1华东师范大学生态与环境科学学院, 上海 2002412浙江天童森林生态系统国家野外科学观测研究站, 浙江宁波 3151143上海动物园, 上海 2000504Department of Environmental Sciences, Abdul Wali Khan University Mardan 23200, Pakistan
Hydraulic architecture of evergreen broad-leaved woody plants at different successional stages in Tiantong National Forest Park, Zhejiang Province, China
ZHAO Yan-Tao1, 2, XU Ming-Shan1, 2, ZHANG Zhi-Hao3, ZHOU Liu-Li1, 2, ZHANG Qing-Qing1, 2, Ali ARSHAD1, 2, 4, SONG Yan-Jun1, 2, YAN En-Rong1, 2, *,
1School of Ecological and Environment Sciences, East China Normal University, Shanghai 200241, China2Tiantong National Forest Ecosystem Observation and Research Station, Ningbo, Zhejiang 315114, China 3Shanghai Zoo Garden, Shanghai 200050, China4Department of Environmental Sciences, Abdul Wali Khan University Mardan 23200, Pakistan
Aims Hydraulic architecture is a morphological strategy in plants to transport water in coping with environmental conditions. Change of hydraulic architecture for plants occupying different canopy layers within community and for the same plant at different successional stages reflect existence and adaptation in plant's water transportation strategies. The objective of this study was to examine how hydraulic architecture varies with canopy layers within a community and with forest succession.Methods The study site is located in Tiantong National Forest Park, Zhejiang Province, China. Hydraulic architectural traits studied include sapwood-specific hydraulic conductivity, leaf-specific hydraulic conductivity, Huber value, sapwood channel area of twigs, total leaf area per terminal twig, and water potential of twigs. We measured those traits for species that occur in multiple successional stages (we called it "overlapping species") and for species that occur only in one successional stage (we called it "turnover species") along a successional series of evergreen broadleaved forests. For a given species, we sampled both overstory and understory trees. Hydraulic architectural traits between overstory and understory trees in the same community and at successional stages were compared. Pearson correlation was used to exam the relationship between hydraulic architectural traits and the twig/leaf traits.Important findings Sapwood-specific hydraulic conductivities and leaf-specific hydraulic conductivities were significantly higher in overstory trees than those in understory trees, but did not significantly differ from successional stages. Huber value decreased significantly for understory trees, but did not change for overstory trees through forest successional stages. For overstory trees, a trend of decreasing sapwood-specific hydraulic conductivity was observed for overlapping species but not for turnover species with successional stages. In contrast, for understory trees, a trend of decreasing Huber values was observed for turner species but not for overlapping species with successional stages. Across tree species, sapwood-specific hydraulic conductivity was positively correlated with sapwood channel area and total leaf area per terminal twig size. Huber value was negatively correlated to water potential of twigs and total leaf area per terminal twig size. These results suggest that water transportation capacity and efficiency are higher in overstory trees than in understory trees across successional stages in evergreen broadleaved forests in Tiantong region. The contrasting trends of sapwood-specific hydraulic conductivity between overlapping species and turnover species indicate that shift of microenvironment conditions might lead to changes of hydraulic architecture in overstory trees, whereas species replacement might result in changes of hydraulic architecture in understory trees.
Keywords:hydraulic architecture
;
overlapping species
;
overstory trees
;
turnover species
;
understory trees
ZHAOYan-Tao, XUMing-Shan, ZHANGZhi-Hao, ZHOULiu-Li, ZHANGQing-Qing, AliARSHAD, SONGYan-Jun, YANEn-Rong. Hydraulic architecture of evergreen broad-leaved woody plants at different successional stages in Tiantong National Forest Park, Zhejiang Province, China[J]. Chinese Journal of Plant Ecology, 2016, 40(2): 116-126 https://doi.org/10.17521/cjpe.2015.0258
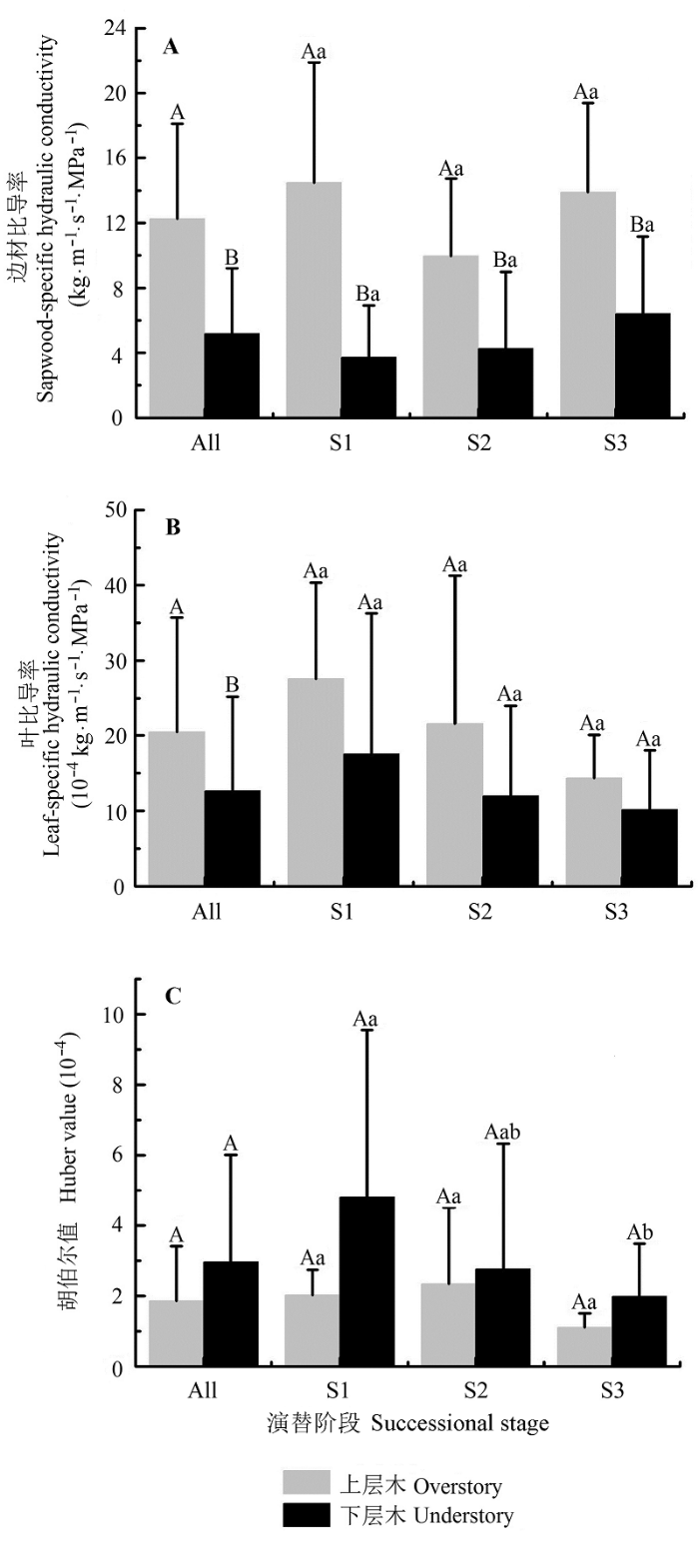
Fig. 1 Hydraulic architecture of trees between overstory and understory species along a successional series in evergreen broad-leaved forest in Tiantong, Zhejiang (mean ± SE). All means were combined for three successional stages. S1, S2 and S3 represent successional stages. Different capital letters indicate significant difference between canopy layers at the same successional stage (p < 0.05). Different lowercase letters indicate significant difference between successional stages for the same canopy layer (p < 0.05).
随演替进行, 上层木和下层木的3种水力结构参数都表现出了一定的变化趋势(图1).上层木3个边材比导率表现出先减小后增大的趋势, 但不显著(F = 1.83, p > 0.05), 下层木虽然随演替进行逐渐增大, 但演替阶段间差异不显著(F = 2.14, p > 0.05).上层木(F = 1.45, p > 0.05)和下层木(F = 1.35, p > 0.05)的叶比导率均随演替进行降低, 但不显著.上层木的胡伯尔值随演替先增后降, 但不显著(F = 1.75, p > 0.05), 下层木的胡伯尔值随演替进行显著下降(F = 3.67, p < 0.05).
Table 2 Results of two-way ANOVA for testing the main effects of canopy layers, succession stages and their interactions on hydraulic architecture of tree species
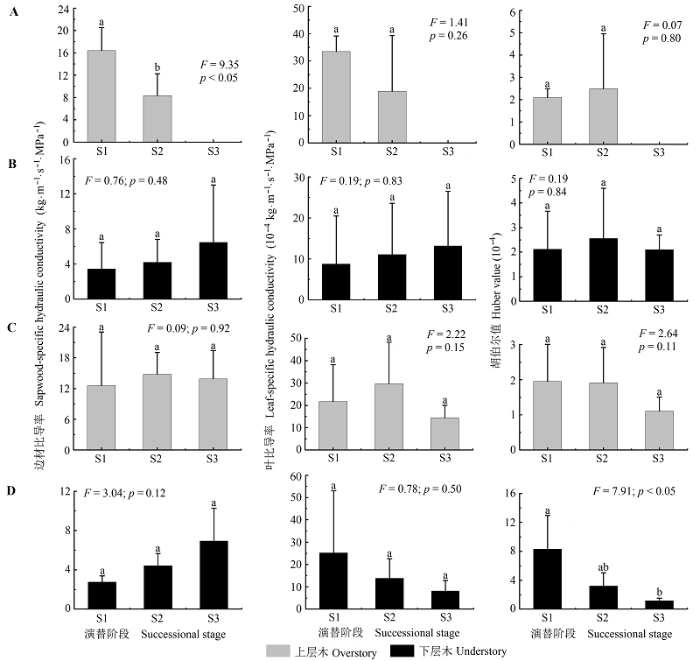
Fig. 2 Differences of hydraulic architecture for over lapping and turnover species in both overstory and understory layers along a successional series in evergreen broad-leaved forest in Tiantong, Zhejiang (mean ± SE). A, Overstory over lapping species. B, Understory over lapping species. C, Overstory turnover species. D, Understory turnover species. S1, S2 and S3 represent successional stages. Different letters among successional stages indicate significant difference (p < 0.05). F and p are results of one-way ANOVA.
Table 3 Coefficients of Pearson correlation between plant hydraulic architecture and branch and leaf traits in evergreen broad-leaved forests in Tiantong, Zhejiang
Inter-species variation of photosynthetic and xylem hydraulic traits in the deciduous and evergreen Euphorbiaceae tree species from a seasonally tropical forest in southwestern China.
A comparison of light environmental characteristics for evergreen broad-leaved forest communities from different successional stages in Tiantong National Forest Park.
Acta Ecologica Sinica, 25, 2862-2867(in Chinese with English abstract) .[丁圣彦, 卢训令, 李昊民 (2005).
Stem hydraulic traits and leaf water-stress tolerance are co-ordinated with the leaf phenology of angiosperm trees in an Asian tropical dry karst forest.
Water use by a warm-temperate deciduous forest under the influence of the Asian monsoon: Contributions of the overstory and understory to forest water use.
The ecological significance of long-distance water transport: Short-term regulation, long-term acclimation and the hydraulic costs of stature across plant life forms.
SongYC, WangXR (1995).Vegetation and Flora of Tiantong National Forest Park, Zhejiang Province, China. Shanghai Scientific and Technological Literature Publishing House, Shanghai.(in Chinese) [宋永昌, 王祥荣 (1995). 浙江天童国家森林公园的植被和区系. 上海科学技术文献出版社, 上海.]
Water relations and hydraulic architecture of a tropical tree (Schefflera morototoni): Data, models, and a comparison with two temperate species (Acer saccharum and Thuja occidentalis).
Plant Physiology, 96, 1105-1113.
[38]
TyreeMT, ZimmermannMH (2002).
Xylem Structure and the Ascent of Sap. Springer-Verlag
Pressure probe measurements of the driving forces for water transport in intact higher plants: Effects of transpiration and salinity. In: Läuchli A, Lüttge U eds. Salinity: Environment, Plants, Molecules
. Kluwer Academic Publishers, Dordrecht, the Netherlands.249-270.
Inter-species variation of photosynthetic and xylem hydraulic traits in the deciduous and evergreen Euphorbiaceae tree species from a seasonally tropical forest in southwestern China.
1
2009
... 水力结构是植物适应不同的水分供应以及生存竞争所形成的不同形态结构和水分运输策略(李吉跃和翟洪波, 2000; Mencuccini, 2003), 在保证植物体内长距离的水分运输(Thomas, 1997; 万贤崇和孟平, 2007),气孔调节和光合作用(Tyree & Ewers, 1991)等方面起着重要作用, 甚至它能限制植物的生长高度(Ryan & Yoder, 1997; Becker et al., 2000; McCulloh et al., 2015).通常情况下, 植物的水力结构采用导水率(hydraulic conductivity),边材比导率(sapwood-specific hydraulic conductivity),叶比导率(leaf-specific hydraulic conductivity)和胡伯尔值(Haber value)来表征(Tyree & Ewers,1991).这4个水力结构参数分别从不同的角度反映了植物茎段的水分运输能力,输水效率以及叶片供水情况.其中, 导水率反映了植物枝条或茎干的导水能力; 边材比导率是指单位茎段边材疏导面积的导水率, 边材比导率越大说明单位有效面积输送水分的能力越强, 输水效率越高; 叶比导率是指单位叶大小(如面积)的导水率, 表征了叶片的供水状况; 胡伯尔值表示支撑单位叶面积或叶干物质量上的枝条投入量, 胡伯尔值越大, 说明维持单位叶面积水分供给所需的枝条组织投入越多.植物水力结构除受物种遗传特性决定外, 在不同生活型间也表现不同, 例如, 裸子植物与被子植物(Zimmermann, 1978; Zimmermann & Sperry, 1983; Ewers & Zimmermann, 1984),木本植物与藤本植物(Zhu & Cao, 2009), 以及乔木与灌木(Tyree et al., 1983;1991),常绿物种与落叶物种(Drake et al., 2015)间均存在不同的输水策略.另外, 植物水力结构还受生境状况的影响, 尤其是受土壤水分与养分的影响(Carvajal et al., 1996).近年来, 有关亚热带,热带地区森林植物水力结构的研究越来越多地引起关注, 例如: 对植物水力结构与光合作用的耦合研究(Brodribb & Field, 2000; Brodribb et al., 2002; Chen et al., 2009; Fu et al., 2012; Zhu et al., 2013), 以及植物水力结构与枝叶性状关系(Hoeber et al., 2014; Zhu et al., 2015)的研究. ...
Xylem embolism and drought-induced stomatal closure in maize.
Isometric partitioning of hydraulic conductance between leaves and stems: Balancing safety and efficiency in different growth forms and habitats.
1
2015
... 水力结构是植物适应不同的水分供应以及生存竞争所形成的不同形态结构和水分运输策略(李吉跃和翟洪波, 2000; Mencuccini, 2003), 在保证植物体内长距离的水分运输(Thomas, 1997; 万贤崇和孟平, 2007),气孔调节和光合作用(Tyree & Ewers, 1991)等方面起着重要作用, 甚至它能限制植物的生长高度(Ryan & Yoder, 1997; Becker et al., 2000; McCulloh et al., 2015).通常情况下, 植物的水力结构采用导水率(hydraulic conductivity),边材比导率(sapwood-specific hydraulic conductivity),叶比导率(leaf-specific hydraulic conductivity)和胡伯尔值(Haber value)来表征(Tyree & Ewers,1991).这4个水力结构参数分别从不同的角度反映了植物茎段的水分运输能力,输水效率以及叶片供水情况.其中, 导水率反映了植物枝条或茎干的导水能力; 边材比导率是指单位茎段边材疏导面积的导水率, 边材比导率越大说明单位有效面积输送水分的能力越强, 输水效率越高; 叶比导率是指单位叶大小(如面积)的导水率, 表征了叶片的供水状况; 胡伯尔值表示支撑单位叶面积或叶干物质量上的枝条投入量, 胡伯尔值越大, 说明维持单位叶面积水分供给所需的枝条组织投入越多.植物水力结构除受物种遗传特性决定外, 在不同生活型间也表现不同, 例如, 裸子植物与被子植物(Zimmermann, 1978; Zimmermann & Sperry, 1983; Ewers & Zimmermann, 1984),木本植物与藤本植物(Zhu & Cao, 2009), 以及乔木与灌木(Tyree et al., 1983;1991),常绿物种与落叶物种(Drake et al., 2015)间均存在不同的输水策略.另外, 植物水力结构还受生境状况的影响, 尤其是受土壤水分与养分的影响(Carvajal et al., 1996).近年来, 有关亚热带,热带地区森林植物水力结构的研究越来越多地引起关注, 例如: 对植物水力结构与光合作用的耦合研究(Brodribb & Field, 2000; Brodribb et al., 2002; Chen et al., 2009; Fu et al., 2012; Zhu et al., 2013), 以及植物水力结构与枝叶性状关系(Hoeber et al., 2014; Zhu et al., 2015)的研究. ...
The hydraulic architecture of balsam fir (Abies balsamea).
1
1984
... 水力结构是植物适应不同的水分供应以及生存竞争所形成的不同形态结构和水分运输策略(李吉跃和翟洪波, 2000; Mencuccini, 2003), 在保证植物体内长距离的水分运输(Thomas, 1997; 万贤崇和孟平, 2007),气孔调节和光合作用(Tyree & Ewers, 1991)等方面起着重要作用, 甚至它能限制植物的生长高度(Ryan & Yoder, 1997; Becker et al., 2000; McCulloh et al., 2015).通常情况下, 植物的水力结构采用导水率(hydraulic conductivity),边材比导率(sapwood-specific hydraulic conductivity),叶比导率(leaf-specific hydraulic conductivity)和胡伯尔值(Haber value)来表征(Tyree & Ewers,1991).这4个水力结构参数分别从不同的角度反映了植物茎段的水分运输能力,输水效率以及叶片供水情况.其中, 导水率反映了植物枝条或茎干的导水能力; 边材比导率是指单位茎段边材疏导面积的导水率, 边材比导率越大说明单位有效面积输送水分的能力越强, 输水效率越高; 叶比导率是指单位叶大小(如面积)的导水率, 表征了叶片的供水状况; 胡伯尔值表示支撑单位叶面积或叶干物质量上的枝条投入量, 胡伯尔值越大, 说明维持单位叶面积水分供给所需的枝条组织投入越多.植物水力结构除受物种遗传特性决定外, 在不同生活型间也表现不同, 例如, 裸子植物与被子植物(Zimmermann, 1978; Zimmermann & Sperry, 1983; Ewers & Zimmermann, 1984),木本植物与藤本植物(Zhu & Cao, 2009), 以及乔木与灌木(Tyree et al., 1983;1991),常绿物种与落叶物种(Drake et al., 2015)间均存在不同的输水策略.另外, 植物水力结构还受生境状况的影响, 尤其是受土壤水分与养分的影响(Carvajal et al., 1996).近年来, 有关亚热带,热带地区森林植物水力结构的研究越来越多地引起关注, 例如: 对植物水力结构与光合作用的耦合研究(Brodribb & Field, 2000; Brodribb et al., 2002; Chen et al., 2009; Fu et al., 2012; Zhu et al., 2013), 以及植物水力结构与枝叶性状关系(Hoeber et al., 2014; Zhu et al., 2015)的研究. ...
Some anatomical adaptations of desert plants.
1
1964
... 蒸腾-内聚力-张力学说常被用来解释植物水力结构随树高变化的现象, 即: 叶片蒸腾产生拉力, 通过木质部中的连续水柱将水分不断提升到树冠部分, 进行水气交换.内聚力学说认为: 蒸腾拉力是水分在植物体内向上运输的主要驱动力(Zimmer- mann et al., 2002).随树高的增加, 植物水分胁迫越来越严重(Koch et al., 2004).解剖观察(Fahn, 1964)和理论推测(West et al., 1999)均表明, 导管或管胞直径随树高的增加而逐渐变小.根据Hagen- Poiseuille理论, 水分输导能力随管道直径成4次方指数的变化, 除了水柱重力越来越大外, 输水路径的阻力也越来越大, 因此, 乔木在水分运输过程中, 要克服更大的水分重力和输导阻力.水分蒸散产生的拉力大小主要取决于蒸腾通量, 而植物蒸腾通量主要由边材解剖特征决定, 如导管直径的大小,导管的数量及密度等(樊大勇和谢宗强, 2004).乔木与灌木相比往往具有更大的导管直径(Carlquist & Hoekman, 1985),较粗的末端枝条, 因此上层木表现出了较大的边材比导率和叶比导率(图1), 即较大的输水能力和较高的输水效率, 从而弥补了长距离水分运输较大阻力带来的影响, 以确保足够的水分供应, 满足正常的生理活动需要. ...
木质部导管空穴化研究中的几个热点问题
1
2004
... 蒸腾-内聚力-张力学说常被用来解释植物水力结构随树高变化的现象, 即: 叶片蒸腾产生拉力, 通过木质部中的连续水柱将水分不断提升到树冠部分, 进行水气交换.内聚力学说认为: 蒸腾拉力是水分在植物体内向上运输的主要驱动力(Zimmer- mann et al., 2002).随树高的增加, 植物水分胁迫越来越严重(Koch et al., 2004).解剖观察(Fahn, 1964)和理论推测(West et al., 1999)均表明, 导管或管胞直径随树高的增加而逐渐变小.根据Hagen- Poiseuille理论, 水分输导能力随管道直径成4次方指数的变化, 除了水柱重力越来越大外, 输水路径的阻力也越来越大, 因此, 乔木在水分运输过程中, 要克服更大的水分重力和输导阻力.水分蒸散产生的拉力大小主要取决于蒸腾通量, 而植物蒸腾通量主要由边材解剖特征决定, 如导管直径的大小,导管的数量及密度等(樊大勇和谢宗强, 2004).乔木与灌木相比往往具有更大的导管直径(Carlquist & Hoekman, 1985),较粗的末端枝条, 因此上层木表现出了较大的边材比导率和叶比导率(图1), 即较大的输水能力和较高的输水效率, 从而弥补了长距离水分运输较大阻力带来的影响, 以确保足够的水分供应, 满足正常的生理活动需要. ...
Stem hydraulic traits and leaf water-stress tolerance are co-ordinated with the leaf phenology of angiosperm trees in an Asian tropical dry karst forest.
1
2012
... 水力结构是植物适应不同的水分供应以及生存竞争所形成的不同形态结构和水分运输策略(李吉跃和翟洪波, 2000; Mencuccini, 2003), 在保证植物体内长距离的水分运输(Thomas, 1997; 万贤崇和孟平, 2007),气孔调节和光合作用(Tyree & Ewers, 1991)等方面起着重要作用, 甚至它能限制植物的生长高度(Ryan & Yoder, 1997; Becker et al., 2000; McCulloh et al., 2015).通常情况下, 植物的水力结构采用导水率(hydraulic conductivity),边材比导率(sapwood-specific hydraulic conductivity),叶比导率(leaf-specific hydraulic conductivity)和胡伯尔值(Haber value)来表征(Tyree & Ewers,1991).这4个水力结构参数分别从不同的角度反映了植物茎段的水分运输能力,输水效率以及叶片供水情况.其中, 导水率反映了植物枝条或茎干的导水能力; 边材比导率是指单位茎段边材疏导面积的导水率, 边材比导率越大说明单位有效面积输送水分的能力越强, 输水效率越高; 叶比导率是指单位叶大小(如面积)的导水率, 表征了叶片的供水状况; 胡伯尔值表示支撑单位叶面积或叶干物质量上的枝条投入量, 胡伯尔值越大, 说明维持单位叶面积水分供给所需的枝条组织投入越多.植物水力结构除受物种遗传特性决定外, 在不同生活型间也表现不同, 例如, 裸子植物与被子植物(Zimmermann, 1978; Zimmermann & Sperry, 1983; Ewers & Zimmermann, 1984),木本植物与藤本植物(Zhu & Cao, 2009), 以及乔木与灌木(Tyree et al., 1983;1991),常绿物种与落叶物种(Drake et al., 2015)间均存在不同的输水策略.另外, 植物水力结构还受生境状况的影响, 尤其是受土壤水分与养分的影响(Carvajal et al., 1996).近年来, 有关亚热带,热带地区森林植物水力结构的研究越来越多地引起关注, 例如: 对植物水力结构与光合作用的耦合研究(Brodribb & Field, 2000; Brodribb et al., 2002; Chen et al., 2009; Fu et al., 2012; Zhu et al., 2013), 以及植物水力结构与枝叶性状关系(Hoeber et al., 2014; Zhu et al., 2015)的研究. ...
Canopy closure exerts weak controls on understory dynamics: A 30-year study of overstory-understory interactions.
The importance of hydraulic conductivity and wood density to growth performance in eight tree species from a tropical semi-dry climate.
1
2014
... 水力结构是植物适应不同的水分供应以及生存竞争所形成的不同形态结构和水分运输策略(李吉跃和翟洪波, 2000; Mencuccini, 2003), 在保证植物体内长距离的水分运输(Thomas, 1997; 万贤崇和孟平, 2007),气孔调节和光合作用(Tyree & Ewers, 1991)等方面起着重要作用, 甚至它能限制植物的生长高度(Ryan & Yoder, 1997; Becker et al., 2000; McCulloh et al., 2015).通常情况下, 植物的水力结构采用导水率(hydraulic conductivity),边材比导率(sapwood-specific hydraulic conductivity),叶比导率(leaf-specific hydraulic conductivity)和胡伯尔值(Haber value)来表征(Tyree & Ewers,1991).这4个水力结构参数分别从不同的角度反映了植物茎段的水分运输能力,输水效率以及叶片供水情况.其中, 导水率反映了植物枝条或茎干的导水能力; 边材比导率是指单位茎段边材疏导面积的导水率, 边材比导率越大说明单位有效面积输送水分的能力越强, 输水效率越高; 叶比导率是指单位叶大小(如面积)的导水率, 表征了叶片的供水状况; 胡伯尔值表示支撑单位叶面积或叶干物质量上的枝条投入量, 胡伯尔值越大, 说明维持单位叶面积水分供给所需的枝条组织投入越多.植物水力结构除受物种遗传特性决定外, 在不同生活型间也表现不同, 例如, 裸子植物与被子植物(Zimmermann, 1978; Zimmermann & Sperry, 1983; Ewers & Zimmermann, 1984),木本植物与藤本植物(Zhu & Cao, 2009), 以及乔木与灌木(Tyree et al., 1983;1991),常绿物种与落叶物种(Drake et al., 2015)间均存在不同的输水策略.另外, 植物水力结构还受生境状况的影响, 尤其是受土壤水分与养分的影响(Carvajal et al., 1996).近年来, 有关亚热带,热带地区森林植物水力结构的研究越来越多地引起关注, 例如: 对植物水力结构与光合作用的耦合研究(Brodribb & Field, 2000; Brodribb et al., 2002; Chen et al., 2009; Fu et al., 2012; Zhu et al., 2013), 以及植物水力结构与枝叶性状关系(Hoeber et al., 2014; Zhu et al., 2015)的研究. ...
Water use by a warm-temperate deciduous forest under the influence of the Asian monsoon: Contributions of the overstory and understory to forest water use.
The ecological significance of long-distance water transport: Short-term regulation, long-term acclimation and the hydraulic costs of stature across plant life forms.
1
2003
... 水力结构是植物适应不同的水分供应以及生存竞争所形成的不同形态结构和水分运输策略(李吉跃和翟洪波, 2000; Mencuccini, 2003), 在保证植物体内长距离的水分运输(Thomas, 1997; 万贤崇和孟平, 2007),气孔调节和光合作用(Tyree & Ewers, 1991)等方面起着重要作用, 甚至它能限制植物的生长高度(Ryan & Yoder, 1997; Becker et al., 2000; McCulloh et al., 2015).通常情况下, 植物的水力结构采用导水率(hydraulic conductivity),边材比导率(sapwood-specific hydraulic conductivity),叶比导率(leaf-specific hydraulic conductivity)和胡伯尔值(Haber value)来表征(Tyree & Ewers,1991).这4个水力结构参数分别从不同的角度反映了植物茎段的水分运输能力,输水效率以及叶片供水情况.其中, 导水率反映了植物枝条或茎干的导水能力; 边材比导率是指单位茎段边材疏导面积的导水率, 边材比导率越大说明单位有效面积输送水分的能力越强, 输水效率越高; 叶比导率是指单位叶大小(如面积)的导水率, 表征了叶片的供水状况; 胡伯尔值表示支撑单位叶面积或叶干物质量上的枝条投入量, 胡伯尔值越大, 说明维持单位叶面积水分供给所需的枝条组织投入越多.植物水力结构除受物种遗传特性决定外, 在不同生活型间也表现不同, 例如, 裸子植物与被子植物(Zimmermann, 1978; Zimmermann & Sperry, 1983; Ewers & Zimmermann, 1984),木本植物与藤本植物(Zhu & Cao, 2009), 以及乔木与灌木(Tyree et al., 1983;1991),常绿物种与落叶物种(Drake et al., 2015)间均存在不同的输水策略.另外, 植物水力结构还受生境状况的影响, 尤其是受土壤水分与养分的影响(Carvajal et al., 1996).近年来, 有关亚热带,热带地区森林植物水力结构的研究越来越多地引起关注, 例如: 对植物水力结构与光合作用的耦合研究(Brodribb & Field, 2000; Brodribb et al., 2002; Chen et al., 2009; Fu et al., 2012; Zhu et al., 2013), 以及植物水力结构与枝叶性状关系(Hoeber et al., 2014; Zhu et al., 2015)的研究. ...
How do traits vary across ecological scales? A case for trait-based ecology.
Simulation of water flow in the branched tree architecture.
1
1997
... 水力结构是植物适应不同的水分供应以及生存竞争所形成的不同形态结构和水分运输策略(李吉跃和翟洪波, 2000; Mencuccini, 2003), 在保证植物体内长距离的水分运输(Thomas, 1997; 万贤崇和孟平, 2007),气孔调节和光合作用(Tyree & Ewers, 1991)等方面起着重要作用, 甚至它能限制植物的生长高度(Ryan & Yoder, 1997; Becker et al., 2000; McCulloh et al., 2015).通常情况下, 植物的水力结构采用导水率(hydraulic conductivity),边材比导率(sapwood-specific hydraulic conductivity),叶比导率(leaf-specific hydraulic conductivity)和胡伯尔值(Haber value)来表征(Tyree & Ewers,1991).这4个水力结构参数分别从不同的角度反映了植物茎段的水分运输能力,输水效率以及叶片供水情况.其中, 导水率反映了植物枝条或茎干的导水能力; 边材比导率是指单位茎段边材疏导面积的导水率, 边材比导率越大说明单位有效面积输送水分的能力越强, 输水效率越高; 叶比导率是指单位叶大小(如面积)的导水率, 表征了叶片的供水状况; 胡伯尔值表示支撑单位叶面积或叶干物质量上的枝条投入量, 胡伯尔值越大, 说明维持单位叶面积水分供给所需的枝条组织投入越多.植物水力结构除受物种遗传特性决定外, 在不同生活型间也表现不同, 例如, 裸子植物与被子植物(Zimmermann, 1978; Zimmermann & Sperry, 1983; Ewers & Zimmermann, 1984),木本植物与藤本植物(Zhu & Cao, 2009), 以及乔木与灌木(Tyree et al., 1983;1991),常绿物种与落叶物种(Drake et al., 2015)间均存在不同的输水策略.另外, 植物水力结构还受生境状况的影响, 尤其是受土壤水分与养分的影响(Carvajal et al., 1996).近年来, 有关亚热带,热带地区森林植物水力结构的研究越来越多地引起关注, 例如: 对植物水力结构与光合作用的耦合研究(Brodribb & Field, 2000; Brodribb et al., 2002; Chen et al., 2009; Fu et al., 2012; Zhu et al., 2013), 以及植物水力结构与枝叶性状关系(Hoeber et al., 2014; Zhu et al., 2015)的研究. ...
The hydraulic architecture of trees and other woody plants.
2
1991
... 水力结构是植物适应不同的水分供应以及生存竞争所形成的不同形态结构和水分运输策略(李吉跃和翟洪波, 2000; Mencuccini, 2003), 在保证植物体内长距离的水分运输(Thomas, 1997; 万贤崇和孟平, 2007),气孔调节和光合作用(Tyree & Ewers, 1991)等方面起着重要作用, 甚至它能限制植物的生长高度(Ryan & Yoder, 1997; Becker et al., 2000; McCulloh et al., 2015).通常情况下, 植物的水力结构采用导水率(hydraulic conductivity),边材比导率(sapwood-specific hydraulic conductivity),叶比导率(leaf-specific hydraulic conductivity)和胡伯尔值(Haber value)来表征(Tyree & Ewers,1991).这4个水力结构参数分别从不同的角度反映了植物茎段的水分运输能力,输水效率以及叶片供水情况.其中, 导水率反映了植物枝条或茎干的导水能力; 边材比导率是指单位茎段边材疏导面积的导水率, 边材比导率越大说明单位有效面积输送水分的能力越强, 输水效率越高; 叶比导率是指单位叶大小(如面积)的导水率, 表征了叶片的供水状况; 胡伯尔值表示支撑单位叶面积或叶干物质量上的枝条投入量, 胡伯尔值越大, 说明维持单位叶面积水分供给所需的枝条组织投入越多.植物水力结构除受物种遗传特性决定外, 在不同生活型间也表现不同, 例如, 裸子植物与被子植物(Zimmermann, 1978; Zimmermann & Sperry, 1983; Ewers & Zimmermann, 1984),木本植物与藤本植物(Zhu & Cao, 2009), 以及乔木与灌木(Tyree et al., 1983;1991),常绿物种与落叶物种(Drake et al., 2015)间均存在不同的输水策略.另外, 植物水力结构还受生境状况的影响, 尤其是受土壤水分与养分的影响(Carvajal et al., 1996).近年来, 有关亚热带,热带地区森林植物水力结构的研究越来越多地引起关注, 例如: 对植物水力结构与光合作用的耦合研究(Brodribb & Field, 2000; Brodribb et al., 2002; Chen et al., 2009; Fu et al., 2012; Zhu et al., 2013), 以及植物水力结构与枝叶性状关系(Hoeber et al., 2014; Zhu et al., 2015)的研究. ...
... ).通常情况下, 植物的水力结构采用导水率(hydraulic conductivity),边材比导率(sapwood-specific hydraulic conductivity),叶比导率(leaf-specific hydraulic conductivity)和胡伯尔值(Haber value)来表征(Tyree & Ewers,1991).这4个水力结构参数分别从不同的角度反映了植物茎段的水分运输能力,输水效率以及叶片供水情况.其中, 导水率反映了植物枝条或茎干的导水能力; 边材比导率是指单位茎段边材疏导面积的导水率, 边材比导率越大说明单位有效面积输送水分的能力越强, 输水效率越高; 叶比导率是指单位叶大小(如面积)的导水率, 表征了叶片的供水状况; 胡伯尔值表示支撑单位叶面积或叶干物质量上的枝条投入量, 胡伯尔值越大, 说明维持单位叶面积水分供给所需的枝条组织投入越多.植物水力结构除受物种遗传特性决定外, 在不同生活型间也表现不同, 例如, 裸子植物与被子植物(Zimmermann, 1978; Zimmermann & Sperry, 1983; Ewers & Zimmermann, 1984),木本植物与藤本植物(Zhu & Cao, 2009), 以及乔木与灌木(Tyree et al., 1983;1991),常绿物种与落叶物种(Drake et al., 2015)间均存在不同的输水策略.另外, 植物水力结构还受生境状况的影响, 尤其是受土壤水分与养分的影响(Carvajal et al., 1996).近年来, 有关亚热带,热带地区森林植物水力结构的研究越来越多地引起关注, 例如: 对植物水力结构与光合作用的耦合研究(Brodribb & Field, 2000; Brodribb et al., 2002; Chen et al., 2009; Fu et al., 2012; Zhu et al., 2013), 以及植物水力结构与枝叶性状关系(Hoeber et al., 2014; Zhu et al., 2015)的研究. ...
The hydraulic architecture of Thuja occidentalis.
1
1983
... 水力结构是植物适应不同的水分供应以及生存竞争所形成的不同形态结构和水分运输策略(李吉跃和翟洪波, 2000; Mencuccini, 2003), 在保证植物体内长距离的水分运输(Thomas, 1997; 万贤崇和孟平, 2007),气孔调节和光合作用(Tyree & Ewers, 1991)等方面起着重要作用, 甚至它能限制植物的生长高度(Ryan & Yoder, 1997; Becker et al., 2000; McCulloh et al., 2015).通常情况下, 植物的水力结构采用导水率(hydraulic conductivity),边材比导率(sapwood-specific hydraulic conductivity),叶比导率(leaf-specific hydraulic conductivity)和胡伯尔值(Haber value)来表征(Tyree & Ewers,1991).这4个水力结构参数分别从不同的角度反映了植物茎段的水分运输能力,输水效率以及叶片供水情况.其中, 导水率反映了植物枝条或茎干的导水能力; 边材比导率是指单位茎段边材疏导面积的导水率, 边材比导率越大说明单位有效面积输送水分的能力越强, 输水效率越高; 叶比导率是指单位叶大小(如面积)的导水率, 表征了叶片的供水状况; 胡伯尔值表示支撑单位叶面积或叶干物质量上的枝条投入量, 胡伯尔值越大, 说明维持单位叶面积水分供给所需的枝条组织投入越多.植物水力结构除受物种遗传特性决定外, 在不同生活型间也表现不同, 例如, 裸子植物与被子植物(Zimmermann, 1978; Zimmermann & Sperry, 1983; Ewers & Zimmermann, 1984),木本植物与藤本植物(Zhu & Cao, 2009), 以及乔木与灌木(Tyree et al., 1983;1991),常绿物种与落叶物种(Drake et al., 2015)间均存在不同的输水策略.另外, 植物水力结构还受生境状况的影响, 尤其是受土壤水分与养分的影响(Carvajal et al., 1996).近年来, 有关亚热带,热带地区森林植物水力结构的研究越来越多地引起关注, 例如: 对植物水力结构与光合作用的耦合研究(Brodribb & Field, 2000; Brodribb et al., 2002; Chen et al., 2009; Fu et al., 2012; Zhu et al., 2013), 以及植物水力结构与枝叶性状关系(Hoeber et al., 2014; Zhu et al., 2015)的研究. ...
Water relations and hydraulic architecture of a tropical tree (Schefflera morototoni): Data, models, and a comparison with two temperate species (Acer saccharum and Thuja occidentalis).
1991
Xylem Structure and the Ascent of Sap. Springer-Verlag
Anatomy of the palm Rhapis excelsa. IX. Xylem structure of the leaf insertion.
1
1983
... 水力结构是植物适应不同的水分供应以及生存竞争所形成的不同形态结构和水分运输策略(李吉跃和翟洪波, 2000; Mencuccini, 2003), 在保证植物体内长距离的水分运输(Thomas, 1997; 万贤崇和孟平, 2007),气孔调节和光合作用(Tyree & Ewers, 1991)等方面起着重要作用, 甚至它能限制植物的生长高度(Ryan & Yoder, 1997; Becker et al., 2000; McCulloh et al., 2015).通常情况下, 植物的水力结构采用导水率(hydraulic conductivity),边材比导率(sapwood-specific hydraulic conductivity),叶比导率(leaf-specific hydraulic conductivity)和胡伯尔值(Haber value)来表征(Tyree & Ewers,1991).这4个水力结构参数分别从不同的角度反映了植物茎段的水分运输能力,输水效率以及叶片供水情况.其中, 导水率反映了植物枝条或茎干的导水能力; 边材比导率是指单位茎段边材疏导面积的导水率, 边材比导率越大说明单位有效面积输送水分的能力越强, 输水效率越高; 叶比导率是指单位叶大小(如面积)的导水率, 表征了叶片的供水状况; 胡伯尔值表示支撑单位叶面积或叶干物质量上的枝条投入量, 胡伯尔值越大, 说明维持单位叶面积水分供给所需的枝条组织投入越多.植物水力结构除受物种遗传特性决定外, 在不同生活型间也表现不同, 例如, 裸子植物与被子植物(Zimmermann, 1978; Zimmermann & Sperry, 1983; Ewers & Zimmermann, 1984),木本植物与藤本植物(Zhu & Cao, 2009), 以及乔木与灌木(Tyree et al., 1983;1991),常绿物种与落叶物种(Drake et al., 2015)间均存在不同的输水策略.另外, 植物水力结构还受生境状况的影响, 尤其是受土壤水分与养分的影响(Carvajal et al., 1996).近年来, 有关亚热带,热带地区森林植物水力结构的研究越来越多地引起关注, 例如: 对植物水力结构与光合作用的耦合研究(Brodribb & Field, 2000; Brodribb et al., 2002; Chen et al., 2009; Fu et al., 2012; Zhu et al., 2013), 以及植物水力结构与枝叶性状关系(Hoeber et al., 2014; Zhu et al., 2015)的研究. ...
Pressure probe measurements of the driving forces for water transport in intact higher plants: Effects of transpiration and salinity. In: Läuchli A, Lüttge U eds. Salinity: Environment, Plants, Molecules
1
... 蒸腾-内聚力-张力学说常被用来解释植物水力结构随树高变化的现象, 即: 叶片蒸腾产生拉力, 通过木质部中的连续水柱将水分不断提升到树冠部分, 进行水气交换.内聚力学说认为: 蒸腾拉力是水分在植物体内向上运输的主要驱动力(Zimmer- mann et al., 2002).随树高的增加, 植物水分胁迫越来越严重(Koch et al., 2004).解剖观察(Fahn, 1964)和理论推测(West et al., 1999)均表明, 导管或管胞直径随树高的增加而逐渐变小.根据Hagen- Poiseuille理论, 水分输导能力随管道直径成4次方指数的变化, 除了水柱重力越来越大外, 输水路径的阻力也越来越大, 因此, 乔木在水分运输过程中, 要克服更大的水分重力和输导阻力.水分蒸散产生的拉力大小主要取决于蒸腾通量, 而植物蒸腾通量主要由边材解剖特征决定, 如导管直径的大小,导管的数量及密度等(樊大勇和谢宗强, 2004).乔木与灌木相比往往具有更大的导管直径(Carlquist & Hoekman, 1985),较粗的末端枝条, 因此上层木表现出了较大的边材比导率和叶比导率(图1), 即较大的输水能力和较高的输水效率, 从而弥补了长距离水分运输较大阻力带来的影响, 以确保足够的水分供应, 满足正常的生理活动需要. ...



{kind=link}
{kind=link}
{kind=link}
{kind=link}