and log vein density, y A = 0.39x + 1.50 represents the logarithms of the linear relationship between stomatal density and vein density, the rest can be done in the same manner.
2.4 不同坡向上刺槐叶脉密度与气孔大小的关系
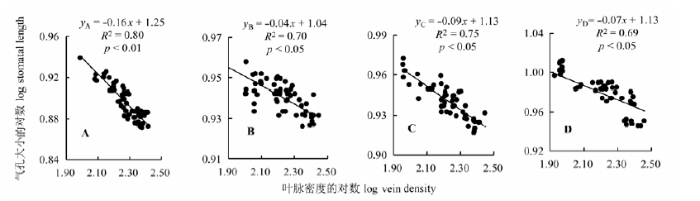
如图2 所示, 在4个坡向上刺槐叶脉密度和气孔大小均呈显著负相关关系(p < 0.05), 但在南坡呈现极显著相关关系(p < 0.01), 刺槐叶脉密度与气孔大小在北坡、东坡和西坡呈显著负相关关系(p < 0.05)。通过SMA分析得出, 叶脉密度与气孔大小呈显著的负的异速生长关系, 表明在各个坡向上随着叶脉密度的降低, 气孔逐渐增大, 在不同坡向生境中有不同的权衡关系。
图2 不同坡向上刺槐叶脉密度与气孔大小的关系。A, 南坡。B, 东坡。C, 西坡。D, 北坡。y 和x 指两个特征参数: 气孔大小对数和叶脉密度对数, y A = -0.16x + 1.25代表南坡气孔大小与叶脉密度对数间的线性关系, 以此类推。
Fig. 2 Relationship between vein density and stomatal length of Robinia pseudoacacia in different aspects. A, South slope. B, East slope. C, West slope. D, North slope. y and x is two parameter: log stomatal length and log vein density, y A = -0.16x + 1.25 represents the logarithms of the linear relationship between stomatal length and vein density, the rest can be done in the same manner.
3 讨论
植物的光合作用是生态系统生产力形成与演化的基础, 叶片作为光合产物形成的主要器官, 既是整株植物的水力瓶颈, 又是植株应对水力失调的重要枢纽, 在很大程度上影响着植物水分和养分吸收、运移和储存功能(金鹰和王传宽, 2015 )。在长期的环境选择进化中, 植物对水分的需求驱动着叶脉网络的变化, 气孔是植物体蒸腾失水的门户(左应梅等, 2011 ), 植物叶片选择发育更有效的叶脉灌输系统和密集的气孔提高光合效率和水分利用效率(张亚等, 2014 ), 对植物生理功能的维持具有重要意义。本研究发现, 各个坡向刺槐的叶脉密度与气孔密度呈正相关关系, 与气孔大小呈负相关关系, 且在南坡达到极显著水平(p < 0.01, 图1 , 图2 ), 表明坡向梯度上刺槐叶片结构不同功能之间的相互匹配与权衡关系是植物叶片的形态结构和特定的水分供应需求分配策略对生境异质性适应的结果。
环境胁迫驱动下的自然选择使得植物倾向于以最小的水分散失来获得最大的碳同化速率(Hether- ington & Woodward, 2003 ), 植物通过调整自身生理特征和解剖结构来适应不同的生境条件(王瑞丽等, 2016 )。生长在南坡刺槐的叶脉密度与气孔密度呈极显著正相关关系(p < 0.01, 图1 ), 与气孔大小存在极显著负相关关系(p < 0.01, 图2 )。研究区位于北半球中纬度地区山地, 南坡太阳直射角较大, 光照时间长, 接受的光照充足, 总的辐射总量和温度均高于北坡(党晶晶等, 2014 ), 南坡植被稀疏, 接受的光合有效辐射最大(表2 ), 高温、强辐射造成刺槐叶片蒸腾加剧, 较强的蒸腾作用使植物面临着水分胁迫, 为了适应高温下的蒸腾速率, 植物倾向于发育小而厚的叶片(史元春等, 2015 ), 将更多的光合产物用于增加叶脉密度以保证高效率的水分输导系统。叶片在进行光合作用时不可避免地伴随着水分的蒸发损耗(Brodribb & Jordan, 2011 ), 在叶片增加运输管道时也应考虑到水分的散失, 气孔作为水分蒸腾的通道, 在土壤中的水分减少时植物供水能力降低(表1 ), 会影响气孔的发生、分化和发育等, 单位面积内叶脉密度的增加使叶脉间距变小, 气孔变窄、气孔密度加大(陈伟月等, 2014 ), 如果气孔不能感知这些变化并做出响应, 就提高了木质部栓塞的可能性, 进而增加了叶片液压功能失调至脱水的风险(Maseda & Fernández, 2006 )。当植株叶片获得的光合有效辐射较大时, 叶片气孔及表皮细胞迅速对光照做出反应, 直接向大气蒸发散水, 此时叶片其他部位并未发生水分亏缺, 小的气孔可以迅速关闭, 防止水分进一步流失, 避免水分亏缺在全叶中发生, 以此减少植物整叶枯萎的损失(高春娟等, 2012 )。因此, 生长在南坡的刺槐选择高的叶脉密度、气孔密度和小的气孔, 验证了Zhang (2012)的叶脉密度与气孔特征协同进化的理论。
植物叶片构型构建与植物生理功能所需的光照、碳投入和水分运输的投资收益相契合, 是植物提高光合收益的一个重要的生活史策略(Westoby et al ., 2002 ; 孙素静等, 2015 )。生长在北坡的刺槐的叶脉密度与气孔密度呈显著正相关关系(p < 0.05, 图1 ), 与气孔大小存在显著负相关关系(p < 0.05, 图2 )。随着资源环境的改变, 北坡植被群落郁闭程度和拥挤程度达到最大, 来自邻株植物的遮阴改变了植物所处光环境, 限制了叶片可获得性光资源, 光合有效辐射减弱(表2 )。植株为了获得更多的光资源, 选择了增大叶面积的生长策略, 使得部分叶肉组织与叶脉接触面积减少, 相对减少叶脉构建投资; 另一方面, 在土壤含水量较充沛的条件下, 植物供水较充足, 气孔张合度受到影响, 单位叶面积内气孔数量减少且CO2 扩散阻力增加, 叶片吸收CO2 受到限制, 光合速率显著降低, 引起植物地上部分对水的需求减少, 使得植物不必投资更多叶脉导管就可以满足叶片水分的供应, 有利于提高植物在资源丰富的环境中的竞争力(Beerling & Franks, 2010 ; Field et al ., 2011 )。气孔直接影响着植物的CO2 交换速率、蒸腾速率和光合速率, 调节气孔密度和气孔长度可适应时刻发生变化的环境状况, 使植物在水碳循环中的平衡调节作用达到最优(高春娟等, 2012 ; 高冠龙等, 2016 ), 当土壤水分较充足时, 根系吸收的水分足以保持体内水分平衡, 气孔进一步开放虽然使得蒸腾速率增加, 但气孔对蒸腾速率的敏感度降低, 吸水速率与蒸腾速率的平衡机制对气孔反馈减少(左应梅等, 2011 ), 采取了“疏而大”的适应特征, 从而保证叶片能吸收足量的CO2 , 进行正常的光合作用, 这与Dow等(2014)对拟南芥(Arabidopsis thaliana )气孔的研究结果一致。刺槐在北坡选择了降低叶脉密度与气孔密度、增大气孔的策略, 使叶含水量维持在动态平衡范围内, 这对于理解植物对环境的适应策略、叶性状之间资源的分配与利用具有重要意义。
形态特征和生长特性的改变是植物适应不同环境和资源水平的重要策略(Maherali & Pelucia, 2001 ), 叶片形态和生理特性在植物碳同化、水分关系和能量平衡等方面具有重要作用, 决定了叶片功能属性在不同环境条件下的差异(Wright et al ., 2004 ; Poorter et al ., 2006 )。东坡和西坡位于阴坡和阳坡过渡区域, 属于植被类型的交错地带, 群落组成较复杂, 群落郁闭度和高度都介于南坡和北坡之间。自然条件下刺槐选择折中了叶脉密度与气孔密度、气孔大小的稳步投资, 兼顾了植株的水分输送、利用与保持等功能的需求, 选择了气孔与叶脉的均衡生长模式, 可以达到更大限度地利用地上部分资源和拓展空间的目的, 充分利用水资源与光资源, 减少植株内部的资源竞争。因此, 生长在东西坡的刺槐的气孔密度、气孔大小与叶脉密度的相关性之间没有明显差异, 这是光合产物在不同构件或功能间权衡分配的结果。
植物叶片气孔性状与叶脉密度的关系是植物叶片生理生态功能对环境适应过程的表现, 体现了不同坡向上刺槐叶片内部构件之间的资源协调分配。本研究发现: 随着坡向的改变, 刺槐的群落环境和土壤水分发生了明显变化, 刺槐通过调整叶片的结构特征来适应环境的变化。叶脉密度与气孔性状之间存在一种协调性, 为了提高种群优势, 满足植株获取光合产物和提高水分利用效率的需求, 生长在南坡的刺槐选择了较高的叶脉密度、气孔密度和小的气孔, 以应对水分胁迫的威胁, 提高水分利用效率; 而生长在北坡的刺槐选择了较低的叶脉密度、气孔密度和大的气孔, 弥补因邻体干扰造成的光抑制, 增大气孔张合度最大化获取光照, 这种结果表明生长在南、北坡刺槐的投资模式表现出截然相反的趋势, 较好地验证了植物应对环境变异的可塑性调节机制。本文仅从叶片内部结构的角度分析了刺槐的水分利用策略, 今后还可以从植株的根和茎分析刺槐的水分传输, 为建立植物根、茎、叶水力机理模型奠定良好的理论基础。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Bazzaz FA Chiariello NR Coley PD Pitelka LF 1987 ). Allocating resources to reproduction and defense
.BioScience 37 , 58 -67 .
[本文引用: 1]
[2]
Beerling DJ Franks PJ 2010 ). Plant science: The hidden cost of transpiration
.Nature 464 , 495 -496 .
[本文引用: 1]
[3]
Berlyn GP Miksche JP 1976). Botanical Microtechnique and Cytochemistry . Iowa State University Press, Ames, USA .
[4]
Blonder B Violle C Bentley LP Enquist BJ 2011 ). Venation networks and the origin of the leaf economics spectrum
.Ecology Letters 14 , 91 -100 .
[本文引用: 1]
[5]
Brodribb TJ Feild TS Sack L 2010 ). Viewing leaf structure and evolution from a hydraulic perspective
.Functional Plant Biology 37 , 488 -498 .
[本文引用: 2]
[6]
Brodribb TJ Jordan GJ 2011 ). Water supply and demand remain balanced during leaf acclimation of
Nothofagus cunninghamii trees. New Phytologist 192 , 437 -448 .
[本文引用: 3]
[7]
Cantón Y del Barrio G Solé-Benet A Lázaro R 2004 ). Topographic controls on the spatial distribution of ground cover in the Tabernas badlands of SE Spain
.Catena 55 , 341 -365 .
[8]
Chen WY LIU CH LI YY Min DH 2014 ). Flag leaf vein traits in winter wheat varieties (lines) and their correlation with stomatal traits
.Chinese Journal of Ecology 33 , 1839 -1846 .(in English with Chinese abstract) [陈伟月 , 刘存海 , 李秧秧 , 闵东红 (2014 ). 冬小麦品种(系)旗叶叶脉性状及其与气孔性状间的关联性. 生态学杂志,
33 , 1839 -1846 .]
[本文引用: 1]
[9]
Chen Y Xu X Zhang DR Wei Y 2006 ). Correlations between vegetation distribution and topographical factors in the northwest of Longmen Mountain, Sichuan Province
.Chinese Journal of Ecology 25 , 1052 -1055 .(in English with Chinese abstract) [陈瑶 , 胥晓 , 张德然 , 魏勇 (2006 ). 四川龙门山西北部植被分布与地形因子的相关性. 生态学杂志,
25 , 1052 -1055 .]
[本文引用: 1]
[10]
Dang JJ Zhao CZ Li Y Hou ZJ Dong XG 2014 ). Variations with slope in stem and leaf traits of
Melica przewalskyi in alpine grassland. Chinese Journal of Plant Ecology 38 , 1307 -1314 .(in English with Chinese abstract) [党晶晶 , 赵成章 , 李钰 , 侯兆疆 , 董小刚 (2014 ). 高寒草地甘肃臭草茎-叶性状的坡度差异性. 植物生态学报,
38 , 1307 -1314 .]
[本文引用: 2]
[11]
Dow GJ Berry JA Bergann DC 2014 ). The physiological importance of developmental mechanisms that enforce proper stomatal spacing in
Arabidopsis thaliana . New Phytologist 201 , 1205 -1217 .
[12]
Du J Zhao CZ Song QH Shi YC Wang JW Chen J 2016 ). Plant size differences with twig and leaf traits of
Zygophyllum xanthoxylum in the northern slope of Qilian Mountains, China. Chinese Journal of Plant Ecology 40 , 212 -220 .(in English with Chinese abstract) [杜晶 , 赵成章 , 宋清华 , 史元春 , 王继伟 , 陈静 (2016 ). 祁连山北坡霸王枝-叶性状关系的个体大小差异. 植物生态学报,
40 , 212 -220 .]
[本文引用: 1]
[13]
Falster DS Warton DI Wright IJ
URL
[本文引用: 1]
[14]
Field TS Brodribb TJ Iglesias A Chatelet DS Baresch A Upchurch GR Jr. Gomez B Mohr BAR Coiffard C Kvacek J Jaramillo C 2011 ). Fossil evidence for Cretaceous escalation in angiosperm leaf vein evolution
.Proceedings of the National Academy of Sciences of the United States of America 108 , 8363 -8366 .
[本文引用: 2]
[15]
Fiorin L Brodribb TJ Anfodillo T 2016 ). Transport efficiency through uniformity: Organization of veins and stomata in angiosperm leaves
.New Phytologist 209 , 216 -227 .
[本文引用: 2]
[16]
Gao CJ Xia XJ Shi K Zhou YH Yu JQ 2012 ). Response of stomata to global climate changes and the underlying regulation mechanism of stress responses
.Plant Physiology Journal 48 , 19 -28 .(in English with Chinese abstract) [高春娟 , 夏晓剑 , 师恺 , 周艳虹 , 喻景权 (2012 ). 植物气孔对全球环境变化的响应及其调控防御机制. 植物生理学报,
48 , 19 -28 .]
[本文引用: 1]
[17]
Gao GL Zhang XY Chang ZQ Yu TF Zhao H 2016 ). Environmental response simulation and the up-scaling of plant stomatal conductance
. Acta Ecologica Sinica 36 , 1491 -1500 .(in English with Chinese abstract) [高冠龙 , 张小由 , 常宗强 , 鱼腾飞 , 赵虹 (2016 ). 植物气孔导度的环境响应模拟及其尺度扩展. 生态学报,
36 , 1491 -1500 .]
[本文引用: 1]
[18]
Gao J Cao KF Wang HX 2004 ). Water relations and stomatal conductance in nine tree species during a dry period grown in a hot and dry valley
.Acta Phytoecologica Sinica 28 , 186 -190 .(in English with Chinese abstract) [高洁 , 曹坤芳 , 王焕校 (2004 ). 干热河谷9种造林树种在旱季的水分关系和气孔导度. 植物生态学报,
28 , 186 -190 .]
[19]
Hethrington AM Woodward FI 2003 ). The role of stomata in sensing and driving environmental change
.Nature 424 , 901 -908 .
[本文引用: 1]
[20]
Jin Y Wang CK 2015 ). Trade-offs between plant leaf hydraulic and economic traits
.Chinese Journal of Plant Ecology 39 , 1021 -1032 .(in Chinese with English abstract) [金鹰 , 王传宽 (2015 ). 植物叶片水力与经济性状权衡关系的研究进展. 植物生态学报,
39 , 1021 -1032 .]
[本文引用: 1]
[21]
Li J Wang XC Shao MA Zhao YJ Li XF 2010 ). Simulation of biomass and soil desiccation of
Robinia pseudoacacia forestlands on semi-arid and semi-humid regions of China’s Loess Plateau. Chinese Journal of Plant Ecology 34 , 330 -339 .(in Chinese with English abstract) [李军 , 王学春 , 邵明安 , 赵玉娟 , 李小芳 (2010 ). 黄土高原半干旱和半湿润地区刺槐林地生物量与土壤干燥化效应的模拟. 植物生态学报,
34 , 330 -339 .]
[本文引用: 1]
[22]
Li L Zeng H Guo DL 2013 ). Leaf venation functional traits and their ecological significance
.Chinese Journal of Plant Ecology 37 , 691 -698 .(in Chinese with English abstract) [李乐 , 曾辉 , 郭大立 (2013 ). 叶脉网络功能性状及其生态学意义. 植物生态学报,
37 , 691 -698 .]
[23]
Maherali H Delucia EH 2001 ). Influence of climate-driven shifts in biomass allocation on water transport and storage in ponderosa pine
.Oecologia 129 , 481 -491 .
[本文引用: 1]
[24]
Maricle BR Koteyeva NK Voznesenskaya EV Thomasson JR Edwards GE 2009 ). Diversity in leaf anatomy, and stomatal distribution and conductance, between salt marsh and freshwater species in the C4 genus
Spartina Poaceae). New Phytologist ,184 , 216 -233 .
[本文引用: 1]
[25]
Maseda PH Fernández RJ 2006 ). Stay wet or else: Three ways in which plants can adjust hydraulically to their environment
.Journal of Experimental Botany 57 , 3963 -3977 .
[26]
Pitman EJG 1939 ). A note on normal correlation
.Biometrika 31 , 9 -12 .
[本文引用: 1]
[27]
Poorter L Bongers L Bongers F 2006 ). Architecture of 54 moist forest tree species: Traits, trade-offs, and functional groups
.Ecology 87 , 1289 -1301 .
[本文引用: 1]
[28]
Ren QJ Li HL Bu HY 2015 ). Comparison of physiological and leaf morphological traits for photosynthesis of the 51 plant species in the Maqu alpine swamp meadow
.Chinese Journal of Plant Ecology 39 , 593 -603 .(in English with Chinese abstract) [任青吉 , 李宏林 , 卜海燕 (2015 ). 玛曲高寒沼泽化草甸51种植物光合生理和叶片形态特征的比较. 植物生态学报,
39 , 593 -603 .]
[本文引用: 1]
[29]
Sack L Frole K 2006 ). Leaf structural diversity is related to hydraulic capacity in tropical rain forest trees
.Ecology 87 , 483 -491 .
[本文引用: 2]
[30]
Sack L Scoffoni C 2013 ). Leaf venation: Structure, function, development, evolution, ecology and applications in the past, present and future
.New Phytologist 198 , 983 -1000 .
[本文引用: 1]
[31]
Sack L Scoffoni C McKown AD Frole K Havran JC Tran H Tran T 2012 ). Developmentally based scaling of leaf venation architecture explains global ecological patterns
.Nature Communications 3 , 837 .
[本文引用: 1]
[32]
Scoffoni C Kunkle J Pasquet-Kok J Vuong C Patel AJ Montgomery RA Thomas J Givnish TJ Sack L 2015 ). Light-induced plasticity in leaf hydraulics, venation, anatomy, and gas exchange in ecologically diverse
Hawaiian lobeliads . New Phytologist 207 , 43 -58 .
[本文引用: 1]
[33]
Shen JP Zhang WH 2014 ). Characteristics of carbon storage and sequestration of
Robinia pseudoacacia forest land converted by farmland in the Hilly Loess Plateau Region. Acta Ecologica Sinica 34 , 2746 -2754 .(in English with Chinese abstract) [申家朋 , 张文辉 (2014 ). 黄土丘陵区退耕还林地刺槐人工林碳储量及分配规律. 生态学报,
34 , 2746 -2754 .]
[本文引用: 2]
[34]
Shi YC Zhao CZ Song QH Du J Chen J Wang JW 2015 ). Slope-related variations in twig and leaf traits of
Robinia pseudoacacia in the northern mountains of Lanzhou. Chinese Journal of Plant Ecology 39 , 362 -370 .(in English with Chinese abstract) [史元春 , 赵成章 , 宋清华 , 杜晶 , 陈静 , 王继伟 (2015 ). 兰州北山刺槐枝叶性状的坡向差异性. 植物生态学报,
39 , 362 -370 .]
[本文引用: 1]
[35]
Skidmore AK 1989 ). A comparison of techniques for calculating gradient and aspect from a gridded digital elevation model
.International Journal of Geographical Information Science 3 , 323 -334 .
[本文引用: 1]
[36]
Sun SJ Li FL Bao WK 2015 ), Advances on construction of leaf venation system and its significance
.Journal of Tropical and Subtropical Botany of Phylogeny 23 , 353 -360 .(in English with Chinese abstract) [孙素静 , 李芳兰 , 包维楷 (2015 ). 叶脉网络系统的构建和系统学意义研究进展. 热带亚热带植物学报,
23 , 353 -360 .]
[本文引用: 1]
[37]
Wang N Zhang Y Qian WL Wang ZQ Gu JC 2016 ). Effects of elevated CO2 concentration on root and needle anatomy and physiological functions in
Pinus koraiensis seedlings. Chinese Journal of Plant Ecology 40 , 60 -68 .(in English with Chinese abstract) [王娜 , 张韫 , 钱文丽 , 王政权 , 谷加存 (2016 ). CO2 浓度倍增对红松幼苗根尖和叶解剖结构及生理功能的影响. 植物生态学报,
40 , 60 -68 .]
[本文引用: 1]
[38]
Wang RL Yu GR He NP Wang QF Zhao N Xu ZW 2016 ). Altitudinal variation in the covariation of stomatal traits with leaf functional traits in Changbai Mountain
.Acta Ecologica Sinica 36 , 1 -10 .(in English with Chinese abstract) [王瑞丽 , 于贵瑞 , 何念鹏 , 王秋凤 , 赵宁 , 徐志伟 (2016 ). 气孔特征与叶片功能性状之间关联性沿海拔梯度的变化规律: 以长白山为例. 生态学报,
36 , 1 -10 .]
[本文引用: 1]
[39]
Wang SG Li ZQ Jia SS Sun DZ Shi YG Fan H Liang ZH Jing RL 2013 ). Relationships of wheat leaf stomatal traits with wheat yield and drought-resistance
.Chinese Journal of Applied Ecology 24 , 1609 -1614 .(in English with Chinese abstract) [王曙光 , 李中青 , 贾寿山 , 孙黛珍 , 史雨刚 , 范华 , 梁增浩 , 景蕊莲 (2013 ). 小麦叶片气孔性状与产量和抗旱性的关系. 应用生态学报,
24 , 1609 -1614 .]
[40]
Warton DI Weber NC 2002 ). Common slope tests for bivariate errors-in-variables models
.Biometrical Journal 44 , 161 -174 .
[本文引用: 1]
[41]
Warton DI Wright IJ Falster DS 2006 ). Bivariate line-fitting methods for allometry
.Biological Reviews 81 , 259 -291 .
[本文引用: 1]
[42]
Wei LL Zhang XQ Hou ZH Xu DY Yu XB 2005 ). Effects of water stress on photosynthesis and carbon allocation in
Cunninghamia lancealata seedlings. Acta Phytoecologica Sinica 29 , 394 -402 .(in Chinese with English abstract) [韦莉莉 , 张小全 , 侯振宏 , 徐德应 , 余雪标 (2005 ). 杉木苗木光合作用及其产物分配对水分胁迫的响应. 植物生态学报,
29 , 394 -402 .]
[本文引用: 1]
[43]
Westoby M Falster DS Moles AT Vesk PA Wright LJ 2002 ). Plant ecological strategies: Some leading dimensions of variation between species
.Annual Review of Ecology Systematics 33 , 125 -159 .
[本文引用: 1]
[44]
Wright IJ Reich PB Westoby M Ackerly DD Baruch Z Bongers F Cavender-Bares J Chapin T Cornelissen JH Diemer M Flexas J Garnier E Groom PK Gulias J Hikosaka K Lamont BB Lee T Lee W Lusk C Midgley JJ Navas ML Niinemets U Oleksyn J Osada N Poorter H Poot P Prior L Pyankov VI Roumet C Thomas SC Tjoelker MG Veneklaas EJ Villar R 2004 ). The worldwide leaf economics spectrum
.Nature 428 , 821 -827 .
[本文引用: 1]
[45]
Xu T Zhao CZ Duan BB Han L Zheng HL Feng W 2016 ). Slope-related variations of different levels of vein density and leaf size in
Robinia pseudoacacia in the northern mountains of Lanzhou. Chinese Journal of Ecology 35 , 41 -47 .(in English with Chinese abstract) [徐婷 , 赵成章 , 段贝贝 , 韩玲 , 郑慧玲 , 冯威 (2016 ). 兰州北山刺槐不同等级叶脉密度与叶大小关系的坡向差异性. 生态学杂志,
35 , 41 -47 .]
[本文引用: 1]
[46]
Xu Z Zhou G 2008 ). Responses of leaf stomatal density to water status and its relationship with photosynthesis in a grass
.Journal of Experimental Botany 59 , 3317 -3325 .
[本文引用: 2]
[47]
Yang HM Wang GX 2001 ). Leaf stomatal densities and distribution in
Triticum Aestivum under drought and CO2 enrichment. Acta Phytoecologica Sinica 25 , 312 -316 .(in English with Chinese abstract) [杨惠敏 , 王根轩 (2001 ). 干旱和CO2 浓度升高对干旱区春小麦气孔密度及分布的影响. 植物生态学报,
25 , 312 -316 .]
[本文引用: 1]
[48]
Yang LM Han M Zhou GS Li JD 2007 ). The changes of water-use efficiency and stoma density of
Leymus chinensis a long Northeast China Transect. Acta Ecologica Sinica 27 , 16 -24 .(in Chinese with English abstract) [杨利民 , 韩梅 , 周广胜 , 李建东 (2007 ). 中国东北样带关键种羊草水分利用效率与气孔密度. 生态学报,
27 , 16 -24 .]
[本文引用: 1]
[49]
Yu ZH Chen YM Du S 2009 ). Sap flow dynamics in the leaf-flushing period of a
Robinia pseudoacacia plantation in semi-arid region of Loess Plateau. Scientia Silvae Sinicae 45 (4 ), 53 -59 .(in Chinese with English abstract) [于占辉 , 陈云明 , 杜盛 (2009 ). 黄土高原半干旱区人工林刺槐展叶期树干液流动态分析. 林业科学,
45(4) , 53 -59 .]
[本文引用: 1]
[50]
Zhang SB Guan ZJ Sun M Zhang JJ Cao KF Hu H 2012 ). Evolutionary association of stomatal traits with leaf vein density in
Paphiopedilum, Orchidaceae. PLOS ONE 7 , e40080 , doi: 10.1371/journal.pone.0040080 .
[本文引用: 1]
[51]
Zhang Y Yang SJ Sun M Cao KF 2014 ). Stomatal traits are evolutionarily associated with vein density in basal angiosperms
.Plant Science Journal 32 , 320 -328 .(in English with Chinese abstract) [张亚 , 杨石建 , 孙梅 , 曹坤芳 (2014 ). 基部被子植物气孔性状与叶脉密度的关联进化. 植物科学学报,
32 , 320 -328 .]
[本文引用: 3]
[52]
Zuo YM Chen QB Deng QQ Tang J Luo HW Wu TK Yang ZF 2011 ). Effects of soil moisture, light, and air humidity on stomatal conductance of cassava (
Manihot esculenta Crantz). Chinese Journal of Ecology 30 , 689 -693 .(in English with Chinese abstract) [左应梅 , 陈秋波 , 邓权权 , 唐建 , 罗海伟 , 巫铁凯 , 杨重法 (2011 ). 土壤水分、光照和空气湿度对木薯气孔导度的影响. 生态学杂志,
30 , 689 -693 .]
Allocating resources to reproduction and defense
1
1987
... 作为植物碳水耦合权衡的重要器官, 叶片在进化过程中对环境的变化较敏感且可塑性较大(Scoffoni et al ., 2015 ), 从叶片形态解剖结构了解植物生态策略在异质生境中的变化对植物的结构和生理变化具有重要意义(Maricle et al ., 2009 ).叶脉是影响叶片水分供应和利用的重要结构, 在一定意义上反映了叶片资源流通能力(Sack & Frole, 2006 ; Sack & Scoffoni, 2013 ).叶脉密度作为叶脉网络功能性状的重要指标(Brodribb et al ., 2010 ; Blonder et al ., 2011 ), 可反映叶脉系统的水分、养分和光合产物等物质运输的效率和安全性以及抵抗外界干扰的能力, 是理解植物环境可塑性机制的重要途径(李乐等, 2013 ).气孔是植物叶片与外界进行水气交换的唯一可调节通道, 通过保卫细胞对环境和内源信号的感知, 与植物的光合作用、蒸腾作用等生理生态功能密切相关(杨惠敏和王根轩, 2001 ).叶脉与气孔是叶片水分供给和散失的重要因素, 两者的大小和数量的配置直接影响着叶片的水分利用效率, 进而影响叶片结构及生理功能的调节机制(Fiorin et al ., 2016 ).当植株处于高光强条件下时, 相对湿度下降, 叶片受到的水分胁迫增加, 叶片在获取光合收益的同时也需兼顾内部水分动态平衡(Fiorin et al ., 2016 ), 此时, 植株往往通过调整净光合速率与蒸腾速率来应对不利环境, 较高的蒸腾速率要求叶片发育较多的叶脉以提供供水系统, 在叶片的蒸腾作用中通过控制气孔密度和气孔大小决定进入大气的水分和CO2 , 降低植物的呼吸和蒸腾成本.叶脉密度与气孔性状共同作用体现了气孔蒸腾对水分的需求和叶脉系统对水分的供给相互匹配与权衡(Brodribb & Jordan, 2011 ).坡向作为重要的地形因素, 控制了太阳辐射和降水的空间再分配(Cartón et al ., 2004 ), 由坡向引起的土壤水分、光辐射、光照时长和大气温度等环境因子形成的生境梯度变化影响着群落组成(Bazzaz et al ., 1987 ; 韦莉莉等, 2005 ), 致使群落环境及其密度制约下的种群内植株的生态位存在差异, 群落内部郁闭度和干扰程度不同, 所接受的光合有效辐射也不同, 从而影响植物的空间分布格局及光照、温度等环境(党晶晶等, 2014 ; 杜晶等, 2016 ), 进而影响植物的形态.为了提高光合能力并保证水分供需间的平衡, 植株叶片会适当调整叶脉密度与气孔密度、气孔大小的生物量分配, 补充蒸腾带来的大量水分和能量的消耗(Hetherington & Woodward, 2003 ), 同时也使叶片中单位碳对光合作用的投资效益最大(Brodribb & Jordan, 2011 ), 从而提高生存适合度和竞争能力.因此, 研究微地形影响下叶脉密度与气孔性状的关系, 有助于探讨植物叶片内部水分供应与散失之间的平衡关系, 对植物生理生态功能的维持具有指导意义. ...
Plant science: The hidden cost of transpiration
1
2010
... 植物叶片构型构建与植物生理功能所需的光照、碳投入和水分运输的投资收益相契合, 是植物提高光合收益的一个重要的生活史策略(Westoby et al ., 2002 ; 孙素静等, 2015 ).生长在北坡的刺槐的叶脉密度与气孔密度呈显著正相关关系(p < 0.05, 图1 ), 与气孔大小存在显著负相关关系(p < 0.05, 图2 ).随着资源环境的改变, 北坡植被群落郁闭程度和拥挤程度达到最大, 来自邻株植物的遮阴改变了植物所处光环境, 限制了叶片可获得性光资源, 光合有效辐射减弱(表2 ).植株为了获得更多的光资源, 选择了增大叶面积的生长策略, 使得部分叶肉组织与叶脉接触面积减少, 相对减少叶脉构建投资; 另一方面, 在土壤含水量较充沛的条件下, 植物供水较充足, 气孔张合度受到影响, 单位叶面积内气孔数量减少且CO2 扩散阻力增加, 叶片吸收CO2 受到限制, 光合速率显著降低, 引起植物地上部分对水的需求减少, 使得植物不必投资更多叶脉导管就可以满足叶片水分的供应, 有利于提高植物在资源丰富的环境中的竞争力(Beerling & Franks, 2010 ; Field et al ., 2011 ).气孔直接影响着植物的CO2 交换速率、蒸腾速率和光合速率, 调节气孔密度和气孔长度可适应时刻发生变化的环境状况, 使植物在水碳循环中的平衡调节作用达到最优(高春娟等, 2012 ; 高冠龙等, 2016 ), 当土壤水分较充足时, 根系吸收的水分足以保持体内水分平衡, 气孔进一步开放虽然使得蒸腾速率增加, 但气孔对蒸腾速率的敏感度降低, 吸水速率与蒸腾速率的平衡机制对气孔反馈减少(左应梅等, 2011 ), 采取了“疏而大”的适应特征, 从而保证叶片能吸收足量的CO2 , 进行正常的光合作用, 这与Dow等(2014)对拟南芥(Arabidopsis thaliana )气孔的研究结果一致.刺槐在北坡选择了降低叶脉密度与气孔密度、增大气孔的策略, 使叶含水量维持在动态平衡范围内, 这对于理解植物对环境的适应策略、叶性状之间资源的分配与利用具有重要意义. ...
Venation networks and the origin of the leaf economics spectrum
1
2011
... 作为植物碳水耦合权衡的重要器官, 叶片在进化过程中对环境的变化较敏感且可塑性较大(Scoffoni et al ., 2015 ), 从叶片形态解剖结构了解植物生态策略在异质生境中的变化对植物的结构和生理变化具有重要意义(Maricle et al ., 2009 ).叶脉是影响叶片水分供应和利用的重要结构, 在一定意义上反映了叶片资源流通能力(Sack & Frole, 2006 ; Sack & Scoffoni, 2013 ).叶脉密度作为叶脉网络功能性状的重要指标(Brodribb et al ., 2010 ; Blonder et al ., 2011 ), 可反映叶脉系统的水分、养分和光合产物等物质运输的效率和安全性以及抵抗外界干扰的能力, 是理解植物环境可塑性机制的重要途径(李乐等, 2013 ).气孔是植物叶片与外界进行水气交换的唯一可调节通道, 通过保卫细胞对环境和内源信号的感知, 与植物的光合作用、蒸腾作用等生理生态功能密切相关(杨惠敏和王根轩, 2001 ).叶脉与气孔是叶片水分供给和散失的重要因素, 两者的大小和数量的配置直接影响着叶片的水分利用效率, 进而影响叶片结构及生理功能的调节机制(Fiorin et al ., 2016 ).当植株处于高光强条件下时, 相对湿度下降, 叶片受到的水分胁迫增加, 叶片在获取光合收益的同时也需兼顾内部水分动态平衡(Fiorin et al ., 2016 ), 此时, 植株往往通过调整净光合速率与蒸腾速率来应对不利环境, 较高的蒸腾速率要求叶片发育较多的叶脉以提供供水系统, 在叶片的蒸腾作用中通过控制气孔密度和气孔大小决定进入大气的水分和CO2 , 降低植物的呼吸和蒸腾成本.叶脉密度与气孔性状共同作用体现了气孔蒸腾对水分的需求和叶脉系统对水分的供给相互匹配与权衡(Brodribb & Jordan, 2011 ).坡向作为重要的地形因素, 控制了太阳辐射和降水的空间再分配(Cartón et al ., 2004 ), 由坡向引起的土壤水分、光辐射、光照时长和大气温度等环境因子形成的生境梯度变化影响着群落组成(Bazzaz et al ., 1987 ; 韦莉莉等, 2005 ), 致使群落环境及其密度制约下的种群内植株的生态位存在差异, 群落内部郁闭度和干扰程度不同, 所接受的光合有效辐射也不同, 从而影响植物的空间分布格局及光照、温度等环境(党晶晶等, 2014 ; 杜晶等, 2016 ), 进而影响植物的形态.为了提高光合能力并保证水分供需间的平衡, 植株叶片会适当调整叶脉密度与气孔密度、气孔大小的生物量分配, 补充蒸腾带来的大量水分和能量的消耗(Hetherington & Woodward, 2003 ), 同时也使叶片中单位碳对光合作用的投资效益最大(Brodribb & Jordan, 2011 ), 从而提高生存适合度和竞争能力.因此, 研究微地形影响下叶脉密度与气孔性状的关系, 有助于探讨植物叶片内部水分供应与散失之间的平衡关系, 对植物生理生态功能的维持具有指导意义. ...
Viewing leaf structure and evolution from a hydraulic perspective
2
2010
... 作为植物碳水耦合权衡的重要器官, 叶片在进化过程中对环境的变化较敏感且可塑性较大(Scoffoni et al ., 2015 ), 从叶片形态解剖结构了解植物生态策略在异质生境中的变化对植物的结构和生理变化具有重要意义(Maricle et al ., 2009 ).叶脉是影响叶片水分供应和利用的重要结构, 在一定意义上反映了叶片资源流通能力(Sack & Frole, 2006 ; Sack & Scoffoni, 2013 ).叶脉密度作为叶脉网络功能性状的重要指标(Brodribb et al ., 2010 ; Blonder et al ., 2011 ), 可反映叶脉系统的水分、养分和光合产物等物质运输的效率和安全性以及抵抗外界干扰的能力, 是理解植物环境可塑性机制的重要途径(李乐等, 2013 ).气孔是植物叶片与外界进行水气交换的唯一可调节通道, 通过保卫细胞对环境和内源信号的感知, 与植物的光合作用、蒸腾作用等生理生态功能密切相关(杨惠敏和王根轩, 2001 ).叶脉与气孔是叶片水分供给和散失的重要因素, 两者的大小和数量的配置直接影响着叶片的水分利用效率, 进而影响叶片结构及生理功能的调节机制(Fiorin et al ., 2016 ).当植株处于高光强条件下时, 相对湿度下降, 叶片受到的水分胁迫增加, 叶片在获取光合收益的同时也需兼顾内部水分动态平衡(Fiorin et al ., 2016 ), 此时, 植株往往通过调整净光合速率与蒸腾速率来应对不利环境, 较高的蒸腾速率要求叶片发育较多的叶脉以提供供水系统, 在叶片的蒸腾作用中通过控制气孔密度和气孔大小决定进入大气的水分和CO2 , 降低植物的呼吸和蒸腾成本.叶脉密度与气孔性状共同作用体现了气孔蒸腾对水分的需求和叶脉系统对水分的供给相互匹配与权衡(Brodribb & Jordan, 2011 ).坡向作为重要的地形因素, 控制了太阳辐射和降水的空间再分配(Cartón et al ., 2004 ), 由坡向引起的土壤水分、光辐射、光照时长和大气温度等环境因子形成的生境梯度变化影响着群落组成(Bazzaz et al ., 1987 ; 韦莉莉等, 2005 ), 致使群落环境及其密度制约下的种群内植株的生态位存在差异, 群落内部郁闭度和干扰程度不同, 所接受的光合有效辐射也不同, 从而影响植物的空间分布格局及光照、温度等环境(党晶晶等, 2014 ; 杜晶等, 2016 ), 进而影响植物的形态.为了提高光合能力并保证水分供需间的平衡, 植株叶片会适当调整叶脉密度与气孔密度、气孔大小的生物量分配, 补充蒸腾带来的大量水分和能量的消耗(Hetherington & Woodward, 2003 ), 同时也使叶片中单位碳对光合作用的投资效益最大(Brodribb & Jordan, 2011 ), 从而提高生存适合度和竞争能力.因此, 研究微地形影响下叶脉密度与气孔性状的关系, 有助于探讨植物叶片内部水分供应与散失之间的平衡关系, 对植物生理生态功能的维持具有指导意义. ...
... 刺槐(Robinia pseudoacacia )是温带落叶乔木, 是我国华北和西北地区绿化荒山和防风固沙的主要植物, 具有固氮能力强和碳储存量大的优势, 作为黄土丘陵区的主要人工造林树种, 其根系发达、生长迅速、成活率高和环境适应性强等特点在保持水土和生态防护中发挥着重要作用.近年来, 关于刺槐在干旱半干旱地区的种植范围和生态适应性等问题引起了学界的广泛关注(于占辉等, 2009 ; 李军等, 2010 ; 申家朋和张文辉, 2014 ).学术界对气孔性状之间的关系(杨惠敏和王根轩, 2001 ; 王曙光等, 2013 )、气孔与环境因子的关系(左应梅等, 2011 ; 王娜等, 2016 )、气孔性状对水分利用的影响(高洁等, 2004 ; 杨利民等, 2007 )、叶脉网络功能性状与叶片光合作用(Brodribb et al ., 2010 ; Sack et al ., 2012 )、水分利用能力(Zhang et al ., 2012 ; Sack & Scoffoni, 2013 )等问题进行了研究, 阐述了植物叶片在适应特定生境中各种性状的塑造, 而将叶脉与气孔性状结合起来, 探讨二者在异质性生境中叶片内部水分传输及水分平衡适应策略的分析和认识目前还比较薄弱.鉴于此, 本文以兰州北山的人工刺槐林为研究对象, 试图揭示以下两个问题: (1)不同坡向上刺槐叶脉密度与气孔密度、气孔大小之间存在何种关系?(2)这种关系的形成原因有哪些?理解异质生境中植物叶片的物理构建方式, 可为提高半干旱人工林资源的利用率和绿化覆盖率提供理论依据. ...
Water supply and demand remain balanced during leaf acclimation of
3
2011
... 作为植物碳水耦合权衡的重要器官, 叶片在进化过程中对环境的变化较敏感且可塑性较大(Scoffoni et al ., 2015 ), 从叶片形态解剖结构了解植物生态策略在异质生境中的变化对植物的结构和生理变化具有重要意义(Maricle et al ., 2009 ).叶脉是影响叶片水分供应和利用的重要结构, 在一定意义上反映了叶片资源流通能力(Sack & Frole, 2006 ; Sack & Scoffoni, 2013 ).叶脉密度作为叶脉网络功能性状的重要指标(Brodribb et al ., 2010 ; Blonder et al ., 2011 ), 可反映叶脉系统的水分、养分和光合产物等物质运输的效率和安全性以及抵抗外界干扰的能力, 是理解植物环境可塑性机制的重要途径(李乐等, 2013 ).气孔是植物叶片与外界进行水气交换的唯一可调节通道, 通过保卫细胞对环境和内源信号的感知, 与植物的光合作用、蒸腾作用等生理生态功能密切相关(杨惠敏和王根轩, 2001 ).叶脉与气孔是叶片水分供给和散失的重要因素, 两者的大小和数量的配置直接影响着叶片的水分利用效率, 进而影响叶片结构及生理功能的调节机制(Fiorin et al ., 2016 ).当植株处于高光强条件下时, 相对湿度下降, 叶片受到的水分胁迫增加, 叶片在获取光合收益的同时也需兼顾内部水分动态平衡(Fiorin et al ., 2016 ), 此时, 植株往往通过调整净光合速率与蒸腾速率来应对不利环境, 较高的蒸腾速率要求叶片发育较多的叶脉以提供供水系统, 在叶片的蒸腾作用中通过控制气孔密度和气孔大小决定进入大气的水分和CO2 , 降低植物的呼吸和蒸腾成本.叶脉密度与气孔性状共同作用体现了气孔蒸腾对水分的需求和叶脉系统对水分的供给相互匹配与权衡(Brodribb & Jordan, 2011 ).坡向作为重要的地形因素, 控制了太阳辐射和降水的空间再分配(Cartón et al ., 2004 ), 由坡向引起的土壤水分、光辐射、光照时长和大气温度等环境因子形成的生境梯度变化影响着群落组成(Bazzaz et al ., 1987 ; 韦莉莉等, 2005 ), 致使群落环境及其密度制约下的种群内植株的生态位存在差异, 群落内部郁闭度和干扰程度不同, 所接受的光合有效辐射也不同, 从而影响植物的空间分布格局及光照、温度等环境(党晶晶等, 2014 ; 杜晶等, 2016 ), 进而影响植物的形态.为了提高光合能力并保证水分供需间的平衡, 植株叶片会适当调整叶脉密度与气孔密度、气孔大小的生物量分配, 补充蒸腾带来的大量水分和能量的消耗(Hetherington & Woodward, 2003 ), 同时也使叶片中单位碳对光合作用的投资效益最大(Brodribb & Jordan, 2011 ), 从而提高生存适合度和竞争能力.因此, 研究微地形影响下叶脉密度与气孔性状的关系, 有助于探讨植物叶片内部水分供应与散失之间的平衡关系, 对植物生理生态功能的维持具有指导意义. ...
... ), 同时也使叶片中单位碳对光合作用的投资效益最大(Brodribb & Jordan, 2011 ), 从而提高生存适合度和竞争能力.因此, 研究微地形影响下叶脉密度与气孔性状的关系, 有助于探讨植物叶片内部水分供应与散失之间的平衡关系, 对植物生理生态功能的维持具有指导意义. ...
... 环境胁迫驱动下的自然选择使得植物倾向于以最小的水分散失来获得最大的碳同化速率(Hether- ington & Woodward, 2003 ), 植物通过调整自身生理特征和解剖结构来适应不同的生境条件(王瑞丽等, 2016 ).生长在南坡刺槐的叶脉密度与气孔密度呈极显著正相关关系(p < 0.01, 图1 ), 与气孔大小存在极显著负相关关系(p < 0.01, 图2 ).研究区位于北半球中纬度地区山地, 南坡太阳直射角较大, 光照时间长, 接受的光照充足, 总的辐射总量和温度均高于北坡(党晶晶等, 2014 ), 南坡植被稀疏, 接受的光合有效辐射最大(表2 ), 高温、强辐射造成刺槐叶片蒸腾加剧, 较强的蒸腾作用使植物面临着水分胁迫, 为了适应高温下的蒸腾速率, 植物倾向于发育小而厚的叶片(史元春等, 2015 ), 将更多的光合产物用于增加叶脉密度以保证高效率的水分输导系统.叶片在进行光合作用时不可避免地伴随着水分的蒸发损耗(Brodribb & Jordan, 2011 ), 在叶片增加运输管道时也应考虑到水分的散失, 气孔作为水分蒸腾的通道, 在土壤中的水分减少时植物供水能力降低(表1 ), 会影响气孔的发生、分化和发育等, 单位面积内叶脉密度的增加使叶脉间距变小, 气孔变窄、气孔密度加大(陈伟月等, 2014 ), 如果气孔不能感知这些变化并做出响应, 就提高了木质部栓塞的可能性, 进而增加了叶片液压功能失调至脱水的风险(Maseda & Fernández, 2006 ).当植株叶片获得的光合有效辐射较大时, 叶片气孔及表皮细胞迅速对光照做出反应, 直接向大气蒸发散水, 此时叶片其他部位并未发生水分亏缺, 小的气孔可以迅速关闭, 防止水分进一步流失, 避免水分亏缺在全叶中发生, 以此减少植物整叶枯萎的损失(高春娟等, 2012 ).因此, 生长在南坡的刺槐选择高的叶脉密度、气孔密度和小的气孔, 验证了Zhang (2012)的叶脉密度与气孔特征协同进化的理论. ...
Topographic controls on the spatial distribution of ground cover in the Tabernas badlands of SE Spain
2004
冬小麦品种(系)旗叶叶脉性状及其与气孔性状间的关联性. 生态学杂志,
1
2014
... 环境胁迫驱动下的自然选择使得植物倾向于以最小的水分散失来获得最大的碳同化速率(Hether- ington & Woodward, 2003 ), 植物通过调整自身生理特征和解剖结构来适应不同的生境条件(王瑞丽等, 2016 ).生长在南坡刺槐的叶脉密度与气孔密度呈极显著正相关关系(p < 0.01, 图1 ), 与气孔大小存在极显著负相关关系(p < 0.01, 图2 ).研究区位于北半球中纬度地区山地, 南坡太阳直射角较大, 光照时间长, 接受的光照充足, 总的辐射总量和温度均高于北坡(党晶晶等, 2014 ), 南坡植被稀疏, 接受的光合有效辐射最大(表2 ), 高温、强辐射造成刺槐叶片蒸腾加剧, 较强的蒸腾作用使植物面临着水分胁迫, 为了适应高温下的蒸腾速率, 植物倾向于发育小而厚的叶片(史元春等, 2015 ), 将更多的光合产物用于增加叶脉密度以保证高效率的水分输导系统.叶片在进行光合作用时不可避免地伴随着水分的蒸发损耗(Brodribb & Jordan, 2011 ), 在叶片增加运输管道时也应考虑到水分的散失, 气孔作为水分蒸腾的通道, 在土壤中的水分减少时植物供水能力降低(表1 ), 会影响气孔的发生、分化和发育等, 单位面积内叶脉密度的增加使叶脉间距变小, 气孔变窄、气孔密度加大(陈伟月等, 2014 ), 如果气孔不能感知这些变化并做出响应, 就提高了木质部栓塞的可能性, 进而增加了叶片液压功能失调至脱水的风险(Maseda & Fernández, 2006 ).当植株叶片获得的光合有效辐射较大时, 叶片气孔及表皮细胞迅速对光照做出反应, 直接向大气蒸发散水, 此时叶片其他部位并未发生水分亏缺, 小的气孔可以迅速关闭, 防止水分进一步流失, 避免水分亏缺在全叶中发生, 以此减少植物整叶枯萎的损失(高春娟等, 2012 ).因此, 生长在南坡的刺槐选择高的叶脉密度、气孔密度和小的气孔, 验证了Zhang (2012)的叶脉密度与气孔特征协同进化的理论. ...
四川龙门山西北部植被分布与地形因子的相关性. 生态学杂志,
1
2006
... 对研究区1:50000地形图利用ArcGIS进行数字化分析, 得到栅格大小为25 m的数字高程模型(digital elevation model, DEM).然后在DEM基础上利用Spatial Analyst模块中的Surface Analysis命令提取坡向, 其中坡向采用3次有限差分方法计算得到(Skidmore, 1989 ), 表示地表面上一点的切平面的法线矢量n 在水平面的投影n xoy 与过该点的正北方向的夹角, 最后利用提取出的坡向指标建立每块样地的数据查找表, 其中坡向组别以国际划分法(陈瑶等, 2006 )确定(圆周角属于315°-360°和0-45°定为正北坡向). ...
高寒草地甘肃臭草茎-叶性状的坡度差异性. 植物生态学报,
2
2014
... 作为植物碳水耦合权衡的重要器官, 叶片在进化过程中对环境的变化较敏感且可塑性较大(Scoffoni et al ., 2015 ), 从叶片形态解剖结构了解植物生态策略在异质生境中的变化对植物的结构和生理变化具有重要意义(Maricle et al ., 2009 ).叶脉是影响叶片水分供应和利用的重要结构, 在一定意义上反映了叶片资源流通能力(Sack & Frole, 2006 ; Sack & Scoffoni, 2013 ).叶脉密度作为叶脉网络功能性状的重要指标(Brodribb et al ., 2010 ; Blonder et al ., 2011 ), 可反映叶脉系统的水分、养分和光合产物等物质运输的效率和安全性以及抵抗外界干扰的能力, 是理解植物环境可塑性机制的重要途径(李乐等, 2013 ).气孔是植物叶片与外界进行水气交换的唯一可调节通道, 通过保卫细胞对环境和内源信号的感知, 与植物的光合作用、蒸腾作用等生理生态功能密切相关(杨惠敏和王根轩, 2001 ).叶脉与气孔是叶片水分供给和散失的重要因素, 两者的大小和数量的配置直接影响着叶片的水分利用效率, 进而影响叶片结构及生理功能的调节机制(Fiorin et al ., 2016 ).当植株处于高光强条件下时, 相对湿度下降, 叶片受到的水分胁迫增加, 叶片在获取光合收益的同时也需兼顾内部水分动态平衡(Fiorin et al ., 2016 ), 此时, 植株往往通过调整净光合速率与蒸腾速率来应对不利环境, 较高的蒸腾速率要求叶片发育较多的叶脉以提供供水系统, 在叶片的蒸腾作用中通过控制气孔密度和气孔大小决定进入大气的水分和CO2 , 降低植物的呼吸和蒸腾成本.叶脉密度与气孔性状共同作用体现了气孔蒸腾对水分的需求和叶脉系统对水分的供给相互匹配与权衡(Brodribb & Jordan, 2011 ).坡向作为重要的地形因素, 控制了太阳辐射和降水的空间再分配(Cartón et al ., 2004 ), 由坡向引起的土壤水分、光辐射、光照时长和大气温度等环境因子形成的生境梯度变化影响着群落组成(Bazzaz et al ., 1987 ; 韦莉莉等, 2005 ), 致使群落环境及其密度制约下的种群内植株的生态位存在差异, 群落内部郁闭度和干扰程度不同, 所接受的光合有效辐射也不同, 从而影响植物的空间分布格局及光照、温度等环境(党晶晶等, 2014 ; 杜晶等, 2016 ), 进而影响植物的形态.为了提高光合能力并保证水分供需间的平衡, 植株叶片会适当调整叶脉密度与气孔密度、气孔大小的生物量分配, 补充蒸腾带来的大量水分和能量的消耗(Hetherington & Woodward, 2003 ), 同时也使叶片中单位碳对光合作用的投资效益最大(Brodribb & Jordan, 2011 ), 从而提高生存适合度和竞争能力.因此, 研究微地形影响下叶脉密度与气孔性状的关系, 有助于探讨植物叶片内部水分供应与散失之间的平衡关系, 对植物生理生态功能的维持具有指导意义. ...
... 环境胁迫驱动下的自然选择使得植物倾向于以最小的水分散失来获得最大的碳同化速率(Hether- ington & Woodward, 2003 ), 植物通过调整自身生理特征和解剖结构来适应不同的生境条件(王瑞丽等, 2016 ).生长在南坡刺槐的叶脉密度与气孔密度呈极显著正相关关系(p < 0.01, 图1 ), 与气孔大小存在极显著负相关关系(p < 0.01, 图2 ).研究区位于北半球中纬度地区山地, 南坡太阳直射角较大, 光照时间长, 接受的光照充足, 总的辐射总量和温度均高于北坡(党晶晶等, 2014 ), 南坡植被稀疏, 接受的光合有效辐射最大(表2 ), 高温、强辐射造成刺槐叶片蒸腾加剧, 较强的蒸腾作用使植物面临着水分胁迫, 为了适应高温下的蒸腾速率, 植物倾向于发育小而厚的叶片(史元春等, 2015 ), 将更多的光合产物用于增加叶脉密度以保证高效率的水分输导系统.叶片在进行光合作用时不可避免地伴随着水分的蒸发损耗(Brodribb & Jordan, 2011 ), 在叶片增加运输管道时也应考虑到水分的散失, 气孔作为水分蒸腾的通道, 在土壤中的水分减少时植物供水能力降低(表1 ), 会影响气孔的发生、分化和发育等, 单位面积内叶脉密度的增加使叶脉间距变小, 气孔变窄、气孔密度加大(陈伟月等, 2014 ), 如果气孔不能感知这些变化并做出响应, 就提高了木质部栓塞的可能性, 进而增加了叶片液压功能失调至脱水的风险(Maseda & Fernández, 2006 ).当植株叶片获得的光合有效辐射较大时, 叶片气孔及表皮细胞迅速对光照做出反应, 直接向大气蒸发散水, 此时叶片其他部位并未发生水分亏缺, 小的气孔可以迅速关闭, 防止水分进一步流失, 避免水分亏缺在全叶中发生, 以此减少植物整叶枯萎的损失(高春娟等, 2012 ).因此, 生长在南坡的刺槐选择高的叶脉密度、气孔密度和小的气孔, 验证了Zhang (2012)的叶脉密度与气孔特征协同进化的理论. ...
The physiological importance of developmental mechanisms that enforce proper stomatal spacing in
2014
祁连山北坡霸王枝-叶性状关系的个体大小差异. 植物生态学报,
1
2016
... 作为植物碳水耦合权衡的重要器官, 叶片在进化过程中对环境的变化较敏感且可塑性较大(Scoffoni et al ., 2015 ), 从叶片形态解剖结构了解植物生态策略在异质生境中的变化对植物的结构和生理变化具有重要意义(Maricle et al ., 2009 ).叶脉是影响叶片水分供应和利用的重要结构, 在一定意义上反映了叶片资源流通能力(Sack & Frole, 2006 ; Sack & Scoffoni, 2013 ).叶脉密度作为叶脉网络功能性状的重要指标(Brodribb et al ., 2010 ; Blonder et al ., 2011 ), 可反映叶脉系统的水分、养分和光合产物等物质运输的效率和安全性以及抵抗外界干扰的能力, 是理解植物环境可塑性机制的重要途径(李乐等, 2013 ).气孔是植物叶片与外界进行水气交换的唯一可调节通道, 通过保卫细胞对环境和内源信号的感知, 与植物的光合作用、蒸腾作用等生理生态功能密切相关(杨惠敏和王根轩, 2001 ).叶脉与气孔是叶片水分供给和散失的重要因素, 两者的大小和数量的配置直接影响着叶片的水分利用效率, 进而影响叶片结构及生理功能的调节机制(Fiorin et al ., 2016 ).当植株处于高光强条件下时, 相对湿度下降, 叶片受到的水分胁迫增加, 叶片在获取光合收益的同时也需兼顾内部水分动态平衡(Fiorin et al ., 2016 ), 此时, 植株往往通过调整净光合速率与蒸腾速率来应对不利环境, 较高的蒸腾速率要求叶片发育较多的叶脉以提供供水系统, 在叶片的蒸腾作用中通过控制气孔密度和气孔大小决定进入大气的水分和CO2 , 降低植物的呼吸和蒸腾成本.叶脉密度与气孔性状共同作用体现了气孔蒸腾对水分的需求和叶脉系统对水分的供给相互匹配与权衡(Brodribb & Jordan, 2011 ).坡向作为重要的地形因素, 控制了太阳辐射和降水的空间再分配(Cartón et al ., 2004 ), 由坡向引起的土壤水分、光辐射、光照时长和大气温度等环境因子形成的生境梯度变化影响着群落组成(Bazzaz et al ., 1987 ; 韦莉莉等, 2005 ), 致使群落环境及其密度制约下的种群内植株的生态位存在差异, 群落内部郁闭度和干扰程度不同, 所接受的光合有效辐射也不同, 从而影响植物的空间分布格局及光照、温度等环境(党晶晶等, 2014 ; 杜晶等, 2016 ), 进而影响植物的形态.为了提高光合能力并保证水分供需间的平衡, 植株叶片会适当调整叶脉密度与气孔密度、气孔大小的生物量分配, 补充蒸腾带来的大量水分和能量的消耗(Hetherington & Woodward, 2003 ), 同时也使叶片中单位碳对光合作用的投资效益最大(Brodribb & Jordan, 2011 ), 从而提高生存适合度和竞争能力.因此, 研究微地形影响下叶脉密度与气孔性状的关系, 有助于探讨植物叶片内部水分供应与散失之间的平衡关系, 对植物生理生态功能的维持具有指导意义. ...
1
... 植物叶片构型构建与植物生理功能所需的光照、碳投入和水分运输的投资收益相契合, 是植物提高光合收益的一个重要的生活史策略(Westoby et al ., 2002 ; 孙素静等, 2015 ).生长在北坡的刺槐的叶脉密度与气孔密度呈显著正相关关系(p < 0.05, 图1 ), 与气孔大小存在显著负相关关系(p < 0.05, 图2 ).随着资源环境的改变, 北坡植被群落郁闭程度和拥挤程度达到最大, 来自邻株植物的遮阴改变了植物所处光环境, 限制了叶片可获得性光资源, 光合有效辐射减弱(表2 ).植株为了获得更多的光资源, 选择了增大叶面积的生长策略, 使得部分叶肉组织与叶脉接触面积减少, 相对减少叶脉构建投资; 另一方面, 在土壤含水量较充沛的条件下, 植物供水较充足, 气孔张合度受到影响, 单位叶面积内气孔数量减少且CO2 扩散阻力增加, 叶片吸收CO2 受到限制, 光合速率显著降低, 引起植物地上部分对水的需求减少, 使得植物不必投资更多叶脉导管就可以满足叶片水分的供应, 有利于提高植物在资源丰富的环境中的竞争力(Beerling & Franks, 2010 ; Field et al ., 2011 ).气孔直接影响着植物的CO2 交换速率、蒸腾速率和光合速率, 调节气孔密度和气孔长度可适应时刻发生变化的环境状况, 使植物在水碳循环中的平衡调节作用达到最优(高春娟等, 2012 ; 高冠龙等, 2016 ), 当土壤水分较充足时, 根系吸收的水分足以保持体内水分平衡, 气孔进一步开放虽然使得蒸腾速率增加, 但气孔对蒸腾速率的敏感度降低, 吸水速率与蒸腾速率的平衡机制对气孔反馈减少(左应梅等, 2011 ), 采取了“疏而大”的适应特征, 从而保证叶片能吸收足量的CO2 , 进行正常的光合作用, 这与Dow等(2014)对拟南芥(Arabidopsis thaliana )气孔的研究结果一致.刺槐在北坡选择了降低叶脉密度与气孔密度、增大气孔的策略, 使叶含水量维持在动态平衡范围内, 这对于理解植物对环境的适应策略、叶性状之间资源的分配与利用具有重要意义. ...
Fossil evidence for Cretaceous escalation in angiosperm leaf vein evolution
2
2011
... 作为植物碳水耦合权衡的重要器官, 叶片在进化过程中对环境的变化较敏感且可塑性较大(Scoffoni et al ., 2015 ), 从叶片形态解剖结构了解植物生态策略在异质生境中的变化对植物的结构和生理变化具有重要意义(Maricle et al ., 2009 ).叶脉是影响叶片水分供应和利用的重要结构, 在一定意义上反映了叶片资源流通能力(Sack & Frole, 2006 ; Sack & Scoffoni, 2013 ).叶脉密度作为叶脉网络功能性状的重要指标(Brodribb et al ., 2010 ; Blonder et al ., 2011 ), 可反映叶脉系统的水分、养分和光合产物等物质运输的效率和安全性以及抵抗外界干扰的能力, 是理解植物环境可塑性机制的重要途径(李乐等, 2013 ).气孔是植物叶片与外界进行水气交换的唯一可调节通道, 通过保卫细胞对环境和内源信号的感知, 与植物的光合作用、蒸腾作用等生理生态功能密切相关(杨惠敏和王根轩, 2001 ).叶脉与气孔是叶片水分供给和散失的重要因素, 两者的大小和数量的配置直接影响着叶片的水分利用效率, 进而影响叶片结构及生理功能的调节机制(Fiorin et al ., 2016 ).当植株处于高光强条件下时, 相对湿度下降, 叶片受到的水分胁迫增加, 叶片在获取光合收益的同时也需兼顾内部水分动态平衡(Fiorin et al ., 2016 ), 此时, 植株往往通过调整净光合速率与蒸腾速率来应对不利环境, 较高的蒸腾速率要求叶片发育较多的叶脉以提供供水系统, 在叶片的蒸腾作用中通过控制气孔密度和气孔大小决定进入大气的水分和CO2 , 降低植物的呼吸和蒸腾成本.叶脉密度与气孔性状共同作用体现了气孔蒸腾对水分的需求和叶脉系统对水分的供给相互匹配与权衡(Brodribb & Jordan, 2011 ).坡向作为重要的地形因素, 控制了太阳辐射和降水的空间再分配(Cartón et al ., 2004 ), 由坡向引起的土壤水分、光辐射、光照时长和大气温度等环境因子形成的生境梯度变化影响着群落组成(Bazzaz et al ., 1987 ; 韦莉莉等, 2005 ), 致使群落环境及其密度制约下的种群内植株的生态位存在差异, 群落内部郁闭度和干扰程度不同, 所接受的光合有效辐射也不同, 从而影响植物的空间分布格局及光照、温度等环境(党晶晶等, 2014 ; 杜晶等, 2016 ), 进而影响植物的形态.为了提高光合能力并保证水分供需间的平衡, 植株叶片会适当调整叶脉密度与气孔密度、气孔大小的生物量分配, 补充蒸腾带来的大量水分和能量的消耗(Hetherington & Woodward, 2003 ), 同时也使叶片中单位碳对光合作用的投资效益最大(Brodribb & Jordan, 2011 ), 从而提高生存适合度和竞争能力.因此, 研究微地形影响下叶脉密度与气孔性状的关系, 有助于探讨植物叶片内部水分供应与散失之间的平衡关系, 对植物生理生态功能的维持具有指导意义. ...
... ).当植株处于高光强条件下时, 相对湿度下降, 叶片受到的水分胁迫增加, 叶片在获取光合收益的同时也需兼顾内部水分动态平衡(Fiorin et al ., 2016 ), 此时, 植株往往通过调整净光合速率与蒸腾速率来应对不利环境, 较高的蒸腾速率要求叶片发育较多的叶脉以提供供水系统, 在叶片的蒸腾作用中通过控制气孔密度和气孔大小决定进入大气的水分和CO2 , 降低植物的呼吸和蒸腾成本.叶脉密度与气孔性状共同作用体现了气孔蒸腾对水分的需求和叶脉系统对水分的供给相互匹配与权衡(Brodribb & Jordan, 2011 ).坡向作为重要的地形因素, 控制了太阳辐射和降水的空间再分配(Cartón et al ., 2004 ), 由坡向引起的土壤水分、光辐射、光照时长和大气温度等环境因子形成的生境梯度变化影响着群落组成(Bazzaz et al ., 1987 ; 韦莉莉等, 2005 ), 致使群落环境及其密度制约下的种群内植株的生态位存在差异, 群落内部郁闭度和干扰程度不同, 所接受的光合有效辐射也不同, 从而影响植物的空间分布格局及光照、温度等环境(党晶晶等, 2014 ; 杜晶等, 2016 ), 进而影响植物的形态.为了提高光合能力并保证水分供需间的平衡, 植株叶片会适当调整叶脉密度与气孔密度、气孔大小的生物量分配, 补充蒸腾带来的大量水分和能量的消耗(Hetherington & Woodward, 2003 ), 同时也使叶片中单位碳对光合作用的投资效益最大(Brodribb & Jordan, 2011 ), 从而提高生存适合度和竞争能力.因此, 研究微地形影响下叶脉密度与气孔性状的关系, 有助于探讨植物叶片内部水分供应与散失之间的平衡关系, 对植物生理生态功能的维持具有指导意义. ...
Transport efficiency through uniformity: Organization of veins and stomata in angiosperm leaves
2
2016
... 环境胁迫驱动下的自然选择使得植物倾向于以最小的水分散失来获得最大的碳同化速率(Hether- ington & Woodward, 2003 ), 植物通过调整自身生理特征和解剖结构来适应不同的生境条件(王瑞丽等, 2016 ).生长在南坡刺槐的叶脉密度与气孔密度呈极显著正相关关系(p < 0.01, 图1 ), 与气孔大小存在极显著负相关关系(p < 0.01, 图2 ).研究区位于北半球中纬度地区山地, 南坡太阳直射角较大, 光照时间长, 接受的光照充足, 总的辐射总量和温度均高于北坡(党晶晶等, 2014 ), 南坡植被稀疏, 接受的光合有效辐射最大(表2 ), 高温、强辐射造成刺槐叶片蒸腾加剧, 较强的蒸腾作用使植物面临着水分胁迫, 为了适应高温下的蒸腾速率, 植物倾向于发育小而厚的叶片(史元春等, 2015 ), 将更多的光合产物用于增加叶脉密度以保证高效率的水分输导系统.叶片在进行光合作用时不可避免地伴随着水分的蒸发损耗(Brodribb & Jordan, 2011 ), 在叶片增加运输管道时也应考虑到水分的散失, 气孔作为水分蒸腾的通道, 在土壤中的水分减少时植物供水能力降低(表1 ), 会影响气孔的发生、分化和发育等, 单位面积内叶脉密度的增加使叶脉间距变小, 气孔变窄、气孔密度加大(陈伟月等, 2014 ), 如果气孔不能感知这些变化并做出响应, 就提高了木质部栓塞的可能性, 进而增加了叶片液压功能失调至脱水的风险(Maseda & Fernández, 2006 ).当植株叶片获得的光合有效辐射较大时, 叶片气孔及表皮细胞迅速对光照做出反应, 直接向大气蒸发散水, 此时叶片其他部位并未发生水分亏缺, 小的气孔可以迅速关闭, 防止水分进一步流失, 避免水分亏缺在全叶中发生, 以此减少植物整叶枯萎的损失(高春娟等, 2012 ).因此, 生长在南坡的刺槐选择高的叶脉密度、气孔密度和小的气孔, 验证了Zhang (2012)的叶脉密度与气孔特征协同进化的理论. ...
... 植物叶片构型构建与植物生理功能所需的光照、碳投入和水分运输的投资收益相契合, 是植物提高光合收益的一个重要的生活史策略(Westoby et al ., 2002 ; 孙素静等, 2015 ).生长在北坡的刺槐的叶脉密度与气孔密度呈显著正相关关系(p < 0.05, 图1 ), 与气孔大小存在显著负相关关系(p < 0.05, 图2 ).随着资源环境的改变, 北坡植被群落郁闭程度和拥挤程度达到最大, 来自邻株植物的遮阴改变了植物所处光环境, 限制了叶片可获得性光资源, 光合有效辐射减弱(表2 ).植株为了获得更多的光资源, 选择了增大叶面积的生长策略, 使得部分叶肉组织与叶脉接触面积减少, 相对减少叶脉构建投资; 另一方面, 在土壤含水量较充沛的条件下, 植物供水较充足, 气孔张合度受到影响, 单位叶面积内气孔数量减少且CO2 扩散阻力增加, 叶片吸收CO2 受到限制, 光合速率显著降低, 引起植物地上部分对水的需求减少, 使得植物不必投资更多叶脉导管就可以满足叶片水分的供应, 有利于提高植物在资源丰富的环境中的竞争力(Beerling & Franks, 2010 ; Field et al ., 2011 ).气孔直接影响着植物的CO2 交换速率、蒸腾速率和光合速率, 调节气孔密度和气孔长度可适应时刻发生变化的环境状况, 使植物在水碳循环中的平衡调节作用达到最优(高春娟等, 2012 ; 高冠龙等, 2016 ), 当土壤水分较充足时, 根系吸收的水分足以保持体内水分平衡, 气孔进一步开放虽然使得蒸腾速率增加, 但气孔对蒸腾速率的敏感度降低, 吸水速率与蒸腾速率的平衡机制对气孔反馈减少(左应梅等, 2011 ), 采取了“疏而大”的适应特征, 从而保证叶片能吸收足量的CO2 , 进行正常的光合作用, 这与Dow等(2014)对拟南芥(Arabidopsis thaliana )气孔的研究结果一致.刺槐在北坡选择了降低叶脉密度与气孔密度、增大气孔的策略, 使叶含水量维持在动态平衡范围内, 这对于理解植物对环境的适应策略、叶性状之间资源的分配与利用具有重要意义. ...
植物气孔对全球环境变化的响应及其调控防御机制. 植物生理学报,
1
2012
... 植物叶片构型构建与植物生理功能所需的光照、碳投入和水分运输的投资收益相契合, 是植物提高光合收益的一个重要的生活史策略(Westoby et al ., 2002 ; 孙素静等, 2015 ).生长在北坡的刺槐的叶脉密度与气孔密度呈显著正相关关系(p < 0.05, 图1 ), 与气孔大小存在显著负相关关系(p < 0.05, 图2 ).随着资源环境的改变, 北坡植被群落郁闭程度和拥挤程度达到最大, 来自邻株植物的遮阴改变了植物所处光环境, 限制了叶片可获得性光资源, 光合有效辐射减弱(表2 ).植株为了获得更多的光资源, 选择了增大叶面积的生长策略, 使得部分叶肉组织与叶脉接触面积减少, 相对减少叶脉构建投资; 另一方面, 在土壤含水量较充沛的条件下, 植物供水较充足, 气孔张合度受到影响, 单位叶面积内气孔数量减少且CO2 扩散阻力增加, 叶片吸收CO2 受到限制, 光合速率显著降低, 引起植物地上部分对水的需求减少, 使得植物不必投资更多叶脉导管就可以满足叶片水分的供应, 有利于提高植物在资源丰富的环境中的竞争力(Beerling & Franks, 2010 ; Field et al ., 2011 ).气孔直接影响着植物的CO2 交换速率、蒸腾速率和光合速率, 调节气孔密度和气孔长度可适应时刻发生变化的环境状况, 使植物在水碳循环中的平衡调节作用达到最优(高春娟等, 2012 ; 高冠龙等, 2016 ), 当土壤水分较充足时, 根系吸收的水分足以保持体内水分平衡, 气孔进一步开放虽然使得蒸腾速率增加, 但气孔对蒸腾速率的敏感度降低, 吸水速率与蒸腾速率的平衡机制对气孔反馈减少(左应梅等, 2011 ), 采取了“疏而大”的适应特征, 从而保证叶片能吸收足量的CO2 , 进行正常的光合作用, 这与Dow等(2014)对拟南芥(Arabidopsis thaliana )气孔的研究结果一致.刺槐在北坡选择了降低叶脉密度与气孔密度、增大气孔的策略, 使叶含水量维持在动态平衡范围内, 这对于理解植物对环境的适应策略、叶性状之间资源的分配与利用具有重要意义. ...
植物气孔导度的环境响应模拟及其尺度扩展. 生态学报,
1
2016
... 刺槐(Robinia pseudoacacia )是温带落叶乔木, 是我国华北和西北地区绿化荒山和防风固沙的主要植物, 具有固氮能力强和碳储存量大的优势, 作为黄土丘陵区的主要人工造林树种, 其根系发达、生长迅速、成活率高和环境适应性强等特点在保持水土和生态防护中发挥着重要作用.近年来, 关于刺槐在干旱半干旱地区的种植范围和生态适应性等问题引起了学界的广泛关注(于占辉等, 2009 ; 李军等, 2010 ; 申家朋和张文辉, 2014 ).学术界对气孔性状之间的关系(杨惠敏和王根轩, 2001 ; 王曙光等, 2013 )、气孔与环境因子的关系(左应梅等, 2011 ; 王娜等, 2016 )、气孔性状对水分利用的影响(高洁等, 2004 ; 杨利民等, 2007 )、叶脉网络功能性状与叶片光合作用(Brodribb et al ., 2010 ; Sack et al ., 2012 )、水分利用能力(Zhang et al ., 2012 ; Sack & Scoffoni, 2013 )等问题进行了研究, 阐述了植物叶片在适应特定生境中各种性状的塑造, 而将叶脉与气孔性状结合起来, 探讨二者在异质性生境中叶片内部水分传输及水分平衡适应策略的分析和认识目前还比较薄弱.鉴于此, 本文以兰州北山的人工刺槐林为研究对象, 试图揭示以下两个问题: (1)不同坡向上刺槐叶脉密度与气孔密度、气孔大小之间存在何种关系?(2)这种关系的形成原因有哪些?理解异质生境中植物叶片的物理构建方式, 可为提高半干旱人工林资源的利用率和绿化覆盖率提供理论依据. ...
干热河谷9种造林树种在旱季的水分关系和气孔导度. 植物生态学报,
2004
The role of stomata in sensing and driving environmental change
1
2003
... 植物的光合作用是生态系统生产力形成与演化的基础, 叶片作为光合产物形成的主要器官, 既是整株植物的水力瓶颈, 又是植株应对水力失调的重要枢纽, 在很大程度上影响着植物水分和养分吸收、运移和储存功能(金鹰和王传宽, 2015 ).在长期的环境选择进化中, 植物对水分的需求驱动着叶脉网络的变化, 气孔是植物体蒸腾失水的门户(左应梅等, 2011 ), 植物叶片选择发育更有效的叶脉灌输系统和密集的气孔提高光合效率和水分利用效率(张亚等, 2014 ), 对植物生理功能的维持具有重要意义.本研究发现, 各个坡向刺槐的叶脉密度与气孔密度呈正相关关系, 与气孔大小呈负相关关系, 且在南坡达到极显著水平(p < 0.01, 图1 , 图2 ), 表明坡向梯度上刺槐叶片结构不同功能之间的相互匹配与权衡关系是植物叶片的形态结构和特定的水分供应需求分配策略对生境异质性适应的结果. ...
植物叶片水力与经济性状权衡关系的研究进展. 植物生态学报,
1
2015
... 刺槐(Robinia pseudoacacia )是温带落叶乔木, 是我国华北和西北地区绿化荒山和防风固沙的主要植物, 具有固氮能力强和碳储存量大的优势, 作为黄土丘陵区的主要人工造林树种, 其根系发达、生长迅速、成活率高和环境适应性强等特点在保持水土和生态防护中发挥着重要作用.近年来, 关于刺槐在干旱半干旱地区的种植范围和生态适应性等问题引起了学界的广泛关注(于占辉等, 2009 ; 李军等, 2010 ; 申家朋和张文辉, 2014 ).学术界对气孔性状之间的关系(杨惠敏和王根轩, 2001 ; 王曙光等, 2013 )、气孔与环境因子的关系(左应梅等, 2011 ; 王娜等, 2016 )、气孔性状对水分利用的影响(高洁等, 2004 ; 杨利民等, 2007 )、叶脉网络功能性状与叶片光合作用(Brodribb et al ., 2010 ; Sack et al ., 2012 )、水分利用能力(Zhang et al ., 2012 ; Sack & Scoffoni, 2013 )等问题进行了研究, 阐述了植物叶片在适应特定生境中各种性状的塑造, 而将叶脉与气孔性状结合起来, 探讨二者在异质性生境中叶片内部水分传输及水分平衡适应策略的分析和认识目前还比较薄弱.鉴于此, 本文以兰州北山的人工刺槐林为研究对象, 试图揭示以下两个问题: (1)不同坡向上刺槐叶脉密度与气孔密度、气孔大小之间存在何种关系?(2)这种关系的形成原因有哪些?理解异质生境中植物叶片的物理构建方式, 可为提高半干旱人工林资源的利用率和绿化覆盖率提供理论依据. ...
黄土高原半干旱和半湿润地区刺槐林地生物量与土壤干燥化效应的模拟. 植物生态学报,
1
2010
... 作为植物碳水耦合权衡的重要器官, 叶片在进化过程中对环境的变化较敏感且可塑性较大(Scoffoni et al ., 2015 ), 从叶片形态解剖结构了解植物生态策略在异质生境中的变化对植物的结构和生理变化具有重要意义(Maricle et al ., 2009 ).叶脉是影响叶片水分供应和利用的重要结构, 在一定意义上反映了叶片资源流通能力(Sack & Frole, 2006 ; Sack & Scoffoni, 2013 ).叶脉密度作为叶脉网络功能性状的重要指标(Brodribb et al ., 2010 ; Blonder et al ., 2011 ), 可反映叶脉系统的水分、养分和光合产物等物质运输的效率和安全性以及抵抗外界干扰的能力, 是理解植物环境可塑性机制的重要途径(李乐等, 2013 ).气孔是植物叶片与外界进行水气交换的唯一可调节通道, 通过保卫细胞对环境和内源信号的感知, 与植物的光合作用、蒸腾作用等生理生态功能密切相关(杨惠敏和王根轩, 2001 ).叶脉与气孔是叶片水分供给和散失的重要因素, 两者的大小和数量的配置直接影响着叶片的水分利用效率, 进而影响叶片结构及生理功能的调节机制(Fiorin et al ., 2016 ).当植株处于高光强条件下时, 相对湿度下降, 叶片受到的水分胁迫增加, 叶片在获取光合收益的同时也需兼顾内部水分动态平衡(Fiorin et al ., 2016 ), 此时, 植株往往通过调整净光合速率与蒸腾速率来应对不利环境, 较高的蒸腾速率要求叶片发育较多的叶脉以提供供水系统, 在叶片的蒸腾作用中通过控制气孔密度和气孔大小决定进入大气的水分和CO2 , 降低植物的呼吸和蒸腾成本.叶脉密度与气孔性状共同作用体现了气孔蒸腾对水分的需求和叶脉系统对水分的供给相互匹配与权衡(Brodribb & Jordan, 2011 ).坡向作为重要的地形因素, 控制了太阳辐射和降水的空间再分配(Cartón et al ., 2004 ), 由坡向引起的土壤水分、光辐射、光照时长和大气温度等环境因子形成的生境梯度变化影响着群落组成(Bazzaz et al ., 1987 ; 韦莉莉等, 2005 ), 致使群落环境及其密度制约下的种群内植株的生态位存在差异, 群落内部郁闭度和干扰程度不同, 所接受的光合有效辐射也不同, 从而影响植物的空间分布格局及光照、温度等环境(党晶晶等, 2014 ; 杜晶等, 2016 ), 进而影响植物的形态.为了提高光合能力并保证水分供需间的平衡, 植株叶片会适当调整叶脉密度与气孔密度、气孔大小的生物量分配, 补充蒸腾带来的大量水分和能量的消耗(Hetherington & Woodward, 2003 ), 同时也使叶片中单位碳对光合作用的投资效益最大(Brodribb & Jordan, 2011 ), 从而提高生存适合度和竞争能力.因此, 研究微地形影响下叶脉密度与气孔性状的关系, 有助于探讨植物叶片内部水分供应与散失之间的平衡关系, 对植物生理生态功能的维持具有指导意义. ...
叶脉网络功能性状及其生态学意义. 植物生态学报,
2013
Influence of climate-driven shifts in biomass allocation on water transport and storage in ponderosa pine
1
2001
... 作为植物碳水耦合权衡的重要器官, 叶片在进化过程中对环境的变化较敏感且可塑性较大(Scoffoni et al ., 2015 ), 从叶片形态解剖结构了解植物生态策略在异质生境中的变化对植物的结构和生理变化具有重要意义(Maricle et al ., 2009 ).叶脉是影响叶片水分供应和利用的重要结构, 在一定意义上反映了叶片资源流通能力(Sack & Frole, 2006 ; Sack & Scoffoni, 2013 ).叶脉密度作为叶脉网络功能性状的重要指标(Brodribb et al ., 2010 ; Blonder et al ., 2011 ), 可反映叶脉系统的水分、养分和光合产物等物质运输的效率和安全性以及抵抗外界干扰的能力, 是理解植物环境可塑性机制的重要途径(李乐等, 2013 ).气孔是植物叶片与外界进行水气交换的唯一可调节通道, 通过保卫细胞对环境和内源信号的感知, 与植物的光合作用、蒸腾作用等生理生态功能密切相关(杨惠敏和王根轩, 2001 ).叶脉与气孔是叶片水分供给和散失的重要因素, 两者的大小和数量的配置直接影响着叶片的水分利用效率, 进而影响叶片结构及生理功能的调节机制(Fiorin et al ., 2016 ).当植株处于高光强条件下时, 相对湿度下降, 叶片受到的水分胁迫增加, 叶片在获取光合收益的同时也需兼顾内部水分动态平衡(Fiorin et al ., 2016 ), 此时, 植株往往通过调整净光合速率与蒸腾速率来应对不利环境, 较高的蒸腾速率要求叶片发育较多的叶脉以提供供水系统, 在叶片的蒸腾作用中通过控制气孔密度和气孔大小决定进入大气的水分和CO2 , 降低植物的呼吸和蒸腾成本.叶脉密度与气孔性状共同作用体现了气孔蒸腾对水分的需求和叶脉系统对水分的供给相互匹配与权衡(Brodribb & Jordan, 2011 ).坡向作为重要的地形因素, 控制了太阳辐射和降水的空间再分配(Cartón et al ., 2004 ), 由坡向引起的土壤水分、光辐射、光照时长和大气温度等环境因子形成的生境梯度变化影响着群落组成(Bazzaz et al ., 1987 ; 韦莉莉等, 2005 ), 致使群落环境及其密度制约下的种群内植株的生态位存在差异, 群落内部郁闭度和干扰程度不同, 所接受的光合有效辐射也不同, 从而影响植物的空间分布格局及光照、温度等环境(党晶晶等, 2014 ; 杜晶等, 2016 ), 进而影响植物的形态.为了提高光合能力并保证水分供需间的平衡, 植株叶片会适当调整叶脉密度与气孔密度、气孔大小的生物量分配, 补充蒸腾带来的大量水分和能量的消耗(Hetherington & Woodward, 2003 ), 同时也使叶片中单位碳对光合作用的投资效益最大(Brodribb & Jordan, 2011 ), 从而提高生存适合度和竞争能力.因此, 研究微地形影响下叶脉密度与气孔性状的关系, 有助于探讨植物叶片内部水分供应与散失之间的平衡关系, 对植物生理生态功能的维持具有指导意义. ...
Diversity in leaf anatomy, and stomatal distribution and conductance, between salt marsh and freshwater species in the C4 genus
1
2009
... 环境胁迫驱动下的自然选择使得植物倾向于以最小的水分散失来获得最大的碳同化速率(Hether- ington & Woodward, 2003 ), 植物通过调整自身生理特征和解剖结构来适应不同的生境条件(王瑞丽等, 2016 ).生长在南坡刺槐的叶脉密度与气孔密度呈极显著正相关关系(p < 0.01, 图1 ), 与气孔大小存在极显著负相关关系(p < 0.01, 图2 ).研究区位于北半球中纬度地区山地, 南坡太阳直射角较大, 光照时间长, 接受的光照充足, 总的辐射总量和温度均高于北坡(党晶晶等, 2014 ), 南坡植被稀疏, 接受的光合有效辐射最大(表2 ), 高温、强辐射造成刺槐叶片蒸腾加剧, 较强的蒸腾作用使植物面临着水分胁迫, 为了适应高温下的蒸腾速率, 植物倾向于发育小而厚的叶片(史元春等, 2015 ), 将更多的光合产物用于增加叶脉密度以保证高效率的水分输导系统.叶片在进行光合作用时不可避免地伴随着水分的蒸发损耗(Brodribb & Jordan, 2011 ), 在叶片增加运输管道时也应考虑到水分的散失, 气孔作为水分蒸腾的通道, 在土壤中的水分减少时植物供水能力降低(表1 ), 会影响气孔的发生、分化和发育等, 单位面积内叶脉密度的增加使叶脉间距变小, 气孔变窄、气孔密度加大(陈伟月等, 2014 ), 如果气孔不能感知这些变化并做出响应, 就提高了木质部栓塞的可能性, 进而增加了叶片液压功能失调至脱水的风险(Maseda & Fernández, 2006 ).当植株叶片获得的光合有效辐射较大时, 叶片气孔及表皮细胞迅速对光照做出反应, 直接向大气蒸发散水, 此时叶片其他部位并未发生水分亏缺, 小的气孔可以迅速关闭, 防止水分进一步流失, 避免水分亏缺在全叶中发生, 以此减少植物整叶枯萎的损失(高春娟等, 2012 ).因此, 生长在南坡的刺槐选择高的叶脉密度、气孔密度和小的气孔, 验证了Zhang (2012)的叶脉密度与气孔特征协同进化的理论. ...
Stay wet or else: Three ways in which plants can adjust hydraulically to their environment
2006
A note on normal correlation
1
1939
... 形态特征和生长特性的改变是植物适应不同环境和资源水平的重要策略(Maherali & Pelucia, 2001 ), 叶片形态和生理特性在植物碳同化、水分关系和能量平衡等方面具有重要作用, 决定了叶片功能属性在不同环境条件下的差异(Wright et al ., 2004 ; Poorter et al ., 2006 ).东坡和西坡位于阴坡和阳坡过渡区域, 属于植被类型的交错地带, 群落组成较复杂, 群落郁闭度和高度都介于南坡和北坡之间.自然条件下刺槐选择折中了叶脉密度与气孔密度、气孔大小的稳步投资, 兼顾了植株的水分输送、利用与保持等功能的需求, 选择了气孔与叶脉的均衡生长模式, 可以达到更大限度地利用地上部分资源和拓展空间的目的, 充分利用水资源与光资源, 减少植株内部的资源竞争.因此, 生长在东西坡的刺槐的气孔密度、气孔大小与叶脉密度的相关性之间没有明显差异, 这是光合产物在不同构件或功能间权衡分配的结果. ...
Architecture of 54 moist forest tree species: Traits, trade-offs, and functional groups
1
2006
... (1)植物光合参数的测定: 选择晴朗天气的9:00-12:00进行气体交换参数的测定.光合测定使用GFS-3000便携式光合测量系统(Heinz Walz GmbH, Bavaria, Germany), 测量过程中使用人工红蓝光源, 光合有效辐射(PAR )为1200 μmol·m-2 ·s-1 , CO2 浓度约为340 μmol·mol-1 , 相对湿度(RH )保持在40%-50%, 流速设定为750 μmol·s-1 , 叶室面积为8 cm2 , 对做好标记的每个叶片记录5组数据用于统计, 分析分别测定的叶片净光合速率(P n )、蒸腾速率(T r )等参数(任青吉等, 2015 ).对于不能充满叶室的叶片, 则保存于湿润的封口袋中, 带回实验室, 用便携式激光叶面积仪(CI-202, Walz, Camas, USA)测定叶面积, 进而计算P n 和T r . ...
玛曲高寒沼泽化草甸51种植物光合生理和叶片形态特征的比较. 植物生态学报,
1
2015
... 作为植物碳水耦合权衡的重要器官, 叶片在进化过程中对环境的变化较敏感且可塑性较大(Scoffoni et al ., 2015 ), 从叶片形态解剖结构了解植物生态策略在异质生境中的变化对植物的结构和生理变化具有重要意义(Maricle et al ., 2009 ).叶脉是影响叶片水分供应和利用的重要结构, 在一定意义上反映了叶片资源流通能力(Sack & Frole, 2006 ; Sack & Scoffoni, 2013 ).叶脉密度作为叶脉网络功能性状的重要指标(Brodribb et al ., 2010 ; Blonder et al ., 2011 ), 可反映叶脉系统的水分、养分和光合产物等物质运输的效率和安全性以及抵抗外界干扰的能力, 是理解植物环境可塑性机制的重要途径(李乐等, 2013 ).气孔是植物叶片与外界进行水气交换的唯一可调节通道, 通过保卫细胞对环境和内源信号的感知, 与植物的光合作用、蒸腾作用等生理生态功能密切相关(杨惠敏和王根轩, 2001 ).叶脉与气孔是叶片水分供给和散失的重要因素, 两者的大小和数量的配置直接影响着叶片的水分利用效率, 进而影响叶片结构及生理功能的调节机制(Fiorin et al ., 2016 ).当植株处于高光强条件下时, 相对湿度下降, 叶片受到的水分胁迫增加, 叶片在获取光合收益的同时也需兼顾内部水分动态平衡(Fiorin et al ., 2016 ), 此时, 植株往往通过调整净光合速率与蒸腾速率来应对不利环境, 较高的蒸腾速率要求叶片发育较多的叶脉以提供供水系统, 在叶片的蒸腾作用中通过控制气孔密度和气孔大小决定进入大气的水分和CO2 , 降低植物的呼吸和蒸腾成本.叶脉密度与气孔性状共同作用体现了气孔蒸腾对水分的需求和叶脉系统对水分的供给相互匹配与权衡(Brodribb & Jordan, 2011 ).坡向作为重要的地形因素, 控制了太阳辐射和降水的空间再分配(Cartón et al ., 2004 ), 由坡向引起的土壤水分、光辐射、光照时长和大气温度等环境因子形成的生境梯度变化影响着群落组成(Bazzaz et al ., 1987 ; 韦莉莉等, 2005 ), 致使群落环境及其密度制约下的种群内植株的生态位存在差异, 群落内部郁闭度和干扰程度不同, 所接受的光合有效辐射也不同, 从而影响植物的空间分布格局及光照、温度等环境(党晶晶等, 2014 ; 杜晶等, 2016 ), 进而影响植物的形态.为了提高光合能力并保证水分供需间的平衡, 植株叶片会适当调整叶脉密度与气孔密度、气孔大小的生物量分配, 补充蒸腾带来的大量水分和能量的消耗(Hetherington & Woodward, 2003 ), 同时也使叶片中单位碳对光合作用的投资效益最大(Brodribb & Jordan, 2011 ), 从而提高生存适合度和竞争能力.因此, 研究微地形影响下叶脉密度与气孔性状的关系, 有助于探讨植物叶片内部水分供应与散失之间的平衡关系, 对植物生理生态功能的维持具有指导意义. ...
Leaf structural diversity is related to hydraulic capacity in tropical rain forest trees
2
2006
... 作为植物碳水耦合权衡的重要器官, 叶片在进化过程中对环境的变化较敏感且可塑性较大(Scoffoni et al ., 2015 ), 从叶片形态解剖结构了解植物生态策略在异质生境中的变化对植物的结构和生理变化具有重要意义(Maricle et al ., 2009 ).叶脉是影响叶片水分供应和利用的重要结构, 在一定意义上反映了叶片资源流通能力(Sack & Frole, 2006 ; Sack & Scoffoni, 2013 ).叶脉密度作为叶脉网络功能性状的重要指标(Brodribb et al ., 2010 ; Blonder et al ., 2011 ), 可反映叶脉系统的水分、养分和光合产物等物质运输的效率和安全性以及抵抗外界干扰的能力, 是理解植物环境可塑性机制的重要途径(李乐等, 2013 ).气孔是植物叶片与外界进行水气交换的唯一可调节通道, 通过保卫细胞对环境和内源信号的感知, 与植物的光合作用、蒸腾作用等生理生态功能密切相关(杨惠敏和王根轩, 2001 ).叶脉与气孔是叶片水分供给和散失的重要因素, 两者的大小和数量的配置直接影响着叶片的水分利用效率, 进而影响叶片结构及生理功能的调节机制(Fiorin et al ., 2016 ).当植株处于高光强条件下时, 相对湿度下降, 叶片受到的水分胁迫增加, 叶片在获取光合收益的同时也需兼顾内部水分动态平衡(Fiorin et al ., 2016 ), 此时, 植株往往通过调整净光合速率与蒸腾速率来应对不利环境, 较高的蒸腾速率要求叶片发育较多的叶脉以提供供水系统, 在叶片的蒸腾作用中通过控制气孔密度和气孔大小决定进入大气的水分和CO2 , 降低植物的呼吸和蒸腾成本.叶脉密度与气孔性状共同作用体现了气孔蒸腾对水分的需求和叶脉系统对水分的供给相互匹配与权衡(Brodribb & Jordan, 2011 ).坡向作为重要的地形因素, 控制了太阳辐射和降水的空间再分配(Cartón et al ., 2004 ), 由坡向引起的土壤水分、光辐射、光照时长和大气温度等环境因子形成的生境梯度变化影响着群落组成(Bazzaz et al ., 1987 ; 韦莉莉等, 2005 ), 致使群落环境及其密度制约下的种群内植株的生态位存在差异, 群落内部郁闭度和干扰程度不同, 所接受的光合有效辐射也不同, 从而影响植物的空间分布格局及光照、温度等环境(党晶晶等, 2014 ; 杜晶等, 2016 ), 进而影响植物的形态.为了提高光合能力并保证水分供需间的平衡, 植株叶片会适当调整叶脉密度与气孔密度、气孔大小的生物量分配, 补充蒸腾带来的大量水分和能量的消耗(Hetherington & Woodward, 2003 ), 同时也使叶片中单位碳对光合作用的投资效益最大(Brodribb & Jordan, 2011 ), 从而提高生存适合度和竞争能力.因此, 研究微地形影响下叶脉密度与气孔性状的关系, 有助于探讨植物叶片内部水分供应与散失之间的平衡关系, 对植物生理生态功能的维持具有指导意义. ...
... 刺槐(Robinia pseudoacacia )是温带落叶乔木, 是我国华北和西北地区绿化荒山和防风固沙的主要植物, 具有固氮能力强和碳储存量大的优势, 作为黄土丘陵区的主要人工造林树种, 其根系发达、生长迅速、成活率高和环境适应性强等特点在保持水土和生态防护中发挥着重要作用.近年来, 关于刺槐在干旱半干旱地区的种植范围和生态适应性等问题引起了学界的广泛关注(于占辉等, 2009 ; 李军等, 2010 ; 申家朋和张文辉, 2014 ).学术界对气孔性状之间的关系(杨惠敏和王根轩, 2001 ; 王曙光等, 2013 )、气孔与环境因子的关系(左应梅等, 2011 ; 王娜等, 2016 )、气孔性状对水分利用的影响(高洁等, 2004 ; 杨利民等, 2007 )、叶脉网络功能性状与叶片光合作用(Brodribb et al ., 2010 ; Sack et al ., 2012 )、水分利用能力(Zhang et al ., 2012 ; Sack & Scoffoni, 2013 )等问题进行了研究, 阐述了植物叶片在适应特定生境中各种性状的塑造, 而将叶脉与气孔性状结合起来, 探讨二者在异质性生境中叶片内部水分传输及水分平衡适应策略的分析和认识目前还比较薄弱.鉴于此, 本文以兰州北山的人工刺槐林为研究对象, 试图揭示以下两个问题: (1)不同坡向上刺槐叶脉密度与气孔密度、气孔大小之间存在何种关系?(2)这种关系的形成原因有哪些?理解异质生境中植物叶片的物理构建方式, 可为提高半干旱人工林资源的利用率和绿化覆盖率提供理论依据. ...
Leaf venation: Structure, function, development, evolution, ecology and applications in the past, present and future
1
2013
... 刺槐(Robinia pseudoacacia )是温带落叶乔木, 是我国华北和西北地区绿化荒山和防风固沙的主要植物, 具有固氮能力强和碳储存量大的优势, 作为黄土丘陵区的主要人工造林树种, 其根系发达、生长迅速、成活率高和环境适应性强等特点在保持水土和生态防护中发挥着重要作用.近年来, 关于刺槐在干旱半干旱地区的种植范围和生态适应性等问题引起了学界的广泛关注(于占辉等, 2009 ; 李军等, 2010 ; 申家朋和张文辉, 2014 ).学术界对气孔性状之间的关系(杨惠敏和王根轩, 2001 ; 王曙光等, 2013 )、气孔与环境因子的关系(左应梅等, 2011 ; 王娜等, 2016 )、气孔性状对水分利用的影响(高洁等, 2004 ; 杨利民等, 2007 )、叶脉网络功能性状与叶片光合作用(Brodribb et al ., 2010 ; Sack et al ., 2012 )、水分利用能力(Zhang et al ., 2012 ; Sack & Scoffoni, 2013 )等问题进行了研究, 阐述了植物叶片在适应特定生境中各种性状的塑造, 而将叶脉与气孔性状结合起来, 探讨二者在异质性生境中叶片内部水分传输及水分平衡适应策略的分析和认识目前还比较薄弱.鉴于此, 本文以兰州北山的人工刺槐林为研究对象, 试图揭示以下两个问题: (1)不同坡向上刺槐叶脉密度与气孔密度、气孔大小之间存在何种关系?(2)这种关系的形成原因有哪些?理解异质生境中植物叶片的物理构建方式, 可为提高半干旱人工林资源的利用率和绿化覆盖率提供理论依据. ...
Developmentally based scaling of leaf venation architecture explains global ecological patterns
1
2012
... 作为植物碳水耦合权衡的重要器官, 叶片在进化过程中对环境的变化较敏感且可塑性较大(Scoffoni et al ., 2015 ), 从叶片形态解剖结构了解植物生态策略在异质生境中的变化对植物的结构和生理变化具有重要意义(Maricle et al ., 2009 ).叶脉是影响叶片水分供应和利用的重要结构, 在一定意义上反映了叶片资源流通能力(Sack & Frole, 2006 ; Sack & Scoffoni, 2013 ).叶脉密度作为叶脉网络功能性状的重要指标(Brodribb et al ., 2010 ; Blonder et al ., 2011 ), 可反映叶脉系统的水分、养分和光合产物等物质运输的效率和安全性以及抵抗外界干扰的能力, 是理解植物环境可塑性机制的重要途径(李乐等, 2013 ).气孔是植物叶片与外界进行水气交换的唯一可调节通道, 通过保卫细胞对环境和内源信号的感知, 与植物的光合作用、蒸腾作用等生理生态功能密切相关(杨惠敏和王根轩, 2001 ).叶脉与气孔是叶片水分供给和散失的重要因素, 两者的大小和数量的配置直接影响着叶片的水分利用效率, 进而影响叶片结构及生理功能的调节机制(Fiorin et al ., 2016 ).当植株处于高光强条件下时, 相对湿度下降, 叶片受到的水分胁迫增加, 叶片在获取光合收益的同时也需兼顾内部水分动态平衡(Fiorin et al ., 2016 ), 此时, 植株往往通过调整净光合速率与蒸腾速率来应对不利环境, 较高的蒸腾速率要求叶片发育较多的叶脉以提供供水系统, 在叶片的蒸腾作用中通过控制气孔密度和气孔大小决定进入大气的水分和CO2 , 降低植物的呼吸和蒸腾成本.叶脉密度与气孔性状共同作用体现了气孔蒸腾对水分的需求和叶脉系统对水分的供给相互匹配与权衡(Brodribb & Jordan, 2011 ).坡向作为重要的地形因素, 控制了太阳辐射和降水的空间再分配(Cartón et al ., 2004 ), 由坡向引起的土壤水分、光辐射、光照时长和大气温度等环境因子形成的生境梯度变化影响着群落组成(Bazzaz et al ., 1987 ; 韦莉莉等, 2005 ), 致使群落环境及其密度制约下的种群内植株的生态位存在差异, 群落内部郁闭度和干扰程度不同, 所接受的光合有效辐射也不同, 从而影响植物的空间分布格局及光照、温度等环境(党晶晶等, 2014 ; 杜晶等, 2016 ), 进而影响植物的形态.为了提高光合能力并保证水分供需间的平衡, 植株叶片会适当调整叶脉密度与气孔密度、气孔大小的生物量分配, 补充蒸腾带来的大量水分和能量的消耗(Hetherington & Woodward, 2003 ), 同时也使叶片中单位碳对光合作用的投资效益最大(Brodribb & Jordan, 2011 ), 从而提高生存适合度和竞争能力.因此, 研究微地形影响下叶脉密度与气孔性状的关系, 有助于探讨植物叶片内部水分供应与散失之间的平衡关系, 对植物生理生态功能的维持具有指导意义. ...
Light-induced plasticity in leaf hydraulics, venation, anatomy, and gas exchange in ecologically diverse
1
2015
... 刺槐(Robinia pseudoacacia )是温带落叶乔木, 是我国华北和西北地区绿化荒山和防风固沙的主要植物, 具有固氮能力强和碳储存量大的优势, 作为黄土丘陵区的主要人工造林树种, 其根系发达、生长迅速、成活率高和环境适应性强等特点在保持水土和生态防护中发挥着重要作用.近年来, 关于刺槐在干旱半干旱地区的种植范围和生态适应性等问题引起了学界的广泛关注(于占辉等, 2009 ; 李军等, 2010 ; 申家朋和张文辉, 2014 ).学术界对气孔性状之间的关系(杨惠敏和王根轩, 2001 ; 王曙光等, 2013 )、气孔与环境因子的关系(左应梅等, 2011 ; 王娜等, 2016 )、气孔性状对水分利用的影响(高洁等, 2004 ; 杨利民等, 2007 )、叶脉网络功能性状与叶片光合作用(Brodribb et al ., 2010 ; Sack et al ., 2012 )、水分利用能力(Zhang et al ., 2012 ; Sack & Scoffoni, 2013 )等问题进行了研究, 阐述了植物叶片在适应特定生境中各种性状的塑造, 而将叶脉与气孔性状结合起来, 探讨二者在异质性生境中叶片内部水分传输及水分平衡适应策略的分析和认识目前还比较薄弱.鉴于此, 本文以兰州北山的人工刺槐林为研究对象, 试图揭示以下两个问题: (1)不同坡向上刺槐叶脉密度与气孔密度、气孔大小之间存在何种关系?(2)这种关系的形成原因有哪些?理解异质生境中植物叶片的物理构建方式, 可为提高半干旱人工林资源的利用率和绿化覆盖率提供理论依据. ...
黄土丘陵区退耕还林地刺槐人工林碳储量及分配规律. 生态学报,
2
2014
... 试验区位于兰州市北山九州台西北师范大学绿化基地(36.17°-36.23° N, 103.21°-103.25° E), 区内海拔1536-1914 m, 温带大陆性季风气候显著, 年平均气温5-9 ℃, ≥10 ℃的活动积温3385.4 ℃, 年日照时间为2607.6 h, 年太阳总辐射529.6-614.0 kJ·cm-2 , 年降水量327.7 mm, 且主要集中在6-9月, 约占全年降水量的60%, 相反, 年蒸发量很大, 达1650 mm, 为降水量的5倍多(史元春等, 2015 ).土壤为黄土母质上发育起来的灰钙土.供试的刺槐林建植于2002年, 栽培的刺槐为同一品种的同龄幼苗, 平均造林密度为2500株·hm-2 , 每年灌溉4-6次, 灌溉量约800 m3 ·hm-2 .植被群落以刺槐和侧柏(Platycladus orientalis )人工林为主, 主要林下植物有红砂(Reaumuria songarica )、枸杞(Lycium chinense )、碱蓬(Suaeda glauca )、白茎盐生草(Halogeton arachnoideus )、柽柳(Tamarix chinensis )、柠条锦鸡儿(Caragana korshinskii )等. ...
... 环境胁迫驱动下的自然选择使得植物倾向于以最小的水分散失来获得最大的碳同化速率(Hether- ington & Woodward, 2003 ), 植物通过调整自身生理特征和解剖结构来适应不同的生境条件(王瑞丽等, 2016 ).生长在南坡刺槐的叶脉密度与气孔密度呈极显著正相关关系(p < 0.01, 图1 ), 与气孔大小存在极显著负相关关系(p < 0.01, 图2 ).研究区位于北半球中纬度地区山地, 南坡太阳直射角较大, 光照时间长, 接受的光照充足, 总的辐射总量和温度均高于北坡(党晶晶等, 2014 ), 南坡植被稀疏, 接受的光合有效辐射最大(表2 ), 高温、强辐射造成刺槐叶片蒸腾加剧, 较强的蒸腾作用使植物面临着水分胁迫, 为了适应高温下的蒸腾速率, 植物倾向于发育小而厚的叶片(史元春等, 2015 ), 将更多的光合产物用于增加叶脉密度以保证高效率的水分输导系统.叶片在进行光合作用时不可避免地伴随着水分的蒸发损耗(Brodribb & Jordan, 2011 ), 在叶片增加运输管道时也应考虑到水分的散失, 气孔作为水分蒸腾的通道, 在土壤中的水分减少时植物供水能力降低(表1 ), 会影响气孔的发生、分化和发育等, 单位面积内叶脉密度的增加使叶脉间距变小, 气孔变窄、气孔密度加大(陈伟月等, 2014 ), 如果气孔不能感知这些变化并做出响应, 就提高了木质部栓塞的可能性, 进而增加了叶片液压功能失调至脱水的风险(Maseda & Fernández, 2006 ).当植株叶片获得的光合有效辐射较大时, 叶片气孔及表皮细胞迅速对光照做出反应, 直接向大气蒸发散水, 此时叶片其他部位并未发生水分亏缺, 小的气孔可以迅速关闭, 防止水分进一步流失, 避免水分亏缺在全叶中发生, 以此减少植物整叶枯萎的损失(高春娟等, 2012 ).因此, 生长在南坡的刺槐选择高的叶脉密度、气孔密度和小的气孔, 验证了Zhang (2012)的叶脉密度与气孔特征协同进化的理论. ...
兰州北山刺槐枝叶性状的坡向差异性. 植物生态学报,
1
2015
... 对研究区1:50000地形图利用ArcGIS进行数字化分析, 得到栅格大小为25 m的数字高程模型(digital elevation model, DEM).然后在DEM基础上利用Spatial Analyst模块中的Surface Analysis命令提取坡向, 其中坡向采用3次有限差分方法计算得到(Skidmore, 1989 ), 表示地表面上一点的切平面的法线矢量n 在水平面的投影n xoy 与过该点的正北方向的夹角, 最后利用提取出的坡向指标建立每块样地的数据查找表, 其中坡向组别以国际划分法(陈瑶等, 2006 )确定(圆周角属于315°-360°和0-45°定为正北坡向). ...
A comparison of techniques for calculating gradient and aspect from a gridded digital elevation model
1
1989
... 植物叶片构型构建与植物生理功能所需的光照、碳投入和水分运输的投资收益相契合, 是植物提高光合收益的一个重要的生活史策略(Westoby et al ., 2002 ; 孙素静等, 2015 ).生长在北坡的刺槐的叶脉密度与气孔密度呈显著正相关关系(p < 0.05, 图1 ), 与气孔大小存在显著负相关关系(p < 0.05, 图2 ).随着资源环境的改变, 北坡植被群落郁闭程度和拥挤程度达到最大, 来自邻株植物的遮阴改变了植物所处光环境, 限制了叶片可获得性光资源, 光合有效辐射减弱(表2 ).植株为了获得更多的光资源, 选择了增大叶面积的生长策略, 使得部分叶肉组织与叶脉接触面积减少, 相对减少叶脉构建投资; 另一方面, 在土壤含水量较充沛的条件下, 植物供水较充足, 气孔张合度受到影响, 单位叶面积内气孔数量减少且CO2 扩散阻力增加, 叶片吸收CO2 受到限制, 光合速率显著降低, 引起植物地上部分对水的需求减少, 使得植物不必投资更多叶脉导管就可以满足叶片水分的供应, 有利于提高植物在资源丰富的环境中的竞争力(Beerling & Franks, 2010 ; Field et al ., 2011 ).气孔直接影响着植物的CO2 交换速率、蒸腾速率和光合速率, 调节气孔密度和气孔长度可适应时刻发生变化的环境状况, 使植物在水碳循环中的平衡调节作用达到最优(高春娟等, 2012 ; 高冠龙等, 2016 ), 当土壤水分较充足时, 根系吸收的水分足以保持体内水分平衡, 气孔进一步开放虽然使得蒸腾速率增加, 但气孔对蒸腾速率的敏感度降低, 吸水速率与蒸腾速率的平衡机制对气孔反馈减少(左应梅等, 2011 ), 采取了“疏而大”的适应特征, 从而保证叶片能吸收足量的CO2 , 进行正常的光合作用, 这与Dow等(2014)对拟南芥(Arabidopsis thaliana )气孔的研究结果一致.刺槐在北坡选择了降低叶脉密度与气孔密度、增大气孔的策略, 使叶含水量维持在动态平衡范围内, 这对于理解植物对环境的适应策略、叶性状之间资源的分配与利用具有重要意义. ...
叶脉网络系统的构建和系统学意义研究进展. 热带亚热带植物学报,
1
2015
... 刺槐(Robinia pseudoacacia )是温带落叶乔木, 是我国华北和西北地区绿化荒山和防风固沙的主要植物, 具有固氮能力强和碳储存量大的优势, 作为黄土丘陵区的主要人工造林树种, 其根系发达、生长迅速、成活率高和环境适应性强等特点在保持水土和生态防护中发挥着重要作用.近年来, 关于刺槐在干旱半干旱地区的种植范围和生态适应性等问题引起了学界的广泛关注(于占辉等, 2009 ; 李军等, 2010 ; 申家朋和张文辉, 2014 ).学术界对气孔性状之间的关系(杨惠敏和王根轩, 2001 ; 王曙光等, 2013 )、气孔与环境因子的关系(左应梅等, 2011 ; 王娜等, 2016 )、气孔性状对水分利用的影响(高洁等, 2004 ; 杨利民等, 2007 )、叶脉网络功能性状与叶片光合作用(Brodribb et al ., 2010 ; Sack et al ., 2012 )、水分利用能力(Zhang et al ., 2012 ; Sack & Scoffoni, 2013 )等问题进行了研究, 阐述了植物叶片在适应特定生境中各种性状的塑造, 而将叶脉与气孔性状结合起来, 探讨二者在异质性生境中叶片内部水分传输及水分平衡适应策略的分析和认识目前还比较薄弱.鉴于此, 本文以兰州北山的人工刺槐林为研究对象, 试图揭示以下两个问题: (1)不同坡向上刺槐叶脉密度与气孔密度、气孔大小之间存在何种关系?(2)这种关系的形成原因有哪些?理解异质生境中植物叶片的物理构建方式, 可为提高半干旱人工林资源的利用率和绿化覆盖率提供理论依据. ...
CO2 浓度倍增对红松幼苗根尖和叶解剖结构及生理功能的影响. 植物生态学报,
1
2016
... 环境胁迫驱动下的自然选择使得植物倾向于以最小的水分散失来获得最大的碳同化速率(Hether- ington & Woodward, 2003 ), 植物通过调整自身生理特征和解剖结构来适应不同的生境条件(王瑞丽等, 2016 ).生长在南坡刺槐的叶脉密度与气孔密度呈极显著正相关关系(p < 0.01, 图1 ), 与气孔大小存在极显著负相关关系(p < 0.01, 图2 ).研究区位于北半球中纬度地区山地, 南坡太阳直射角较大, 光照时间长, 接受的光照充足, 总的辐射总量和温度均高于北坡(党晶晶等, 2014 ), 南坡植被稀疏, 接受的光合有效辐射最大(表2 ), 高温、强辐射造成刺槐叶片蒸腾加剧, 较强的蒸腾作用使植物面临着水分胁迫, 为了适应高温下的蒸腾速率, 植物倾向于发育小而厚的叶片(史元春等, 2015 ), 将更多的光合产物用于增加叶脉密度以保证高效率的水分输导系统.叶片在进行光合作用时不可避免地伴随着水分的蒸发损耗(Brodribb & Jordan, 2011 ), 在叶片增加运输管道时也应考虑到水分的散失, 气孔作为水分蒸腾的通道, 在土壤中的水分减少时植物供水能力降低(表1 ), 会影响气孔的发生、分化和发育等, 单位面积内叶脉密度的增加使叶脉间距变小, 气孔变窄、气孔密度加大(陈伟月等, 2014 ), 如果气孔不能感知这些变化并做出响应, 就提高了木质部栓塞的可能性, 进而增加了叶片液压功能失调至脱水的风险(Maseda & Fernández, 2006 ).当植株叶片获得的光合有效辐射较大时, 叶片气孔及表皮细胞迅速对光照做出反应, 直接向大气蒸发散水, 此时叶片其他部位并未发生水分亏缺, 小的气孔可以迅速关闭, 防止水分进一步流失, 避免水分亏缺在全叶中发生, 以此减少植物整叶枯萎的损失(高春娟等, 2012 ).因此, 生长在南坡的刺槐选择高的叶脉密度、气孔密度和小的气孔, 验证了Zhang (2012)的叶脉密度与气孔特征协同进化的理论. ...
气孔特征与叶片功能性状之间关联性沿海拔梯度的变化规律: 以长白山为例. 生态学报,
1
2016
... 刺槐(Robinia pseudoacacia )是温带落叶乔木, 是我国华北和西北地区绿化荒山和防风固沙的主要植物, 具有固氮能力强和碳储存量大的优势, 作为黄土丘陵区的主要人工造林树种, 其根系发达、生长迅速、成活率高和环境适应性强等特点在保持水土和生态防护中发挥着重要作用.近年来, 关于刺槐在干旱半干旱地区的种植范围和生态适应性等问题引起了学界的广泛关注(于占辉等, 2009 ; 李军等, 2010 ; 申家朋和张文辉, 2014 ).学术界对气孔性状之间的关系(杨惠敏和王根轩, 2001 ; 王曙光等, 2013 )、气孔与环境因子的关系(左应梅等, 2011 ; 王娜等, 2016 )、气孔性状对水分利用的影响(高洁等, 2004 ; 杨利民等, 2007 )、叶脉网络功能性状与叶片光合作用(Brodribb et al ., 2010 ; Sack et al ., 2012 )、水分利用能力(Zhang et al ., 2012 ; Sack & Scoffoni, 2013 )等问题进行了研究, 阐述了植物叶片在适应特定生境中各种性状的塑造, 而将叶脉与气孔性状结合起来, 探讨二者在异质性生境中叶片内部水分传输及水分平衡适应策略的分析和认识目前还比较薄弱.鉴于此, 本文以兰州北山的人工刺槐林为研究对象, 试图揭示以下两个问题: (1)不同坡向上刺槐叶脉密度与气孔密度、气孔大小之间存在何种关系?(2)这种关系的形成原因有哪些?理解异质生境中植物叶片的物理构建方式, 可为提高半干旱人工林资源的利用率和绿化覆盖率提供理论依据. ...
小麦叶片气孔性状与产量和抗旱性的关系. 应用生态学报,
2013
Common slope tests for bivariate errors-in-variables models
1
2002
... 将坡向划分为北坡(315°-360°和0°-45°)、东坡(45°-135°)、西坡(225°-315°)和南坡(135°-225°), 数据分析前将每个样点刺槐叶片的气孔密度、气孔大小和叶脉密度的平均值进行对数(以10为底)转换后使之符合正态分布.对叶脉密度与气孔性状关系的研究, 采用y = bxa , 线性转换成log(y ) = log(b ) + a log(x ), 式中x 和y 是两个特征参数, b 代表性状关系的截距, a 表示相关性的斜率, 即异速生长参数或相对生长的指数, 当a = 1时, 表示两者是等速关系; 当a > 1时, 表示y 的增加程度大于x 的增加程度; 当a < 1时, 表示y 的增加程度小于x 的增加程度.数据分析主要采用标准化主轴估计(SMA)的方法(Warton et al ., 2006 ), 由软件(S)MATR Version 2.0 (Falster et al ., 2012 )计算完成.每一个回归斜率的置信区间根据Pitman (1939)的方法计算, 并采用Warton和Weber (2002)的方法对每个密度的斜率进行异质性测试.不同坡向间植物功能性状平均值的差异比较采用单因素方差分析(α = 0.05), 实验所用数据采用SPSS 16.0软件进行处理分析. ...
Bivariate line-fitting methods for allometry
1
2006
... 作为植物碳水耦合权衡的重要器官, 叶片在进化过程中对环境的变化较敏感且可塑性较大(Scoffoni et al ., 2015 ), 从叶片形态解剖结构了解植物生态策略在异质生境中的变化对植物的结构和生理变化具有重要意义(Maricle et al ., 2009 ).叶脉是影响叶片水分供应和利用的重要结构, 在一定意义上反映了叶片资源流通能力(Sack & Frole, 2006 ; Sack & Scoffoni, 2013 ).叶脉密度作为叶脉网络功能性状的重要指标(Brodribb et al ., 2010 ; Blonder et al ., 2011 ), 可反映叶脉系统的水分、养分和光合产物等物质运输的效率和安全性以及抵抗外界干扰的能力, 是理解植物环境可塑性机制的重要途径(李乐等, 2013 ).气孔是植物叶片与外界进行水气交换的唯一可调节通道, 通过保卫细胞对环境和内源信号的感知, 与植物的光合作用、蒸腾作用等生理生态功能密切相关(杨惠敏和王根轩, 2001 ).叶脉与气孔是叶片水分供给和散失的重要因素, 两者的大小和数量的配置直接影响着叶片的水分利用效率, 进而影响叶片结构及生理功能的调节机制(Fiorin et al ., 2016 ).当植株处于高光强条件下时, 相对湿度下降, 叶片受到的水分胁迫增加, 叶片在获取光合收益的同时也需兼顾内部水分动态平衡(Fiorin et al ., 2016 ), 此时, 植株往往通过调整净光合速率与蒸腾速率来应对不利环境, 较高的蒸腾速率要求叶片发育较多的叶脉以提供供水系统, 在叶片的蒸腾作用中通过控制气孔密度和气孔大小决定进入大气的水分和CO2 , 降低植物的呼吸和蒸腾成本.叶脉密度与气孔性状共同作用体现了气孔蒸腾对水分的需求和叶脉系统对水分的供给相互匹配与权衡(Brodribb & Jordan, 2011 ).坡向作为重要的地形因素, 控制了太阳辐射和降水的空间再分配(Cartón et al ., 2004 ), 由坡向引起的土壤水分、光辐射、光照时长和大气温度等环境因子形成的生境梯度变化影响着群落组成(Bazzaz et al ., 1987 ; 韦莉莉等, 2005 ), 致使群落环境及其密度制约下的种群内植株的生态位存在差异, 群落内部郁闭度和干扰程度不同, 所接受的光合有效辐射也不同, 从而影响植物的空间分布格局及光照、温度等环境(党晶晶等, 2014 ; 杜晶等, 2016 ), 进而影响植物的形态.为了提高光合能力并保证水分供需间的平衡, 植株叶片会适当调整叶脉密度与气孔密度、气孔大小的生物量分配, 补充蒸腾带来的大量水分和能量的消耗(Hetherington & Woodward, 2003 ), 同时也使叶片中单位碳对光合作用的投资效益最大(Brodribb & Jordan, 2011 ), 从而提高生存适合度和竞争能力.因此, 研究微地形影响下叶脉密度与气孔性状的关系, 有助于探讨植物叶片内部水分供应与散失之间的平衡关系, 对植物生理生态功能的维持具有指导意义. ...
杉木苗木光合作用及其产物分配对水分胁迫的响应. 植物生态学报,
1
2005
... 植物叶片构型构建与植物生理功能所需的光照、碳投入和水分运输的投资收益相契合, 是植物提高光合收益的一个重要的生活史策略(Westoby et al ., 2002 ; 孙素静等, 2015 ).生长在北坡的刺槐的叶脉密度与气孔密度呈显著正相关关系(p < 0.05, 图1 ), 与气孔大小存在显著负相关关系(p < 0.05, 图2 ).随着资源环境的改变, 北坡植被群落郁闭程度和拥挤程度达到最大, 来自邻株植物的遮阴改变了植物所处光环境, 限制了叶片可获得性光资源, 光合有效辐射减弱(表2 ).植株为了获得更多的光资源, 选择了增大叶面积的生长策略, 使得部分叶肉组织与叶脉接触面积减少, 相对减少叶脉构建投资; 另一方面, 在土壤含水量较充沛的条件下, 植物供水较充足, 气孔张合度受到影响, 单位叶面积内气孔数量减少且CO2 扩散阻力增加, 叶片吸收CO2 受到限制, 光合速率显著降低, 引起植物地上部分对水的需求减少, 使得植物不必投资更多叶脉导管就可以满足叶片水分的供应, 有利于提高植物在资源丰富的环境中的竞争力(Beerling & Franks, 2010 ; Field et al ., 2011 ).气孔直接影响着植物的CO2 交换速率、蒸腾速率和光合速率, 调节气孔密度和气孔长度可适应时刻发生变化的环境状况, 使植物在水碳循环中的平衡调节作用达到最优(高春娟等, 2012 ; 高冠龙等, 2016 ), 当土壤水分较充足时, 根系吸收的水分足以保持体内水分平衡, 气孔进一步开放虽然使得蒸腾速率增加, 但气孔对蒸腾速率的敏感度降低, 吸水速率与蒸腾速率的平衡机制对气孔反馈减少(左应梅等, 2011 ), 采取了“疏而大”的适应特征, 从而保证叶片能吸收足量的CO2 , 进行正常的光合作用, 这与Dow等(2014)对拟南芥(Arabidopsis thaliana )气孔的研究结果一致.刺槐在北坡选择了降低叶脉密度与气孔密度、增大气孔的策略, 使叶含水量维持在动态平衡范围内, 这对于理解植物对环境的适应策略、叶性状之间资源的分配与利用具有重要意义. ...
Plant ecological strategies: Some leading dimensions of variation between species
1
2002
... 形态特征和生长特性的改变是植物适应不同环境和资源水平的重要策略(Maherali & Pelucia, 2001 ), 叶片形态和生理特性在植物碳同化、水分关系和能量平衡等方面具有重要作用, 决定了叶片功能属性在不同环境条件下的差异(Wright et al ., 2004 ; Poorter et al ., 2006 ).东坡和西坡位于阴坡和阳坡过渡区域, 属于植被类型的交错地带, 群落组成较复杂, 群落郁闭度和高度都介于南坡和北坡之间.自然条件下刺槐选择折中了叶脉密度与气孔密度、气孔大小的稳步投资, 兼顾了植株的水分输送、利用与保持等功能的需求, 选择了气孔与叶脉的均衡生长模式, 可以达到更大限度地利用地上部分资源和拓展空间的目的, 充分利用水资源与光资源, 减少植株内部的资源竞争.因此, 生长在东西坡的刺槐的气孔密度、气孔大小与叶脉密度的相关性之间没有明显差异, 这是光合产物在不同构件或功能间权衡分配的结果. ...
The worldwide leaf economics spectrum
1
2004
... (2)叶脉密度的测定: 在测完光合参数的植株上采集已标记的叶片, 用含有37%甲醛溶液、50%乙醇和13%冰醋酸溶液的福尔马林-乙酸溶液固定, 然后用含有5% NaOH的乙醇进行化学清理, 用0.1%番红-固绿染色液进行双重染色, 用蒸馏水清洗3遍, 在数码显微镜(Pro 2009a, Regent Instruments, Quebec, Canada)下观察、扫描并拍照(Berlyn & Miksche, 1976 ; 徐婷等, 2016 ).每个叶片拍摄3个视野(叶片两端及中间部位, 包含中脉), 利用CAD软件描绘叶脉长度, 计算所选图像叶脉密度的平均值, 叶脉密度=叶脉长度/叶面积. ...
兰州北山刺槐不同等级叶脉密度与叶大小关系的坡向差异性. 生态学杂志,
1
2016
... (1)气孔密度和气孔大小的测定: 采用印记法, 将采集的叶片用纸巾擦净后, 选取叶片背面中部采集气孔样品(1 cm × 1 cm), 用透明指甲油涂抹在样品上, 待晾干后取出印记叶片放在载玻片上, 置于Motic SFC-18生物显微镜下观察并拍照, 每个叶片拍摄10个视野, 放大100倍观察气孔密度, 在400×放大倍数下观察气孔长度, 用图形软件ImageJ对含有气孔的照片进行编辑处理, 统计样品内的气孔个数.由于气孔的张开程度随时在变, 选用保卫细胞的长度代表气孔的最大张开程度, 即气孔的大小, 而不是测定气孔的实际张开程度(Xu & Zhou, 2008 ).计算气孔密度: 气孔密度=气孔数/面积. ...
Responses of leaf stomatal density to water status and its relationship with photosynthesis in a grass
2
2008
... 作为植物碳水耦合权衡的重要器官, 叶片在进化过程中对环境的变化较敏感且可塑性较大(Scoffoni et al ., 2015 ), 从叶片形态解剖结构了解植物生态策略在异质生境中的变化对植物的结构和生理变化具有重要意义(Maricle et al ., 2009 ).叶脉是影响叶片水分供应和利用的重要结构, 在一定意义上反映了叶片资源流通能力(Sack & Frole, 2006 ; Sack & Scoffoni, 2013 ).叶脉密度作为叶脉网络功能性状的重要指标(Brodribb et al ., 2010 ; Blonder et al ., 2011 ), 可反映叶脉系统的水分、养分和光合产物等物质运输的效率和安全性以及抵抗外界干扰的能力, 是理解植物环境可塑性机制的重要途径(李乐等, 2013 ).气孔是植物叶片与外界进行水气交换的唯一可调节通道, 通过保卫细胞对环境和内源信号的感知, 与植物的光合作用、蒸腾作用等生理生态功能密切相关(杨惠敏和王根轩, 2001 ).叶脉与气孔是叶片水分供给和散失的重要因素, 两者的大小和数量的配置直接影响着叶片的水分利用效率, 进而影响叶片结构及生理功能的调节机制(Fiorin et al ., 2016 ).当植株处于高光强条件下时, 相对湿度下降, 叶片受到的水分胁迫增加, 叶片在获取光合收益的同时也需兼顾内部水分动态平衡(Fiorin et al ., 2016 ), 此时, 植株往往通过调整净光合速率与蒸腾速率来应对不利环境, 较高的蒸腾速率要求叶片发育较多的叶脉以提供供水系统, 在叶片的蒸腾作用中通过控制气孔密度和气孔大小决定进入大气的水分和CO2 , 降低植物的呼吸和蒸腾成本.叶脉密度与气孔性状共同作用体现了气孔蒸腾对水分的需求和叶脉系统对水分的供给相互匹配与权衡(Brodribb & Jordan, 2011 ).坡向作为重要的地形因素, 控制了太阳辐射和降水的空间再分配(Cartón et al ., 2004 ), 由坡向引起的土壤水分、光辐射、光照时长和大气温度等环境因子形成的生境梯度变化影响着群落组成(Bazzaz et al ., 1987 ; 韦莉莉等, 2005 ), 致使群落环境及其密度制约下的种群内植株的生态位存在差异, 群落内部郁闭度和干扰程度不同, 所接受的光合有效辐射也不同, 从而影响植物的空间分布格局及光照、温度等环境(党晶晶等, 2014 ; 杜晶等, 2016 ), 进而影响植物的形态.为了提高光合能力并保证水分供需间的平衡, 植株叶片会适当调整叶脉密度与气孔密度、气孔大小的生物量分配, 补充蒸腾带来的大量水分和能量的消耗(Hetherington & Woodward, 2003 ), 同时也使叶片中单位碳对光合作用的投资效益最大(Brodribb & Jordan, 2011 ), 从而提高生存适合度和竞争能力.因此, 研究微地形影响下叶脉密度与气孔性状的关系, 有助于探讨植物叶片内部水分供应与散失之间的平衡关系, 对植物生理生态功能的维持具有指导意义. ...
... 刺槐(Robinia pseudoacacia )是温带落叶乔木, 是我国华北和西北地区绿化荒山和防风固沙的主要植物, 具有固氮能力强和碳储存量大的优势, 作为黄土丘陵区的主要人工造林树种, 其根系发达、生长迅速、成活率高和环境适应性强等特点在保持水土和生态防护中发挥着重要作用.近年来, 关于刺槐在干旱半干旱地区的种植范围和生态适应性等问题引起了学界的广泛关注(于占辉等, 2009 ; 李军等, 2010 ; 申家朋和张文辉, 2014 ).学术界对气孔性状之间的关系(杨惠敏和王根轩, 2001 ; 王曙光等, 2013 )、气孔与环境因子的关系(左应梅等, 2011 ; 王娜等, 2016 )、气孔性状对水分利用的影响(高洁等, 2004 ; 杨利民等, 2007 )、叶脉网络功能性状与叶片光合作用(Brodribb et al ., 2010 ; Sack et al ., 2012 )、水分利用能力(Zhang et al ., 2012 ; Sack & Scoffoni, 2013 )等问题进行了研究, 阐述了植物叶片在适应特定生境中各种性状的塑造, 而将叶脉与气孔性状结合起来, 探讨二者在异质性生境中叶片内部水分传输及水分平衡适应策略的分析和认识目前还比较薄弱.鉴于此, 本文以兰州北山的人工刺槐林为研究对象, 试图揭示以下两个问题: (1)不同坡向上刺槐叶脉密度与气孔密度、气孔大小之间存在何种关系?(2)这种关系的形成原因有哪些?理解异质生境中植物叶片的物理构建方式, 可为提高半干旱人工林资源的利用率和绿化覆盖率提供理论依据. ...
干旱和CO2 浓度升高对干旱区春小麦气孔密度及分布的影响. 植物生态学报,
1
2001
... 刺槐(Robinia pseudoacacia )是温带落叶乔木, 是我国华北和西北地区绿化荒山和防风固沙的主要植物, 具有固氮能力强和碳储存量大的优势, 作为黄土丘陵区的主要人工造林树种, 其根系发达、生长迅速、成活率高和环境适应性强等特点在保持水土和生态防护中发挥着重要作用.近年来, 关于刺槐在干旱半干旱地区的种植范围和生态适应性等问题引起了学界的广泛关注(于占辉等, 2009 ; 李军等, 2010 ; 申家朋和张文辉, 2014 ).学术界对气孔性状之间的关系(杨惠敏和王根轩, 2001 ; 王曙光等, 2013 )、气孔与环境因子的关系(左应梅等, 2011 ; 王娜等, 2016 )、气孔性状对水分利用的影响(高洁等, 2004 ; 杨利民等, 2007 )、叶脉网络功能性状与叶片光合作用(Brodribb et al ., 2010 ; Sack et al ., 2012 )、水分利用能力(Zhang et al ., 2012 ; Sack & Scoffoni, 2013 )等问题进行了研究, 阐述了植物叶片在适应特定生境中各种性状的塑造, 而将叶脉与气孔性状结合起来, 探讨二者在异质性生境中叶片内部水分传输及水分平衡适应策略的分析和认识目前还比较薄弱.鉴于此, 本文以兰州北山的人工刺槐林为研究对象, 试图揭示以下两个问题: (1)不同坡向上刺槐叶脉密度与气孔密度、气孔大小之间存在何种关系?(2)这种关系的形成原因有哪些?理解异质生境中植物叶片的物理构建方式, 可为提高半干旱人工林资源的利用率和绿化覆盖率提供理论依据. ...
中国东北样带关键种羊草水分利用效率与气孔密度. 生态学报,
1
2007
... 刺槐(Robinia pseudoacacia )是温带落叶乔木, 是我国华北和西北地区绿化荒山和防风固沙的主要植物, 具有固氮能力强和碳储存量大的优势, 作为黄土丘陵区的主要人工造林树种, 其根系发达、生长迅速、成活率高和环境适应性强等特点在保持水土和生态防护中发挥着重要作用.近年来, 关于刺槐在干旱半干旱地区的种植范围和生态适应性等问题引起了学界的广泛关注(于占辉等, 2009 ; 李军等, 2010 ; 申家朋和张文辉, 2014 ).学术界对气孔性状之间的关系(杨惠敏和王根轩, 2001 ; 王曙光等, 2013 )、气孔与环境因子的关系(左应梅等, 2011 ; 王娜等, 2016 )、气孔性状对水分利用的影响(高洁等, 2004 ; 杨利民等, 2007 )、叶脉网络功能性状与叶片光合作用(Brodribb et al ., 2010 ; Sack et al ., 2012 )、水分利用能力(Zhang et al ., 2012 ; Sack & Scoffoni, 2013 )等问题进行了研究, 阐述了植物叶片在适应特定生境中各种性状的塑造, 而将叶脉与气孔性状结合起来, 探讨二者在异质性生境中叶片内部水分传输及水分平衡适应策略的分析和认识目前还比较薄弱.鉴于此, 本文以兰州北山的人工刺槐林为研究对象, 试图揭示以下两个问题: (1)不同坡向上刺槐叶脉密度与气孔密度、气孔大小之间存在何种关系?(2)这种关系的形成原因有哪些?理解异质生境中植物叶片的物理构建方式, 可为提高半干旱人工林资源的利用率和绿化覆盖率提供理论依据. ...
黄土高原半干旱区人工林刺槐展叶期树干液流动态分析. 林业科学,
1
2009
... 刺槐(Robinia pseudoacacia )是温带落叶乔木, 是我国华北和西北地区绿化荒山和防风固沙的主要植物, 具有固氮能力强和碳储存量大的优势, 作为黄土丘陵区的主要人工造林树种, 其根系发达、生长迅速、成活率高和环境适应性强等特点在保持水土和生态防护中发挥着重要作用.近年来, 关于刺槐在干旱半干旱地区的种植范围和生态适应性等问题引起了学界的广泛关注(于占辉等, 2009 ; 李军等, 2010 ; 申家朋和张文辉, 2014 ).学术界对气孔性状之间的关系(杨惠敏和王根轩, 2001 ; 王曙光等, 2013 )、气孔与环境因子的关系(左应梅等, 2011 ; 王娜等, 2016 )、气孔性状对水分利用的影响(高洁等, 2004 ; 杨利民等, 2007 )、叶脉网络功能性状与叶片光合作用(Brodribb et al ., 2010 ; Sack et al ., 2012 )、水分利用能力(Zhang et al ., 2012 ; Sack & Scoffoni, 2013 )等问题进行了研究, 阐述了植物叶片在适应特定生境中各种性状的塑造, 而将叶脉与气孔性状结合起来, 探讨二者在异质性生境中叶片内部水分传输及水分平衡适应策略的分析和认识目前还比较薄弱.鉴于此, 本文以兰州北山的人工刺槐林为研究对象, 试图揭示以下两个问题: (1)不同坡向上刺槐叶脉密度与气孔密度、气孔大小之间存在何种关系?(2)这种关系的形成原因有哪些?理解异质生境中植物叶片的物理构建方式, 可为提高半干旱人工林资源的利用率和绿化覆盖率提供理论依据. ...
Evolutionary association of stomatal traits with leaf vein density in
1
2012
... 植物的光合作用是生态系统生产力形成与演化的基础, 叶片作为光合产物形成的主要器官, 既是整株植物的水力瓶颈, 又是植株应对水力失调的重要枢纽, 在很大程度上影响着植物水分和养分吸收、运移和储存功能(金鹰和王传宽, 2015 ).在长期的环境选择进化中, 植物对水分的需求驱动着叶脉网络的变化, 气孔是植物体蒸腾失水的门户(左应梅等, 2011 ), 植物叶片选择发育更有效的叶脉灌输系统和密集的气孔提高光合效率和水分利用效率(张亚等, 2014 ), 对植物生理功能的维持具有重要意义.本研究发现, 各个坡向刺槐的叶脉密度与气孔密度呈正相关关系, 与气孔大小呈负相关关系, 且在南坡达到极显著水平(p < 0.01, 图1 , 图2 ), 表明坡向梯度上刺槐叶片结构不同功能之间的相互匹配与权衡关系是植物叶片的形态结构和特定的水分供应需求分配策略对生境异质性适应的结果. ...
基部被子植物气孔性状与叶脉密度的关联进化. 植物科学学报,
3
2014
... 刺槐(Robinia pseudoacacia )是温带落叶乔木, 是我国华北和西北地区绿化荒山和防风固沙的主要植物, 具有固氮能力强和碳储存量大的优势, 作为黄土丘陵区的主要人工造林树种, 其根系发达、生长迅速、成活率高和环境适应性强等特点在保持水土和生态防护中发挥着重要作用.近年来, 关于刺槐在干旱半干旱地区的种植范围和生态适应性等问题引起了学界的广泛关注(于占辉等, 2009 ; 李军等, 2010 ; 申家朋和张文辉, 2014 ).学术界对气孔性状之间的关系(杨惠敏和王根轩, 2001 ; 王曙光等, 2013 )、气孔与环境因子的关系(左应梅等, 2011 ; 王娜等, 2016 )、气孔性状对水分利用的影响(高洁等, 2004 ; 杨利民等, 2007 )、叶脉网络功能性状与叶片光合作用(Brodribb et al ., 2010 ; Sack et al ., 2012 )、水分利用能力(Zhang et al ., 2012 ; Sack & Scoffoni, 2013 )等问题进行了研究, 阐述了植物叶片在适应特定生境中各种性状的塑造, 而将叶脉与气孔性状结合起来, 探讨二者在异质性生境中叶片内部水分传输及水分平衡适应策略的分析和认识目前还比较薄弱.鉴于此, 本文以兰州北山的人工刺槐林为研究对象, 试图揭示以下两个问题: (1)不同坡向上刺槐叶脉密度与气孔密度、气孔大小之间存在何种关系?(2)这种关系的形成原因有哪些?理解异质生境中植物叶片的物理构建方式, 可为提高半干旱人工林资源的利用率和绿化覆盖率提供理论依据. ...
... 植物的光合作用是生态系统生产力形成与演化的基础, 叶片作为光合产物形成的主要器官, 既是整株植物的水力瓶颈, 又是植株应对水力失调的重要枢纽, 在很大程度上影响着植物水分和养分吸收、运移和储存功能(金鹰和王传宽, 2015 ).在长期的环境选择进化中, 植物对水分的需求驱动着叶脉网络的变化, 气孔是植物体蒸腾失水的门户(左应梅等, 2011 ), 植物叶片选择发育更有效的叶脉灌输系统和密集的气孔提高光合效率和水分利用效率(张亚等, 2014 ), 对植物生理功能的维持具有重要意义.本研究发现, 各个坡向刺槐的叶脉密度与气孔密度呈正相关关系, 与气孔大小呈负相关关系, 且在南坡达到极显著水平(p < 0.01, 图1 , 图2 ), 表明坡向梯度上刺槐叶片结构不同功能之间的相互匹配与权衡关系是植物叶片的形态结构和特定的水分供应需求分配策略对生境异质性适应的结果. ...
... 植物叶片构型构建与植物生理功能所需的光照、碳投入和水分运输的投资收益相契合, 是植物提高光合收益的一个重要的生活史策略(Westoby et al ., 2002 ; 孙素静等, 2015 ).生长在北坡的刺槐的叶脉密度与气孔密度呈显著正相关关系(p < 0.05, 图1 ), 与气孔大小存在显著负相关关系(p < 0.05, 图2 ).随着资源环境的改变, 北坡植被群落郁闭程度和拥挤程度达到最大, 来自邻株植物的遮阴改变了植物所处光环境, 限制了叶片可获得性光资源, 光合有效辐射减弱(表2 ).植株为了获得更多的光资源, 选择了增大叶面积的生长策略, 使得部分叶肉组织与叶脉接触面积减少, 相对减少叶脉构建投资; 另一方面, 在土壤含水量较充沛的条件下, 植物供水较充足, 气孔张合度受到影响, 单位叶面积内气孔数量减少且CO2 扩散阻力增加, 叶片吸收CO2 受到限制, 光合速率显著降低, 引起植物地上部分对水的需求减少, 使得植物不必投资更多叶脉导管就可以满足叶片水分的供应, 有利于提高植物在资源丰富的环境中的竞争力(Beerling & Franks, 2010 ; Field et al ., 2011 ).气孔直接影响着植物的CO2 交换速率、蒸腾速率和光合速率, 调节气孔密度和气孔长度可适应时刻发生变化的环境状况, 使植物在水碳循环中的平衡调节作用达到最优(高春娟等, 2012 ; 高冠龙等, 2016 ), 当土壤水分较充足时, 根系吸收的水分足以保持体内水分平衡, 气孔进一步开放虽然使得蒸腾速率增加, 但气孔对蒸腾速率的敏感度降低, 吸水速率与蒸腾速率的平衡机制对气孔反馈减少(左应梅等, 2011 ), 采取了“疏而大”的适应特征, 从而保证叶片能吸收足量的CO2 , 进行正常的光合作用, 这与Dow等(2014)对拟南芥(Arabidopsis thaliana )气孔的研究结果一致.刺槐在北坡选择了降低叶脉密度与气孔密度、增大气孔的策略, 使叶含水量维持在动态平衡范围内, 这对于理解植物对环境的适应策略、叶性状之间资源的分配与利用具有重要意义. ...
土壤水分、光照和空气湿度对木薯气孔导度的影响. 生态学杂志,
2011


{kind=link}
{kind=link}
{kind=link}
{kind=link}