“胎生”一词起源于哺乳动物, 然而在植物中也发现了类似的繁殖方式, 其定义为后代在母体上萌发并生长至一定阶段才离开母体(Elmqvist & Cox, 1996 )。根据这个定义, 胎生可分为真胎生与假胎生。真胎生是指通过有性生殖产生的后代在母体萌发的现象; 而假胎生是指植物通过无融合生殖或在有性生殖器官上产生无性繁殖体的现象, 与营养繁殖类似。真胎生与假胎生有着遗传和进化上的本质区别, 应当区分开来。
胎生现象在植物界中以红树植物最为常见。红树植物是指生活在热带、亚热带海岸潮间带滩涂的木本植物。红树植物的胎生类型有显胎生(vivipary)和隐胎生(cryptovivipary)两种。显胎生是指繁殖体(主要是下胚轴)突破了种皮和果皮两层结构并伸出果实外的过程, 主要集中在红树科的红树属(Rhizophora )、秋茄树属(Kandelia )、角果木属(Ceriops )和木榄属(Bruguiera )。隐胎生是指种子发育只突破了种皮但仍包被在果实内的现象, 如分布在不同科的海榄雌属(Avicennia )、蜡烛果属(Aegiceras )、水椰属(Nypa )和Pelliciera 。胚胎生长到一定阶段后, 繁殖体从树上掉落到泥土中, 就地生长或随海水飘到异地。繁殖体储存了大量营养物质, 能有效地克服落地后滩涂的恶劣环境, 如海浪冲击、高盐、淹水等, 在接触到松软的泥土后, 能在短时间内定植, 继续生长发育。显胎生和隐胎生的胚胎虽有多个胚珠能受精, 但通常只有一个或偶见两个繁殖体能发育成功。而非胎生红树植物如海桑属(Sonneratia )是多种子果实, 果实掉落后在滩涂上开裂, 种子散出后才开始发育(王文卿和王瑁, 2007 )。
红树植物生长的环境是海洋与陆地交错的潮间带, 具有海洋生态系统和陆地生态系统的双重特性。在这样的生境中, 红树植物胎生繁殖方式具有重要的环境适应性, 因而引起众多植物学家和生态学家的关注。红树植物的胚胎发育在形态发育、生理生化、分子水平、生态适应性4个层次上均进化出与传统繁殖方式不同的特征, 以此提高对海岸潮间带复杂生境的适应性, 有助于幼苗落地后成功定植。目前关于红树植物胎生发育的研究在4个层次都有涉及, 但仍有诸多问题没有解决, 本文将在这4个层次上进行综述与展望。
1 形态发育比较
大部分陆生植物种子发育成熟后往往要经历一个脱水、休眠的阶段(Angelovici et al ., 2010 ), 脱离母体后, 待外界条件如温度、湿度适宜时种子才逐渐萌发, 利用贮存的碳水化合物、脂肪和蛋白质等大分子化合物水解产生的小分子物质和能量供应幼苗生长发育。胎生植物则不然, 它们的种子没有休眠期或休眠期很短, 直接在母体植株的果实中萌发, 形成胚芽、下胚轴、胚根结构, 脱离母体定植后继续生长。这种繁殖模式显然与它们生活的热带、亚热带温暖、湿润的气候环境有关。
真胎生现象在4类植物中已有报道(Farnsworth, 1997 )。首先是在遗传上以胎生为繁殖方式的植物, 如分布在8个属的红树植物(表1 )及两个属的海草(Tomlinson, 1986 ), 它们在自然生长环境条件下以胎生为最主要的繁殖方式, 繁殖体在母体上突破种皮或果皮并生长; 二是生长于热带、亚热带的某些顽拗性种子植物, 如少数仙人掌科植物(Cota-Sánchez, 2004 ); 三是某些处于特殊环境中的植物, 如水稻(Oryza sativa )、玉米(Zea mays )等, 其种子在高温、高湿条件下提前萌发, 但它们不从母体中吸收营养(叶能干, 1979 ); 四是与休眠有关的植物激素脱落酸(ABA)合成或响应通路有关的突变体植株, 或因种子中休眠基因功能的缺失不能进入休眠期而在母体直接开始萌发(Robertson, 1955 ), 后两类植物由于受外界条件诱导而形成提前萌发的性状, 在遗传本质上不是真正意义上的胎生, 但对于红树植物胎生现象研究仍有一定的借鉴作用。表1 列出已发表的以胎生为繁殖方式的植物。
1.1 显胎生红树植物胚胎发育过程
红树植物胎生过程包含胚胎形成和胚胎萌发两个过程。以显胎生为繁殖策略的红树胚胎发育又可分为3个阶段: 子叶的第一次生长; 下胚轴的生长; 子叶的第二次生长(Cook, 1907 )。红树属、秋茄树属、角果木属和木榄属4个属间的发育各有特点, 在进化地位上有所差异。
首先是子叶的形态不同。木榄属的子叶是二裂或三裂, 厚肉质, 成熟时不与下胚轴分开, 一直包被在果皮内与之同时掉落。角果木属、红树属及秋茄树属的子叶都是合生性, 呈圆柱体形, 前两个属子叶中的维管组织比秋茄树属的多(Juncosa & Tomlinson, 1988 )。待胚轴成熟, 子叶进行第二次生长后, 子叶基部的细胞分裂产生一个离区, 有助于下胚轴与子叶分离、掉落, 而子叶及果皮仍留在母体上(Juncosa, 1982 )。其次是促使突破种皮的原因也不一样。早期文献报道除木榄属的胎生发育是由于下胚轴伸长外, 红树属、角果木属、秋茄树属中都是由大量生长的胚乳突破种皮的(Juncosa & Tomlinson, 1988)。在种皮被突破时, 胚仍处于较早期的阶段(在红树(Rhizophora apiculata )中甚至处于心形胚, 图1A ), 即由于胚乳的增加, 胚孔被打开, 胚吸收胚乳的营养, 开始生长。我们最近的研究发现秋茄树(Kandelia obovata )的胎生发育不是像Juncosa和Tomlinson (1988)所描述的依靠胚乳的大量生长突破种皮, 而是由于下胚轴的伸长, 即由位于茎尖分生组织下方的居间分生组织细胞进行分裂生长而成(图1F )。因此, 在秋茄树、木榄属中, 胎生萌发起始点为因下胚轴伸长使胚根伸出种皮的时刻, 这与非胎生的种子发育相似。胚乳在这一过程中的作用并不像在红树属中那样显著, 类似地, 水稻突变体endospermless1 (enl1 )缺乏胚乳, 表现出胎生现象(Hara et al ., 2015 )。胚乳存在与否及其功能可能影响胎生发育, 由此我们推测在红树植物中显胎生发育可能存在两种机制。一旦开始发育, 在这4个属中, 下胚轴的伸长均是居间分生组织细胞的快速分裂与细胞伸长导致的, 有的胚轴能达几十厘米悬挂在母树上。
图1 显胎生的红树(A-C)与秋茄树(D-F)胚胎发育过程比较, 表明红树由于胚乳的大量生长而突破种皮, 秋茄树中种皮的打开则是依靠下胚轴的伸长。A, 心形胚阶段, 此时种皮明显因胚乳的大量生长而打开。B, 胚开始分化, 上端为子叶, 下端将分化出茎、下胚轴、根等结构。C, 分化出明显的茎尖分生组织, 胚轴进入快速伸长阶段。D, 秋茄树胚的生长首先集中在子叶部位, 随后分化出下胚轴, 此时未见胚乳, 胚孔端为连接种子和胎座的珠柄细胞, 外种皮有很多单宁成分, 内种皮则细胞质更加丰富。E, 比D阶段靠后, 但胚未伸出种皮。F, 胚伸出种皮, 下胚轴分化出茎尖分生组织, 由于居间分生组织的生长, 胚轴将迅速伸长。a, 胚轴; co, 子叶; em, 胚; en, 胚乳; f, 珠柄; i, 种皮; ii, 内种皮; IM, 居间分生组织; oi, 外种皮; SAM, 茎尖分生组织。
Fig. 1 Comparisons of embryo developmental processes between true-viviparous mangroves Rhizophora apiculata (A-C) and Kandelia obovata (D-F), showing that breaking of the seed coat in R . apiculata is due to the massive growth of the endosperm while it is because of hypocotyl elongation in K . obovata . A, Embryo in the heart-shaped stage, in which the integument is opened by the intrusive growth of the endosperm. B, The embryo starts to differentiate, and cotyledon is on the upper, while shoot, hypocotyl and root are below. C, An embryo in rapid elongation stage because of cell division and elongation of the intercalary meristem zone below shoot apical meristem. D, The embryo first grows in the fused cotyledon; also shown is the axis; no endosperm is seen at this stage. The funicle connects the seed and the placenta. E, A stage later than D, but the whole embryo is still enclosed by integument. F, The embryo (axis) protrudes the seed by the growth of the intercalary meristem, and the embryo will enter a rapid growth stage. a, axis; co, cotyledon; em, embryo; en, endosperm; f, funicle; i, integument; ii, inner integument; IM, intercalary meristem; oi, outer integument; SAM, shoot apical meristem.
另外, 在显胎生红树植物发育的过程中, 有许多与功能相适应的结构、特征引起人们的注意: 首先是在红树属中, 与种皮相邻的最外层胚乳细胞特化出向内皱褶的壁, 被称为转运组织, 通过转运细胞, 水分和养料得以从种皮向胚转运, 但随着胚的不断生长, 这层转运细胞被挤压最后失去功能。在胎生海草Amphibolis 和Thalassodendron 中发现连接幼苗和母体的基部组织最外层也存在着运输养分的转运组织(Kuo & Kirkman, 1990 )。其次是胚轴在母体上生长时主根并不分化, 而是由数量较多的侧根进行分化, 并且在胚轴落地后的定植过程中起锚定作用。最后, 红树科中除了以显胎生为繁殖方式的4个属生活在滨海潮间带外, 其他的类群均为陆生非胎生种(Juncosa & Tomlinson, 1988 )。同时, Juncosa和Tomlinson (1988)认为在红树科中, 显胎生的许多繁殖特征, 如果实不开裂, 合生性子叶, 具有突出种子的胚乳, 由胚乳起始的发育过程, 胚乳中特化的转运细胞, 由侧根而不是主根进行胚轴(幼苗)定植等都是衍生的特征。由胚珠、种子、果实的形态特征推测显胎生的4个属是逐渐进化而适应水环境传播的(Tobe & Raven, 1988 )。
图2 显胎生、隐胎生与非胎生红树植物胚胎发育过程对比示意图。A-E为红树显胎生胚胎发育图。A, 胚仍在种皮内。B, 胚乳突破种皮, 胚开始快速分化生长。C, 合生性的子叶充满了种子内部, 下胚轴分化明显并开始伸长。D, 胚胎突破了种皮进行生长, 但仍在果实内部。E, 下胚轴突破果皮。F-J为秋茄树显胎生胚胎发育图。F, 胚胎呈线形且绿色, 并有一定程度的弯曲。G, 合生性的子叶占据了种子内的大部分空间, 下胚轴开始伸长。H, 由于下胚轴的伸长胚突破种皮。I, 与红树D时期相似。J, 与红树E时期相似。K-N为海榄雌隐胎生胚胎发育图。K, 胚乳与早期的胚在胚珠内生长。L, 胚孔吸根萎缩, 胚乳连同心形胚进入子房腔内生长。M, 折叠的子叶包裹着胚轴, 此时种子成熟, 但仍包被在果皮内。N, 种子落地后萌发。O, P分别为无瓣海桑非胎生胚胎发育的球形胚和心形胚阶段。Q, 离开母体植株后的萌发种子, 此前, 胚胎不突破种皮或果皮。a, 胚轴; co, 子叶; em, 胚; en, 胚乳; i, 种皮; mh, 胚孔吸根; p, 果皮; pl, 胚芽; r, 根; SM, 茎尖分生区。各图没有按比例尺绘图。
Fig. 2 Comparative illustrations in embryo developments in true-vivipary, crypto-vivipary and non-vivipary in mangrove plants. A-E represent true-vivipary embryo development in Rhizophora apiculata . A, The embryo is still enclosed in the integument. B, Integument has been forced open by large amount of endosperm. C, The embryo sac is filled with fused cotyledon, and axis starts its growth. D, The embryo grows out of the integument while still inside the fruit. E, The hypocotyl elongates out of the pericarp. F-J represent the true-vivipary embryo development in Kandelia obovata . F, After fertilization, when the liner embryo lays in the seed and curves to a certain extent. G, The fused cotyledon occupies the majority of the seed while axis starts to extend. H, The axis extends out of the integument. I, The same stage as D of R . apiculata . J, The same stage as E of R . apiculata . K-N, Crypto-vivipary embryogenesis and seed germination in Avivennia marina . K, Endosperm surrounds the globular pro-embryo. L, Micropylar haustorium becomes atrophied, and then the endosperm and the heart-shaped embryo grow out of the ovular tissue, with subsequent embryo development occurring outside of the ovule. M, Mature seed with folded cotyledons and embryonic axis while closely contiguous with the pericarp. N, Seed germinate in the intertidal mud. O, P, Non-vivipary embryo development in Sonneratia apetala . O, In globular stage. P, In heart-shaped stage. Q, Germinating seed after leaving the parent plant. a, axis; co, cotyledon; em, embryo; en, endosperm; i, integument; mh, micropylar haustorium; p, pericarp; pl, plumule; r, root; SM, shoot meristem. Figures are not drawn to scale.
1.2 隐胎生与非胎生红树植物胚胎发育过程
隐胎生红树植物的胚胎发育方式与显胎生有所差别。海榄雌(Avicennia marina )胚胎发育至球形胚时, 胚孔吸根(micropylar haustorium)萎缩, 胚乳连同胚从珠被(种皮)中伸出, 进入到子房腔生长, 胚进而充满子房, 成熟时果实掉落(Farrant et al ., 1992 )。在蜡烛果属植物Aegiceras majus 中的情况较为不同, 胚珠受精后具有分生性的胚乳先行分裂, 待大量胚乳形成后, 合子才在胚孔端开始发育(Carey & Fraser, 1932 )。胚乳在胚孔处的分裂产生皱褶, 这些皱褶增加了吸收养分的面积, 相当于吸器的功能。经过一段时间的生长后, 胚占主导地位, 为了适应胚的这一变化, 胎座也伸长以维持胚的相对位置不变, 并保证了养料的持续供给, 而后胚根及胚轴的下部伸出种皮。随着胚逐渐增大, 种皮形成了类似帽子的结构, 罩在子叶上端。果实连同发育了的胚一起掉落, 在适当的条件下胚轴继续伸长, 直至胚根突破果皮, 侧根生长在松软的泥土上完成定植, 子叶也从果实中露出, 与包裹在其中的幼叶一起进行光合作用, 幼苗开始生长。
非胎生红树植物无瓣海桑(Sonneratia apetala ) 是多种子果实, 其胚胎发育过程与传统的荠菜胚一致, 但胚囊的中央部分始终不被胚乳填满, 胚乳在胚孔端有极性积累, 且到后期胚乳完全被胚吸收, 为无胚乳种子(Venkateswarlu, 1937 )。果实成熟后从树上掉落, 果皮开裂, 释放种子。
红树植物发育过程中的特化结构具有独特的功能适应性, 如转运细胞、居间分生组织、胚柄、吸器等。显胎生与显胎生、隐胎生与隐胎生、显胎生与隐胎生、显胎生与隐胎生、非胎生之间的不同之处表明了红树植物丰富的生物多样性, 对其发育机制的深入理解不仅有利于追溯胎生的起源进化, 而且对探究以不同发育方式适应相似生境的过程也有重要意义。
2 生理生化水平
2.1 胚胎的含水量
胎生红树植物胚胎发育具有两个重要的特点, 首先是种子不经脱水且不经休眠或短暂休眠就开始发育生长。其次, 研究发现胎生植物繁殖器官都有很高的含水量, 胚胎、种皮、果皮含水量均高于54% (Sussex, 1975 ), 在发育过程中Rhizophora mangle 的胚胎含水量始终高于种皮和果皮, 充足的水分供应为发育和新陈代谢提供了必要条件。相反, 进行休眠的种子成熟后干燥水分会降低至8%左右(Vertucci, 1989 ), 以便于储藏, 但新陈代谢、生长发育各个生理过程均处于停止状态, 等待新水源与其他条件成熟以后再行萌发, 这样可以避免冬天或干旱等不利的季节条件。由于红树植物生长在环境水分、温度都适宜生长的地方, 它们进化出不用经过休眠的方式来适宜这种环境, 以期得到最高的生长量和最短的繁殖周期。
2.2 胚胎发育过程中繁殖器官盐离子动态
胎生红树植物胚胎发育的另一个重要的特征是发育过程中繁殖器官盐离子浓度的变化。对于胚胎如何适应落地后的高盐生境, 有两种不同的结论, 即胎生是低盐化过程, 以及与此对立的胎生是盐分累积过程。郑文教和林鹏(1997)测定了秋茄树、木榄(Bruguiera gymnorhiza )和海莲(Bruguiera sexangula )胚轴发育过程中的盐离子, 发现随着胚轴成熟, 其Mg2+ 、Ca2+ 、Na+ 、K+ 、Cl- 含量呈下降趋势, 胚轴的盐含量明显低于母树果皮、叶片的盐含量, 且果皮、叶片的组织液Cl- 浓度与海水Cl- 浓度接近, 故认为胎生过程是一个低盐化的过程, 后代是通过与母体的长期接触来获得抗盐锻炼, 进而适应落地后的环境(郑文教等, 1992 ; 郑文教和林鹏, 1997 )。红树植物很可能起源于淡水环境, 胚胎发育是其生活史的重演, 这种胚胎发育低盐化的过程很可能是一种“返祖”现象。有趣的是, Joshi等(1972)发现显胎生植物胚轴发育过程中Mg2+ 、Ca2+ 、Na+ 、K+ 含量也逐渐下降, 但Cl- 含量逐渐增加。然而, 秋茄树成熟胚轴的Mg2+ 、Ca2+ 、Na+ 、K+ 、Cl- 含量与早期胚轴相比呈倍性增加(Wang et al ., 2002 )。部分研究者认为胎生繁殖体的盐抗性是逐渐适应从母体输送到幼苗的盐分而获得的(Lin, 1988 )。这两种不同的结论可能是由于对胎生发育过程阶段的划分不同及盐离子含量、浓度的高低定义不同, 甚至是物种不同而导致的。相比于胎生红树植物, 非胎生红树植物繁殖体发育过程中盐离子动态的研究相对较少, 但有证据表明非胎生的老鼠簕(Acanthus ilicifolius )幼苗Cl- 含量高于胎生物种Ceriops candolleana 、Avicennia officinalis 、Aegiceras majus (Joshi et al ., 1972 ), 其机制仍有待阐明。Mg2+ 、Ca2 、Na+ 、K+ 和Cl- 这5种离子是细胞渗透势的主要成分, 除了构成盐分浓度外, 还可能作为一种信号在种子成熟过程中起其他作用, 如Ca2+ 作为二级信使传导信号调节基因表达。
长久以来胎生被认为是一个针对盐度适应的特征(Joshi, 1933 ), 但这一认识并没有被广泛接受, 因为其他的盐生植物并没有胎生现象。Wang等(2002)的研究发现秋茄树胚轴发育后期组织液离子浓度并没有明显变化, 推测胎生与抗盐性并没有直接的联系, 认为胎生并不是对某一个环境因子的适应, 而是对海岸潮间带多个生态因子综合响应的结果(闫中正等, 2004 )。
2.3 胚胎发育过程中激素的变化
植物激素在种子发育和休眠中具有广泛的作用。脱落酸(ABA)作为一个内源因子在调节种子成熟和发育过程中能够促进种子休眠, 抑制萌发。与非胎生物种相比, 胎生红树植物胚胎的ABA含量较低, 且在红树科中发现母体ABA含量高于胚胎, 因此低含量的ABA可能是红树植物胎生发生时种子没有脱水休眠的重要原因之一(Farnsworth & Farrant, 1998 ; Farnsworth, 2000 )。对秋茄树外源施加ABA发现果实个体及胚轴长度均小于对照组(洪礼伟等, 未发表资料), 表明其发育受到抑制, 进一步说明胚胎发育受到ABA影响, 推测胎生过程中ABA合成减少, 而不是胚胎发育对ABA不敏感。
脱水素蛋白(dehydrin protein)如late embryogenesis abundant (LEA)等与ABA之间存在密切的联系。Farrant等(1996)在非胎生物种成熟、未干燥的种子中检测到脱水素蛋白, 但在显胎生红树植物红树属、角果木属、木榄属和隐胎生红树植物海榄雌的成熟胚轴中则不能检测到脱水素, 然而在红茄苳(Rhizophora mucronata )早期胚胎发育时却有类脱水素蛋白的存在(Ismail et al ., 2010 )。脱水素在不同发育方式中的存在与否可能与生境有关。
另外, 与ABA有拮抗作用的赤霉素GA3在种子发育过程中始终维持较高的水平, 卢磊(2012) 推测GA3与ABA通过调控α淀粉酶影响种子中糖分的含量和比例, 从而影响胎生种子的萌发。与其他植物类似, 激素之间的相互作用在红树植物胎生发育过程中必不可少, 但仍存在许多问题, 如产生部位、运输方式、具体作用部位、作用机制、不同激素如何互作等。
2.4 胚胎发育过程中的能量动态
热值(caloric value)指单位质量干物质在完全燃烧后所释放出来的热量值, 可以反映出胎生繁殖体的能量水平(鲍雅静等, 2008 )。在繁殖体的发育过程中, 从花、果至成熟胚轴, 其热值逐渐降低, 且始终低于母体。这可能是由于生长发育过程中胚轴已经开始消耗能量, 以达到迅速突破果皮, 进而伸长的结果, 同时胚轴中低能量物质如纤维素等增加(林鹏等, 2000 )。此外, 灰分含量与热值变化趋势类似, 例如蜡烛果(Aegiceras corniculatum )胚轴在成熟过程中灰分含量降低, 但脱离母树后灰分含量逐步累积提高, 一些研究人员将灰分含量与盐离子联系起来, 认为蜡烛果胚轴脱离母树开始独立生长是一个大量吸收盐分的过程(林益明等, 2004 ; 赵胡等, 2004 )。虽然热值和灰分随着发育过程而降低, 但随着生物量的增加, 单个胚轴所含能量总体呈上升趋势, 为后续的传播和定植提供了足够的能耗支持。
无论是哪个因子单独起作用或是多个因子综合起作用, 这些因子在红树植物长期的进化过程中不断积累效应, 引起一系列分子、细胞、生理上的代谢变化, 这些变化使得红树植物的胎生过程在形态解剖、生理生化上有别于其他胚胎、种子发育, 使得胎生这一特殊繁殖方式适应海岸潮间带的复杂生境。
3 分子水平研究
3.1 红树植物的种群遗传多样性
遗传多样性在生物进化过程中起着重要作用, 因此研究红树植物的种群遗传结构对于理解红树植物的进化以及完善保育工作都具有重要的意义。Kado等(2004)通过分子标记技术, 对显胎生红树植物秋茄树、隐胎生红树植物海榄雌和非胎生红树植物榄李(Lumnitzera racemosa )在越南和日本的种群遗传多样性进行比较分析后发现, 繁殖体特性、地理分布以及自然干扰可能会影响红树植物种群的遗传结构, 但该研究未能解释不同的繁殖方式是如何影响种群遗传结构的。胎生物种分布在5个科8个属中, 这些科中的其他非红树植物不以胎生为繁殖方式, 说明红树植物的胎生在适应滨海特殊环境时以趋同进化的方式发展。Shi等(2005)分析了26个属的红树植物(包括胎生、非胎生植物类型)的rbc L、18S核糖体、mat R基因数据, 并构建分子系统进化树证实了胎生与泌盐这两个特征在红树植物中是独立起源的, 且胎生现象是多起源的, 但以显胎生为繁殖方式的红树科4个属在进化过程中有共同的祖先(Setoguchi et al ., 1999 ; Shi et al ., 2005 )。然而, 目前对显胎生、隐胎生和非胎生之间如何进化的关系却鲜有研究。
3.2 红树植物胎生发育过程转录组研究现状
红树植物在抗盐、抗重金属污染等方面的逆境生理生态特征及其分子生态学机制已有报道(张凤琴等, 2005 ; Miyama & Hanagata, 2007 ; Wong et al ., 2007 ; 程皓等, 2009 ; Parida & Jha, 2010 ), 但对红树植物独特的繁殖方式——胎生的分子机制研究甚少。转录组研究基本集中在红树属植物Rhizophora mangle 中, 目前仅此物种的花蕾、幼嫩及成熟的胚轴有转录组数据。Dassanayake等(2009)对R . mangle 不同阶段的营养器官、繁殖器官的转录组进行基因功能注释(gene ontology)分析, 在对所有表达的基因功能进行分类(分子功能、生物过程、细胞组分)比较后发现, “分子功能(molecular function)+生物过程(biological process)”共同表达的基因是“分子功能(molecular function)+细胞组分(cellular component)”共同表达的基因数量的两倍, 说明通过转录水平调控生理过程(而非改变细胞组分)有可能与红树植物胎生的生长发育过程存在密切关系。然而,由于采样阶段、基因组资源、信息分析、缺乏基因功能验证实验等的局限, 高达55%的序列没有功能注释, 而且未对花蕾、胚轴的发育进行深入分析, 此实验未能回答红树植物胎生发育的一系列问题。
3.3 其他植物中发现的胎生调控基因
如前所述, 某些作物中也存在着胎生现象, 表现为直接在植株上生根发芽, 称为成熟前萌发。这种在谷类作物中的胎生现象导致了大量减产, 因此对其分子研究较为深入。虽然这种作物提前萌发的现象在遗传进化上和红树植物的胎生有本质的区别, 但对研究红树植物胎生的分子机理有重要的借鉴意义。在玉米、水稻、拟南芥(Arabidopsis thaliana )等植物中发现的胎生调控基因基本都与ABA合成或响应通路有关(表2 ), 这与红树植物胎生过程中胚胎的ABA含量较低且能受外源ABA激素影响的现象相吻合。我们将这一系列与ABA合成及信号感受的基因(表2 )在玉米、水稻、拟南芥, 以及显胎生红树植物红树、隐胎生红树植物海榄雌和非胎生红树植物杯萼海桑(Sonneratia alba )的基因组数据中进行同源比对, 发现许多ABA通路中关键基因的蛋白序列在红树物种中具有高度保守性, 例如编码ABA生物合成第一步反应所需的玉米黄质环氧化酶的ABA1 基因(洪礼伟等, 未发表资料), 这也从另一个方面暗示着ABA很可能从分子水平上调控红树植物的胎生发育过程。值得一提的是, 在水稻中发现了对ABA信号不敏感并表现出胎生的突变体riv1-1 (rice vivipary 1-1 )、riv1-2 、riv2 及缺乏胚乳结构的enl1 (Kurata et al ., 2005 )。前3个突变体必须在外界湿润的条件下才能表现出提前萌发或提前萌发率大幅上升的现象, 而enl1 突变体由于缺乏在种子休眠中起重要作用的胚乳, 即使不施加水分也能提前萌发, 但在有水分的条件下会表现出更高的胎生率(Miyoshi et al ., 2000 ), 这说明胎生是内源基因与外界环境(很可能是湿度)共同作用的结果。因此, 在研究红树植物胎生现象时应当同时考虑基因与环境之间相互作用的可能性及重要性。
4 生态适应性
红树植物胎生现象的生态适应性长期以来被归结为两点: 有效定植及对海岸带环境的高度适应。
4.1 胎生繁殖体的有效定植
胎生意味着种子在母体上萌发, 直至生长为成熟的繁殖体, 是一个能量不断积累的过程(闫中正等, 2004 )。此时的繁殖体储存大量的碳水化合物, 并且因为表皮含叶绿素能进行光合作用, 能够为独立生长初期和漂流过程提供足够的营养供给(Lin & Sternberg, 1995 ; Smith & Snedaker, 2000 )。
胎生红树植物由于下胚轴在母体上已经发育生长到一定长度, 脱离母体后有一定概率直接插入滩涂与沙地, 其侧根能在落地后几小时内萌发, 又由于次生木质部等的作用, 胚轴能由水平到直立生长(Cheeseman, 2012 )。同时, 红树植物繁殖体主要是被动地依靠水体和泥沙流动进行传播, 在这种随波而流的前提下, 显胎生红树植物(例如木榄、秋茄树)胚轴所具有的厚角质层、发达的皮层通气组织以及表皮缺乏气孔等系列特征为长时间漂流提供支持, 并可避免海水的渗入(陈月琴等, 1995 )。而且繁殖体在漂流过程中, 能应对水体缺氧状况, 降低呼吸速率, 相应地减少能耗(Brown et al ., 1969 )。胎生繁殖体单宁含量比花期高, 在成熟过程中虽略有波动, 但总体还是呈积累趋势(Lin et al ., 2006 )。高含量的单宁使胎生繁殖体在漂流过程中不被海水腐蚀和动物啃食, 保持长时间的生命力, 有利于进行长距离扩散(林益明等, 2005 ), 这即是红树植物得以漂洋过海的秘密。
但漂流的远近与显胎生、非胎生繁殖方式并没有直接关联, 而是与果实大小、形状、密度紧密相关(Ryck et al ., 2012 )。胚轴密度与生境中海水密度是否接近导致不一样的漂流模式, 例如显胎生的木榄成熟胚轴密度接近海水密度, 适应随潮水传播, 而隐胎生的蜡烛果胚轴密度远大于生境海水, 仅适合短时间内漂流(张宜辉, 2003 )。个体较大的红茄苳繁殖体比起角果木来说传播得更远, 而根系萌发迅速的角果木在向陆带(landward)有定植优势(Robert et al ., 2015 )。这些不同的扩散模式可能最终决定了红树林的分带(Kadoya & Inoue, 2015 )。如上所述, 胎生种子在成熟过程中一直保持高水分含量。代谢水的丢失会对种子造成不可挽回的损伤, 但是在胎生繁殖体中这种损失并不存在, 能够最大限度地保持种子活力。胎生繁殖体(胚轴)作为具有相对完整结构的幼苗, 其定植明显比休眠的种子更加迅速而有效, 是海岸潮间带复杂环境的优势适应性特征。
4.2 胎生繁殖体对环境因子的响应
红树植物定植的海岸潮间带主要具有高盐、海水浸淹缺氧等特点, 胎生作为协同进化的结果, 对此类生境具有高度特异适应性。
胎生对适应高盐环境具有一定意义。在胚轴发育过程中细胞液的盐分浓度变化不明显(Wang et al ., 2002 ), 但是胚轴在母体上发育成熟, 最终达到与母树生境的离子浓度一致(张宜辉, 2003 )。繁殖体掉落在滩涂后迅速形成侧根, 并与外界进行离子交换, 以快速适应盐度变化。Joshi (1933)认为, 种子的胎生是为了避免受到高盐生境的迫害, 使其顺利发芽乃至定植在海岸潮间带。实际上, 在不同盐度培养条件下, 红树植物幼苗均表现出低盐促进、高盐抑制的特征。邱广龙(2005)发现低盐度有利于隐胎生白骨壤(即海榄雌)繁殖体的萌根与幼苗生长, 胎生和非胎生红树植物繁殖体同样偏好低盐环境(Jayatissa et al ., 2008 ; Robert et al ., 2015 )。然而, 有报道指出胎生繁殖体的萌发需要一定的盐度, 某些红树繁殖体在淡水中无法正常生长(Wang et al ., 2011 )。
光照是繁殖体定植的重要因素。前文提到胎生繁殖体表皮含有叶绿素, 能够进行光合作用为早期生长提供能量, 促进根茎生长。研究表明叶绿素缺失的繁殖体因缺乏色素而表现出白化特性, 无法进行光合作用, 只能存活数月, 直至自身含有的能量耗尽(Duke & Watkinson, 2002 )。繁殖体对不同光照强度有不同反应, 弱光条件下繁殖体通过扩大叶片面积和叶绿素含量增强光吸收, 强光条件下则相反, 最适的中度光照利于固碳作用(Ulqodry et al ., 2014 )。也有一些个例, 如分布范围狭窄的Pelliciera rhizophorae 在阴蔽处生长旺盛, 受到光照反而生长不良, 这一发现丰富了普遍认可的红树植物不耐阴的观念(Dangremond et al ., 2015 )。另外, 全黑条件下沙培的白骨壤茎生长的优势更加明显, 但生成的茎细长脆弱, 机械组织不发达, 对复杂潮间带的适应力减弱(邱广龙, 2005 )。不难解释, 林窗处的繁殖体幼苗生长更加迅速而强壮。
不仅成年红树植物需要面对潮汐的冲击, 而且胎生繁殖体受到更大干扰。胎生繁殖体可以忍受长期浸淹的缺氧条件生长发育成幼苗, 合适的潮汐(水淹时间长度及间隔)能在一定程度上促进繁殖体生长(Chen et al ., 2004 )。随海水漂流的繁殖体经长时间浸淹, 根伸长推迟, 茎叶生长表现得相当缓慢, 所以相对性缺水(例如进入潮落时的滩涂)促使繁殖体萌根及定植(Robert et al ., 2015 )。当然, 水体中的盐度和pH值处于一个不断变化的过程(Marchand et al ., 2004 ), 不同种类红树植物对水淹也具有不同的适应特征(例如多种类的根系、叶片结构), 使红树植物对潮汐的响应复杂化。
4.3 胎生适应潮间带环境的优势
与非胎生种子相比, 繁殖体特别是显胎生胚轴已经是相对完整的幼苗, 其所具有的一系列形态生理特征高度适应潮间带环境, 能够远距离传播和迅速定植。迅速定植是应对复杂多变潮间带环境的有力适应性手段, 红树植物胎生繁殖体在这一点上具有巨大优势。另外, Kadoya和Inoue (2015) 认为繁殖体在传播时, 其胎生特性具有更高的“可塑性”, 即胚轴能更快地适应水体中盐分的变化, 及时调整比重, 遇到合适的生境时下沉而定植。也就是说, 胎生繁殖体作为幼苗, 全部细胞处于活化状态, 因而对外界环境变化更加灵敏。
然而, 现有对胎生现象适应意义的了解并不能解释所有红树植物对其生长环境的适应性, 毕竟只有大约1/3的红树植物具有胎生现象。显然, 我们需要进行更全面系统的研究以阐明这一特殊繁殖方式对红树植物生态适应性机制的贡献。
5 研究不足与展望
对红树植物独特的胎生繁殖现象在形态结构、生理生态、分子层次、生态适应与进化等方面已经有了一定程度的了解, 但仍存在许多不足, 因为胎生现象的本质问题没有得到解答。第一, 红树植物的胎生现象是长期进化形成的繁殖方式, 那么它是如何受到那些未知遗传因子调控的?第二, 这种胎生现象对红树植物适应它们的生活环境起到多大的作用?第三, 哪些是促进红树植物胎生进化的关键因素?从形态结构方面来说, 过去的研究侧重简单地描述发育过程的结构, 而对具有重要适应性的结构缺乏更深层的探究。红树植物胎生发育并不是一个线性过程, 前人研究未将胎生发育过程的重要阶段或限速阶段进行定义, 这很可能掩盖了红树植物胎生的最关键的阶段问题, 阻碍我们对其进行正确的探究。对胎生发育关键转变阶段的界定将有利于有针对性地开展更高层次的研究, 把握发育过程的主要变化, 解决核心问题。在进行生理生态研究时, 仅笼统地区分花、果、成熟胚轴等阶段, 虽然能部分反映出发育过程的特性, 但容易因阶段划分过大, 忽略了主要转折点而导致不同甚至相反的结论。相比于其他层次的研究, 分子水平的进展最为缓慢, 目前尚无任何参与红树植物胎生发育过程基因的相关报道。虽然有对胎生物种如Rhizophora mangle 、角果木(Ceriops tagal )进行转录组测序, 但都集中在根、茎、叶等营养结构, 即使有选取花、胚轴作为转录组测序的组织, 但仍未就胎生这一发育过程进行数据挖掘。因此, 红树植物胎生发育的分子机制还有待阐明。
红树植物的胎生发育过程没有休眠期且不经脱水, 这与顽拗性种子相似。然而, 其发育场所为母体而非土壤这一特征则大大区别于一般的顽拗性种子, 因此, 在研究其机制时必须考虑母体(营养器官、种皮、果皮等)与后代(胚、胚轴)的能量流、物质流、信息流的传递与交换。从形态发育来看, 胎生现象是在不同阶段不同分生组织进行细胞分裂和增殖的结果, 这个过程包含着多种活性物质的协调作用, 如抑制发育、促进休眠的ABA在胚胎发育过程中呈低含量, 而作为各种生理生化反应所必需的水分始终维持在较高水平。促进与抑制萌发的物质在不同阶段的含量或信号传导敏感性的变化是导致胎生发育的重要原因。然而, 6种常见的植物激素对胎生的发育可能都起到一些生理作用, 因为许多生理效应是多种激素共同作用的结果。如在ABA浓度低的情况下, 其他激素如生长素、激动素等在胚胎发育、胎生萌发时都必须维持在高的水平。然而, 胎生(下胚轴)生长并不是无止境的, 而是到一定长度就停止, 这又要靠其他激素来调控。激素如何协调合作调控红树植物胎生发育这一过程目前尚未清楚。
红树胎生现象区别于其他植物繁殖方式的另一特征在于其复杂的生境。海岸潮间带具有高温、高湿、高盐、潮汐水淹、风浪冲击等特点, 在红树植物生活史的早期, 即胚胎萌发时就不可避免地受这些环境因子的影响。温度、水分、光照、盐度等因子作为环境信号在特定的时期引起植株内部一系列的变化, 诱导胚胎发育, 研究这些环境因子在这一过程中的具体作用对理解胎生发育的根本机制至关重要。显然, 环境因子作为外因引起胚胎内部的一系列形态结构、生理生化、基因层次的变化是红树植物胎生现象的本质。将红树植物胎生发育过程中的形态解剖与生理、分子、生态各个层次有机地结合起来是阐明胎生发育机理、机制的关键。红树植物的分子生态学、分子生物学研究仍留有许多空白, 尤其是在挖掘和利用红树植物独特的基因资源方面有很大研究空间。日益发展的基因组、转录组技术, 为大范围发掘植物基因和探索更广泛的生物学问题提供了强大的技术手段。对目标物种进行全基因组测序, 不仅可以获得该植物基因组和重要功能基因的序列信息, 而且能为从分子水平全面研究植物的分子进化、基因组成和基因调控等提供一定的基础和依据。同时, 基于新一代高通量测序技术的转录组测序目前已广泛应用到植物分子生物学研究中, 并且在研究植物胚胎发育、环境响应等方面已取得惊人的成果。通过获取不同关键阶段及组织的转录组数据并进行分析, 可得到基因表达差异、基因结构优化、可变剪接、单核苷酸多态性等信息, 这将有效地帮助我们找出胎生发育过程中与适应环境有关的基因, 从而探究胎生适应海岸潮间带特殊生境的机制。
从进化角度看, 现有的研究表明显胎生与隐胎生是分别进化而来的(Shi et al ., 2005 ), 而且分布在不同的科属中。更有趣的是, 这些红树植物的陆生近缘种(同科不同属)不是胎生物种。因此, 可以推断胎生的进化与红树植物的生境有关。更重要的是, 由于红树植物胎生是个自然的遗传程序, 需要一系列来自基因组的遗传信息, 即基因及其表达产物, 因此, 从基因组中找到这些遗传信息是将来胎生研究的重要方向。比较基因组的研究不仅可以找到胎生红树植物特有的基因簇, 而且也能得到在进化过程中产生的基因的特殊表达、调控方式及其转录组、蛋白组特征。在漫长的进化过程中, 积累在遗传物质中的信息决定了这一现象的发生, 不同基因在不同阶段、不同结构的表达, 各种转录因子的调控乃至这些基因架构成的网络是各种形态结构、生理生化变化的本质。这些组学水平的分析结果将对指导分子、生理乃至生态实验起到重要的启发作用。要理解红树植物胎生现象的遗传、生态、进化的本质, 需要多个领域许多科学家的共同努力。
我国红树林面积在历史上曾达25 × 104 hm2 , 到2013年总面积不足2.5 × 104 hm2 (廖宝文和张乔民, 2014 )。目前我国红树林恢复造林的主要方法是利用显胎生或隐胎生繁殖体进行滩涂扦插(彭逸生等, 2008 ), 具有操作便易、成本较低、幼苗存活率高等优点。在深入理解胎生发育机制后, 对造林技术进行优化, 能有效地帮助恢复红树林湿地面积。同时, 在全球气候变化的影响下, 极端温度频繁出现、海平面上升等可能威胁红树植物的生存状态。而作为独特的繁殖方式, 胎生如何产生变化适应新的环境因素也是未来研究的重要方向。
致谢 野外采样得到了海南清澜港省级红树林保护区及福建漳江口国家级红树林保护区工作人员的大力支持, 厦门大学环境与生态学院, 滨海湿地生态系统教育部重点实验室同仁苏文悦、陆文佳、曲海东、于志波协助完成实验样品采集, 吴鼎勋、田惠桥在制作石蜡切片过程中给予热心指导, 一并致谢。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Angelovici R Galili G Fernie AR Fait A 2010 ). Seed desiccation: A bridge between maturation and germination
.Trends in Plant Science 15 , 211 -218 .
[本文引用: 1]
[2]
Bao YJ Li ZH Han XG Song GB Yang XH Lü HY 2008 ). Plant caloric value and its bio-ecological attributes
.Chinese Journal of Ecology 25 , 1095 -1103 .(in Chinese with English abstract) [鲍雅静 , 李政海 , 韩兴国 , 宋国宝 , 杨晓慧 , 吕海燕 (2008 ). 植物热值及其生物生态学属性
. 生态学杂志, 25 , 1095 -1103 .]
[本文引用: 1]
[3]
Bertoni G 2010 ). Maize viviparous 14: Structure meets function
.The Plant Cell 22 , 2925 -2925 .
[4]
Brown J Outred H Hill C 1969 ). Respiratory metabolism in mangrove seedlings
.Plant Physiology 44 , 287 -294 .
[本文引用: 1]
[5]
Burrows CJ 1995 ). Germination behavior of the seeds of the New Zealand species
Aristotelia serrata, Coprosma robusta, Cordyline australis, Myrtus obcordata, and Schefflera digitata . New Zealand Journal of Botany 33 , 257 -264 .
[6]
Burrows CJ 1997 ). Reproductive ecology of New Zealand forests: 1. Natural seed storage phenomena
.New Zealand Natural Sciences 23 , 31 -52 .
[7]
Carey G Fraser L 1932 ). The embryology and seedling development ofAegiceras majus Gaertn
. Proceedings of the Linnean Society of New South Wales 57 , 341 -360 .
[本文引用: 1]
[8]
Cheeseman JM 2012 ). How red mangrove seedlings stand up
.Plant and Soil 355 , 395 -406 .
[本文引用: 1]
[9]
Chen L Wang W Lin P 2004 ). Influence of water logging time on the growth of
Kandelia candel seedlings. Acta Oceanologica Sinica 23 , 149 -157 .
[本文引用: 1]
[10]
Chen YQ Lan CY Huang YS Tan FY 1995 ). Anatomical structures and ecological adaptations of mangrove propagules
.Acta Scientiarum Naturalium Universitatis Sunyatseni 34 (4 ), 70 -75 .(in Chinese) [陈月琴 , 蓝崇钰 , 黄玉山 , 谭凤仪 (1995 ). 秋茄木榄繁殖体的结构及其生态特异性. 中山大学学报(自然科学版),
34(4) , 70 -75 .]
[本文引用: 1]
[11]
Cheng H Chen GZ Ye ZH 2009 ). Research progress of heavy metal pollution in mangrove wetlands
.Acta Ecologica Sinica 29 , 3893 -3900 .(in Chinese with English abstract) [程皓 , 陈桂珠 , 叶志鸿 (2009 ). 红树林重金属污染生态学研究进展. 生态学报,
29 , 3893 -3900 .]
[本文引用: 1]
[12]
Conde FL 1975 ). Vivipary in
Epiphyllum . Cactus and Succulent Journal 47 , 38 -39 .
[13]
Conti A Pancaldi S Fambrini M Michelotti V Bonora A Salvini M Pugliesi C 2004 ). A deficiency at the gene coding for ζ-carotene desaturase characterizes the sunflower
non dormant-1 mutant. Plant and Cell Physiology 45 , 445 -455 .
[14]
Cook MT 1907 ). The embryology of
Rhizophora mangle . Bulletin of the Torrey Botanical Club 34 , 271 -277 .
[本文引用: 1]
[15]
Corner EJH 1976). The Seeds of Dicotyledons . Cambridge University Press, New York .
[16]
Cota-Sánchez JH 2004 ). Vivipary in the Cactaceae: Its taxonomic occurrence and biological significance
.Flora- Morphology, Distribution, Functional Ecology of Plants 199 , 481 -490 .
[本文引用: 1]
[17]
Dahlgren R Clifford HT Yeo P 1985 ). The Families of the Monocotyledons: Structure, Evolution, and Taxonomy . Springer-Verlag
, New York.
[18]
Dangremond EM Feller IC Sousa WP 2015 ). Environmental tolerances of rare and common mangroves along light and salinity gradients
.Oecologia 179 , 1187 -1198 .
[本文引用: 1]
[19]
Dassanayake M Haas J Bohnert H Cheeseman J 2009 ). Shedding light on an extremophile life style through transcriptomics
.New Phytologist 183 , 764 -775 .
[20]
Deore AC Johnson TS 2008 ). Occurrence of vivipary in
Jatropha curcas L. Current Science 95 , 321 -322 .
[21]
Dintu K Sibi C Ravichandran P Satheeshkumar K 2015 ). Vivipary in
Ophiorrhiza mungos L.—A rare phenomenon in angiosperms. Plant Biology 17 , 294 -295 .
[22]
Dransfield S Bulletin K 1981 ). The genus
Dinochloa Gramineae-Bambusoideae) in Sabah. Kew Bulletin , 36 , 613 -633 .
[23]
Ducker S Knox R 1976 ). Submarine pollination in seagrasses
.Nature 263 , 705 -706 .
[24]
Duke NC Watkinson AJ 2002 ). Chlorophyll-deficient propagules of
Avicennia marina and apparent longer term deterioration of mangrove fitness in oil-polluted sediments. Marine Pollution Bulletin 44 , 1269 -1276 .
[本文引用: 1]
[25]
Ellis RH Hong T Roberts EH 1985 ). Handbook of Seed Technology for Genebanks . Volume II . Compendium of Specific Germination Information and Test Recommendations . International Board for Plant Genetic Resources, Rome
, Italy.
[26]
Elmqvist T Cox PA 1996 ). The evolution of vivipary in flowering plants
.Oikos 77 , 3 -9 .
[本文引用: 1]
[27]
Fang J Chai C Qian Q Li C Tang J Sun L Huang Z Guo X Sun C Liu M 2008 ). Mutations of genes in synthesis of the carotenoid precursors of ABA lead to pre-harvest sprouting and photo-oxidation in rice
.The Plant Journal 54 , 177 -189 .
[28]
Farnsworth E 2000 ). The ecology and physiology of viviparous and recalcitrant seeds
.Annual Review of Ecology and Systematics 31 , 107 -138 .
[本文引用: 1]
[29]
Farnsworth E Farrant J 1998 ). Reductions in abscisic acid are linked with viviparous reproduction in mangroves
.American Journal of Botany 85 , 760 .
[本文引用: 1]
[30]
Farnsworth EJ 1997 ). Evolutionary and Ecological Physiology of Mangrove Seedlings: Correlates, Costs, and Consequences of Viviparous Reproduction . Harvard University, Cambridge
, UK.
[本文引用: 1]
[31]
Farrant JM Berjak P Cutting J Pammenter N 1993 ). The role of plant growth regulators in the development and germination of the desiccation-sensitive (recalcitrant) seeds of
Avicennia marina . Seed Science Research 3 , 55 -63 .
[32]
Farrant JM Pammenter NW Berjak P 1992 ). Development of the recalcitrant (homoiohydrous) seeds of
Avicennia marina: Anatomical, ultrastructural and biochemical events associated with development from histodifferentiation to maturation. Annals of Botany 70 , 75 -86 .
[本文引用: 1]
[33]
Farrant JM Pammenter NW Berjak P Farnsworth EJ Vertucci CW 1996 ). Presence of dehydrin-like proteins and levels of abscisic acid in recalcitrant (desiccation sensitive) seeds may be related to habitat
.Seed Science Research 6 , 175 -182 .
[34]
Fountain DW Outred HA 1991 ). Germination requirements of New Zealand native plants: A review
.New Zealand Journal of Botany 29 , 311 -316 .
[35]
Guppy HB 1912 ). Studies in Seeds and Fruits: An Investigation with the Balance . Williams and Norgate
, London.
[36]
Hable W Oishi K Schumaker K 1998 ). Viviparous-5 encodes phytoene desaturase, an enzyme essential for abscisic acid (ABA) accumulation and seed development in maize
.Molecular and General Genetics 257 , 167 -176 .
[37]
Hara T Katoh H Ogawa D Kagaya Y Sato Y Kitano H Nagato Y Ishikawa R Ono A Kinoshita T 2015 ). Rice SNF2 family helicase ENL1 is essential for syncytial endosperm development
.The Plant Journal 81 , 1 -12 .
[本文引用: 1]
[38]
Ismail FA Nitsch LM Wolters-Arts MM Mariani C Derksen JW 2010 ). Semi-viviparous embryo development and dehydrin expression in the mangrove
Rhizophora mucronata Lam. Sexual Plant Reproduction 23 , 95 -103 .
[本文引用: 1]
[39]
Janzen DH 1983). Costa Rican Natural History . University of Chicago Press, Chicago .
[40]
Jayatissa L Wickramasinghe W Dahdouh-Guebas F Huxham M 2008 ). Research paper interspecific variations in responses of mangrove seedlings to two contrasting salinities
.Internationale Revue Der Gesamten Hydrobiologie Und Hydrographie 93 , 700 -710 .
[本文引用: 1]
[41]
Joshi A 1933 ). A suggested explanation of the prevalence of vivipary on the sea-shore
.Journal of Ecology 21 , 209 -212 .
[本文引用: 1]
[42]
Joshi BD Paroda RS 1991 ). Buckwheat in India . National Bureau of Plant Genetic Resources Regional Station, Shimla
, India.
[43]
Joshi G Pimplaskar M Bhosale LJ 1972 ). Physiological studies in germination of mangroves
.Botanica Marina 15 , 91 -95 .
[本文引用: 1]
[44]
Juncosa AM 1982 ). Developmental morphology of the embryo and seedling of
Rhizophora mangle L.Rhizophoraceae). American Journal of Botany , 69 , 1599 -1611 .
[本文引用: 1]
[45]
Juncosa AM Tomlinson PB 1988 ). Systematic comparison and some biological characteristics of Rhizophoraceae and Anisophylleaceae
.Annals of the Missouri Botanical Garden 75 , 1296 -1318 .
[本文引用: 2]
[46]
Kado T Fujimoto A Giang LH Tuan M Hong PN Harada K Tachida H 2004 ). Genetic structures of natural populations of three mangrove species,
Avicennia marina, Kandelia candel and Lumnitzera racemosa, in Vietnam revealed by maturase sequences of plastid DNA. Plant Species Biology 19 , 91 -99 .
[47]
Kadoya T Inoue T 2015 ). Spatio-temporal pattern of specific gravity of mangrove diaspore: Implications for upstream dispersal
.Ecography 38 , 472 -479 .
[本文引用: 1]
[48]
Kuo J Kirkman H 1990 ). Anatomy of viviparous seagrasses seedlings of Amphibolis and Thalassodendron and their nutrient supply
. Botanica Marina, 33, 117-126.
[本文引用: 1]
[49]
Kurata N Miyoshi K Nonomura KI Yamazaki Y Ito Y 2005 ). Rice mutants and genes related to organ development, morphogenesis and physiological traits
.Plant and Cell Physiology 46 , 48 -62 .
[本文引用: 1]
[50]
Leck MA Parker VT Simpson RL 2008). Seedling Ecology and Evolution . Cambridge University Press, New York .
[51]
Leite A Rankin J 1981 ). Ecology of seeds of
Pithecolobium racemosum Ducke. Acta Amazonica 11 , 309 -318 .]
[52]
Liao BW Zhang QM 2014 ). Area, distribution and species composition of mangroves in China
.Wetland Science 12 , 435 -440 .(in Chinese with English abstract) [廖宝文 , 张乔民 (2014 ). 中国红树林的分布、面积和树种组成. 湿地科学,
12 , 435 -440 .]
[本文引用: 1]
[53]
Lin G Sternberg LDSL 1995 ). Variation in propagule mass and its effect on carbon assimilation and seedling growth of red mangrove (
Rhizophora mangle ) in Florida, USA. Journal of Tropical Ecology 11 , 109 -119 .
[本文引用: 1]
[54]
Lin P 1988). Mangrove Vegetation . China Ocean Press, Beijing .
[55]
Lin P Wu SJ Lin YM 2000 ). Changes in energy with the development of the propagules of some mangrove species
.Marine Science 24 (9 ), 46 -50 .(in Chinese with English abstract) [林鹏 , 吴世军 , 林益明 (2000 ). 红树植物繁殖体发育过程的能量变化. 海洋科学,
24(9) , 46 -50 .]
[本文引用: 1]
[56]
Lin YM Liu J Xiang P Lin P Ye G Da Sternberg L 2006 ). Tannin dynamics of propagules and leaves of
Kandelia candel and Bruguiera gymnorrhiza in the Jiulong River Estuary, Fujian, China. Biogeochemistry 78 , 343 -359 .
[本文引用: 1]
[57]
Lin YM Xiang P Lin P 2004 ). Caloric values of propagules and leaves at the different development stages of mangrove species at Futian, Shenzhen
.Marine Science 28 (2 ), 43 -47 .(in Chinese with English abstract) [林益明 , 向平 , 林鹏 (2004 ). 深圳福田几种红树植物繁殖体与不同发育阶段叶片热值研究. 海洋科学,
28(2) , 43 -47 .]
[本文引用: 1]
[58]
Lin YM Xiang P Lin P 2005 ). Studies on tannins of mangroves—A review
.Marine Science 29 (3 ), 59 -64 .(in Chinese) [林益明 , 向平 , 林鹏 (2005 ). 红树林单宁的研究进展. 海洋科学,
29(3) , 59 -63 .]
[本文引用: 1]
[59]
Lombardi JA 1993 ). Viviparity in
Rhipsalis pilocarpa . Science and Culture 45 , 407 .
[60]
Lu L 2012 ). Function of Change of GA3, ABA and Sugar in the Viviparity of Mangrove Plant Species . Master degree dissertation. Xiamen University
, Xiamen. (in Chinese with English abstract) [卢磊 (2012 ). 红树植物胎生过程中ABA、GA3和糖分含量的变化
. 硕士学位论文, 厦门大学, 厦门.]
[本文引用: 1]
[61]
Marchand C Baltzer F Lallier-Vergès E Albéric P 2004 ). Pore-water chemistry in mangrove sediments: Relationship with species composition and developmental stages (French Guiana)
.Marine Geology 208 , 361 -381 .
[本文引用: 1]
[62]
Marilyn CB Graham DF 1984 ). Photosynthetic and stomatal responses of two mangrove species,
Aegiceras corniculatum and Avicennia marina, to long term salinity and humidity conditions. Plant Physiology 74 , 1 -6 .
[63]
Marrero J 1942 ). A seed storage study of maga
.Caribbean Forester 3 (4 ), 173 -183 .
[64]
Matthews PD Luo R Wurtzel ET 2003 ). Maize phytoene desaturase and ζ-carotene desaturase catalyse a poly-Z desaturation pathway: Implications for genetic engineering of carotenoid content among cereal crops
.Journal of Experimental Botany 54 , 2215 -2230 .
[65]
McCarty DR Carson CB Stinard PS Robertson DS 1989 ). Molecular analysis of
viviparous-1: An abscisic acid insensitive mutant of maize. The Plant Cell 1 , 523 -532 .
[66]
McCarty DR Hattori T Carson CB Vasil V Lazar M Vasil IK 1991 ). The viviparous-1 developmental gene of maize encodes a novel transcriptional activator
.Cell 66 , 895 -905 .
[67]
Meinke DW Franzmann LH Nickle TC Yeung EC 1994 ). Leafy cotyledon mutants of
Arabidopsis . The Plant Cell 6 , 1049 -1064 .
[68]
Miyama M Hanagata N 2007 ). Microarray analysis of 7029 gene expression patterns in Burma mangrove under high-salinity stress
.Plant Science 172 , 948 -957 .
[本文引用: 1]
[69]
Miyoshi K Nakata E Nagato Y 2000 ). Characterization of viviparous mutants in rice
.Breeding Science 50 , 207 -213 .
[本文引用: 1]
[70]
Parida AK Jha B 2010 ). Salt tolerance mechanisms in mangroves: A review
.Trees 24 , 199 -217 .
[本文引用: 1]
[71]
Peng YS Zhou YW Chen GZ 2008 ). The restoration of mangrove wetland: A review
.Acta Ecologica Sinica 28 , 786 -797 .(in Chinese with English abstract) [彭逸生 , 周炎武 , 陈桂珠 (2008 ). 红树林湿地恢复研究进展. 生态学报,
28 , 786 -797 .]
[本文引用: 1]
[72]
Pijl L 1972 ). Principles of Dispersal in Higher Plants . Springer-Verlag
, New York.
[73]
Porch TG Tseung CW Schmelz EA Mark Settles A 2006 ). The maize viviparous10/viviparous13 locus encodes the
Cnx1 gene required for molybdenum cofactor biosynthesis. The Plant Journal 45 , 250 -263 .
[74]
Qiu GL 2005 ). Research on Reproductive Ecology and Analysis on Fruit Quality of Mangrove Avicenna marina . Master degree dissertation. Guangxi University
, Guangxi. (in Chinese with English Abstract) [邱广龙 (2005 ). 红树植物白骨壤繁殖生态研究与果实品质分析
. 硕士学位论文, 广西大学, 广西.]
[本文引用: 1]
[75]
Ramanayake SMSD Weerawardene TE 2003 ). Flowering in a bamboo,
Melocanna baccifera Bambusoideae: Poaceae). Botanical Journal of the Linnean Society ,143 , 287 -291 .
[76]
Robert EM Oste J van der Stocken T de Ryck DJ Quisthoudt K Kairo JG Dahdouh-Guebas F Koedam N Schmitz N 2015 ). Viviparous mangrove propagules of
Ceriops tagal and Rhizophora mucronata, where both Rhizophoraceae show different dispersal and establishment strategies. Journal of Experimental Marine Biology and Ecology 468 , 45 -54 .
[本文引用: 3]
[77]
Robertson DS 1955 ). The genetics of vivipary in maize
. Genetics 40 , 745 -760 .
[本文引用: 1]
[78]
Ryck DJRD Robert EMR Nele S Stocken TVD Nitto DD Guebas FD Koedam N 2012 ). Size does matter, but not only size: Two alternative dispersal strategies for viviparous mangrove propagules
.Aquatic Botany 103 , 66 -73 .
[本文引用: 1]
[79]
Sankaran M Damodaran D Singh DR Jerard BA 2012 ). Vivipary in
Cocos nucifera L. var. Andaman Green Dwarf. Current Science 103 , 1139 -1140 .
[80]
Setoguchi H Kosuge K Tobe H 1999 ). Molecular phylogeny of Rhizophoraceae based on
rbc L gene sequences. Journal of Plant Research 112 , 443 -455 .
[本文引用: 1]
[81]
Sheue CR Liu HY Yong JW 2003 ). Kandelia obovata (Rhizophoraceae), a new mangrove species from Eastern Asia
.Taxon 52 , 287 -294 .
[82]
Shi SH Huang YL Zeng K Tan FX He HH Huang JZ Fu YX 2005 ). Molecular phylogenetic analysis of mangroves: Independent evolutionary origins of vivipary and salt secretion
.Molecular Phylogenetics and Evolution 34 , 159 -166 .
[本文引用: 2]
[83]
Simpson MJA 1977 ). Fruit characters and seed germination of
Euphrasia disperma Hook f. New Zealand Journal of Botany 15 , 181 -183 .
[84]
Simpson MJA 1979 ). Lack of dormancy in seeds of New Zealand plants
.Canterbury Botanical Society 13 , 36 -37 .
[85]
Singh D Mathur SB 2004). Histopathology of Seed-Borne Infections . CRC Press, Florida .
[86]
Singh M Lewis PE Hardeman K Bai L Rose JKC Mazourek M Chomet P Brutnell TP 2003 ). Activator mutagenesis of the
pink scutellum1 /viviparous7 locus of maize. The Plant Cell 15 , 874 -884 .
[87]
Smith SM Snedaker SC 2000 ). Hypocotyl function in seedling development of the red mangrove,
Rhizophora mangle L. Biotropica 32 , 677 -685 .
[本文引用: 1]
[88]
Sussex I 1975 ). Growth and metabolism of the embryo and attached seedling of the viviparous mangrove,
Rhizophora mangle . American Journal of Botany 62 , 948 -953 .
[本文引用: 1]
[89]
Suzuki M Mark Settles A Tseung CW Li QB Latshaw S Wu S Porch TG Schmelz EA James MG McCarty DR 2006 ). The maize viviparous 15 locus encodes the molybdopterin synthase small subunit
.The Plant Journal 45 , 264 -274 .
[90]
Tan H Rao A 1981 ). Vivipary in
Ophiorrhiza tomentosa Jack (Rubiaceae). Biotropica 13 , 232 -233 .
[91]
Teichman I Wyk AE 1991 ). Trends in the evolution of dicotyledonous seeds based on character associations, with special reference to pachychalazy and recalcitrance
.Botanical Journal of the Linnean Society 105 , 211 -237 .
[92]
Tobe H Raven PH 1988 ). Seed morphology and anatomy of Rhizophoraceae, inter- and infrafamilial relationships
.Annals of the Missouri Botanical Garden 75 ,1319 -1342 .
[本文引用: 1]
[93]
Tomlinson P 1986). The Botany of Mangroves . Cambridge University Press, New York .
[94]
Ulqodry TZ Matsumoto F Okimoto Y Nose A Zheng SH 2014 ). Study on photosynthetic responses and chlorophyll fluorescence in
Rhizophora mucronata seedlings under shade regimes. Acta Physiologiae Plantarum 36 , 1903 -1917 .
[本文引用: 1]
[95]
Venkateswarlu J 1937 ). A contribution to the embryology of Sonneratiaceae
.Proceedings: Plant Sciences 5 , 206 -223 .
[本文引用: 1]
[96]
Vertucci CW 1989 ). The effects of low water contents on physiological activities of seeds
.Physiologia Plantarum 77 , 172 -176 .
[本文引用: 1]
[97]
Wagner WL Weller SG Sakai AK Medeiros AC 1999 ). An autogamous rainforest species of
Schiedea Caryophyllaceae) from East Maui, Hawaiian Islands, Novon , 9 , 284 -287 .
[98]
Wang W Ke L Tam N Wong Y 2002 ). Changes in the main osmotica during the development of
Kandelia candel hypocotyls and after mature hypocotyls were transplanted in solutions with different salinities. Marine Biology 141 , 1029 -1034 .
[本文引用: 2]
[99]
Wang W Yan Z You S Zhang Y Chen L Lin G 2011 ). Mangroves: Obligate or facultative halophytes? A review
.Trees 25 , 953 -963 .
[本文引用: 1]
[100]
Wang WQ Wang M 2007). The Mangroves of China . Science Press, Beijing .(in Chinese) [王文卿 , 王瑁 (2007). 中国红树林. 科学出版社 , 北京.]
[101]
Wong YY Ho CL Nguyen PD Teo SS Harikrishna JA Rahim RA Wong MC 2007 ). Isolation of salinity tolerant genes from the mangrove plant,
Bruguiera cylindrica by using suppression subtractive hybridization (SSH) and bacterial functional screening. Aquatic Botany 86 , 117 -122 .
[本文引用: 1]
[102]
Yan ZZ Wang WQ Huang WB 2004 ). Development of the viviparous hypocotyl of mangrove and its adaptation to inter-tidal habitats: A review
.Acta Ecologica Sinica 24 , 2317 -2323 .(in Chinese with English abstract) [闫中正 , 王文卿 , 黄伟滨 (2004 ). 红树胎生现象及其对潮间带生境适应性研究进展. 生态学报,
24 , 2317 -2323 .]
[本文引用: 2]
[103]
Ye NG 1979 ). How does vivipary happen in viviparous plant?
Plants 6 ), 20 -21 .(in Chinese) [叶能干 (1979 ). 胎生植物是怎样 “胎生” 的? 植物杂志,
(6 ), 20 -21 .]
[本文引用: 1]
[104]
Zhang FQ Wang YS Yin JP Dong JD 2005 ). Research advances on the resistance of mangrove plants to heavy metal pollution
.Acta Botanica Yunnanica 27 , 225 -231 .(in Chinese with English abstract) [张凤琴 , 王友绍 , 殷建平 , 董俊德 (2005 ). 红树植物抗重金属污染研究进展. 云南植物研究,
27 , 225 -231 .]
[本文引用: 1]
[105]
Zhang YH 2003 ). The Study of Propagule Development and Seedling Growth in Some Mangrove Species . PhD dissertation. Xiamen University
, Xiamen. (in Chinese) [张宜辉 (2003 ). 几种红树植物繁殖体发育和幼苗成长过程的生理生态学研究
. 博士学位论文, 厦门大学, 厦门.]
[本文引用: 2]
[106]
Zhao H Zheng WJ Sun J Lin P 2004 ). Dynamics of element levels and adaptation to saline environment during the development in
Aegiceras corniculatum mangrove. Marine Science 28 (9 ), 1 -5 .(in Chinese with English abstract) [赵胡 , 郑文教 , 孙娟 , 林鹏 (2004 ). 红树植物桐花树生长发育过程的元素动态与抗盐适应性. 海洋科学,
28 (9) , 1 -5 .]
[本文引用: 1]
[107]
Zheng WJ Lin P 1997 ). The element dynamics of propagule development and leaf growth in some mangrove plants
.Acta Oceanologica Sinica 19 (1 ), 96 -103 .(in Chinese) [郑文教 , 林鹏 (1997 ). 红树胚轴和叶片生长发育的元素动态. 海洋学报,
19 , 96 -103 .]
[本文引用: 1]
[108]
Zheng WJ Lin P Zheng FZ 1992 ). Dynamic of Cl content in development of viviparous hypocotyl and leaf of mangrove
.Journal of Xiamen University (Natural Science )31 , 537 -542 .(in Chinese with English abstract) [ 郑文教 , 林鹏 , 郑逢中 (1992 ). 红树胎生胚轴发育和叶片发育的 C1量动态. 厦门大学学报(自然科学版),
31 , 537 -542 .]
[本文引用: 1]
Seed desiccation: A bridge between maturation and germination
1
2010
... 大部分陆生植物种子发育成熟后往往要经历一个脱水、休眠的阶段(Angelovici et al ., 2010 ), 脱离母体后, 待外界条件如温度、湿度适宜时种子才逐渐萌发, 利用贮存的碳水化合物、脂肪和蛋白质等大分子化合物水解产生的小分子物质和能量供应幼苗生长发育.胎生植物则不然, 它们的种子没有休眠期或休眠期很短, 直接在母体植株的果实中萌发, 形成胚芽、下胚轴、胚根结构, 脱离母体定植后继续生长.这种繁殖模式显然与它们生活的热带、亚热带温暖、湿润的气候环境有关. ...
植物热值及其生物生态学属性
1
2008
... 热值(caloric value)指单位质量干物质在完全燃烧后所释放出来的热量值, 可以反映出胎生繁殖体的能量水平(鲍雅静等, 2008 ).在繁殖体的发育过程中, 从花、果至成熟胚轴, 其热值逐渐降低, 且始终低于母体.这可能是由于生长发育过程中胚轴已经开始消耗能量, 以达到迅速突破果皮, 进而伸长的结果, 同时胚轴中低能量物质如纤维素等增加(林鹏等, 2000 ).此外, 灰分含量与热值变化趋势类似, 例如蜡烛果(Aegiceras corniculatum )胚轴在成熟过程中灰分含量降低, 但脱离母树后灰分含量逐步累积提高, 一些研究人员将灰分含量与盐离子联系起来, 认为蜡烛果胚轴脱离母树开始独立生长是一个大量吸收盐分的过程(林益明等, 2004 ; 赵胡等, 2004 ).虽然热值和灰分随着发育过程而降低, 但随着生物量的增加, 单个胚轴所含能量总体呈上升趋势, 为后续的传播和定植提供了足够的能耗支持. ...
Maize viviparous 14: Structure meets function
2010
Respiratory metabolism in mangrove seedlings
1
1969
... 胎生红树植物由于下胚轴在母体上已经发育生长到一定长度, 脱离母体后有一定概率直接插入滩涂与沙地, 其侧根能在落地后几小时内萌发, 又由于次生木质部等的作用, 胚轴能由水平到直立生长(Cheeseman, 2012 ).同时, 红树植物繁殖体主要是被动地依靠水体和泥沙流动进行传播, 在这种随波而流的前提下, 显胎生红树植物(例如木榄、秋茄树)胚轴所具有的厚角质层、发达的皮层通气组织以及表皮缺乏气孔等系列特征为长时间漂流提供支持, 并可避免海水的渗入(陈月琴等, 1995 ).而且繁殖体在漂流过程中, 能应对水体缺氧状况, 降低呼吸速率, 相应地减少能耗(Brown et al ., 1969 ).胎生繁殖体单宁含量比花期高, 在成熟过程中虽略有波动, 但总体还是呈积累趋势(Lin et al ., 2006 ).高含量的单宁使胎生繁殖体在漂流过程中不被海水腐蚀和动物啃食, 保持长时间的生命力, 有利于进行长距离扩散(林益明等, 2005 ), 这即是红树植物得以漂洋过海的秘密. ...
Germination behavior of the seeds of the New Zealand species
1995
Reproductive ecology of New Zealand forests: 1. Natural seed storage phenomena
1997
The embryology and seedling development ofAegiceras majus Gaertn
1
1932
... 隐胎生红树植物的胚胎发育方式与显胎生有所差别.海榄雌(Avicennia marina )胚胎发育至球形胚时, 胚孔吸根(micropylar haustorium)萎缩, 胚乳连同胚从珠被(种皮)中伸出, 进入到子房腔生长, 胚进而充满子房, 成熟时果实掉落(Farrant et al ., 1992 ).在蜡烛果属植物Aegiceras majus 中的情况较为不同, 胚珠受精后具有分生性的胚乳先行分裂, 待大量胚乳形成后, 合子才在胚孔端开始发育(Carey & Fraser, 1932 ).胚乳在胚孔处的分裂产生皱褶, 这些皱褶增加了吸收养分的面积, 相当于吸器的功能.经过一段时间的生长后, 胚占主导地位, 为了适应胚的这一变化, 胎座也伸长以维持胚的相对位置不变, 并保证了养料的持续供给, 而后胚根及胚轴的下部伸出种皮.随着胚逐渐增大, 种皮形成了类似帽子的结构, 罩在子叶上端.果实连同发育了的胚一起掉落, 在适当的条件下胚轴继续伸长, 直至胚根突破果皮, 侧根生长在松软的泥土上完成定植, 子叶也从果实中露出, 与包裹在其中的幼叶一起进行光合作用, 幼苗开始生长. ...
How red mangrove seedlings stand up
1
2012
... 胎生红树植物由于下胚轴在母体上已经发育生长到一定长度, 脱离母体后有一定概率直接插入滩涂与沙地, 其侧根能在落地后几小时内萌发, 又由于次生木质部等的作用, 胚轴能由水平到直立生长(Cheeseman, 2012 ).同时, 红树植物繁殖体主要是被动地依靠水体和泥沙流动进行传播, 在这种随波而流的前提下, 显胎生红树植物(例如木榄、秋茄树)胚轴所具有的厚角质层、发达的皮层通气组织以及表皮缺乏气孔等系列特征为长时间漂流提供支持, 并可避免海水的渗入(陈月琴等, 1995 ).而且繁殖体在漂流过程中, 能应对水体缺氧状况, 降低呼吸速率, 相应地减少能耗(Brown et al ., 1969 ).胎生繁殖体单宁含量比花期高, 在成熟过程中虽略有波动, 但总体还是呈积累趋势(Lin et al ., 2006 ).高含量的单宁使胎生繁殖体在漂流过程中不被海水腐蚀和动物啃食, 保持长时间的生命力, 有利于进行长距离扩散(林益明等, 2005 ), 这即是红树植物得以漂洋过海的秘密. ...
Influence of water logging time on the growth of
1
2004
... 不仅成年红树植物需要面对潮汐的冲击, 而且胎生繁殖体受到更大干扰.胎生繁殖体可以忍受长期浸淹的缺氧条件生长发育成幼苗, 合适的潮汐(水淹时间长度及间隔)能在一定程度上促进繁殖体生长(Chen et al ., 2004 ).随海水漂流的繁殖体经长时间浸淹, 根伸长推迟, 茎叶生长表现得相当缓慢, 所以相对性缺水(例如进入潮落时的滩涂)促使繁殖体萌根及定植(Robert et al ., 2015 ).当然, 水体中的盐度和pH值处于一个不断变化的过程(Marchand et al ., 2004 ), 不同种类红树植物对水淹也具有不同的适应特征(例如多种类的根系、叶片结构), 使红树植物对潮汐的响应复杂化. ...
秋茄木榄繁殖体的结构及其生态特异性. 中山大学学报(自然科学版),
1
1995
... 胎生红树植物由于下胚轴在母体上已经发育生长到一定长度, 脱离母体后有一定概率直接插入滩涂与沙地, 其侧根能在落地后几小时内萌发, 又由于次生木质部等的作用, 胚轴能由水平到直立生长(Cheeseman, 2012 ).同时, 红树植物繁殖体主要是被动地依靠水体和泥沙流动进行传播, 在这种随波而流的前提下, 显胎生红树植物(例如木榄、秋茄树)胚轴所具有的厚角质层、发达的皮层通气组织以及表皮缺乏气孔等系列特征为长时间漂流提供支持, 并可避免海水的渗入(陈月琴等, 1995 ).而且繁殖体在漂流过程中, 能应对水体缺氧状况, 降低呼吸速率, 相应地减少能耗(Brown et al ., 1969 ).胎生繁殖体单宁含量比花期高, 在成熟过程中虽略有波动, 但总体还是呈积累趋势(Lin et al ., 2006 ).高含量的单宁使胎生繁殖体在漂流过程中不被海水腐蚀和动物啃食, 保持长时间的生命力, 有利于进行长距离扩散(林益明等, 2005 ), 这即是红树植物得以漂洋过海的秘密. ...
红树林重金属污染生态学研究进展. 生态学报,
1
2009
... 红树植物在抗盐、抗重金属污染等方面的逆境生理生态特征及其分子生态学机制已有报道(张凤琴等, 2005 ; Miyama & Hanagata, 2007 ; Wong et al ., 2007 ; 程皓等, 2009 ; Parida & Jha, 2010 ), 但对红树植物独特的繁殖方式——胎生的分子机制研究甚少.转录组研究基本集中在红树属植物Rhizophora mangle 中, 目前仅此物种的花蕾、幼嫩及成熟的胚轴有转录组数据.Dassanayake等(2009)对R . mangle 不同阶段的营养器官、繁殖器官的转录组进行基因功能注释(gene ontology)分析, 在对所有表达的基因功能进行分类(分子功能、生物过程、细胞组分)比较后发现, “分子功能(molecular function)+生物过程(biological process)”共同表达的基因是“分子功能(molecular function)+细胞组分(cellular component)”共同表达的基因数量的两倍, 说明通过转录水平调控生理过程(而非改变细胞组分)有可能与红树植物胎生的生长发育过程存在密切关系.然而,由于采样阶段、基因组资源、信息分析、缺乏基因功能验证实验等的局限, 高达55%的序列没有功能注释, 而且未对花蕾、胚轴的发育进行深入分析, 此实验未能回答红树植物胎生发育的一系列问题. ...
A deficiency at the gene coding for ζ-carotene desaturase characterizes the sunflower
2004
The embryology of
1
1907
... 红树植物胎生过程包含胚胎形成和胚胎萌发两个过程.以显胎生为繁殖策略的红树胚胎发育又可分为3个阶段: 子叶的第一次生长; 下胚轴的生长; 子叶的第二次生长(Cook, 1907 ).红树属、秋茄树属、角果木属和木榄属4个属间的发育各有特点, 在进化地位上有所差异. ...
Vivipary in the Cactaceae: Its taxonomic occurrence and biological significance
1
2004
... 真胎生现象在4类植物中已有报道(Farnsworth, 1997 ).首先是在遗传上以胎生为繁殖方式的植物, 如分布在8个属的红树植物(表1 )及两个属的海草(Tomlinson, 1986 ), 它们在自然生长环境条件下以胎生为最主要的繁殖方式, 繁殖体在母体上突破种皮或果皮并生长; 二是生长于热带、亚热带的某些顽拗性种子植物, 如少数仙人掌科植物(Cota-Sánchez, 2004 ); 三是某些处于特殊环境中的植物, 如水稻(Oryza sativa )、玉米(Zea mays )等, 其种子在高温、高湿条件下提前萌发, 但它们不从母体中吸收营养(叶能干, 1979 ); 四是与休眠有关的植物激素脱落酸(ABA)合成或响应通路有关的突变体植株, 或因种子中休眠基因功能的缺失不能进入休眠期而在母体直接开始萌发(Robertson, 1955 ), 后两类植物由于受外界条件诱导而形成提前萌发的性状, 在遗传本质上不是真正意义上的胎生, 但对于红树植物胎生现象研究仍有一定的借鉴作用.表1 列出已发表的以胎生为繁殖方式的植物. ...
The Families of the Monocotyledons: Structure, Evolution, and Taxonomy . Springer-Verlag
1985
Environmental tolerances of rare and common mangroves along light and salinity gradients
1
2015
... 光照是繁殖体定植的重要因素.前文提到胎生繁殖体表皮含有叶绿素, 能够进行光合作用为早期生长提供能量, 促进根茎生长.研究表明叶绿素缺失的繁殖体因缺乏色素而表现出白化特性, 无法进行光合作用, 只能存活数月, 直至自身含有的能量耗尽(Duke & Watkinson, 2002 ).繁殖体对不同光照强度有不同反应, 弱光条件下繁殖体通过扩大叶片面积和叶绿素含量增强光吸收, 强光条件下则相反, 最适的中度光照利于固碳作用(Ulqodry et al ., 2014 ).也有一些个例, 如分布范围狭窄的Pelliciera rhizophorae 在阴蔽处生长旺盛, 受到光照反而生长不良, 这一发现丰富了普遍认可的红树植物不耐阴的观念(Dangremond et al ., 2015 ).另外, 全黑条件下沙培的白骨壤茎生长的优势更加明显, 但生成的茎细长脆弱, 机械组织不发达, 对复杂潮间带的适应力减弱(邱广龙, 2005 ).不难解释, 林窗处的繁殖体幼苗生长更加迅速而强壮. ...
Shedding light on an extremophile life style through transcriptomics
2009
Occurrence of vivipary in
2008
Submarine pollination in seagrasses
1976
Chlorophyll-deficient propagules of
1
2002
... 光照是繁殖体定植的重要因素.前文提到胎生繁殖体表皮含有叶绿素, 能够进行光合作用为早期生长提供能量, 促进根茎生长.研究表明叶绿素缺失的繁殖体因缺乏色素而表现出白化特性, 无法进行光合作用, 只能存活数月, 直至自身含有的能量耗尽(Duke & Watkinson, 2002 ).繁殖体对不同光照强度有不同反应, 弱光条件下繁殖体通过扩大叶片面积和叶绿素含量增强光吸收, 强光条件下则相反, 最适的中度光照利于固碳作用(Ulqodry et al ., 2014 ).也有一些个例, 如分布范围狭窄的Pelliciera rhizophorae 在阴蔽处生长旺盛, 受到光照反而生长不良, 这一发现丰富了普遍认可的红树植物不耐阴的观念(Dangremond et al ., 2015 ).另外, 全黑条件下沙培的白骨壤茎生长的优势更加明显, 但生成的茎细长脆弱, 机械组织不发达, 对复杂潮间带的适应力减弱(邱广龙, 2005 ).不难解释, 林窗处的繁殖体幼苗生长更加迅速而强壮. ...
Handbook of Seed Technology for Genebanks . Volume II . Compendium of Specific Germination Information and Test Recommendations . International Board for Plant Genetic Resources, Rome
1985
The evolution of vivipary in flowering plants
1
1996
... “胎生”一词起源于哺乳动物, 然而在植物中也发现了类似的繁殖方式, 其定义为后代在母体上萌发并生长至一定阶段才离开母体(Elmqvist & Cox, 1996 ).根据这个定义, 胎生可分为真胎生与假胎生.真胎生是指通过有性生殖产生的后代在母体萌发的现象; 而假胎生是指植物通过无融合生殖或在有性生殖器官上产生无性繁殖体的现象, 与营养繁殖类似.真胎生与假胎生有着遗传和进化上的本质区别, 应当区分开来. ...
Mutations of genes in synthesis of the carotenoid precursors of ABA lead to pre-harvest sprouting and photo-oxidation in rice
2008
The ecology and physiology of viviparous and recalcitrant seeds
1
2000
... 植物激素在种子发育和休眠中具有广泛的作用.脱落酸(ABA)作为一个内源因子在调节种子成熟和发育过程中能够促进种子休眠, 抑制萌发.与非胎生物种相比, 胎生红树植物胚胎的ABA含量较低, 且在红树科中发现母体ABA含量高于胚胎, 因此低含量的ABA可能是红树植物胎生发生时种子没有脱水休眠的重要原因之一(Farnsworth & Farrant, 1998 ; Farnsworth, 2000 ).对秋茄树外源施加ABA发现果实个体及胚轴长度均小于对照组(洪礼伟等, 未发表资料), 表明其发育受到抑制, 进一步说明胚胎发育受到ABA影响, 推测胎生过程中ABA合成减少, 而不是胚胎发育对ABA不敏感. ...
Reductions in abscisic acid are linked with viviparous reproduction in mangroves
1
1998
... 植物激素在种子发育和休眠中具有广泛的作用.脱落酸(ABA)作为一个内源因子在调节种子成熟和发育过程中能够促进种子休眠, 抑制萌发.与非胎生物种相比, 胎生红树植物胚胎的ABA含量较低, 且在红树科中发现母体ABA含量高于胚胎, 因此低含量的ABA可能是红树植物胎生发生时种子没有脱水休眠的重要原因之一(Farnsworth & Farrant, 1998 ; Farnsworth, 2000 ).对秋茄树外源施加ABA发现果实个体及胚轴长度均小于对照组(洪礼伟等, 未发表资料), 表明其发育受到抑制, 进一步说明胚胎发育受到ABA影响, 推测胎生过程中ABA合成减少, 而不是胚胎发育对ABA不敏感. ...
Evolutionary and Ecological Physiology of Mangrove Seedlings: Correlates, Costs, and Consequences of Viviparous Reproduction . Harvard University, Cambridge
1
1997
... 真胎生现象在4类植物中已有报道(Farnsworth, 1997 ).首先是在遗传上以胎生为繁殖方式的植物, 如分布在8个属的红树植物(表1 )及两个属的海草(Tomlinson, 1986 ), 它们在自然生长环境条件下以胎生为最主要的繁殖方式, 繁殖体在母体上突破种皮或果皮并生长; 二是生长于热带、亚热带的某些顽拗性种子植物, 如少数仙人掌科植物(Cota-Sánchez, 2004 ); 三是某些处于特殊环境中的植物, 如水稻(Oryza sativa )、玉米(Zea mays )等, 其种子在高温、高湿条件下提前萌发, 但它们不从母体中吸收营养(叶能干, 1979 ); 四是与休眠有关的植物激素脱落酸(ABA)合成或响应通路有关的突变体植株, 或因种子中休眠基因功能的缺失不能进入休眠期而在母体直接开始萌发(Robertson, 1955 ), 后两类植物由于受外界条件诱导而形成提前萌发的性状, 在遗传本质上不是真正意义上的胎生, 但对于红树植物胎生现象研究仍有一定的借鉴作用.表1 列出已发表的以胎生为繁殖方式的植物. ...
The role of plant growth regulators in the development and germination of the desiccation-sensitive (recalcitrant) seeds of
1993
Development of the recalcitrant (homoiohydrous) seeds of
1
1992
... 隐胎生红树植物的胚胎发育方式与显胎生有所差别.海榄雌(Avicennia marina )胚胎发育至球形胚时, 胚孔吸根(micropylar haustorium)萎缩, 胚乳连同胚从珠被(种皮)中伸出, 进入到子房腔生长, 胚进而充满子房, 成熟时果实掉落(Farrant et al ., 1992 ).在蜡烛果属植物Aegiceras majus 中的情况较为不同, 胚珠受精后具有分生性的胚乳先行分裂, 待大量胚乳形成后, 合子才在胚孔端开始发育(Carey & Fraser, 1932 ).胚乳在胚孔处的分裂产生皱褶, 这些皱褶增加了吸收养分的面积, 相当于吸器的功能.经过一段时间的生长后, 胚占主导地位, 为了适应胚的这一变化, 胎座也伸长以维持胚的相对位置不变, 并保证了养料的持续供给, 而后胚根及胚轴的下部伸出种皮.随着胚逐渐增大, 种皮形成了类似帽子的结构, 罩在子叶上端.果实连同发育了的胚一起掉落, 在适当的条件下胚轴继续伸长, 直至胚根突破果皮, 侧根生长在松软的泥土上完成定植, 子叶也从果实中露出, 与包裹在其中的幼叶一起进行光合作用, 幼苗开始生长. ...
Presence of dehydrin-like proteins and levels of abscisic acid in recalcitrant (desiccation sensitive) seeds may be related to habitat
1996
Germination requirements of New Zealand native plants: A review
1991
Studies in Seeds and Fruits: An Investigation with the Balance . Williams and Norgate
1912
Viviparous-5 encodes phytoene desaturase, an enzyme essential for abscisic acid (ABA) accumulation and seed development in maize
1998
Rice SNF2 family helicase ENL1 is essential for syncytial endosperm development
1
2015
... 首先是子叶的形态不同.木榄属的子叶是二裂或三裂, 厚肉质, 成熟时不与下胚轴分开, 一直包被在果皮内与之同时掉落.角果木属、红树属及秋茄树属的子叶都是合生性, 呈圆柱体形, 前两个属子叶中的维管组织比秋茄树属的多(Juncosa & Tomlinson, 1988 ).待胚轴成熟, 子叶进行第二次生长后, 子叶基部的细胞分裂产生一个离区, 有助于下胚轴与子叶分离、掉落, 而子叶及果皮仍留在母体上(Juncosa, 1982 ).其次是促使突破种皮的原因也不一样.早期文献报道除木榄属的胎生发育是由于下胚轴伸长外, 红树属、角果木属、秋茄树属中都是由大量生长的胚乳突破种皮的(Juncosa & Tomlinson, 1988).在种皮被突破时, 胚仍处于较早期的阶段(在红树(Rhizophora apiculata )中甚至处于心形胚, 图1A ), 即由于胚乳的增加, 胚孔被打开, 胚吸收胚乳的营养, 开始生长.我们最近的研究发现秋茄树(Kandelia obovata )的胎生发育不是像Juncosa和Tomlinson (1988)所描述的依靠胚乳的大量生长突破种皮, 而是由于下胚轴的伸长, 即由位于茎尖分生组织下方的居间分生组织细胞进行分裂生长而成(图1F ).因此, 在秋茄树、木榄属中, 胎生萌发起始点为因下胚轴伸长使胚根伸出种皮的时刻, 这与非胎生的种子发育相似.胚乳在这一过程中的作用并不像在红树属中那样显著, 类似地, 水稻突变体endospermless1 (enl1 )缺乏胚乳, 表现出胎生现象(Hara et al ., 2015 ).胚乳存在与否及其功能可能影响胎生发育, 由此我们推测在红树植物中显胎生发育可能存在两种机制.一旦开始发育, 在这4个属中, 下胚轴的伸长均是居间分生组织细胞的快速分裂与细胞伸长导致的, 有的胚轴能达几十厘米悬挂在母树上. ...
Semi-viviparous embryo development and dehydrin expression in the mangrove
1
2010
... 脱水素蛋白(dehydrin protein)如late embryogenesis abundant (LEA)等与ABA之间存在密切的联系.Farrant等(1996)在非胎生物种成熟、未干燥的种子中检测到脱水素蛋白, 但在显胎生红树植物红树属、角果木属、木榄属和隐胎生红树植物海榄雌的成熟胚轴中则不能检测到脱水素, 然而在红茄苳(Rhizophora mucronata )早期胚胎发育时却有类脱水素蛋白的存在(Ismail et al ., 2010 ).脱水素在不同发育方式中的存在与否可能与生境有关. ...
Research paper interspecific variations in responses of mangrove seedlings to two contrasting salinities
1
2008
... 胎生对适应高盐环境具有一定意义.在胚轴发育过程中细胞液的盐分浓度变化不明显(Wang et al ., 2002 ), 但是胚轴在母体上发育成熟, 最终达到与母树生境的离子浓度一致(张宜辉, 2003 ).繁殖体掉落在滩涂后迅速形成侧根, 并与外界进行离子交换, 以快速适应盐度变化.Joshi (1933)认为, 种子的胎生是为了避免受到高盐生境的迫害, 使其顺利发芽乃至定植在海岸潮间带.实际上, 在不同盐度培养条件下, 红树植物幼苗均表现出低盐促进、高盐抑制的特征.邱广龙(2005)发现低盐度有利于隐胎生白骨壤(即海榄雌)繁殖体的萌根与幼苗生长, 胎生和非胎生红树植物繁殖体同样偏好低盐环境(Jayatissa et al ., 2008 ; Robert et al ., 2015 ).然而, 有报道指出胎生繁殖体的萌发需要一定的盐度, 某些红树繁殖体在淡水中无法正常生长(Wang et al ., 2011 ). ...
A suggested explanation of the prevalence of vivipary on the sea-shore
1
1933
... 长久以来胎生被认为是一个针对盐度适应的特征(Joshi, 1933 ), 但这一认识并没有被广泛接受, 因为其他的盐生植物并没有胎生现象.Wang等(2002)的研究发现秋茄树胚轴发育后期组织液离子浓度并没有明显变化, 推测胎生与抗盐性并没有直接的联系, 认为胎生并不是对某一个环境因子的适应, 而是对海岸潮间带多个生态因子综合响应的结果(闫中正等, 2004 ). ...
Buckwheat in India . National Bureau of Plant Genetic Resources Regional Station, Shimla
1991
Physiological studies in germination of mangroves
1
1972
... 胎生红树植物胚胎发育的另一个重要的特征是发育过程中繁殖器官盐离子浓度的变化.对于胚胎如何适应落地后的高盐生境, 有两种不同的结论, 即胎生是低盐化过程, 以及与此对立的胎生是盐分累积过程.郑文教和林鹏(1997)测定了秋茄树、木榄(Bruguiera gymnorhiza )和海莲(Bruguiera sexangula )胚轴发育过程中的盐离子, 发现随着胚轴成熟, 其Mg2+ 、Ca2+ 、Na+ 、K+ 、Cl- 含量呈下降趋势, 胚轴的盐含量明显低于母树果皮、叶片的盐含量, 且果皮、叶片的组织液Cl- 浓度与海水Cl- 浓度接近, 故认为胎生过程是一个低盐化的过程, 后代是通过与母体的长期接触来获得抗盐锻炼, 进而适应落地后的环境(郑文教等, 1992 ; 郑文教和林鹏, 1997 ).红树植物很可能起源于淡水环境, 胚胎发育是其生活史的重演, 这种胚胎发育低盐化的过程很可能是一种“返祖”现象.有趣的是, Joshi等(1972)发现显胎生植物胚轴发育过程中Mg2+ 、Ca2+ 、Na+ 、K+ 含量也逐渐下降, 但Cl- 含量逐渐增加.然而, 秋茄树成熟胚轴的Mg2+ 、Ca2+ 、Na+ 、K+ 、Cl- 含量与早期胚轴相比呈倍性增加(Wang et al ., 2002 ).部分研究者认为胎生繁殖体的盐抗性是逐渐适应从母体输送到幼苗的盐分而获得的(Lin, 1988 ).这两种不同的结论可能是由于对胎生发育过程阶段的划分不同及盐离子含量、浓度的高低定义不同, 甚至是物种不同而导致的.相比于胎生红树植物, 非胎生红树植物繁殖体发育过程中盐离子动态的研究相对较少, 但有证据表明非胎生的老鼠簕(Acanthus ilicifolius )幼苗Cl- 含量高于胎生物种Ceriops candolleana 、Avicennia officinalis 、Aegiceras majus (Joshi et al ., 1972 ), 其机制仍有待阐明.Mg2+ 、Ca2 、Na+ 、K+ 和Cl- 这5种离子是细胞渗透势的主要成分, 除了构成盐分浓度外, 还可能作为一种信号在种子成熟过程中起其他作用, 如Ca2+ 作为二级信使传导信号调节基因表达. ...
Developmental morphology of the embryo and seedling of
1
1982
... 首先是子叶的形态不同.木榄属的子叶是二裂或三裂, 厚肉质, 成熟时不与下胚轴分开, 一直包被在果皮内与之同时掉落.角果木属、红树属及秋茄树属的子叶都是合生性, 呈圆柱体形, 前两个属子叶中的维管组织比秋茄树属的多(Juncosa & Tomlinson, 1988 ).待胚轴成熟, 子叶进行第二次生长后, 子叶基部的细胞分裂产生一个离区, 有助于下胚轴与子叶分离、掉落, 而子叶及果皮仍留在母体上(Juncosa, 1982 ).其次是促使突破种皮的原因也不一样.早期文献报道除木榄属的胎生发育是由于下胚轴伸长外, 红树属、角果木属、秋茄树属中都是由大量生长的胚乳突破种皮的(Juncosa & Tomlinson, 1988).在种皮被突破时, 胚仍处于较早期的阶段(在红树(Rhizophora apiculata )中甚至处于心形胚, 图1A ), 即由于胚乳的增加, 胚孔被打开, 胚吸收胚乳的营养, 开始生长.我们最近的研究发现秋茄树(Kandelia obovata )的胎生发育不是像Juncosa和Tomlinson (1988)所描述的依靠胚乳的大量生长突破种皮, 而是由于下胚轴的伸长, 即由位于茎尖分生组织下方的居间分生组织细胞进行分裂生长而成(图1F ).因此, 在秋茄树、木榄属中, 胎生萌发起始点为因下胚轴伸长使胚根伸出种皮的时刻, 这与非胎生的种子发育相似.胚乳在这一过程中的作用并不像在红树属中那样显著, 类似地, 水稻突变体endospermless1 (enl1 )缺乏胚乳, 表现出胎生现象(Hara et al ., 2015 ).胚乳存在与否及其功能可能影响胎生发育, 由此我们推测在红树植物中显胎生发育可能存在两种机制.一旦开始发育, 在这4个属中, 下胚轴的伸长均是居间分生组织细胞的快速分裂与细胞伸长导致的, 有的胚轴能达几十厘米悬挂在母树上. ...
Systematic comparison and some biological characteristics of Rhizophoraceae and Anisophylleaceae
2
1988
... 首先是子叶的形态不同.木榄属的子叶是二裂或三裂, 厚肉质, 成熟时不与下胚轴分开, 一直包被在果皮内与之同时掉落.角果木属、红树属及秋茄树属的子叶都是合生性, 呈圆柱体形, 前两个属子叶中的维管组织比秋茄树属的多(Juncosa & Tomlinson, 1988 ).待胚轴成熟, 子叶进行第二次生长后, 子叶基部的细胞分裂产生一个离区, 有助于下胚轴与子叶分离、掉落, 而子叶及果皮仍留在母体上(Juncosa, 1982 ).其次是促使突破种皮的原因也不一样.早期文献报道除木榄属的胎生发育是由于下胚轴伸长外, 红树属、角果木属、秋茄树属中都是由大量生长的胚乳突破种皮的(Juncosa & Tomlinson, 1988).在种皮被突破时, 胚仍处于较早期的阶段(在红树(Rhizophora apiculata )中甚至处于心形胚, 图1A ), 即由于胚乳的增加, 胚孔被打开, 胚吸收胚乳的营养, 开始生长.我们最近的研究发现秋茄树(Kandelia obovata )的胎生发育不是像Juncosa和Tomlinson (1988)所描述的依靠胚乳的大量生长突破种皮, 而是由于下胚轴的伸长, 即由位于茎尖分生组织下方的居间分生组织细胞进行分裂生长而成(图1F ).因此, 在秋茄树、木榄属中, 胎生萌发起始点为因下胚轴伸长使胚根伸出种皮的时刻, 这与非胎生的种子发育相似.胚乳在这一过程中的作用并不像在红树属中那样显著, 类似地, 水稻突变体endospermless1 (enl1 )缺乏胚乳, 表现出胎生现象(Hara et al ., 2015 ).胚乳存在与否及其功能可能影响胎生发育, 由此我们推测在红树植物中显胎生发育可能存在两种机制.一旦开始发育, 在这4个属中, 下胚轴的伸长均是居间分生组织细胞的快速分裂与细胞伸长导致的, 有的胚轴能达几十厘米悬挂在母树上. ...
... 另外, 在显胎生红树植物发育的过程中, 有许多与功能相适应的结构、特征引起人们的注意: 首先是在红树属中, 与种皮相邻的最外层胚乳细胞特化出向内皱褶的壁, 被称为转运组织, 通过转运细胞, 水分和养料得以从种皮向胚转运, 但随着胚的不断生长, 这层转运细胞被挤压最后失去功能.在胎生海草Amphibolis 和Thalassodendron 中发现连接幼苗和母体的基部组织最外层也存在着运输养分的转运组织(Kuo & Kirkman, 1990 ).其次是胚轴在母体上生长时主根并不分化, 而是由数量较多的侧根进行分化, 并且在胚轴落地后的定植过程中起锚定作用.最后, 红树科中除了以显胎生为繁殖方式的4个属生活在滨海潮间带外, 其他的类群均为陆生非胎生种(Juncosa & Tomlinson, 1988 ).同时, Juncosa和Tomlinson (1988)认为在红树科中, 显胎生的许多繁殖特征, 如果实不开裂, 合生性子叶, 具有突出种子的胚乳, 由胚乳起始的发育过程, 胚乳中特化的转运细胞, 由侧根而不是主根进行胚轴(幼苗)定植等都是衍生的特征.由胚珠、种子、果实的形态特征推测显胎生的4个属是逐渐进化而适应水环境传播的(Tobe & Raven, 1988 ). ...
Genetic structures of natural populations of three mangrove species,
2004
Spatio-temporal pattern of specific gravity of mangrove diaspore: Implications for upstream dispersal
1
2015
... 但漂流的远近与显胎生、非胎生繁殖方式并没有直接关联, 而是与果实大小、形状、密度紧密相关(Ryck et al ., 2012 ).胚轴密度与生境中海水密度是否接近导致不一样的漂流模式, 例如显胎生的木榄成熟胚轴密度接近海水密度, 适应随潮水传播, 而隐胎生的蜡烛果胚轴密度远大于生境海水, 仅适合短时间内漂流(张宜辉, 2003 ).个体较大的红茄苳繁殖体比起角果木来说传播得更远, 而根系萌发迅速的角果木在向陆带(landward)有定植优势(Robert et al ., 2015 ).这些不同的扩散模式可能最终决定了红树林的分带(Kadoya & Inoue, 2015 ).如上所述, 胎生种子在成熟过程中一直保持高水分含量.代谢水的丢失会对种子造成不可挽回的损伤, 但是在胎生繁殖体中这种损失并不存在, 能够最大限度地保持种子活力.胎生繁殖体(胚轴)作为具有相对完整结构的幼苗, 其定植明显比休眠的种子更加迅速而有效, 是海岸潮间带复杂环境的优势适应性特征. ...
Anatomy of viviparous seagrasses seedlings of Amphibolis and Thalassodendron and their nutrient supply
1
1990
... 另外, 在显胎生红树植物发育的过程中, 有许多与功能相适应的结构、特征引起人们的注意: 首先是在红树属中, 与种皮相邻的最外层胚乳细胞特化出向内皱褶的壁, 被称为转运组织, 通过转运细胞, 水分和养料得以从种皮向胚转运, 但随着胚的不断生长, 这层转运细胞被挤压最后失去功能.在胎生海草Amphibolis 和Thalassodendron 中发现连接幼苗和母体的基部组织最外层也存在着运输养分的转运组织(Kuo & Kirkman, 1990 ).其次是胚轴在母体上生长时主根并不分化, 而是由数量较多的侧根进行分化, 并且在胚轴落地后的定植过程中起锚定作用.最后, 红树科中除了以显胎生为繁殖方式的4个属生活在滨海潮间带外, 其他的类群均为陆生非胎生种(Juncosa & Tomlinson, 1988 ).同时, Juncosa和Tomlinson (1988)认为在红树科中, 显胎生的许多繁殖特征, 如果实不开裂, 合生性子叶, 具有突出种子的胚乳, 由胚乳起始的发育过程, 胚乳中特化的转运细胞, 由侧根而不是主根进行胚轴(幼苗)定植等都是衍生的特征.由胚珠、种子、果实的形态特征推测显胎生的4个属是逐渐进化而适应水环境传播的(Tobe & Raven, 1988 ). ...
Rice mutants and genes related to organ development, morphogenesis and physiological traits
1
2005
... 如前所述, 某些作物中也存在着胎生现象, 表现为直接在植株上生根发芽, 称为成熟前萌发.这种在谷类作物中的胎生现象导致了大量减产, 因此对其分子研究较为深入.虽然这种作物提前萌发的现象在遗传进化上和红树植物的胎生有本质的区别, 但对研究红树植物胎生的分子机理有重要的借鉴意义.在玉米、水稻、拟南芥(Arabidopsis thaliana )等植物中发现的胎生调控基因基本都与ABA合成或响应通路有关(表2 ), 这与红树植物胎生过程中胚胎的ABA含量较低且能受外源ABA激素影响的现象相吻合.我们将这一系列与ABA合成及信号感受的基因(表2 )在玉米、水稻、拟南芥, 以及显胎生红树植物红树、隐胎生红树植物海榄雌和非胎生红树植物杯萼海桑(Sonneratia alba )的基因组数据中进行同源比对, 发现许多ABA通路中关键基因的蛋白序列在红树物种中具有高度保守性, 例如编码ABA生物合成第一步反应所需的玉米黄质环氧化酶的ABA1 基因(洪礼伟等, 未发表资料), 这也从另一个方面暗示着ABA很可能从分子水平上调控红树植物的胎生发育过程.值得一提的是, 在水稻中发现了对ABA信号不敏感并表现出胎生的突变体riv1-1 (rice vivipary 1-1 )、riv1-2 、riv2 及缺乏胚乳结构的enl1 (Kurata et al ., 2005 ).前3个突变体必须在外界湿润的条件下才能表现出提前萌发或提前萌发率大幅上升的现象, 而enl1 突变体由于缺乏在种子休眠中起重要作用的胚乳, 即使不施加水分也能提前萌发, 但在有水分的条件下会表现出更高的胎生率(Miyoshi et al ., 2000 ), 这说明胎生是内源基因与外界环境(很可能是湿度)共同作用的结果.因此, 在研究红树植物胎生现象时应当同时考虑基因与环境之间相互作用的可能性及重要性. ...
中国红树林的分布、面积和树种组成. 湿地科学,
1
2014
... 我国红树林面积在历史上曾达25 × 104 hm2 , 到2013年总面积不足2.5 × 104 hm2 (廖宝文和张乔民, 2014 ).目前我国红树林恢复造林的主要方法是利用显胎生或隐胎生繁殖体进行滩涂扦插(彭逸生等, 2008 ), 具有操作便易、成本较低、幼苗存活率高等优点.在深入理解胎生发育机制后, 对造林技术进行优化, 能有效地帮助恢复红树林湿地面积.同时, 在全球气候变化的影响下, 极端温度频繁出现、海平面上升等可能威胁红树植物的生存状态.而作为独特的繁殖方式, 胎生如何产生变化适应新的环境因素也是未来研究的重要方向. ...
Variation in propagule mass and its effect on carbon assimilation and seedling growth of red mangrove (
1
1995
... 胎生意味着种子在母体上萌发, 直至生长为成熟的繁殖体, 是一个能量不断积累的过程(闫中正等, 2004 ).此时的繁殖体储存大量的碳水化合物, 并且因为表皮含叶绿素能进行光合作用, 能够为独立生长初期和漂流过程提供足够的营养供给(Lin & Sternberg, 1995 ; Smith & Snedaker, 2000 ). ...
红树植物繁殖体发育过程的能量变化. 海洋科学,
1
2000
... 热值(caloric value)指单位质量干物质在完全燃烧后所释放出来的热量值, 可以反映出胎生繁殖体的能量水平(鲍雅静等, 2008 ).在繁殖体的发育过程中, 从花、果至成熟胚轴, 其热值逐渐降低, 且始终低于母体.这可能是由于生长发育过程中胚轴已经开始消耗能量, 以达到迅速突破果皮, 进而伸长的结果, 同时胚轴中低能量物质如纤维素等增加(林鹏等, 2000 ).此外, 灰分含量与热值变化趋势类似, 例如蜡烛果(Aegiceras corniculatum )胚轴在成熟过程中灰分含量降低, 但脱离母树后灰分含量逐步累积提高, 一些研究人员将灰分含量与盐离子联系起来, 认为蜡烛果胚轴脱离母树开始独立生长是一个大量吸收盐分的过程(林益明等, 2004 ; 赵胡等, 2004 ).虽然热值和灰分随着发育过程而降低, 但随着生物量的增加, 单个胚轴所含能量总体呈上升趋势, 为后续的传播和定植提供了足够的能耗支持. ...
Tannin dynamics of propagules and leaves of
1
2006
... 胎生红树植物由于下胚轴在母体上已经发育生长到一定长度, 脱离母体后有一定概率直接插入滩涂与沙地, 其侧根能在落地后几小时内萌发, 又由于次生木质部等的作用, 胚轴能由水平到直立生长(Cheeseman, 2012 ).同时, 红树植物繁殖体主要是被动地依靠水体和泥沙流动进行传播, 在这种随波而流的前提下, 显胎生红树植物(例如木榄、秋茄树)胚轴所具有的厚角质层、发达的皮层通气组织以及表皮缺乏气孔等系列特征为长时间漂流提供支持, 并可避免海水的渗入(陈月琴等, 1995 ).而且繁殖体在漂流过程中, 能应对水体缺氧状况, 降低呼吸速率, 相应地减少能耗(Brown et al ., 1969 ).胎生繁殖体单宁含量比花期高, 在成熟过程中虽略有波动, 但总体还是呈积累趋势(Lin et al ., 2006 ).高含量的单宁使胎生繁殖体在漂流过程中不被海水腐蚀和动物啃食, 保持长时间的生命力, 有利于进行长距离扩散(林益明等, 2005 ), 这即是红树植物得以漂洋过海的秘密. ...
深圳福田几种红树植物繁殖体与不同发育阶段叶片热值研究. 海洋科学,
1
2004
... 热值(caloric value)指单位质量干物质在完全燃烧后所释放出来的热量值, 可以反映出胎生繁殖体的能量水平(鲍雅静等, 2008 ).在繁殖体的发育过程中, 从花、果至成熟胚轴, 其热值逐渐降低, 且始终低于母体.这可能是由于生长发育过程中胚轴已经开始消耗能量, 以达到迅速突破果皮, 进而伸长的结果, 同时胚轴中低能量物质如纤维素等增加(林鹏等, 2000 ).此外, 灰分含量与热值变化趋势类似, 例如蜡烛果(Aegiceras corniculatum )胚轴在成熟过程中灰分含量降低, 但脱离母树后灰分含量逐步累积提高, 一些研究人员将灰分含量与盐离子联系起来, 认为蜡烛果胚轴脱离母树开始独立生长是一个大量吸收盐分的过程(林益明等, 2004 ; 赵胡等, 2004 ).虽然热值和灰分随着发育过程而降低, 但随着生物量的增加, 单个胚轴所含能量总体呈上升趋势, 为后续的传播和定植提供了足够的能耗支持. ...
红树林单宁的研究进展. 海洋科学,
1
2005
... 胎生红树植物由于下胚轴在母体上已经发育生长到一定长度, 脱离母体后有一定概率直接插入滩涂与沙地, 其侧根能在落地后几小时内萌发, 又由于次生木质部等的作用, 胚轴能由水平到直立生长(Cheeseman, 2012 ).同时, 红树植物繁殖体主要是被动地依靠水体和泥沙流动进行传播, 在这种随波而流的前提下, 显胎生红树植物(例如木榄、秋茄树)胚轴所具有的厚角质层、发达的皮层通气组织以及表皮缺乏气孔等系列特征为长时间漂流提供支持, 并可避免海水的渗入(陈月琴等, 1995 ).而且繁殖体在漂流过程中, 能应对水体缺氧状况, 降低呼吸速率, 相应地减少能耗(Brown et al ., 1969 ).胎生繁殖体单宁含量比花期高, 在成熟过程中虽略有波动, 但总体还是呈积累趋势(Lin et al ., 2006 ).高含量的单宁使胎生繁殖体在漂流过程中不被海水腐蚀和动物啃食, 保持长时间的生命力, 有利于进行长距离扩散(林益明等, 2005 ), 这即是红树植物得以漂洋过海的秘密. ...
红树植物胎生过程中ABA、GA3和糖分含量的变化
1
2012
... 另外, 与ABA有拮抗作用的赤霉素GA3在种子发育过程中始终维持较高的水平, 卢磊(2012) 推测GA3与ABA通过调控α淀粉酶影响种子中糖分的含量和比例, 从而影响胎生种子的萌发.与其他植物类似, 激素之间的相互作用在红树植物胎生发育过程中必不可少, 但仍存在许多问题, 如产生部位、运输方式、具体作用部位、作用机制、不同激素如何互作等. ...
Pore-water chemistry in mangrove sediments: Relationship with species composition and developmental stages (French Guiana)
1
2004
... 不仅成年红树植物需要面对潮汐的冲击, 而且胎生繁殖体受到更大干扰.胎生繁殖体可以忍受长期浸淹的缺氧条件生长发育成幼苗, 合适的潮汐(水淹时间长度及间隔)能在一定程度上促进繁殖体生长(Chen et al ., 2004 ).随海水漂流的繁殖体经长时间浸淹, 根伸长推迟, 茎叶生长表现得相当缓慢, 所以相对性缺水(例如进入潮落时的滩涂)促使繁殖体萌根及定植(Robert et al ., 2015 ).当然, 水体中的盐度和pH值处于一个不断变化的过程(Marchand et al ., 2004 ), 不同种类红树植物对水淹也具有不同的适应特征(例如多种类的根系、叶片结构), 使红树植物对潮汐的响应复杂化. ...
Photosynthetic and stomatal responses of two mangrove species,
1984
A seed storage study of maga
1942
Maize phytoene desaturase and ζ-carotene desaturase catalyse a poly-Z desaturation pathway: Implications for genetic engineering of carotenoid content among cereal crops
2003
Molecular analysis of
1989
The viviparous-1 developmental gene of maize encodes a novel transcriptional activator
1991
Leafy cotyledon mutants of
1994
Microarray analysis of 7029 gene expression patterns in Burma mangrove under high-salinity stress
1
2007
... 红树植物在抗盐、抗重金属污染等方面的逆境生理生态特征及其分子生态学机制已有报道(张凤琴等, 2005 ; Miyama & Hanagata, 2007 ; Wong et al ., 2007 ; 程皓等, 2009 ; Parida & Jha, 2010 ), 但对红树植物独特的繁殖方式——胎生的分子机制研究甚少.转录组研究基本集中在红树属植物Rhizophora mangle 中, 目前仅此物种的花蕾、幼嫩及成熟的胚轴有转录组数据.Dassanayake等(2009)对R . mangle 不同阶段的营养器官、繁殖器官的转录组进行基因功能注释(gene ontology)分析, 在对所有表达的基因功能进行分类(分子功能、生物过程、细胞组分)比较后发现, “分子功能(molecular function)+生物过程(biological process)”共同表达的基因是“分子功能(molecular function)+细胞组分(cellular component)”共同表达的基因数量的两倍, 说明通过转录水平调控生理过程(而非改变细胞组分)有可能与红树植物胎生的生长发育过程存在密切关系.然而,由于采样阶段、基因组资源、信息分析、缺乏基因功能验证实验等的局限, 高达55%的序列没有功能注释, 而且未对花蕾、胚轴的发育进行深入分析, 此实验未能回答红树植物胎生发育的一系列问题. ...
Characterization of viviparous mutants in rice
1
2000
... 如前所述, 某些作物中也存在着胎生现象, 表现为直接在植株上生根发芽, 称为成熟前萌发.这种在谷类作物中的胎生现象导致了大量减产, 因此对其分子研究较为深入.虽然这种作物提前萌发的现象在遗传进化上和红树植物的胎生有本质的区别, 但对研究红树植物胎生的分子机理有重要的借鉴意义.在玉米、水稻、拟南芥(Arabidopsis thaliana )等植物中发现的胎生调控基因基本都与ABA合成或响应通路有关(表2 ), 这与红树植物胎生过程中胚胎的ABA含量较低且能受外源ABA激素影响的现象相吻合.我们将这一系列与ABA合成及信号感受的基因(表2 )在玉米、水稻、拟南芥, 以及显胎生红树植物红树、隐胎生红树植物海榄雌和非胎生红树植物杯萼海桑(Sonneratia alba )的基因组数据中进行同源比对, 发现许多ABA通路中关键基因的蛋白序列在红树物种中具有高度保守性, 例如编码ABA生物合成第一步反应所需的玉米黄质环氧化酶的ABA1 基因(洪礼伟等, 未发表资料), 这也从另一个方面暗示着ABA很可能从分子水平上调控红树植物的胎生发育过程.值得一提的是, 在水稻中发现了对ABA信号不敏感并表现出胎生的突变体riv1-1 (rice vivipary 1-1 )、riv1-2 、riv2 及缺乏胚乳结构的enl1 (Kurata et al ., 2005 ).前3个突变体必须在外界湿润的条件下才能表现出提前萌发或提前萌发率大幅上升的现象, 而enl1 突变体由于缺乏在种子休眠中起重要作用的胚乳, 即使不施加水分也能提前萌发, 但在有水分的条件下会表现出更高的胎生率(Miyoshi et al ., 2000 ), 这说明胎生是内源基因与外界环境(很可能是湿度)共同作用的结果.因此, 在研究红树植物胎生现象时应当同时考虑基因与环境之间相互作用的可能性及重要性. ...
Salt tolerance mechanisms in mangroves: A review
1
2010
... 红树植物在抗盐、抗重金属污染等方面的逆境生理生态特征及其分子生态学机制已有报道(张凤琴等, 2005 ; Miyama & Hanagata, 2007 ; Wong et al ., 2007 ; 程皓等, 2009 ; Parida & Jha, 2010 ), 但对红树植物独特的繁殖方式——胎生的分子机制研究甚少.转录组研究基本集中在红树属植物Rhizophora mangle 中, 目前仅此物种的花蕾、幼嫩及成熟的胚轴有转录组数据.Dassanayake等(2009)对R . mangle 不同阶段的营养器官、繁殖器官的转录组进行基因功能注释(gene ontology)分析, 在对所有表达的基因功能进行分类(分子功能、生物过程、细胞组分)比较后发现, “分子功能(molecular function)+生物过程(biological process)”共同表达的基因是“分子功能(molecular function)+细胞组分(cellular component)”共同表达的基因数量的两倍, 说明通过转录水平调控生理过程(而非改变细胞组分)有可能与红树植物胎生的生长发育过程存在密切关系.然而,由于采样阶段、基因组资源、信息分析、缺乏基因功能验证实验等的局限, 高达55%的序列没有功能注释, 而且未对花蕾、胚轴的发育进行深入分析, 此实验未能回答红树植物胎生发育的一系列问题. ...
红树林湿地恢复研究进展. 生态学报,
1
2008
... 我国红树林面积在历史上曾达25 × 104 hm2 , 到2013年总面积不足2.5 × 104 hm2 (廖宝文和张乔民, 2014 ).目前我国红树林恢复造林的主要方法是利用显胎生或隐胎生繁殖体进行滩涂扦插(彭逸生等, 2008 ), 具有操作便易、成本较低、幼苗存活率高等优点.在深入理解胎生发育机制后, 对造林技术进行优化, 能有效地帮助恢复红树林湿地面积.同时, 在全球气候变化的影响下, 极端温度频繁出现、海平面上升等可能威胁红树植物的生存状态.而作为独特的繁殖方式, 胎生如何产生变化适应新的环境因素也是未来研究的重要方向. ...
Principles of Dispersal in Higher Plants . Springer-Verlag
1972
The maize viviparous10/viviparous13 locus encodes the
2006
红树植物白骨壤繁殖生态研究与果实品质分析
1
2005
... 光照是繁殖体定植的重要因素.前文提到胎生繁殖体表皮含有叶绿素, 能够进行光合作用为早期生长提供能量, 促进根茎生长.研究表明叶绿素缺失的繁殖体因缺乏色素而表现出白化特性, 无法进行光合作用, 只能存活数月, 直至自身含有的能量耗尽(Duke & Watkinson, 2002 ).繁殖体对不同光照强度有不同反应, 弱光条件下繁殖体通过扩大叶片面积和叶绿素含量增强光吸收, 强光条件下则相反, 最适的中度光照利于固碳作用(Ulqodry et al ., 2014 ).也有一些个例, 如分布范围狭窄的Pelliciera rhizophorae 在阴蔽处生长旺盛, 受到光照反而生长不良, 这一发现丰富了普遍认可的红树植物不耐阴的观念(Dangremond et al ., 2015 ).另外, 全黑条件下沙培的白骨壤茎生长的优势更加明显, 但生成的茎细长脆弱, 机械组织不发达, 对复杂潮间带的适应力减弱(邱广龙, 2005 ).不难解释, 林窗处的繁殖体幼苗生长更加迅速而强壮. ...
Flowering in a bamboo,
2003
Viviparous mangrove propagules of
3
2015
... 但漂流的远近与显胎生、非胎生繁殖方式并没有直接关联, 而是与果实大小、形状、密度紧密相关(Ryck et al ., 2012 ).胚轴密度与生境中海水密度是否接近导致不一样的漂流模式, 例如显胎生的木榄成熟胚轴密度接近海水密度, 适应随潮水传播, 而隐胎生的蜡烛果胚轴密度远大于生境海水, 仅适合短时间内漂流(张宜辉, 2003 ).个体较大的红茄苳繁殖体比起角果木来说传播得更远, 而根系萌发迅速的角果木在向陆带(landward)有定植优势(Robert et al ., 2015 ).这些不同的扩散模式可能最终决定了红树林的分带(Kadoya & Inoue, 2015 ).如上所述, 胎生种子在成熟过程中一直保持高水分含量.代谢水的丢失会对种子造成不可挽回的损伤, 但是在胎生繁殖体中这种损失并不存在, 能够最大限度地保持种子活力.胎生繁殖体(胚轴)作为具有相对完整结构的幼苗, 其定植明显比休眠的种子更加迅速而有效, 是海岸潮间带复杂环境的优势适应性特征. ...
... 胎生对适应高盐环境具有一定意义.在胚轴发育过程中细胞液的盐分浓度变化不明显(Wang et al ., 2002 ), 但是胚轴在母体上发育成熟, 最终达到与母树生境的离子浓度一致(张宜辉, 2003 ).繁殖体掉落在滩涂后迅速形成侧根, 并与外界进行离子交换, 以快速适应盐度变化.Joshi (1933)认为, 种子的胎生是为了避免受到高盐生境的迫害, 使其顺利发芽乃至定植在海岸潮间带.实际上, 在不同盐度培养条件下, 红树植物幼苗均表现出低盐促进、高盐抑制的特征.邱广龙(2005)发现低盐度有利于隐胎生白骨壤(即海榄雌)繁殖体的萌根与幼苗生长, 胎生和非胎生红树植物繁殖体同样偏好低盐环境(Jayatissa et al ., 2008 ; Robert et al ., 2015 ).然而, 有报道指出胎生繁殖体的萌发需要一定的盐度, 某些红树繁殖体在淡水中无法正常生长(Wang et al ., 2011 ). ...
... 不仅成年红树植物需要面对潮汐的冲击, 而且胎生繁殖体受到更大干扰.胎生繁殖体可以忍受长期浸淹的缺氧条件生长发育成幼苗, 合适的潮汐(水淹时间长度及间隔)能在一定程度上促进繁殖体生长(Chen et al ., 2004 ).随海水漂流的繁殖体经长时间浸淹, 根伸长推迟, 茎叶生长表现得相当缓慢, 所以相对性缺水(例如进入潮落时的滩涂)促使繁殖体萌根及定植(Robert et al ., 2015 ).当然, 水体中的盐度和pH值处于一个不断变化的过程(Marchand et al ., 2004 ), 不同种类红树植物对水淹也具有不同的适应特征(例如多种类的根系、叶片结构), 使红树植物对潮汐的响应复杂化. ...
The genetics of vivipary in maize
1
1955
... 真胎生现象在4类植物中已有报道(Farnsworth, 1997 ).首先是在遗传上以胎生为繁殖方式的植物, 如分布在8个属的红树植物(表1 )及两个属的海草(Tomlinson, 1986 ), 它们在自然生长环境条件下以胎生为最主要的繁殖方式, 繁殖体在母体上突破种皮或果皮并生长; 二是生长于热带、亚热带的某些顽拗性种子植物, 如少数仙人掌科植物(Cota-Sánchez, 2004 ); 三是某些处于特殊环境中的植物, 如水稻(Oryza sativa )、玉米(Zea mays )等, 其种子在高温、高湿条件下提前萌发, 但它们不从母体中吸收营养(叶能干, 1979 ); 四是与休眠有关的植物激素脱落酸(ABA)合成或响应通路有关的突变体植株, 或因种子中休眠基因功能的缺失不能进入休眠期而在母体直接开始萌发(Robertson, 1955 ), 后两类植物由于受外界条件诱导而形成提前萌发的性状, 在遗传本质上不是真正意义上的胎生, 但对于红树植物胎生现象研究仍有一定的借鉴作用.表1 列出已发表的以胎生为繁殖方式的植物. ...
Size does matter, but not only size: Two alternative dispersal strategies for viviparous mangrove propagules
1
2012
... 但漂流的远近与显胎生、非胎生繁殖方式并没有直接关联, 而是与果实大小、形状、密度紧密相关(Ryck et al ., 2012 ).胚轴密度与生境中海水密度是否接近导致不一样的漂流模式, 例如显胎生的木榄成熟胚轴密度接近海水密度, 适应随潮水传播, 而隐胎生的蜡烛果胚轴密度远大于生境海水, 仅适合短时间内漂流(张宜辉, 2003 ).个体较大的红茄苳繁殖体比起角果木来说传播得更远, 而根系萌发迅速的角果木在向陆带(landward)有定植优势(Robert et al ., 2015 ).这些不同的扩散模式可能最终决定了红树林的分带(Kadoya & Inoue, 2015 ).如上所述, 胎生种子在成熟过程中一直保持高水分含量.代谢水的丢失会对种子造成不可挽回的损伤, 但是在胎生繁殖体中这种损失并不存在, 能够最大限度地保持种子活力.胎生繁殖体(胚轴)作为具有相对完整结构的幼苗, 其定植明显比休眠的种子更加迅速而有效, 是海岸潮间带复杂环境的优势适应性特征. ...
Molecular phylogeny of Rhizophoraceae based on
1
1999
... 遗传多样性在生物进化过程中起着重要作用, 因此研究红树植物的种群遗传结构对于理解红树植物的进化以及完善保育工作都具有重要的意义.Kado等(2004)通过分子标记技术, 对显胎生红树植物秋茄树、隐胎生红树植物海榄雌和非胎生红树植物榄李(Lumnitzera racemosa )在越南和日本的种群遗传多样性进行比较分析后发现, 繁殖体特性、地理分布以及自然干扰可能会影响红树植物种群的遗传结构, 但该研究未能解释不同的繁殖方式是如何影响种群遗传结构的.胎生物种分布在5个科8个属中, 这些科中的其他非红树植物不以胎生为繁殖方式, 说明红树植物的胎生在适应滨海特殊环境时以趋同进化的方式发展.Shi等(2005)分析了26个属的红树植物(包括胎生、非胎生植物类型)的rbc L、18S核糖体、mat R基因数据, 并构建分子系统进化树证实了胎生与泌盐这两个特征在红树植物中是独立起源的, 且胎生现象是多起源的, 但以显胎生为繁殖方式的红树科4个属在进化过程中有共同的祖先(Setoguchi et al ., 1999 ; Shi et al ., 2005 ).然而, 目前对显胎生、隐胎生和非胎生之间如何进化的关系却鲜有研究. ...
Kandelia obovata (Rhizophoraceae), a new mangrove species from Eastern Asia
2003
Molecular phylogenetic analysis of mangroves: Independent evolutionary origins of vivipary and salt secretion
2
2005
... 遗传多样性在生物进化过程中起着重要作用, 因此研究红树植物的种群遗传结构对于理解红树植物的进化以及完善保育工作都具有重要的意义.Kado等(2004)通过分子标记技术, 对显胎生红树植物秋茄树、隐胎生红树植物海榄雌和非胎生红树植物榄李(Lumnitzera racemosa )在越南和日本的种群遗传多样性进行比较分析后发现, 繁殖体特性、地理分布以及自然干扰可能会影响红树植物种群的遗传结构, 但该研究未能解释不同的繁殖方式是如何影响种群遗传结构的.胎生物种分布在5个科8个属中, 这些科中的其他非红树植物不以胎生为繁殖方式, 说明红树植物的胎生在适应滨海特殊环境时以趋同进化的方式发展.Shi等(2005)分析了26个属的红树植物(包括胎生、非胎生植物类型)的rbc L、18S核糖体、mat R基因数据, 并构建分子系统进化树证实了胎生与泌盐这两个特征在红树植物中是独立起源的, 且胎生现象是多起源的, 但以显胎生为繁殖方式的红树科4个属在进化过程中有共同的祖先(Setoguchi et al ., 1999 ; Shi et al ., 2005 ).然而, 目前对显胎生、隐胎生和非胎生之间如何进化的关系却鲜有研究. ...
... 从进化角度看, 现有的研究表明显胎生与隐胎生是分别进化而来的(Shi et al ., 2005 ), 而且分布在不同的科属中.更有趣的是, 这些红树植物的陆生近缘种(同科不同属)不是胎生物种.因此, 可以推断胎生的进化与红树植物的生境有关.更重要的是, 由于红树植物胎生是个自然的遗传程序, 需要一系列来自基因组的遗传信息, 即基因及其表达产物, 因此, 从基因组中找到这些遗传信息是将来胎生研究的重要方向.比较基因组的研究不仅可以找到胎生红树植物特有的基因簇, 而且也能得到在进化过程中产生的基因的特殊表达、调控方式及其转录组、蛋白组特征.在漫长的进化过程中, 积累在遗传物质中的信息决定了这一现象的发生, 不同基因在不同阶段、不同结构的表达, 各种转录因子的调控乃至这些基因架构成的网络是各种形态结构、生理生化变化的本质.这些组学水平的分析结果将对指导分子、生理乃至生态实验起到重要的启发作用.要理解红树植物胎生现象的遗传、生态、进化的本质, 需要多个领域许多科学家的共同努力. ...
Fruit characters and seed germination of
1977
Lack of dormancy in seeds of New Zealand plants
1979
Activator mutagenesis of the
2003
Hypocotyl function in seedling development of the red mangrove,
1
2000
... 胎生意味着种子在母体上萌发, 直至生长为成熟的繁殖体, 是一个能量不断积累的过程(闫中正等, 2004 ).此时的繁殖体储存大量的碳水化合物, 并且因为表皮含叶绿素能进行光合作用, 能够为独立生长初期和漂流过程提供足够的营养供给(Lin & Sternberg, 1995 ; Smith & Snedaker, 2000 ). ...
Growth and metabolism of the embryo and attached seedling of the viviparous mangrove,
1
1975
... 胎生红树植物胚胎发育具有两个重要的特点, 首先是种子不经脱水且不经休眠或短暂休眠就开始发育生长.其次, 研究发现胎生植物繁殖器官都有很高的含水量, 胚胎、种皮、果皮含水量均高于54% (Sussex, 1975 ), 在发育过程中Rhizophora mangle 的胚胎含水量始终高于种皮和果皮, 充足的水分供应为发育和新陈代谢提供了必要条件.相反, 进行休眠的种子成熟后干燥水分会降低至8%左右(Vertucci, 1989 ), 以便于储藏, 但新陈代谢、生长发育各个生理过程均处于停止状态, 等待新水源与其他条件成熟以后再行萌发, 这样可以避免冬天或干旱等不利的季节条件.由于红树植物生长在环境水分、温度都适宜生长的地方, 它们进化出不用经过休眠的方式来适宜这种环境, 以期得到最高的生长量和最短的繁殖周期. ...
The maize viviparous 15 locus encodes the molybdopterin synthase small subunit
2006
Trends in the evolution of dicotyledonous seeds based on character associations, with special reference to pachychalazy and recalcitrance
1991
Seed morphology and anatomy of Rhizophoraceae, inter- and infrafamilial relationships
1
1988
... 另外, 在显胎生红树植物发育的过程中, 有许多与功能相适应的结构、特征引起人们的注意: 首先是在红树属中, 与种皮相邻的最外层胚乳细胞特化出向内皱褶的壁, 被称为转运组织, 通过转运细胞, 水分和养料得以从种皮向胚转运, 但随着胚的不断生长, 这层转运细胞被挤压最后失去功能.在胎生海草Amphibolis 和Thalassodendron 中发现连接幼苗和母体的基部组织最外层也存在着运输养分的转运组织(Kuo & Kirkman, 1990 ).其次是胚轴在母体上生长时主根并不分化, 而是由数量较多的侧根进行分化, 并且在胚轴落地后的定植过程中起锚定作用.最后, 红树科中除了以显胎生为繁殖方式的4个属生活在滨海潮间带外, 其他的类群均为陆生非胎生种(Juncosa & Tomlinson, 1988 ).同时, Juncosa和Tomlinson (1988)认为在红树科中, 显胎生的许多繁殖特征, 如果实不开裂, 合生性子叶, 具有突出种子的胚乳, 由胚乳起始的发育过程, 胚乳中特化的转运细胞, 由侧根而不是主根进行胚轴(幼苗)定植等都是衍生的特征.由胚珠、种子、果实的形态特征推测显胎生的4个属是逐渐进化而适应水环境传播的(Tobe & Raven, 1988 ). ...
Study on photosynthetic responses and chlorophyll fluorescence in
1
2014
... 光照是繁殖体定植的重要因素.前文提到胎生繁殖体表皮含有叶绿素, 能够进行光合作用为早期生长提供能量, 促进根茎生长.研究表明叶绿素缺失的繁殖体因缺乏色素而表现出白化特性, 无法进行光合作用, 只能存活数月, 直至自身含有的能量耗尽(Duke & Watkinson, 2002 ).繁殖体对不同光照强度有不同反应, 弱光条件下繁殖体通过扩大叶片面积和叶绿素含量增强光吸收, 强光条件下则相反, 最适的中度光照利于固碳作用(Ulqodry et al ., 2014 ).也有一些个例, 如分布范围狭窄的Pelliciera rhizophorae 在阴蔽处生长旺盛, 受到光照反而生长不良, 这一发现丰富了普遍认可的红树植物不耐阴的观念(Dangremond et al ., 2015 ).另外, 全黑条件下沙培的白骨壤茎生长的优势更加明显, 但生成的茎细长脆弱, 机械组织不发达, 对复杂潮间带的适应力减弱(邱广龙, 2005 ).不难解释, 林窗处的繁殖体幼苗生长更加迅速而强壮. ...
A contribution to the embryology of Sonneratiaceae
1
1937
... 非胎生红树植物无瓣海桑(Sonneratia apetala ) 是多种子果实, 其胚胎发育过程与传统的荠菜胚一致, 但胚囊的中央部分始终不被胚乳填满, 胚乳在胚孔端有极性积累, 且到后期胚乳完全被胚吸收, 为无胚乳种子(Venkateswarlu, 1937 ).果实成熟后从树上掉落, 果皮开裂, 释放种子. ...
The effects of low water contents on physiological activities of seeds
1
1989
... 胎生红树植物胚胎发育具有两个重要的特点, 首先是种子不经脱水且不经休眠或短暂休眠就开始发育生长.其次, 研究发现胎生植物繁殖器官都有很高的含水量, 胚胎、种皮、果皮含水量均高于54% (Sussex, 1975 ), 在发育过程中Rhizophora mangle 的胚胎含水量始终高于种皮和果皮, 充足的水分供应为发育和新陈代谢提供了必要条件.相反, 进行休眠的种子成熟后干燥水分会降低至8%左右(Vertucci, 1989 ), 以便于储藏, 但新陈代谢、生长发育各个生理过程均处于停止状态, 等待新水源与其他条件成熟以后再行萌发, 这样可以避免冬天或干旱等不利的季节条件.由于红树植物生长在环境水分、温度都适宜生长的地方, 它们进化出不用经过休眠的方式来适宜这种环境, 以期得到最高的生长量和最短的繁殖周期. ...
An autogamous rainforest species of
1999
Changes in the main osmotica during the development of
2
2002
... 胎生红树植物胚胎发育的另一个重要的特征是发育过程中繁殖器官盐离子浓度的变化.对于胚胎如何适应落地后的高盐生境, 有两种不同的结论, 即胎生是低盐化过程, 以及与此对立的胎生是盐分累积过程.郑文教和林鹏(1997)测定了秋茄树、木榄(Bruguiera gymnorhiza )和海莲(Bruguiera sexangula )胚轴发育过程中的盐离子, 发现随着胚轴成熟, 其Mg2+ 、Ca2+ 、Na+ 、K+ 、Cl- 含量呈下降趋势, 胚轴的盐含量明显低于母树果皮、叶片的盐含量, 且果皮、叶片的组织液Cl- 浓度与海水Cl- 浓度接近, 故认为胎生过程是一个低盐化的过程, 后代是通过与母体的长期接触来获得抗盐锻炼, 进而适应落地后的环境(郑文教等, 1992 ; 郑文教和林鹏, 1997 ).红树植物很可能起源于淡水环境, 胚胎发育是其生活史的重演, 这种胚胎发育低盐化的过程很可能是一种“返祖”现象.有趣的是, Joshi等(1972)发现显胎生植物胚轴发育过程中Mg2+ 、Ca2+ 、Na+ 、K+ 含量也逐渐下降, 但Cl- 含量逐渐增加.然而, 秋茄树成熟胚轴的Mg2+ 、Ca2+ 、Na+ 、K+ 、Cl- 含量与早期胚轴相比呈倍性增加(Wang et al ., 2002 ).部分研究者认为胎生繁殖体的盐抗性是逐渐适应从母体输送到幼苗的盐分而获得的(Lin, 1988 ).这两种不同的结论可能是由于对胎生发育过程阶段的划分不同及盐离子含量、浓度的高低定义不同, 甚至是物种不同而导致的.相比于胎生红树植物, 非胎生红树植物繁殖体发育过程中盐离子动态的研究相对较少, 但有证据表明非胎生的老鼠簕(Acanthus ilicifolius )幼苗Cl- 含量高于胎生物种Ceriops candolleana 、Avicennia officinalis 、Aegiceras majus (Joshi et al ., 1972 ), 其机制仍有待阐明.Mg2+ 、Ca2 、Na+ 、K+ 和Cl- 这5种离子是细胞渗透势的主要成分, 除了构成盐分浓度外, 还可能作为一种信号在种子成熟过程中起其他作用, 如Ca2+ 作为二级信使传导信号调节基因表达. ...
... 胎生对适应高盐环境具有一定意义.在胚轴发育过程中细胞液的盐分浓度变化不明显(Wang et al ., 2002 ), 但是胚轴在母体上发育成熟, 最终达到与母树生境的离子浓度一致(张宜辉, 2003 ).繁殖体掉落在滩涂后迅速形成侧根, 并与外界进行离子交换, 以快速适应盐度变化.Joshi (1933)认为, 种子的胎生是为了避免受到高盐生境的迫害, 使其顺利发芽乃至定植在海岸潮间带.实际上, 在不同盐度培养条件下, 红树植物幼苗均表现出低盐促进、高盐抑制的特征.邱广龙(2005)发现低盐度有利于隐胎生白骨壤(即海榄雌)繁殖体的萌根与幼苗生长, 胎生和非胎生红树植物繁殖体同样偏好低盐环境(Jayatissa et al ., 2008 ; Robert et al ., 2015 ).然而, 有报道指出胎生繁殖体的萌发需要一定的盐度, 某些红树繁殖体在淡水中无法正常生长(Wang et al ., 2011 ). ...
Mangroves: Obligate or facultative halophytes? A review
1
2011
... 胎生对适应高盐环境具有一定意义.在胚轴发育过程中细胞液的盐分浓度变化不明显(Wang et al ., 2002 ), 但是胚轴在母体上发育成熟, 最终达到与母树生境的离子浓度一致(张宜辉, 2003 ).繁殖体掉落在滩涂后迅速形成侧根, 并与外界进行离子交换, 以快速适应盐度变化.Joshi (1933)认为, 种子的胎生是为了避免受到高盐生境的迫害, 使其顺利发芽乃至定植在海岸潮间带.实际上, 在不同盐度培养条件下, 红树植物幼苗均表现出低盐促进、高盐抑制的特征.邱广龙(2005)发现低盐度有利于隐胎生白骨壤(即海榄雌)繁殖体的萌根与幼苗生长, 胎生和非胎生红树植物繁殖体同样偏好低盐环境(Jayatissa et al ., 2008 ; Robert et al ., 2015 ).然而, 有报道指出胎生繁殖体的萌发需要一定的盐度, 某些红树繁殖体在淡水中无法正常生长(Wang et al ., 2011 ). ...
Isolation of salinity tolerant genes from the mangrove plant,
1
2007
... 红树植物在抗盐、抗重金属污染等方面的逆境生理生态特征及其分子生态学机制已有报道(张凤琴等, 2005 ; Miyama & Hanagata, 2007 ; Wong et al ., 2007 ; 程皓等, 2009 ; Parida & Jha, 2010 ), 但对红树植物独特的繁殖方式——胎生的分子机制研究甚少.转录组研究基本集中在红树属植物Rhizophora mangle 中, 目前仅此物种的花蕾、幼嫩及成熟的胚轴有转录组数据.Dassanayake等(2009)对R . mangle 不同阶段的营养器官、繁殖器官的转录组进行基因功能注释(gene ontology)分析, 在对所有表达的基因功能进行分类(分子功能、生物过程、细胞组分)比较后发现, “分子功能(molecular function)+生物过程(biological process)”共同表达的基因是“分子功能(molecular function)+细胞组分(cellular component)”共同表达的基因数量的两倍, 说明通过转录水平调控生理过程(而非改变细胞组分)有可能与红树植物胎生的生长发育过程存在密切关系.然而,由于采样阶段、基因组资源、信息分析、缺乏基因功能验证实验等的局限, 高达55%的序列没有功能注释, 而且未对花蕾、胚轴的发育进行深入分析, 此实验未能回答红树植物胎生发育的一系列问题. ...
红树胎生现象及其对潮间带生境适应性研究进展. 生态学报,
2
2004
... 长久以来胎生被认为是一个针对盐度适应的特征(Joshi, 1933 ), 但这一认识并没有被广泛接受, 因为其他的盐生植物并没有胎生现象.Wang等(2002)的研究发现秋茄树胚轴发育后期组织液离子浓度并没有明显变化, 推测胎生与抗盐性并没有直接的联系, 认为胎生并不是对某一个环境因子的适应, 而是对海岸潮间带多个生态因子综合响应的结果(闫中正等, 2004 ). ...
... 胎生意味着种子在母体上萌发, 直至生长为成熟的繁殖体, 是一个能量不断积累的过程(闫中正等, 2004 ).此时的繁殖体储存大量的碳水化合物, 并且因为表皮含叶绿素能进行光合作用, 能够为独立生长初期和漂流过程提供足够的营养供给(Lin & Sternberg, 1995 ; Smith & Snedaker, 2000 ). ...
胎生植物是怎样 “胎生” 的? 植物杂志,
1
1979
... 真胎生现象在4类植物中已有报道(Farnsworth, 1997 ).首先是在遗传上以胎生为繁殖方式的植物, 如分布在8个属的红树植物(表1 )及两个属的海草(Tomlinson, 1986 ), 它们在自然生长环境条件下以胎生为最主要的繁殖方式, 繁殖体在母体上突破种皮或果皮并生长; 二是生长于热带、亚热带的某些顽拗性种子植物, 如少数仙人掌科植物(Cota-Sánchez, 2004 ); 三是某些处于特殊环境中的植物, 如水稻(Oryza sativa )、玉米(Zea mays )等, 其种子在高温、高湿条件下提前萌发, 但它们不从母体中吸收营养(叶能干, 1979 ); 四是与休眠有关的植物激素脱落酸(ABA)合成或响应通路有关的突变体植株, 或因种子中休眠基因功能的缺失不能进入休眠期而在母体直接开始萌发(Robertson, 1955 ), 后两类植物由于受外界条件诱导而形成提前萌发的性状, 在遗传本质上不是真正意义上的胎生, 但对于红树植物胎生现象研究仍有一定的借鉴作用.表1 列出已发表的以胎生为繁殖方式的植物. ...
红树植物抗重金属污染研究进展. 云南植物研究,
1
2005
... 红树植物在抗盐、抗重金属污染等方面的逆境生理生态特征及其分子生态学机制已有报道(张凤琴等, 2005 ; Miyama & Hanagata, 2007 ; Wong et al ., 2007 ; 程皓等, 2009 ; Parida & Jha, 2010 ), 但对红树植物独特的繁殖方式——胎生的分子机制研究甚少.转录组研究基本集中在红树属植物Rhizophora mangle 中, 目前仅此物种的花蕾、幼嫩及成熟的胚轴有转录组数据.Dassanayake等(2009)对R . mangle 不同阶段的营养器官、繁殖器官的转录组进行基因功能注释(gene ontology)分析, 在对所有表达的基因功能进行分类(分子功能、生物过程、细胞组分)比较后发现, “分子功能(molecular function)+生物过程(biological process)”共同表达的基因是“分子功能(molecular function)+细胞组分(cellular component)”共同表达的基因数量的两倍, 说明通过转录水平调控生理过程(而非改变细胞组分)有可能与红树植物胎生的生长发育过程存在密切关系.然而,由于采样阶段、基因组资源、信息分析、缺乏基因功能验证实验等的局限, 高达55%的序列没有功能注释, 而且未对花蕾、胚轴的发育进行深入分析, 此实验未能回答红树植物胎生发育的一系列问题. ...
几种红树植物繁殖体发育和幼苗成长过程的生理生态学研究
2
2003
... 但漂流的远近与显胎生、非胎生繁殖方式并没有直接关联, 而是与果实大小、形状、密度紧密相关(Ryck et al ., 2012 ).胚轴密度与生境中海水密度是否接近导致不一样的漂流模式, 例如显胎生的木榄成熟胚轴密度接近海水密度, 适应随潮水传播, 而隐胎生的蜡烛果胚轴密度远大于生境海水, 仅适合短时间内漂流(张宜辉, 2003 ).个体较大的红茄苳繁殖体比起角果木来说传播得更远, 而根系萌发迅速的角果木在向陆带(landward)有定植优势(Robert et al ., 2015 ).这些不同的扩散模式可能最终决定了红树林的分带(Kadoya & Inoue, 2015 ).如上所述, 胎生种子在成熟过程中一直保持高水分含量.代谢水的丢失会对种子造成不可挽回的损伤, 但是在胎生繁殖体中这种损失并不存在, 能够最大限度地保持种子活力.胎生繁殖体(胚轴)作为具有相对完整结构的幼苗, 其定植明显比休眠的种子更加迅速而有效, 是海岸潮间带复杂环境的优势适应性特征. ...
... 胎生对适应高盐环境具有一定意义.在胚轴发育过程中细胞液的盐分浓度变化不明显(Wang et al ., 2002 ), 但是胚轴在母体上发育成熟, 最终达到与母树生境的离子浓度一致(张宜辉, 2003 ).繁殖体掉落在滩涂后迅速形成侧根, 并与外界进行离子交换, 以快速适应盐度变化.Joshi (1933)认为, 种子的胎生是为了避免受到高盐生境的迫害, 使其顺利发芽乃至定植在海岸潮间带.实际上, 在不同盐度培养条件下, 红树植物幼苗均表现出低盐促进、高盐抑制的特征.邱广龙(2005)发现低盐度有利于隐胎生白骨壤(即海榄雌)繁殖体的萌根与幼苗生长, 胎生和非胎生红树植物繁殖体同样偏好低盐环境(Jayatissa et al ., 2008 ; Robert et al ., 2015 ).然而, 有报道指出胎生繁殖体的萌发需要一定的盐度, 某些红树繁殖体在淡水中无法正常生长(Wang et al ., 2011 ). ...
红树植物桐花树生长发育过程的元素动态与抗盐适应性. 海洋科学,
1
2004
... 热值(caloric value)指单位质量干物质在完全燃烧后所释放出来的热量值, 可以反映出胎生繁殖体的能量水平(鲍雅静等, 2008 ).在繁殖体的发育过程中, 从花、果至成熟胚轴, 其热值逐渐降低, 且始终低于母体.这可能是由于生长发育过程中胚轴已经开始消耗能量, 以达到迅速突破果皮, 进而伸长的结果, 同时胚轴中低能量物质如纤维素等增加(林鹏等, 2000 ).此外, 灰分含量与热值变化趋势类似, 例如蜡烛果(Aegiceras corniculatum )胚轴在成熟过程中灰分含量降低, 但脱离母树后灰分含量逐步累积提高, 一些研究人员将灰分含量与盐离子联系起来, 认为蜡烛果胚轴脱离母树开始独立生长是一个大量吸收盐分的过程(林益明等, 2004 ; 赵胡等, 2004 ).虽然热值和灰分随着发育过程而降低, 但随着生物量的增加, 单个胚轴所含能量总体呈上升趋势, 为后续的传播和定植提供了足够的能耗支持. ...
红树胚轴和叶片生长发育的元素动态. 海洋学报,
1
1997
... 胎生红树植物胚胎发育的另一个重要的特征是发育过程中繁殖器官盐离子浓度的变化.对于胚胎如何适应落地后的高盐生境, 有两种不同的结论, 即胎生是低盐化过程, 以及与此对立的胎生是盐分累积过程.郑文教和林鹏(1997)测定了秋茄树、木榄(Bruguiera gymnorhiza )和海莲(Bruguiera sexangula )胚轴发育过程中的盐离子, 发现随着胚轴成熟, 其Mg2+ 、Ca2+ 、Na+ 、K+ 、Cl- 含量呈下降趋势, 胚轴的盐含量明显低于母树果皮、叶片的盐含量, 且果皮、叶片的组织液Cl- 浓度与海水Cl- 浓度接近, 故认为胎生过程是一个低盐化的过程, 后代是通过与母体的长期接触来获得抗盐锻炼, 进而适应落地后的环境(郑文教等, 1992 ; 郑文教和林鹏, 1997 ).红树植物很可能起源于淡水环境, 胚胎发育是其生活史的重演, 这种胚胎发育低盐化的过程很可能是一种“返祖”现象.有趣的是, Joshi等(1972)发现显胎生植物胚轴发育过程中Mg2+ 、Ca2+ 、Na+ 、K+ 含量也逐渐下降, 但Cl- 含量逐渐增加.然而, 秋茄树成熟胚轴的Mg2+ 、Ca2+ 、Na+ 、K+ 、Cl- 含量与早期胚轴相比呈倍性增加(Wang et al ., 2002 ).部分研究者认为胎生繁殖体的盐抗性是逐渐适应从母体输送到幼苗的盐分而获得的(Lin, 1988 ).这两种不同的结论可能是由于对胎生发育过程阶段的划分不同及盐离子含量、浓度的高低定义不同, 甚至是物种不同而导致的.相比于胎生红树植物, 非胎生红树植物繁殖体发育过程中盐离子动态的研究相对较少, 但有证据表明非胎生的老鼠簕(Acanthus ilicifolius )幼苗Cl- 含量高于胎生物种Ceriops candolleana 、Avicennia officinalis 、Aegiceras majus (Joshi et al ., 1972 ), 其机制仍有待阐明.Mg2+ 、Ca2 、Na+ 、K+ 和Cl- 这5种离子是细胞渗透势的主要成分, 除了构成盐分浓度外, 还可能作为一种信号在种子成熟过程中起其他作用, 如Ca2+ 作为二级信使传导信号调节基因表达. ...
红树胎生胚轴发育和叶片发育的 C1量动态. 厦门大学学报(自然科学版),
1
1992
... 胎生红树植物胚胎发育的另一个重要的特征是发育过程中繁殖器官盐离子浓度的变化.对于胚胎如何适应落地后的高盐生境, 有两种不同的结论, 即胎生是低盐化过程, 以及与此对立的胎生是盐分累积过程.郑文教和林鹏(1997)测定了秋茄树、木榄(Bruguiera gymnorhiza )和海莲(Bruguiera sexangula )胚轴发育过程中的盐离子, 发现随着胚轴成熟, 其Mg2+ 、Ca2+ 、Na+ 、K+ 、Cl- 含量呈下降趋势, 胚轴的盐含量明显低于母树果皮、叶片的盐含量, 且果皮、叶片的组织液Cl- 浓度与海水Cl- 浓度接近, 故认为胎生过程是一个低盐化的过程, 后代是通过与母体的长期接触来获得抗盐锻炼, 进而适应落地后的环境(郑文教等, 1992 ; 郑文教和林鹏, 1997 ).红树植物很可能起源于淡水环境, 胚胎发育是其生活史的重演, 这种胚胎发育低盐化的过程很可能是一种“返祖”现象.有趣的是, Joshi等(1972)发现显胎生植物胚轴发育过程中Mg2+ 、Ca2+ 、Na+ 、K+ 含量也逐渐下降, 但Cl- 含量逐渐增加.然而, 秋茄树成熟胚轴的Mg2+ 、Ca2+ 、Na+ 、K+ 、Cl- 含量与早期胚轴相比呈倍性增加(Wang et al ., 2002 ).部分研究者认为胎生繁殖体的盐抗性是逐渐适应从母体输送到幼苗的盐分而获得的(Lin, 1988 ).这两种不同的结论可能是由于对胎生发育过程阶段的划分不同及盐离子含量、浓度的高低定义不同, 甚至是物种不同而导致的.相比于胎生红树植物, 非胎生红树植物繁殖体发育过程中盐离子动态的研究相对较少, 但有证据表明非胎生的老鼠簕(Acanthus ilicifolius )幼苗Cl- 含量高于胎生物种Ceriops candolleana 、Avicennia officinalis 、Aegiceras majus (Joshi et al ., 1972 ), 其机制仍有待阐明.Mg2+ 、Ca2 、Na+ 、K+ 和Cl- 这5种离子是细胞渗透势的主要成分, 除了构成盐分浓度外, 还可能作为一种信号在种子成熟过程中起其他作用, 如Ca2+ 作为二级信使传导信号调节基因表达. ...


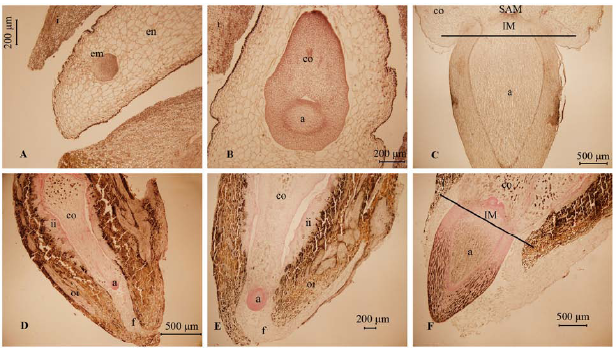

{kind=link}
{kind=link}
{kind=link}
{kind=link}