Spatial patterns of species diversity in the herb layer of early post-fire regeneration in mixed Pinus yunnanensis forests
HAN Jie1, YING Ling-Xiao1, LI Gui-Xiang2, SHEN Ze-Hao1, *,
1Department of Ecology, College of Urban & Environmental Sciences, the Key Laboratory of the Ministry of Education on Earth Surface Processes, Peking University, Beijing 100871, China2Forest Ecology Institute, Yunnan Academy of Forestry, Kunming 650204, China
通讯作者: * 通讯作者Author for correspondence (E-mail: shzh@pku.edu.cn
Herb layer plays an important role in maintaining ecosystem functioning of forests. The aims of this study were to determine the pattern of species diversity in early post-fire regeneration of the herb layer in mixed Yunnan pine forests and to identify the effects of topography, fire severity, pre-fire vegetation and herb species life-history feature on post-fire regeneration.
Methods
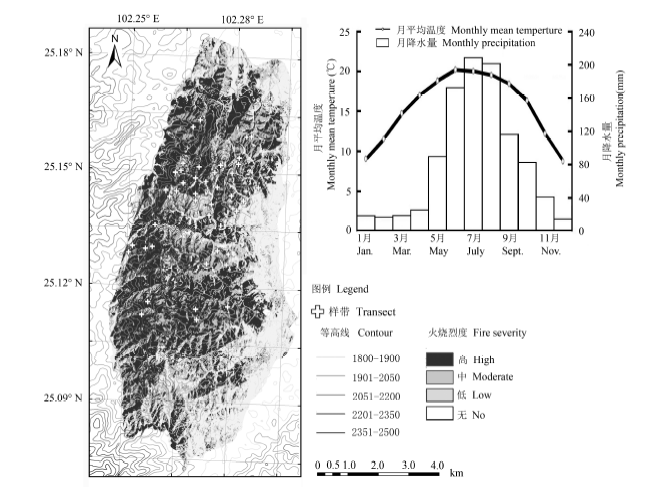
We conducted field investigations of post-fire regeneration by randomly setting 47 transects of 10 m × 100 m in a burned forest stand in Qinfeng Township, Lufeng County, Yunnan Province, from September to October, 2013. The Two-Way Indicator Species Analysis (TWINSPAN) was used to classify the herb community types from the 47 transects, and species richness and two indexes of β-diversity were used to analyze herb species diversity and compositional change within sampling transects. Generalized linear model and hierarchical variation partitioning were applied to estimate factors affecting the pattern of herb species diversity of the post-fire communities.
Important findings
Four herb community types were identified for the 47 transects, which are differentiated by topographic features and fire severities. The species richness in the herb layer of early post-fire regeneration was primarily affected by elevation, topography, fire severity and the pre-fire forest canopy. Specifically, the species richness decreased with increasing elevation, and was positively correlated with biomass of the standing trees and steepness of site slope and negatively correlated with slope position. However, the species richness of herbs was not correlated with fire severity significantly. The within community β-diversity of the post-fire herb layer had a positive correlation with both basal area of pre-fire canopy trees and fire severity. Moreover, the β-diversity was lower at higher elevations. The results also highlighted the differences between annual and perennial species in their distribution patterns.
Keywords:post-fire regeneration
;
herb diversity
;
species composition
;
pre-fire vegetation
;
topography
;
fire severity
HANJie, YINGLing-Xiao, LIGui-Xiang, SHENZe-Hao. Spatial patterns of species diversity in the herb layer of early post-fire regeneration in mixed Pinus yunnanensis forests[J]. Chinese Journal of Plant Ecology, 2016, 40(3): 200-211 https://doi.org/10.17521/cjpe.2015.0161
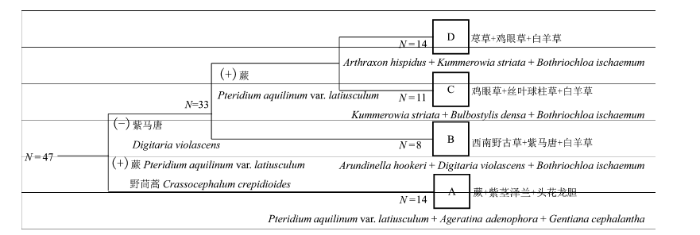
Fig. 2 The classification diagram of TWINSPAN for the herb layer of post-fire regenerated communities. N represents the number of transects. A, B, C, and D designate four types of post-fire regenerated herb communities. A, Pteridium aquilinum var. latiusculum + Ageratina adenophora + Gentiana cephalantha community; B, Arundinella hookeri + Digitaria violascens + Bothriochloa ischaemum community; C, Kummerowia striata + Bulbostylis densa + Bothriochloa ischaemum community; D, Arthraxon hispidus + Kummerowia striata + Bothriochloa ischaemum community. (+) means it is positive with classification; (-) means it is negetive with classification.
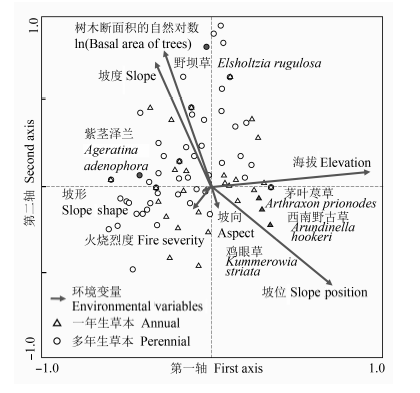
Fig. 3 The canonical correspondence analysis ordination pattern for the annual (∆) and perennial (○) plant species in the early post-fire regenerated communities.
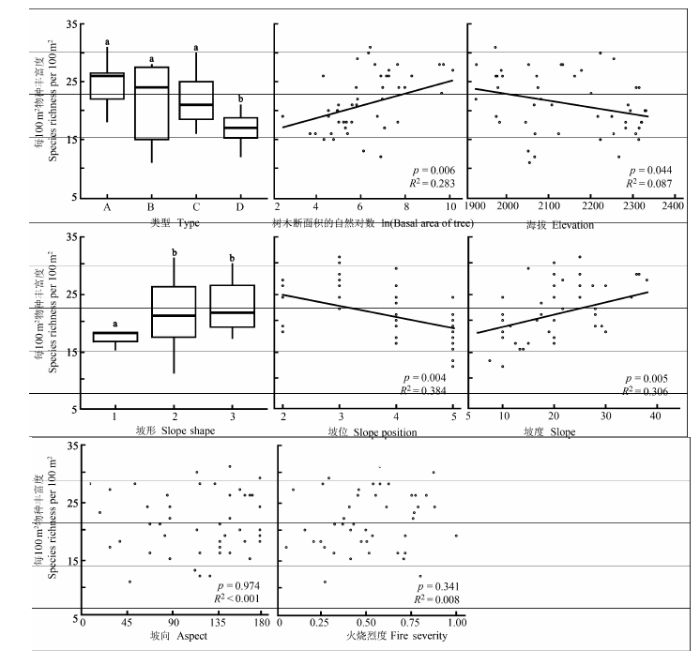
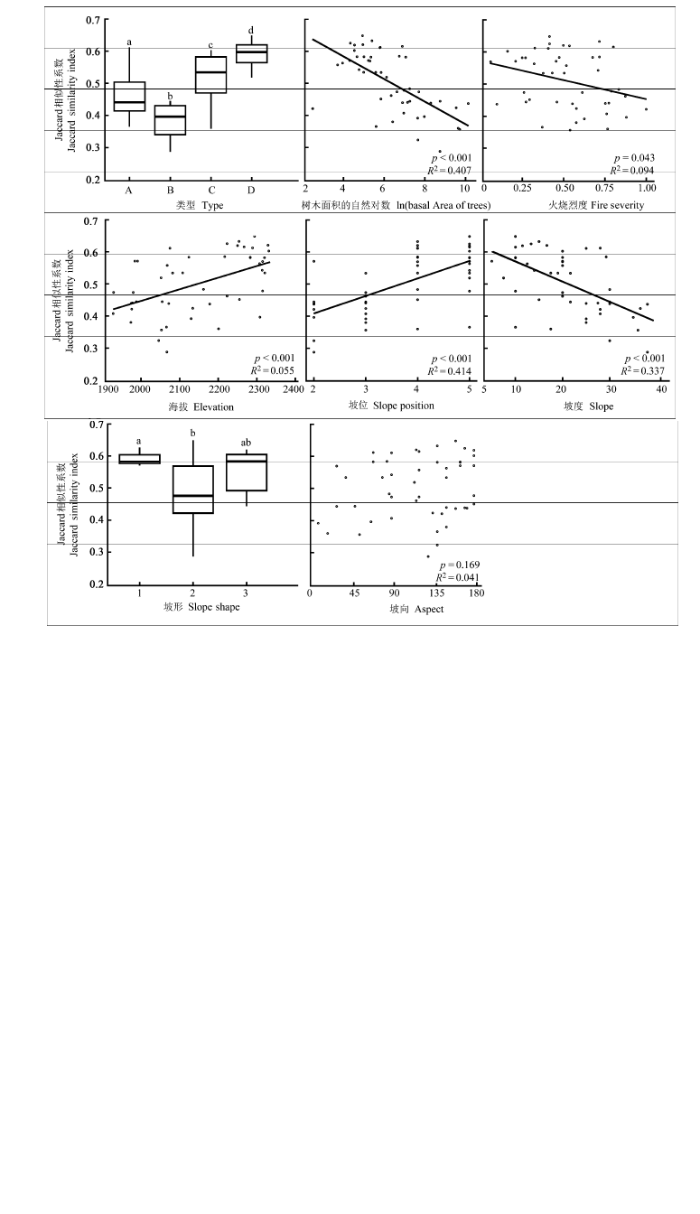
Fig. 4 The relationships between habitat factors and the species richness in herb layer of the transects of post-fire plant communities. The letters a and b above all box-plots refer to significantly different levels in t-test. A, B, C, and D designate four types of post-fire regenerated herb communities. A, Pteridium aquilinum var. latiusculum + Ageratina adenophora + Gentiana cephalantha community; B, Arundinella hookeri + Digitaria violascens + Bothriochloa ischaemum community; C, Kummerowia striata + Bulbostylis densa + Bothriochloa ischaemum community; D, Arthraxon hispidus + Kummerowia striata + Bothriochloa ischaemum community.
Fig. 5 The patterns of β-diversity in herb layer of post-fire plant communities. The letters a and b above all box-plots refer to significantly different levels in t-test. A, B, C, and D in the first graph panel designate four types of post-fire regenerated herb communities. A, Pteridium aquilinum var. latiusculum + Ageratina adenophora + Gentiana cephalantha community; B, Arundinella hookeri + Digitaria violascens + Bothriochloa ischaemum community; C, Kummerowia striata + Bulbostylis densa + Bothriochloa ischaemum community; D, Arthraxon hispidus + Kummerowia striata + Bothriochloa ischaemum community.
Table 2
表2
表2 生境因子对火后植物群落草本层物种多样性的影响
Table 2 Effects of habitat factors on the species diversity in herb layer of post-fire plant communities
Pattern and Process in a Forested Ecosystem: Disturbance, Development and the Steady State Based on the Hubbard Brook Ecosystem Study. Springer Science & Business Media
Topography and forest composition affecting the variability in fire severity and post-fire regeneration occurring after a large fire in the Mediterranean basin.
International Journal of Wildland Fire, 13, 209-216.
The relationship between the severely affected forest fire season in Yunnan and the abnormal change of the Pacific Ocean surface temperature. Journal of Yunnan University (
Natural Sciences), 33(S1), 188-191. (in Chinese with English abstract)[戴丛蕊, 顾本文, 李金槐 (2011).
Early post-fire regeneration of a fire-prone subtropical Pinus mixed forest in Yunnan, southwest China: The effects of pre-fire vegetation, fire severity and topographic factors.
HillMO (1979). TWINSPAN—A FORTRAN Program for Arranging Multivariate Data in an Ordered Two-Way Table by Classification of the Individuals and Attributes. Cornell University Press, New York.
[19]
HurttGC, PacalaSW (1995).
The consequences of recruitment limitation: reconciling chance, history and competitive differences between plants.
Effects of heterogeneity of pre-fire forests and vegetation burn severity on short-term post-fire vegetation density and regeneration in Samcheok, Korea.
Landscape and Ecological Engineering, 10, 215-228.
Post-fire plant diversity and abundance in pine and eucalypt stands in Portugal: Effects of biogeography, topography, forest type and post-fire management.
Species richness patterns in the understory of Pyrenean Pinus sylvestris forest.
Journal of Vegetation Science, 5, 517-524.
[33]
RoderA, HillJ, DuguyB, AllozaJ, VallejoR (2008).
Using long time series of Landsat data to monitor fire events and post-fire dynamics and identify driving factors. A case study in the Ayora region (eastern Spain).
Pattern and Process in a Forested Ecosystem: Disturbance, Development and the Steady State Based on the Hubbard Brook Ecosystem Study. Springer Science & Business Media
Salvage logging affects early post-fire tree composition in Canadian boreal forest.
1
2014
... 本研究中火后更新的草本植物α多样性和β多样性受火烧强度的影响并不十分显著, 与其他研究结果相近(Keeley et al., 2005).火烧强度加剧导致更新草本α多样性降低, 但只反映在样方而非样带尺度上, 对β多样性的影响则仅反映在不考虑物种多度的Jaccard指数值中.但是, 火烧强度与其他生境因子之间存在复杂的相互作用, 如: 1)火烧的烈度本身有可能受到地形、环境的影响, 在不同的地形上火烧强度差异会很大(Keeley et al., 2005), 从而造成不同地形部位更新草本的多样性不同; 2)火烧强度的不同会导致木本层的毁坏程度不同, 改变上层林木冠层结构, 类似砍伐的效果, 从而使地表接受到的光照效果不同(Boucher et al., 2014); 3)不同的火烧强度导致土壤的理化性质发生了不同的变化, 造成了草本微生境的不同, 增加了其β多样性.因此, 火烧强度与生境因子之间存在某种协同的变化. ...
Fire in the Earth system.
1
2009
... 火是陆地生态系统的主要干扰类型, 它不仅改变局部生境的植被结构与动态, 而且能改变景观格局与生态过程(Bond & Keeley, 2005; Kurkowski et al., 2008; Bowman et al., 2009).在过去几十年里, 由于气候变化和人类活动加剧, 全球范围内森林火灾发生的频率及火烧面积都有了显著增加(Roder et al., 2008 ; Bowman et al., 2011; Oliveira et al., 2014).植被对火干扰的响应及火后植被更新动态有助于我们理解火的作用, 及预测火后森林生态系统的结构功能变化(Mayer & Swetnam, 2000; Casady et al., 2010), 因此, 它是火生态研究的核心问题之一(Ackerly, 2004; Forrestel et al., 2014). ...
The human dimension of fire regimes on Earth.
1
2011
... 火是陆地生态系统的主要干扰类型, 它不仅改变局部生境的植被结构与动态, 而且能改变景观格局与生态过程(Bond & Keeley, 2005; Kurkowski et al., 2008; Bowman et al., 2009).在过去几十年里, 由于气候变化和人类活动加剧, 全球范围内森林火灾发生的频率及火烧面积都有了显著增加(Roder et al., 2008 ; Bowman et al., 2011; Oliveira et al., 2014).植被对火干扰的响应及火后植被更新动态有助于我们理解火的作用, 及预测火后森林生态系统的结构功能变化(Mayer & Swetnam, 2000; Casady et al., 2010), 因此, 它是火生态研究的核心问题之一(Ackerly, 2004; Forrestel et al., 2014). ...
Topography and forest composition affecting the variability in fire severity and post-fire regeneration occurring after a large fire in the Mediterranean basin.
1
2004
... 已有研究表明, 火后更新群落的物种多样性主要受到4个因素的影响, 即地形和土壤环境(Lobo, 2001; Abella & Covington, 2006)、火烧强度(Vallejo & Alloza, 2012; Crotteau et al., 2013)、火前植被(Broncano & Retana, 2004; Lee et al., 2014)和物种生活史特征(Hurtt & Pacala, 1995).但是, 这4个因素之间并非完全独立.如阳坡和阴坡所接受的太阳辐射和水分蒸散量不同会导致不同的土壤环境, 因而火前植被群落也不同(Mouillot et al., 2005; Fox et al., 2008).目前, 关于各个因素对火后植被更新的影响大小尚未达成共识.如Pausas (1994)发现, 随着海拔增加, 火后更新层中木本植物的多样性在减少; 但Lobo (2001)在伊比利亚半岛的研究结果却相反.此外, 大多数的研究表明火烧强度决定了最初的种子库, 从而影响了火后更新植被的多样性(Maia et al., 2012; Crotteau et al., 2013; Lee et al., 2014).然而, 最近有研究指出火烧强度对火后更新物种多样性没有显著影响(Camac et al., 2013).显然, 关于火后更新物种多样性的格局及其影响机制有待更多的研究积累和深入探讨. ...
Post-fire regeneration in alpine heathland: Does fire severity matter?
1
2013
... 已有研究表明, 火后更新群落的物种多样性主要受到4个因素的影响, 即地形和土壤环境(Lobo, 2001; Abella & Covington, 2006)、火烧强度(Vallejo & Alloza, 2012; Crotteau et al., 2013)、火前植被(Broncano & Retana, 2004; Lee et al., 2014)和物种生活史特征(Hurtt & Pacala, 1995).但是, 这4个因素之间并非完全独立.如阳坡和阴坡所接受的太阳辐射和水分蒸散量不同会导致不同的土壤环境, 因而火前植被群落也不同(Mouillot et al., 2005; Fox et al., 2008).目前, 关于各个因素对火后植被更新的影响大小尚未达成共识.如Pausas (1994)发现, 随着海拔增加, 火后更新层中木本植物的多样性在减少; 但Lobo (2001)在伊比利亚半岛的研究结果却相反.此外, 大多数的研究表明火烧强度决定了最初的种子库, 从而影响了火后更新植被的多样性(Maia et al., 2012; Crotteau et al., 2013; Lee et al., 2014).然而, 最近有研究指出火烧强度对火后更新物种多样性没有显著影响(Camac et al., 2013).显然, 关于火后更新物种多样性的格局及其影响机制有待更多的研究积累和深入探讨. ...
Evaluating post-wildfire vegetation regeneration as a response to multiple environmental determinants.
1
2010
... 火是陆地生态系统的主要干扰类型, 它不仅改变局部生境的植被结构与动态, 而且能改变景观格局与生态过程(Bond & Keeley, 2005; Kurkowski et al., 2008; Bowman et al., 2009).在过去几十年里, 由于气候变化和人类活动加剧, 全球范围内森林火灾发生的频率及火烧面积都有了显著增加(Roder et al., 2008 ; Bowman et al., 2011; Oliveira et al., 2014).植被对火干扰的响应及火后植被更新动态有助于我们理解火的作用, 及预测火后森林生态系统的结构功能变化(Mayer & Swetnam, 2000; Casady et al., 2010), 因此, 它是火生态研究的核心问题之一(Ackerly, 2004; Forrestel et al., 2014). ...
Post-fire regeneration across a fire severity gradient in the southern Cascades.
1
2013
... 已有研究表明, 火后更新群落的物种多样性主要受到4个因素的影响, 即地形和土壤环境(Lobo, 2001; Abella & Covington, 2006)、火烧强度(Vallejo & Alloza, 2012; Crotteau et al., 2013)、火前植被(Broncano & Retana, 2004; Lee et al., 2014)和物种生活史特征(Hurtt & Pacala, 1995).但是, 这4个因素之间并非完全独立.如阳坡和阴坡所接受的太阳辐射和水分蒸散量不同会导致不同的土壤环境, 因而火前植被群落也不同(Mouillot et al., 2005; Fox et al., 2008).目前, 关于各个因素对火后植被更新的影响大小尚未达成共识.如Pausas (1994)发现, 随着海拔增加, 火后更新层中木本植物的多样性在减少; 但Lobo (2001)在伊比利亚半岛的研究结果却相反.此外, 大多数的研究表明火烧强度决定了最初的种子库, 从而影响了火后更新植被的多样性(Maia et al., 2012; Crotteau et al., 2013; Lee et al., 2014).然而, 最近有研究指出火烧强度对火后更新物种多样性没有显著影响(Camac et al., 2013).显然, 关于火后更新物种多样性的格局及其影响机制有待更多的研究积累和深入探讨. ...
Early post-fire regeneration of a fire-prone subtropical Pinus mixed forest in Yunnan, southwest China: The effects of pre-fire vegetation, fire severity and topographic factors.
1
2015
... 火后更新的草本物种多样性格局与地形的关系是密切的(Lobo, 2001; Abella & Covington, 2006).在本研究中, 草本层物种丰富度与物种构成的变化均受到地形因子的显著影响.其中坡位和坡度在样方和样带尺度上均为效应最突出的因子.大致上较高坡位的草本种类较少, 而坡度较陡地段的物种丰富度却较高, 主要因为较陡地形一般在中下坡位, 水分条件较好, 草本层恢复较快, 覆盖度较高.一般来说, 海拔与水热条件联系紧密, 但在本研究中, 一方面海拔范围并不太大, 而海拔变化与坡位变化有一致性, 主要体现了土壤水分条件的变化, 这在之前的研究中也得到了验证(Han et al., 2015).中高纬度地区火后植被恢复往往受坡向的影响较显著, 这主要与所接受的太阳辐射有关(Mouillot et al., 2005; Fox et al., 2008).本研究中坡向对恢复草本层α多样性的影响并不显著(图2; 表2), 这可能是因为研究区域所处纬度较低(25.13° N), 特别是夏、秋季节南北坡的太阳辐射高度角相差不大. ...
The consequences of recruitment limitation: reconciling chance, history and competitive differences between plants.
1
1995
... 已有研究表明, 火后更新群落的物种多样性主要受到4个因素的影响, 即地形和土壤环境(Lobo, 2001; Abella & Covington, 2006)、火烧强度(Vallejo & Alloza, 2012; Crotteau et al., 2013)、火前植被(Broncano & Retana, 2004; Lee et al., 2014)和物种生活史特征(Hurtt & Pacala, 1995).但是, 这4个因素之间并非完全独立.如阳坡和阴坡所接受的太阳辐射和水分蒸散量不同会导致不同的土壤环境, 因而火前植被群落也不同(Mouillot et al., 2005; Fox et al., 2008).目前, 关于各个因素对火后植被更新的影响大小尚未达成共识.如Pausas (1994)发现, 随着海拔增加, 火后更新层中木本植物的多样性在减少; 但Lobo (2001)在伊比利亚半岛的研究结果却相反.此外, 大多数的研究表明火烧强度决定了最初的种子库, 从而影响了火后更新植被的多样性(Maia et al., 2012; Crotteau et al., 2013; Lee et al., 2014).然而, 最近有研究指出火烧强度对火后更新物种多样性没有显著影响(Camac et al., 2013).显然, 关于火后更新物种多样性的格局及其影响机制有待更多的研究积累和深入探讨. ...
The distribution of the flora in the alpine zone.
1
1912
... Jaccard相似性系数(J) (Jaccard, 1912): ...
Factors affecting plant diversity during post-fire recovery and succession of Mediterranean-climate shrublands in California, USA.
5
2005
... 本研究表明, 对于云南松混交林而言, 在森林火灾后的更新群落, 火前冠层植被、火烧强度以及生境地形条件均对草本层物种多样性及其构成产生显著影响, 与其他研究火后更新的多样性结果(Guo, 2001; Keeley et al., 2005; Maia et al., 2014)相比, 具体影响更新结果的因子不尽相同. ...
Relative importance of different secondary successional pathways in an Alaskan boreal forest.
1
2008
... 火是陆地生态系统的主要干扰类型, 它不仅改变局部生境的植被结构与动态, 而且能改变景观格局与生态过程(Bond & Keeley, 2005; Kurkowski et al., 2008; Bowman et al., 2009).在过去几十年里, 由于气候变化和人类活动加剧, 全球范围内森林火灾发生的频率及火烧面积都有了显著增加(Roder et al., 2008 ; Bowman et al., 2011; Oliveira et al., 2014).植被对火干扰的响应及火后植被更新动态有助于我们理解火的作用, 及预测火后森林生态系统的结构功能变化(Mayer & Swetnam, 2000; Casady et al., 2010), 因此, 它是火生态研究的核心问题之一(Ackerly, 2004; Forrestel et al., 2014). ...
Effects of heterogeneity of pre-fire forests and vegetation burn severity on short-term post-fire vegetation density and regeneration in Samcheok, Korea.
2
2014
... 已有研究表明, 火后更新群落的物种多样性主要受到4个因素的影响, 即地形和土壤环境(Lobo, 2001; Abella & Covington, 2006)、火烧强度(Vallejo & Alloza, 2012; Crotteau et al., 2013)、火前植被(Broncano & Retana, 2004; Lee et al., 2014)和物种生活史特征(Hurtt & Pacala, 1995).但是, 这4个因素之间并非完全独立.如阳坡和阴坡所接受的太阳辐射和水分蒸散量不同会导致不同的土壤环境, 因而火前植被群落也不同(Mouillot et al., 2005; Fox et al., 2008).目前, 关于各个因素对火后植被更新的影响大小尚未达成共识.如Pausas (1994)发现, 随着海拔增加, 火后更新层中木本植物的多样性在减少; 但Lobo (2001)在伊比利亚半岛的研究结果却相反.此外, 大多数的研究表明火烧强度决定了最初的种子库, 从而影响了火后更新植被的多样性(Maia et al., 2012; Crotteau et al., 2013; Lee et al., 2014).然而, 最近有研究指出火烧强度对火后更新物种多样性没有显著影响(Camac et al., 2013).显然, 关于火后更新物种多样性的格局及其影响机制有待更多的研究积累和深入探讨. ...
... ; Crotteau et al., 2013; Lee et al., 2014).然而, 最近有研究指出火烧强度对火后更新物种多样性没有显著影响(Camac et al., 2013).显然, 关于火后更新物种多样性的格局及其影响机制有待更多的研究积累和深入探讨. ...
Spatial and environmental determinants of vascular plant species richness distribution in the Iberian Peninsula and Balearic Islands.
2
2001
... 已有研究表明, 火后更新群落的物种多样性主要受到4个因素的影响, 即地形和土壤环境(Lobo, 2001; Abella & Covington, 2006)、火烧强度(Vallejo & Alloza, 2012; Crotteau et al., 2013)、火前植被(Broncano & Retana, 2004; Lee et al., 2014)和物种生活史特征(Hurtt & Pacala, 1995).但是, 这4个因素之间并非完全独立.如阳坡和阴坡所接受的太阳辐射和水分蒸散量不同会导致不同的土壤环境, 因而火前植被群落也不同(Mouillot et al., 2005; Fox et al., 2008).目前, 关于各个因素对火后植被更新的影响大小尚未达成共识.如Pausas (1994)发现, 随着海拔增加, 火后更新层中木本植物的多样性在减少; 但Lobo (2001)在伊比利亚半岛的研究结果却相反.此外, 大多数的研究表明火烧强度决定了最初的种子库, 从而影响了火后更新植被的多样性(Maia et al., 2012; Crotteau et al., 2013; Lee et al., 2014).然而, 最近有研究指出火烧强度对火后更新物种多样性没有显著影响(Camac et al., 2013).显然, 关于火后更新物种多样性的格局及其影响机制有待更多的研究积累和深入探讨. ...
... 火后更新的草本物种多样性格局与地形的关系是密切的(Lobo, 2001; Abella & Covington, 2006).在本研究中, 草本层物种丰富度与物种构成的变化均受到地形因子的显著影响.其中坡位和坡度在样方和样带尺度上均为效应最突出的因子.大致上较高坡位的草本种类较少, 而坡度较陡地段的物种丰富度却较高, 主要因为较陡地形一般在中下坡位, 水分条件较好, 草本层恢复较快, 覆盖度较高.一般来说, 海拔与水热条件联系紧密, 但在本研究中, 一方面海拔范围并不太大, 而海拔变化与坡位变化有一致性, 主要体现了土壤水分条件的变化, 这在之前的研究中也得到了验证(Han et al., 2015).中高纬度地区火后植被恢复往往受坡向的影响较显著, 这主要与所接受的太阳辐射有关(Mouillot et al., 2005; Fox et al., 2008).本研究中坡向对恢复草本层α多样性的影响并不显著(图2; 表2), 这可能是因为研究区域所处纬度较低(25.13° N), 特别是夏、秋季节南北坡的太阳辐射高度角相差不大. ...
Post-fire plant diversity and abundance in pine and eucalypt stands in Portugal: Effects of biogeography, topography, forest type and post-fire management.
1
2014
... 本研究表明, 对于云南松混交林而言, 在森林火灾后的更新群落, 火前冠层植被、火烧强度以及生境地形条件均对草本层物种多样性及其构成产生显著影响, 与其他研究火后更新的多样性结果(Guo, 2001; Keeley et al., 2005; Maia et al., 2014)相比, 具体影响更新结果的因子不尽相同. ...
Fire severity as a key factor in post-fire regeneration of Pinus pinaster (Ait.) in Central Portugal.
1
2012
... 已有研究表明, 火后更新群落的物种多样性主要受到4个因素的影响, 即地形和土壤环境(Lobo, 2001; Abella & Covington, 2006)、火烧强度(Vallejo & Alloza, 2012; Crotteau et al., 2013)、火前植被(Broncano & Retana, 2004; Lee et al., 2014)和物种生活史特征(Hurtt & Pacala, 1995).但是, 这4个因素之间并非完全独立.如阳坡和阴坡所接受的太阳辐射和水分蒸散量不同会导致不同的土壤环境, 因而火前植被群落也不同(Mouillot et al., 2005; Fox et al., 2008).目前, 关于各个因素对火后植被更新的影响大小尚未达成共识.如Pausas (1994)发现, 随着海拔增加, 火后更新层中木本植物的多样性在减少; 但Lobo (2001)在伊比利亚半岛的研究结果却相反.此外, 大多数的研究表明火烧强度决定了最初的种子库, 从而影响了火后更新植被的多样性(Maia et al., 2012; Crotteau et al., 2013; Lee et al., 2014).然而, 最近有研究指出火烧强度对火后更新物种多样性没有显著影响(Camac et al., 2013).显然, 关于火后更新物种多样性的格局及其影响机制有待更多的研究积累和深入探讨. ...
Century scale climate forcing of fire regimes in the American Southwest.
1
2000
... 火是陆地生态系统的主要干扰类型, 它不仅改变局部生境的植被结构与动态, 而且能改变景观格局与生态过程(Bond & Keeley, 2005; Kurkowski et al., 2008; Bowman et al., 2009).在过去几十年里, 由于气候变化和人类活动加剧, 全球范围内森林火灾发生的频率及火烧面积都有了显著增加(Roder et al., 2008 ; Bowman et al., 2011; Oliveira et al., 2014).植被对火干扰的响应及火后植被更新动态有助于我们理解火的作用, 及预测火后森林生态系统的结构功能变化(Mayer & Swetnam, 2000; Casady et al., 2010), 因此, 它是火生态研究的核心问题之一(Ackerly, 2004; Forrestel et al., 2014). ...
Long-term forest dynamic after land abandonment in a fire prone Mediterranean landscape (central Corsica, France).
3
2005
... 已有研究表明, 火后更新群落的物种多样性主要受到4个因素的影响, 即地形和土壤环境(Lobo, 2001; Abella & Covington, 2006)、火烧强度(Vallejo & Alloza, 2012; Crotteau et al., 2013)、火前植被(Broncano & Retana, 2004; Lee et al., 2014)和物种生活史特征(Hurtt & Pacala, 1995).但是, 这4个因素之间并非完全独立.如阳坡和阴坡所接受的太阳辐射和水分蒸散量不同会导致不同的土壤环境, 因而火前植被群落也不同(Mouillot et al., 2005; Fox et al., 2008).目前, 关于各个因素对火后植被更新的影响大小尚未达成共识.如Pausas (1994)发现, 随着海拔增加, 火后更新层中木本植物的多样性在减少; 但Lobo (2001)在伊比利亚半岛的研究结果却相反.此外, 大多数的研究表明火烧强度决定了最初的种子库, 从而影响了火后更新植被的多样性(Maia et al., 2012; Crotteau et al., 2013; Lee et al., 2014).然而, 最近有研究指出火烧强度对火后更新物种多样性没有显著影响(Camac et al., 2013).显然, 关于火后更新物种多样性的格局及其影响机制有待更多的研究积累和深入探讨. ...
... 火后更新的草本物种多样性格局与地形的关系是密切的(Lobo, 2001; Abella & Covington, 2006).在本研究中, 草本层物种丰富度与物种构成的变化均受到地形因子的显著影响.其中坡位和坡度在样方和样带尺度上均为效应最突出的因子.大致上较高坡位的草本种类较少, 而坡度较陡地段的物种丰富度却较高, 主要因为较陡地形一般在中下坡位, 水分条件较好, 草本层恢复较快, 覆盖度较高.一般来说, 海拔与水热条件联系紧密, 但在本研究中, 一方面海拔范围并不太大, 而海拔变化与坡位变化有一致性, 主要体现了土壤水分条件的变化, 这在之前的研究中也得到了验证(Han et al., 2015).中高纬度地区火后植被恢复往往受坡向的影响较显著, 这主要与所接受的太阳辐射有关(Mouillot et al., 2005; Fox et al., 2008).本研究中坡向对恢复草本层α多样性的影响并不显著(图2; 表2), 这可能是因为研究区域所处纬度较低(25.13° N), 特别是夏、秋季节南北坡的太阳辐射高度角相差不大. ...
... 对于β多样性, 地形影响主要反映在海拔、坡位和坡度方面.随着海拔和坡位上升、坡度变平缓, 生境因受土壤水分限制而逐渐变得单一, 草本的构成更为简单、相似(Mouillot et al., 2005; Fox et al., 2008), β多样性减小. ...
Exploring the spatial patterns of fire density in Southern Europe using Geographically Weighted Regression.
1
2014
... 火是陆地生态系统的主要干扰类型, 它不仅改变局部生境的植被结构与动态, 而且能改变景观格局与生态过程(Bond & Keeley, 2005; Kurkowski et al., 2008; Bowman et al., 2009).在过去几十年里, 由于气候变化和人类活动加剧, 全球范围内森林火灾发生的频率及火烧面积都有了显著增加(Roder et al., 2008 ; Bowman et al., 2011; Oliveira et al., 2014).植被对火干扰的响应及火后植被更新动态有助于我们理解火的作用, 及预测火后森林生态系统的结构功能变化(Mayer & Swetnam, 2000; Casady et al., 2010), 因此, 它是火生态研究的核心问题之一(Ackerly, 2004; Forrestel et al., 2014). ...
Species richness patterns in the understory of Pyrenean Pinus sylvestris forest.
1994
Using long time series of Landsat data to monitor fire events and post-fire dynamics and identify driving factors. A case study in the Ayora region (eastern Spain).
1
2008
... 火是陆地生态系统的主要干扰类型, 它不仅改变局部生境的植被结构与动态, 而且能改变景观格局与生态过程(Bond & Keeley, 2005; Kurkowski et al., 2008; Bowman et al., 2009).在过去几十年里, 由于气候变化和人类活动加剧, 全球范围内森林火灾发生的频率及火烧面积都有了显著增加(Roder et al., 2008 ; Bowman et al., 2011; Oliveira et al., 2014).植被对火干扰的响应及火后植被更新动态有助于我们理解火的作用, 及预测火后森林生态系统的结构功能变化(Mayer & Swetnam, 2000; Casady et al., 2010), 因此, 它是火生态研究的核心问题之一(Ackerly, 2004; Forrestel et al., 2014). ...
Modified TWINSPAN classification in which the hierarchy respects cluster heterogeneity.
1
2009
... 为探究在不同条件下火后更新的草本群落差异, 使用双向指示种分析(TWINSPAN, Hill, 1979)对47条样带的草本层物种构成进行植物群落分类.TWINSPAN仍是目前最常用的植物群落分类方法(沈泽昊等, 2000b).分析在软件PC-ORD (Roleček et al., 2009)中完成. ...
Geographical patterns of community-based tree species richness in Chinese mountain forests: The effects of contemporary climate and regional history.
1
2012
... 为探究不同环境因子在影响火后更新草本层多样性上的不同表现, 使用广义线性回归模型(GLM)与变量解释的等级方差分离法.采用广义线性回归模型来拟合多个自变量和因变量的关系, 用赤池信息准则评价和筛选最优模型.为了控制模型解释变量之间的共线性, 采用等级方差分离法来定量评价诸因子对模型的贡献(Shen et al., 2012).等级方差分离法将每个变量的独立和协同贡献表达为占解释变化总量的百分比.变量独立贡献的统计显著性用模型的随机化重复来估计, 并基于Z检验的0.95置信上限来评价(Walsh & Mac Nally, 2007).利用统计软件R中的等级方差分离程序包(hier.part)来完成这一步分析. ...
a). 三峡大老岭森林物种多样性的空间格局分析及其地形解释
2000
b). 三峡大老岭地区森林群落的数量分类研究
2000
Thr role of fire in the Central Yunnan Plateau ecosystem, southwestern China.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}