版权声明: 2016 植物生态学报编辑部 本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。
基金资助:
展开
摘要
采用盆栽试验模拟干旱胁迫(土壤相对含水量40%-45%)在小麦(Triticum aestivum)拔节孕穗期胁迫12天, 测定其生长速率、光合特征及关键代谢产物含量, 以探讨干旱胁迫对拔节孕穗期小麦叶片初生及次生代谢产物的影响及其涉及的代谢途径, 讨论小麦生长代谢变化规律及应答机制。研究表明: 干旱胁迫使小麦叶片气孔受限制导致光合速率下降; 使叶绿素含量下降直接影响光系统II活性, 最终导致生长率降低。检测出的初级代谢产物组包括有机酸、氨基酸、碳水化合物、嘧啶和嘌呤等64个代谢产物, 其中29个代谢产物在干旱胁迫下发生明显的变化。主成分分析(PCA)结果显示全部样本均分布在95%的置信区间内, 两个主成分得分为64%。单因素方差分析结果表明, 干旱胁迫导致苹果酸、柠檬酸、乌头酸等参与三羧酸(TCA)循环的代谢产物消耗明显, 且引起大部分氨基酸(如脯氨酸、丝氨酸、缬氨酸)和碳水化合物(肌醇、果糖、葡萄糖)大量积累的同时转氨基代谢(天冬酰胺、谷氨酰胺和γ氨基丁酸)产物消耗, 研究证明干旱胁迫明显地促进小麦叶片的糖酵解和氨基酸合成途径, 但抑制了TCA循环和转氨基反应, 加速氨基酸代谢网络向脯氨酸合成转变过程。这些结果表明干旱胁迫引起了转氨基反应、TCA循环、糖酵解/糖异生、谷氨酸介导的脯氨酸合成, 以及嘧啶和嘌呤等代谢网络系统广泛的变化, 说明小麦在合成大量的氨基酸和碳水化合物类物质的同时也消耗了大量的能量, 暗示了糖异生到脯氨酸合成的转变。
关键词:
Abstract
Keywords:
随着工业快速发展、人口迅速增加和用水需求增加, 水资源供需矛盾日益突出, 农业用水更加紧缺, 已成为制约粮食生产的重要因素(赵雅静等, 2009; 康绍忠, 2014)。我国干旱缺水地区占到国土面积的52%左右, 涉及全国25个省、自治区和直辖市, 每年因旱灾粮食减产达1.00 × 1011 kg以上(陈雪, 2015)。因此, 大力发掘和利用对干旱胁迫具有耐性和适应性的优良农作物, 才能做到充分利用土地与自然资源, 稳定发展农业生产。
在农业生态系统中, 理论上, 干旱可能发生在作物的任何生长发育阶段。首先, 植物体受到干旱胁迫时, 个体生长受到抑制, 体内含水量下降致使气孔关闭, 细胞膨压遭到破坏导致生长延伸受到抑制(Jaleel et al., 2008; 张仁和等, 2011)。干旱胁迫打破植物细胞内水分平衡及干扰植物激素的合成, 进而影响到整个代谢网络(Bray, 1997; 吴杨等, 2009)。已有报道显示, 干旱会强烈影响植物光合作用、矿质元素代谢和次生代谢。目前关于植物对干旱响应的研究不断深入, 涉及蛋白激酶(MAPK)、Ca2+依赖型蛋白激酶(CDPK)、超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、组氨酸激酶(HPK)等基于激酶的信号转导途径(赵琳琳等, 2008; Banuelos et al., 2011; 王丽等, 2012)。同样, 蛋白组和转录组相关研究也在不同物种中展开。如在拟南芥(Arabidopsis spp.)中的研究表明, 干旱胁迫引起拟南芥2000多个基因发生改变, 占整个拟南芥基因组的30% (Kreps et al., 2002); 在不同抗旱性的小麦(Triticum aestivum)品种旗叶和籽粒中, 发现干旱胁迫下各品种的小麦多表现为蛋白质带增强, 极少出现新的蛋白质带(徐民俊等, 2002)。然而, 目前植物对干旱的代谢响应研究较少。代谢组学技术应用到干旱的研究中去, 不但可以揭示细胞渗透调节物质的种类, 也可发现植物适应干旱过程的特殊的代谢途径变化, 其结果不但可以为进一步的分子机制研究提供依据和线索, 也可与蛋白组和转录组关联分析, 为进一步确定关键基因奠定基础。
小麦是我国主要的粮食作物, 种植面积占耕地总面积的22%-30%。我国小麦品种分为冬小麦和春小麦, 北方地区是冬小麦的主要种植区, 其小麦产量占全国小麦总产量的71%, 对保障国家粮食安全至关重要。拔节孕穗期是冬小麦由冬前营养生长转变为营养与生殖生长同时并进的时期, 是决定冬小麦最终产量的关键时期(杨文平等, 2008)。因此, 土壤水分的变化势必引起该时期冬小麦生长和生理上的响应。目前, 小麦干旱胁迫下的代谢研究主要集中在根、茎等器官, 研究某个或者某几个代谢产物, 例如脯氨酸、甜菜碱的含量变化, 而全面系统地研究小麦拔节孕穗期干旱胁迫下代谢应答的报道目前还很少。本研究利用气相色谱-质谱(GC-MS)的代谢组学方法结合多元数据统计分析, 系统研究小麦叶片干旱胁迫的代谢应答, 确定关键特征代谢产物及代谢途径, 为小麦栽培和育种提供理论依据。
采用盆栽试验, 盆口直径36 cm, 高30 cm, 底直径30 cm, 每盆装土20.0 kg。选取华北地区常用的冬小麦品种‘石家庄8号’作为供试材料, 试验以壤质潮土为基质, 土壤采自河北衡水实验地, 自然风干后过筛。试验土壤pH值为7.2, 有效N、P和K总含量分别为2.2 mg·kg-1、33.1 mg·kg-1和27.4 mg·kg-1, 平均每盆装16 kg土。用Wilkes法测定盆栽最大土壤持水量为26.6%。
试验在中国农业科学院试验站防雨棚中进行, 试验站位于40.10° N, 116.92° E, 海拔35.19 m, 白天温度一般为20.0-25.5 ℃, 夜间温度一般为18.5- 21.5 ℃, 相对湿度55%。本研究共种植15盆冬小麦,出苗后每天透灌1次, 待生长到三叶期每盆定苗5株。拔节孕穗期时将15盆分为3组, 每组为5盆, 1盆为1个重复。有1组作为处理前基础生物量指标的测试样品, 1组充分灌水为对照组(CK), 1组土壤相对含水量为40%-45%, 连续胁迫处理12天, 为处理组(DS)。采用电子秤(精度0.001 kg)称质量法, 严格控制土壤含水量, 每天称质量一次。土壤相对含水量= (土壤质量–土壤干质量)/(土壤饱和质量-土壤干质量) × 100%。
处理结束时, 将整盆土壤取出, 使用大量清水将根系冲洗干净, 稍后再用蒸馏水洗净全株, 并用吸水纸吸去附着的水分, 将根与地上部分从子叶痕处剪开, 分别称取地上、地下两部分鲜质量并记录。称取部分鲜样用于测定代谢产物。然后将植株置于105 ℃烘箱内杀青15 min, 杀青后转置80 ℃的真空干燥器内干燥至恒质量, 然后测定干质量。
1.3.1 生长指标的测定
相对生长率(RGR) = (ln干旱处理后干质量- ln干旱处理前干质量)/处理天数; 组织含水量(WC) = (鲜质量-干质量)/鲜质量×100% (Kingsbury & Epstein, 1984) 。
1.3.2 光合特性指标的测定
光合特性: 在收获之前利用LI-6400XT便携式光合仪(LI-COR, Lincoln, USA)对倒数第二片叶进行净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr)的测定, 测定时间为9:00-11:00, 利用LED模拟光源, 光量子通量设定为1200 μmol·m-2·s-1 , 每个叶片进行3次重复测量。
荧光特性: 利用IMAGING-PAM荧光仪(Walz, Effeltrich, Germany)测定叶绿素荧光参数: 光系统II (PSII)原初光能转换效率= Fv/Fm、光化学淬灭系数(qP) = (Fm′ - F)/( Fm′ - Fo′)、PSII的表观光合电子传递速率(ETR) = (Fm′ - F)/ Fm′ × PAR × 0.5 × 0.84, 其中, F为光化光照射下打开饱和脉冲前记录的荧光, Fm为充足暗适应后打开饱和脉冲得出的最大荧光, Fv为可变荧光产量, Fm′为作用光打开时的最大荧光产量, Fo′为作用光关闭时的原初荧光产量, PAR为光合有效辐射。测定前, 叶片暗适应20 min, 每个处理测定3个叶片, 所有操作均在背景光强低于1 μmol·m-2·s-1的环境中进行。仪器的饱和脉冲光的强度为2400 μmol·m-2·s-1, 光化光强设为默认值185 μmol·m-2·s-1, 测量光低于0.5 μmol·m-2·s-1 (Hill et al., 2004)。
叶绿素含量: 待荧光测量完毕后, 将叶片采下洗净磨碎, 称取样品0.1 g, 用1:1的丙酮和无水乙醇作为萃取液进行萃取。在波长440、645和663 nm下测定吸光度, 应用Arnon (1949)方程计算叶绿素含量: 叶绿素a含量= 9.784 × A663 - 0.99 × A645; 叶绿素b含量= 21.426 × A663 - 4.65 × A645。
1.3.3 代谢产物检测和筛选
萃取衍生化: 取100 mg小麦叶片样本置于2 mL离心管里, 加入60 μL核糖醇漩涡混匀; 再加入0.35 mL甲醇漩涡混匀。70 Hz 5 min研磨仪处理, 70 ℃烘箱中放置10 min; 然后将样本离心10 min (4 ℃, 12000 r·m-1); 取出0.35 mL上清液放于2 mL进样瓶中(甲烷硅基化)。在真空浓缩器中干燥提取代谢产物(45 ℃, 1.5 h); 向干燥后的代谢产物加入80 μL甲氧胺盐试剂(甲氧胺盐酸盐, 溶于吡啶20 mg·mL-1), 轻轻混匀后, 放入烘箱中37 ℃孵育2 h; 向每个样品中迅速加入100 μL衍生化试剂N,O-双三甲基硅基三氟乙酰胺(BSTFA) + 1%三甲基氯硅烷(TCMS), 将混合物70 ℃下反应1 h, 冷却至室温(Lisec et al., 2006)。
上机检测: 使用Agilent 7890气相色谱仪质谱仪(Agilent, Palo Alto, USA)联机进行GC-MS分析。GC-MS分析条件: DN-5MS毛细管柱(30 m × 250 μm × 0.25 μm, J & W Scientific, Folsom, USA); 进样量1 μL; 载气: 氦气; 前进样口吹扫流速: 3 mL·min-1; 流速: 1 mL·min-1; 柱温: 初始温度90 ℃保持15 s, 以10 ℃·min-1升至180 ℃, 再以5 ℃·min-1升至240 ℃, 再以20 ℃·min-1继续升至285 ℃保持11.5 min。前进样口温度、传输线温度和离子源温度分别为280、245和220 ℃。电离电压: -70 eV; 扫描方式(全扫描): 20-600 m/z; 扫描速度: 100 spectra·s-1; 溶剂延迟: 492 s。
定性定量: 将所得的GC-MS原始数据采取LECO公司的Chroma TOF4.3X软件和LECO-Fiehn Rtx5数据库, 进行峰定性和定量, 使用内标标准化法处理导出的数据, 内标物为核糖醇(纯度≥98%)。利用SIMCA 14.0软件进行分析。
利用SPSS 17.0和Microsoft Excel对数据进行统计和分析, 全部数据是5次重复实验的平均值±标准误差。实验结果用t检验检测统计学差异, 检验水平为5%。利用SIMCA 14.0对代谢产物进行分析, 首先采用主成分分析(PCA)对数据进行总览, 观察样本分布和异常值。相似度> 700、变量重要性投影 (VIP) > 1。
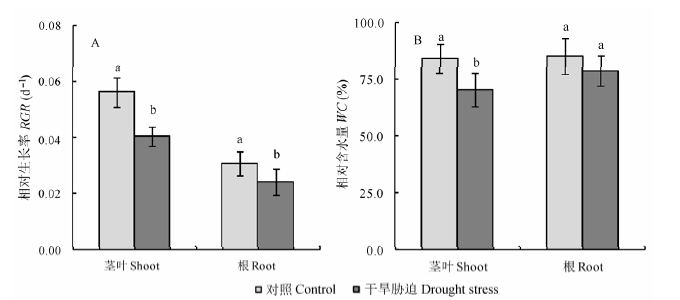
播种180天后小麦拔节孕穗期, 在干旱胁迫下小麦茎叶和根系的RGR和WC均呈现下降的趋势, 而茎叶受影响的程度明显大于根系, 与对照组相比较茎叶中分别下降约28%和16% (图1, p < 0.05)。尽管干旱胁迫也抑制了小麦根的WC, 但其影响并没有达到显著性差异(图1, p < 0.05)。
图1 干旱胁迫对冬小麦地上部分和地下部分相对生长速率(RGR) (A)和含水量(WC) (B)的影响(平均值±标准误差, n = 5)。不同小写字母表示处理间差异显著(p < 0.05, t-test)。
Fig. 1 Effects of drought stress on the relative growth rate (RGR) (A) and relative water content (WC) (B) of shoots and roots of wheat at the jointing-booting stage (mean ± SE, n = 5). Different lowercase letters indicate significant difference between treatments (p < 0.05, t-test).
与对照相比, 干旱胁迫使叶片光合作用受到抑制, 其光合作用参数Pn和Gs下降显著(表1, p < 0.05)。干旱胁迫下Fv/Fm、qP和ETR下降明显, 表明PSII反应中心利用光能的效率和潜在活性受到显著抑制(表1, p < 0.05)。与对照植株相比, 干旱胁迫显著降低了小麦叶片叶绿素含量和Chl a/Chl b (表1, p < 0.05)。
表1 干旱胁迫对冬小麦光合参数(Pn, Gs), 荧光参数(Fv/Fm, qP, ETR)和叶绿素含量(Chl a, Chl b, Car)的影响(平均值±标准误差, n = 5)
Table 1 The photosynthetic indices (Pn, Gs), chlorophyll fluorescence (Fv/Fm, qP, ETR), chlorophyll and carotenoid contents and Chl a/Chl b of wheat seedling leaves under drought stress at the jointing-booting stage (mean ± SE, n = 5)
| 水分处理 Water treatment | 光合参数 Photosynthetic indices | 荧光参数 Chlorophyll fluorescence | 叶绿素含量 Chlorophyll content | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 净光合速率 Pn (μmol·m-2·s-1) | 气孔导度 Gs (μmol·m-2·s-1) | PSII原初光能 转换效率 Fv/Fm | 光化学 淬灭系数qP | PSII的表观光合 电子传递速率 ETR | 叶绿素含量 Chlorophyll contents (g·kg-1 fresh mass) | 叶绿素a/ 叶绿素b Chl a/Chl b | |||
| 对照 Control | 14.98 ± 0.63a | 0.29 ± 0.01a | 0.73 ± 0.02a | 0.22 ± 0.05a | 28.28 ± 3.36a | 4.29 ± 0.88a | 2.16 ± 0.23a | ||
| 干旱胁迫 Drought stress | 3.15 ± 0.37b | 0.04 ± 0.00b | 0.31 ± 0.03b | 0.09 ± 0.00b | 12.49 ± 1.33b | 2.01 ± 0.63b | 1.31 ± 0.14b | ||
根据单因素方差分析结果, 在拔节孕穗期干旱胁迫下小麦叶片和对照相比有30个代谢产物发生了明显的变化。总体而言, 21个代谢产物显著增加, 9个代谢产物显著减少(表2, p < 0.05)。在三羧酸(TCA)循环中干旱胁迫促使草酸、异柠檬酸、延胡索酸和苹果酸的积累, 但却抑制柠檬酸和乌头酸的合成(表2, p < 0.05)。干旱胁迫使参与糖酵解的代谢产物明显增加, 包括磷酸烯醇式丙酮酸、葡萄糖-6-磷酸、果糖-6-磷酸和葡萄糖, 结果表明干旱胁迫明显促进了叶片中糖酵解代谢过程(表2, p < 0.05)。与对照相比, 干旱胁迫下除了蔗糖含量下降, 叶片中大多数的糖含量呈现上升趋势, 包括果糖、海藻糖、龙胆二糖和景天庚糖(表2, p < 0.05)。与此同时,干旱胁迫下叶片中也积累大量的氨基酸, 包括脯氨酸、天冬酰胺、甘氨酸、丝氨酸、谷氨酸、缬氨酸、异亮氨酸和亮氨酸(表2, p < 0.05)。但是干旱胁迫导致小麦叶片转氨基代谢产物, 如天冬酰胺、谷氨酰胺和γ-氨基丁酸, 以及核苷酸衍生物, 如胸腺嘧啶和次黄嘌呤含量大量消耗 (表2, p < 0.05)。
表2 拔节孕穗期干旱胁迫下小麦叶片提取物中代谢产物相对含量和变化倍数
Table 2 Relative concentration and the change of major metabolites in leaves of wheat seedlings under drought stress (DS) treatment at the jointing-booting stage
| 代谢通路 Metabolic Pathway | 代谢产物 Metabolite | 相对含量 Relative concentration | 变化率 Rate of change Log2 (DS/CK) | ||
|---|---|---|---|---|---|
| 对照组 Control (CK) | 干旱胁迫组 DS | ||||
| 三羧酸循环 The tricarboxylic acid cycle | 草酸 Oxalic acid | 1.85 | 1.03 | -0.84 | |
| 柠檬酸 Citric acid | 6.39 | 2.24 | -1.51* | ||
| 乌头酸 Aconitic acid | 3.37 | 1.65 | -1.03* | ||
| 异柠檬酸 Isocitric acid | 1.01 | 0.64 | -0.66 | ||
| Α-酮戊二酸 α-Ketoglutaric acid | 0.31 | 0.12 | -1.37* | ||
| 琥珀酸 Succinic acid | 1.46 | 0.99 | -0.56 | ||
| 延胡索酸 Fumaric acid | 0.59 | 0.27 | -1.13* | ||
| 苹果酸 Malic acid | 44.28 | 15.78 | -1.49* | ||
| 糖酵解 Glycolysis | 丙酮酸 Pyruvate | 0.36 | 0.42 | 0.21 | |
| 磷酸烯醇式丙酮酸 Phosphoenolpyruvate | 0.00 | 0.00 | 1.52* | ||
| 葡萄糖-6-磷酸 Fructose-6-phosphate | 0.00 | 0.01 | 1.05* | ||
| 果糖-6-磷酸 Glucose-6-phosphate | 0.05 | 0.11 | 1.09* | ||
| 葡萄糖 Glucose | 0.02 | 0.05 | 1.41* | ||
| 氨基酸类 Amino acids | 脯氨酸 Proline | 1.76 | 58.64 | 5.06* | |
| 丙氨酸 Alanine | 3.13 | 2.93 | -0.10 | ||
| 苯丙氨酸 Phenylalanine | 0.88 | 1.14 | 0.38 | ||
| 天冬氨酸 Aspartic acid | 14.38 | 6.41 | -1.17* | ||
| 天冬酰胺 Asparagine | 0.11 | 0.39 | 1.83* | ||
| 甘氨酸 Glycine | 0.26 | 0.53 | 1.03* | ||
| 丝氨酸 Serine | 5.90 | 11.90 | 1.01* | ||
| 苏氨酸 Threonine | 0.01 | 0.01 | 0.59 | ||
| 谷氨酸 Glutamate | 0.47 | 1.41 | 1.60* | ||
| 谷氨酰胺 Glutamine | 1.41 | 0.37 | -1.93* | ||
| 缬氨酸 Valine | 3.62 | 12.09 | 1.74* | ||
| 半胱氨酸 Cysteine | 0.03 | 0.05 | 0.72 | ||
| 异亮氨酸 Isoleucine | 1.61 | 7.09 | 2.14* | ||
| 亮氨酸 Leucine | 0.36 | 1.52 | 2.06* | ||
| 赖氨酸 Lysine | 0.46 | 0.48 | 0.07 | ||
| 甲硫氨酸 Methionine | 0.50 | 0.38 | -0.40 | ||
| 色氨酸 Tryptophan | 0.05 | 0.05 | 0.05 | ||
| 酪氨酸 Tyrosine | 0.01 | 0.01 | -0.12 | ||
| 鸟氨酸 Ornithine | 0.22 | 0.19 | -0.18 | ||
| 瓜氨酸 Citrulline | 0.06 | 0.10 | 0.85 | ||
| 糖类及多元醇 Sugars and polyols | 蔗糖 Sucrose | 12.77 | 6.37 | -1.00* | |
| 果糖 Fructose | 1.09 | 2.21 | 1.03* | ||
| 半乳糖 Galactose | 0.19 | 0.29 | 0.63 | ||
| 木糖 Xylose | 0.06 | 0.09 | 0.62 | ||
| 海藻糖 Trehalose | 0.02 | 0.04 | 0.97* | ||
| 肌醇 Myo-inositol | 15.06 | 30.85 | 1.03* | ||
| 甘露糖 Mannose | 21.27 | 20.53 | -0.05 | ||
| 纤维二糖 Cellobiose | 0.04 | 0.03 | -0.48 | ||
| 阿卓糖 Altrose | 1.03 | 1.34 | 0.39 | ||
| 葡庚糖 Glucoheptose | 0.38 | 0.48 | 0.35 | ||
| 夫糖 Fucose | 0.08 | 0.09 | 0.13 | ||
| 表2 (续) Table 2 (continued) | |||||
| 代谢通路 Metabolic Pathway | 代谢产物 Metabolite | 相对含量 Relative concentration | 变化率 Rate of change Log2 (DS/CK) | ||
| 对照组 Control (CK) | 干旱胁迫组 DS | ||||
| 糖类及多元醇 Sugars and polyols | 半乳糖苷 Galactinol | 1.57 | 1.34 | -0.23 | |
| 龙胆二糖 Gentiobiose | 0.09 | 0.20 | 1.14* | ||
| 乳糖 Lactose | 0.07 | 0.05 | -0.68 | ||
| 苏糖 Threose | 0.03 | 0.03 | 0.15 | ||
| 来苏糖 Lyxose | 20.14 | 27.01 | 0.42 | ||
| 景天庚糖 Sedoheptulose | 0.02 | 0.04 | 1.04* | ||
| 塔格糖 Tagatose | 1.74 | 2.73 | 0.65 | ||
| 核苷酸衍生物 Nucleotides derivatives | 尿嘧啶 Uridine | 0.03 | 0.03 | -0.06 | |
| 胸腺嘧啶 Thymidine | 1.82 | 0.68 | -1.42* | ||
| 鸟嘌呤 Guanosine | 0.27 | 0.17 | -0.65 | ||
| 次黄嘌呤 Hypoxanthine | 0.25 | 0.11 | -1.14* | ||
| 有机酸及其他代谢产物 Organic acids and others | γ-氨基丁酸 γ-aminobutyric acid | 28.23 | 15.43 | -0.87* | |
| 莽草酸 Shikimic acid | 11.89 | 12.61 | 0.08 | ||
| 奎尼酸 Quinic acid | 2.77 | 2.70 | -0.04 | ||
| 乙醇酸 Glyceric acid | 1.34 | 1.42 | 0.09 | ||
| 乙醇胺 Ethanolamine | 0.81 | 0.36 | -1.16* | ||
| 肉桂酸 Cinnamic acid | 0.03 | 0.04 | 0.25 | ||
| 绿原酸 Chlorogenic acid | 0.28 | 0.18 | -0.66 | ||
| 阿魏酸 Ferulic acid | 0.06 | 0.06 | 0.12 | ||
| 琥珀酸半醛 Succinate semialdehyde | 0.04 | 0.06 | 0.63 | ||
RGR和WC可反映植物体受到外界环境胁迫时的基本生理反应(Yang et al., 2008)。本实验结果表明, 干旱胁迫下小麦茎叶和根系的RGR和WC受到了明显的抑制, 且对茎叶的影响明显大于根系, 这说明小麦地上、地下部分对干旱胁迫可能有着不同的响应方式(图1)。干旱胁迫导致小麦叶片中叶绿素含量降低, 使得光系统II原初光能转换效率及光系统II潜在活性受到抑制, 干旱胁迫直接影响了光合作用的电子传递和CO2同化过程(表2) (Lawlor & Cornic, 2002; 吴永成等, 2004)。
在长期的进化过程中, 植物生存的土壤和气候条件并非一成不变, 植物甚至经常遭受极端的气候和土壤条件。植物可能已经进化出多种复杂的代谢调节机制, 其适应胁迫的代谢响应可能是迅速和多样的, 甚至动员多个或者全部主要的代谢途径。代谢组学分析技术可以全面地揭示植物响应胁迫的特殊的代谢调控网络。当前研究利用GC-MS代谢组学分析方法, 系统描述干旱胁迫下代谢产物组成的变化以及变化过程中涉及的作物适应干旱环境的可能的重要代谢途径。研究结果表明, 干旱胁迫引起小麦代谢组改变, 包括许多代谢过程如光合作用、TCA循环、糖酵解/糖异生和氨基酸代谢。
干旱胁迫诱导植物自身产生大量活性氧(ROS) 激活蛋白酶, 导致植物细胞内部产生过量的NH4+, 植物细胞通常会把多余的NH4+通过转氨作用转化成谷氨酸/谷氨酰胺, 或者通过谷氨酸脱氢酶转化为α-酮戊二酸进入TCA循环 (Cui et al., 2008; Daiet al., 2010)。本研究结果表明, 在拔节孕穗期小麦叶片中的脯氨酸是干旱胁迫12天条件下变化最为明显的代谢产物, 与对照组相比较积累超过25倍, 约占总游离氨基酸的55% (表2)(吕丽华等, 2006; 姜淑欣等, 2014)。而脯氨酸的增长伴随着转氨作用的产物谷氨酰胺和天冬氨酸含量降低以及天冬酰胺的增加, 这种变化与氨基酸代谢网络向脯氨酸合成转变过程相一致。胸腺嘧啶和次黄嘌呤含量的下降以及谷氨酸的增加, 可能暗示了嘧啶和嘌呤代谢产物通过鸟氨酸循环转化成谷氨酸, 大量的谷氨酸通过Δ1-吡咯啉-5-羧酸转化为脯氨酸进而促进脯氨酸的合成(表2)(LaRosa et al., 1991)。甜菜碱是高等植物在逆境下的另一种重要的有机渗透调节物质, 本研究并没有在小麦叶片中发现甜菜碱及其生物合成前体, 这可能是因为甜菜碱是短期应答代谢反应, 但是干旱胁迫抑制甜菜碱合成酶活性导致其含量降低。缬氨酸、异亮氨酸和亮氨酸属于支链氨基酸, 是参与丙酮酸代谢相关的生糖氨基酸, 它们的明显增加可能作为减少转氨基产物的额外途径, 另外它们还是多酚类化合物的前体, 可以作为内源性植物抗氧化剂(表2)(王镜岩等, 2002)。丝氨酸和甘氨酸可能与糖酵解途径中的转氨基产物有关, 因此干旱胁迫下它们含量的增加有利于促进糖酵解过程, 进而增加小麦的抗旱性(表2)。此外, 我们的研究表明干旱胁迫明显抑制一种非结构氨基酸-γ-氨基丁酸积累, 这可能是一种适应性响应, 降解非结构氨基酸可能是为了进一步节省氮源, 保证蛋白质的合成。
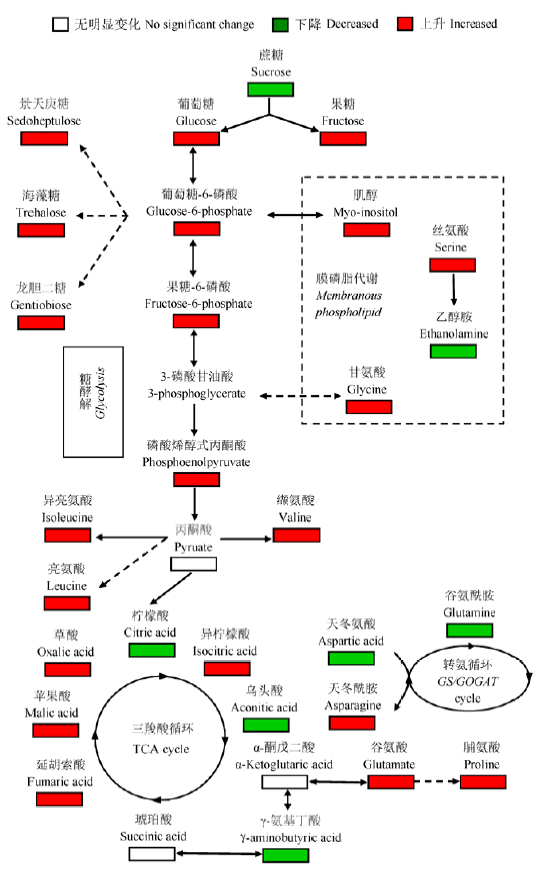
在植物细胞中糖类主要来自光合作用、糖异生途径和多糖降解。在本研究中干旱胁迫下小麦叶片中蔗糖含量显著性下降, 但葡萄糖、果糖、海藻糖等含量显著性上升, 结合小麦光合特性结果, 干旱胁迫使小麦光合速率显著降低并限制了氮代谢导致蔗糖含量降低, 同时, 加速蔗糖水解为葡萄糖和果糖, 不但可以降低细胞水势, 也可为细胞的各种化合物代谢合成提供碳骨架和能量。肌醇是最为有效的羟自由基清除剂, 在清除干旱条件下产生的毒害物质方面起到重要作用(侯彩霞和汤章城, 1999)。本研究发现, 肌醇在总可溶性碳水化合物中所占比例较大, 且含量在干旱胁迫下显著增加, 这说明干旱胁迫下肌醇是小麦的主要渗透调节物质, 同时它也可以被看作是一种比较好的碳储存物。研究还显示小麦叶片中乙醇胺含量在干旱胁迫条件下明显降低, 乙醇胺是植物细胞质膜的重要代谢产物, 它的降低可能反映了小麦细胞膜降解的抑制。研究发现, 在拔节孕穗期干旱胁迫导致小麦叶片中的苹果酸、柠檬酸、乌头酸、α-酮戊二酸和延胡索酸显著减少, 这表明了干旱胁迫明显抑制了TCA循环, 暗示了小麦在受到干旱胁迫时消耗大量的能量(表2; 图2)。但是与此同时, 磷酸烯醇式丙酮酸、葡萄糖-6-磷酸和果糖-6-磷酸大量积累, 表明干旱胁迫可能增强小麦的糖酵解途径, 进而促进糖类物质积累, 不但可以降低水势, 也可能为其他类型的适应性响应提供物质和能量供应(图2)。
图2 干旱胁迫下小麦叶片代谢途径网络变化图。红色代表显著性增加, 绿色代表显著性降低 (p < 0.05)。
Fig. 2 Proposed changes of metabolic network of wheat leaves under drought stress. Red boxes denote significant increases while green ones denote significant decreases (p < 0.05).
致谢 感谢上海阿趣生物公司科技有限公司邓军亮先生提供的数据测量和分析的帮助。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
| [1] |
Copper enzymes in isolated chlorop lasts phenoloxidases in
|
| [2] |
Selenium accumula- tion, distribution, and speciation in spineless prickly pear cactus: A drought- and salt-tolerant, selenium-enriched nutraceutical fruit crop for biofortified foods . |
| [3] |
Plant responses to water deficit . |
| [4] |
Effects of Drought Stress on Growth, Yield and Quality of Different Barley Genotypes. Master degree dissertation, Zhejiang University ,干旱胁迫对不同大麦生长发育、产量和品质的影响 . |
| [5] |
Metabolite identification via the madison metabolomics consortium database . |
| [6] |
Combined NMR and LC-MS analysis reveals the metabonomic changes in |
| [7] |
Spatial heterogeneity of photosynthesis and the effect of temperature-induced bleaching conditions in three species of corals . |
| [8] |
Function and mechanism of com- patible solutes .细胞相容性物质的生理功能及其作用机制. 植物生理学通讯, |
| [9] |
Differential responses in water use efficiency in two varieties of |
| [10] |
Effects of PEG stress and recovery on activities of key enzymes involved in proline metabolism in wheat cultivars with difference in drought tolerance .PEG胁迫及复水对不同抗旱性小麦幼苗脯氨酸代谢关键酶活性的影响. 西北植物学报, |
| [11] |
Towards water and food security in china .水安全与粮食安全. 中国生态农业学报, |
| [12] |
Selection for salt resistant in spring wheat . |
| [13] |
Transcriptome changes for |
| [14] |
Elevated accumulation of proline in NaCl-adapted tobacco cells is not due to altered 1-pyrroline-5- carboxylate reductase . |
| [15] |
Photosynthetic carbon assimila- tion and associated metabolism in relation to water deficits in higher plants . |
| [16] |
Gas chromatography mass spectrometry-based metabolite profiling in plants . |
| [17] |
Dynamics of proline accumulation in winter wheat under different water stress .水分胁迫下不同抗旱性冬小麦脯氨酸积累动态. 华北农学报, |
| [18] |
|
| [19] |
Drought stress signal transduction and regulation mechanism in plants .植物干旱胁迫信号转导及其调控机制研究进展. 生物技术通报, |
| [20] |
Advances in regulatory metabolic networks in drought stress responses .植物干旱胁迫下的调控代谢网络研究进展. 湖北农业科学, |
| [21] |
Review on genetic im- provement in root related to drought-resistance in wheat .小麦 |
| [22] |
|
| [23] |
|
| [24] |
Comparative effects of salt stress and alkali stress on growth, osmotic adjustment and ionic balance of an alkali resistant halophyte |
| [25] |
Effects of soil drought on carbon metabolism of winter wheat during jointing stage .土壤干旱对冬小麦拔节期叶片碳代谢的影响. 河南农业科学, |
| [26] |
Effects of drought stress on photosynthetic traits and protective enzyme activity in maize seeding .干旱胁迫对玉米苗期叶片光合作用和保护酶的影响. 生态学报, |
| [27] |
The mitogen- activated protein kinase signal transduction in plant cell under biotic and abiotic stress conditions .生物和非生物胁迫下的植物细胞中丝裂原活化蛋白激酶(MAPK)信号转导. 植物生理学通讯, |
| [28] |
Physiological and ecological responses of plants to drought stress and its research progress .植物对干旱胁迫的生理生态响应及其研究进展. 福建稻麦科技,
|

/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}