根据《中国统计年鉴》, 近年来全国桃园面积及产量稳步上升.桃(Amygdalus persica )是继苹果(Malus pumila ),梨(Pyrus ),葡萄(Vitis )之后的第四大水果.桃树枝条萌芽率高, 成枝力强, 枝叶生长量大, 生长旺的枝条一年可萌发2-3次副梢(Desmond & Daniele, 2008 ).栽培管理过程中需对桃树进行大量修剪以保证树体通风透光, 进而带来了剪落枝条大量堆积处理困难的问题, 这不仅影响桃园环境, 而且容易引起病虫害的发生.有关研究发现, 小麦(Triticum aestivum )和玉米(Zea mays )秸秆还田均有增产的效果(韩新忠等, 2012 ; 慕平等, 2012 ).因此, 枝条还田的提出, 在杜绝枝条杂乱占用果园面积影响生产的同时还可能有增肥增产作用, 为处理枝条提供了一种可供选择的方案.
桃树再植当年基本不生新根, 发枝短长势弱, 造成桃园减产, 连作障碍严重(刘嘉彬等, 2006 ).桃树根系,枝条,叶片,果肉及桃仁中都有不同含量的苦杏仁苷(赵宇瑛等, 2006 ), 桃树残根在土壤中降解产生大量的苦杏仁苷, 其水解生成的氢氰酸和苯甲醛对根系造成伤害(杨兴洪和罗新书, 1991 ), 植株残体在淹水缺氧条件下分解产生酚酸类物质(Patrick, 1971 ; Tang & Young, 1982 ).有研究发现: 地黄(Rehmannia glutinosa )连作土壤中酚酸类物质含量提高, 导致脲酶,蔗糖酶活性升高, 过氧化氢酶活性降低(陈慧等, 2007 ); 西瓜园酚酸类物质含量升高后, 土壤过氧化氢酶,脲酶活性均提高(吕卫光等, 2006 ).酚酸类物质含量增加, 导致杉木(Cunningh- amia lanceolata )苗叶绿素合成受阻, 光合作用减弱(陈秀华等, 2002 ), 对绿豆(Vigna radiata ),大豆(Glycine max ),黄瓜(Cucumis sativus )和玉米种子发芽和生长有抑制作用, 且随浓度升高抑制作用增强(曹光球等, 2003 ).酚酸类物质和苦杏仁苷都是造成果树连作障碍的重要因素(Patrick, 1955 ; Bӧrner, 1960 ).有研究表明, 棉田土壤生物活性受到作物连作障碍的负面效应和秸秆长期还田培肥地力的正面效应的双重影响(刘建国等, 2008 ).因此桃树枝条还田也可能会对桃树植株生长产生有利的影响.
目前, 桃树枝条还田对土壤酚酸类物质,苦杏仁苷,土壤微生物,土壤酶活性和桃树生长的影响尚未见报道.因此, 本试验以一年生盆栽'春美/毛桃' (Amygdalus persica "Chunmei/Maotao")为试材, 设计不同桃树枝条还田量及还田方式, 研究桃树枝条还田对土壤酚酸类物质,苦杏仁苷,土壤微生物,土壤酶活性及植株生长的影响, 以期确定合适的桃树枝条还田方式和用量, 为桃树枝条还田提供理论依据.
1 材料和方法
1.1 试验设计
2013年3月至2014年7月在山东农业大学园艺试验站进行试验.以一年生盆栽'春美/毛桃'为试材, 花盆为圆柱形, 直径32 cm, 高50 cm, 土质量约20 kg, 于3月初进行定植.自2013年5月29日起处理, 共设4个处理, T1: 每株桃树30 g枝条; 枝条剪为1-2 cm小段, 每次10 g, 分3次每周1次施入花盆中, 覆盖在土壤表面; T2: 每株桃树450 g枝条; 枝条剪为1-2 cm小段, 每次90 g, 共5次每周1次施入花盆中, 覆盖在土壤表面; T3: 每株桃树30 g枝条; 枝条剪为1-2 cm小段, 取30 g于1500 mL灌溉水中振荡3天获得浸提液, 分3次每周一次施入花盆中, 每次500 mL; T4: 每株桃树450 g枝条, 枝条剪为1-2 cm小段, 取450 g于2500 mL灌溉水中振荡3天获得浸提液, 共5次每周1次施入花盆中, 每次500 mL; 以无枝条添加为对照(CK).其中T2和T4处理每周添加90 g, 6月7日起每周监测桃树植株生长, 直到新梢生长量和叶片SPAD值与CK出现显著差异后停止添加, 此时枝条添加量为450 g.根据称量, 大田两年生桃树夏剪和冬剪总修剪量一年约为2.5 kg, 单棵桃树覆盖面积为6 m2 , 根据盆栽表面积可计算出试验盆栽桃树一年常规修剪量约为30 g, 450 g则约为15倍的常规修剪量.对各处理选9株长势基本一致的盆栽苗进行处理, 单株小区, 小区完全随机排列.每株桃树30 g换算为土壤中枝条含量为1.5 g·kg-1 , 每株桃树450 g枝条换算为土壤中枝条含量为22.5 g·kg-1 , 在全文中通用1.5 g·kg-1 和22.5 g·kg-1 表示每株桃树30 g枝条和每棵桃树450 g枝条的处理.试验处理中所添加的枝条为大田2年生'春美/毛桃'修剪所得.
1.2 取样与测定方法
1.2.1 桃树枝条中苦杏仁苷和酚酸类物质含量测定
枝条烘干磨碎过20目筛后取0.5 g, 参照索氏提取法和高效液相色谱法(叶晶晶, 2011 )测得枝条中苦杏仁苷含量; 取T4处理中枝条浸提液5 mL过0.22 μm有机相滤膜备用, 参照高效液相色谱法(尹承苗等, 2013 )测得枝条浸提液中的酚酸类物质含量.
1.2.2 土壤中酚酸类物质的测定
测定土壤酚酸所用土样为2014年6月所取, 准确称取过12目筛的风干土样80 g, 加入适量的硅藻土于烧杯中混匀.在100 mL萃取池底部垫上1片纤维素膜, 将混匀的样品装入萃取池中, 按优化好的ASE条件萃取: 提取溶剂为甲醇和无水乙醇, 提取温度为120 ℃, 提取2次, 加热5 min, 静态提取时间为5 min, 吹扫体积为60%, 吹扫90 s.萃取完成后, 34 ℃减压经RE201D型旋转蒸发仪(RE201D, 上海耀裕仪器设备有限公司, 上海)浓缩近干, 加入3 mL甲醇复溶, 过0.22 μm有机相滤膜, 待高效液相色谱(HPLC)分析.色谱条件: 色谱柱是Acclaim 120 C18 (3 μm, 150 mm × 3 mm), 柱温30 ℃.流动相是 A-乙腈, B-水(乙酸调pH值至2.6), 乙腈:水= 95:5, 流速0.5 mL·min-1 ; 进样方式及体积: 自动进样5 μL; 检测波长280 nm (尹承苗等, 2013 ).
1.2.3 土壤中苦杏仁苷的测定
测定土壤中苦杏仁苷所用土样为2014年6月所取, 准确称取过20目筛的风干土样1.0 g, 后采用索氏提取法(乙醚2 h去杂质和甲醇6 h提取)得到100 mL提取液, 用50%甲醇溶液溶解至刻度200 mL, 摇匀后超声萃取, 过0.22 μm有机相滤膜, 待高效液相色谱分析.色谱条件: 色谱柱为Acclaim 120 C18 (5 μm, 250 mm × 4.6 mm), 柱温25 ℃.流动相为乙腈:水= 17:83, 流速1.0 mL·min-1 ; 进样方式及体积: 自动进样10 μL; 检测波长210 nm (叶晶晶, 2011 ).
1.2.4 土壤微生物高通量测序和分析
2014年6月采用四分法用无菌取土器在距离桃树主干0-5 cm处取样, 深度为0-20 cm, 每处理取3株桃树, 土样立即过12目筛, 混匀, 至少10 g保存于液氮中, 通过高通量测序方法主要测定土壤细菌及真菌的种类和数量(Caporaso et al ., 2011 ; Amato et al ., 2013 ).
1.2.5 土壤酶活性的测定
采用四分法用土钻在距离桃树主干5-10 cm处随机取样, 深度0-20 cm, 每处理取3棵桃树, 共取样5次(2013年6月,7月,8月和2014年5月,6月).土壤蔗糖酶活性用3,5-二硝基水杨酸比色法测定, 37 ℃培养24 h; 土壤脲酶用苯酚钠比色法测定, 37 ℃培养24 h; 土壤过氧化氢酶活性用高锰酸钾滴定(0.1 mol·L-1 KMnO4 , 20 min) (关松荫, 1986 ).
1.2.6 植株生长指标和叶片净光合速率测定
于2014年6月7日-8月8日每15天使用卷尺测定一次桃树新梢长度(cm), 每株选取3个新梢测定, 作为一次重复, 重复5次.于2013年和2014年6月中旬, 使用游标卡尺测定地径, 即各处理距地面5 cm处桃幼树直径(mm), 单株重复, 重复5次.分别于2013年6月,7月,8月和2014年5月,6月, 各处理每株标记5片枝条中位叶, 测定叶绿素含量及叶片净光合速率, 重复5次.叶绿素含量用叶绿素仪SPAD-502 (SPAD-502, Konica Minolta, Tokyo, Japan)测定, 叶绿素仪读数与叶绿素含量成正比; 光合速率用CIRAS-3型光合仪(CIRAS-3, 汉莎科学仪器有限公司, 山东泰安)测定.
1.3 数据统计分析
试验数据采用Microsoft Excel 2003进行计算和作图, 通过DPS 7.05软件进行Duncan新复极差法数据差异性分析.
2 结果分析
2.1 桃树枝条中苦杏仁苷和酚酸类物质的含量
检测结果表明, 桃树枝条含有一定量的自毒物质.其中, 苦杏仁苷含量最高, 其次是丁香酸,苯甲酸,儿茶素(表1 ).
2.2 枝条还田对土壤酚酸类物质含量的影响
表2 为2014年6月所测桃树枝条还田后土壤中11种酚酸类物质的含量, 结果表明各处理均为没食子酸和儿茶素的含量最高.土壤总酚酸含量大小顺序为T2 > T4 > T1 > T3 > CK, 其中T2和T4处理显著高于其他处理, T1和T3处理显著高于CK.T2和T4处理除香豆酸,阿魏酸,对羟基苯甲酸外, 其余8种酚酸含量均显著提高.与对照相比, 各处理土壤酚酸类物质总量分别提高了11.9%,32.0%,13.0%和29.4%.可见, 加量枝条浸提液处理和枝条剪碎处理均增加了土壤酚酸类物质含量.
2.3 枝条还田对土壤苦杏仁苷含量的影响
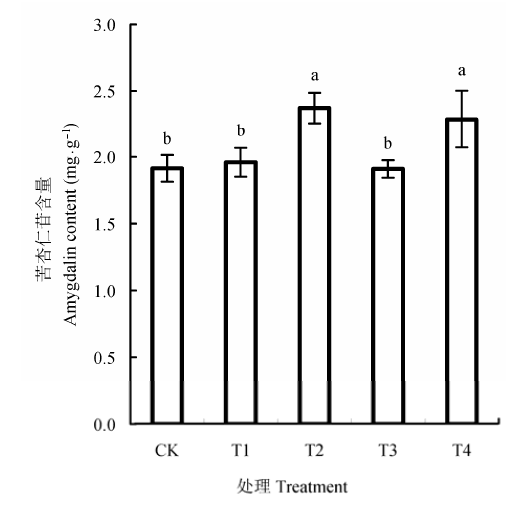
2014年6月, T1和T3处理土壤的苦杏仁苷含量与对照无显著差异, T2和T4处理均显著增加了土壤中苦杏仁苷的含量(图1 ).
图1 枝条还田对桃树土壤苦杏仁苷含量的影响(平均值±标准偏差).CK, 无枝条添加对照; T1,T2,T3,T4分别代表1 .5 g·kg-1 枝条剪碎还田处理,22.5 g·kg-1 枝条剪碎还田处理,1.5 g·kg-1 枝条浸提液还田处理,22.5 g·kg-1 枝条浸提液还田处理.图中不同字母表示处理间显著性差异达5%显著水平.
Fig. 1 Effects of branch returning on the amygdalin content of soils under peach tree (means ± SD ). CK, no branch addition as control; T1, T2, T3 and T4 represent treatment of coverage by fragmented peach tree branches in 1.5 g·kg-1 and 22.5 g·kg-1 , and treatment of peach tree branch leachate application of 1.5 g·kg-1 and 22.5 g·kg-1 , respectively. Different letters in the figure denote significant difference at a level of p < 0.05.
2.4 加量枝条还田对土壤微生物的影响
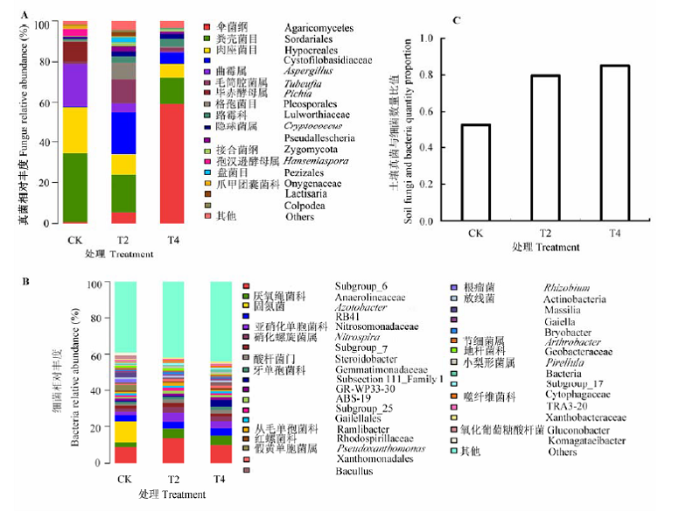
由图2A 可以看出, 土壤真菌主要由伞菌纲,粪壳菌纲粪壳菌目和肉座菌目,异担子菌纲银耳亚纲Cystofilobasidiaceae,不整子囊菌纲曲霉属(Asper- gillus ),座囊菌纲毛筒腔菌属(Tubeufia ),半子囊菌纲毕赤酵母属(Pichia )和座囊菌纲格孢菌目组成.加量枝条还田后, T2和T4处理土壤中粪壳菌目,肉座菌目,曲霉属和毕赤酵母属相对丰度低于CK; 伞菌纲相对丰度高于CK, 且T4处理远远大于T2处理; T2处理中Cystofilobasidiaceae和毛筒腔菌属的相对丰度大于T4处理, 远远大于CK.
由图2B 可以看出, 土壤细菌主要由Subgroup_ 6, 厌氧绳菌科,固氮菌,RB41, 亚硝化单胞菌科,硝化螺旋菌科硝化螺旋菌属(Nitrospira )组成.加量枝条还田后, T2和T4处理土壤中没有检测到固氮菌.土壤中固氮菌吸取氮转化为氮肥供植株生长使用, T2和T4处理土壤中固氮菌的缺失不利于幼龄桃树生长.
图2C 为土壤真菌与细菌数量比例, T2和T4处理真菌比例明显升高, 细菌比例下降, 土壤由细菌型向真菌型转化.
图2 加量枝条还田对土壤真菌(A),细菌(B)以及两者比例(C)的影响.CK, 无枝条添加对照; T2,T4分别代表22 .5 g·kg-1 枝条剪碎还田处理和22.5 g·kg-1 枝条浸提液还田处理.
Fig. 2 Effects of high-quantity returning treatment on soil fungi (A), bacteria (B) and the fungi:bacteria ratio (C). CK, no branches addition as control; T2 and T4 represent branch fragment application and branch leachate application in 22.5 g·kg-1 , respectively.
2.5 枝条还田对土壤基础酶活性的影响
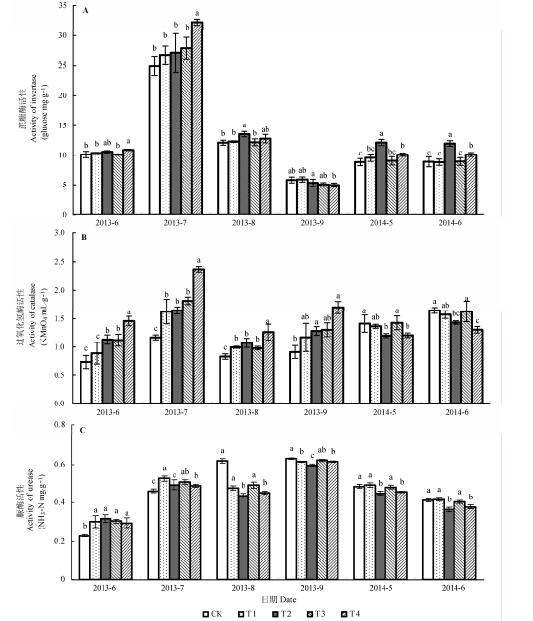
由图3A 可以看出, 土壤蔗糖酶活性在2013年7月显著高于其他月.2013年7,8月T4处理显著高于CK, 其他处理略高于CK, 6月和9月各处理间差异不显著.而2014年5,6月T2处理酶活性最高, T4处理次之, T1和T3处理与CK无显著差异.T4处理比T2处理更早表现出促进作用, 这是因为T4处理是枝条浸提液处理.这一结果说明, 土壤中酚酸类物质和苦杏仁苷达到一定浓度时提高了土壤中蔗糖酶的活性.
由图3B 可以看出, 2013年枝条还田处理土壤过氧化氢酶活性均显著高于CK, 其中以T4处理最为显著.2014年5,6月, T2和T4处理土壤过氧化氢酶活性最低, 而T1和T3处理与CK无显著差异.表明桃树枝条中酚酸类物质和苦杏仁苷进入土壤后浓度不断累积, 而这两类物质对过氧化氢酶活性表现出低浓度促进,高浓度抑制.T2和T4处理中土壤过氧化氢酶活性减弱, 不利于消除土壤中过量的过氧化氢, 从而对桃幼树生长产生负面影响.
由图3C 可以看出, 2013年6,7月, 各处理土壤脲酶活性高于CK但无显著差异, 而8月各处理脲酶活性显著低于CK.2014年5,6月, T2和T4处理土壤脲酶活性显著低于CK, T1和T3处理与CK无显著差异.这说明桃树枝条中酚酸类物质和苦杏仁苷进入土壤, 浓度累积, 而这两类物质对脲酶活性的影响与过氧化氢酶活性类似, 表现为低促高抑.土壤脲酶活性与土壤尿素供应能力,有机质含量,微生物数量关系密切, T2和T4处理降低了土壤脲酶的活性, 可能对桃幼树的生长有很大的影响.
图3 枝条还田对桃树土壤蔗糖酶(A),过氧化氢酶(B),脲酶(C)活性的影响(平均值±标准偏差).CK, 无枝条添加对照; T1,T2,T3,T4分别代表1 .5 g·kg-1 枝条剪碎还田处理,22.5 g·kg-1 枝条剪碎还田处理,1.5 g·kg-1 枝条浸提液还田处理,22.5 g·kg-1 枝条浸提液还田处理.不同字母表示处理间显著性差异达5%显著水平.
Fig. 3 Effects of branch returning on the soil invertase (A), catalase (B) and urease (C) activity (means ± SD ). CK, no branches addition as control; T1, T2, T3 and T4 represent treatment of coverage by fragmented peach tree branches in 1.5 g·kg-1 and 22.5 g·kg-1 , and treatment of peach tree branch leachate application of 1.5 g·kg-1 and 22.5 g·kg-1 , respectively. Different letters in the figure denote significant difference at a level of p < 0.05.
2.6 枝条还田对桃树净光合速率的影响
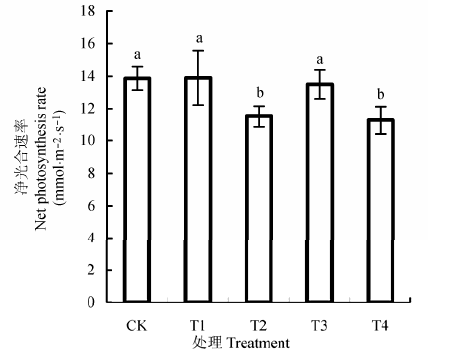
由图4 可以看出, 2014年6月中旬, T1,T3处理桃树叶片净光合速率与对照无显著差异, 而T2和T4处理显著降低了桃幼树叶片的净光合速率, 分别降低了16.67%和18.84%.可见, 加量枝条处理中酚酸类物质和苦杏仁苷进入土壤累积, 改变了土壤微生物结构和土壤酶活性, 可能进一步抑制了植株叶片的净光合速率.
图4 枝条还田对桃树叶片净光合速率的影响(平均值±标准偏差).CK, 无枝条添加对照; T1,T2,T3,T4分别代表1 .5 g·kg-1 枝条剪碎还田处理,22.5 g·kg-1 枝条剪碎还田处理,1.5 g·kg-1 枝条浸提液还田处理,22.5 g·kg-1 枝条浸提液还田处理.图中不同字母表示处理间显著性差异达5%显著水平.
Fig. 4 Effects of branch returning on the net photosynthesis rate of peach trees (means ± SD ). CK, no branches addition as control; T1, T2, T3 and T4 represent treatment of coverage by fragmented peach tree branches in 1.5 g·kg-1 and 22.5 g·kg-1 , and treatment of peach tree branch leachate application of 1.5 g·kg-1 and 22.5 g·kg-1 , respectively. Different letters in the figure denote significant difference at a level of p < 0.05.
2.7 枝条还田对桃幼树营养生长的影响
由表3 和表4 可以看出, T1和T3处理新梢生长量与CK无显著差异.与CK相比, T2和T4处理在7月初表现出新梢生长缓慢的现象, 8月新梢生长几乎停滞.2013年6月底, SPAD叶绿素仪测得T4处理叶片SPAD值显著低于其他处理, 7月,8月各处理间差异不显著.2014年5月和6月, T1和T3叶片SPAD值与CK无显著差异, T2和T4处理与CK相比均显著降低.2013年6月, 各处理间桃幼树地径无显著差异; 2014年6月, T1和T3处理桃幼树地径与CK无显著差异; 与对照相比, T2和T4处理地径分别减少10.39%和9.72%, 差异显著.可见, T2和T4处理抑制了桃幼树营养生长.
3 讨论
前人研究结果显示, 连作或秸秆还田的土壤中, 土壤酚酸类物质有明显提高.麦秸还田40天后, 玉米土壤耕作层酚酸含量达到最高峰(郑皓皓等, 2001 ; 胡晓军等, 2011 ), 花生(Arachis hypogaea )和黄瓜连作后土壤酚酸含量均显著升高(马云华等, 2005 ; 李培栋等, 2010 ), 连作苹果园土壤中酚酸物质总量显著提高(王青青, 2012 ).与其他作物不同, 桃树残根降解会产生苦杏仁苷, 经土壤酶分解产生的氢氰酸,苯甲醛和苯甲酸, 对新茬桃树根系产生巨大毒害(Baziramakenga et al ., 1995 ).本试验结果表明, 枝条浸提液中含有酚酸类物质和高量的苦杏仁苷, 加量枝条还田后, 土壤中酚酸类物质和苦杏仁苷含量与对照相比分别提高约30%和20%, 加量桃树枝条(22.5 g·kg-1 )的加入同样增加自毒物质的累积.因为盆栽桃树添加枝条后, 露天放置, 剪碎枝条经过雨水和灌溉水浸泡后同样会产生大量此类物质, 这与有关研究中植株残体淹水缺氧可分解产生挥发性脂肪酸和酚酸类(Patrick, 1971 ; Tang & Young, 1982 )的结果是一致的.
土壤微生物数量多, 种类多, 参与土壤有机质分解,养分转化等过程, 对植株生长发育有重要影响.研究发现, 连作障碍能够影响土壤中细菌和真菌的比例, 如连作棉田秸秆还田5-10年连作障碍明显, 细菌放线菌数量下降, 真菌比例提高(刘建国等, 2008 ; 刘素慧等, 2010 ), 马铃薯(Solanum tuberosum )连作后, 土壤由细菌型向真菌型转化(马琨等, 2010 ).马铃薯连作土壤中主要土传病原真菌为镰孢菌(Fusariun ),轮枝菌(Verticillium )和立枯丝核菌(Rhizoctonia solani )(李继平等, 2013 ), 黄瓜连作土壤真菌优势菌群主要是丝孢纲,外生菌根真菌,接合菌纲,伞菌纲,担子菌纲(乔蓬蕾等, 2014 ).本试验结果表明, 加量枝条处理后, 盆栽桃树土壤中真菌粪壳菌目,肉座菌目,曲霉属和比赤酵母减少, 伞菌纲,银耳亚纲Cystofilobasidiaceae和毛筒腔菌属增多, 这7种真菌均属于子囊菌门和担子菌门, 由此推断抑制桃树生长的优势真菌为子囊菌门和担子菌门.伞菌纲的最大功能是腐化植物残体, 它的增多加速了土壤中枝条的腐化.加量枝条处理后, 土壤中固氮菌缺失, 土壤中固氮菌能够吸取氮转化为氮肥供植株生长使用, 固氮菌的缺失不利于桃幼树生长.加量枝条处理后土壤中真菌比例升高, 是地力衰竭的标志(Larkin, 2003 ; 林茂兹等, 2012 ).桃园连作可能是自毒物质直接进入植株或通过影响土壤微生物(李小蓉和李蕾, 2013 )改变土壤环境, 进而影响植株生长引起的.
土壤酶活性是研究土壤质量的重要指标.在小麦-玉米轮作情况下, 秸秆还田提高了土壤酶活性(路文涛等, 2011 ; 甄丽莎等, 2012 ).杨树林土壤酚酸浓度升高后, 土壤脲酶和碱性磷酸酶活性降低, 过氧化氢酶活性则升高(王延平等, 2013 ).肖宏(2004)研究发现连作苹果土壤中酚酸类物质含量升高后, 基础酶活性降低.本试验结果表明, T1和T3处理的土壤酶活性与CK无显著差异.T2和T4处理的土壤蔗糖酶活性始终高于CK.处理前期, 土壤过氧化氢酶和脲酶活性均显著高于CK, 处理后期均显著低于CK.蔗糖酶又称转化酶, 与土壤有机质含量呈正相关, 可能是因为枝条腐解产生有机质, 引起蔗糖酶活性升高.脲酶促进尿素氮肥转化成氨, 过氧化氢酶可以清除土壤中的过氧化氢.处理后期脲酶和过氧化氢酶活性降低, 减少了植株的直接氮素来源, 造成土壤中过氧化氢的积累而不利于桃树植株的 生长.
前人研究结果显示, 土壤自毒物质对不同植株表现出类似的抑制作用.外源添加肉桂酸,苯甲酸,对羟基苯甲酸, 抑制了杉木幼苗胚根和胚芽的生长(汪思龙等, 2002 ).土壤酚酸含量增加, 玉米幼苗的株高和根系生长均受到抑制(郑皓皓等, 2001 ; 胡晓军等, 2011 ).对羟基苯甲酸,香草酸和香豆酸的累积能够抑制花生幼苗生长, 提高花生发病率(李培栋等, 2010 ).桃树根系浸提液对桃树生长指标表现为低浓度促进,高浓度抑制的质量浓度效应(张斌斌等, 2012 ).本试验结果表明, 常规修剪量(1.5 g·kg-1 )还田对桃幼树生长无显著影响, 而加量枝条(22.5 g·kg-1 )还田时, 土壤酚酸总量达到1700 ng·g-1 , 土壤苦杏仁苷含量2.30 mg·g-1 , 出现了显著抑制现象, 如叶绿素含量降低,新梢停长期提前,地径生长量减小和净光合速率降低.
常规量枝条添加与对照无显著差异, 而加量枝条的加入为盆栽土壤提供了大量的苦杏仁苷和酚酸类自毒物质, 自毒物质直接抑制桃幼树生长(Patrick, 1955 ), 同时能够通过间接途径改变土壤微生物结构和土壤基础酶活性, 引起地下部分变化而调控桃幼树地上部分生长, 导致叶绿素含量降低, 新梢停长期提前, 地径生长减缓, 叶片净光合速率降低等现象.但究竟是酚酸类物质还是苦杏仁苷在这个过程中起关键作用仍有待进一步研究.大田两年桃树枝条(2012年12月至2014年7月)还田试验表明, 与对照相比, 常规修剪量枝条还田处理, 土壤自毒物质,微生物,酶活性及桃树生长指标均无显著差异.初步推测, 常规修剪量枝条还田对植株生长没有不利影响; 但加量枝条集中还田, 会对桃树生长产生抑制作用.
致谢 本研究得到山东省高校优秀科研创新团队计划项目资助, 特此致谢.
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Amato KR Yeoman CJ Kent A Righini N Carbonero F Estrada A Gaskins HR Stumpf RM Yildirim S Torralba M Gillis M Wilson BA Nelson KE White BA Leigh SR 2013 ). Habitat degradation impacts black howler monkey (Alouatta pigra) gastrointestinal microbiomes.
International Society for Microbial Ecology , 7 , 1344 -1353 .
[本文引用: 1]
[2]
Baziramakenga R Leroux GD Simard RR 1995 ). Effects of benzoic and cinnamic acids on membrane permeability of soybean roots.
Journal of Chemical Ecology , 21 , 1271 -1285 .
[本文引用: 1]
[3]
Börner H 1960 ). Liberation of organic substances from higher plants and their role in the soil sickness problem.
Botanical Review , 26 , 393 -424 .
[本文引用: 1]
[4]
Cao GQ Lin SZ Du L Lin GL Liu Y 2003 ). The bioassay of ferulic acid and cinnamic acid allelopathic to Chinese fir.
Chinese Journal of Eco-Agriculture , 11 (2 ), 8 -10 .(in Chinese with English abstract) [曹光球 , 林思祖 , 杜玲 , 林桂莲 , 刘雁 (2003 ). 阿魏酸与肉桂酸对杉木化感作用的生物评价
. 中国生态农业学报 ,11 (2 )8 -10 .]
[本文引用: 1]
[5]
Caporaso JG Lauber CL Walters WA Berg-Lyons D Lozupone CA Turnbaugh PJ Fierer N Knight R 2011 ). Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample.
Proceedings of the National Academy of Sciences of the United States of America , 108 (Suppl. 1 ), 4516 -4522 .
[本文引用: 1]
[6]
Chen H Hao HR Xiong J Qi XH Zhang ZY Lin WX 2007 ). Effects of successive cropping Rehmannia glutinosa on rhizosphere soil microbial flora and enzyme activities.
Chinese Journal of Applied Ecology , 18 , 2755 -2759 .(in Chinese with English abstract) [陈慧 , 郝慧荣 , 熊君 , 齐晓辉 , 张重义 , 林文雄 (2007 ). 地黄连作对根际微生物区系及土壤酶活性的影响
. 应用生态学报 ,18 , 2755 -2759 .]
[本文引用: 1]
[7]
Chen XH Li CH He SJ 2002 ). Preliminary study on phenolic acids movement in Soil-Chinese fir seedlings system.
Journal of Huazhong Agricultural University , 21 , 235 -237 .(in Chinese with English abstract) [陈秀华 , 李传涵 , 何绍江 (2002 ). 酚酸在土壤--杉木苗间运移的初步探讨
. 华中农业大学学报 ,21 , 235 -237 .]
[本文引用: 1]
[8]
Desmond RL Daniele B 2008). The Peach: Botany , Production and Uses . CABI Publishing Press , London . 4 -6 .
[本文引用: 1]
[9]
Guan SY Soil Enzyme and Its Methodology .Agriculture Press , Beijing . 260 -360 .(in Chinese) [关松荫 (1986). 土壤酶及其研究法 . 中国农业出版社 , 北京 . 260 -360 .]
[本文引用: 1]
[10]
Han XZ Zhu LQ Yang MF Yu Q Bian XM 2012 ). Effects of different amount of wheat straw returning on rice growth, soil microbial biomass and enzyme activity.
Journal of Agro-Environment Science , 31 , 2129 -2199 .(in Chinese with English abstract) [韩新忠 , 朱利群 , 杨敏芳 , 俞琦 , 卞新民 (2012 ). 不同小麦秸秆还田量对水稻生长,土壤微生物生物量及酶活性的影响
. 农业环境科学学报 , 31 , 2129 -2199 .]
[本文引用: 1]
[11]
Hu XJ Zheng HH Jia JY Wu E Xing JJ Zhang KY 2011 ). Temporal change of the phenolic acids in soil with returning wheat residues and their effects on the seedling growth and the yield of summer corn.
Chinese Journal of Ecology , 20 (3 ), 9 -11 .(in Chinese with English abstract) [胡晓军 , 郑皓皓 , 贾敬业 , 吴萼 , 邢建军 , 张克银 (2011 ). 麦秸还田耕层酚酸的时间变化及其对夏玉米幼苗生长和产量的影响
. 生态学杂志 , 20 (3 ), 9 -11 .]
[本文引用: 2]
[12]
Larkin RP 2003 ). Characterization of soil microbial communities under different potato cropping systems by microbial population dynamics, substrate utilization, and fatty acid profiles.
Soil Biology and Biochemistry , 35 , 1451 -1466 .
[本文引用: 1]
[13]
Li JP Li MQ Hui NN Wang L Ma YQ Qi YH 2013 ). Population dynamics of main fungal pathogens in soil of continuously cropped potato.
Acta Prataculturae Sinica , 22 (4 ), 147 -152 .(in Chinese with English abstract) [李继平 , 李敏权 , 惠娜娜 , 王立 , 马永强 , 漆永红 (2013 ). 马铃薯连作田土壤中主要病原真菌的种群动态变化规律
. 草业学报 ,22 (4 ), 147 -152 .]
[本文引用: 1]
[14]
Li PD Wang XX Li YL Wang HW Liang FY Dai CC 2010 ). The contents of phenolic acids in continuous cropping peanut and their allelopathy.
Acta Ecologica Sinica , 20 , 2128 -2134 .(in Chinese with English abstract) [李培栋 , 王兴祥 , 李奕林 , 王宏伟 , 梁飞燕 , 戴传超 (2010 ). 连作花生土壤中酚酸类物质的检测及其对花生的化感作用
. 生态学报 ,20 , 2128 -2134 .]
[本文引用: 2]
[15]
Li XR Li L 2013 ). Review of the advances on plant allelochemicals and soil microorganisms.
Guangdong Agricultural Sciences , 40 , 178 -181 .(in Chinese with English abstract) [李小蓉 , 李蕾 (2013 ). 植物化感物质与土壤微生物的研究进展
. 广东农业科学 , 40 , 178 -181 .]
[本文引用: 1]
[16]
Lin MZ Wang HB Lin HF 2012 ). Effects of Pseudostellariae heterophylla continuous cropping on rhizosphere soil microorganisms.
Chinese Journal of Ecology , 31 , 106 -111 .(in Chinese with English abstract) [林茂兹 , 王海斌 , 林辉锋 (2012 ). 太子参连作对根际土壤微生物的影响
. 生态学杂志 ,31 , 106 -111 .]
[本文引用: 1]
[17]
Liu JB Zhang ZY Liu ZJ 2006 ). Peach continuous cropping disease and its prevention and control technology.
The Journal of Hebei Forestry Science and Technology , (2 ), 67 .(in Chinese) [刘嘉彬 , 张泽勇 , 刘振京 (2006 ). 桃树重茬病及其防治技术
. 河北林业科技 ,(2 ), 67 .]
[本文引用: 1]
[18]
Liu JG Bian XM Li YB Zhang W Li S 2008 ). Effects of long-term continuous cropping of cotton and returning cotton stalk into field on soil biological activities.
Chinese Journal of Applied Ecology , 19 , 1027 -1032 .(in Chinese with English abstract) [刘建国 , 卞新民 , 李彦斌 , 张伟 , 李崧 (2008 ). 长期连作和秸秆还田对棉田土壤生物活性的影响
. 应用生态学报 , (19 ), 1027 -1032 .]
[本文引用: 2]
[19]
Liu SH Liu SQ Zhang ZK Wei H Qi JJ Duan JF 2010 ). Influence of garlic continuous cropping on rhizosphere soil microorganisms and enzyme activities.
Scientia Agricultura Sinica , 43 , 1000 -1006 .(in Chinese with English abstract) [刘素慧 , 刘世琦 , 张自坤 , 尉辉 , 齐建建 , 段吉锋 (2010 ). 大蒜连作对其根际土壤微生物和酶活性的影响
. 中国农业科学 , 43 , 1000 -1006 .]
[本文引用: 1]
[20]
Lü WG Shen QR Yu TY Zhu HT 2006 ). The effect of added phenolic acids on soil enzyme activities and nutrients.
Plant Nutrition and Fertilizer Science , 12 , 845 -849 .(in Chinese with English abstract) [吕卫光 , 沈其荣 , 余廷园 , 诸海涛 (2006 ). 酚酸化合物对土壤酶活性和土壤养分的影响
. 植物营养与肥料学报 , 12 , 845 -849 .]
[本文引用: 1]
[21]
Lu WT Jia ZK Zhang P Wang W Hou XQ Yang BP Li YP 2011 ). Effects of straw returning on soil labile organic carbon and enzyme activity in semi-arid areas of southern Ningxia, China.
Journal of Agro-Environment Science , 30 , 522 -528 .(in Chinese with English abstract) [路文涛 , 贾志宽 , 张鹏 , 王维 , 侯贤清 , 杨保平 , 李永平 (2011 ). 秸秆还田对宁南旱作农田土壤活性有机碳及酶活性的影响
. 农业环境科学学报 , 30 , 522 -528 .]
[本文引用: 1]
[22]
Ma K Zhang L Du Q Song NL 2010 ). Effect of potato continuous cropping on soil microorganism community structure and function.
Journal of Soil and Water Conservation , 24 , 229 -233 .[马琨 , 张丽 , 杜茜 , 宋乃平 (2010 ). 马铃薯连作栽培对土壤微生物群落的影响
. 水土保持学报 ,24 , 229 -233 .]
[本文引用: 1]
[23]
Ma YH Wang XF Wei M Qi YF Li TL 2005 ). Accumulation of phenolic acids in continuously cropped cucumber soil and their effects on soil microbes and enzyme activities.
Chinese Journal of Applied Ecology , 16 , 2149 -2153 .[马云华 , 王秀峰 , 魏珉 , 亓延凤 , 李天来 (2005 ). 黄瓜连作土壤酚酸类物质积累对土壤微生物和酶活性的影响
. 应用生态学报 , 16 , 2149 -2153 .]
[本文引用: 1]
[24]
Mu P Zhang EH Wang HN Fang YF 2012 ). Effects of continuous straw return to soil on maize growth and soil chemical and physical characteristics.
Chinese Journal of Eco-Agriculture , 20 , 291 -296 .(in Chinese with English abstract) [慕平 , 张恩和 , 王汉宁 , 方永丰 (2012 ). 不同年限全量玉米秸秆还田对玉米生长发育及土壤理化性状的影响
. 中国生态农业学报 , 20 , 291 -296 .]
[本文引用: 1]
[25]
Patrick ZA 1955 ). The peach replant problem in Ontario: II Toxic substances from microbial decomposition products of peach root residues.
Canadian Journal of Botany , 33 , 461 -486 .
[本文引用: 2]
[26]
Patrick ZA 1971 ). Phytotoxic substances associated with the decomposition in soil of plant residues.
Soil Science , 111 , 13 -18 .
[本文引用: 2]
[27]
Qiao PL Zhou XG Wu FZ 2014 ). T-RFLP analysis of cucumber rhizosphere microbial communities in different cropping seasons.
Chinese Journal of Ecology , 33 , 2640 -2649 .(in Chinese with English abstract) [乔蓬蕾 , 周新刚 , 吴凤芝 (2014 ). 不同连作茬次黄瓜根际土壤微生物群落的T-RFLP分析
. 生态学杂志 , 33 , 2640 -2649 .]
[本文引用: 1]
[28]
Tang CS Young CC 1982 ). Collection and identification of allelopathic compounds from the undisturbed root system of bigalta limpograss (Hemarthria altissima).
Plant Physiology , 69 , 155 -160 .
[本文引用: 2]
[29]
Wang QQ 2012 ). Variations of Phenolic Acids in Soil of Apple Orchards and the Effect of Phloridzin on the Tricarboxylic Acid Cycle. Master degree dissertation, Shandong Agricultural University
, Taian. (in Chinese with English abstract) [王青青 (2012 ). 苹果园土壤酚酸类物质变化及根皮苷对TCA循环影响的研究
. 硕士学位论文, 山东农业大学, 泰安. ]
[本文引用: 1]
[30]
Wang SL Chen LC Liao LP Huang ZQ 2002 ). Effects of three kinds of allelochemicals on growth of Chinese fir seedlings.
Chinese Journal of Applied & Environmental Biology , 8 , 588 -591 .(in Chinese with English abstract) [汪思龙 , 陈龙池 , 廖利平 , 黄志群 (2002 ). 几种化感物质对杉木幼苗生长的影响
. 应用与环境生物学报 , 8 , 588 -591 .]
[本文引用: 1]
[31]
Wang YP Wang HT Xu T Ni GP Jiang YZ 2013 ). Effects of exogenous phenolic acid on soil nutrient availability and enzyme activities in a poplar plantation.
Chinese Journal of Applied Ecology , 24 , 667 -674 .(in Chinese with English abstract) [王延平 , 王华田 , 许坛 , 倪桂萍 , 姜岳忠 (2013 ). 酚酸对杨树人工林土壤养分有效性及酶活性的影响
. 应用生态学报 , 24 , 667 -674 .]
[本文引用: 1]
[32]
Xiao H 2004 ). Effect of Soil Pasteurization and Rotation on Apple Replant Problem. Master degree dissertation, Shandong Agricultural University
, Taian. (in Chinese) [肖宏 (2004 ). 土壤消毒和轮作对克服苹果连作障碍效果的研究
. 硕士学位论文, 山东农业大学, 泰安.]
[33]
Yang XH Luo XS 1991 ). The research progress of fruit tree problem of replantation.
Journal of Fruit Science , 8 , 239 -244 .(in Chinese) [杨兴洪 , 罗新书 (1991 ). 果树再植问题研究进展
. 果树科学 , 8 , 239 -244 .]
[本文引用: 1]
[34]
Ye JJ 2011 ). Determination of amygdal in Prunus persical (L.) Batsch produced in different areas by HPLC.
Chinese Archives of Traditional Chinese Medicine , 29 , 206 -207 .(in Chinese with English abstract) [叶晶晶 (2011 ). HPLC法测定不同产地桃仁中苦杏仁苷的含量
. 中华中医药学刊 , 29 , 206 -207 .]
[本文引用: 2]
[35]
Yin CM Wang GS Li YY Che JS Shen X Chen XS Mao ZQ Wu SJ 2013 ). A new method for analysis of phenolic acids in the soil--Soil from replanted apple orchards was investigated.
Scientia Agricultura Sinica , 46 , 4612 -4619 .(in Chinese with English abstract) [尹承苗 , 王功帅 , 李园园 , 车金水 , 沈向 , 陈学森 , 毛志泉 , 吴树敬 (2013 ). 一种分析土壤中酚酸类物质含量的新方法--以连作苹果园土壤为试材
. 中国农业科学 ,46 , 4612 -4619 .]
[本文引用: 2]
[36]
Zhang BB Ma RJ Cai ZX Song HF Shen JH 2012 ). Effect of continuous cultivation on growth and leaf traits of replanted peach seedling. Southwest China Journal of Agricultural Sciences
, 25 , 1388 -1392 .(in Chinese with English abstract) [张斌斌 , 马瑞娟 , 蔡志翔 , 宋宏峰 , 沈江海 (2012 ). 连作对再植桃树幼苗生长及叶片性状的影响
. 西南农业学报 , 25 , 1388 -1392 .]
[本文引用: 1]
[37]
Zhao YY Cheng LQ Shang B 2006 ). Studies on yearly change of amygdalin content in peach. Journal of Yangtze University (
Natural Science Edition ), 3 , 137 -138, 141 .(in Chinese with English abstract) [赵宇瑛 , 程丽琴 , 尚冰 (2006 ). 桃树体内苦杏仁甙含量年变化研究
. 长江大学学报(自然版) , 3 , 137 -138, 141 .]
[本文引用: 1]
[38]
Zhen LS Gu J Gao H Qin QJ Chen QL 2012 ). Effect of straws, manure and chemical fertilizer on soil properties and crop yields.
Acta Botanica Boreali-Occidentalia Sinica , 32 , 1811 -1818 .(in Chinese with English abstract) [甄丽莎 , 谷洁 , 高华 , 秦清军 , 陈强龙 (2012 ). 秸秆还田与施肥对土壤酶活性和作物产量的影响
. 西北植物学报 , 32 ,1811 -1818 .]
[本文引用: 1]
[39]
Zheng HH Hu XJ Jia JY Wu E Xing JJ 2001 ). Changes of the phenolic acid in plough layer and its effects on the growth and yield of summer corn with returning wheat straw.
Eco-Agriculture Research , 9 (4 ), 79 -81 .(in Chinese with English abstract) [郑皓皓 , 胡晓军 , 贾敬业 , 吴萼 , 邢建军 (2001 ). 麦秸还田耕层酚酸变化及其对夏玉米生长的影响
. 中国生态农业学报 ,9 (4 ), 79 -81 .]
[本文引用: 2]
Habitat degradation impacts black howler monkey (Alouatta pigra) gastrointestinal microbiomes.
1
2013
... 2014年6月采用四分法用无菌取土器在距离桃树主干0-5 cm处取样, 深度为0-20 cm, 每处理取3株桃树, 土样立即过12目筛, 混匀, 至少10 g保存于液氮中, 通过高通量测序方法主要测定土壤细菌及真菌的种类和数量(Caporaso et al ., 2011 ; Amato et al ., 2013 ). ...
Effects of benzoic and cinnamic acids on membrane permeability of soybean roots.
1
1995
... 前人研究结果显示, 连作或秸秆还田的土壤中, 土壤酚酸类物质有明显提高.麦秸还田40天后, 玉米土壤耕作层酚酸含量达到最高峰(郑皓皓等, 2001 ; 胡晓军等, 2011 ), 花生(Arachis hypogaea )和黄瓜连作后土壤酚酸含量均显著升高(马云华等, 2005 ; 李培栋等, 2010 ), 连作苹果园土壤中酚酸物质总量显著提高(王青青, 2012 ).与其他作物不同, 桃树残根降解会产生苦杏仁苷, 经土壤酶分解产生的氢氰酸,苯甲醛和苯甲酸, 对新茬桃树根系产生巨大毒害(Baziramakenga et al ., 1995 ).本试验结果表明, 枝条浸提液中含有酚酸类物质和高量的苦杏仁苷, 加量枝条还田后, 土壤中酚酸类物质和苦杏仁苷含量与对照相比分别提高约30%和20%, 加量桃树枝条(22.5 g·kg-1 )的加入同样增加自毒物质的累积.因为盆栽桃树添加枝条后, 露天放置, 剪碎枝条经过雨水和灌溉水浸泡后同样会产生大量此类物质, 这与有关研究中植株残体淹水缺氧可分解产生挥发性脂肪酸和酚酸类(Patrick, 1971 ; Tang & Young, 1982 )的结果是一致的. ...
Liberation of organic substances from higher plants and their role in the soil sickness problem.
1
1960
... 桃树再植当年基本不生新根, 发枝短长势弱, 造成桃园减产, 连作障碍严重(刘嘉彬等, 2006 ).桃树根系,枝条,叶片,果肉及桃仁中都有不同含量的苦杏仁苷(赵宇瑛等, 2006 ), 桃树残根在土壤中降解产生大量的苦杏仁苷, 其水解生成的氢氰酸和苯甲醛对根系造成伤害(杨兴洪和罗新书, 1991 ), 植株残体在淹水缺氧条件下分解产生酚酸类物质(Patrick, 1971 ; Tang & Young, 1982 ).有研究发现: 地黄(Rehmannia glutinosa )连作土壤中酚酸类物质含量提高, 导致脲酶,蔗糖酶活性升高, 过氧化氢酶活性降低(陈慧等, 2007 ); 西瓜园酚酸类物质含量升高后, 土壤过氧化氢酶,脲酶活性均提高(吕卫光等, 2006 ).酚酸类物质含量增加, 导致杉木(Cunningh- amia lanceolata )苗叶绿素合成受阻, 光合作用减弱(陈秀华等, 2002 ), 对绿豆(Vigna radiata ),大豆(Glycine max ),黄瓜(Cucumis sativus )和玉米种子发芽和生长有抑制作用, 且随浓度升高抑制作用增强(曹光球等, 2003 ).酚酸类物质和苦杏仁苷都是造成果树连作障碍的重要因素(Patrick, 1955 ; Bӧrner, 1960 ).有研究表明, 棉田土壤生物活性受到作物连作障碍的负面效应和秸秆长期还田培肥地力的正面效应的双重影响(刘建国等, 2008 ).因此桃树枝条还田也可能会对桃树植株生长产生有利的影响. ...
阿魏酸与肉桂酸对杉木化感作用的生物评价
1
2003
... 桃树再植当年基本不生新根, 发枝短长势弱, 造成桃园减产, 连作障碍严重(刘嘉彬等, 2006 ).桃树根系,枝条,叶片,果肉及桃仁中都有不同含量的苦杏仁苷(赵宇瑛等, 2006 ), 桃树残根在土壤中降解产生大量的苦杏仁苷, 其水解生成的氢氰酸和苯甲醛对根系造成伤害(杨兴洪和罗新书, 1991 ), 植株残体在淹水缺氧条件下分解产生酚酸类物质(Patrick, 1971 ; Tang & Young, 1982 ).有研究发现: 地黄(Rehmannia glutinosa )连作土壤中酚酸类物质含量提高, 导致脲酶,蔗糖酶活性升高, 过氧化氢酶活性降低(陈慧等, 2007 ); 西瓜园酚酸类物质含量升高后, 土壤过氧化氢酶,脲酶活性均提高(吕卫光等, 2006 ).酚酸类物质含量增加, 导致杉木(Cunningh- amia lanceolata )苗叶绿素合成受阻, 光合作用减弱(陈秀华等, 2002 ), 对绿豆(Vigna radiata ),大豆(Glycine max ),黄瓜(Cucumis sativus )和玉米种子发芽和生长有抑制作用, 且随浓度升高抑制作用增强(曹光球等, 2003 ).酚酸类物质和苦杏仁苷都是造成果树连作障碍的重要因素(Patrick, 1955 ; Bӧrner, 1960 ).有研究表明, 棉田土壤生物活性受到作物连作障碍的负面效应和秸秆长期还田培肥地力的正面效应的双重影响(刘建国等, 2008 ).因此桃树枝条还田也可能会对桃树植株生长产生有利的影响. ...
Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample.
1
2011
... 2014年6月采用四分法用无菌取土器在距离桃树主干0-5 cm处取样, 深度为0-20 cm, 每处理取3株桃树, 土样立即过12目筛, 混匀, 至少10 g保存于液氮中, 通过高通量测序方法主要测定土壤细菌及真菌的种类和数量(Caporaso et al ., 2011 ; Amato et al ., 2013 ). ...
地黄连作对根际微生物区系及土壤酶活性的影响
1
2007
... 桃树再植当年基本不生新根, 发枝短长势弱, 造成桃园减产, 连作障碍严重(刘嘉彬等, 2006 ).桃树根系,枝条,叶片,果肉及桃仁中都有不同含量的苦杏仁苷(赵宇瑛等, 2006 ), 桃树残根在土壤中降解产生大量的苦杏仁苷, 其水解生成的氢氰酸和苯甲醛对根系造成伤害(杨兴洪和罗新书, 1991 ), 植株残体在淹水缺氧条件下分解产生酚酸类物质(Patrick, 1971 ; Tang & Young, 1982 ).有研究发现: 地黄(Rehmannia glutinosa )连作土壤中酚酸类物质含量提高, 导致脲酶,蔗糖酶活性升高, 过氧化氢酶活性降低(陈慧等, 2007 ); 西瓜园酚酸类物质含量升高后, 土壤过氧化氢酶,脲酶活性均提高(吕卫光等, 2006 ).酚酸类物质含量增加, 导致杉木(Cunningh- amia lanceolata )苗叶绿素合成受阻, 光合作用减弱(陈秀华等, 2002 ), 对绿豆(Vigna radiata ),大豆(Glycine max ),黄瓜(Cucumis sativus )和玉米种子发芽和生长有抑制作用, 且随浓度升高抑制作用增强(曹光球等, 2003 ).酚酸类物质和苦杏仁苷都是造成果树连作障碍的重要因素(Patrick, 1955 ; Bӧrner, 1960 ).有研究表明, 棉田土壤生物活性受到作物连作障碍的负面效应和秸秆长期还田培肥地力的正面效应的双重影响(刘建国等, 2008 ).因此桃树枝条还田也可能会对桃树植株生长产生有利的影响. ...
酚酸在土壤--杉木苗间运移的初步探讨
1
2002
... 桃树再植当年基本不生新根, 发枝短长势弱, 造成桃园减产, 连作障碍严重(刘嘉彬等, 2006 ).桃树根系,枝条,叶片,果肉及桃仁中都有不同含量的苦杏仁苷(赵宇瑛等, 2006 ), 桃树残根在土壤中降解产生大量的苦杏仁苷, 其水解生成的氢氰酸和苯甲醛对根系造成伤害(杨兴洪和罗新书, 1991 ), 植株残体在淹水缺氧条件下分解产生酚酸类物质(Patrick, 1971 ; Tang & Young, 1982 ).有研究发现: 地黄(Rehmannia glutinosa )连作土壤中酚酸类物质含量提高, 导致脲酶,蔗糖酶活性升高, 过氧化氢酶活性降低(陈慧等, 2007 ); 西瓜园酚酸类物质含量升高后, 土壤过氧化氢酶,脲酶活性均提高(吕卫光等, 2006 ).酚酸类物质含量增加, 导致杉木(Cunningh- amia lanceolata )苗叶绿素合成受阻, 光合作用减弱(陈秀华等, 2002 ), 对绿豆(Vigna radiata ),大豆(Glycine max ),黄瓜(Cucumis sativus )和玉米种子发芽和生长有抑制作用, 且随浓度升高抑制作用增强(曹光球等, 2003 ).酚酸类物质和苦杏仁苷都是造成果树连作障碍的重要因素(Patrick, 1955 ; Bӧrner, 1960 ).有研究表明, 棉田土壤生物活性受到作物连作障碍的负面效应和秸秆长期还田培肥地力的正面效应的双重影响(刘建国等, 2008 ).因此桃树枝条还田也可能会对桃树植株生长产生有利的影响. ...
1
... 根据《中国统计年鉴》, 近年来全国桃园面积及产量稳步上升.桃(Amygdalus persica )是继苹果(Malus pumila ),梨(Pyrus ),葡萄(Vitis )之后的第四大水果.桃树枝条萌芽率高, 成枝力强, 枝叶生长量大, 生长旺的枝条一年可萌发2-3次副梢(Desmond & Daniele, 2008 ).栽培管理过程中需对桃树进行大量修剪以保证树体通风透光, 进而带来了剪落枝条大量堆积处理困难的问题, 这不仅影响桃园环境, 而且容易引起病虫害的发生.有关研究发现, 小麦(Triticum aestivum )和玉米(Zea mays )秸秆还田均有增产的效果(韩新忠等, 2012 ; 慕平等, 2012 ).因此, 枝条还田的提出, 在杜绝枝条杂乱占用果园面积影响生产的同时还可能有增肥增产作用, 为处理枝条提供了一种可供选择的方案. ...
1
... 采用四分法用土钻在距离桃树主干5-10 cm处随机取样, 深度0-20 cm, 每处理取3棵桃树, 共取样5次(2013年6月,7月,8月和2014年5月,6月).土壤蔗糖酶活性用3,5-二硝基水杨酸比色法测定, 37 ℃培养24 h; 土壤脲酶用苯酚钠比色法测定, 37 ℃培养24 h; 土壤过氧化氢酶活性用高锰酸钾滴定(0.1 mol·L-1 KMnO4 , 20 min) (关松荫, 1986 ). ...
不同小麦秸秆还田量对水稻生长,土壤微生物生物量及酶活性的影响
1
2012
... 根据《中国统计年鉴》, 近年来全国桃园面积及产量稳步上升.桃(Amygdalus persica )是继苹果(Malus pumila ),梨(Pyrus ),葡萄(Vitis )之后的第四大水果.桃树枝条萌芽率高, 成枝力强, 枝叶生长量大, 生长旺的枝条一年可萌发2-3次副梢(Desmond & Daniele, 2008 ).栽培管理过程中需对桃树进行大量修剪以保证树体通风透光, 进而带来了剪落枝条大量堆积处理困难的问题, 这不仅影响桃园环境, 而且容易引起病虫害的发生.有关研究发现, 小麦(Triticum aestivum )和玉米(Zea mays )秸秆还田均有增产的效果(韩新忠等, 2012 ; 慕平等, 2012 ).因此, 枝条还田的提出, 在杜绝枝条杂乱占用果园面积影响生产的同时还可能有增肥增产作用, 为处理枝条提供了一种可供选择的方案. ...
麦秸还田耕层酚酸的时间变化及其对夏玉米幼苗生长和产量的影响
2
2011
... 前人研究结果显示, 连作或秸秆还田的土壤中, 土壤酚酸类物质有明显提高.麦秸还田40天后, 玉米土壤耕作层酚酸含量达到最高峰(郑皓皓等, 2001 ; 胡晓军等, 2011 ), 花生(Arachis hypogaea )和黄瓜连作后土壤酚酸含量均显著升高(马云华等, 2005 ; 李培栋等, 2010 ), 连作苹果园土壤中酚酸物质总量显著提高(王青青, 2012 ).与其他作物不同, 桃树残根降解会产生苦杏仁苷, 经土壤酶分解产生的氢氰酸,苯甲醛和苯甲酸, 对新茬桃树根系产生巨大毒害(Baziramakenga et al ., 1995 ).本试验结果表明, 枝条浸提液中含有酚酸类物质和高量的苦杏仁苷, 加量枝条还田后, 土壤中酚酸类物质和苦杏仁苷含量与对照相比分别提高约30%和20%, 加量桃树枝条(22.5 g·kg-1 )的加入同样增加自毒物质的累积.因为盆栽桃树添加枝条后, 露天放置, 剪碎枝条经过雨水和灌溉水浸泡后同样会产生大量此类物质, 这与有关研究中植株残体淹水缺氧可分解产生挥发性脂肪酸和酚酸类(Patrick, 1971 ; Tang & Young, 1982 )的结果是一致的. ...
... 前人研究结果显示, 土壤自毒物质对不同植株表现出类似的抑制作用.外源添加肉桂酸,苯甲酸,对羟基苯甲酸, 抑制了杉木幼苗胚根和胚芽的生长(汪思龙等, 2002 ).土壤酚酸含量增加, 玉米幼苗的株高和根系生长均受到抑制(郑皓皓等, 2001 ; 胡晓军等, 2011 ).对羟基苯甲酸,香草酸和香豆酸的累积能够抑制花生幼苗生长, 提高花生发病率(李培栋等, 2010 ).桃树根系浸提液对桃树生长指标表现为低浓度促进,高浓度抑制的质量浓度效应(张斌斌等, 2012 ).本试验结果表明, 常规修剪量(1.5 g·kg-1 )还田对桃幼树生长无显著影响, 而加量枝条(22.5 g·kg-1 )还田时, 土壤酚酸总量达到1700 ng·g-1 , 土壤苦杏仁苷含量2.30 mg·g-1 , 出现了显著抑制现象, 如叶绿素含量降低,新梢停长期提前,地径生长量减小和净光合速率降低. ...
Characterization of soil microbial communities under different potato cropping systems by microbial population dynamics, substrate utilization, and fatty acid profiles.
1
2003
... 土壤微生物数量多, 种类多, 参与土壤有机质分解,养分转化等过程, 对植株生长发育有重要影响.研究发现, 连作障碍能够影响土壤中细菌和真菌的比例, 如连作棉田秸秆还田5-10年连作障碍明显, 细菌放线菌数量下降, 真菌比例提高(刘建国等, 2008 ; 刘素慧等, 2010 ), 马铃薯(Solanum tuberosum )连作后, 土壤由细菌型向真菌型转化(马琨等, 2010 ).马铃薯连作土壤中主要土传病原真菌为镰孢菌(Fusariun ),轮枝菌(Verticillium )和立枯丝核菌(Rhizoctonia solani )(李继平等, 2013 ), 黄瓜连作土壤真菌优势菌群主要是丝孢纲,外生菌根真菌,接合菌纲,伞菌纲,担子菌纲(乔蓬蕾等, 2014 ).本试验结果表明, 加量枝条处理后, 盆栽桃树土壤中真菌粪壳菌目,肉座菌目,曲霉属和比赤酵母减少, 伞菌纲,银耳亚纲Cystofilobasidiaceae和毛筒腔菌属增多, 这7种真菌均属于子囊菌门和担子菌门, 由此推断抑制桃树生长的优势真菌为子囊菌门和担子菌门.伞菌纲的最大功能是腐化植物残体, 它的增多加速了土壤中枝条的腐化.加量枝条处理后, 土壤中固氮菌缺失, 土壤中固氮菌能够吸取氮转化为氮肥供植株生长使用, 固氮菌的缺失不利于桃幼树生长.加量枝条处理后土壤中真菌比例升高, 是地力衰竭的标志(Larkin, 2003 ; 林茂兹等, 2012 ).桃园连作可能是自毒物质直接进入植株或通过影响土壤微生物(李小蓉和李蕾, 2013 )改变土壤环境, 进而影响植株生长引起的. ...
马铃薯连作田土壤中主要病原真菌的种群动态变化规律
1
2013
... 土壤微生物数量多, 种类多, 参与土壤有机质分解,养分转化等过程, 对植株生长发育有重要影响.研究发现, 连作障碍能够影响土壤中细菌和真菌的比例, 如连作棉田秸秆还田5-10年连作障碍明显, 细菌放线菌数量下降, 真菌比例提高(刘建国等, 2008 ; 刘素慧等, 2010 ), 马铃薯(Solanum tuberosum )连作后, 土壤由细菌型向真菌型转化(马琨等, 2010 ).马铃薯连作土壤中主要土传病原真菌为镰孢菌(Fusariun ),轮枝菌(Verticillium )和立枯丝核菌(Rhizoctonia solani )(李继平等, 2013 ), 黄瓜连作土壤真菌优势菌群主要是丝孢纲,外生菌根真菌,接合菌纲,伞菌纲,担子菌纲(乔蓬蕾等, 2014 ).本试验结果表明, 加量枝条处理后, 盆栽桃树土壤中真菌粪壳菌目,肉座菌目,曲霉属和比赤酵母减少, 伞菌纲,银耳亚纲Cystofilobasidiaceae和毛筒腔菌属增多, 这7种真菌均属于子囊菌门和担子菌门, 由此推断抑制桃树生长的优势真菌为子囊菌门和担子菌门.伞菌纲的最大功能是腐化植物残体, 它的增多加速了土壤中枝条的腐化.加量枝条处理后, 土壤中固氮菌缺失, 土壤中固氮菌能够吸取氮转化为氮肥供植株生长使用, 固氮菌的缺失不利于桃幼树生长.加量枝条处理后土壤中真菌比例升高, 是地力衰竭的标志(Larkin, 2003 ; 林茂兹等, 2012 ).桃园连作可能是自毒物质直接进入植株或通过影响土壤微生物(李小蓉和李蕾, 2013 )改变土壤环境, 进而影响植株生长引起的. ...
连作花生土壤中酚酸类物质的检测及其对花生的化感作用
2
2010
... 前人研究结果显示, 连作或秸秆还田的土壤中, 土壤酚酸类物质有明显提高.麦秸还田40天后, 玉米土壤耕作层酚酸含量达到最高峰(郑皓皓等, 2001 ; 胡晓军等, 2011 ), 花生(Arachis hypogaea )和黄瓜连作后土壤酚酸含量均显著升高(马云华等, 2005 ; 李培栋等, 2010 ), 连作苹果园土壤中酚酸物质总量显著提高(王青青, 2012 ).与其他作物不同, 桃树残根降解会产生苦杏仁苷, 经土壤酶分解产生的氢氰酸,苯甲醛和苯甲酸, 对新茬桃树根系产生巨大毒害(Baziramakenga et al ., 1995 ).本试验结果表明, 枝条浸提液中含有酚酸类物质和高量的苦杏仁苷, 加量枝条还田后, 土壤中酚酸类物质和苦杏仁苷含量与对照相比分别提高约30%和20%, 加量桃树枝条(22.5 g·kg-1 )的加入同样增加自毒物质的累积.因为盆栽桃树添加枝条后, 露天放置, 剪碎枝条经过雨水和灌溉水浸泡后同样会产生大量此类物质, 这与有关研究中植株残体淹水缺氧可分解产生挥发性脂肪酸和酚酸类(Patrick, 1971 ; Tang & Young, 1982 )的结果是一致的. ...
... 前人研究结果显示, 土壤自毒物质对不同植株表现出类似的抑制作用.外源添加肉桂酸,苯甲酸,对羟基苯甲酸, 抑制了杉木幼苗胚根和胚芽的生长(汪思龙等, 2002 ).土壤酚酸含量增加, 玉米幼苗的株高和根系生长均受到抑制(郑皓皓等, 2001 ; 胡晓军等, 2011 ).对羟基苯甲酸,香草酸和香豆酸的累积能够抑制花生幼苗生长, 提高花生发病率(李培栋等, 2010 ).桃树根系浸提液对桃树生长指标表现为低浓度促进,高浓度抑制的质量浓度效应(张斌斌等, 2012 ).本试验结果表明, 常规修剪量(1.5 g·kg-1 )还田对桃幼树生长无显著影响, 而加量枝条(22.5 g·kg-1 )还田时, 土壤酚酸总量达到1700 ng·g-1 , 土壤苦杏仁苷含量2.30 mg·g-1 , 出现了显著抑制现象, 如叶绿素含量降低,新梢停长期提前,地径生长量减小和净光合速率降低. ...
植物化感物质与土壤微生物的研究进展
1
2013
... 土壤微生物数量多, 种类多, 参与土壤有机质分解,养分转化等过程, 对植株生长发育有重要影响.研究发现, 连作障碍能够影响土壤中细菌和真菌的比例, 如连作棉田秸秆还田5-10年连作障碍明显, 细菌放线菌数量下降, 真菌比例提高(刘建国等, 2008 ; 刘素慧等, 2010 ), 马铃薯(Solanum tuberosum )连作后, 土壤由细菌型向真菌型转化(马琨等, 2010 ).马铃薯连作土壤中主要土传病原真菌为镰孢菌(Fusariun ),轮枝菌(Verticillium )和立枯丝核菌(Rhizoctonia solani )(李继平等, 2013 ), 黄瓜连作土壤真菌优势菌群主要是丝孢纲,外生菌根真菌,接合菌纲,伞菌纲,担子菌纲(乔蓬蕾等, 2014 ).本试验结果表明, 加量枝条处理后, 盆栽桃树土壤中真菌粪壳菌目,肉座菌目,曲霉属和比赤酵母减少, 伞菌纲,银耳亚纲Cystofilobasidiaceae和毛筒腔菌属增多, 这7种真菌均属于子囊菌门和担子菌门, 由此推断抑制桃树生长的优势真菌为子囊菌门和担子菌门.伞菌纲的最大功能是腐化植物残体, 它的增多加速了土壤中枝条的腐化.加量枝条处理后, 土壤中固氮菌缺失, 土壤中固氮菌能够吸取氮转化为氮肥供植株生长使用, 固氮菌的缺失不利于桃幼树生长.加量枝条处理后土壤中真菌比例升高, 是地力衰竭的标志(Larkin, 2003 ; 林茂兹等, 2012 ).桃园连作可能是自毒物质直接进入植株或通过影响土壤微生物(李小蓉和李蕾, 2013 )改变土壤环境, 进而影响植株生长引起的. ...
太子参连作对根际土壤微生物的影响
1
2012
... 土壤微生物数量多, 种类多, 参与土壤有机质分解,养分转化等过程, 对植株生长发育有重要影响.研究发现, 连作障碍能够影响土壤中细菌和真菌的比例, 如连作棉田秸秆还田5-10年连作障碍明显, 细菌放线菌数量下降, 真菌比例提高(刘建国等, 2008 ; 刘素慧等, 2010 ), 马铃薯(Solanum tuberosum )连作后, 土壤由细菌型向真菌型转化(马琨等, 2010 ).马铃薯连作土壤中主要土传病原真菌为镰孢菌(Fusariun ),轮枝菌(Verticillium )和立枯丝核菌(Rhizoctonia solani )(李继平等, 2013 ), 黄瓜连作土壤真菌优势菌群主要是丝孢纲,外生菌根真菌,接合菌纲,伞菌纲,担子菌纲(乔蓬蕾等, 2014 ).本试验结果表明, 加量枝条处理后, 盆栽桃树土壤中真菌粪壳菌目,肉座菌目,曲霉属和比赤酵母减少, 伞菌纲,银耳亚纲Cystofilobasidiaceae和毛筒腔菌属增多, 这7种真菌均属于子囊菌门和担子菌门, 由此推断抑制桃树生长的优势真菌为子囊菌门和担子菌门.伞菌纲的最大功能是腐化植物残体, 它的增多加速了土壤中枝条的腐化.加量枝条处理后, 土壤中固氮菌缺失, 土壤中固氮菌能够吸取氮转化为氮肥供植株生长使用, 固氮菌的缺失不利于桃幼树生长.加量枝条处理后土壤中真菌比例升高, 是地力衰竭的标志(Larkin, 2003 ; 林茂兹等, 2012 ).桃园连作可能是自毒物质直接进入植株或通过影响土壤微生物(李小蓉和李蕾, 2013 )改变土壤环境, 进而影响植株生长引起的. ...
桃树重茬病及其防治技术
1
2006
... 桃树再植当年基本不生新根, 发枝短长势弱, 造成桃园减产, 连作障碍严重(刘嘉彬等, 2006 ).桃树根系,枝条,叶片,果肉及桃仁中都有不同含量的苦杏仁苷(赵宇瑛等, 2006 ), 桃树残根在土壤中降解产生大量的苦杏仁苷, 其水解生成的氢氰酸和苯甲醛对根系造成伤害(杨兴洪和罗新书, 1991 ), 植株残体在淹水缺氧条件下分解产生酚酸类物质(Patrick, 1971 ; Tang & Young, 1982 ).有研究发现: 地黄(Rehmannia glutinosa )连作土壤中酚酸类物质含量提高, 导致脲酶,蔗糖酶活性升高, 过氧化氢酶活性降低(陈慧等, 2007 ); 西瓜园酚酸类物质含量升高后, 土壤过氧化氢酶,脲酶活性均提高(吕卫光等, 2006 ).酚酸类物质含量增加, 导致杉木(Cunningh- amia lanceolata )苗叶绿素合成受阻, 光合作用减弱(陈秀华等, 2002 ), 对绿豆(Vigna radiata ),大豆(Glycine max ),黄瓜(Cucumis sativus )和玉米种子发芽和生长有抑制作用, 且随浓度升高抑制作用增强(曹光球等, 2003 ).酚酸类物质和苦杏仁苷都是造成果树连作障碍的重要因素(Patrick, 1955 ; Bӧrner, 1960 ).有研究表明, 棉田土壤生物活性受到作物连作障碍的负面效应和秸秆长期还田培肥地力的正面效应的双重影响(刘建国等, 2008 ).因此桃树枝条还田也可能会对桃树植株生长产生有利的影响. ...
长期连作和秸秆还田对棉田土壤生物活性的影响
2
2008
... 桃树再植当年基本不生新根, 发枝短长势弱, 造成桃园减产, 连作障碍严重(刘嘉彬等, 2006 ).桃树根系,枝条,叶片,果肉及桃仁中都有不同含量的苦杏仁苷(赵宇瑛等, 2006 ), 桃树残根在土壤中降解产生大量的苦杏仁苷, 其水解生成的氢氰酸和苯甲醛对根系造成伤害(杨兴洪和罗新书, 1991 ), 植株残体在淹水缺氧条件下分解产生酚酸类物质(Patrick, 1971 ; Tang & Young, 1982 ).有研究发现: 地黄(Rehmannia glutinosa )连作土壤中酚酸类物质含量提高, 导致脲酶,蔗糖酶活性升高, 过氧化氢酶活性降低(陈慧等, 2007 ); 西瓜园酚酸类物质含量升高后, 土壤过氧化氢酶,脲酶活性均提高(吕卫光等, 2006 ).酚酸类物质含量增加, 导致杉木(Cunningh- amia lanceolata )苗叶绿素合成受阻, 光合作用减弱(陈秀华等, 2002 ), 对绿豆(Vigna radiata ),大豆(Glycine max ),黄瓜(Cucumis sativus )和玉米种子发芽和生长有抑制作用, 且随浓度升高抑制作用增强(曹光球等, 2003 ).酚酸类物质和苦杏仁苷都是造成果树连作障碍的重要因素(Patrick, 1955 ; Bӧrner, 1960 ).有研究表明, 棉田土壤生物活性受到作物连作障碍的负面效应和秸秆长期还田培肥地力的正面效应的双重影响(刘建国等, 2008 ).因此桃树枝条还田也可能会对桃树植株生长产生有利的影响. ...
... 土壤微生物数量多, 种类多, 参与土壤有机质分解,养分转化等过程, 对植株生长发育有重要影响.研究发现, 连作障碍能够影响土壤中细菌和真菌的比例, 如连作棉田秸秆还田5-10年连作障碍明显, 细菌放线菌数量下降, 真菌比例提高(刘建国等, 2008 ; 刘素慧等, 2010 ), 马铃薯(Solanum tuberosum )连作后, 土壤由细菌型向真菌型转化(马琨等, 2010 ).马铃薯连作土壤中主要土传病原真菌为镰孢菌(Fusariun ),轮枝菌(Verticillium )和立枯丝核菌(Rhizoctonia solani )(李继平等, 2013 ), 黄瓜连作土壤真菌优势菌群主要是丝孢纲,外生菌根真菌,接合菌纲,伞菌纲,担子菌纲(乔蓬蕾等, 2014 ).本试验结果表明, 加量枝条处理后, 盆栽桃树土壤中真菌粪壳菌目,肉座菌目,曲霉属和比赤酵母减少, 伞菌纲,银耳亚纲Cystofilobasidiaceae和毛筒腔菌属增多, 这7种真菌均属于子囊菌门和担子菌门, 由此推断抑制桃树生长的优势真菌为子囊菌门和担子菌门.伞菌纲的最大功能是腐化植物残体, 它的增多加速了土壤中枝条的腐化.加量枝条处理后, 土壤中固氮菌缺失, 土壤中固氮菌能够吸取氮转化为氮肥供植株生长使用, 固氮菌的缺失不利于桃幼树生长.加量枝条处理后土壤中真菌比例升高, 是地力衰竭的标志(Larkin, 2003 ; 林茂兹等, 2012 ).桃园连作可能是自毒物质直接进入植株或通过影响土壤微生物(李小蓉和李蕾, 2013 )改变土壤环境, 进而影响植株生长引起的. ...
大蒜连作对其根际土壤微生物和酶活性的影响
1
2010
... 土壤微生物数量多, 种类多, 参与土壤有机质分解,养分转化等过程, 对植株生长发育有重要影响.研究发现, 连作障碍能够影响土壤中细菌和真菌的比例, 如连作棉田秸秆还田5-10年连作障碍明显, 细菌放线菌数量下降, 真菌比例提高(刘建国等, 2008 ; 刘素慧等, 2010 ), 马铃薯(Solanum tuberosum )连作后, 土壤由细菌型向真菌型转化(马琨等, 2010 ).马铃薯连作土壤中主要土传病原真菌为镰孢菌(Fusariun ),轮枝菌(Verticillium )和立枯丝核菌(Rhizoctonia solani )(李继平等, 2013 ), 黄瓜连作土壤真菌优势菌群主要是丝孢纲,外生菌根真菌,接合菌纲,伞菌纲,担子菌纲(乔蓬蕾等, 2014 ).本试验结果表明, 加量枝条处理后, 盆栽桃树土壤中真菌粪壳菌目,肉座菌目,曲霉属和比赤酵母减少, 伞菌纲,银耳亚纲Cystofilobasidiaceae和毛筒腔菌属增多, 这7种真菌均属于子囊菌门和担子菌门, 由此推断抑制桃树生长的优势真菌为子囊菌门和担子菌门.伞菌纲的最大功能是腐化植物残体, 它的增多加速了土壤中枝条的腐化.加量枝条处理后, 土壤中固氮菌缺失, 土壤中固氮菌能够吸取氮转化为氮肥供植株生长使用, 固氮菌的缺失不利于桃幼树生长.加量枝条处理后土壤中真菌比例升高, 是地力衰竭的标志(Larkin, 2003 ; 林茂兹等, 2012 ).桃园连作可能是自毒物质直接进入植株或通过影响土壤微生物(李小蓉和李蕾, 2013 )改变土壤环境, 进而影响植株生长引起的. ...
酚酸化合物对土壤酶活性和土壤养分的影响
1
2006
... 桃树再植当年基本不生新根, 发枝短长势弱, 造成桃园减产, 连作障碍严重(刘嘉彬等, 2006 ).桃树根系,枝条,叶片,果肉及桃仁中都有不同含量的苦杏仁苷(赵宇瑛等, 2006 ), 桃树残根在土壤中降解产生大量的苦杏仁苷, 其水解生成的氢氰酸和苯甲醛对根系造成伤害(杨兴洪和罗新书, 1991 ), 植株残体在淹水缺氧条件下分解产生酚酸类物质(Patrick, 1971 ; Tang & Young, 1982 ).有研究发现: 地黄(Rehmannia glutinosa )连作土壤中酚酸类物质含量提高, 导致脲酶,蔗糖酶活性升高, 过氧化氢酶活性降低(陈慧等, 2007 ); 西瓜园酚酸类物质含量升高后, 土壤过氧化氢酶,脲酶活性均提高(吕卫光等, 2006 ).酚酸类物质含量增加, 导致杉木(Cunningh- amia lanceolata )苗叶绿素合成受阻, 光合作用减弱(陈秀华等, 2002 ), 对绿豆(Vigna radiata ),大豆(Glycine max ),黄瓜(Cucumis sativus )和玉米种子发芽和生长有抑制作用, 且随浓度升高抑制作用增强(曹光球等, 2003 ).酚酸类物质和苦杏仁苷都是造成果树连作障碍的重要因素(Patrick, 1955 ; Bӧrner, 1960 ).有研究表明, 棉田土壤生物活性受到作物连作障碍的负面效应和秸秆长期还田培肥地力的正面效应的双重影响(刘建国等, 2008 ).因此桃树枝条还田也可能会对桃树植株生长产生有利的影响. ...
秸秆还田对宁南旱作农田土壤活性有机碳及酶活性的影响
1
2011
... 土壤酶活性是研究土壤质量的重要指标.在小麦-玉米轮作情况下, 秸秆还田提高了土壤酶活性(路文涛等, 2011 ; 甄丽莎等, 2012 ).杨树林土壤酚酸浓度升高后, 土壤脲酶和碱性磷酸酶活性降低, 过氧化氢酶活性则升高(王延平等, 2013 ).肖宏(2004)研究发现连作苹果土壤中酚酸类物质含量升高后, 基础酶活性降低.本试验结果表明, T1和T3处理的土壤酶活性与CK无显著差异.T2和T4处理的土壤蔗糖酶活性始终高于CK.处理前期, 土壤过氧化氢酶和脲酶活性均显著高于CK, 处理后期均显著低于CK.蔗糖酶又称转化酶, 与土壤有机质含量呈正相关, 可能是因为枝条腐解产生有机质, 引起蔗糖酶活性升高.脲酶促进尿素氮肥转化成氨, 过氧化氢酶可以清除土壤中的过氧化氢.处理后期脲酶和过氧化氢酶活性降低, 减少了植株的直接氮素来源, 造成土壤中过氧化氢的积累而不利于桃树植株的 生长. ...
马铃薯连作栽培对土壤微生物群落的影响
1
2010
... 土壤微生物数量多, 种类多, 参与土壤有机质分解,养分转化等过程, 对植株生长发育有重要影响.研究发现, 连作障碍能够影响土壤中细菌和真菌的比例, 如连作棉田秸秆还田5-10年连作障碍明显, 细菌放线菌数量下降, 真菌比例提高(刘建国等, 2008 ; 刘素慧等, 2010 ), 马铃薯(Solanum tuberosum )连作后, 土壤由细菌型向真菌型转化(马琨等, 2010 ).马铃薯连作土壤中主要土传病原真菌为镰孢菌(Fusariun ),轮枝菌(Verticillium )和立枯丝核菌(Rhizoctonia solani )(李继平等, 2013 ), 黄瓜连作土壤真菌优势菌群主要是丝孢纲,外生菌根真菌,接合菌纲,伞菌纲,担子菌纲(乔蓬蕾等, 2014 ).本试验结果表明, 加量枝条处理后, 盆栽桃树土壤中真菌粪壳菌目,肉座菌目,曲霉属和比赤酵母减少, 伞菌纲,银耳亚纲Cystofilobasidiaceae和毛筒腔菌属增多, 这7种真菌均属于子囊菌门和担子菌门, 由此推断抑制桃树生长的优势真菌为子囊菌门和担子菌门.伞菌纲的最大功能是腐化植物残体, 它的增多加速了土壤中枝条的腐化.加量枝条处理后, 土壤中固氮菌缺失, 土壤中固氮菌能够吸取氮转化为氮肥供植株生长使用, 固氮菌的缺失不利于桃幼树生长.加量枝条处理后土壤中真菌比例升高, 是地力衰竭的标志(Larkin, 2003 ; 林茂兹等, 2012 ).桃园连作可能是自毒物质直接进入植株或通过影响土壤微生物(李小蓉和李蕾, 2013 )改变土壤环境, 进而影响植株生长引起的. ...
黄瓜连作土壤酚酸类物质积累对土壤微生物和酶活性的影响
1
2005
... 前人研究结果显示, 连作或秸秆还田的土壤中, 土壤酚酸类物质有明显提高.麦秸还田40天后, 玉米土壤耕作层酚酸含量达到最高峰(郑皓皓等, 2001 ; 胡晓军等, 2011 ), 花生(Arachis hypogaea )和黄瓜连作后土壤酚酸含量均显著升高(马云华等, 2005 ; 李培栋等, 2010 ), 连作苹果园土壤中酚酸物质总量显著提高(王青青, 2012 ).与其他作物不同, 桃树残根降解会产生苦杏仁苷, 经土壤酶分解产生的氢氰酸,苯甲醛和苯甲酸, 对新茬桃树根系产生巨大毒害(Baziramakenga et al ., 1995 ).本试验结果表明, 枝条浸提液中含有酚酸类物质和高量的苦杏仁苷, 加量枝条还田后, 土壤中酚酸类物质和苦杏仁苷含量与对照相比分别提高约30%和20%, 加量桃树枝条(22.5 g·kg-1 )的加入同样增加自毒物质的累积.因为盆栽桃树添加枝条后, 露天放置, 剪碎枝条经过雨水和灌溉水浸泡后同样会产生大量此类物质, 这与有关研究中植株残体淹水缺氧可分解产生挥发性脂肪酸和酚酸类(Patrick, 1971 ; Tang & Young, 1982 )的结果是一致的. ...
不同年限全量玉米秸秆还田对玉米生长发育及土壤理化性状的影响
1
2012
... 根据《中国统计年鉴》, 近年来全国桃园面积及产量稳步上升.桃(Amygdalus persica )是继苹果(Malus pumila ),梨(Pyrus ),葡萄(Vitis )之后的第四大水果.桃树枝条萌芽率高, 成枝力强, 枝叶生长量大, 生长旺的枝条一年可萌发2-3次副梢(Desmond & Daniele, 2008 ).栽培管理过程中需对桃树进行大量修剪以保证树体通风透光, 进而带来了剪落枝条大量堆积处理困难的问题, 这不仅影响桃园环境, 而且容易引起病虫害的发生.有关研究发现, 小麦(Triticum aestivum )和玉米(Zea mays )秸秆还田均有增产的效果(韩新忠等, 2012 ; 慕平等, 2012 ).因此, 枝条还田的提出, 在杜绝枝条杂乱占用果园面积影响生产的同时还可能有增肥增产作用, 为处理枝条提供了一种可供选择的方案. ...
The peach replant problem in Ontario: II Toxic substances from microbial decomposition products of peach root residues.
2
1955
... 桃树再植当年基本不生新根, 发枝短长势弱, 造成桃园减产, 连作障碍严重(刘嘉彬等, 2006 ).桃树根系,枝条,叶片,果肉及桃仁中都有不同含量的苦杏仁苷(赵宇瑛等, 2006 ), 桃树残根在土壤中降解产生大量的苦杏仁苷, 其水解生成的氢氰酸和苯甲醛对根系造成伤害(杨兴洪和罗新书, 1991 ), 植株残体在淹水缺氧条件下分解产生酚酸类物质(Patrick, 1971 ; Tang & Young, 1982 ).有研究发现: 地黄(Rehmannia glutinosa )连作土壤中酚酸类物质含量提高, 导致脲酶,蔗糖酶活性升高, 过氧化氢酶活性降低(陈慧等, 2007 ); 西瓜园酚酸类物质含量升高后, 土壤过氧化氢酶,脲酶活性均提高(吕卫光等, 2006 ).酚酸类物质含量增加, 导致杉木(Cunningh- amia lanceolata )苗叶绿素合成受阻, 光合作用减弱(陈秀华等, 2002 ), 对绿豆(Vigna radiata ),大豆(Glycine max ),黄瓜(Cucumis sativus )和玉米种子发芽和生长有抑制作用, 且随浓度升高抑制作用增强(曹光球等, 2003 ).酚酸类物质和苦杏仁苷都是造成果树连作障碍的重要因素(Patrick, 1955 ; Bӧrner, 1960 ).有研究表明, 棉田土壤生物活性受到作物连作障碍的负面效应和秸秆长期还田培肥地力的正面效应的双重影响(刘建国等, 2008 ).因此桃树枝条还田也可能会对桃树植株生长产生有利的影响. ...
... 常规量枝条添加与对照无显著差异, 而加量枝条的加入为盆栽土壤提供了大量的苦杏仁苷和酚酸类自毒物质, 自毒物质直接抑制桃幼树生长(Patrick, 1955 ), 同时能够通过间接途径改变土壤微生物结构和土壤基础酶活性, 引起地下部分变化而调控桃幼树地上部分生长, 导致叶绿素含量降低, 新梢停长期提前, 地径生长减缓, 叶片净光合速率降低等现象.但究竟是酚酸类物质还是苦杏仁苷在这个过程中起关键作用仍有待进一步研究.大田两年桃树枝条(2012年12月至2014年7月)还田试验表明, 与对照相比, 常规修剪量枝条还田处理, 土壤自毒物质,微生物,酶活性及桃树生长指标均无显著差异.初步推测, 常规修剪量枝条还田对植株生长没有不利影响; 但加量枝条集中还田, 会对桃树生长产生抑制作用. ...
Phytotoxic substances associated with the decomposition in soil of plant residues.
2
1971
... 桃树再植当年基本不生新根, 发枝短长势弱, 造成桃园减产, 连作障碍严重(刘嘉彬等, 2006 ).桃树根系,枝条,叶片,果肉及桃仁中都有不同含量的苦杏仁苷(赵宇瑛等, 2006 ), 桃树残根在土壤中降解产生大量的苦杏仁苷, 其水解生成的氢氰酸和苯甲醛对根系造成伤害(杨兴洪和罗新书, 1991 ), 植株残体在淹水缺氧条件下分解产生酚酸类物质(Patrick, 1971 ; Tang & Young, 1982 ).有研究发现: 地黄(Rehmannia glutinosa )连作土壤中酚酸类物质含量提高, 导致脲酶,蔗糖酶活性升高, 过氧化氢酶活性降低(陈慧等, 2007 ); 西瓜园酚酸类物质含量升高后, 土壤过氧化氢酶,脲酶活性均提高(吕卫光等, 2006 ).酚酸类物质含量增加, 导致杉木(Cunningh- amia lanceolata )苗叶绿素合成受阻, 光合作用减弱(陈秀华等, 2002 ), 对绿豆(Vigna radiata ),大豆(Glycine max ),黄瓜(Cucumis sativus )和玉米种子发芽和生长有抑制作用, 且随浓度升高抑制作用增强(曹光球等, 2003 ).酚酸类物质和苦杏仁苷都是造成果树连作障碍的重要因素(Patrick, 1955 ; Bӧrner, 1960 ).有研究表明, 棉田土壤生物活性受到作物连作障碍的负面效应和秸秆长期还田培肥地力的正面效应的双重影响(刘建国等, 2008 ).因此桃树枝条还田也可能会对桃树植株生长产生有利的影响. ...
... 前人研究结果显示, 连作或秸秆还田的土壤中, 土壤酚酸类物质有明显提高.麦秸还田40天后, 玉米土壤耕作层酚酸含量达到最高峰(郑皓皓等, 2001 ; 胡晓军等, 2011 ), 花生(Arachis hypogaea )和黄瓜连作后土壤酚酸含量均显著升高(马云华等, 2005 ; 李培栋等, 2010 ), 连作苹果园土壤中酚酸物质总量显著提高(王青青, 2012 ).与其他作物不同, 桃树残根降解会产生苦杏仁苷, 经土壤酶分解产生的氢氰酸,苯甲醛和苯甲酸, 对新茬桃树根系产生巨大毒害(Baziramakenga et al ., 1995 ).本试验结果表明, 枝条浸提液中含有酚酸类物质和高量的苦杏仁苷, 加量枝条还田后, 土壤中酚酸类物质和苦杏仁苷含量与对照相比分别提高约30%和20%, 加量桃树枝条(22.5 g·kg-1 )的加入同样增加自毒物质的累积.因为盆栽桃树添加枝条后, 露天放置, 剪碎枝条经过雨水和灌溉水浸泡后同样会产生大量此类物质, 这与有关研究中植株残体淹水缺氧可分解产生挥发性脂肪酸和酚酸类(Patrick, 1971 ; Tang & Young, 1982 )的结果是一致的. ...
不同连作茬次黄瓜根际土壤微生物群落的T-RFLP分析
1
2014
... 土壤微生物数量多, 种类多, 参与土壤有机质分解,养分转化等过程, 对植株生长发育有重要影响.研究发现, 连作障碍能够影响土壤中细菌和真菌的比例, 如连作棉田秸秆还田5-10年连作障碍明显, 细菌放线菌数量下降, 真菌比例提高(刘建国等, 2008 ; 刘素慧等, 2010 ), 马铃薯(Solanum tuberosum )连作后, 土壤由细菌型向真菌型转化(马琨等, 2010 ).马铃薯连作土壤中主要土传病原真菌为镰孢菌(Fusariun ),轮枝菌(Verticillium )和立枯丝核菌(Rhizoctonia solani )(李继平等, 2013 ), 黄瓜连作土壤真菌优势菌群主要是丝孢纲,外生菌根真菌,接合菌纲,伞菌纲,担子菌纲(乔蓬蕾等, 2014 ).本试验结果表明, 加量枝条处理后, 盆栽桃树土壤中真菌粪壳菌目,肉座菌目,曲霉属和比赤酵母减少, 伞菌纲,银耳亚纲Cystofilobasidiaceae和毛筒腔菌属增多, 这7种真菌均属于子囊菌门和担子菌门, 由此推断抑制桃树生长的优势真菌为子囊菌门和担子菌门.伞菌纲的最大功能是腐化植物残体, 它的增多加速了土壤中枝条的腐化.加量枝条处理后, 土壤中固氮菌缺失, 土壤中固氮菌能够吸取氮转化为氮肥供植株生长使用, 固氮菌的缺失不利于桃幼树生长.加量枝条处理后土壤中真菌比例升高, 是地力衰竭的标志(Larkin, 2003 ; 林茂兹等, 2012 ).桃园连作可能是自毒物质直接进入植株或通过影响土壤微生物(李小蓉和李蕾, 2013 )改变土壤环境, 进而影响植株生长引起的. ...
Collection and identification of allelopathic compounds from the undisturbed root system of bigalta limpograss (Hemarthria altissima).
2
1982
... 桃树再植当年基本不生新根, 发枝短长势弱, 造成桃园减产, 连作障碍严重(刘嘉彬等, 2006 ).桃树根系,枝条,叶片,果肉及桃仁中都有不同含量的苦杏仁苷(赵宇瑛等, 2006 ), 桃树残根在土壤中降解产生大量的苦杏仁苷, 其水解生成的氢氰酸和苯甲醛对根系造成伤害(杨兴洪和罗新书, 1991 ), 植株残体在淹水缺氧条件下分解产生酚酸类物质(Patrick, 1971 ; Tang & Young, 1982 ).有研究发现: 地黄(Rehmannia glutinosa )连作土壤中酚酸类物质含量提高, 导致脲酶,蔗糖酶活性升高, 过氧化氢酶活性降低(陈慧等, 2007 ); 西瓜园酚酸类物质含量升高后, 土壤过氧化氢酶,脲酶活性均提高(吕卫光等, 2006 ).酚酸类物质含量增加, 导致杉木(Cunningh- amia lanceolata )苗叶绿素合成受阻, 光合作用减弱(陈秀华等, 2002 ), 对绿豆(Vigna radiata ),大豆(Glycine max ),黄瓜(Cucumis sativus )和玉米种子发芽和生长有抑制作用, 且随浓度升高抑制作用增强(曹光球等, 2003 ).酚酸类物质和苦杏仁苷都是造成果树连作障碍的重要因素(Patrick, 1955 ; Bӧrner, 1960 ).有研究表明, 棉田土壤生物活性受到作物连作障碍的负面效应和秸秆长期还田培肥地力的正面效应的双重影响(刘建国等, 2008 ).因此桃树枝条还田也可能会对桃树植株生长产生有利的影响. ...
... 前人研究结果显示, 连作或秸秆还田的土壤中, 土壤酚酸类物质有明显提高.麦秸还田40天后, 玉米土壤耕作层酚酸含量达到最高峰(郑皓皓等, 2001 ; 胡晓军等, 2011 ), 花生(Arachis hypogaea )和黄瓜连作后土壤酚酸含量均显著升高(马云华等, 2005 ; 李培栋等, 2010 ), 连作苹果园土壤中酚酸物质总量显著提高(王青青, 2012 ).与其他作物不同, 桃树残根降解会产生苦杏仁苷, 经土壤酶分解产生的氢氰酸,苯甲醛和苯甲酸, 对新茬桃树根系产生巨大毒害(Baziramakenga et al ., 1995 ).本试验结果表明, 枝条浸提液中含有酚酸类物质和高量的苦杏仁苷, 加量枝条还田后, 土壤中酚酸类物质和苦杏仁苷含量与对照相比分别提高约30%和20%, 加量桃树枝条(22.5 g·kg-1 )的加入同样增加自毒物质的累积.因为盆栽桃树添加枝条后, 露天放置, 剪碎枝条经过雨水和灌溉水浸泡后同样会产生大量此类物质, 这与有关研究中植株残体淹水缺氧可分解产生挥发性脂肪酸和酚酸类(Patrick, 1971 ; Tang & Young, 1982 )的结果是一致的. ...
苹果园土壤酚酸类物质变化及根皮苷对TCA循环影响的研究
1
2012
... 前人研究结果显示, 连作或秸秆还田的土壤中, 土壤酚酸类物质有明显提高.麦秸还田40天后, 玉米土壤耕作层酚酸含量达到最高峰(郑皓皓等, 2001 ; 胡晓军等, 2011 ), 花生(Arachis hypogaea )和黄瓜连作后土壤酚酸含量均显著升高(马云华等, 2005 ; 李培栋等, 2010 ), 连作苹果园土壤中酚酸物质总量显著提高(王青青, 2012 ).与其他作物不同, 桃树残根降解会产生苦杏仁苷, 经土壤酶分解产生的氢氰酸,苯甲醛和苯甲酸, 对新茬桃树根系产生巨大毒害(Baziramakenga et al ., 1995 ).本试验结果表明, 枝条浸提液中含有酚酸类物质和高量的苦杏仁苷, 加量枝条还田后, 土壤中酚酸类物质和苦杏仁苷含量与对照相比分别提高约30%和20%, 加量桃树枝条(22.5 g·kg-1 )的加入同样增加自毒物质的累积.因为盆栽桃树添加枝条后, 露天放置, 剪碎枝条经过雨水和灌溉水浸泡后同样会产生大量此类物质, 这与有关研究中植株残体淹水缺氧可分解产生挥发性脂肪酸和酚酸类(Patrick, 1971 ; Tang & Young, 1982 )的结果是一致的. ...
几种化感物质对杉木幼苗生长的影响
1
2002
... 前人研究结果显示, 土壤自毒物质对不同植株表现出类似的抑制作用.外源添加肉桂酸,苯甲酸,对羟基苯甲酸, 抑制了杉木幼苗胚根和胚芽的生长(汪思龙等, 2002 ).土壤酚酸含量增加, 玉米幼苗的株高和根系生长均受到抑制(郑皓皓等, 2001 ; 胡晓军等, 2011 ).对羟基苯甲酸,香草酸和香豆酸的累积能够抑制花生幼苗生长, 提高花生发病率(李培栋等, 2010 ).桃树根系浸提液对桃树生长指标表现为低浓度促进,高浓度抑制的质量浓度效应(张斌斌等, 2012 ).本试验结果表明, 常规修剪量(1.5 g·kg-1 )还田对桃幼树生长无显著影响, 而加量枝条(22.5 g·kg-1 )还田时, 土壤酚酸总量达到1700 ng·g-1 , 土壤苦杏仁苷含量2.30 mg·g-1 , 出现了显著抑制现象, 如叶绿素含量降低,新梢停长期提前,地径生长量减小和净光合速率降低. ...
酚酸对杨树人工林土壤养分有效性及酶活性的影响
1
2013
... 土壤酶活性是研究土壤质量的重要指标.在小麦-玉米轮作情况下, 秸秆还田提高了土壤酶活性(路文涛等, 2011 ; 甄丽莎等, 2012 ).杨树林土壤酚酸浓度升高后, 土壤脲酶和碱性磷酸酶活性降低, 过氧化氢酶活性则升高(王延平等, 2013 ).肖宏(2004)研究发现连作苹果土壤中酚酸类物质含量升高后, 基础酶活性降低.本试验结果表明, T1和T3处理的土壤酶活性与CK无显著差异.T2和T4处理的土壤蔗糖酶活性始终高于CK.处理前期, 土壤过氧化氢酶和脲酶活性均显著高于CK, 处理后期均显著低于CK.蔗糖酶又称转化酶, 与土壤有机质含量呈正相关, 可能是因为枝条腐解产生有机质, 引起蔗糖酶活性升高.脲酶促进尿素氮肥转化成氨, 过氧化氢酶可以清除土壤中的过氧化氢.处理后期脲酶和过氧化氢酶活性降低, 减少了植株的直接氮素来源, 造成土壤中过氧化氢的积累而不利于桃树植株的 生长. ...
土壤消毒和轮作对克服苹果连作障碍效果的研究
2004
果树再植问题研究进展
1
1991
... 桃树再植当年基本不生新根, 发枝短长势弱, 造成桃园减产, 连作障碍严重(刘嘉彬等, 2006 ).桃树根系,枝条,叶片,果肉及桃仁中都有不同含量的苦杏仁苷(赵宇瑛等, 2006 ), 桃树残根在土壤中降解产生大量的苦杏仁苷, 其水解生成的氢氰酸和苯甲醛对根系造成伤害(杨兴洪和罗新书, 1991 ), 植株残体在淹水缺氧条件下分解产生酚酸类物质(Patrick, 1971 ; Tang & Young, 1982 ).有研究发现: 地黄(Rehmannia glutinosa )连作土壤中酚酸类物质含量提高, 导致脲酶,蔗糖酶活性升高, 过氧化氢酶活性降低(陈慧等, 2007 ); 西瓜园酚酸类物质含量升高后, 土壤过氧化氢酶,脲酶活性均提高(吕卫光等, 2006 ).酚酸类物质含量增加, 导致杉木(Cunningh- amia lanceolata )苗叶绿素合成受阻, 光合作用减弱(陈秀华等, 2002 ), 对绿豆(Vigna radiata ),大豆(Glycine max ),黄瓜(Cucumis sativus )和玉米种子发芽和生长有抑制作用, 且随浓度升高抑制作用增强(曹光球等, 2003 ).酚酸类物质和苦杏仁苷都是造成果树连作障碍的重要因素(Patrick, 1955 ; Bӧrner, 1960 ).有研究表明, 棉田土壤生物活性受到作物连作障碍的负面效应和秸秆长期还田培肥地力的正面效应的双重影响(刘建国等, 2008 ).因此桃树枝条还田也可能会对桃树植株生长产生有利的影响. ...
HPLC法测定不同产地桃仁中苦杏仁苷的含量
2
2011
... 枝条烘干磨碎过20目筛后取0.5 g, 参照索氏提取法和高效液相色谱法(叶晶晶, 2011 )测得枝条中苦杏仁苷含量; 取T4处理中枝条浸提液5 mL过0.22 μm有机相滤膜备用, 参照高效液相色谱法(尹承苗等, 2013 )测得枝条浸提液中的酚酸类物质含量. ...
... 测定土壤中苦杏仁苷所用土样为2014年6月所取, 准确称取过20目筛的风干土样1.0 g, 后采用索氏提取法(乙醚2 h去杂质和甲醇6 h提取)得到100 mL提取液, 用50%甲醇溶液溶解至刻度200 mL, 摇匀后超声萃取, 过0.22 μm有机相滤膜, 待高效液相色谱分析.色谱条件: 色谱柱为Acclaim 120 C18 (5 μm, 250 mm × 4.6 mm), 柱温25 ℃.流动相为乙腈:水= 17:83, 流速1.0 mL·min-1 ; 进样方式及体积: 自动进样10 μL; 检测波长210 nm (叶晶晶, 2011 ). ...
一种分析土壤中酚酸类物质含量的新方法--以连作苹果园土壤为试材
2
2013
... 枝条烘干磨碎过20目筛后取0.5 g, 参照索氏提取法和高效液相色谱法(叶晶晶, 2011 )测得枝条中苦杏仁苷含量; 取T4处理中枝条浸提液5 mL过0.22 μm有机相滤膜备用, 参照高效液相色谱法(尹承苗等, 2013 )测得枝条浸提液中的酚酸类物质含量. ...
... 测定土壤酚酸所用土样为2014年6月所取, 准确称取过12目筛的风干土样80 g, 加入适量的硅藻土于烧杯中混匀.在100 mL萃取池底部垫上1片纤维素膜, 将混匀的样品装入萃取池中, 按优化好的ASE条件萃取: 提取溶剂为甲醇和无水乙醇, 提取温度为120 ℃, 提取2次, 加热5 min, 静态提取时间为5 min, 吹扫体积为60%, 吹扫90 s.萃取完成后, 34 ℃减压经RE201D型旋转蒸发仪(RE201D, 上海耀裕仪器设备有限公司, 上海)浓缩近干, 加入3 mL甲醇复溶, 过0.22 μm有机相滤膜, 待高效液相色谱(HPLC)分析.色谱条件: 色谱柱是Acclaim 120 C18 (3 μm, 150 mm × 3 mm), 柱温30 ℃.流动相是 A-乙腈, B-水(乙酸调pH值至2.6), 乙腈:水= 95:5, 流速0.5 mL·min-1 ; 进样方式及体积: 自动进样5 μL; 检测波长280 nm (尹承苗等, 2013 ). ...
连作对再植桃树幼苗生长及叶片性状的影响
1
2012
... 前人研究结果显示, 土壤自毒物质对不同植株表现出类似的抑制作用.外源添加肉桂酸,苯甲酸,对羟基苯甲酸, 抑制了杉木幼苗胚根和胚芽的生长(汪思龙等, 2002 ).土壤酚酸含量增加, 玉米幼苗的株高和根系生长均受到抑制(郑皓皓等, 2001 ; 胡晓军等, 2011 ).对羟基苯甲酸,香草酸和香豆酸的累积能够抑制花生幼苗生长, 提高花生发病率(李培栋等, 2010 ).桃树根系浸提液对桃树生长指标表现为低浓度促进,高浓度抑制的质量浓度效应(张斌斌等, 2012 ).本试验结果表明, 常规修剪量(1.5 g·kg-1 )还田对桃幼树生长无显著影响, 而加量枝条(22.5 g·kg-1 )还田时, 土壤酚酸总量达到1700 ng·g-1 , 土壤苦杏仁苷含量2.30 mg·g-1 , 出现了显著抑制现象, 如叶绿素含量降低,新梢停长期提前,地径生长量减小和净光合速率降低. ...
桃树体内苦杏仁甙含量年变化研究
1
2006
... 桃树再植当年基本不生新根, 发枝短长势弱, 造成桃园减产, 连作障碍严重(刘嘉彬等, 2006 ).桃树根系,枝条,叶片,果肉及桃仁中都有不同含量的苦杏仁苷(赵宇瑛等, 2006 ), 桃树残根在土壤中降解产生大量的苦杏仁苷, 其水解生成的氢氰酸和苯甲醛对根系造成伤害(杨兴洪和罗新书, 1991 ), 植株残体在淹水缺氧条件下分解产生酚酸类物质(Patrick, 1971 ; Tang & Young, 1982 ).有研究发现: 地黄(Rehmannia glutinosa )连作土壤中酚酸类物质含量提高, 导致脲酶,蔗糖酶活性升高, 过氧化氢酶活性降低(陈慧等, 2007 ); 西瓜园酚酸类物质含量升高后, 土壤过氧化氢酶,脲酶活性均提高(吕卫光等, 2006 ).酚酸类物质含量增加, 导致杉木(Cunningh- amia lanceolata )苗叶绿素合成受阻, 光合作用减弱(陈秀华等, 2002 ), 对绿豆(Vigna radiata ),大豆(Glycine max ),黄瓜(Cucumis sativus )和玉米种子发芽和生长有抑制作用, 且随浓度升高抑制作用增强(曹光球等, 2003 ).酚酸类物质和苦杏仁苷都是造成果树连作障碍的重要因素(Patrick, 1955 ; Bӧrner, 1960 ).有研究表明, 棉田土壤生物活性受到作物连作障碍的负面效应和秸秆长期还田培肥地力的正面效应的双重影响(刘建国等, 2008 ).因此桃树枝条还田也可能会对桃树植株生长产生有利的影响. ...
秸秆还田与施肥对土壤酶活性和作物产量的影响
1
2012
... 土壤酶活性是研究土壤质量的重要指标.在小麦-玉米轮作情况下, 秸秆还田提高了土壤酶活性(路文涛等, 2011 ; 甄丽莎等, 2012 ).杨树林土壤酚酸浓度升高后, 土壤脲酶和碱性磷酸酶活性降低, 过氧化氢酶活性则升高(王延平等, 2013 ).肖宏(2004)研究发现连作苹果土壤中酚酸类物质含量升高后, 基础酶活性降低.本试验结果表明, T1和T3处理的土壤酶活性与CK无显著差异.T2和T4处理的土壤蔗糖酶活性始终高于CK.处理前期, 土壤过氧化氢酶和脲酶活性均显著高于CK, 处理后期均显著低于CK.蔗糖酶又称转化酶, 与土壤有机质含量呈正相关, 可能是因为枝条腐解产生有机质, 引起蔗糖酶活性升高.脲酶促进尿素氮肥转化成氨, 过氧化氢酶可以清除土壤中的过氧化氢.处理后期脲酶和过氧化氢酶活性降低, 减少了植株的直接氮素来源, 造成土壤中过氧化氢的积累而不利于桃树植株的 生长. ...
麦秸还田耕层酚酸变化及其对夏玉米生长的影响
2
2001
... 前人研究结果显示, 连作或秸秆还田的土壤中, 土壤酚酸类物质有明显提高.麦秸还田40天后, 玉米土壤耕作层酚酸含量达到最高峰(郑皓皓等, 2001 ; 胡晓军等, 2011 ), 花生(Arachis hypogaea )和黄瓜连作后土壤酚酸含量均显著升高(马云华等, 2005 ; 李培栋等, 2010 ), 连作苹果园土壤中酚酸物质总量显著提高(王青青, 2012 ).与其他作物不同, 桃树残根降解会产生苦杏仁苷, 经土壤酶分解产生的氢氰酸,苯甲醛和苯甲酸, 对新茬桃树根系产生巨大毒害(Baziramakenga et al ., 1995 ).本试验结果表明, 枝条浸提液中含有酚酸类物质和高量的苦杏仁苷, 加量枝条还田后, 土壤中酚酸类物质和苦杏仁苷含量与对照相比分别提高约30%和20%, 加量桃树枝条(22.5 g·kg-1 )的加入同样增加自毒物质的累积.因为盆栽桃树添加枝条后, 露天放置, 剪碎枝条经过雨水和灌溉水浸泡后同样会产生大量此类物质, 这与有关研究中植株残体淹水缺氧可分解产生挥发性脂肪酸和酚酸类(Patrick, 1971 ; Tang & Young, 1982 )的结果是一致的. ...
... 前人研究结果显示, 土壤自毒物质对不同植株表现出类似的抑制作用.外源添加肉桂酸,苯甲酸,对羟基苯甲酸, 抑制了杉木幼苗胚根和胚芽的生长(汪思龙等, 2002 ).土壤酚酸含量增加, 玉米幼苗的株高和根系生长均受到抑制(郑皓皓等, 2001 ; 胡晓军等, 2011 ).对羟基苯甲酸,香草酸和香豆酸的累积能够抑制花生幼苗生长, 提高花生发病率(李培栋等, 2010 ).桃树根系浸提液对桃树生长指标表现为低浓度促进,高浓度抑制的质量浓度效应(张斌斌等, 2012 ).本试验结果表明, 常规修剪量(1.5 g·kg-1 )还田对桃幼树生长无显著影响, 而加量枝条(22.5 g·kg-1 )还田时, 土壤酚酸总量达到1700 ng·g-1 , 土壤苦杏仁苷含量2.30 mg·g-1 , 出现了显著抑制现象, 如叶绿素含量降低,新梢停长期提前,地径生长量减小和净光合速率降低. ...
 , 蒋晓梅
, 蒋晓梅

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}