版权声明: 2016 植物生态学报编辑部 本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。
基金资助:
展开
摘要
通过对光合-光响应曲线的研究来探索光合有效辐射与净光合速率的关系是一种非常重要的手段。合适的模型才能较好地反映植株的光合特性。分析由于生育期与叶位的不同而导致各模型拟合值与实测值差异的变化性以及不同光合有效辐射强度下各模型的适用性, 可为夏玉米(Zea mays)光合模型的选择和光合-光响应曲线模型的进一步优化提供参考。该研究运用3种典型的光合模型对夏玉米大喇叭口期与乳熟期上部叶、穗位叶与下部叶做光合-光响应曲线拟合, 对比各模型的拟合度以及对实测数据的反映情况, 并将光合有效辐射分为3段, 分析3种模型在每段的拟合残差。结果表明: 最大净光合速率(Pnmax)、饱和光强(Isat)、暗呼吸速率(Rd)等光合特征参数值随叶位由上而下呈降低趋势, 乳熟期普遍小于大喇叭口期; 不同生育期和叶位3种模型均可以拟合, 直角双曲线模型、非直角双曲线模型对Pnmax、Isat的拟合值与实测值有一定的偏差; 通过残差分析表明叶子飘模型的拟合值与实测值最为相符, 尤其对高光部分的拟合表现出独有的优越性。
关键词:
Abstract
Aim The photosynthesis-light response curve is the most commonly used method to explore the relationship between photosynthetically active radiation and the net photosynthetic rate, because it is more effective to reflect the plant photosynthetic characteristics. And it is very meaningful for researchers to choose a suitable summer corn (Zea mays) photosynthetic model and optimal the models of photosynthesis-light response curve by analyzing the differences between simulation and observation in each growth period of some plant. So the object of this paper was to propose some useful suggestions for the choice of summer corn photosynthetic modes and the optimization of the photosynthetic light response curves model in further.Methods In this paper, three typical photosynthetic models were used to fitting the photosynthetic light response curve for upper leaf, leaf at ear of grain and lower leaf of summer corn during bell and milk period. And then the fitting degree of each model was compared to the measured data. Photosynthetic active radiation was divided into three parts, and the fitting residual errors of these three models were analyzed individually.Important findings The photosynthetic characteristic parameters such as maximum net photosynthetic rate (Pnmax), saturated light intensity (Isat) and dark respiration rate (Rd) decreased constantly with a top-down leaf position and the parameters at milk stage were generally lower than the bell stage. Each growth period and leaf position could fit the curve, but some deviation exists for the Pnmax and Isat in the rectangular hyperbolic model and the non-rectangular hyperbolic model. The results of curve fitting residual showed that the simulation values from Ye Zi-Piao model were closest to the actual values, and especially for the high photosynthetically active radiation section.
Keywords:
光合作用是生物界赖以生存的基础, 光是光合作用的主导因子, 对每种绿色植物均可做出光合作用对光的响应曲线(刘宇锋等, 2005)。通过对光合-光响应曲线的研究来探索光合有效辐射与净光合速率的关系是一种非常重要的手段。Baly (1935)提出了用直角双曲线模型拟合光合光响应曲线, 随后出现非直角双曲线模型(Thornley, 1976)、指数函数模型(Bassman & Zwier, 1991)以及由我国学者叶子飘提出的模型(Ye, 2007, 以下称叶子飘模型)等。广大学者应用这些模型对各类植物的光合-光响应曲线进行拟合从而研究植物的光合特性。如用指数函数模型对扁蓿豆(Melissitus ruthenica)光合特性的研究(王照兰等, 2009)、用叶子飘模型研究温度变化对烟草(Nicotiana tabacum)光合作用光响应特征的影响(钟楚等, 2012)等等。在拟合过程中不同模型具有其各自的优缺点, 合适的模型才能更好地拟合光合特征参数, 在以往对某种植物做不同的水分、施肥量或品种下各模型的光合-光响应曲线拟合时, 忽略了由于生育期与叶位的不同而导致各模型拟合值与实测值差异的变化性以及不同光合有效辐射下各模型的适用性; 残差作为实测数据与回归估计值的差值, 通过拟合残差所提供的信息, 分析拟合数据的可靠性、周期性或其他干扰已被广泛应用于天文、气象和光学等各个领域(周朴等, 2010; 陈华华等, 2013; 孙乐强等, 2014)。为了探讨光合-光响应模型对不同生育期不同叶位拟合情况的差异性, 采用3种模型对夏玉米(Zea mays)做光合-光响应曲线拟合, 对比分析不同生育期与叶位的拟合结果, 并结合残差分析不同光合有效辐射下各模型的优缺点与适应性, 为夏玉米拟合模型的选择和光合-光响应曲线模型的进一步优化提供参考。
实验场地位于西北农林科技大学旱区农业水土工程教育部重点实验室塑胶大棚内, 选择抗虫性高的夏玉米品种‘农大375’, 于2015年6月18日在塑料膜透光大棚内种植20盆, 塑料盆上部内口直径31.8 cm, 高34 cm。全生育期123天, 幼苗期与抽雄期进行两次施肥, 采用传统的称质量法与EM50土壤水分监测仪(DECAGON, Pullman, USA)对盆内水分监测。浇水上限为90%田间持水量, 下限为70%田间持水量, 为充分供水环境。分别在2015年7月31日至8月2日和9月13日至9月15日进行光合-光响应曲线的测定, 随机选择3株长势均匀一致的健壮植株进行测定。根据拔节期测量的叶倾角度数自上往下分别选取叶倾角小于10°、小于20°、小于30°的第一个叶片, 依次标记为上部叶、穗位叶、下部叶。测量仪器采用LI-6400便携式光合测定仪(LI-COR, Lincoln, USA), CO2浓度设置为380 μmol·mol-1, 温度设置为33 ℃, 光合有效辐射梯度设置为2000、1800、1600、1400、1200、1000、800、600、400、200、100、50、0 μmol·m-2·s-1。测量前让待测定叶片在光合光量子通量密度为1200 μmol·m-2·s-1的光下、空气中诱导约60 min, 待叶片光合诱导期结束后, 开始测量。
不同模型的数学方程以及计算光合特征参数的方法是不同的, 在运用这些模型拟合时, 需要先对数学模型以及计算参数时所做出的假设做简单介绍。
直角双曲线模型(Baly, 1935):

式中Pn为净光合速率, I为光强, α为植物光合作用对光响应曲线在I = 0时的斜率, 即初始量子效率, Pnmax为最大净光合速率, Rd为暗呼吸速率, θ为反映光响应曲线弯曲程度的曲角参数, 取值0 < θ ≤ 1, β为光抑制项, γ为光饱和项。用公式(1)、(2)计算时需要估算植物的饱和光强, 做法为: 用直线方程拟合弱光强条件下(≤ 200 μmol·m-2·s-1)的光响应数据得到表观量子效率(AQE), 利用非线性最小二乘法估算Pnmax, 而对于饱和光强(Isat), 需要由直线方程Pnmax = AQE × Isat - Rd (叶子飘, 2010)求解。方程(3)是有极值的函数, 可以直接求解Pnmax、Isat的解析解。
实验数据处理由Excel完成, 作图软件为Origin 8.0, 显著性分析由SPSS 19完成。
叶龄是生命活动长短的标尺, 叶龄以及生长状况不同, 会直接影响植株光合作用与光合特征参数(魏雅芬等, 2011; 陈凯利等, 2013), 取两生育期各部位叶片多组实测数据的平均值对其光合特征参数进行分析(表1)。
表1 玉米大喇叭口期与乳熟期光合特征参数
Table 1 Photosynthetic characteristic parameters of Zea mays at bell stage and milk stage respectively
| 时期 Period | 叶位 Leaf position | 表观量子效率 AQE (μmol CO2·μmol-1 photons) | 最大净光合速率 Pnmax (μmol·m-2·s-1) | 饱和光强 Isat (μmol·m-2·s-1) | 光补偿点 Ic (μmol·m-2·s-1) | 暗呼吸速率 Rd (μmol·m-2·s-1) |
|---|---|---|---|---|---|---|
| 大喇叭口期 Bell stage | 上部叶 Upper leaf | 0.061a | 36.78a | 2 000a | 48.87a | 3.36a |
| 穗位叶 Leaf at ear of grain | 0.057b | 36.99a | 2 000a | 49.14a | 3.25a | |
| 下部叶 Lower leaf | 0.051c | 24.20b | 1 800b | 43.40b | 2.63b | |
| 上部叶 Upper leaf | 0.053a | 19.70a | 1 800a | 32.29a | 2.24a | |
| 乳熟期 Milk stage | 穗位叶 Leaf at ear of grain | 0.042b | 18.66a | 1 800a | 32.32a | 1.74b |
| 下部叶 Lower leaf | 0.040c | 16.44b | 1 800a | 31.43a | 1.50c |
两生育期不同部位叶片光合特征参数有明显的差异: 在大喇叭口期, AQE表现为上部叶>穗位叶>下部叶, 降幅依次为6.6%、10.5%; Pnmax上部叶与穗位叶差别不大, 下部叶较穗位叶降低34.7%; Rd表现为上部叶>穗位叶>下部叶。在乳熟期, AQE表现为上部叶>穗位叶>下部叶, 降幅依次为20.8%、4.8%; Pnmax表现为上部叶>穗位叶>下部叶, 降幅依次为5.3%、11.9%; Rd表现为上部叶>穗位叶>下部叶, 降幅依次为22.3%、13.8%。不同生育期叶片的光合特征参数也存在差异, 乳熟期较大喇叭口期各部位叶片的光合特征参数基本均呈现下降趋势, 上部叶AQE下降13.1%, 穗位叶AQE下降26.3%, 下部叶AQE下降21.6%; 上部叶Pnmax下降46.4%, 穗位叶Pnmax下降49.6%, 下部叶Pnmax下降32.5%; 上部叶Rd下降33.3%、穗位叶Rd下降46.5%, 下部叶Rd下降43.0%。
分析发现: 叶片的AQE、Pnmax与Rd等光合特征参数均是由叶位自上而下逐渐降低的, 乳熟期到大喇叭口期叶片的光合特征参数值也是下降的, 后者降幅更大, AQE表示植株对光能尤其是弱光的利用效率(郎莹等, 2011), 一定环境条件下, 叶片Pnmax代表了其最大光合能力(陆佩玲等, 2000), 说明叶位自上而下叶片的光能利用率与最大光合能力都在降低, 大喇叭口期光能的利用率与最大光合能力大于乳熟期。暗呼吸产生能量(腺苷三磷酸、还原型烟酰胺腺嘌呤二核苷酸)、碳骨架为植物生理活动提供了能量和物质基础(蒋高明, 2004), 乳熟期较低的Rd可以降低呼吸作用对光合产物的消耗, 有利于干物质的累积(张其德等, 2001)。
在时间上(同部位叶片不同生育期)与空间上(同生育期不同部位)叶片的光合特性均有差异, 那么这种差异对光合-光响应曲线拟合模型有何影响? 为了探讨这一问题, 运用直角双曲线模型、非直角双曲线模型及叶子飘模型分别对大喇叭口期与乳熟期不同部位叶片进行光合-光响应曲线拟合, 拟合结果见表2。
表2 3种光响应曲线模型拟合大喇叭口期与乳熟期光响应曲线的实测值与拟合结果
Table 2 Simulation results of light response parameters fitted by three models and the measured data at bell stage and milk stage respectively
| 时期 Period | 叶位 Leaf position | 模型 Model | 初始量子效率 α (μmol CO2· μmol-1 photons) | 最大净光合速率 Pnmax (μmol·m-2·s-1) | 饱和光强 Isat (μmol·m-2·s-1) | 光补偿点 Ic (μmol·m-2·s-1) | 暗呼吸速率 Rd (μmol·m-2·s-1) | 决定系数 Determination coefficient |
|---|---|---|---|---|---|---|---|---|
| 上部叶 Upper leaf | 直角双曲线 Rectangular hyperbolic | 0.083a | 55.89Aa | 997Aa | 48.35Aa | 3.70Aa | 0.998 | |
| 非直角双曲线 Non-rectangular hyperbolic | 0.064b | 48.70Ab | 846Ab | 49.33Aa | 3.06Ab | 0.998 | ||
| 叶子飘模型 Ye Zi-Piao model | 0.068c | 37.12Bc | 2 235Bc | 48.71Aa | 3.15Ab | 0.999 | ||
| 测量值 Measured data | - | 36.78B | 2 000B | 48.87A | 3.36B | - | ||
| 直角双曲线 Rectangular hyperbolic | 0.079a | 57.44Aa | 1 071Aa | 50.01Aa | 3.65Aa | 0.997 | ||
| 大喇叭口期 Bell stage | 穗位叶 Leaf at ear of | 非直角双曲线 Non-rectangular hyperbolic | 0.057b | 47.35Ab | 880Ab | 50.75Aa | 2.83Ab | 0.998 |
| grain | 叶子飘模型 Ye Zi-Piao model | 0.063c | 37.10Bc | 2 061Ac | 50.23Aa | 3.01Ac | 0.999 | |
| 测量值 Measured data | - | 36.99B | 2 000B | 49.14A | 3.25B | - | ||
| 直角双曲线 Rectangular hyperbolic | 0.090a | 33.93Aa | 729Aa | 41.46Aa | 3.27Aa | 0.996 | ||
| 下部叶 Lower leaf | 非直角双曲线 Non-rectangular hyperbolic | 0.059b | 29.56Ab | 628Ab | 44.14Aa | 2.50Ba | 0.999 | |
| 叶子飘模型 Ye Zi-Piao model | 0.070c | 24.18Bc | 1 789Bc | 42.98Aa | 2.76Bb | 0.999 | ||
| 测量值 Measured data | - | 24.20B | 1 800B | 43.40A | 2.63B | - | ||
| 直角双曲线 Rectangular hyperbolic | 0.102a | 25.30Aa | 540Aa | 31.31Aa | 2.71Aa | 0.997 | ||
| 上部叶 | 非直角双曲线 Non-rectangular hyperbolic | 0.076b | 23.90Bb | 499Aa | 32.29Aa | 2.29Bb | 0.999 | |
| Upper leaf | 叶子飘模型 Ye Zi-Piao model | 0.089c | 22.71Bb | 1 973Bb | 31.63Aa | 2.45Ba | 0.998 | |
| 测量值 Measured data | - | 19.70B | 1 800B | 32.29Aa | 2.24B | - | ||
| 直角双曲线 Rectangular hyperbolic | 0.076a | 24.94Aa | 645Aa | 32.09Aa | 2.16Aa | 0.996 | ||
| 乳熟期 Milk stage | 穗位叶 Leaf at ear of | 非直角双曲线 Non-rectangular hyperbolic | 0.050b | 20.02Ab | 561Ab | 32.88Aa | 1.59Bb | 0.998 |
| grain | 叶子飘模型 Ye Zi-Piao model | 0.062c | 18.55Bc | 1 819Bc | 32.36Aa | 1.83Bb | 0.999 | |
| 测量值 Measured data | - | 18.66B | 1 800B | 32.32A | 1.74B | - | ||
| 直角双曲线 Rectangular hyperbolic | 0.073a | 21.88Aa | 595Aa | 30.41Aa | 1.92Aa | 0.996 | ||
| 下部叶 Lower leaf | 非直角双曲线 Non-rectangular hyperbolic | 0.049b | 19.67Ab | 528Ab | 30.81Aa | 1.45Bb | 0.998 | |
| 叶子飘模型 Ye Zi-Piao model | 0.059c | 16.45Bc | 1 777Bc | 30.40Aa | 1.61Bb | 0.999 | ||
| 测量值 Measured data | - | 16.44B | 1 800B | 31.43A | 1.50B | - |
3种模型的决定系数均较高, 最低值0.996, 初始量子效率(α)在0-0.125理论范围值之内(陆佩玲等, 2000)。说明直角双曲线模型、非直角双曲线模型、叶子飘模型都可以用来拟合玉米不同生育期不同部位叶片的光合-光响应曲线(图1)。但衡量一个模型的适用性不是仅仅看拟合度的高低, 还要结合拟合参数对实测值的反映情况, 对比分析拟合结果, 各模型拟合的参数值有差异, 对实测值的反映情况也不尽相同, 由叶子飘模型得到的Pnmax的拟合值与实测值最接近, 直角双曲线模型与非直角双曲线模型的拟合值均远大于实测值, 但与实测值的偏离度是不一样的, 直角双曲线模型在大喇叭口期上部叶、穗位叶、下部叶较实测值的偏离度依次为52%、55%、26%, 大喇叭口期依次为40%、34%、33%; 非直角双曲线模型在大喇叭口期的偏离度依次为32%、28%、22%, 乳熟期依次为21%、7%、20%, 可以看出乳熟期的偏离度整体是小于大喇叭口期的, 叶片由上而下偏离度也是降低的。对Isat的拟合值叶子飘模型与实测值最接近, 直角双曲线模型与非直角双曲线模型远小于实测值, 直角双曲线模型在大喇叭口期上部叶、穗位叶、下部叶的偏离度依次为50%、46%、60%, 在乳熟期的偏离度依次为58%、56%、65%; 非直角双曲线模型在大喇叭口期的偏离度依次为70%、64%、67%, 乳熟期的偏离度依次为72%、69%、70%, 乳熟期的偏差整体大于大喇叭口期, 不同部位叶片之间无明显差异。3种模型对光补偿点的拟合结果均较好。直角双曲线模型对Rd的拟合值均大于实测值, 在大喇叭口期上部叶、穗位叶、下部叶较实测值的偏离度依次为10%、12%、24%, 乳熟期依次为21%、24%、28%, 乳熟期的偏离度整体上大于大喇叭口期的, 叶片由上而下偏离度也是增加的; 非直角双曲线模型和叶子飘模型对乳熟期Rd的拟合整体较好, 而在大喇叭口期只对呼吸作用相对较弱的下部叶可以较好地拟合。从各拟合参数来看, 对于光补偿点, 3种模型均可以较好地反映实测值; 对于Rd, 非直角双曲线模型和叶子飘模型优于直角双曲线模型; 对于Pnmax和Isat, 叶子飘模型优于其余两种模型, 这也反映了决定系数高只能说明曲线拟合程度好, 并不能保证拟合结果一定与实测值相符。
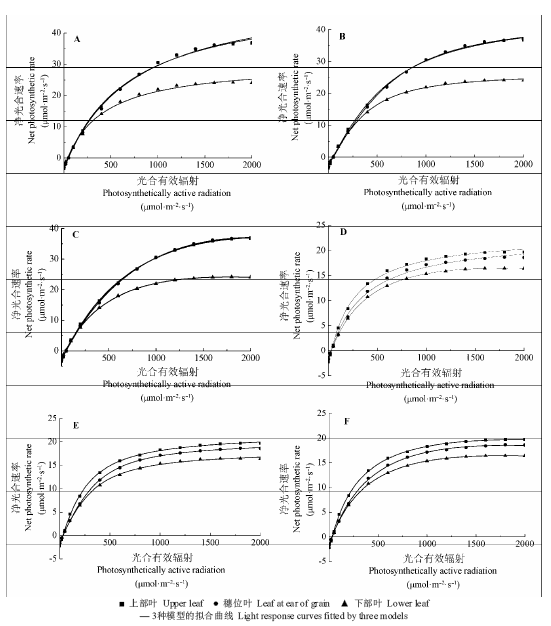
图1 夏玉米大喇叭口期与乳熟期3种模型的光合-光响应曲线拟合图。A, 直角双曲线模型对大喇叭口期叶片的光合-光响应曲线。B, 非直角双曲线模型对大喇叭口期叶片的光合-光响应曲线。C, 叶子飘模型对大喇叭口期叶片的光合-光响应曲线。D, 直角双曲线模型对乳熟期叶片的光合-光响应曲线。E, 非直角双曲线模型对乳熟期叶片的光合-光响应曲线。F, 叶子飘模型对乳熟期叶片的光合-光响应曲线。
Fig. 1 Light response curves fitted by three models at bell stage and milk stage for summer maize. A, Light response curves fitted by rectangular hyperbolic model at bell stage. B, Light response curves fitted by non-rectangular hyperbolic model at bell stage. C, Light response curves fitted by Ye Zi-Piao model at bell stage. D, Light response curves fitted by rectangular hyperbolic model at milk stage. E, Light response curves fitted by non-rectangular hyperbolic model at milk stage. F, Light response curves fitted by Ye Zi-Piao model at milk stage.
从拟合图(图1)可以看出: 各模型对两生育期三部位的叶片均是可以拟合的, 拟合图更加直观地表现出两时期各部位叶片的光合情况: 大喇叭口期上部叶与穗位叶的拟合曲线基本重合, 上部叶净光合速率大于下部叶, 乳熟期净光合速率上部叶>穗位叶>下部叶, 大喇叭口期曲线变平缓的光强点相对于乳熟期明显靠后, 表明大喇叭口期的Isat大于乳熟期, 曲线所表现出的规律与上文对实测数据的分析结果是一致的。这是由于叶龄的原因, 在大喇叭口期, 玉米上部叶和穗位叶均较新, 光合作用强烈, 拟合图中便出现上部叶与穗位叶的拟合曲线重合且整体位于下部叶拟合曲线的上方, 而到了乳熟期, 三处叶片均开始衰老, 但上部叶较穗位叶叶龄小, 穗位叶较下部叶叶龄小, 图中便出现由上而下光合递减的阶梯状分布, 大喇叭口期的叶片比乳熟期要新, 其净光合速率整体便大于乳熟期。
图1中各拟合曲线的坡度均由陡变缓, 依据光强分为坡度较大阶段(0-200 μmol·m-2·s-1)、过渡阶段(400-1000 μmol·m-2·s-1)和平缓阶段(1200-2000 μmol·m-2·s-1)。在坡度较大阶段, 净光合速率快速增高, 3种模型的拟合曲线均可以较好地穿过实测点, 过渡阶段曲线处于一个由陡变缓的过程, 说明净光合速率仍在增加, 但增速放缓, 图1中直角双曲线模型有部分实测点没有落在拟合曲线上, 平缓阶段曲线坡度变平缓, 净光合速率基本不变, 直角双曲线模型与非直角双曲线模型均有部分点没有落在拟合曲线上, 非直角双曲线模型优于直角双曲线模型。对此, 分别求取所有拟合曲线在坡度较大阶段、过渡阶段和平缓阶段的残差平方和, 结果见表3。在坡度较大阶段, 3种模型的残差平方和均较小, 此阶段对光合特征参数(光补偿点、暗呼吸速率)的拟合(表2)也均较好; 过渡阶段的残差平方和只有直角双曲线模型大于1, 在图1中表现出部分实测数据没有落在拟合曲线上; 平缓阶段高光部分叶子飘模型的残差最小, 对应的实测值对拟合曲线的符合度(图1)和对光合特征参数(Pnmax、Isat)的拟合(表2)均为最优。
表3 3种模型拟合曲线的残差平方和
Table 3 Residual sum of squares of light-response curves fitted by three models
| 光合有效辐射 PAR (μmol·m-2·s-1) | 0-200 | 400-1 000 | 1 200-2 000 |
|---|---|---|---|
| 直角双曲线 Rectangular hyperbolic | 0.75 | 1.15 | 1.92 |
| 非直角双曲线 Non-rectangular hyperbolic | 0.40 | 0.31 | 0.50 |
| 叶子飘模型 Ye Zi-Piao model | 0.28 | 0.14 | 0.05 |
随着叶龄增加, 叶片逐渐衰老, 气孔导度下降, Rubisco含量和活性开始下降, 光合电子传递能力降低, 因而光合能力逐渐减弱(Dai et al., 2004)。无论哪种模型拟合均可以反映出夏玉米在叶龄的影响下随着叶位靠下与生育期的靠后, 净光合速率会下降并随PAR变化的拐点会靠前。在PAR为0-2000 μmol·m-2·s-1时, 光合特性的变化是有一定规律可循的, 在大喇叭口期PAR为0-200 μmol·m-2·s-1时, 3个部位叶片的净光合速率基本一致, 而乳熟期PAR为0-100 μmol·m-2·s-1时, 3个部位叶片的净光合速率基本一致。以此为分界点, 随着PAR的增大, 大喇叭口期上部叶与穗位叶净光合速率的增速明显大于下部叶, 乳熟期净光合速率的增速为上部叶>穗位叶>下部叶。大喇叭口期上部叶与穗位叶在PAR为1400 μmol·m-2·s-1时, 净光合速率的增速放缓, 下部叶在PAR为1000 μmol·m-2·s-1时增速放缓; 乳熟期净光合速率均在PAR为1000 μmol·m-2·s-1时增速放缓。任丽丽和高辉远(2008)对酸模(Rumex acetosa)、师生波等(2011)对不同叶龄拟南芥(Arabidopsis thaliana)的研究均表明叶片充分生长后随着叶龄的增长植株光合能力下降, 本研究中玉米乳熟期光合能力小于大喇叭口期, 叶位由上而下光合能力逐步降低, 与他们的研究结论一致。叶龄是影响植株暗呼吸的一个主要因素, 叶片的Rd均随着叶龄的增加而降低(Villar et al., 1995; 孙金伟等, 2013), 这是因为快速生长期的植株需要更多的物质和能量, 以满足生长需求, 大喇叭口期叶片比乳熟期叶片新, 越靠上部叶位的叶龄越小, 随着生育期的靠后以及叶位由上而下, Rd逐步降低。AQE与Rd呈现相同的变化规律。我们综合各光合特征参数的变化规律可以发现, 其与拟合曲线在坡度较大阶段是吻合的, 因为新叶片的AQE高, 其在低光强阶段的光合作用强, 但其暗呼吸作用也强, 较老的叶片AQE低暗呼吸也弱, 在两者的共同作用下拟合曲线在低光阶段便出现重合的现象; 随着光强的升高, 光合作用速率增加, 暗呼吸的影响相对减弱, 不同叶龄的拟合曲线在坡度较大阶段后便呈现不同变化趋势。
对于植物光合-光响应曲线的拟合, 常见的模型有直角双曲线模型、非直角双曲线模型、指数函数模型和叶子飘模型, 指数函数模型对于玉米等C4植物应用较少, 本文中并没有采用。Yu等(2004)关于冬小麦(Triticum aestivum)气孔导度与叶片光合、蒸腾等关系的研究表明直角双曲线模型极大地估算了叶片的Pnmax; Junior等(2013)对亚马孙河10种树木进行了大量的光响应曲线拟合, 表明直角双曲线与非直角双曲线模型会过大估算Pnmax; Lobo等(2013)用Microsoft Excel拟合光合-光响应曲线时对比分析了各拟合模型, 肯定了叶子飘模型在Pnmax和Isat方面拟合的优势。本文的结论与上述研究成果是一致的, 但Lobo等(2013)的研究结果同时也表明该模型对模型各参数的生理学意义没有作深远的阐述。这一问题现在已解决, β为光抑制项, γ为光饱和项, 当γ一定时, β值越大, 光响应曲线弯曲程度就越大, 植物受光抑制就越强。当β一定时, γ值越小, 表示光响应曲线弯曲程度就越小, 植物就越不容易发生光饱和现象(叶子飘和康华靖, 2012)。然而, 各模型的差异在不同生育期不同部位叶片之间是否有所变化, 前人并没有给出结果, 通过对拟合参数与实测数据的对比, 发现对于Pnmax, 直角双曲线模型与非直角双曲线模型与实测数据的偏离度在大喇叭口期上部叶与穗位叶一致, 而到下部叶是降低的, 乳熟期的偏离度上部叶>穗位叶>下部叶, 大喇叭口期的偏差整体大于乳熟期, 与净光合速率的变化是一致的, 光合作用越强, 偏离度越大; 对Isat拟合的偏离情况不同部位叶片之间未发现统一规律, 乳熟期整体大于大喇叭口期, 叶子飘模型在不同生育期不同叶位拟合中并无这种偏差的情况, 有很好的稳定性, 与实测值也最为接近。从拟合图形与残差分析可以看出, 3种模型对于0-200 μmol·m-2·s-1低光强阶段的拟合均较好, 与表2中3种模型对低光阶段的暗呼吸速率与光补偿点较好的拟合是相对应的; PAR为400-1000 μmol·m-2·s-1时, 直角双曲线模型实测点与拟合曲线偏离较大; 对1200-2000 μmol·m-2·s-1高光强部分, 直角双曲线模型和非直角双曲线模型的拟合曲线与光合特征参数(Isat、Pnmax)均与实测值有所差异, 拟合残差大于叶子飘模型, 这是由模型的机制所决定的, 由各模型的公式可以知道, 直角双曲线模型与非直角双曲线模型均是没有极值的方程, 所以只能拟合植物不存在光抑制时的光响应曲线; 叶子飘模型便体现出它独有的优越性, 它存在极值, 可以在不做任何假设的情况下由方程本身直接求出Pnmax与Isat, 文中在此基础上, 用该模型对夏玉米叶片做光合-光响应曲线拟合, 结合残差分析, 表明该模型对夏玉米叶片0-2000 μmol·m-2·s-1各阶段光强均有较好的适应性。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
| [1] |
The kinetics of photosynthesis . |
| [2] |
Gas exchange characteristics ofPopulus trichocarpa, Populus deltoides and Populus trichocarpa × . |
| [3] |
Image super-resolution reconstruction based on residual error .基于残差的图像超分辨率重建. 中国图象图形学报, ( |
| [4] |
Effects of water on photosynthesis in different age of tomato leaves .水分对番茄不同叶龄叶片光合作用的影响 . 生态学报, |
| [5] |
Changes in activity of energy dissipating mechanisms in wheat flag leaves during senescence . |
| [6] |
|
| [7] |
Measuring the impact of flooding on Amazonian trees: Photosynthetic response models for ten species flooded by hydroelectric dams .
|
| [8] |
Light response of photosynthesis and its simulation in leaves of 不同土壤水分下山杏光合作用光响应过程及其模拟. 生态学报, |
| [9] |
Primary application on the non-rectangular hyperbola model for photosynthetic light-response curve .非直线双曲线模型在光合光响应曲线数据分析中的应用. 中国农学通报, |
| [10] |
Fitting net photosynthetic light-response curves with Microsoft Excel — A critical look at the models .
|
| [11] |
Characteristic parameters of light response curves of photosynthesis of winter wheat in north China .华北地区冬小麦光合作用的光响应曲线的特征参数. 应用气象学报, |
| [12] |
Effects of NaCl stress on induction of photosynthesis and PSII photochemistry efficiency of 不同叶龄杂交酸模叶片光合速率及PSII光化学效率诱导对NaCl胁迫的响应. 西北植物学报,
|
| [13] |
Analysis of chlorophyll fluorescence parameters in 不同叶龄拟南芥荧光参数分析及对强光的响应. 草原与草坪,
|
| [14] |
Dark respiration of terrestrial vegetations: A review .陆地植被暗呼吸的研究进展. 应用生态学报, |
| [15] |
Assessment and correction of TMPA products 3B43RT and 3B42V6 .TMPA卫星降水数据的评估与校正. 水利学报, |
| [16] |
|
| [17] |
Dark leaf respiration in light and darkness of an evergreen and a deciduous plant species . |
| [18] |
Photosynthetic characteristics and photo-adaptability of four 不同生态型扁蓿豆光合特性和光适应能力. 生态学杂志, |
| [19] |
Eco-physiological traits of different-age needles of 科尔沁沙地樟子松人工林不同年龄针叶生理生态性状. 植物生态学报, |
| [20] |
A new model for relationship between irradiance and the rate of photosynthesis in
|
| [21] |
A review on modeling of responses of photosynthesis to light and CO2 .光合作用对光和CO2响应模型的研究进展. 植物生态学报, |
| [22] |
Study on biological significance of coefficient in modified model of photosynthesis- irradiance .植物光响应修正模型中系数的生物学意义研究. 扬州大学学报(农业与生命科学版), |
| [23] |
Simulation of the stomatal conductance of winter wheat in response to light, temperature and CO2 changes .
|
| [24] |
Photosynthetic capability of 12 genotypes of 12个不同基因型冬小麦的光合能力. 植物生态学报, |
| [25] |
Effects of temperature variation on the light-response characteristics of tobacco leaf photosynthesis .温度变化对烟草光合作用光响应特征的影响. 生态学杂志, |
| [26] |
Analysis on residual error for adaptive optical system based on stochastic parallel gradient descent control algorithm .基于随机并行梯度下降算法自适应光学系统的校正残差分析. 光学学报, |

/
| 〈 |
|
〉 |

{kind=link}
{kind=link}