Responses of growth of four desert species to different N addition levels
HUANG Ju-Ying1, *, , YU Hai-Long2
1Center of New Technology Application and Research, Ningxia University, Yinchuan 750021, China2College of Resources and Environment, Ningxia University, Yinchuan 750021, China
Aims The increase in atmospheric N deposition has accelerated N cycling of ecosystems, thus altering the structure and function of ecosystems, especially in those limited by N availability. Studies on the response of plant growth to artificial N addition could provide basic data for a better understanding of how the structure of grasslands in northern China responds to increasing N deposition. Methods We investigated the seasonal dynamics of plant growth of four species after 2-year multi-level N addition in a field experiment conducted in a desert steppe of Ningxia in 2011. Plant biomass and the relative growth rate (RGR) of the studied species were measured and their relationships with C:N:P ratios of plants (community and leaf levels) and soils were analyzed. Important findings Results in 2012 showed that 2-year N addition promoted the growth of the four species and the effects were different among growth forms and were species-specific. In general, the plant biomass of the studied species was significantly correlated with leaf N concentration, leaf N:P ratio, community N pool, soil total N content and soil N:P ratio, while only weak relationships were observed between plant biomass and C:N and C:P ratios of plants and soils. In contrast, there was a significant linear relationship between RGR and N:P ratios both of plants and soils.Our results suggest that short-term N addition promoted the accumulation of plant biomass, and the species-specific responses to stimulated N addition can directly affect the structure of the desert steppe ecosystem. Plant N:P ratio and soil N:P ratio could indicate nutrient limitation of plant growth to a certain extent: N addition increased soil N content and N:P ratio, and thus relieved N limitation gradually. Once more N is available to plants, the growth of plants and the accumulation of community N was stimulated in turn.
Keywords:C:N:P ecological stoichiometry
;
desert steppe
;
relative growth rate
;
increase in atmospheric N deposition
;
population biomass
HUANGJu-Ying, YUHai-Long. Responses of growth of four desert species to different N addition levels[J]. Chinese Journal of Plant Ecology, 2016, 40(2): 165- https://doi.org/10.17521/cjpe.2015.0210
Fig. 1 Effects of N addition on plant biomass of the four species (mean ± SE, n = 5). F0, F2.5, F5, F10, F20, and F40 represent N addition level at 0.00, 2.50, 5.00, 10.00, 20.00, and 40.00 g·m-2·a-1, respectively. Different lowercase letters indicate significant differences (p < 0.05) between population biomass within N levels. The same lowercase letters indicate insignificant differences (p > 0.05). A, B and C are data of Lespedeza potaninii in June, July, and August, respectively. D, E and F are data of Cynanchum komarovii in June, July, and August, respectively. G, H and I are data of Stipa capillata in June, July, and August, respectively. J, K and L are data of Agropyron cristatum in June, July, and August, respectively.
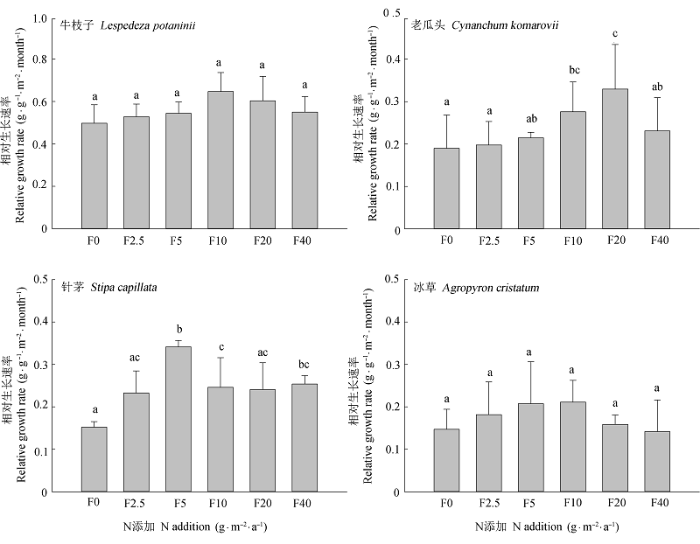
Fig. 2 Effects of N addition on the relative growth rate (from June to August) of the four species (mean ± SE, n = 5). F0, F2.5, F5, F10, F20, and F40 represent N addition level at 0.00,2.50,5.00,10.00,20.00, and 40.00 g·m-2·a-1, respectively. Different lowercase letters indicate significant differences (p < 0.05) between relative growth rate among N addition levels. The same lowercase letters indicate insignificant differences (p > 0.05).
Table 1 Correlation coefficients between the C:N:P ratio of plant leaves and the plant biomass or the relative growth rate from June to August of the four species
指标 Index
全C浓度 Total C
全N浓度 Total N
全P浓度 Total P
C:N
C:P
N:P
牛枝子 Lespedeza potaninii
种群生物量 Population biomass
-0.647
0.585
0.178
-0.734
-0.583
0.824*
相对生长速率 Relative growth rate
-0.564
0.461
0.031
-0.618
-0.443
0.889*
老瓜头 Cynanchum komarovii
种群生物量 Population biomass
-0.541
0.887*
-0.494
-0.865*
-0.314
0.988**
相对生长速率 Relative growth rate
-0.558
0.555
-0.709
-0.708
-0.169
0.802*
冰草 Agropyron cristatum
种群生物量 Population biomass
-0.276
0.499
0.667
-0.561
-0.655
0.245
相对生长速率 Relative growth rate
0.669
-0.321
0.049
0.483
0.390
-0.585
针茅 Stipa capillata
种群生物量 Population biomass
-0.477
0.950**
0.655
-0.836*
-0.508
-0.589
相对生长速率 Relative growth rate
-0.869*
0.288
0.691
-0.686
-0.920**
-0.426
*, p < 0.05; ** p < 0.01. Total C, total N, and total P: mg·g-1; Population biomass: g·m-2; Relative growth rate: g·g-1·m-2·month-1.*, p < 0.05; ** p < 0.01.全C,全N和全P浓度单位: mg·g-1; 种群生物量单位: g·m-2; 相对生长速率单位: g·g-1·m-2·month-1.
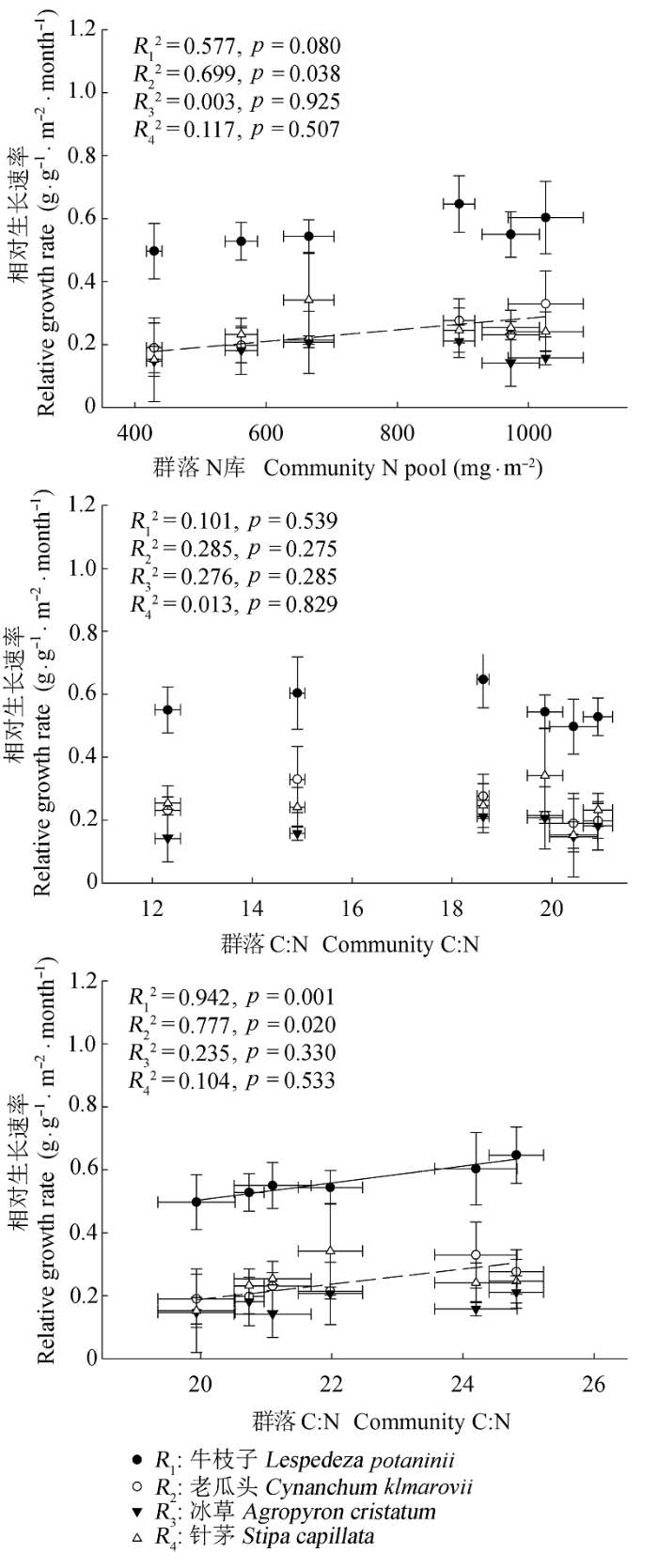
图5 4个物种6-8月相对生长速率与群落N库C:N和N:P的关系(平均值±标准误差, n = 5).
Fig. 5 Relationships between the relative growth rate of the four specie (from June to August) and community N pool, C:N ratio, and N:P ratio (mean ± SE, n = 5).
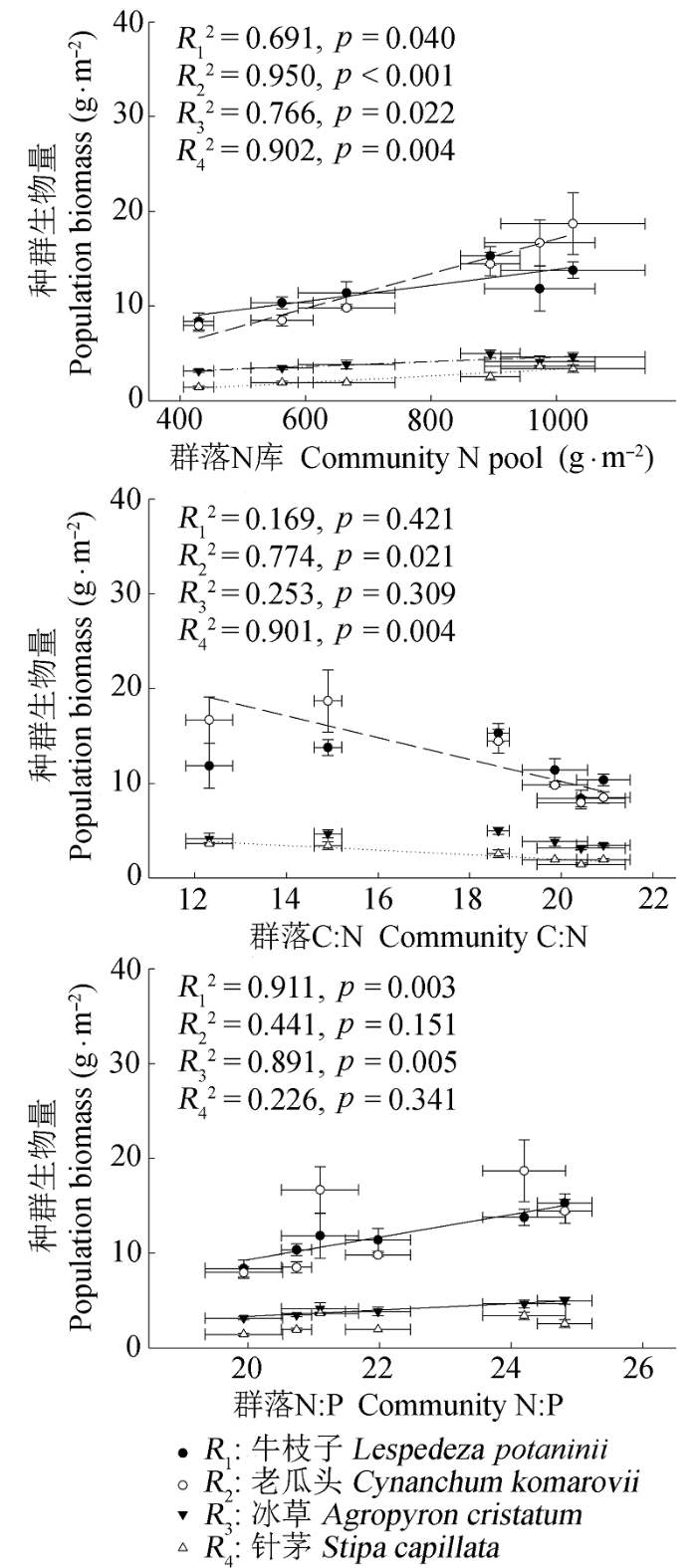
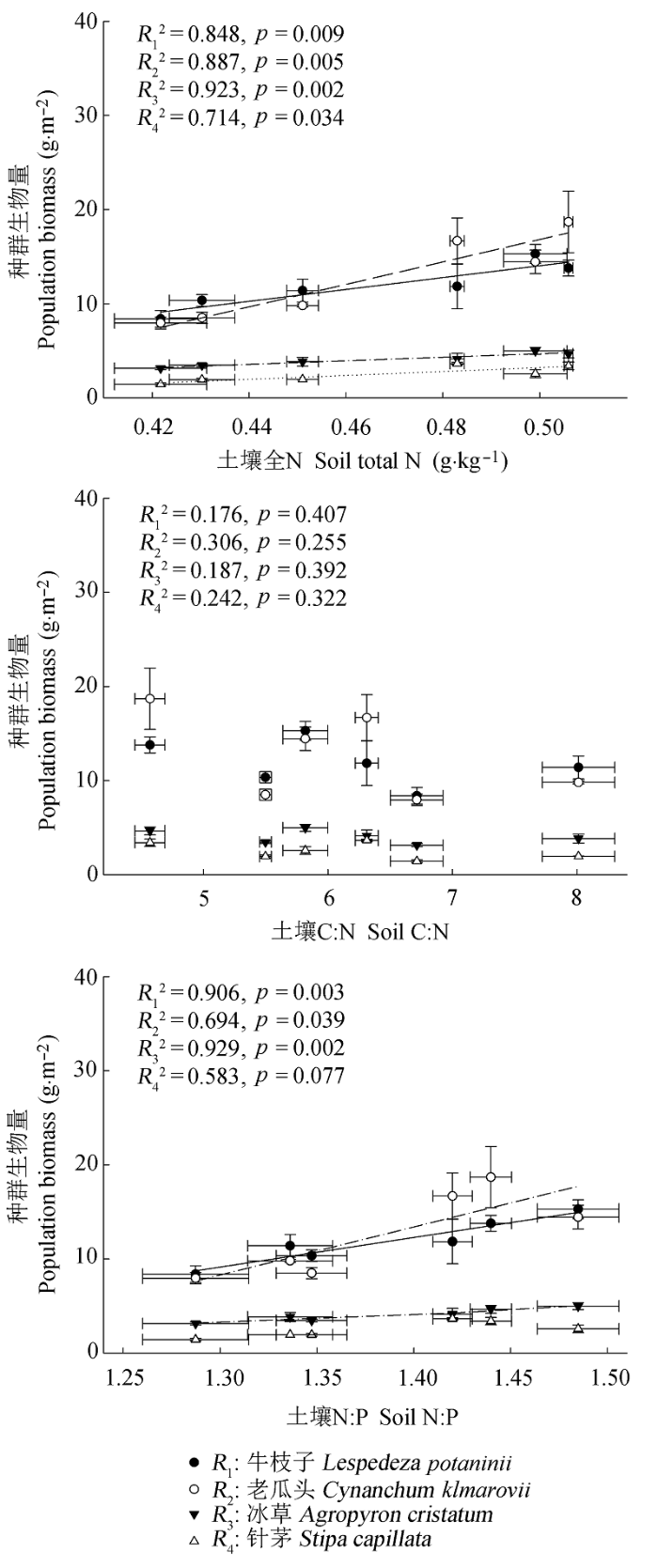
图6 8月份4个物种种群生物量与土壤全N含量,C:N和N:P的线性关系(平均值±标准误差, n = 5).
Fig. 6 Relationships between the population biomass of the four species and soil total N content, C:N ratio and N:P ratio in August (mean ± SE, n = 5).
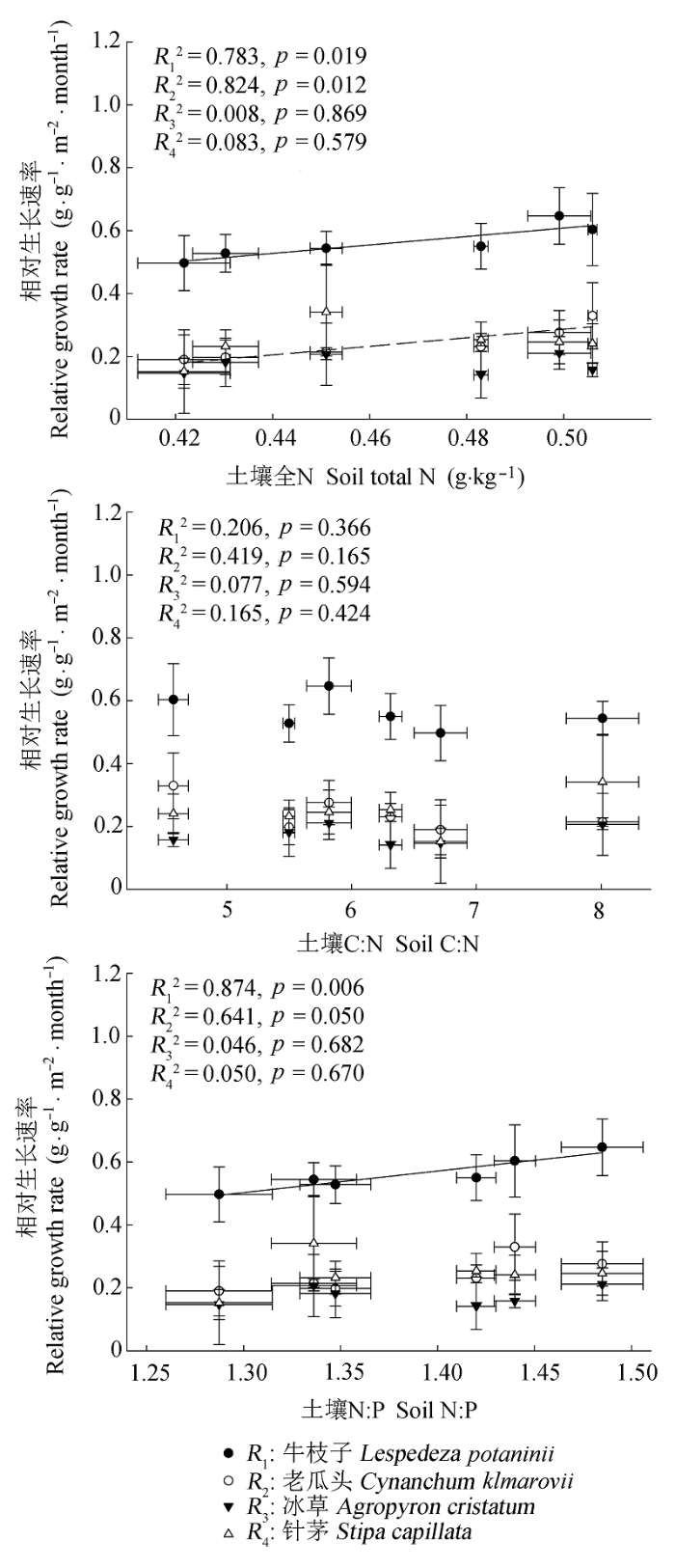
图7 4个物种6-8月相对生长速率与土壤全N含量,C:N和N:P的线性关系(平均值±标准误差, n = 5).
Fig. 7 Relationships between the relative growth rate of the four species (from June to August) and soil total N content, C:N ratio, and N:P ratio (mean ± SE, n = 5).
Delgado-Baquerizo M, Maestre FT, Gallardol A, Bowker MA, Wallenstein MD, Quero JL, Ochoa V, Gozalo B, García-Gómez M, Soliveres S, García-Palacios P, Berdugo M, Valencia E, Escolar C, Arredondo T, Barraza-Zepeda C, Bran D, Carreira JA, Chaieb M, Conceicão AA, Derak M, Eldridge DJ, Escudero A, Espinosa CI, Gaitán J, Gatica MG, Gómez-González S, Guzman E, Gutiérrez JR, Florentino A, Hepper E, Hernández RM, Huber-Sannwald E, Jankju M, Liu J, Mau RL, Miriti M, Monerris J, Naseri K, Noumi Z, Polo V, Prina A, Pucheta E, Ramírez ERamírez-Collantes DA, Romão R, Tighe M, Torres D, Torres-Díaz C, Ungar ED, Val J, Wamiti W, Wang D, Zaady E(2013).
Decoupling of soil nutrient cycles as a function of aridity in global drylands.
2012). Shifts in plant community structure of a threatened sandy grassland over a 9-yr period under experimentally induced nutrient regimes: Is there a lag phase?
Journal of Vegetation Science, Journal of Vegetation Science, 23, 372-386.
Contribution of acidification and eutrophication to declines in species richness of calcifuge grasslands along a gradient of atmospheric nitrogen deposition.
Combined effects of simulated nitrogen deposition and drought stress on growth and photosynthetic physiological responses of two annual desert plants in Junggar Basin, China.
Chinese Journal of Plant Ecology, 34, 1394-1403. (in Chinese with English abstract).[周晓兵, 张元明, 王莎莎, 张丙昌 (2010).
Nutrient limitation in three lowland tropical forests in southern China receiving high nitrogen deposition: Insights from fine root responses to nutrient additions.
PLoS ONE, 8, e82661.
The C: N: P stoichiometry of autotrophs: Theory and observations.
Biological stoichiometry of plant production: Metabolism, scaling and ecological response to global change.
1
2010
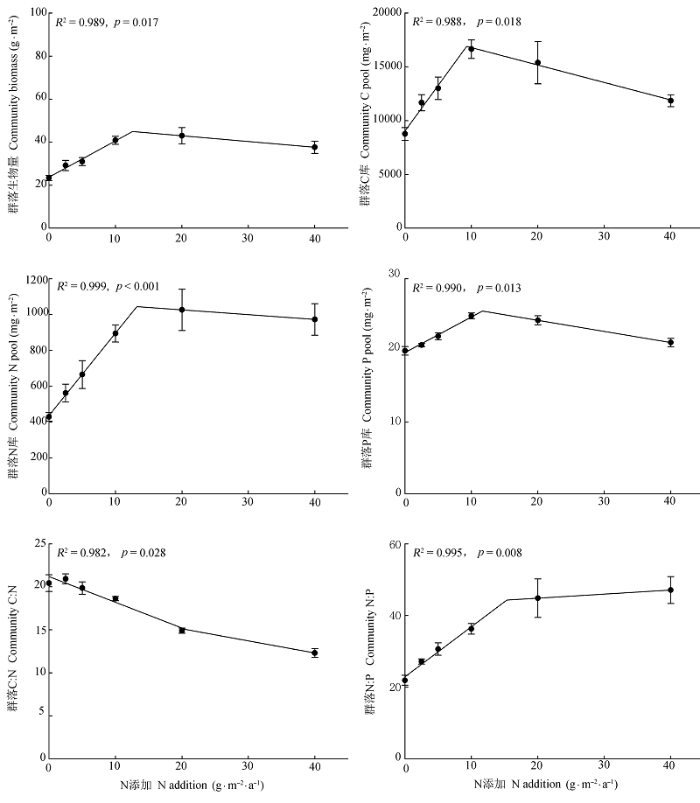
... 生态化学计量学作为研究C,N,P循环的新思路和新手段, 已引起国内外研究者的广泛关注.植物和土壤C,N,P及其生态化学计量比可以作为生态系统C,N,P饱和的诊断和有效性预测的指标.植物C:N:P在一定程度上可指示所在生态系统的C积累动态和N,P养分限制格局(Elser et al., 2010).其中, 叶片C:N和C:P代表植物吸收营养元素时所能同化C的能力, 反映了植物对N和P的利用效率, 同时也代表着不同群落或植物固C效率的高低.对植物来说, 高的生长速率对应低的C:N和C:P (He et al., 2008).这种相对应的关系与养分限制类型有一定的关系: 当受N限制时, 生长速率与C:N负相关; 当受P限制时, 生长速率与C:P负相关(Yu et al., 2012).土壤C:N:P是反映土壤内部C,N,P循环的主要指标, 综合了生态系统功能的变异性, 而且有助于确定生态过程对全球变化的响应, 因而成为确定土壤C,N,P平衡特征的一个重要参数(王绍强和于贵瑞, 2008; Tian et al., 2010).一般而言, 土壤C:N和C:P可以表征有机质状况,凋落物分解质量及其对土壤肥力的潜在贡献.土壤N:P则由于对外源N或者P增加反应敏感, 被认为可以指示生态系统养分受限性(Peňuelas et al., 2012).Tian等(2010)统计了我国2473份0-10 cm层土壤C:N,C:P和N:P等的变化特征, 认为表层土壤C:N和N:P可以很好地指示土壤N供给状况.然而到目前为止, 土壤C:N:P还没有被广泛地应用于生态系统养分受限性的预测中, 尤其是对于大气N沉降增加敏感的荒漠草原. ...
2012). Shifts in plant community structure of a threatened sandy grassland over a 9-yr period under experimentally induced nutrient regimes: Is there a lag phase?
... 通过对我国0-10 cm层土壤C:N:P计量比变化特征的综合分析, Tian等(2010)认为表层土壤C:N和N:P能够很好地指示土壤N的供给状况.Yang等(2014)的研究进一步表明, 表层土壤C:N相对稳定, 而N:P的变异性相对较大, 证实了全球气候变化会逐渐改变土壤N和P之间的耦合关系(Delgado- Baquerizo et al., 2013).因此, 与C:N相比, 土壤N:P能够较好地指示生态系统的养分受限性.本研究中, 几个物种种群生物量和6-8月相对生长速率均与0-10 cm土壤全N含量和N:P存在不同程度的正相关, 而与土壤C:N不存在相关性, 表明随着土壤N有效性的增加, 土壤N含量和N:P有相继升高的趋势, 几个荒漠草原物种受N的限制性逐渐缓解, 因而可供植物摄取的N增多, 促进了植物群落生长和生物量积累.因此, 土壤N:P的变化状况将直接影响到荒漠草原植物生长和养分利用策略.由于长期,大量大气N沉降增加会显著提高土壤N:P (Bobbink et al. 1998; Menge & Field 2007; Vitousek et al., 2010), 如果能够进行适当人为干扰(例如P添加), 可能会通过改变土壤N:P, 削弱大气N沉降增加对生态系统群落结构的不利影响. ...
鼎湖山苗圃和主要森林土壤CO2排放和CH4吸收对模拟N沉降的短期响应
1
2005
... 工业革命以来, 化石矿物焚烧,干物质燃烧,毁林开荒,化肥施用和豆科植物广泛栽植等人类活动产生了大量含氮(N)化合物, 导致全球大气N沉降增加(IPCC, 2013).据估计, 1860-2000年全球活性N生产提高了11倍(Galloway et al., 2003), 且随着社会经济和工农业的进一步发展, 将保持持续升高的趋势.在一些发达国家, 活性N沉降速率较工业革命前提高了7倍(Clark & Tilman, 2008).在我国, 1980-2010年N沉降平均以0.41 kg·hm-2·a-1的速率增加, 并且在未来数十年内还将继续增加(Liu et al., 2013).国内外相关研究表明, 在受N限制的生态系统, 少量或短期N沉降增加对系统各组分具有明显的促进作用.例如, 促进了植物N摄取能力和光合作用, 提高了植被生产力和碳(C)储备(李林森等, 2015), 刺激了N矿化和磷酸酶活性, 因此提高了土壤N和磷(P)的有效性(Jones & Power, 2012), 促进了微生物-酶系统对土壤有机质,N和P的分解, 加速了凋落物分解(Liu et al., 2010).然而, 随着过多N的输入, 不仅其他资源(主要为P)将成为限制植物生长的因子(Bobbink et al., 2010), 且由于土壤中保持了大量的硝态N, 增加了养分淋溶和土壤酸化的潜在危险, 导致生物多样性降低,生态系统退化(Stevens et al., 2010; Wardle et al., 2013; Stark et al., 2014). ...
The human-induced imbalance between C, N and P in Earth's life system.
2
2012
... 生态化学计量学作为研究C,N,P循环的新思路和新手段, 已引起国内外研究者的广泛关注.植物和土壤C,N,P及其生态化学计量比可以作为生态系统C,N,P饱和的诊断和有效性预测的指标.植物C:N:P在一定程度上可指示所在生态系统的C积累动态和N,P养分限制格局(Elser et al., 2010).其中, 叶片C:N和C:P代表植物吸收营养元素时所能同化C的能力, 反映了植物对N和P的利用效率, 同时也代表着不同群落或植物固C效率的高低.对植物来说, 高的生长速率对应低的C:N和C:P (He et al., 2008).这种相对应的关系与养分限制类型有一定的关系: 当受N限制时, 生长速率与C:N负相关; 当受P限制时, 生长速率与C:P负相关(Yu et al., 2012).土壤C:N:P是反映土壤内部C,N,P循环的主要指标, 综合了生态系统功能的变异性, 而且有助于确定生态过程对全球变化的响应, 因而成为确定土壤C,N,P平衡特征的一个重要参数(王绍强和于贵瑞, 2008; Tian et al., 2010).一般而言, 土壤C:N和C:P可以表征有机质状况,凋落物分解质量及其对土壤肥力的潜在贡献.土壤N:P则由于对外源N或者P增加反应敏感, 被认为可以指示生态系统养分受限性(Peňuelas et al., 2012).Tian等(2010)统计了我国2473份0-10 cm层土壤C:N,C:P和N:P等的变化特征, 认为表层土壤C:N和N:P可以很好地指示土壤N供给状况.然而到目前为止, 土壤C:N:P还没有被广泛地应用于生态系统养分受限性的预测中, 尤其是对于大气N沉降增加敏感的荒漠草原. ...
... 大气N沉降增加, 是人类正面临的一个全球性气候变化问题, 其生态学效应受到越来越多科研工作者的关注.对于我国草原生态系统而言, 尽管已监测到的N沉降量低于其他生态系统, 但是草原生态系统本身N沉降临界负荷低(段雷等, 2002), 而且即使是低剂量的N添加, 其长期的影响也是巨大的(Clark & Tilman, 2008).对于一些受P限制的生态系统(例如热带森林和酸性泥炭沼泽), 随着过多N的输入, 不仅导致这些生态系统N富集, 而且可能会引起N和P不平衡, 造成系统P压力增加(Phuyal et al., 2008; Zhu et al., 2013),植物生长P受限性增强(Yu et al., 2012), 进而对生态系统结构和功能产生深远影响.与这些生态系统不同, 荒漠草原具有较低土壤N:P (Delgado-Baquerizo et al., 2013), 大气N沉降增加将有利于该生态系统稳定性的维持, 还是会导致系统P压力增加, 这是值得我们深入研究的一个问题.因此, 今后的研究工作需要重点考虑大气N沉降增加对荒漠草原结构和功能的长期影响. ...
Long-term nitrogen deposition increases phosphorus limitation of bryophytes in an ombrotrophic bog.
1
2008
... 工业革命以来, 化石矿物焚烧,干物质燃烧,毁林开荒,化肥施用和豆科植物广泛栽植等人类活动产生了大量含氮(N)化合物, 导致全球大气N沉降增加(IPCC, 2013).据估计, 1860-2000年全球活性N生产提高了11倍(Galloway et al., 2003), 且随着社会经济和工农业的进一步发展, 将保持持续升高的趋势.在一些发达国家, 活性N沉降速率较工业革命前提高了7倍(Clark & Tilman, 2008).在我国, 1980-2010年N沉降平均以0.41 kg·hm-2·a-1的速率增加, 并且在未来数十年内还将继续增加(Liu et al., 2013).国内外相关研究表明, 在受N限制的生态系统, 少量或短期N沉降增加对系统各组分具有明显的促进作用.例如, 促进了植物N摄取能力和光合作用, 提高了植被生产力和碳(C)储备(李林森等, 2015), 刺激了N矿化和磷酸酶活性, 因此提高了土壤N和磷(P)的有效性(Jones & Power, 2012), 促进了微生物-酶系统对土壤有机质,N和P的分解, 加速了凋落物分解(Liu et al., 2010).然而, 随着过多N的输入, 不仅其他资源(主要为P)将成为限制植物生长的因子(Bobbink et al., 2010), 且由于土壤中保持了大量的硝态N, 增加了养分淋溶和土壤酸化的潜在危险, 导致生物多样性降低,生态系统退化(Stevens et al., 2010; Wardle et al., 2013; Stark et al., 2014). ...
Nutrient availab- ility and pH jointly constrain microbial extracellular enzyme activities in nutrient-poor tundra soils.
1
2014
... 工业革命以来, 化石矿物焚烧,干物质燃烧,毁林开荒,化肥施用和豆科植物广泛栽植等人类活动产生了大量含氮(N)化合物, 导致全球大气N沉降增加(IPCC, 2013).据估计, 1860-2000年全球活性N生产提高了11倍(Galloway et al., 2003), 且随着社会经济和工农业的进一步发展, 将保持持续升高的趋势.在一些发达国家, 活性N沉降速率较工业革命前提高了7倍(Clark & Tilman, 2008).在我国, 1980-2010年N沉降平均以0.41 kg·hm-2·a-1的速率增加, 并且在未来数十年内还将继续增加(Liu et al., 2013).国内外相关研究表明, 在受N限制的生态系统, 少量或短期N沉降增加对系统各组分具有明显的促进作用.例如, 促进了植物N摄取能力和光合作用, 提高了植被生产力和碳(C)储备(李林森等, 2015), 刺激了N矿化和磷酸酶活性, 因此提高了土壤N和磷(P)的有效性(Jones & Power, 2012), 促进了微生物-酶系统对土壤有机质,N和P的分解, 加速了凋落物分解(Liu et al., 2010).然而, 随着过多N的输入, 不仅其他资源(主要为P)将成为限制植物生长的因子(Bobbink et al., 2010), 且由于土壤中保持了大量的硝态N, 增加了养分淋溶和土壤酸化的潜在危险, 导致生物多样性降低,生态系统退化(Stevens et al., 2010; Wardle et al., 2013; Stark et al., 2014). ...
Contribution of acidification and eutrophication to declines in species richness of calcifuge grasslands along a gradient of atmospheric nitrogen deposition.
... 大气N沉降增加, 是人类正面临的一个全球性气候变化问题, 其生态学效应受到越来越多科研工作者的关注.对于我国草原生态系统而言, 尽管已监测到的N沉降量低于其他生态系统, 但是草原生态系统本身N沉降临界负荷低(段雷等, 2002), 而且即使是低剂量的N添加, 其长期的影响也是巨大的(Clark & Tilman, 2008).对于一些受P限制的生态系统(例如热带森林和酸性泥炭沼泽), 随着过多N的输入, 不仅导致这些生态系统N富集, 而且可能会引起N和P不平衡, 造成系统P压力增加(Phuyal et al., 2008; Zhu et al., 2013),植物生长P受限性增强(Yu et al., 2012), 进而对生态系统结构和功能产生深远影响.与这些生态系统不同, 荒漠草原具有较低土壤N:P (Delgado-Baquerizo et al., 2013), 大气N沉降增加将有利于该生态系统稳定性的维持, 还是会导致系统P压力增加, 这是值得我们深入研究的一个问题.因此, 今后的研究工作需要重点考虑大气N沉降增加对荒漠草原结构和功能的长期影响. ...
Nutrient limitation in three lowland tropical forests in southern China receiving high nitrogen deposition: Insights from fine root responses to nutrient additions.
 , 余海龙
, 余海龙

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}