The morphology of fine root branching of woody plants is highly variable in their forms and functions. In the past two decades, researchers have increasingly recognized that the root-diameter-based method, using an arbitrary size of root diameter, failed to precisely characterize the physiological and ecological processes involved in finest roots. The number of publications using root-order-based approaches has increased regardless the fact that root trait-measurements based on root order are time-consuming and labor-intensive. A new approach—root functional classification method—was proposed and had been applied in the literature. The functional classification of fine roots separates roots of < 2 mm to absorptive and transport pools, making it more feasible for studies on root biomass and turnover. This new concept redefines fine root guild and has great potentials for future studies. Our literature review of the topic indicates that less is known about the inter-specific differences in estimates of biomass of absorptive and/or transport roots, with a large variation of absorptive roots on global scale. In addition, our review emphasizes the importance in: a) precision estimating of the absorptive biomass of fine roots, and b) proper definition of the range of the transport roots within and among forest ecosystems. Finally, after compare the strengths and weaknesses of the functional classification method, we propose several specific suggestions to improve the applications of this approach.
Keywords:absorptive root
;
transport root
;
fine root
;
root order
;
root turnover
GUJia-Cun, WANGDong-Nan, XIAXiu-Xue, WANGShao-Zhong. Applications of functional classification methods for tree fine root biomass estimation: Advancements and synthesis. Chinese Journal of Plant Ecology, 2016, 40(12): 1344-1351 https://doi.org/10.17521/cjpe.2016.0167
b). Endogenous and exogenous controls of root life span, mortality and nitrogen flux in a longleaf pine forest: Root branch order predominates
.Journal of Ecology ,96, 737-745.
[19]
GuoDL, XiaMX, WeiX, ChangWJ, LiuY, WangZQ (2008
c). Anatomical traits associated with absorption and mycorrhizal colonization are linked to root branch order in twenty-three Chinese temperate tree species
.New Phytologist, 180, 673-683.
[20]
HeJS, WangZQ, FangJY (2004).
Issues and prospects of belowground ecology with special reference to global climate change
Fine root biomass and dynamics in beech forests across a precipitation gradient—Is optimal resource partitioning theory applicable to water-limited mature trees?
Journal of Ecology, 101, 1183-1200.
[23]
JacksonRB, MooneyHA, SchulzeED (1997).
A global budget for fine root biomass, surface area, and nutrient contents
.Proceedings of the National Academy of Sciences of the United States of America, 94, 7362-7366.
Acquisition of ephemeral module in roots: A new view and test
.Scientific Reports, 4, 1-4.
[27]
KouL, GuoDL, YangH, GaoWL, LiSG (2015).
Growth, morphological traits and mycorrhizal colonization of fine roots respond differently to nitrogen addition in a slash pine plantation in subtropical China
Complementarity in nutrient foraging strategies of absorptive fine roots and arbuscular mycorrhizal fungi across 14 coexisting subtropical tree species
Effects of thinning on fine root biomass and carbon storage of subalpine
Picea asperata plantation in Western Sichuan Province, China. Chinese Journal of Plant Ecology, 36, 645-654.(in Chinese with English abstract) [刘运科, 范川, 李贤伟, 凌银花, 周义贵, 冯茂松, 黄从德 (2012).
间伐对川西亚高山粗枝云杉人工林细根生物量及碳储量的影响
. 植物生态学报, 36, 645-654.]
[33]
LongYQ, KongDL, ChenZX, ZengH (2013).
Variation of the linkage of root function with root branch order
.PLOS ONE ,8, e57153. doi: 10.1371/journal.pone.0057153.
Aggregated and comple- mentary: Symmetric proliferation, overyielding, and mass effects explain fine-root biomass in soil patches in a diverse temperate deciduous forest landscape
Fine root biomass, production, turnover rates, and nutrient contents in boreal forest ecosystems in relation to species, climate, fertility, and stand age: Literature review and meta-analyses
Scots pine fine roots adjust along a 2000-km latitudinal climatic gradient
.New Phytologist ,212, 389-399.
A trade-off between scale and precision in resource foraging
1
1991
... 最后, 本文主要关注了采用功能划分方法开展细根生物量与周转研究的进展, 但是功能划分方法在其他研究领域中的应用亟待开展.例如, 细根分解是树木向土壤中输入养分和有机质的重要途径(Silver & Miya, 2001; Freschet et al., 2013), 但是目前采用功能划分方法进行的相关研究较少(Fan & Guo, 2010; Goebel et al., 2011); 根系觅食策略(foraging strategy)及其对土壤资源异质性的响应, 也是长期以来受关注的研究问题(Campbell et al., 1991; McNickle et al., 2009; Liu et al., 2015), 吸收根概念的引入将有助于准确地理解根系功能属性与养分的互作机制.因此, 在根系生物量分配、生产和周转相关的研究中, 都有必要考虑功能划分方法的适用性. ...
Intrinsic and extrinsic controls of fine root life span
1
2013
... 其次, 因为吸收根周转率高, 准确估计吸收根的生物量是构建森林生态系统物质循环模型的重要基础.采用微根管法(minirhizotron)观测的研究证实, 树木吸收根的中值寿命短则几周, 长则数月, 而最长也不超过2.5年(周转率至少为1年0.4次)(With- ington et al., 2006; McCormack et al., 2012; Liu et al., 2016).然而, 通过同位素或模型模拟等方法估计的运输根(在一些研究中称为“长寿命根”)的周转率远低于吸收根, 大部分为1年0.1-0.2次(Matamala et al., 2003; Lynch et al., 2013).更重要的是, 当排除研究方法上的差异时, Xia等(2010)采用根窗法对吸收根和运输根的研究证实, 水曲柳吸收根(1-3级根)的中值寿命为510-692 d (死亡率> 90%), 而运输根(4-5级根)的死亡率<10%; Liu等(2016)通过根窗法对我国新疆5种树木细根寿命的研究发现, 前3级根中值寿命为95-154 d, 而4-5级根均超过了550 d.因此, 吸收根是树木细根中最为活跃的组分, 在C和养分循环中发挥着重要的作用.由此可见, McCormack等(2015)曾假定吸收根周转率是运输根的5至10倍, 在很大程度上是合理的.基于这样的假定, 只有运输根的生物量现存量是吸收根的5-10倍时, 二者对细根生物量周转的贡献才相等.但是, 现有实测吸收根与运输根生物量的研究中, 运输根与吸收根的比例变化范围仅为0.80 (水曲柳, Wang et al., 2006)至1.85 (粗枝云杉, 刘运科等 2012), 远小于可能出现的理论值上限(如5-10).鉴于以往通过微根管法获得了较多树木吸收根寿命(或周转率)的数据(如Withington et al., 2006; Espeleta et al., 2009; McCormack et al., 2012; Chen & Brassard, 2013), 吸收根生物量的准确量化成为评价细根周转对森林地下生态学过程调控作用的关键. ...
Root morphology, anatomy and chemistry of nine fern species (pteridophyta) in a temperate forest
1
2015
... 其次, 在研究吸收根生物量和其他功能属性时, 一定要对不同树种的吸收根范围进行定义和检验(卫星等, 2008; McCormack et al., 2015), 这应该作为一个标准的研究环节而受到重视.可以先通过根解剖属性的测定判断哪些类群具有明显的吸收功能, 因为对于不同树种而言, 具有吸收能力的根系所包括的根序范围可能是不同的(Long et al., 2013).例如, Valenzuela-Estrada等(2008)对根直径很细的高丛蓝莓(Vaccinium corymbosum)研究发现, 前3级根均未出现次生生长, 且明显受到真菌侵染; Long等(2013)研究证实, 根直径较粗的合果木(Paramichelia baillonii)中1-5级根均存在完整的皮层组织并具有菌根侵染特征.但是Guo等(2008c)研究的23个树种前3级根中均存在皮层组织, 且受树种吸收根直径大小的影响不明显.因此, 树种吸收根直径大小可能并不是影响吸收根界定范围(包括几级根序)的重要因子, 表明树种水平上根系解剖学检验是必要的, 尤其是当研究中包括其他类群植物(如草本植物)时更应如此(Dong et al., 2015).值得注意的是, 目前并不清楚树木吸收根类群是否因地理区域的不同而发生明显的变化, 例如对一些广布种(如栎类), 是否在不同的环境梯度下包括了相同的低级根类群.因此, 即使对于同一物种, 也有必要在地理位置相距较远的不同地点上检验其吸收根所包括的根序范围(Zadworny et al., 2016), 并以此为基础研究根生物量和功能属性的变化. ...
Linking root traits to nutrient foraging in arbuscular mycorrhizal trees in a temperate forest
2
2015
... 再次, 由于吸收根实测数据的缺乏, 吸收根生物量在区域和全球尺度上的变异和影响机制还有待进一步研究.以往对细根(如直径≤2 mm)生物量现存量和生产量变异所做的较多综述性研究(如Vogt et al., 1996; Jackson et al., 1997; Finér et al., 2007, 2011a, 2011b; Yuan & Chen, 2010), 为我们提供了很好的启示.早期的研究显示, 在不同的生物区系、气候区或林型间, 细根生物量变异很大, 但是并没有表现出有规律的变化(如Vogt et al., 1996; Jackson et al., 1997).近年来的研究显示, 气候因子或林分特征能够较好地解释细根生物量的变异.在区域尺度上, Yuan和Chen (2010)在分析北方森林(boreal forest)细根生物量的格局时指出, 细根生物量与年降水量和温度之间显著正相关(r = 0.4, p < 0.001); Meier和Leuschner (2008)对欧洲中部沙质土壤样带上14个山毛榉(Fagus sylvatica)林分的研究也发现, 细根(直径<2 mm)生物量随年降水量的增加而增加, 但是Hertel等(2013)的结果是在砂岩土壤样带上山毛榉细根生物量随着降水量增加而降低.在全球尺度上, 尽管气候因子无法很好地解释细根生物量变异, 但是林分平均基面积(basal area)和林分年龄分别能够解释林分水平细根生物量30%和单株水平细根生物量45%的变异(Finér et al., 2011a).与以上研究相比, 目前还无法获得全球尺度上吸收根生物量的实测数据, 但是仍有少量针对单个树种的根尖(或共生真菌侵染的1-2级根)生物量大尺度空间变异的研究(Helmisaari et al., 2009; Ostonen et al., 2011).由于根尖(或菌根)在根系资源吸收中的重要作用(如Guo et al., 2008c), 这些研究结果可以在一定程度上帮助我们了解吸收根生物量大尺度的变异格局.Ostonen等(2011)对欧洲12个挪威云杉(Picea abies)林分(纬度相差20°)的研究证实, 林分水平的菌根(大部分属于1、2级根)生物量在高纬度地区比低纬度地区大, 而且能够被N沉降量、年平均气温和土壤C:N比较好地解释; 但是Helmisaari等(2009)对相对较小的空间范围(纬度相差9°)的研究显示, 芬兰南部挪威云杉和欧洲赤松(Pinus sylvestris)根尖生物量与其芬兰北部生物量均没有显著的差别.最近,Zadworny等(2016)研究了北欧地区2000 km的梯度(纬度相差19°)上欧洲赤松吸收根比例的变异规律,发现2-5级根中具有吸收功能个体根的比例均随着年平均温度升高而降低(1级根全部是吸收根),证明寒冷地区树木分配了更多的根系生物量用于养分和水分的吸收.由于气候、土壤和人为干扰等因子在大尺度空间上存在着显著的变异性, 而且不同树种根系生长对环境变异的响应也存在差异(Eissenstat et al., 2015), 了解吸收根生物量在区域或全球尺度的变异规律和影响因素, 仍需在更广泛的地理区域上开展林分尺度上的研究. ...
... 首先, 采用根系功能划分的方法在获取吸收根和运输根生物量上展现出了很大的优势(例如Kong & Ma, 2014), 而且这一方法也可以应用到估计不同类群根系形态、组织化学和寿命与周转的研究中(Eissenstat et al., 2015; Liu et al., 2015).但是, 在测定某些根系功能属性时采用根序划分的方法可能是必要的, 例如根系解剖特征(如Guo et al., 2008c; Gu et al., 2014; Kong et al., 2014), 这是因为将不同根序解剖特征的数据混合并取平均数, 没有实质上的生物学意义, 而且可能引入新的测量误差. ...
Tree species fine-root demography parallels habitat specialization across a sandhill soil resource gradient
1
2009
... 其次, 因为吸收根周转率高, 准确估计吸收根的生物量是构建森林生态系统物质循环模型的重要基础.采用微根管法(minirhizotron)观测的研究证实, 树木吸收根的中值寿命短则几周, 长则数月, 而最长也不超过2.5年(周转率至少为1年0.4次)(With- ington et al., 2006; McCormack et al., 2012; Liu et al., 2016).然而, 通过同位素或模型模拟等方法估计的运输根(在一些研究中称为“长寿命根”)的周转率远低于吸收根, 大部分为1年0.1-0.2次(Matamala et al., 2003; Lynch et al., 2013).更重要的是, 当排除研究方法上的差异时, Xia等(2010)采用根窗法对吸收根和运输根的研究证实, 水曲柳吸收根(1-3级根)的中值寿命为510-692 d (死亡率> 90%), 而运输根(4-5级根)的死亡率<10%; Liu等(2016)通过根窗法对我国新疆5种树木细根寿命的研究发现, 前3级根中值寿命为95-154 d, 而4-5级根均超过了550 d.因此, 吸收根是树木细根中最为活跃的组分, 在C和养分循环中发挥着重要的作用.由此可见, McCormack等(2015)曾假定吸收根周转率是运输根的5至10倍, 在很大程度上是合理的.基于这样的假定, 只有运输根的生物量现存量是吸收根的5-10倍时, 二者对细根生物量周转的贡献才相等.但是, 现有实测吸收根与运输根生物量的研究中, 运输根与吸收根的比例变化范围仅为0.80 (水曲柳, Wang et al., 2006)至1.85 (粗枝云杉, 刘运科等 2012), 远小于可能出现的理论值上限(如5-10).鉴于以往通过微根管法获得了较多树木吸收根寿命(或周转率)的数据(如Withington et al., 2006; Espeleta et al., 2009; McCormack et al., 2012; Chen & Brassard, 2013), 吸收根生物量的准确量化成为评价细根周转对森林地下生态学过程调控作用的关键. ...
Slow decomposition of lower order roots: A key mechanism of root carbon and nutrient retention in the soil
1
2010
... 最后, 本文主要关注了采用功能划分方法开展细根生物量与周转研究的进展, 但是功能划分方法在其他研究领域中的应用亟待开展.例如, 细根分解是树木向土壤中输入养分和有机质的重要途径(Silver & Miya, 2001; Freschet et al., 2013), 但是目前采用功能划分方法进行的相关研究较少(Fan & Guo, 2010; Goebel et al., 2011); 根系觅食策略(foraging strategy)及其对土壤资源异质性的响应, 也是长期以来受关注的研究问题(Campbell et al., 1991; McNickle et al., 2009; Liu et al., 2015), 吸收根概念的引入将有助于准确地理解根系功能属性与养分的互作机制.因此, 在根系生物量分配、生产和周转相关的研究中, 都有必要考虑功能划分方法的适用性. ...
Variation in fine root biomass of three European tree species: Beech (
1
2007
... 再次, 由于吸收根实测数据的缺乏, 吸收根生物量在区域和全球尺度上的变异和影响机制还有待进一步研究.以往对细根(如直径≤2 mm)生物量现存量和生产量变异所做的较多综述性研究(如Vogt et al., 1996; Jackson et al., 1997; Finér et al., 2007, 2011a, 2011b; Yuan & Chen, 2010), 为我们提供了很好的启示.早期的研究显示, 在不同的生物区系、气候区或林型间, 细根生物量变异很大, 但是并没有表现出有规律的变化(如Vogt et al., 1996; Jackson et al., 1997).近年来的研究显示, 气候因子或林分特征能够较好地解释细根生物量的变异.在区域尺度上, Yuan和Chen (2010)在分析北方森林(boreal forest)细根生物量的格局时指出, 细根生物量与年降水量和温度之间显著正相关(r = 0.4, p < 0.001); Meier和Leuschner (2008)对欧洲中部沙质土壤样带上14个山毛榉(Fagus sylvatica)林分的研究也发现, 细根(直径<2 mm)生物量随年降水量的增加而增加, 但是Hertel等(2013)的结果是在砂岩土壤样带上山毛榉细根生物量随着降水量增加而降低.在全球尺度上, 尽管气候因子无法很好地解释细根生物量变异, 但是林分平均基面积(basal area)和林分年龄分别能够解释林分水平细根生物量30%和单株水平细根生物量45%的变异(Finér et al., 2011a).与以上研究相比, 目前还无法获得全球尺度上吸收根生物量的实测数据, 但是仍有少量针对单个树种的根尖(或共生真菌侵染的1-2级根)生物量大尺度空间变异的研究(Helmisaari et al., 2009; Ostonen et al., 2011).由于根尖(或菌根)在根系资源吸收中的重要作用(如Guo et al., 2008c), 这些研究结果可以在一定程度上帮助我们了解吸收根生物量大尺度的变异格局.Ostonen等(2011)对欧洲12个挪威云杉(Picea abies)林分(纬度相差20°)的研究证实, 林分水平的菌根(大部分属于1、2级根)生物量在高纬度地区比低纬度地区大, 而且能够被N沉降量、年平均气温和土壤C:N比较好地解释; 但是Helmisaari等(2009)对相对较小的空间范围(纬度相差9°)的研究显示, 芬兰南部挪威云杉和欧洲赤松(Pinus sylvestris)根尖生物量与其芬兰北部生物量均没有显著的差别.最近,Zadworny等(2016)研究了北欧地区2000 km的梯度(纬度相差19°)上欧洲赤松吸收根比例的变异规律,发现2-5级根中具有吸收功能个体根的比例均随着年平均温度升高而降低(1级根全部是吸收根),证明寒冷地区树木分配了更多的根系生物量用于养分和水分的吸收.由于气候、土壤和人为干扰等因子在大尺度空间上存在着显著的变异性, 而且不同树种根系生长对环境变异的响应也存在差异(Eissenstat et al., 2015), 了解吸收根生物量在区域或全球尺度的变异规律和影响因素, 仍需在更广泛的地理区域上开展林分尺度上的研究. ...
a). Factors causing variation in fine root biomass in forest ecosystems
2011
b). Fine root production and turnover in forest ecosystems in relation to stand and environmental characteristics
2011
Linking litter decomposi- tion of above- and below-ground organs to plant-soil feed- backs worldwide
1
2013
... 最后, 本文主要关注了采用功能划分方法开展细根生物量与周转研究的进展, 但是功能划分方法在其他研究领域中的应用亟待开展.例如, 细根分解是树木向土壤中输入养分和有机质的重要途径(Silver & Miya, 2001; Freschet et al., 2013), 但是目前采用功能划分方法进行的相关研究较少(Fan & Guo, 2010; Goebel et al., 2011); 根系觅食策略(foraging strategy)及其对土壤资源异质性的响应, 也是长期以来受关注的研究问题(Campbell et al., 1991; McNickle et al., 2009; Liu et al., 2015), 吸收根概念的引入将有助于准确地理解根系功能属性与养分的互作机制.因此, 在根系生物量分配、生产和周转相关的研究中, 都有必要考虑功能划分方法的适用性. ...
Measuring and modeling the spectrum of fine-root turnover times in three forests using isotopes, minirhizotrons, and the Radix model
1
2010
... 综合以往研究证据可见, 传统的细根由两类功能不同的根群组成, 即吸收根群(absorptive fine roots)和运输根群(transport fine roots) (Joslin et al., 2006; McCormack et al., 2015).前者由低级根组成, 是资源吸收的主体, 寿命短而周转快; 后者由高级根组成, 主要负责运输与存储, 寿命长而周转慢(图1).从物质循环的角度看, 将传统的细根按照根序把细根分为不同的库的方法(order-based method), 或采用功能划分的方法(functional-classification method)区分为吸收根和运输根2个库, 可以更准确地评价细根在生态系统中的重要作用(Guo et al., 2008b; Gaudinski et al., 2010; Parton et al., 2010; McCormack et al., 2015).然而, 将细根区分成单个根序进行研究, 如生物量估计, 需要消耗大量的时间和人力; 比较之下, 功能划分的方法既能减少工作量, 又能有效地进行不同处理间或树种间根系生物量及其他根系过程(如根呼吸和周转)的比较, 在根系研究的准确性和可行性上达到了更好的平衡(McCormack et al., 2015).因此, 采用功能划分方法能充分刻化树木根系统内在结构功能的异质性, 从而更加客观地评价根系主导的生态学过程及其影响.McCormack等(2015)最近基于功能划分的方法, 将传统的细根(直径≤2 mm)区分为吸收根和运输根2个库, 并估算相应的生物量和周转率, 结果显示细根周转消耗陆地生态系统NPP的比例为22%, 明显低于以往将细根按照直径阈值区分为1个库估算的结果(约33%, Jackson et al., 1997).然而, 与传统直径阈值的研究相比, 基于功能划分方法进行的根系研究是非常缺乏的. ...
Global patterns of root turnover for terrestrial ecosystems
Decomposi- tion of the finest root branching orders: Linking carbon and nutrient dynamics belowground to fine root function and structure
1
2011
... 最后, 本文主要关注了采用功能划分方法开展细根生物量与周转研究的进展, 但是功能划分方法在其他研究领域中的应用亟待开展.例如, 细根分解是树木向土壤中输入养分和有机质的重要途径(Silver & Miya, 2001; Freschet et al., 2013), 但是目前采用功能划分方法进行的相关研究较少(Fan & Guo, 2010; Goebel et al., 2011); 根系觅食策略(foraging strategy)及其对土壤资源异质性的响应, 也是长期以来受关注的研究问题(Campbell et al., 1991; McNickle et al., 2009; Liu et al., 2015), 吸收根概念的引入将有助于准确地理解根系功能属性与养分的互作机制.因此, 在根系生物量分配、生产和周转相关的研究中, 都有必要考虑功能划分方法的适用性. ...
Influence of root structure on fine root survivorship: An analysis of 18 tree species using a minirhizotron method
1
2011
... 为了准确估计细根在生态系统物质分配和养分循环上的作用, 研究者需要知道细根的生物量现存量(单位面积上的干质量)和周转率(Norby & Jackson 2000; Majdi et al., 2005; Guo et al., 2008b).因此, 如果采用功能划分的方法进行根系生物量和周转的估计, 需要分别获得吸收根和运输根对应的参数(McCormack et al., 2015).然而, 由于概念理解和方法学上的限制, 相应的根系参数非常缺乏.在McCormack等(2015)的研究中, 是在以往Jackson等(1997)基于直径阈值方法的基础上, 将直径≤2 mm的生物量按照以往研究中吸收根和运输根的生物量比例(木本植物为10:90、33:67和60:40等3种不同比例), 并假定吸收根周转率为1年周转1次, 而运输根周转率为1年0.1次和0.2次两种情境, 来计算细根周转消耗的NPP.McCormack等(2015)的研究结果显示了采用功能划分方法来研究细根生物量和相关生态学过程的重要性.然而, 将根系结构与其功能, 特别是与生物量和周转联系起来, 只是在最近10余年才得到应有的重视(Wells et al., 2002; Guo et al., 2008c; Strand et al., 2008; Gu et al., 2011), 如今, 我们对吸收根和运输根生物量的变异规律、潜在机制的认识和理解并不全面.为此, 我们对近年来国内外采用根系功能划分方法开展根系生物量的研究进行了总结, 以期发现当前研究中存在的不足和亟待解决的问题, 在应用该方法时值得注意的问题, 以促进我国树木根系生态学的进一步发展. ...
Root diameter variations explained by anatomy and phylogeny of 50 tropical and temperate tree species
1
2014
... 首先, 采用根系功能划分的方法在获取吸收根和运输根生物量上展现出了很大的优势(例如Kong & Ma, 2014), 而且这一方法也可以应用到估计不同类群根系形态、组织化学和寿命与周转的研究中(Eissenstat et al., 2015; Liu et al., 2015).但是, 在测定某些根系功能属性时采用根序划分的方法可能是必要的, 例如根系解剖特征(如Guo et al., 2008c; Gu et al., 2014; Kong et al., 2014), 这是因为将不同根序解剖特征的数据混合并取平均数, 没有实质上的生物学意义, 而且可能引入新的测量误差. ...
a). Fine root heterogeneity by branch order: Exploring the discrepancy in root turnover estimates between minirhizotron and carbon isotopic methods
2008
Fine root branch orders respond differentially to carbon source-sink manipulations in a longleaf pine forest
2004
b). Endogenous and exogenous controls of root life span, mortality and nitrogen flux in a longleaf pine forest: Root branch order predominates
2008
c). Anatomical traits associated with absorption and mycorrhizal colonization are linked to root branch order in twenty-three Chinese temperate tree species
2008
Issues and prospects of belowground ecology with special reference to global climate change
Ectomycorrhizal root tips in relation to site and stand characteristics in Norway spruce and Scots pine stands in boreal forests
1
2009
... 再次, 由于吸收根实测数据的缺乏, 吸收根生物量在区域和全球尺度上的变异和影响机制还有待进一步研究.以往对细根(如直径≤2 mm)生物量现存量和生产量变异所做的较多综述性研究(如Vogt et al., 1996; Jackson et al., 1997; Finér et al., 2007, 2011a, 2011b; Yuan & Chen, 2010), 为我们提供了很好的启示.早期的研究显示, 在不同的生物区系、气候区或林型间, 细根生物量变异很大, 但是并没有表现出有规律的变化(如Vogt et al., 1996; Jackson et al., 1997).近年来的研究显示, 气候因子或林分特征能够较好地解释细根生物量的变异.在区域尺度上, Yuan和Chen (2010)在分析北方森林(boreal forest)细根生物量的格局时指出, 细根生物量与年降水量和温度之间显著正相关(r = 0.4, p < 0.001); Meier和Leuschner (2008)对欧洲中部沙质土壤样带上14个山毛榉(Fagus sylvatica)林分的研究也发现, 细根(直径<2 mm)生物量随年降水量的增加而增加, 但是Hertel等(2013)的结果是在砂岩土壤样带上山毛榉细根生物量随着降水量增加而降低.在全球尺度上, 尽管气候因子无法很好地解释细根生物量变异, 但是林分平均基面积(basal area)和林分年龄分别能够解释林分水平细根生物量30%和单株水平细根生物量45%的变异(Finér et al., 2011a).与以上研究相比, 目前还无法获得全球尺度上吸收根生物量的实测数据, 但是仍有少量针对单个树种的根尖(或共生真菌侵染的1-2级根)生物量大尺度空间变异的研究(Helmisaari et al., 2009; Ostonen et al., 2011).由于根尖(或菌根)在根系资源吸收中的重要作用(如Guo et al., 2008c), 这些研究结果可以在一定程度上帮助我们了解吸收根生物量大尺度的变异格局.Ostonen等(2011)对欧洲12个挪威云杉(Picea abies)林分(纬度相差20°)的研究证实, 林分水平的菌根(大部分属于1、2级根)生物量在高纬度地区比低纬度地区大, 而且能够被N沉降量、年平均气温和土壤C:N比较好地解释; 但是Helmisaari等(2009)对相对较小的空间范围(纬度相差9°)的研究显示, 芬兰南部挪威云杉和欧洲赤松(Pinus sylvestris)根尖生物量与其芬兰北部生物量均没有显著的差别.最近,Zadworny等(2016)研究了北欧地区2000 km的梯度(纬度相差19°)上欧洲赤松吸收根比例的变异规律,发现2-5级根中具有吸收功能个体根的比例均随着年平均温度升高而降低(1级根全部是吸收根),证明寒冷地区树木分配了更多的根系生物量用于养分和水分的吸收.由于气候、土壤和人为干扰等因子在大尺度空间上存在着显著的变异性, 而且不同树种根系生长对环境变异的响应也存在差异(Eissenstat et al., 2015), 了解吸收根生物量在区域或全球尺度的变异规律和影响因素, 仍需在更广泛的地理区域上开展林分尺度上的研究. ...
Fine root biomass and dynamics in beech forests across a precipitation gradient—Is optimal resource partitioning theory applicable to water-limited mature trees?
2013
A global budget for fine root biomass, surface area, and nutrient contents
3
1997
... 细根一般指直径≤2 mm的根系组分, 是树木吸收养分和水分的重要器官.由于细根不断地进行生产、衰老、死亡和再生长过程(即周转), 在陆地生态系统碳(C)和养分循环中发挥着重要作用(Norby & Jackson, 2000; Strand et al., 2008).据估计, 全球尺度上, 如果细根每年周转一次, 将会消耗陆地生态系统净初级生产力(NPP)的33% (Jackson et al., 1997).以往大多数研究者将直径小于某一阈值(如1或2 mm)的个体根作为一个结构和功能一致的整体来研究, 然而, 近年来的研究证据显示, 以往采用直径大小对细根进行划分的方法, 很大程度上忽视了个体根在根系统上所处位置的不同, 即根序(branch order)的差异.不同根序个体根在解剖、形态和生理功能上存在明显的差异, 因此采用直径阈值来划分根系组分的方法缺乏对根系功能的充分考虑(虽然假定了根系功能与直径大小有关), 这会严重地阻碍我们对细根结构与功能的准确理解(Pregitzer et al., 2002; Guo et al., 2008a; Valenzuela- Estrada et al., 2008; McCormack et al., 2015). ...
... 再次, 由于吸收根实测数据的缺乏, 吸收根生物量在区域和全球尺度上的变异和影响机制还有待进一步研究.以往对细根(如直径≤2 mm)生物量现存量和生产量变异所做的较多综述性研究(如Vogt et al., 1996; Jackson et al., 1997; Finér et al., 2007, 2011a, 2011b; Yuan & Chen, 2010), 为我们提供了很好的启示.早期的研究显示, 在不同的生物区系、气候区或林型间, 细根生物量变异很大, 但是并没有表现出有规律的变化(如Vogt et al., 1996; Jackson et al., 1997).近年来的研究显示, 气候因子或林分特征能够较好地解释细根生物量的变异.在区域尺度上, Yuan和Chen (2010)在分析北方森林(boreal forest)细根生物量的格局时指出, 细根生物量与年降水量和温度之间显著正相关(r = 0.4, p < 0.001); Meier和Leuschner (2008)对欧洲中部沙质土壤样带上14个山毛榉(Fagus sylvatica)林分的研究也发现, 细根(直径<2 mm)生物量随年降水量的增加而增加, 但是Hertel等(2013)的结果是在砂岩土壤样带上山毛榉细根生物量随着降水量增加而降低.在全球尺度上, 尽管气候因子无法很好地解释细根生物量变异, 但是林分平均基面积(basal area)和林分年龄分别能够解释林分水平细根生物量30%和单株水平细根生物量45%的变异(Finér et al., 2011a).与以上研究相比, 目前还无法获得全球尺度上吸收根生物量的实测数据, 但是仍有少量针对单个树种的根尖(或共生真菌侵染的1-2级根)生物量大尺度空间变异的研究(Helmisaari et al., 2009; Ostonen et al., 2011).由于根尖(或菌根)在根系资源吸收中的重要作用(如Guo et al., 2008c), 这些研究结果可以在一定程度上帮助我们了解吸收根生物量大尺度的变异格局.Ostonen等(2011)对欧洲12个挪威云杉(Picea abies)林分(纬度相差20°)的研究证实, 林分水平的菌根(大部分属于1、2级根)生物量在高纬度地区比低纬度地区大, 而且能够被N沉降量、年平均气温和土壤C:N比较好地解释; 但是Helmisaari等(2009)对相对较小的空间范围(纬度相差9°)的研究显示, 芬兰南部挪威云杉和欧洲赤松(Pinus sylvestris)根尖生物量与其芬兰北部生物量均没有显著的差别.最近,Zadworny等(2016)研究了北欧地区2000 km的梯度(纬度相差19°)上欧洲赤松吸收根比例的变异规律,发现2-5级根中具有吸收功能个体根的比例均随着年平均温度升高而降低(1级根全部是吸收根),证明寒冷地区树木分配了更多的根系生物量用于养分和水分的吸收.由于气候、土壤和人为干扰等因子在大尺度空间上存在着显著的变异性, 而且不同树种根系生长对环境变异的响应也存在差异(Eissenstat et al., 2015), 了解吸收根生物量在区域或全球尺度的变异规律和影响因素, 仍需在更广泛的地理区域上开展林分尺度上的研究. ...
... ; Jackson et al., 1997).近年来的研究显示, 气候因子或林分特征能够较好地解释细根生物量的变异.在区域尺度上, Yuan和Chen (2010)在分析北方森林(boreal forest)细根生物量的格局时指出, 细根生物量与年降水量和温度之间显著正相关(r = 0.4, p < 0.001); Meier和Leuschner (2008)对欧洲中部沙质土壤样带上14个山毛榉(Fagus sylvatica)林分的研究也发现, 细根(直径<2 mm)生物量随年降水量的增加而增加, 但是Hertel等(2013)的结果是在砂岩土壤样带上山毛榉细根生物量随着降水量增加而降低.在全球尺度上, 尽管气候因子无法很好地解释细根生物量变异, 但是林分平均基面积(basal area)和林分年龄分别能够解释林分水平细根生物量30%和单株水平细根生物量45%的变异(Finér et al., 2011a).与以上研究相比, 目前还无法获得全球尺度上吸收根生物量的实测数据, 但是仍有少量针对单个树种的根尖(或共生真菌侵染的1-2级根)生物量大尺度空间变异的研究(Helmisaari et al., 2009; Ostonen et al., 2011).由于根尖(或菌根)在根系资源吸收中的重要作用(如Guo et al., 2008c), 这些研究结果可以在一定程度上帮助我们了解吸收根生物量大尺度的变异格局.Ostonen等(2011)对欧洲12个挪威云杉(Picea abies)林分(纬度相差20°)的研究证实, 林分水平的菌根(大部分属于1、2级根)生物量在高纬度地区比低纬度地区大, 而且能够被N沉降量、年平均气温和土壤C:N比较好地解释; 但是Helmisaari等(2009)对相对较小的空间范围(纬度相差9°)的研究显示, 芬兰南部挪威云杉和欧洲赤松(Pinus sylvestris)根尖生物量与其芬兰北部生物量均没有显著的差别.最近,Zadworny等(2016)研究了北欧地区2000 km的梯度(纬度相差19°)上欧洲赤松吸收根比例的变异规律,发现2-5级根中具有吸收功能个体根的比例均随着年平均温度升高而降低(1级根全部是吸收根),证明寒冷地区树木分配了更多的根系生物量用于养分和水分的吸收.由于气候、土壤和人为干扰等因子在大尺度空间上存在着显著的变异性, 而且不同树种根系生长对环境变异的响应也存在差异(Eissenstat et al., 2015), 了解吸收根生物量在区域或全球尺度的变异规律和影响因素, 仍需在更广泛的地理区域上开展林分尺度上的研究. ...
Fine-root turnover patterns and their relationship to root diameter and soil depth in a 14C-labeled hardwood forest
1
2006
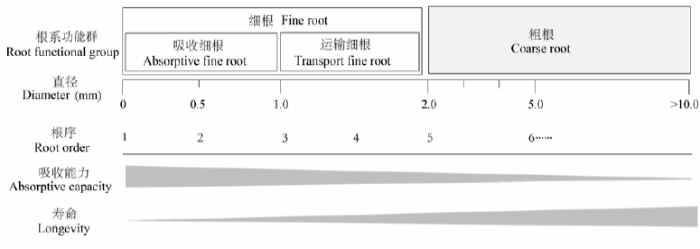
... 综合以往研究证据可见, 传统的细根由两类功能不同的根群组成, 即吸收根群(absorptive fine roots)和运输根群(transport fine roots) (Joslin et al., 2006; McCormack et al., 2015).前者由低级根组成, 是资源吸收的主体, 寿命短而周转快; 后者由高级根组成, 主要负责运输与存储, 寿命长而周转慢(图1).从物质循环的角度看, 将传统的细根按照根序把细根分为不同的库的方法(order-based method), 或采用功能划分的方法(functional-classification method)区分为吸收根和运输根2个库, 可以更准确地评价细根在生态系统中的重要作用(Guo et al., 2008b; Gaudinski et al., 2010; Parton et al., 2010; McCormack et al., 2015).然而, 将细根区分成单个根序进行研究, 如生物量估计, 需要消耗大量的时间和人力; 比较之下, 功能划分的方法既能减少工作量, 又能有效地进行不同处理间或树种间根系生物量及其他根系过程(如根呼吸和周转)的比较, 在根系研究的准确性和可行性上达到了更好的平衡(McCormack et al., 2015).因此, 采用功能划分方法能充分刻化树木根系统内在结构功能的异质性, 从而更加客观地评价根系主导的生态学过程及其影响.McCormack等(2015)最近基于功能划分的方法, 将传统的细根(直径≤2 mm)区分为吸收根和运输根2个库, 并估算相应的生物量和周转率, 结果显示细根周转消耗陆地生态系统NPP的比例为22%, 明显低于以往将细根按照直径阈值区分为1个库估算的结果(约33%, Jackson et al., 1997).然而, 与传统直径阈值的研究相比, 基于功能划分方法进行的根系研究是非常缺乏的. ...
Leading dimensions of root trait variation in subtropical forests
1
2014
... 首先, 采用根系功能划分的方法在获取吸收根和运输根生物量上展现出了很大的优势(例如Kong & Ma, 2014), 而且这一方法也可以应用到估计不同类群根系形态、组织化学和寿命与周转的研究中(Eissenstat et al., 2015; Liu et al., 2015).但是, 在测定某些根系功能属性时采用根序划分的方法可能是必要的, 例如根系解剖特征(如Guo et al., 2008c; Gu et al., 2014; Kong et al., 2014), 这是因为将不同根序解剖特征的数据混合并取平均数, 没有实质上的生物学意义, 而且可能引入新的测量误差. ...
Acquisition of ephemeral module in roots: A new view and test
2014
Growth, morphological traits and mycorrhizal colonization of fine roots respond differently to nitrogen addition in a slash pine plantation in subtropical China
2015
Relation of fine root distribution to soil C in a
2014
Life span and structure of ephemeral root modules of different functional groups from a desert system
1
2016
... 其次, 因为吸收根周转率高, 准确估计吸收根的生物量是构建森林生态系统物质循环模型的重要基础.采用微根管法(minirhizotron)观测的研究证实, 树木吸收根的中值寿命短则几周, 长则数月, 而最长也不超过2.5年(周转率至少为1年0.4次)(With- ington et al., 2006; McCormack et al., 2012; Liu et al., 2016).然而, 通过同位素或模型模拟等方法估计的运输根(在一些研究中称为“长寿命根”)的周转率远低于吸收根, 大部分为1年0.1-0.2次(Matamala et al., 2003; Lynch et al., 2013).更重要的是, 当排除研究方法上的差异时, Xia等(2010)采用根窗法对吸收根和运输根的研究证实, 水曲柳吸收根(1-3级根)的中值寿命为510-692 d (死亡率> 90%), 而运输根(4-5级根)的死亡率<10%; Liu等(2016)通过根窗法对我国新疆5种树木细根寿命的研究发现, 前3级根中值寿命为95-154 d, 而4-5级根均超过了550 d.因此, 吸收根是树木细根中最为活跃的组分, 在C和养分循环中发挥着重要的作用.由此可见, McCormack等(2015)曾假定吸收根周转率是运输根的5至10倍, 在很大程度上是合理的.基于这样的假定, 只有运输根的生物量现存量是吸收根的5-10倍时, 二者对细根生物量周转的贡献才相等.但是, 现有实测吸收根与运输根生物量的研究中, 运输根与吸收根的比例变化范围仅为0.80 (水曲柳, Wang et al., 2006)至1.85 (粗枝云杉, 刘运科等 2012), 远小于可能出现的理论值上限(如5-10).鉴于以往通过微根管法获得了较多树木吸收根寿命(或周转率)的数据(如Withington et al., 2006; Espeleta et al., 2009; McCormack et al., 2012; Chen & Brassard, 2013), 吸收根生物量的准确量化成为评价细根周转对森林地下生态学过程调控作用的关键. ...
Complementarity in nutrient foraging strategies of absorptive fine roots and arbuscular mycorrhizal fungi across 14 coexisting subtropical tree species
2
2015
... 首先, 采用根系功能划分的方法在获取吸收根和运输根生物量上展现出了很大的优势(例如Kong & Ma, 2014), 而且这一方法也可以应用到估计不同类群根系形态、组织化学和寿命与周转的研究中(Eissenstat et al., 2015; Liu et al., 2015).但是, 在测定某些根系功能属性时采用根序划分的方法可能是必要的, 例如根系解剖特征(如Guo et al., 2008c; Gu et al., 2014; Kong et al., 2014), 这是因为将不同根序解剖特征的数据混合并取平均数, 没有实质上的生物学意义, 而且可能引入新的测量误差. ...
... 最后, 本文主要关注了采用功能划分方法开展细根生物量与周转研究的进展, 但是功能划分方法在其他研究领域中的应用亟待开展.例如, 细根分解是树木向土壤中输入养分和有机质的重要途径(Silver & Miya, 2001; Freschet et al., 2013), 但是目前采用功能划分方法进行的相关研究较少(Fan & Guo, 2010; Goebel et al., 2011); 根系觅食策略(foraging strategy)及其对土壤资源异质性的响应, 也是长期以来受关注的研究问题(Campbell et al., 1991; McNickle et al., 2009; Liu et al., 2015), 吸收根概念的引入将有助于准确地理解根系功能属性与养分的互作机制.因此, 在根系生物量分配、生产和周转相关的研究中, 都有必要考虑功能划分方法的适用性. ...
Root Structure of Five Tree Species in Northeast China
2008
间伐对川西亚高山粗枝云杉人工林细根生物量及碳储量的影响
2012
Variation of the linkage of root function with root branch order
1
2013
... 其次, 在研究吸收根生物量和其他功能属性时, 一定要对不同树种的吸收根范围进行定义和检验(卫星等, 2008; McCormack et al., 2015), 这应该作为一个标准的研究环节而受到重视.可以先通过根解剖属性的测定判断哪些类群具有明显的吸收功能, 因为对于不同树种而言, 具有吸收能力的根系所包括的根序范围可能是不同的(Long et al., 2013).例如, Valenzuela-Estrada等(2008)对根直径很细的高丛蓝莓(Vaccinium corymbosum)研究发现, 前3级根均未出现次生生长, 且明显受到真菌侵染; Long等(2013)研究证实, 根直径较粗的合果木(Paramichelia baillonii)中1-5级根均存在完整的皮层组织并具有菌根侵染特征.但是Guo等(2008c)研究的23个树种前3级根中均存在皮层组织, 且受树种吸收根直径大小的影响不明显.因此, 树种吸收根直径大小可能并不是影响吸收根界定范围(包括几级根序)的重要因子, 表明树种水平上根系解剖学检验是必要的, 尤其是当研究中包括其他类群植物(如草本植物)时更应如此(Dong et al., 2015).值得注意的是, 目前并不清楚树木吸收根类群是否因地理区域的不同而发生明显的变化, 例如对一些广布种(如栎类), 是否在不同的环境梯度下包括了相同的低级根类群.因此, 即使对于同一物种, 也有必要在地理位置相距较远的不同地点上检验其吸收根所包括的根序范围(Zadworny et al., 2016), 并以此为基础研究根生物量和功能属性的变化. ...
Stored carbon partly fuels fine-root respiration but is not used for production of new fine roots
1
2013
... 其次, 因为吸收根周转率高, 准确估计吸收根的生物量是构建森林生态系统物质循环模型的重要基础.采用微根管法(minirhizotron)观测的研究证实, 树木吸收根的中值寿命短则几周, 长则数月, 而最长也不超过2.5年(周转率至少为1年0.4次)(With- ington et al., 2006; McCormack et al., 2012; Liu et al., 2016).然而, 通过同位素或模型模拟等方法估计的运输根(在一些研究中称为“长寿命根”)的周转率远低于吸收根, 大部分为1年0.1-0.2次(Matamala et al., 2003; Lynch et al., 2013).更重要的是, 当排除研究方法上的差异时, Xia等(2010)采用根窗法对吸收根和运输根的研究证实, 水曲柳吸收根(1-3级根)的中值寿命为510-692 d (死亡率> 90%), 而运输根(4-5级根)的死亡率<10%; Liu等(2016)通过根窗法对我国新疆5种树木细根寿命的研究发现, 前3级根中值寿命为95-154 d, 而4-5级根均超过了550 d.因此, 吸收根是树木细根中最为活跃的组分, 在C和养分循环中发挥着重要的作用.由此可见, McCormack等(2015)曾假定吸收根周转率是运输根的5至10倍, 在很大程度上是合理的.基于这样的假定, 只有运输根的生物量现存量是吸收根的5-10倍时, 二者对细根生物量周转的贡献才相等.但是, 现有实测吸收根与运输根生物量的研究中, 运输根与吸收根的比例变化范围仅为0.80 (水曲柳, Wang et al., 2006)至1.85 (粗枝云杉, 刘运科等 2012), 远小于可能出现的理论值上限(如5-10).鉴于以往通过微根管法获得了较多树木吸收根寿命(或周转率)的数据(如Withington et al., 2006; Espeleta et al., 2009; McCormack et al., 2012; Chen & Brassard, 2013), 吸收根生物量的准确量化成为评价细根周转对森林地下生态学过程调控作用的关键. ...
Measuring fine root turnover in forest ecosystems
1
2005
... 为了准确估计细根在生态系统物质分配和养分循环上的作用, 研究者需要知道细根的生物量现存量(单位面积上的干质量)和周转率(Norby & Jackson 2000; Majdi et al., 2005; Guo et al., 2008b).因此, 如果采用功能划分的方法进行根系生物量和周转的估计, 需要分别获得吸收根和运输根对应的参数(McCormack et al., 2015).然而, 由于概念理解和方法学上的限制, 相应的根系参数非常缺乏.在McCormack等(2015)的研究中, 是在以往Jackson等(1997)基于直径阈值方法的基础上, 将直径≤2 mm的生物量按照以往研究中吸收根和运输根的生物量比例(木本植物为10:90、33:67和60:40等3种不同比例), 并假定吸收根周转率为1年周转1次, 而运输根周转率为1年0.1次和0.2次两种情境, 来计算细根周转消耗的NPP.McCormack等(2015)的研究结果显示了采用功能划分方法来研究细根生物量和相关生态学过程的重要性.然而, 将根系结构与其功能, 特别是与生物量和周转联系起来, 只是在最近10余年才得到应有的重视(Wells et al., 2002; Guo et al., 2008c; Strand et al., 2008; Gu et al., 2011), 如今, 我们对吸收根和运输根生物量的变异规律、潜在机制的认识和理解并不全面.为此, 我们对近年来国内外采用根系功能划分方法开展根系生物量的研究进行了总结, 以期发现当前研究中存在的不足和亟待解决的问题, 在应用该方法时值得注意的问题, 以促进我国树木根系生态学的进一步发展. ...
Impacts of fine root turnover on forest
1
2003
... 其次, 因为吸收根周转率高, 准确估计吸收根的生物量是构建森林生态系统物质循环模型的重要基础.采用微根管法(minirhizotron)观测的研究证实, 树木吸收根的中值寿命短则几周, 长则数月, 而最长也不超过2.5年(周转率至少为1年0.4次)(With- ington et al., 2006; McCormack et al., 2012; Liu et al., 2016).然而, 通过同位素或模型模拟等方法估计的运输根(在一些研究中称为“长寿命根”)的周转率远低于吸收根, 大部分为1年0.1-0.2次(Matamala et al., 2003; Lynch et al., 2013).更重要的是, 当排除研究方法上的差异时, Xia等(2010)采用根窗法对吸收根和运输根的研究证实, 水曲柳吸收根(1-3级根)的中值寿命为510-692 d (死亡率> 90%), 而运输根(4-5级根)的死亡率<10%; Liu等(2016)通过根窗法对我国新疆5种树木细根寿命的研究发现, 前3级根中值寿命为95-154 d, 而4-5级根均超过了550 d.因此, 吸收根是树木细根中最为活跃的组分, 在C和养分循环中发挥着重要的作用.由此可见, McCormack等(2015)曾假定吸收根周转率是运输根的5至10倍, 在很大程度上是合理的.基于这样的假定, 只有运输根的生物量现存量是吸收根的5-10倍时, 二者对细根生物量周转的贡献才相等.但是, 现有实测吸收根与运输根生物量的研究中, 运输根与吸收根的比例变化范围仅为0.80 (水曲柳, Wang et al., 2006)至1.85 (粗枝云杉, 刘运科等 2012), 远小于可能出现的理论值上限(如5-10).鉴于以往通过微根管法获得了较多树木吸收根寿命(或周转率)的数据(如Withington et al., 2006; Espeleta et al., 2009; McCormack et al., 2012; Chen & Brassard, 2013), 吸收根生物量的准确量化成为评价细根周转对森林地下生态学过程调控作用的关键. ...
Predicting fine root lifespan from plant functional traits in temperate trees
2
2012
... 其次, 因为吸收根周转率高, 准确估计吸收根的生物量是构建森林生态系统物质循环模型的重要基础.采用微根管法(minirhizotron)观测的研究证实, 树木吸收根的中值寿命短则几周, 长则数月, 而最长也不超过2.5年(周转率至少为1年0.4次)(With- ington et al., 2006; McCormack et al., 2012; Liu et al., 2016).然而, 通过同位素或模型模拟等方法估计的运输根(在一些研究中称为“长寿命根”)的周转率远低于吸收根, 大部分为1年0.1-0.2次(Matamala et al., 2003; Lynch et al., 2013).更重要的是, 当排除研究方法上的差异时, Xia等(2010)采用根窗法对吸收根和运输根的研究证实, 水曲柳吸收根(1-3级根)的中值寿命为510-692 d (死亡率> 90%), 而运输根(4-5级根)的死亡率<10%; Liu等(2016)通过根窗法对我国新疆5种树木细根寿命的研究发现, 前3级根中值寿命为95-154 d, 而4-5级根均超过了550 d.因此, 吸收根是树木细根中最为活跃的组分, 在C和养分循环中发挥着重要的作用.由此可见, McCormack等(2015)曾假定吸收根周转率是运输根的5至10倍, 在很大程度上是合理的.基于这样的假定, 只有运输根的生物量现存量是吸收根的5-10倍时, 二者对细根生物量周转的贡献才相等.但是, 现有实测吸收根与运输根生物量的研究中, 运输根与吸收根的比例变化范围仅为0.80 (水曲柳, Wang et al., 2006)至1.85 (粗枝云杉, 刘运科等 2012), 远小于可能出现的理论值上限(如5-10).鉴于以往通过微根管法获得了较多树木吸收根寿命(或周转率)的数据(如Withington et al., 2006; Espeleta et al., 2009; McCormack et al., 2012; Chen & Brassard, 2013), 吸收根生物量的准确量化成为评价细根周转对森林地下生态学过程调控作用的关键. ...
... 近年来的研究揭示了树木细根在结构和功能上存在着高度的复杂性和异质性, 这为我们准确地理解细根在生态系统水平上的作用奠定了基础.早期研究者采用直径阈值的方法将细根和粗根区分开来, 也是考虑到二者在结构和功能上存在差异; 而近年来采用根序划分的方法证实单一直径阈值的方法忽略了个体根在根系分支上位置的不同, 因此, 基于根序开展研究可最大程度上减小取样单元内在的异质性(Pregitzer et al., 2002; Guo et al., 2008c).由于将细根区分为单个根序进行研究, 特别是以获取生物量为目的的研究, 对人力和时间的需求巨大, 导致该方法不能被广泛应用.因此, 采取根系功能划分的方法是一个折中的、高效的方法(McCormack et al., 2015).但是应当认识到, 在具体的根系研究中应用功能划分的方法仍存在着一些限制, 这里我们提出该方法存在的几点不足, 并提出一些建议供研究者参考. ...
Global patterns in root decomposition: Comparisons of climate and litter quality effects
1
2001
... 最后, 本文主要关注了采用功能划分方法开展细根生物量与周转研究的进展, 但是功能划分方法在其他研究领域中的应用亟待开展.例如, 细根分解是树木向土壤中输入养分和有机质的重要途径(Silver & Miya, 2001; Freschet et al., 2013), 但是目前采用功能划分方法进行的相关研究较少(Fan & Guo, 2010; Goebel et al., 2011); 根系觅食策略(foraging strategy)及其对土壤资源异质性的响应, 也是长期以来受关注的研究问题(Campbell et al., 1991; McNickle et al., 2009; Liu et al., 2015), 吸收根概念的引入将有助于准确地理解根系功能属性与养分的互作机制.因此, 在根系生物量分配、生产和周转相关的研究中, 都有必要考虑功能划分方法的适用性. ...
Irreconcilable differences: Fine-root life spans and
2
2008
... 细根一般指直径≤2 mm的根系组分, 是树木吸收养分和水分的重要器官.由于细根不断地进行生产、衰老、死亡和再生长过程(即周转), 在陆地生态系统碳(C)和养分循环中发挥着重要作用(Norby & Jackson, 2000; Strand et al., 2008).据估计, 全球尺度上, 如果细根每年周转一次, 将会消耗陆地生态系统净初级生产力(NPP)的33% (Jackson et al., 1997).以往大多数研究者将直径小于某一阈值(如1或2 mm)的个体根作为一个结构和功能一致的整体来研究, 然而, 近年来的研究证据显示, 以往采用直径大小对细根进行划分的方法, 很大程度上忽视了个体根在根系统上所处位置的不同, 即根序(branch order)的差异.不同根序个体根在解剖、形态和生理功能上存在明显的差异, 因此采用直径阈值来划分根系组分的方法缺乏对根系功能的充分考虑(虽然假定了根系功能与直径大小有关), 这会严重地阻碍我们对细根结构与功能的准确理解(Pregitzer et al., 2002; Guo et al., 2008a; Valenzuela- Estrada et al., 2008; McCormack et al., 2015). ...
... 为了准确估计细根在生态系统物质分配和养分循环上的作用, 研究者需要知道细根的生物量现存量(单位面积上的干质量)和周转率(Norby & Jackson 2000; Majdi et al., 2005; Guo et al., 2008b).因此, 如果采用功能划分的方法进行根系生物量和周转的估计, 需要分别获得吸收根和运输根对应的参数(McCormack et al., 2015).然而, 由于概念理解和方法学上的限制, 相应的根系参数非常缺乏.在McCormack等(2015)的研究中, 是在以往Jackson等(1997)基于直径阈值方法的基础上, 将直径≤2 mm的生物量按照以往研究中吸收根和运输根的生物量比例(木本植物为10:90、33:67和60:40等3种不同比例), 并假定吸收根周转率为1年周转1次, 而运输根周转率为1年0.1次和0.2次两种情境, 来计算细根周转消耗的NPP.McCormack等(2015)的研究结果显示了采用功能划分方法来研究细根生物量和相关生态学过程的重要性.然而, 将根系结构与其功能, 特别是与生物量和周转联系起来, 只是在最近10余年才得到应有的重视(Wells et al., 2002; Guo et al., 2008c; Strand et al., 2008; Gu et al., 2011), 如今, 我们对吸收根和运输根生物量的变异规律、潜在机制的认识和理解并不全面.为此, 我们对近年来国内外采用根系功能划分方法开展根系生物量的研究进行了总结, 以期发现当前研究中存在的不足和亟待解决的问题, 在应用该方法时值得注意的问题, 以促进我国树木根系生态学的进一步发展. ...
1
... 细根一般指直径≤2 mm的根系组分, 是树木吸收养分和水分的重要器官.由于细根不断地进行生产、衰老、死亡和再生长过程(即周转), 在陆地生态系统碳(C)和养分循环中发挥着重要作用(Norby & Jackson, 2000; Strand et al., 2008).据估计, 全球尺度上, 如果细根每年周转一次, 将会消耗陆地生态系统净初级生产力(NPP)的33% (Jackson et al., 1997).以往大多数研究者将直径小于某一阈值(如1或2 mm)的个体根作为一个结构和功能一致的整体来研究, 然而, 近年来的研究证据显示, 以往采用直径大小对细根进行划分的方法, 很大程度上忽视了个体根在根系统上所处位置的不同, 即根序(branch order)的差异.不同根序个体根在解剖、形态和生理功能上存在明显的差异, 因此采用直径阈值来划分根系组分的方法缺乏对根系功能的充分考虑(虽然假定了根系功能与直径大小有关), 这会严重地阻碍我们对细根结构与功能的准确理解(Pregitzer et al., 2002; Guo et al., 2008a; Valenzuela- Estrada et al., 2008; McCormack et al., 2015). ...
Root anatomy, morphology, and longevity among root orders in
2008
Aggregated and comple- mentary: Symmetric proliferation, overyielding, and mass effects explain fine-root biomass in soil patches in a diverse temperate deciduous forest landscape
2015
Review of root dynamics in forest ecosystems grouped by climate, climatic forest type and species
Fine root biomass, production, turnover rates, and nutrient contents in boreal forest ecosystems in relation to species, climate, fertility, and stand age: Literature review and meta-analyses
1
2010
... 其次, 在研究吸收根生物量和其他功能属性时, 一定要对不同树种的吸收根范围进行定义和检验(卫星等, 2008; McCormack et al., 2015), 这应该作为一个标准的研究环节而受到重视.可以先通过根解剖属性的测定判断哪些类群具有明显的吸收功能, 因为对于不同树种而言, 具有吸收能力的根系所包括的根序范围可能是不同的(Long et al., 2013).例如, Valenzuela-Estrada等(2008)对根直径很细的高丛蓝莓(Vaccinium corymbosum)研究发现, 前3级根均未出现次生生长, 且明显受到真菌侵染; Long等(2013)研究证实, 根直径较粗的合果木(Paramichelia baillonii)中1-5级根均存在完整的皮层组织并具有菌根侵染特征.但是Guo等(2008c)研究的23个树种前3级根中均存在皮层组织, 且受树种吸收根直径大小的影响不明显.因此, 树种吸收根直径大小可能并不是影响吸收根界定范围(包括几级根序)的重要因子, 表明树种水平上根系解剖学检验是必要的, 尤其是当研究中包括其他类群植物(如草本植物)时更应如此(Dong et al., 2015).值得注意的是, 目前并不清楚树木吸收根类群是否因地理区域的不同而发生明显的变化, 例如对一些广布种(如栎类), 是否在不同的环境梯度下包括了相同的低级根类群.因此, 即使对于同一物种, 也有必要在地理位置相距较远的不同地点上检验其吸收根所包括的根序范围(Zadworny et al., 2016), 并以此为基础研究根生物量和功能属性的变化. ...
Scots pine fine roots adjust along a 2000-km latitudinal climatic gradient


{kind=link}
{kind=link}