在不同环境下, 植物种群的发育特性和物质分配策略是植物局部特化的形态特征对资源异质性的响应(Kleyer & Minden, 2015 )。在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ; 史元春等, 2015 ), 在植物的整个生长过程中, 功能性状是植物与环境相互作用的结果, 如何调节小枝内各构件的生长及关系, 是植物生存策略和生境适合度调整的重要内容(肖遥等, 2014 )。在植物各器官中, 枝条和叶片是重要的输导和物质生产器官, 在生物力学及功能上具有密切联系。枝条具有导水功能和机械支撑作用(Alméras et al., 2004 ), 其大小往往关系到植物的空间拓展策略, 决定叶片的物质投资方式; 叶片大小和多少直接影响着植株的发育模式, 以及植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ), 枝、叶关系不仅是植物个体发育过程的表现(Brouat et al., 1998 ), 也体现了植物对特定生境的适应性(Violle et al., 2007 )。植物的形态特征和枝、叶功能之间的平衡受年龄和个体大小制约(Heuret et al., 2006 ; Barthélémy & Caraglio, 2007 ), 不同大小株丛在群落中占据的资源生态位存在差异(姚婧等, 2013 ), 其受群落郁闭度和干扰程度不同, 不同层次枝叶接受的有效太阳辐射也不同, 从而影响植物的空间分布格局及光照、温度等环境条件(程栋梁等, 2011 ), 往往导致不同个体大小植株分枝构型出现明显的分化。为了提高植物构型构件的生境适应性和自身的生存适合度, 不同大小等级植株可能需要适时调整其分枝角度, 增加植株所接受的光照强度, 促使植株枝-叶性状生长及其相互关系进行相应的调整(林勇明等, 2007 ), 使种群内每个植株可获得性资源的数量最大化, 构建与生境协同进化的冠层构型(de Kroons & Hutchings, 1995 ; 武高林等, 2010 ), 体现了植物适应特定环境的构型 塑造。
蒺藜科霸王属霸王(Zygophyllum xanthoxylum)是分布于我国西北部干旱荒漠区的强旱生小灌木, 当年生枝条数量多、叶量大、生长快, 具有抗旱、速生、高产、耐沙埋、耐贫瘠、抗风蚀的能力, 具有较好的饲用价值和适口性, 是荒漠灌丛植被的主要优势种和建群种(Chen et al., 2015 )。目前, 许多学者已对植物个体大小构型差异与竞争环境的关系(Aarssen, 2015 )、不同发育阶段分枝格局与叶片生长变化规律(许强等, 2013 )、不同个体大小茎-叶资源分配策略(李钰等, 2013b )等方面进行了大量研究, 解释了植物适应特定环境的生长特性与物质分配规律。霸王在不同环境下的表型特征差异也得到了关注(南江等, 2014 ), 有关霸王枝、叶性状关系的个体大小差异及其对不同光照强度生长特性的研究还不够深入, 影响了植物枝叶功能性状对不同生境适应机制的认识。因此, 本研究从植株个体大小的角度研究了荒漠草地霸王的枝叶构型特征及其光环境响应策略, 试图揭示: (1)不同大小等级霸王小枝内枝大小、叶面积和叶数量资源配置模式; (2)这种资源配置模式对霸王枝长度与叶大小、枝长度和叶数量间异速生长关系的影响。
1 研究地区和研究方法
1.1 研究区和样地概况
研究区位于祁连山北坡中山区的甘肃省肃南县皂矾沟地区, 地理位置为38.91°-38.98° N, 99.81°-99.87° E, 海拔1834-2118 m, 属于山地荒漠草原, 年平均气温6.5 ℃, 7月平均气温19 ℃, 1月平均气温-7.5 ℃, ≥0 ℃积温为2933-3300 ℃, 年降水量173.1 mm, 降水主要集中在6-8月, 年蒸发量2013.7 mm, 是年降水量的11.6倍。土壤为山地灰漠土, 由于气候干燥和长期剥蚀的作用, 形成砾石倾斜和裸岩广布的低山地貌景观。植物以旱生小半灌木为主, 红砂属(Reaumuria)植物占相当比例。主要植物有霸王、红砂(Reaumuria songarica)、合头草(Sympegma regelii)、珍珠猪毛菜(Salsola passer- ina)、盐爪爪(Kalidium foliatum)、驼绒藜(Ceratoides latens), 伴生植物有灌木亚菊(Ajania fruticulosa)、中亚紫菀木(Asterothamnus centrali-asiaticus)、碱韭(Allium polyrhizum)、黄花瓦松(Orostachys spinosus)、黄花补血草(Limonium aureum)、三芒草(Aristida adscensionis)等。
1.2 野外取样和测量
1.2.1 草地群落学调查
于2014年8月10-20日, 在研究区选择地貌相对一致、地势平坦的草地, 沿东西方向设置250 m × 60 m的水平样带1条, 在样带内间隔50 m布置3块50 m × 50 m的样地。采用双对角线法进行草地群落学调查, 在样带内设置6个5 m × 5 m的小样方测定草地群落所有植物的高度和密度。
1.2.2 霸王枝叶性状测定
对每块样地内的所有霸王进行编号, 按编号顺序测量并记录每株霸王的东西和南北冠幅、高度。在每株霸王东、西、南、北4个方位各选择1个枝条(枝条数量≤4则全部选择), 对于选择的每个大枝, 定义枝条末端(最外层的当年生枝)为第1层级枝条, 向内第2层级为支撑枝, 并标记1个无明显损失的小枝, 在现场, 用量角器测量当年生小枝(标记的)与其支撑枝之间的夹角(最小角度), 然后将小枝(标记的)剪取, 放入塑料袋, 编码后带回实验室进一步处理。在实验室6 h内完成以下测量: 对于每个当年生枝条, 首先清点记录叶片数量并摘除, 再测量当年生小枝上每个叶片的叶面积(通过扫描每一个小枝上的所有叶片, 用MapInfo软件进行计算); 然后分别用直尺和游标卡尺测量小枝枝长度和最粗直径; 最后, 将小枝、叶片一起置于60 ℃烘箱中烘干后称量干质量, 以备其他分析。
1.2.3 霸王光合有效辐射测定
选择晴朗无云的天气用手持光量子计(3415F, Walz, Plainfield, USA)于9:00-11:00对3块样地内所有霸王进行光合有效辐射(photosynthetically active radiation, PAR)测定, 测定位置分别在植株上方(距冠层15 cm)、中层(株高1/2处)和地表(距地表15 cm处)。
1.3 植株大小等级划分
植株的体积(植株个体地面高度×长冠幅×短冠幅)是反映其形体大小的重要指标(杨洪晓等, 2006 )。通过Excel 2007软件将植株个数设为纵坐标, 植株体积立方根设为横坐标, 生成一条正态曲线, 依据曲线走向将霸王划分为3个大小等级: I级(d ≤ 60 cm); II级(60 cm < d ≤ 120 cm); III级(d > 120 cm), 其中d为植株体积的立方根。经统计分析, 采集的117株霸王中有I级39株、II级44株、III级34株。
1.4 数据分析
以每个大小等级30株霸王枝、叶性状及各层次光合有效辐射的平均值代表该个体大小霸王的枝、叶性状值和各层次光合有效辐射值, 然后对枝、叶性状的平均值进行对数(以10为底)转换, 使之符合正态分布后再进行分析。枝与叶片属性包括: 枝长度、叶面积、叶数量、枝横截面积。叶面积指当年生小枝上所支撑的所有叶片单叶面积的平均值。对枝叶功能性状关系的研究, 采用y = bxa , 线性转换成log(y) = log(b) + alog(x), 式中, x和y表示两个特征参数, b代表性状关系的截距, a表示相关性的斜率, 即异速生长参数或相对生长的指数, 当a = 1时, 表示两者是等速关系; 当a > 1时, 表示y的增加程度大于x的增加程度; 当a < 1时, 表示y的增加程度小于x的增加程度。数据分析主要采用标准化主轴估计(SMA)的方法(Warton et al., 2006 ), 由软件(S)MATR Version 2.0 (Falster et al., 2006 )计算完成。每一个回归斜率的置信区间根据Pitman (1939)方法计算, 通过Warton和Weber (2002)的方法判断回归斜率的异质性(heterogeneity), 并且在斜率同质(homogeneity)时计算共同斜率, 截距和斜率的差异利用方差分析检验。不同个体大小间功能特征平均值的比较采用单因素方差分析(ANOVA), 这些计算在SPSS 16.0下进行。
2 结果
2.1 草地群落特征和霸王主要功能性状分析
根据草地群落学调查分析, 研究区样地群落高度为(94.80 ± 3.45) cm, 密度为(8.00 ± 1.20)株·m-2 。单因素方差分析表明不同大小等级霸王的高度、分枝角度和枝横截面积均存在极显著差异(p < 0.01, 表1 )。随着大小等级的增加, 霸王高度和枝横截面积呈逐渐增大趋势, II级霸王较I级霸王分别增加了72.83%和13.72%, III级霸王较II级霸王分别增加了20.27%和26.87%; 霸王分枝角度呈逐渐减小趋势, 其中II级霸王较I级霸王减小了7.82%, III级霸王较II级霸王减小了11.04%。
2.2 不同大小等级霸王光合有效辐射分析
由表2 的方差分析可看出, 各层次不同大小等级霸王光合有效辐射存在极显著差异(p < 0.01, 表2 )。由上层、中层至地表, 光合有效辐射依次呈递减趋势, I级霸王较II级霸王上层、中层、地表光合有效辐射分别降低了16.62%、19.55%、23.52%; I级霸王较III级霸王上层、中层、地表光合有效辐射分别降低了24.45%、26.78%、31.23%; II级霸王较III级霸王上层、中层、地表光合有效辐射分别降低了9.39%、8.99%、10.07%。可见, 个体等级越大, 植株上层光照捕获量越大; 个体等级越小, 植株愈易受到群落邻体干扰和上层遮阴, 这不利于光照下透, 影响中下层植物叶片光合作用。
2.3 不同大小等级霸王枝与叶性状
单因素方差分析表明, 不同大小等级霸王的枝长度、叶面积和叶数量均存在极显著差异(p < 0.01, 表3 )。随着大小等级的增加, 霸王枝长度与叶面积均呈逐渐增大趋势, II级霸王较I级霸王分别增加了5.18%和11.02%, III级霸王较II级霸王分别增加了6.43%和6.87%; 霸王叶数量呈逐渐减小趋势, II级霸王较I级霸王减小了5.84%, III级霸王较II级霸王减小了3.94%。
2.4 不同大小等级霸王枝长度与叶面积的关系
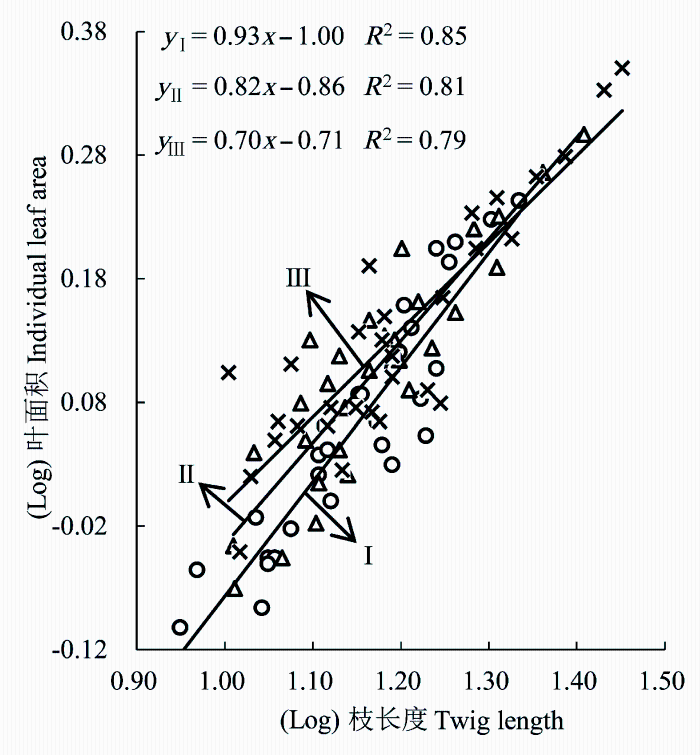
在不同大小等级上, 霸王枝长度与叶面积间均为显著正相关关系, 不同大小等级间的斜率存在显著差异(p < 0.05)。I级霸王斜率为0.93 (95%的置信区间CI = (0.85, 1.06), p = 0.62; 图1 ), 与1.0无显著差异(p > 0.05), 表明I级霸王枝长度和叶面积间存在等速权衡关系。II级和III级霸王斜率分别为0.82 (95%的置信区间CI = (0.74, 0.91), p = 0.73; 图1 )和0.70 (95%的置信区间CI = (0.63, 0.81), p = 0.59; 图1 ), 表明枝长度越大, 叶面积越大, 枝长度增加的速度大于叶面积增加的速度。随着大小等级的增加, 霸王叶面积与枝长度的斜率逐渐减小, 表明在某一给定的枝长度时, 较大等级上霸王具有更小的叶面积与枝长度的比值(图1 )。
图1 霸王枝长度与叶面积的关系。I-III, 个体大小等级。
Fig. 1 The relationship between twig length and leaf area of Zygophyllum xanthoxylum. I-III, plant size classes.
2.5 不同大小等级霸王枝长度与叶数量的关系
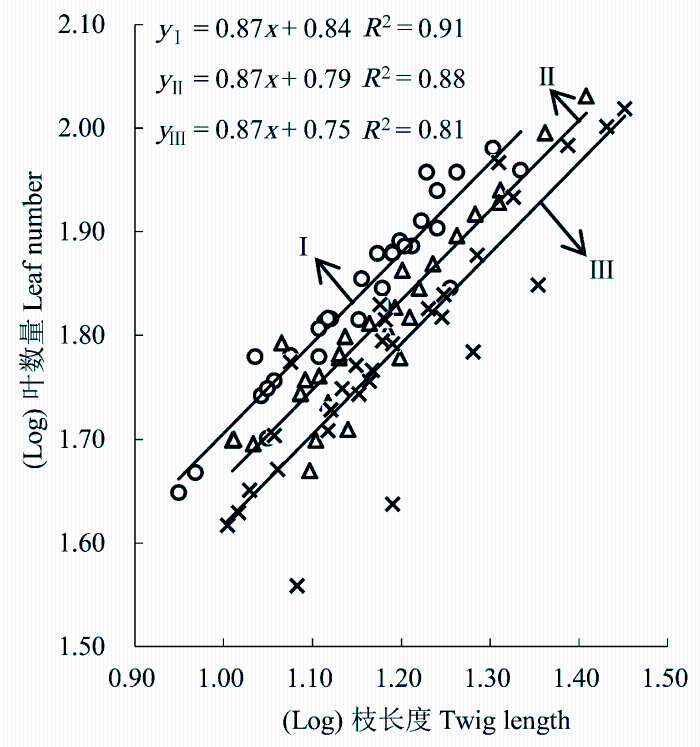
图2 显示, 枝长度与叶数量呈现显著的正相关关系(p < 0.05, 图2 )。通过SMA分析得出3个不同大小等级间的斜率无显著差异(p > 0.05), 枝长度与叶数量回归关系具有共同斜率, 为0.87 (95%的置信区间CI = (0.80, 0.96), 图2 ), 显著小于1 (p < 0.05), 表明枝长度增加比率大于叶数量的增加比率。随着大小等级的增加, 枝长度与叶数量的y轴截距显著下降(p < 0.05), 表明相同的枝长度投入下, 大小等级越大, 叶数量的投入相对越小。
图2 霸王枝长度与叶数量的关系。I-III, 个体大小等级。
Fig. 2 The relationship between twig length and leaf number of Zygophyllum xanthoxylum. I-III, plant size classes.
3 讨论
光是植物生长的能量来源, 不同的光照强度对植物光合作用的影响主要是通过植物对光照的捕获利用能力来实现的(王艺和韦小丽, 2010 )。Valladares等(2002)指出, 叶内与茎内支撑投资之间可能存在某种权衡关系, 进而导致不同的冠层设计, 使得捕光效率在总体上是相似的。叶面积的增大需要更长的枝条来减少个体内部对光照的相互遮挡, 叶片面积和数量的增加则要求枝条具有更高的支撑能力(Niinemets et al., 2007 ), 大个体植株较小个体植株能生长更粗大的枝条用于支持功能, 体现了植物枝-叶生长中的物质和能量转换关系(Enquist & Niklas, 2001 , 2002 )。本研究发现, 随着霸王大小等级的增加, 枝长度与叶面积关系的异速斜率逐渐降低, 即枝长度的生长速度逐渐高于叶面积的生长速度; 枝长度一定的条件下, 相对于小个体霸王, 大个体霸王植株叶数量的增加速率降低, 自然选择使它们以最佳的收益/代价关系以满足不同个体大小植株的生长需求。
3.1 不同大小等级霸王枝长度与叶面积的异速生长关系
植物的枝与叶性状之间始终存在着维持几何大小和重力平衡的需求, 二者间的大小匹配, 只有满足机械力学和水分动力学的要求, 才能保持枝叶机械支撑的平衡以及水分运输和蒸腾的协调(Westoby et al., 2002 )。I级霸王枝长度与叶面积表现出近等速生长关系, II级和III级霸王枝长度增加的速度大于叶面积增加的速度, 即大个体植株生长的大叶导致更高比例的枝长度投入, 霸王枝长度与叶面积间的关系具有个体大小差异性的特点。可能原因如下: 1)光照是影响植物生存和繁衍的关键性生态因子, 光照强度制约着植物的光合作用和有机物产量(Li et al., 2014 )。结合草地群落学调查、表1 和表2 资料认为, 研究区草地群落的高度和密度均较高, I级霸王位于群落中下层, 较II级和III级霸王接受的光合有效辐射降低(表2 ), 光照资源竞争能力弱, 而且群落中其他物种的邻体干扰使之遭受了较强烈的遮阴, 降低了植物光合作用能力。为了更多地捕获群落中下层的光斑, 在枝条分形方式上, I级霸王倾向于增大小枝与其支撑枝的分枝角度进行横向拓展(表1 ), 试图增加小枝空间分布范围和枝叶规模, 增强枝叶空间拓展能力和叶片光合作用, 因而I级霸王枝长度与叶面积近等速生长。II级霸王植株高度增加, 分枝角度减小, 将有限的小枝资源较多的用于机械支撑器官以增强其直立生长的能力, 捕获更多的光照资源, 能够降低邻体的遮挡和群落遮蔽程度; 相对而言III级霸王植株占据群落高度优势(表1 ), 株丛内部分枝数量多, 光照下透量减少, 叶片自遮阴严重, 阻止了小枝在冠内生长, III级霸王进一步降低分枝角度, 试图增加枝长度向高空发展以降低小枝在植冠内的遮挡, 因而II和III级霸王选择生长较长的枝条可能更有利。2) I级霸王小枝的枝横截面积小, 输水阻力大, 在不利环境下小枝更容易出现栓塞(Mcculloh & Sperry, 2005 ), 往往导致传输效率降低, 只能满足较小叶片的营养和水分传输需求, 同时较小的叶面积有助于植物组织规避水分亏缺的风险(Woodward et al., 2004 ); II级和III级霸王植株高度不断增大, 使木质部水分输送的距离增加, 增加枝横截面积(表1 ), 提高植株输水效率, 有助于增强植株向叶片运输水分能力, 因此, 荒漠草地中随着植物大小等级的增加, 植株可能增大枝横截面积以满足水分供给需求。3)研究区常受风力作用, 由风力引起的弯曲力矩与叶片面积和叶柄长度的立方成正比(Niklas, 1999 ), 小枝除了供给叶片水分和养分外, 还需提供支撑由于风等外界因素对叶片的拖曳的力, 这要求枝条需额外的资源配置, 叶片越大, 受到风等外力作用就越大, 枝条相应的投入也越高, 因而较大等级的霸王枝长度较长。值得注意的是, 随着大小等级的增加, 霸王枝长度逐渐增加, 叶面积也逐渐增大, 主要是由于自然选择的压力促使植物器官间沿最优化的方向进化(Enquist & Niklas, 2002 ), 大叶能够平铺排布, 有效地捕获来自垂直角度的光和大部分的光照(Horn, 1971 ), 为满足植株光合作用和水分运输的需求, 霸王植株叶面积与枝的输导面积相对应, 使得枝条所支撑的叶面积与枝条导管横截面积必须成比例。
3.2 不同大小等级霸王枝长度与叶数量的异速生长关系
植物的形态可塑性是适应异质环境的一个重要特征, 使植物能够把资源最佳地分配到不同的器官, 以提高物种生存适合度(李钰等, 2013a )。研究发现: 不同大小等级霸王的枝长度与叶数量呈正的异速生长关系, 回归方程具有共同的SMA斜率且显著小于1 (p<0.05) (图2 ), 表明枝长度的增加速度大于叶片数的增加速度, 这符合“长的当年生小枝比短的当年生小枝具有更小的叶数量与枝长度的比值, 即单位枝长度的叶片数更少”的结论(Yagi, 2004 )。不同大小等级枝长度与叶数量y轴截距的变化则表明霸王投入的枝长度一定时, 大个体植株的叶数量较少, 这种差异性可能归因于以下3个方面: (1)霸王叶片对生或簇生, 均匀地排列在枝条上(周向睿等, 2006 ), 彼此之间相互作用、相互关联, 有利于小枝与叶片内部物质传输和转移, 使得这些小枝与其上着生叶片的生长具有密切的关系。结合表3 中叶面积的变化趋势, 推测可能是随着霸王个体大小等级的增加, 其枝条长度逐渐增大, 叶片数与叶面积均增加, 过多和过大的叶片都会造成上部叶片对下部叶片光照的遮挡, 使植株下部光合有效辐射降低, 产生低效率的光合作用, 同时随着分枝角度的逐渐减小, 枝条之间相互拥挤程度加剧, 造成叶片之间相互遮光严重, 因而较大等级霸王植株叶数量减小以提高植株内部光照资源的获取效率。(2)可能是叶片“大小-数量”的权衡对枝长度与叶片数关系产生影响。Kleiman和Aarssen (2007)以及杨冬梅等(2012)在小枝内部“大小-数量”权衡关系的研究中发现, 叶片“大小-数量”间呈等速负相关关系, 在单位枝条大小上着生的叶片面积越大数量越少, 反之亦然。随着个体大小等级的增大, 霸王增加叶面积的投资意味着植株将通过减少叶数量的方式来弥补大叶构建和扩展增加的成本, 试图使内部的自我荫蔽程度最小化(Niklas, 1999 )。因此, 霸王叶数量与枝长度在y轴截距的差异也可能是叶片大小变异影响的结果。(3)在进化过程中为提高光合收益, 植物应该增大获取限制性资源的器官分配, 而减少获取非限制资源的器官分配(Pickup et al., 2005 ; Niinemets et al., 2006 )。为了保证其生存和发展的光合产物需求, I级霸王生长大量小叶可以提高叶片配置的机动性来接收透射光, 增加植株的光合收益效率; II和III级霸王随着植株高度的增加, 接受的太阳辐射能较多, 若仍生长大量的大叶有可能导致呼吸和蒸腾成本更高, 不利于其光拦截能力和光合碳获取能力最大化(Wright et al., 2002 ), 因而较大等级的霸王选择生长少量的大叶可能更有利。
形态特征和生长特性的改变是植物适应不同环境和资源水平的重要策略(Maherali & Delucia, 2001 ), 植物功能性状与生存环境之间是一种相互协同进化与适应的关系(侯兆疆等, 2014 )。枝、叶作为植物的两个重要器官, 其关系不仅是不同构件单元大小数量关系权衡的结果, 也是适应不同光环境条件下能量交换效率的一个副产物。本研究中霸王不同大小等级植株枝叶性状关系存在差异, 在给定的枝长度下, 霸王小个体植株选择小的叶面积和多的叶数量, 大个体植株则选择了大的叶面积和少的叶数量, 霸王枝叶性状间的个体大小差异性较好地验证了植物通过功能性状的可塑性调整响应光环境变异的协同适应性机制, 反映了植物不同个体大小的冠层构型和环境适应性策略, 通过异速生长分析植物生长速率上的可塑性, 可以更好地了解植物在资源分配上的生境依赖规律, 从而进一步了解植物对环境的响应特征。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Aarssen LW 2015 ). Body size and fitness in plants: Revisiting the selection consequences of competition.
Perspectives in Plant Ecology, Evolution and Systematics , 17 , 236 -242 .
[本文引用: 1]
[2]
Alméras T Costes E Salles JC 2004 ). Identification of biomechanical actors involved in stem shape variability between pricot tree varieties.
Annals of Botany , 93 , 55 -468 .
[本文引用: 1]
[3]
Barthélémy D Caraglio Y 2007 ). Plant architecture: A dynamic, multilevel and comprehensive approach to plant form, structure and ontogeny.
Annuals of Botany , 99 , 375 -407 .
[本文引用: 1]
[4]
Brouat C Gibernau M Amsellem L McKey D 1998 ). Corner’s rules revisited: Ontogenetic and interspecific patterns in leaf-stem allometry.
New Phytologist , 139 , 459 -470 .
[本文引用: 1]
[5]
Chen M Zhao XY Zuo XA Lian J Zhu YC 2015 ). Floral traits and pollination system of Zygophyllum xanthoxylum in the managed and wild populations in an arid region of Northwest China.
Journal of Arid Land , 7 , 488 -500 .
[本文引用: 1]
[6]
Cheng DL Zhong QL Lin MZ Jin MF Qian RF 2011 ). The advance of allometric studies on plant metabolic rates and biomass.
Acta Ecologica Sinica , 31 , 2312 -2320 . (in Chinese with English abstract)[程栋梁 , 钟全林 , 林茂兹 , 金美芳 , 钱瑞芳 (2011 ). 植物代谢速率与个体生物量关系研究进展
. 生态学报 , 31 , 2312 -2320 .]
[本文引用: 1]
[7]
de Kroons H Hutchings MJ 1995 ). Morphological plasticity in clonal plants: The foraging concept reconsidered.
Journal of Ecology , 83 , 143 -152 .
[本文引用: 1]
[8]
Enquist BJ Niklas KJ 2001 ). Invariant scaling relations across tree-dominated communities.
Nature , 410 , 655 -660 .
[本文引用: 1]
[9]
Enquist BJ Niklas KJ 2002 ). Global allocation rules for patterns of biomass partitioning in seed plants.
Science , 295 , 1517 -1520 .
[本文引用: 2]
[10]
Falster DS Warton DI Wright IJ
URL
[本文引用: 1]
[11]
Givnish TJ Vermeij GJ 1976 ). Sizes and shapes of liane leaves.
The American Naturalist , 110 , 743 -778 .
[本文引用: 1]
[12]
Heuret P Meredieu C Coudurier T Courdier F Barthélémy D 2006 ). Ontogenetic trends in the morphological features of main stem annual shoots of Pinus pinaster (Pinaceae).
American Journal of Botany , 93 , 1577 -1587 .
[13]
Horn HS 1971). The Adaptive Geometry of Trees . Vol. 3. Princeton University Press, Princeton, USA .
[本文引用: 1]
[14]
Hou ZJ Zhao CZ Li Y Zhang Q Ma XL 2014 ). Trade-off between height and branch numbers in Stellera chamaejasme on slopes of different aspects in a degraded alpine grassland.
Chinese Journal of Plant Ecology , 38 , 281 -288 . (in Chinese with English abstract)[侯兆疆 , 赵成章 , 李钰 , 张茜 , 马小丽 (2014 ). 不同坡向高寒退化草地狼毒株高和枝条数的权衡关系
. 植物生态学报 , 38 , 281 -288 .]
[15]
Kleiman D Aarssen LW 2007 ). The leaf size/number trade-off in trees.
Journal of Ecology , 95 , 376 -382 .
[本文引用: 1]
[16]
Kleyer M Minden V 2015 ). Why functional ecology should consider all plant organs: An allocation-based perspective.
Basic and Applied Ecology , 16 , 1 -9 .
[本文引用: 1]
[17]
Li L Li XY Xu XW Lin LS Zeng FJ Chen FL 2014 ). Assimilative branches and leaves of the desert plant Alhagi sparsifolia Shap. possesses a different adaptation mechanism to shade.
Plant Physiology and Biochemistry , 74 , 239 -245 .
[18]
Li Y Zhao CZ Dong XG Hou ZJ Ma XL Zhang Q 2013 a). Twig and leaf trait differences in Stellera chamaejasme with slope in alpine grassland.
Chinese Journal of Plant Ecology , 37 , 709 -717 . (in Chinese with English abstract)[李钰 , 赵成章 , 董小刚 , 侯兆疆 , 马小丽 , 张茜 (2013 a). 高寒草地狼毒枝-叶性状的坡度差异性
. 植物生态学报 , 37 , 709 -717 .]
[19]
Li Y Zhao CZ Hou ZJ Ma XL Zhang Q 2013 b). Body size and stem-and leaf allometry of Stellera chamaejasme in degraded alpine grassland.
Chinese Journal of Ecology , 32 , 241 -246 . (in Chinese with English abstract)[李钰 , 赵成章 , 侯兆疆 , 马小丽 , 张茜 (2013 b). 高寒退化草地狼毒种群个体大小与茎、叶的异速生长
. 生态学杂志 , 32 , 241 -246 .]
[本文引用: 1]
[20]
Li YN Yang DM Sun SC Gao XM 2008 ). Effects of twig size on biomass allocation within twigs and on lamina area supporting efficiency in Rhododendron: Allometric scaling analyses. Journal of Plant Ecology (
Chinese Version ), 32 , 1175 -1183 .(in Chinese with English abstract) [李亚男 , 杨冬梅 , 孙书存 , 高贤明 (2008 ). 杜鹃花属植物小枝大小对小枝生物量分配及叶面积支持效率的影响: 异速生长分析. 植物生态学报,
32 , 1175 -1183 .]
[本文引用: 1]
[21]
Lin YM Hong T Wu CZ Hong W Hu XS Fan HL Song P 2007 ). Architectural analysis of Osmanthus fragrans crown geometry.
Journal of Tropical and Subtropical Botany , 15 , 301 -306 . (in Chinese with English abstract)[林勇明 , 洪滔 , 吴承祯 , 洪伟 , 胡喜生 , 范海兰 , 宋萍 (2007 ). 桂花植冠的枝系构型分析
. 热带亚热带植物学报 , 15 , 301 -306 .]
[本文引用: 1]
[22]
Maherali H Delucia EH 2001 ). Influence of climate-driven shifts in biomass allocation on water transport and storage in ponderosa pine.
Oecologia , 129 , 481 -491 .
[本文引用: 1]
[23]
Mcculloh KA Sperry JS 2005 ). Patterns in hydraulic architecture and their implications for transport efficiency.
Tree Physiology , 25 , 257 -267 .
[本文引用: 1]
[24]
Nan J Zhao XY Yuan H Zhang LL 2014 ). Phenotype differences between Zygophyllum xanthoxylum and Salsola arbuscula from open and wind-protected sites.
Acta Ecologica Sinica , 34 , 5758 -5765 . (in Chinese with English abstract)[南江 , 赵晓英 , 原慧 , 张琳琳 (2014 ). 霸王和木本猪毛菜在遮风和不遮风环境下的表型特征差异
. 生态学报 , 34 , 5758 -5765 .]
[本文引用: 1]
[25]
Niinemets Ü Portsmuth A Tobias M 2006 ). Leaf size modifies support biomass distribution among stems, petioles and midribs in temperate plants.
New Phytologist , 171 , 91 -104 .
[本文引用: 1]
[26]
Niinemets Ü Portsmuth A Tobias M 2007 ). Leaf shape and venation pattern alter the support investments within leaf lamina in temperate species: A neglected source of leaf physiological differentiation?
Functional Ecology , 21 , 28 -40 .
[本文引用: 2]
[27]
Niklas KJ 1999 ). A mechanical perspective on foliage leaf form and function.
New Phytologist , 143 , 19 -31 .
[本文引用: 1]
[28]
Parkhurst DF Loucks OL 1972 ). Optimal leaf size in relation to environment.
Journal of Ecology , 60 , 505 -537 .
[本文引用: 1]
[29]
Pickup M Westoby M Basden A 2005 ). Dry mass costs of deploying leaf area in relation to leaf size.
Functional Ecology , 19 , 88 -97 .
[30]
Pitman E 1939 ). A note on normal correlation.
Biometrika , 31 , 9 -12 .
[本文引用: 1]
[31]
Shi YC Zhao CZ Song QH Du J Chen J Wang JW 2015 ). Slope-related variations in twig and leaf traits of Robinia pseudoacacia in the northern mountains of Lanzhou.
Chinese Journal of Plant Ecology , 39 , 362 -370 . (in Chinese with English abstract)[史元春 , 赵成章 , 宋清华 , 杜晶 , 陈静 , 王继伟 (2015 ). 兰州北山刺槐枝叶性状的坡向差异性
. 植物生态学报 , 39 , 362 -370 .]
[32]
Valladares F Skillman JB Pearcy RW 2002 ). Convergence in light capture efficiencies among tropical forest understory plants with contrasting crown architectures: A case of morphological compensation.
American Journal of Botany , 89 , 1275 -1284 .
[本文引用: 1]
[33]
Violle C Navas ML Vile D Kazakou E Fortunel C Hummel I Garnier E 2007 ). Let the concept of trait be functional!
Oikos , 116 , 882 -892 .
[本文引用: 1]
[34]
Wang Y Wei XL 2010 ). Advance on the effects of different light environments on growth, physiological biochemistry and morphostructure of plant.
Journal of Mountain Agriculture and Biology , 29 , 353 -359 . (in Chinese with English abstract)[王艺 , 韦小丽 (2010 ). 不同光照对植物生长、生理生化和形态结构影响的研究进展
. 山地农业生物学报 , 29 , 353 -359 .]
[35]
Warton DI Weber NC 2002 ). Common slope tests for bivariateerrors-in-variables models.
Biometrical Journal , 44 , 161 -174 .
[本文引用: 1]
[36]
Warton DI Wright IJ Falster DS Westoby M 2006 ). Bivariate line-fitting methods for allometry.
Biological Reviews , 81 , 259 -291 .
[本文引用: 1]
[37]
Westoby M Falster DS Moles AT Vesk PA Wright IJ 2002 ). Plant ecological strategies: Some leading dimensions of variation between species.
Annual Review of Ecology and Systematics , 33 , 125 -159 .
[本文引用: 1]
[38]
Woodward FI Lomas MR Kelly CK 2004 ). Global climate and the distribution of plant biomes.
Philosophical Transactions of the Royal Society B: Biological Sciences , 359 , 1465 -1476 .
[本文引用: 1]
[39]
Wright IJ Westoby M Reich PB 2002 ). Convergence towards higher leaf mass per area in dry and nutrient-poor habitats has different consequences for leaf life span.
Journal of Ecology , 90 , 534 -543 .
[本文引用: 1]
[40]
Wu GL Chen M Du GZ 2010 ). Response of biomass allocation and morphological characteristics to light and nutrient resources for seedlings of three alpine species.
Acta Ecologica Sinica , 30 , 60 -66 .(in Chinese with English abstract) [武高林 , 陈敏 , 杜国祯 (2010 ). 三种高寒植物幼苗生物量分配及性状特征对光照和养分的响应. 生态学报,
30 , 60 -66 .]
[本文引用: 1]
[41]
Xiao Y Tao Y Zhang YM 2014 ). Biomass allocation and leaf stoichiometric characteristics in four desert herbaceous plants during different growth periods in the Gurbantünggüt Desert, China.
Chinese Journal of Plant Ecology , 38 , 929 -940 . (in Chinese with English abstract)[肖遥 , 陶冶 , 张元明 (2014 ). 古尔班通古特沙漠4种荒漠草本植物不同生长期的生物量分配与叶片化学计量特征
. 植物生态学报 , 38 , 929 -940 .]
[本文引用: 1]
[42]
Xu Q Yang ZH Guo SJ Wang QQ 2013 ). Branch system configuration features of Haloxylon ammodendron in different growth stages.
Journal of Northwest Forestry University , 28 , 50 -54 . (in Chinese with English abstract)[许强 , 杨自辉 , 郭树江 , 王强强 (2013 ). 梭梭不同生长阶段的枝系构型特征
. 西北林学院学报 , 28 , 50 -54 .]
[本文引用: 1]
[43]
Yagi T 2004 ). Within-tree variations in shoot differentiation patterns of 10 tall tree species in a Japanese cool temperate forest.
Canadian Journal of Botany , 82 , 228 -243 .
[44]
Yang DM Zhan F Zhang HW 2012 ). Trade-off between leaf size and number in current-year twigs of deciduous broad-leaved woody species at different altitudes on Qingliang Mountain, southeastern China.
Chinese Journal of Plant Ecology , 36 , 281 -291 . (in Chinese with English abstract)[杨冬梅 , 占峰 , 张宏伟 (2012 ). 清凉峰不同海拔木本植物小枝内叶大小-数量权衡关系,
植物生态学报 , 36 , 281 -291 .]
[本文引用: 1]
[45]
Yang HX Zhang JT Wu B Li XS Zhang YY 2006 ). Point pattern analysis of Artemisia ordosica population in the Mu Us sandy land. Journal of Plant Ecology (
Chinese Version ), 30 , 563 -570 . (in Chinese with English abstract)[杨洪晓 , 张金屯 , 吴波 , 李晓松 , 张友炎 (2006 ). 毛乌素沙地油蒿种群点格局分析
. 植物生态学报 , 30 , 563 -570 .]
[本文引用: 1]
[46]
Yao J Li Y Wei LP Jiang SS Yang S Hou JH 2013 ). Changes of allometric relationships among leaf traits in different ontogenetic stages of Acer mono from different types of forests in Donglingshan of Beijing.
Acta Ecologica Sinica , 33 , 3907 -3915 . (in Chinese with English abstract)[姚婧 , 李颖 , 魏丽萍 , 蒋思思 , 杨松 , 侯继华 (2013 ). 东灵山不同林型五角枫叶性状异速生长关系随发育阶段的变化
. 生态学报 , 33 , 3907 -3915 .]
[本文引用: 1]
[47]
Zhou XR Zhou ZY Wu CX 2006 ). The research of the breeding characters of
Zygophyllum xanthoxylum. Pratacultural Science , 23 , 38 -41 .(in Chinese with English abstract) [周向睿 , 周志宇 , 吴彩霞 (2006 ). 霸王繁殖特性的研究. 草业科学,
23 , 38 -41 .]
Body size and fitness in plants: Revisiting the selection consequences of competition.
1
2015
... 蒺藜科霸王属霸王(Zygophyllum xanthoxylum)是分布于我国西北部干旱荒漠区的强旱生小灌木, 当年生枝条数量多、叶量大、生长快, 具有抗旱、速生、高产、耐沙埋、耐贫瘠、抗风蚀的能力, 具有较好的饲用价值和适口性, 是荒漠灌丛植被的主要优势种和建群种(Chen et al., 2015 ).目前, 许多学者已对植物个体大小构型差异与竞争环境的关系(Aarssen, 2015 )、不同发育阶段分枝格局与叶片生长变化规律(许强等, 2013 )、不同个体大小茎-叶资源分配策略(李钰等, 2013b )等方面进行了大量研究, 解释了植物适应特定环境的生长特性与物质分配规律.霸王在不同环境下的表型特征差异也得到了关注(南江等, 2014 ), 有关霸王枝、叶性状关系的个体大小差异及其对不同光照强度生长特性的研究还不够深入, 影响了植物枝叶功能性状对不同生境适应机制的认识.因此, 本研究从植株个体大小的角度研究了荒漠草地霸王的枝叶构型特征及其光环境响应策略, 试图揭示: (1)不同大小等级霸王小枝内枝大小、叶面积和叶数量资源配置模式; (2)这种资源配置模式对霸王枝长度与叶大小、枝长度和叶数量间异速生长关系的影响. ...
Identification of biomechanical actors involved in stem shape variability between pricot tree varieties.
1
2004
... 在不同环境下, 植物种群的发育特性和物质分配策略是植物局部特化的形态特征对资源异质性的响应(Kleyer & Minden, 2015 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ; 史元春等, 2015 ), 在植物的整个生长过程中, 功能性状是植物与环境相互作用的结果, 如何调节小枝内各构件的生长及关系, 是植物生存策略和生境适合度调整的重要内容(肖遥等, 2014 ).在植物各器官中, 枝条和叶片是重要的输导和物质生产器官, 在生物力学及功能上具有密切联系.枝条具有导水功能和机械支撑作用(Alméras et al., 2004 ), 其大小往往关系到植物的空间拓展策略, 决定叶片的物质投资方式; 叶片大小和多少直接影响着植株的发育模式, 以及植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ), 枝、叶关系不仅是植物个体发育过程的表现(Brouat et al., 1998 ), 也体现了植物对特定生境的适应性(Violle et al., 2007 ).植物的形态特征和枝、叶功能之间的平衡受年龄和个体大小制约(Heuret et al., 2006 ; Barthélémy & Caraglio, 2007 ), 不同大小株丛在群落中占据的资源生态位存在差异(姚婧等, 2013 ), 其受群落郁闭度和干扰程度不同, 不同层次枝叶接受的有效太阳辐射也不同, 从而影响植物的空间分布格局及光照、温度等环境条件(程栋梁等, 2011 ), 往往导致不同个体大小植株分枝构型出现明显的分化.为了提高植物构型构件的生境适应性和自身的生存适合度, 不同大小等级植株可能需要适时调整其分枝角度, 增加植株所接受的光照强度, 促使植株枝-叶性状生长及其相互关系进行相应的调整(林勇明等, 2007 ), 使种群内每个植株可获得性资源的数量最大化, 构建与生境协同进化的冠层构型(de Kroons & Hutchings, 1995 ; 武高林等, 2010 ), 体现了植物适应特定环境的构型 塑造. ...
Plant architecture: A dynamic, multilevel and comprehensive approach to plant form, structure and ontogeny.
1
2007
... 在不同环境下, 植物种群的发育特性和物质分配策略是植物局部特化的形态特征对资源异质性的响应(Kleyer & Minden, 2015 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ; 史元春等, 2015 ), 在植物的整个生长过程中, 功能性状是植物与环境相互作用的结果, 如何调节小枝内各构件的生长及关系, 是植物生存策略和生境适合度调整的重要内容(肖遥等, 2014 ).在植物各器官中, 枝条和叶片是重要的输导和物质生产器官, 在生物力学及功能上具有密切联系.枝条具有导水功能和机械支撑作用(Alméras et al., 2004 ), 其大小往往关系到植物的空间拓展策略, 决定叶片的物质投资方式; 叶片大小和多少直接影响着植株的发育模式, 以及植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ), 枝、叶关系不仅是植物个体发育过程的表现(Brouat et al., 1998 ), 也体现了植物对特定生境的适应性(Violle et al., 2007 ).植物的形态特征和枝、叶功能之间的平衡受年龄和个体大小制约(Heuret et al., 2006 ; Barthélémy & Caraglio, 2007 ), 不同大小株丛在群落中占据的资源生态位存在差异(姚婧等, 2013 ), 其受群落郁闭度和干扰程度不同, 不同层次枝叶接受的有效太阳辐射也不同, 从而影响植物的空间分布格局及光照、温度等环境条件(程栋梁等, 2011 ), 往往导致不同个体大小植株分枝构型出现明显的分化.为了提高植物构型构件的生境适应性和自身的生存适合度, 不同大小等级植株可能需要适时调整其分枝角度, 增加植株所接受的光照强度, 促使植株枝-叶性状生长及其相互关系进行相应的调整(林勇明等, 2007 ), 使种群内每个植株可获得性资源的数量最大化, 构建与生境协同进化的冠层构型(de Kroons & Hutchings, 1995 ; 武高林等, 2010 ), 体现了植物适应特定环境的构型 塑造. ...
Corner’s rules revisited: Ontogenetic and interspecific patterns in leaf-stem allometry.
1
1998
... 在不同环境下, 植物种群的发育特性和物质分配策略是植物局部特化的形态特征对资源异质性的响应(Kleyer & Minden, 2015 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ; 史元春等, 2015 ), 在植物的整个生长过程中, 功能性状是植物与环境相互作用的结果, 如何调节小枝内各构件的生长及关系, 是植物生存策略和生境适合度调整的重要内容(肖遥等, 2014 ).在植物各器官中, 枝条和叶片是重要的输导和物质生产器官, 在生物力学及功能上具有密切联系.枝条具有导水功能和机械支撑作用(Alméras et al., 2004 ), 其大小往往关系到植物的空间拓展策略, 决定叶片的物质投资方式; 叶片大小和多少直接影响着植株的发育模式, 以及植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ), 枝、叶关系不仅是植物个体发育过程的表现(Brouat et al., 1998 ), 也体现了植物对特定生境的适应性(Violle et al., 2007 ).植物的形态特征和枝、叶功能之间的平衡受年龄和个体大小制约(Heuret et al., 2006 ; Barthélémy & Caraglio, 2007 ), 不同大小株丛在群落中占据的资源生态位存在差异(姚婧等, 2013 ), 其受群落郁闭度和干扰程度不同, 不同层次枝叶接受的有效太阳辐射也不同, 从而影响植物的空间分布格局及光照、温度等环境条件(程栋梁等, 2011 ), 往往导致不同个体大小植株分枝构型出现明显的分化.为了提高植物构型构件的生境适应性和自身的生存适合度, 不同大小等级植株可能需要适时调整其分枝角度, 增加植株所接受的光照强度, 促使植株枝-叶性状生长及其相互关系进行相应的调整(林勇明等, 2007 ), 使种群内每个植株可获得性资源的数量最大化, 构建与生境协同进化的冠层构型(de Kroons & Hutchings, 1995 ; 武高林等, 2010 ), 体现了植物适应特定环境的构型 塑造. ...
Floral traits and pollination system of Zygophyllum xanthoxylum in the managed and wild populations in an arid region of Northwest China.
1
2015
... 蒺藜科霸王属霸王(Zygophyllum xanthoxylum)是分布于我国西北部干旱荒漠区的强旱生小灌木, 当年生枝条数量多、叶量大、生长快, 具有抗旱、速生、高产、耐沙埋、耐贫瘠、抗风蚀的能力, 具有较好的饲用价值和适口性, 是荒漠灌丛植被的主要优势种和建群种(Chen et al., 2015 ).目前, 许多学者已对植物个体大小构型差异与竞争环境的关系(Aarssen, 2015 )、不同发育阶段分枝格局与叶片生长变化规律(许强等, 2013 )、不同个体大小茎-叶资源分配策略(李钰等, 2013b )等方面进行了大量研究, 解释了植物适应特定环境的生长特性与物质分配规律.霸王在不同环境下的表型特征差异也得到了关注(南江等, 2014 ), 有关霸王枝、叶性状关系的个体大小差异及其对不同光照强度生长特性的研究还不够深入, 影响了植物枝叶功能性状对不同生境适应机制的认识.因此, 本研究从植株个体大小的角度研究了荒漠草地霸王的枝叶构型特征及其光环境响应策略, 试图揭示: (1)不同大小等级霸王小枝内枝大小、叶面积和叶数量资源配置模式; (2)这种资源配置模式对霸王枝长度与叶大小、枝长度和叶数量间异速生长关系的影响. ...
植物代谢速率与个体生物量关系研究进展
1
2011
... 在不同环境下, 植物种群的发育特性和物质分配策略是植物局部特化的形态特征对资源异质性的响应(Kleyer & Minden, 2015 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ; 史元春等, 2015 ), 在植物的整个生长过程中, 功能性状是植物与环境相互作用的结果, 如何调节小枝内各构件的生长及关系, 是植物生存策略和生境适合度调整的重要内容(肖遥等, 2014 ).在植物各器官中, 枝条和叶片是重要的输导和物质生产器官, 在生物力学及功能上具有密切联系.枝条具有导水功能和机械支撑作用(Alméras et al., 2004 ), 其大小往往关系到植物的空间拓展策略, 决定叶片的物质投资方式; 叶片大小和多少直接影响着植株的发育模式, 以及植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ), 枝、叶关系不仅是植物个体发育过程的表现(Brouat et al., 1998 ), 也体现了植物对特定生境的适应性(Violle et al., 2007 ).植物的形态特征和枝、叶功能之间的平衡受年龄和个体大小制约(Heuret et al., 2006 ; Barthélémy & Caraglio, 2007 ), 不同大小株丛在群落中占据的资源生态位存在差异(姚婧等, 2013 ), 其受群落郁闭度和干扰程度不同, 不同层次枝叶接受的有效太阳辐射也不同, 从而影响植物的空间分布格局及光照、温度等环境条件(程栋梁等, 2011 ), 往往导致不同个体大小植株分枝构型出现明显的分化.为了提高植物构型构件的生境适应性和自身的生存适合度, 不同大小等级植株可能需要适时调整其分枝角度, 增加植株所接受的光照强度, 促使植株枝-叶性状生长及其相互关系进行相应的调整(林勇明等, 2007 ), 使种群内每个植株可获得性资源的数量最大化, 构建与生境协同进化的冠层构型(de Kroons & Hutchings, 1995 ; 武高林等, 2010 ), 体现了植物适应特定环境的构型 塑造. ...
Morphological plasticity in clonal plants: The foraging concept reconsidered.
1
1995
... 在不同环境下, 植物种群的发育特性和物质分配策略是植物局部特化的形态特征对资源异质性的响应(Kleyer & Minden, 2015 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ; 史元春等, 2015 ), 在植物的整个生长过程中, 功能性状是植物与环境相互作用的结果, 如何调节小枝内各构件的生长及关系, 是植物生存策略和生境适合度调整的重要内容(肖遥等, 2014 ).在植物各器官中, 枝条和叶片是重要的输导和物质生产器官, 在生物力学及功能上具有密切联系.枝条具有导水功能和机械支撑作用(Alméras et al., 2004 ), 其大小往往关系到植物的空间拓展策略, 决定叶片的物质投资方式; 叶片大小和多少直接影响着植株的发育模式, 以及植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ), 枝、叶关系不仅是植物个体发育过程的表现(Brouat et al., 1998 ), 也体现了植物对特定生境的适应性(Violle et al., 2007 ).植物的形态特征和枝、叶功能之间的平衡受年龄和个体大小制约(Heuret et al., 2006 ; Barthélémy & Caraglio, 2007 ), 不同大小株丛在群落中占据的资源生态位存在差异(姚婧等, 2013 ), 其受群落郁闭度和干扰程度不同, 不同层次枝叶接受的有效太阳辐射也不同, 从而影响植物的空间分布格局及光照、温度等环境条件(程栋梁等, 2011 ), 往往导致不同个体大小植株分枝构型出现明显的分化.为了提高植物构型构件的生境适应性和自身的生存适合度, 不同大小等级植株可能需要适时调整其分枝角度, 增加植株所接受的光照强度, 促使植株枝-叶性状生长及其相互关系进行相应的调整(林勇明等, 2007 ), 使种群内每个植株可获得性资源的数量最大化, 构建与生境协同进化的冠层构型(de Kroons & Hutchings, 1995 ; 武高林等, 2010 ), 体现了植物适应特定环境的构型 塑造. ...
Invariant scaling relations across tree-dominated communities.
1
2001
... 光是植物生长的能量来源, 不同的光照强度对植物光合作用的影响主要是通过植物对光照的捕获利用能力来实现的(王艺和韦小丽, 2010 ).Valladares等(2002)指出, 叶内与茎内支撑投资之间可能存在某种权衡关系, 进而导致不同的冠层设计, 使得捕光效率在总体上是相似的.叶面积的增大需要更长的枝条来减少个体内部对光照的相互遮挡, 叶片面积和数量的增加则要求枝条具有更高的支撑能力(Niinemets et al., 2007 ), 大个体植株较小个体植株能生长更粗大的枝条用于支持功能, 体现了植物枝-叶生长中的物质和能量转换关系(Enquist & Niklas, 2001 , 2002 ).本研究发现, 随着霸王大小等级的增加, 枝长度与叶面积关系的异速斜率逐渐降低, 即枝长度的生长速度逐渐高于叶面积的生长速度; 枝长度一定的条件下, 相对于小个体霸王, 大个体霸王植株叶数量的增加速率降低, 自然选择使它们以最佳的收益/代价关系以满足不同个体大小植株的生长需求. ...
Global allocation rules for patterns of biomass partitioning in seed plants.
2
2002
... 光是植物生长的能量来源, 不同的光照强度对植物光合作用的影响主要是通过植物对光照的捕获利用能力来实现的(王艺和韦小丽, 2010 ).Valladares等(2002)指出, 叶内与茎内支撑投资之间可能存在某种权衡关系, 进而导致不同的冠层设计, 使得捕光效率在总体上是相似的.叶面积的增大需要更长的枝条来减少个体内部对光照的相互遮挡, 叶片面积和数量的增加则要求枝条具有更高的支撑能力(Niinemets et al., 2007 ), 大个体植株较小个体植株能生长更粗大的枝条用于支持功能, 体现了植物枝-叶生长中的物质和能量转换关系(Enquist & Niklas, 2001 , 2002 ).本研究发现, 随着霸王大小等级的增加, 枝长度与叶面积关系的异速斜率逐渐降低, 即枝长度的生长速度逐渐高于叶面积的生长速度; 枝长度一定的条件下, 相对于小个体霸王, 大个体霸王植株叶数量的增加速率降低, 自然选择使它们以最佳的收益/代价关系以满足不同个体大小植株的生长需求. ...
... 植物的枝与叶性状之间始终存在着维持几何大小和重力平衡的需求, 二者间的大小匹配, 只有满足机械力学和水分动力学的要求, 才能保持枝叶机械支撑的平衡以及水分运输和蒸腾的协调(Westoby et al., 2002 ).I级霸王枝长度与叶面积表现出近等速生长关系, II级和III级霸王枝长度增加的速度大于叶面积增加的速度, 即大个体植株生长的大叶导致更高比例的枝长度投入, 霸王枝长度与叶面积间的关系具有个体大小差异性的特点.可能原因如下: 1)光照是影响植物生存和繁衍的关键性生态因子, 光照强度制约着植物的光合作用和有机物产量(Li et al., 2014 ).结合草地群落学调查、表1 和表2 资料认为, 研究区草地群落的高度和密度均较高, I级霸王位于群落中下层, 较II级和III级霸王接受的光合有效辐射降低(表2 ), 光照资源竞争能力弱, 而且群落中其他物种的邻体干扰使之遭受了较强烈的遮阴, 降低了植物光合作用能力.为了更多地捕获群落中下层的光斑, 在枝条分形方式上, I级霸王倾向于增大小枝与其支撑枝的分枝角度进行横向拓展(表1 ), 试图增加小枝空间分布范围和枝叶规模, 增强枝叶空间拓展能力和叶片光合作用, 因而I级霸王枝长度与叶面积近等速生长.II级霸王植株高度增加, 分枝角度减小, 将有限的小枝资源较多的用于机械支撑器官以增强其直立生长的能力, 捕获更多的光照资源, 能够降低邻体的遮挡和群落遮蔽程度; 相对而言III级霸王植株占据群落高度优势(表1 ), 株丛内部分枝数量多, 光照下透量减少, 叶片自遮阴严重, 阻止了小枝在冠内生长, III级霸王进一步降低分枝角度, 试图增加枝长度向高空发展以降低小枝在植冠内的遮挡, 因而II和III级霸王选择生长较长的枝条可能更有利.2) I级霸王小枝的枝横截面积小, 输水阻力大, 在不利环境下小枝更容易出现栓塞(Mcculloh & Sperry, 2005 ), 往往导致传输效率降低, 只能满足较小叶片的营养和水分传输需求, 同时较小的叶面积有助于植物组织规避水分亏缺的风险(Woodward et al., 2004 ); II级和III级霸王植株高度不断增大, 使木质部水分输送的距离增加, 增加枝横截面积(表1 ), 提高植株输水效率, 有助于增强植株向叶片运输水分能力, 因此, 荒漠草地中随着植物大小等级的增加, 植株可能增大枝横截面积以满足水分供给需求.3)研究区常受风力作用, 由风力引起的弯曲力矩与叶片面积和叶柄长度的立方成正比(Niklas, 1999 ), 小枝除了供给叶片水分和养分外, 还需提供支撑由于风等外界因素对叶片的拖曳的力, 这要求枝条需额外的资源配置, 叶片越大, 受到风等外力作用就越大, 枝条相应的投入也越高, 因而较大等级的霸王枝长度较长.值得注意的是, 随着大小等级的增加, 霸王枝长度逐渐增加, 叶面积也逐渐增大, 主要是由于自然选择的压力促使植物器官间沿最优化的方向进化(Enquist & Niklas, 2002 ), 大叶能够平铺排布, 有效地捕获来自垂直角度的光和大部分的光照(Horn, 1971 ), 为满足植株光合作用和水分运输的需求, 霸王植株叶面积与枝的输导面积相对应, 使得枝条所支撑的叶面积与枝条导管横截面积必须成比例. ...
1
... 在不同环境下, 植物种群的发育特性和物质分配策略是植物局部特化的形态特征对资源异质性的响应(Kleyer & Minden, 2015 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ; 史元春等, 2015 ), 在植物的整个生长过程中, 功能性状是植物与环境相互作用的结果, 如何调节小枝内各构件的生长及关系, 是植物生存策略和生境适合度调整的重要内容(肖遥等, 2014 ).在植物各器官中, 枝条和叶片是重要的输导和物质生产器官, 在生物力学及功能上具有密切联系.枝条具有导水功能和机械支撑作用(Alméras et al., 2004 ), 其大小往往关系到植物的空间拓展策略, 决定叶片的物质投资方式; 叶片大小和多少直接影响着植株的发育模式, 以及植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ), 枝、叶关系不仅是植物个体发育过程的表现(Brouat et al., 1998 ), 也体现了植物对特定生境的适应性(Violle et al., 2007 ).植物的形态特征和枝、叶功能之间的平衡受年龄和个体大小制约(Heuret et al., 2006 ; Barthélémy & Caraglio, 2007 ), 不同大小株丛在群落中占据的资源生态位存在差异(姚婧等, 2013 ), 其受群落郁闭度和干扰程度不同, 不同层次枝叶接受的有效太阳辐射也不同, 从而影响植物的空间分布格局及光照、温度等环境条件(程栋梁等, 2011 ), 往往导致不同个体大小植株分枝构型出现明显的分化.为了提高植物构型构件的生境适应性和自身的生存适合度, 不同大小等级植株可能需要适时调整其分枝角度, 增加植株所接受的光照强度, 促使植株枝-叶性状生长及其相互关系进行相应的调整(林勇明等, 2007 ), 使种群内每个植株可获得性资源的数量最大化, 构建与生境协同进化的冠层构型(de Kroons & Hutchings, 1995 ; 武高林等, 2010 ), 体现了植物适应特定环境的构型 塑造. ...
Sizes and shapes of liane leaves.
1
1976
... 在不同环境下, 植物种群的发育特性和物质分配策略是植物局部特化的形态特征对资源异质性的响应(Kleyer & Minden, 2015 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ; 史元春等, 2015 ), 在植物的整个生长过程中, 功能性状是植物与环境相互作用的结果, 如何调节小枝内各构件的生长及关系, 是植物生存策略和生境适合度调整的重要内容(肖遥等, 2014 ).在植物各器官中, 枝条和叶片是重要的输导和物质生产器官, 在生物力学及功能上具有密切联系.枝条具有导水功能和机械支撑作用(Alméras et al., 2004 ), 其大小往往关系到植物的空间拓展策略, 决定叶片的物质投资方式; 叶片大小和多少直接影响着植株的发育模式, 以及植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ), 枝、叶关系不仅是植物个体发育过程的表现(Brouat et al., 1998 ), 也体现了植物对特定生境的适应性(Violle et al., 2007 ).植物的形态特征和枝、叶功能之间的平衡受年龄和个体大小制约(Heuret et al., 2006 ; Barthélémy & Caraglio, 2007 ), 不同大小株丛在群落中占据的资源生态位存在差异(姚婧等, 2013 ), 其受群落郁闭度和干扰程度不同, 不同层次枝叶接受的有效太阳辐射也不同, 从而影响植物的空间分布格局及光照、温度等环境条件(程栋梁等, 2011 ), 往往导致不同个体大小植株分枝构型出现明显的分化.为了提高植物构型构件的生境适应性和自身的生存适合度, 不同大小等级植株可能需要适时调整其分枝角度, 增加植株所接受的光照强度, 促使植株枝-叶性状生长及其相互关系进行相应的调整(林勇明等, 2007 ), 使种群内每个植株可获得性资源的数量最大化, 构建与生境协同进化的冠层构型(de Kroons & Hutchings, 1995 ; 武高林等, 2010 ), 体现了植物适应特定环境的构型 塑造. ...
Ontogenetic trends in the morphological features of main stem annual shoots of Pinus pinaster (Pinaceae).
2006
1
... 形态特征和生长特性的改变是植物适应不同环境和资源水平的重要策略(Maherali & Delucia, 2001 ), 植物功能性状与生存环境之间是一种相互协同进化与适应的关系(侯兆疆等, 2014 ).枝、叶作为植物的两个重要器官, 其关系不仅是不同构件单元大小数量关系权衡的结果, 也是适应不同光环境条件下能量交换效率的一个副产物.本研究中霸王不同大小等级植株枝叶性状关系存在差异, 在给定的枝长度下, 霸王小个体植株选择小的叶面积和多的叶数量, 大个体植株则选择了大的叶面积和少的叶数量, 霸王枝叶性状间的个体大小差异性较好地验证了植物通过功能性状的可塑性调整响应光环境变异的协同适应性机制, 反映了植物不同个体大小的冠层构型和环境适应性策略, 通过异速生长分析植物生长速率上的可塑性, 可以更好地了解植物在资源分配上的生境依赖规律, 从而进一步了解植物对环境的响应特征. ...
不同坡向高寒退化草地狼毒株高和枝条数的权衡关系
2014
The leaf size/number trade-off in trees.
1
2007
... 在不同环境下, 植物种群的发育特性和物质分配策略是植物局部特化的形态特征对资源异质性的响应(Kleyer & Minden, 2015 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ; 史元春等, 2015 ), 在植物的整个生长过程中, 功能性状是植物与环境相互作用的结果, 如何调节小枝内各构件的生长及关系, 是植物生存策略和生境适合度调整的重要内容(肖遥等, 2014 ).在植物各器官中, 枝条和叶片是重要的输导和物质生产器官, 在生物力学及功能上具有密切联系.枝条具有导水功能和机械支撑作用(Alméras et al., 2004 ), 其大小往往关系到植物的空间拓展策略, 决定叶片的物质投资方式; 叶片大小和多少直接影响着植株的发育模式, 以及植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ), 枝、叶关系不仅是植物个体发育过程的表现(Brouat et al., 1998 ), 也体现了植物对特定生境的适应性(Violle et al., 2007 ).植物的形态特征和枝、叶功能之间的平衡受年龄和个体大小制约(Heuret et al., 2006 ; Barthélémy & Caraglio, 2007 ), 不同大小株丛在群落中占据的资源生态位存在差异(姚婧等, 2013 ), 其受群落郁闭度和干扰程度不同, 不同层次枝叶接受的有效太阳辐射也不同, 从而影响植物的空间分布格局及光照、温度等环境条件(程栋梁等, 2011 ), 往往导致不同个体大小植株分枝构型出现明显的分化.为了提高植物构型构件的生境适应性和自身的生存适合度, 不同大小等级植株可能需要适时调整其分枝角度, 增加植株所接受的光照强度, 促使植株枝-叶性状生长及其相互关系进行相应的调整(林勇明等, 2007 ), 使种群内每个植株可获得性资源的数量最大化, 构建与生境协同进化的冠层构型(de Kroons & Hutchings, 1995 ; 武高林等, 2010 ), 体现了植物适应特定环境的构型 塑造. ...
Why functional ecology should consider all plant organs: An allocation-based perspective.
1
2015
... 植物的枝与叶性状之间始终存在着维持几何大小和重力平衡的需求, 二者间的大小匹配, 只有满足机械力学和水分动力学的要求, 才能保持枝叶机械支撑的平衡以及水分运输和蒸腾的协调(Westoby et al., 2002 ).I级霸王枝长度与叶面积表现出近等速生长关系, II级和III级霸王枝长度增加的速度大于叶面积增加的速度, 即大个体植株生长的大叶导致更高比例的枝长度投入, 霸王枝长度与叶面积间的关系具有个体大小差异性的特点.可能原因如下: 1)光照是影响植物生存和繁衍的关键性生态因子, 光照强度制约着植物的光合作用和有机物产量(Li et al., 2014 ).结合草地群落学调查、表1 和表2 资料认为, 研究区草地群落的高度和密度均较高, I级霸王位于群落中下层, 较II级和III级霸王接受的光合有效辐射降低(表2 ), 光照资源竞争能力弱, 而且群落中其他物种的邻体干扰使之遭受了较强烈的遮阴, 降低了植物光合作用能力.为了更多地捕获群落中下层的光斑, 在枝条分形方式上, I级霸王倾向于增大小枝与其支撑枝的分枝角度进行横向拓展(表1 ), 试图增加小枝空间分布范围和枝叶规模, 增强枝叶空间拓展能力和叶片光合作用, 因而I级霸王枝长度与叶面积近等速生长.II级霸王植株高度增加, 分枝角度减小, 将有限的小枝资源较多的用于机械支撑器官以增强其直立生长的能力, 捕获更多的光照资源, 能够降低邻体的遮挡和群落遮蔽程度; 相对而言III级霸王植株占据群落高度优势(表1 ), 株丛内部分枝数量多, 光照下透量减少, 叶片自遮阴严重, 阻止了小枝在冠内生长, III级霸王进一步降低分枝角度, 试图增加枝长度向高空发展以降低小枝在植冠内的遮挡, 因而II和III级霸王选择生长较长的枝条可能更有利.2) I级霸王小枝的枝横截面积小, 输水阻力大, 在不利环境下小枝更容易出现栓塞(Mcculloh & Sperry, 2005 ), 往往导致传输效率降低, 只能满足较小叶片的营养和水分传输需求, 同时较小的叶面积有助于植物组织规避水分亏缺的风险(Woodward et al., 2004 ); II级和III级霸王植株高度不断增大, 使木质部水分输送的距离增加, 增加枝横截面积(表1 ), 提高植株输水效率, 有助于增强植株向叶片运输水分能力, 因此, 荒漠草地中随着植物大小等级的增加, 植株可能增大枝横截面积以满足水分供给需求.3)研究区常受风力作用, 由风力引起的弯曲力矩与叶片面积和叶柄长度的立方成正比(Niklas, 1999 ), 小枝除了供给叶片水分和养分外, 还需提供支撑由于风等外界因素对叶片的拖曳的力, 这要求枝条需额外的资源配置, 叶片越大, 受到风等外力作用就越大, 枝条相应的投入也越高, 因而较大等级的霸王枝长度较长.值得注意的是, 随着大小等级的增加, 霸王枝长度逐渐增加, 叶面积也逐渐增大, 主要是由于自然选择的压力促使植物器官间沿最优化的方向进化(Enquist & Niklas, 2002 ), 大叶能够平铺排布, 有效地捕获来自垂直角度的光和大部分的光照(Horn, 1971 ), 为满足植株光合作用和水分运输的需求, 霸王植株叶面积与枝的输导面积相对应, 使得枝条所支撑的叶面积与枝条导管横截面积必须成比例. ...
Assimilative branches and leaves of the desert plant Alhagi sparsifolia Shap. possesses a different adaptation mechanism to shade.
2014
a). 高寒草地狼毒枝-叶性状的坡度差异性
2013
b). 高寒退化草地狼毒种群个体大小与茎、叶的异速生长
1
2013
... 在不同环境下, 植物种群的发育特性和物质分配策略是植物局部特化的形态特征对资源异质性的响应(Kleyer & Minden, 2015 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ; 史元春等, 2015 ), 在植物的整个生长过程中, 功能性状是植物与环境相互作用的结果, 如何调节小枝内各构件的生长及关系, 是植物生存策略和生境适合度调整的重要内容(肖遥等, 2014 ).在植物各器官中, 枝条和叶片是重要的输导和物质生产器官, 在生物力学及功能上具有密切联系.枝条具有导水功能和机械支撑作用(Alméras et al., 2004 ), 其大小往往关系到植物的空间拓展策略, 决定叶片的物质投资方式; 叶片大小和多少直接影响着植株的发育模式, 以及植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ), 枝、叶关系不仅是植物个体发育过程的表现(Brouat et al., 1998 ), 也体现了植物对特定生境的适应性(Violle et al., 2007 ).植物的形态特征和枝、叶功能之间的平衡受年龄和个体大小制约(Heuret et al., 2006 ; Barthélémy & Caraglio, 2007 ), 不同大小株丛在群落中占据的资源生态位存在差异(姚婧等, 2013 ), 其受群落郁闭度和干扰程度不同, 不同层次枝叶接受的有效太阳辐射也不同, 从而影响植物的空间分布格局及光照、温度等环境条件(程栋梁等, 2011 ), 往往导致不同个体大小植株分枝构型出现明显的分化.为了提高植物构型构件的生境适应性和自身的生存适合度, 不同大小等级植株可能需要适时调整其分枝角度, 增加植株所接受的光照强度, 促使植株枝-叶性状生长及其相互关系进行相应的调整(林勇明等, 2007 ), 使种群内每个植株可获得性资源的数量最大化, 构建与生境协同进化的冠层构型(de Kroons & Hutchings, 1995 ; 武高林等, 2010 ), 体现了植物适应特定环境的构型 塑造. ...
杜鹃花属植物小枝大小对小枝生物量分配及叶面积支持效率的影响: 异速生长分析. 植物生态学报,
1
2008
... 在不同环境下, 植物种群的发育特性和物质分配策略是植物局部特化的形态特征对资源异质性的响应(Kleyer & Minden, 2015 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ; 史元春等, 2015 ), 在植物的整个生长过程中, 功能性状是植物与环境相互作用的结果, 如何调节小枝内各构件的生长及关系, 是植物生存策略和生境适合度调整的重要内容(肖遥等, 2014 ).在植物各器官中, 枝条和叶片是重要的输导和物质生产器官, 在生物力学及功能上具有密切联系.枝条具有导水功能和机械支撑作用(Alméras et al., 2004 ), 其大小往往关系到植物的空间拓展策略, 决定叶片的物质投资方式; 叶片大小和多少直接影响着植株的发育模式, 以及植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ), 枝、叶关系不仅是植物个体发育过程的表现(Brouat et al., 1998 ), 也体现了植物对特定生境的适应性(Violle et al., 2007 ).植物的形态特征和枝、叶功能之间的平衡受年龄和个体大小制约(Heuret et al., 2006 ; Barthélémy & Caraglio, 2007 ), 不同大小株丛在群落中占据的资源生态位存在差异(姚婧等, 2013 ), 其受群落郁闭度和干扰程度不同, 不同层次枝叶接受的有效太阳辐射也不同, 从而影响植物的空间分布格局及光照、温度等环境条件(程栋梁等, 2011 ), 往往导致不同个体大小植株分枝构型出现明显的分化.为了提高植物构型构件的生境适应性和自身的生存适合度, 不同大小等级植株可能需要适时调整其分枝角度, 增加植株所接受的光照强度, 促使植株枝-叶性状生长及其相互关系进行相应的调整(林勇明等, 2007 ), 使种群内每个植株可获得性资源的数量最大化, 构建与生境协同进化的冠层构型(de Kroons & Hutchings, 1995 ; 武高林等, 2010 ), 体现了植物适应特定环境的构型 塑造. ...
桂花植冠的枝系构型分析
1
2007
... 形态特征和生长特性的改变是植物适应不同环境和资源水平的重要策略(Maherali & Delucia, 2001 ), 植物功能性状与生存环境之间是一种相互协同进化与适应的关系(侯兆疆等, 2014 ).枝、叶作为植物的两个重要器官, 其关系不仅是不同构件单元大小数量关系权衡的结果, 也是适应不同光环境条件下能量交换效率的一个副产物.本研究中霸王不同大小等级植株枝叶性状关系存在差异, 在给定的枝长度下, 霸王小个体植株选择小的叶面积和多的叶数量, 大个体植株则选择了大的叶面积和少的叶数量, 霸王枝叶性状间的个体大小差异性较好地验证了植物通过功能性状的可塑性调整响应光环境变异的协同适应性机制, 反映了植物不同个体大小的冠层构型和环境适应性策略, 通过异速生长分析植物生长速率上的可塑性, 可以更好地了解植物在资源分配上的生境依赖规律, 从而进一步了解植物对环境的响应特征. ...
Influence of climate-driven shifts in biomass allocation on water transport and storage in ponderosa pine.
1
2001
... 植物的枝与叶性状之间始终存在着维持几何大小和重力平衡的需求, 二者间的大小匹配, 只有满足机械力学和水分动力学的要求, 才能保持枝叶机械支撑的平衡以及水分运输和蒸腾的协调(Westoby et al., 2002 ).I级霸王枝长度与叶面积表现出近等速生长关系, II级和III级霸王枝长度增加的速度大于叶面积增加的速度, 即大个体植株生长的大叶导致更高比例的枝长度投入, 霸王枝长度与叶面积间的关系具有个体大小差异性的特点.可能原因如下: 1)光照是影响植物生存和繁衍的关键性生态因子, 光照强度制约着植物的光合作用和有机物产量(Li et al., 2014 ).结合草地群落学调查、表1 和表2 资料认为, 研究区草地群落的高度和密度均较高, I级霸王位于群落中下层, 较II级和III级霸王接受的光合有效辐射降低(表2 ), 光照资源竞争能力弱, 而且群落中其他物种的邻体干扰使之遭受了较强烈的遮阴, 降低了植物光合作用能力.为了更多地捕获群落中下层的光斑, 在枝条分形方式上, I级霸王倾向于增大小枝与其支撑枝的分枝角度进行横向拓展(表1 ), 试图增加小枝空间分布范围和枝叶规模, 增强枝叶空间拓展能力和叶片光合作用, 因而I级霸王枝长度与叶面积近等速生长.II级霸王植株高度增加, 分枝角度减小, 将有限的小枝资源较多的用于机械支撑器官以增强其直立生长的能力, 捕获更多的光照资源, 能够降低邻体的遮挡和群落遮蔽程度; 相对而言III级霸王植株占据群落高度优势(表1 ), 株丛内部分枝数量多, 光照下透量减少, 叶片自遮阴严重, 阻止了小枝在冠内生长, III级霸王进一步降低分枝角度, 试图增加枝长度向高空发展以降低小枝在植冠内的遮挡, 因而II和III级霸王选择生长较长的枝条可能更有利.2) I级霸王小枝的枝横截面积小, 输水阻力大, 在不利环境下小枝更容易出现栓塞(Mcculloh & Sperry, 2005 ), 往往导致传输效率降低, 只能满足较小叶片的营养和水分传输需求, 同时较小的叶面积有助于植物组织规避水分亏缺的风险(Woodward et al., 2004 ); II级和III级霸王植株高度不断增大, 使木质部水分输送的距离增加, 增加枝横截面积(表1 ), 提高植株输水效率, 有助于增强植株向叶片运输水分能力, 因此, 荒漠草地中随着植物大小等级的增加, 植株可能增大枝横截面积以满足水分供给需求.3)研究区常受风力作用, 由风力引起的弯曲力矩与叶片面积和叶柄长度的立方成正比(Niklas, 1999 ), 小枝除了供给叶片水分和养分外, 还需提供支撑由于风等外界因素对叶片的拖曳的力, 这要求枝条需额外的资源配置, 叶片越大, 受到风等外力作用就越大, 枝条相应的投入也越高, 因而较大等级的霸王枝长度较长.值得注意的是, 随着大小等级的增加, 霸王枝长度逐渐增加, 叶面积也逐渐增大, 主要是由于自然选择的压力促使植物器官间沿最优化的方向进化(Enquist & Niklas, 2002 ), 大叶能够平铺排布, 有效地捕获来自垂直角度的光和大部分的光照(Horn, 1971 ), 为满足植株光合作用和水分运输的需求, 霸王植株叶面积与枝的输导面积相对应, 使得枝条所支撑的叶面积与枝条导管横截面积必须成比例. ...
Patterns in hydraulic architecture and their implications for transport efficiency.
1
2005
... 蒺藜科霸王属霸王(Zygophyllum xanthoxylum)是分布于我国西北部干旱荒漠区的强旱生小灌木, 当年生枝条数量多、叶量大、生长快, 具有抗旱、速生、高产、耐沙埋、耐贫瘠、抗风蚀的能力, 具有较好的饲用价值和适口性, 是荒漠灌丛植被的主要优势种和建群种(Chen et al., 2015 ).目前, 许多学者已对植物个体大小构型差异与竞争环境的关系(Aarssen, 2015 )、不同发育阶段分枝格局与叶片生长变化规律(许强等, 2013 )、不同个体大小茎-叶资源分配策略(李钰等, 2013b )等方面进行了大量研究, 解释了植物适应特定环境的生长特性与物质分配规律.霸王在不同环境下的表型特征差异也得到了关注(南江等, 2014 ), 有关霸王枝、叶性状关系的个体大小差异及其对不同光照强度生长特性的研究还不够深入, 影响了植物枝叶功能性状对不同生境适应机制的认识.因此, 本研究从植株个体大小的角度研究了荒漠草地霸王的枝叶构型特征及其光环境响应策略, 试图揭示: (1)不同大小等级霸王小枝内枝大小、叶面积和叶数量资源配置模式; (2)这种资源配置模式对霸王枝长度与叶大小、枝长度和叶数量间异速生长关系的影响. ...
霸王和木本猪毛菜在遮风和不遮风环境下的表型特征差异
1
2014
... 植物的形态可塑性是适应异质环境的一个重要特征, 使植物能够把资源最佳地分配到不同的器官, 以提高物种生存适合度(李钰等, 2013a ).研究发现: 不同大小等级霸王的枝长度与叶数量呈正的异速生长关系, 回归方程具有共同的SMA斜率且显著小于1 (p<0.05) (图2 ), 表明枝长度的增加速度大于叶片数的增加速度, 这符合“长的当年生小枝比短的当年生小枝具有更小的叶数量与枝长度的比值, 即单位枝长度的叶片数更少”的结论(Yagi, 2004 ).不同大小等级枝长度与叶数量y轴截距的变化则表明霸王投入的枝长度一定时, 大个体植株的叶数量较少, 这种差异性可能归因于以下3个方面: (1)霸王叶片对生或簇生, 均匀地排列在枝条上(周向睿等, 2006 ), 彼此之间相互作用、相互关联, 有利于小枝与叶片内部物质传输和转移, 使得这些小枝与其上着生叶片的生长具有密切的关系.结合表3 中叶面积的变化趋势, 推测可能是随着霸王个体大小等级的增加, 其枝条长度逐渐增大, 叶片数与叶面积均增加, 过多和过大的叶片都会造成上部叶片对下部叶片光照的遮挡, 使植株下部光合有效辐射降低, 产生低效率的光合作用, 同时随着分枝角度的逐渐减小, 枝条之间相互拥挤程度加剧, 造成叶片之间相互遮光严重, 因而较大等级霸王植株叶数量减小以提高植株内部光照资源的获取效率.(2)可能是叶片“大小-数量”的权衡对枝长度与叶片数关系产生影响.Kleiman和Aarssen (2007)以及杨冬梅等(2012)在小枝内部“大小-数量”权衡关系的研究中发现, 叶片“大小-数量”间呈等速负相关关系, 在单位枝条大小上着生的叶片面积越大数量越少, 反之亦然.随着个体大小等级的增大, 霸王增加叶面积的投资意味着植株将通过减少叶数量的方式来弥补大叶构建和扩展增加的成本, 试图使内部的自我荫蔽程度最小化(Niklas, 1999 ).因此, 霸王叶数量与枝长度在y轴截距的差异也可能是叶片大小变异影响的结果.(3)在进化过程中为提高光合收益, 植物应该增大获取限制性资源的器官分配, 而减少获取非限制资源的器官分配(Pickup et al., 2005 ; Niinemets et al., 2006 ).为了保证其生存和发展的光合产物需求, I级霸王生长大量小叶可以提高叶片配置的机动性来接收透射光, 增加植株的光合收益效率; II和III级霸王随着植株高度的增加, 接受的太阳辐射能较多, 若仍生长大量的大叶有可能导致呼吸和蒸腾成本更高, 不利于其光拦截能力和光合碳获取能力最大化(Wright et al., 2002 ), 因而较大等级的霸王选择生长少量的大叶可能更有利. ...
Leaf size modifies support biomass distribution among stems, petioles and midribs in temperate plants.
1
2006
... 光是植物生长的能量来源, 不同的光照强度对植物光合作用的影响主要是通过植物对光照的捕获利用能力来实现的(王艺和韦小丽, 2010 ).Valladares等(2002)指出, 叶内与茎内支撑投资之间可能存在某种权衡关系, 进而导致不同的冠层设计, 使得捕光效率在总体上是相似的.叶面积的增大需要更长的枝条来减少个体内部对光照的相互遮挡, 叶片面积和数量的增加则要求枝条具有更高的支撑能力(Niinemets et al., 2007 ), 大个体植株较小个体植株能生长更粗大的枝条用于支持功能, 体现了植物枝-叶生长中的物质和能量转换关系(Enquist & Niklas, 2001 , 2002 ).本研究发现, 随着霸王大小等级的增加, 枝长度与叶面积关系的异速斜率逐渐降低, 即枝长度的生长速度逐渐高于叶面积的生长速度; 枝长度一定的条件下, 相对于小个体霸王, 大个体霸王植株叶数量的增加速率降低, 自然选择使它们以最佳的收益/代价关系以满足不同个体大小植株的生长需求. ...
Leaf shape and venation pattern alter the support investments within leaf lamina in temperate species: A neglected source of leaf physiological differentiation?
2
2007
... 植物的枝与叶性状之间始终存在着维持几何大小和重力平衡的需求, 二者间的大小匹配, 只有满足机械力学和水分动力学的要求, 才能保持枝叶机械支撑的平衡以及水分运输和蒸腾的协调(Westoby et al., 2002 ).I级霸王枝长度与叶面积表现出近等速生长关系, II级和III级霸王枝长度增加的速度大于叶面积增加的速度, 即大个体植株生长的大叶导致更高比例的枝长度投入, 霸王枝长度与叶面积间的关系具有个体大小差异性的特点.可能原因如下: 1)光照是影响植物生存和繁衍的关键性生态因子, 光照强度制约着植物的光合作用和有机物产量(Li et al., 2014 ).结合草地群落学调查、表1 和表2 资料认为, 研究区草地群落的高度和密度均较高, I级霸王位于群落中下层, 较II级和III级霸王接受的光合有效辐射降低(表2 ), 光照资源竞争能力弱, 而且群落中其他物种的邻体干扰使之遭受了较强烈的遮阴, 降低了植物光合作用能力.为了更多地捕获群落中下层的光斑, 在枝条分形方式上, I级霸王倾向于增大小枝与其支撑枝的分枝角度进行横向拓展(表1 ), 试图增加小枝空间分布范围和枝叶规模, 增强枝叶空间拓展能力和叶片光合作用, 因而I级霸王枝长度与叶面积近等速生长.II级霸王植株高度增加, 分枝角度减小, 将有限的小枝资源较多的用于机械支撑器官以增强其直立生长的能力, 捕获更多的光照资源, 能够降低邻体的遮挡和群落遮蔽程度; 相对而言III级霸王植株占据群落高度优势(表1 ), 株丛内部分枝数量多, 光照下透量减少, 叶片自遮阴严重, 阻止了小枝在冠内生长, III级霸王进一步降低分枝角度, 试图增加枝长度向高空发展以降低小枝在植冠内的遮挡, 因而II和III级霸王选择生长较长的枝条可能更有利.2) I级霸王小枝的枝横截面积小, 输水阻力大, 在不利环境下小枝更容易出现栓塞(Mcculloh & Sperry, 2005 ), 往往导致传输效率降低, 只能满足较小叶片的营养和水分传输需求, 同时较小的叶面积有助于植物组织规避水分亏缺的风险(Woodward et al., 2004 ); II级和III级霸王植株高度不断增大, 使木质部水分输送的距离增加, 增加枝横截面积(表1 ), 提高植株输水效率, 有助于增强植株向叶片运输水分能力, 因此, 荒漠草地中随着植物大小等级的增加, 植株可能增大枝横截面积以满足水分供给需求.3)研究区常受风力作用, 由风力引起的弯曲力矩与叶片面积和叶柄长度的立方成正比(Niklas, 1999 ), 小枝除了供给叶片水分和养分外, 还需提供支撑由于风等外界因素对叶片的拖曳的力, 这要求枝条需额外的资源配置, 叶片越大, 受到风等外力作用就越大, 枝条相应的投入也越高, 因而较大等级的霸王枝长度较长.值得注意的是, 随着大小等级的增加, 霸王枝长度逐渐增加, 叶面积也逐渐增大, 主要是由于自然选择的压力促使植物器官间沿最优化的方向进化(Enquist & Niklas, 2002 ), 大叶能够平铺排布, 有效地捕获来自垂直角度的光和大部分的光照(Horn, 1971 ), 为满足植株光合作用和水分运输的需求, 霸王植株叶面积与枝的输导面积相对应, 使得枝条所支撑的叶面积与枝条导管横截面积必须成比例. ...
... 植物的形态可塑性是适应异质环境的一个重要特征, 使植物能够把资源最佳地分配到不同的器官, 以提高物种生存适合度(李钰等, 2013a ).研究发现: 不同大小等级霸王的枝长度与叶数量呈正的异速生长关系, 回归方程具有共同的SMA斜率且显著小于1 (p<0.05) (图2 ), 表明枝长度的增加速度大于叶片数的增加速度, 这符合“长的当年生小枝比短的当年生小枝具有更小的叶数量与枝长度的比值, 即单位枝长度的叶片数更少”的结论(Yagi, 2004 ).不同大小等级枝长度与叶数量y轴截距的变化则表明霸王投入的枝长度一定时, 大个体植株的叶数量较少, 这种差异性可能归因于以下3个方面: (1)霸王叶片对生或簇生, 均匀地排列在枝条上(周向睿等, 2006 ), 彼此之间相互作用、相互关联, 有利于小枝与叶片内部物质传输和转移, 使得这些小枝与其上着生叶片的生长具有密切的关系.结合表3 中叶面积的变化趋势, 推测可能是随着霸王个体大小等级的增加, 其枝条长度逐渐增大, 叶片数与叶面积均增加, 过多和过大的叶片都会造成上部叶片对下部叶片光照的遮挡, 使植株下部光合有效辐射降低, 产生低效率的光合作用, 同时随着分枝角度的逐渐减小, 枝条之间相互拥挤程度加剧, 造成叶片之间相互遮光严重, 因而较大等级霸王植株叶数量减小以提高植株内部光照资源的获取效率.(2)可能是叶片“大小-数量”的权衡对枝长度与叶片数关系产生影响.Kleiman和Aarssen (2007)以及杨冬梅等(2012)在小枝内部“大小-数量”权衡关系的研究中发现, 叶片“大小-数量”间呈等速负相关关系, 在单位枝条大小上着生的叶片面积越大数量越少, 反之亦然.随着个体大小等级的增大, 霸王增加叶面积的投资意味着植株将通过减少叶数量的方式来弥补大叶构建和扩展增加的成本, 试图使内部的自我荫蔽程度最小化(Niklas, 1999 ).因此, 霸王叶数量与枝长度在y轴截距的差异也可能是叶片大小变异影响的结果.(3)在进化过程中为提高光合收益, 植物应该增大获取限制性资源的器官分配, 而减少获取非限制资源的器官分配(Pickup et al., 2005 ; Niinemets et al., 2006 ).为了保证其生存和发展的光合产物需求, I级霸王生长大量小叶可以提高叶片配置的机动性来接收透射光, 增加植株的光合收益效率; II和III级霸王随着植株高度的增加, 接受的太阳辐射能较多, 若仍生长大量的大叶有可能导致呼吸和蒸腾成本更高, 不利于其光拦截能力和光合碳获取能力最大化(Wright et al., 2002 ), 因而较大等级的霸王选择生长少量的大叶可能更有利. ...
A mechanical perspective on foliage leaf form and function.
1
1999
... 在不同环境下, 植物种群的发育特性和物质分配策略是植物局部特化的形态特征对资源异质性的响应(Kleyer & Minden, 2015 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ; 史元春等, 2015 ), 在植物的整个生长过程中, 功能性状是植物与环境相互作用的结果, 如何调节小枝内各构件的生长及关系, 是植物生存策略和生境适合度调整的重要内容(肖遥等, 2014 ).在植物各器官中, 枝条和叶片是重要的输导和物质生产器官, 在生物力学及功能上具有密切联系.枝条具有导水功能和机械支撑作用(Alméras et al., 2004 ), 其大小往往关系到植物的空间拓展策略, 决定叶片的物质投资方式; 叶片大小和多少直接影响着植株的发育模式, 以及植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ), 枝、叶关系不仅是植物个体发育过程的表现(Brouat et al., 1998 ), 也体现了植物对特定生境的适应性(Violle et al., 2007 ).植物的形态特征和枝、叶功能之间的平衡受年龄和个体大小制约(Heuret et al., 2006 ; Barthélémy & Caraglio, 2007 ), 不同大小株丛在群落中占据的资源生态位存在差异(姚婧等, 2013 ), 其受群落郁闭度和干扰程度不同, 不同层次枝叶接受的有效太阳辐射也不同, 从而影响植物的空间分布格局及光照、温度等环境条件(程栋梁等, 2011 ), 往往导致不同个体大小植株分枝构型出现明显的分化.为了提高植物构型构件的生境适应性和自身的生存适合度, 不同大小等级植株可能需要适时调整其分枝角度, 增加植株所接受的光照强度, 促使植株枝-叶性状生长及其相互关系进行相应的调整(林勇明等, 2007 ), 使种群内每个植株可获得性资源的数量最大化, 构建与生境协同进化的冠层构型(de Kroons & Hutchings, 1995 ; 武高林等, 2010 ), 体现了植物适应特定环境的构型 塑造. ...
Optimal leaf size in relation to environment.
1
1972
... 植物的形态可塑性是适应异质环境的一个重要特征, 使植物能够把资源最佳地分配到不同的器官, 以提高物种生存适合度(李钰等, 2013a ).研究发现: 不同大小等级霸王的枝长度与叶数量呈正的异速生长关系, 回归方程具有共同的SMA斜率且显著小于1 (p<0.05) (图2 ), 表明枝长度的增加速度大于叶片数的增加速度, 这符合“长的当年生小枝比短的当年生小枝具有更小的叶数量与枝长度的比值, 即单位枝长度的叶片数更少”的结论(Yagi, 2004 ).不同大小等级枝长度与叶数量y轴截距的变化则表明霸王投入的枝长度一定时, 大个体植株的叶数量较少, 这种差异性可能归因于以下3个方面: (1)霸王叶片对生或簇生, 均匀地排列在枝条上(周向睿等, 2006 ), 彼此之间相互作用、相互关联, 有利于小枝与叶片内部物质传输和转移, 使得这些小枝与其上着生叶片的生长具有密切的关系.结合表3 中叶面积的变化趋势, 推测可能是随着霸王个体大小等级的增加, 其枝条长度逐渐增大, 叶片数与叶面积均增加, 过多和过大的叶片都会造成上部叶片对下部叶片光照的遮挡, 使植株下部光合有效辐射降低, 产生低效率的光合作用, 同时随着分枝角度的逐渐减小, 枝条之间相互拥挤程度加剧, 造成叶片之间相互遮光严重, 因而较大等级霸王植株叶数量减小以提高植株内部光照资源的获取效率.(2)可能是叶片“大小-数量”的权衡对枝长度与叶片数关系产生影响.Kleiman和Aarssen (2007)以及杨冬梅等(2012)在小枝内部“大小-数量”权衡关系的研究中发现, 叶片“大小-数量”间呈等速负相关关系, 在单位枝条大小上着生的叶片面积越大数量越少, 反之亦然.随着个体大小等级的增大, 霸王增加叶面积的投资意味着植株将通过减少叶数量的方式来弥补大叶构建和扩展增加的成本, 试图使内部的自我荫蔽程度最小化(Niklas, 1999 ).因此, 霸王叶数量与枝长度在y轴截距的差异也可能是叶片大小变异影响的结果.(3)在进化过程中为提高光合收益, 植物应该增大获取限制性资源的器官分配, 而减少获取非限制资源的器官分配(Pickup et al., 2005 ; Niinemets et al., 2006 ).为了保证其生存和发展的光合产物需求, I级霸王生长大量小叶可以提高叶片配置的机动性来接收透射光, 增加植株的光合收益效率; II和III级霸王随着植株高度的增加, 接受的太阳辐射能较多, 若仍生长大量的大叶有可能导致呼吸和蒸腾成本更高, 不利于其光拦截能力和光合碳获取能力最大化(Wright et al., 2002 ), 因而较大等级的霸王选择生长少量的大叶可能更有利. ...
Dry mass costs of deploying leaf area in relation to leaf size.
2005
A note on normal correlation.
1
1939
... 在不同环境下, 植物种群的发育特性和物质分配策略是植物局部特化的形态特征对资源异质性的响应(Kleyer & Minden, 2015 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ; 史元春等, 2015 ), 在植物的整个生长过程中, 功能性状是植物与环境相互作用的结果, 如何调节小枝内各构件的生长及关系, 是植物生存策略和生境适合度调整的重要内容(肖遥等, 2014 ).在植物各器官中, 枝条和叶片是重要的输导和物质生产器官, 在生物力学及功能上具有密切联系.枝条具有导水功能和机械支撑作用(Alméras et al., 2004 ), 其大小往往关系到植物的空间拓展策略, 决定叶片的物质投资方式; 叶片大小和多少直接影响着植株的发育模式, 以及植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ), 枝、叶关系不仅是植物个体发育过程的表现(Brouat et al., 1998 ), 也体现了植物对特定生境的适应性(Violle et al., 2007 ).植物的形态特征和枝、叶功能之间的平衡受年龄和个体大小制约(Heuret et al., 2006 ; Barthélémy & Caraglio, 2007 ), 不同大小株丛在群落中占据的资源生态位存在差异(姚婧等, 2013 ), 其受群落郁闭度和干扰程度不同, 不同层次枝叶接受的有效太阳辐射也不同, 从而影响植物的空间分布格局及光照、温度等环境条件(程栋梁等, 2011 ), 往往导致不同个体大小植株分枝构型出现明显的分化.为了提高植物构型构件的生境适应性和自身的生存适合度, 不同大小等级植株可能需要适时调整其分枝角度, 增加植株所接受的光照强度, 促使植株枝-叶性状生长及其相互关系进行相应的调整(林勇明等, 2007 ), 使种群内每个植株可获得性资源的数量最大化, 构建与生境协同进化的冠层构型(de Kroons & Hutchings, 1995 ; 武高林等, 2010 ), 体现了植物适应特定环境的构型 塑造. ...
Convergence in light capture efficiencies among tropical forest understory plants with contrasting crown architectures: A case of morphological compensation.
1
2002
... 在不同环境下, 植物种群的发育特性和物质分配策略是植物局部特化的形态特征对资源异质性的响应(Kleyer & Minden, 2015 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ; 史元春等, 2015 ), 在植物的整个生长过程中, 功能性状是植物与环境相互作用的结果, 如何调节小枝内各构件的生长及关系, 是植物生存策略和生境适合度调整的重要内容(肖遥等, 2014 ).在植物各器官中, 枝条和叶片是重要的输导和物质生产器官, 在生物力学及功能上具有密切联系.枝条具有导水功能和机械支撑作用(Alméras et al., 2004 ), 其大小往往关系到植物的空间拓展策略, 决定叶片的物质投资方式; 叶片大小和多少直接影响着植株的发育模式, 以及植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ), 枝、叶关系不仅是植物个体发育过程的表现(Brouat et al., 1998 ), 也体现了植物对特定生境的适应性(Violle et al., 2007 ).植物的形态特征和枝、叶功能之间的平衡受年龄和个体大小制约(Heuret et al., 2006 ; Barthélémy & Caraglio, 2007 ), 不同大小株丛在群落中占据的资源生态位存在差异(姚婧等, 2013 ), 其受群落郁闭度和干扰程度不同, 不同层次枝叶接受的有效太阳辐射也不同, 从而影响植物的空间分布格局及光照、温度等环境条件(程栋梁等, 2011 ), 往往导致不同个体大小植株分枝构型出现明显的分化.为了提高植物构型构件的生境适应性和自身的生存适合度, 不同大小等级植株可能需要适时调整其分枝角度, 增加植株所接受的光照强度, 促使植株枝-叶性状生长及其相互关系进行相应的调整(林勇明等, 2007 ), 使种群内每个植株可获得性资源的数量最大化, 构建与生境协同进化的冠层构型(de Kroons & Hutchings, 1995 ; 武高林等, 2010 ), 体现了植物适应特定环境的构型 塑造. ...
Let the concept of trait be functional!
1
2007
... 光是植物生长的能量来源, 不同的光照强度对植物光合作用的影响主要是通过植物对光照的捕获利用能力来实现的(王艺和韦小丽, 2010 ).Valladares等(2002)指出, 叶内与茎内支撑投资之间可能存在某种权衡关系, 进而导致不同的冠层设计, 使得捕光效率在总体上是相似的.叶面积的增大需要更长的枝条来减少个体内部对光照的相互遮挡, 叶片面积和数量的增加则要求枝条具有更高的支撑能力(Niinemets et al., 2007 ), 大个体植株较小个体植株能生长更粗大的枝条用于支持功能, 体现了植物枝-叶生长中的物质和能量转换关系(Enquist & Niklas, 2001 , 2002 ).本研究发现, 随着霸王大小等级的增加, 枝长度与叶面积关系的异速斜率逐渐降低, 即枝长度的生长速度逐渐高于叶面积的生长速度; 枝长度一定的条件下, 相对于小个体霸王, 大个体霸王植株叶数量的增加速率降低, 自然选择使它们以最佳的收益/代价关系以满足不同个体大小植株的生长需求. ...
不同光照对植物生长、生理生化和形态结构影响的研究进展
2010
Common slope tests for bivariateerrors-in-variables models.
1
2002
... 以每个大小等级30株霸王枝、叶性状及各层次光合有效辐射的平均值代表该个体大小霸王的枝、叶性状值和各层次光合有效辐射值, 然后对枝、叶性状的平均值进行对数(以10为底)转换, 使之符合正态分布后再进行分析.枝与叶片属性包括: 枝长度、叶面积、叶数量、枝横截面积.叶面积指当年生小枝上所支撑的所有叶片单叶面积的平均值.对枝叶功能性状关系的研究, 采用y = bxa , 线性转换成log(y) = log(b) + alog(x), 式中, x和y表示两个特征参数, b代表性状关系的截距, a表示相关性的斜率, 即异速生长参数或相对生长的指数, 当a = 1时, 表示两者是等速关系; 当a > 1时, 表示y的增加程度大于x的增加程度; 当a < 1时, 表示y的增加程度小于x的增加程度.数据分析主要采用标准化主轴估计(SMA)的方法(Warton et al., 2006 ), 由软件(S)MATR Version 2.0 (Falster et al., 2006 )计算完成.每一个回归斜率的置信区间根据Pitman (1939)方法计算, 通过Warton和Weber (2002)的方法判断回归斜率的异质性(heterogeneity), 并且在斜率同质(homogeneity)时计算共同斜率, 截距和斜率的差异利用方差分析检验.不同个体大小间功能特征平均值的比较采用单因素方差分析(ANOVA), 这些计算在SPSS 16.0下进行. ...
Bivariate line-fitting methods for allometry.
1
2006
... 植物的枝与叶性状之间始终存在着维持几何大小和重力平衡的需求, 二者间的大小匹配, 只有满足机械力学和水分动力学的要求, 才能保持枝叶机械支撑的平衡以及水分运输和蒸腾的协调(Westoby et al., 2002 ).I级霸王枝长度与叶面积表现出近等速生长关系, II级和III级霸王枝长度增加的速度大于叶面积增加的速度, 即大个体植株生长的大叶导致更高比例的枝长度投入, 霸王枝长度与叶面积间的关系具有个体大小差异性的特点.可能原因如下: 1)光照是影响植物生存和繁衍的关键性生态因子, 光照强度制约着植物的光合作用和有机物产量(Li et al., 2014 ).结合草地群落学调查、表1 和表2 资料认为, 研究区草地群落的高度和密度均较高, I级霸王位于群落中下层, 较II级和III级霸王接受的光合有效辐射降低(表2 ), 光照资源竞争能力弱, 而且群落中其他物种的邻体干扰使之遭受了较强烈的遮阴, 降低了植物光合作用能力.为了更多地捕获群落中下层的光斑, 在枝条分形方式上, I级霸王倾向于增大小枝与其支撑枝的分枝角度进行横向拓展(表1 ), 试图增加小枝空间分布范围和枝叶规模, 增强枝叶空间拓展能力和叶片光合作用, 因而I级霸王枝长度与叶面积近等速生长.II级霸王植株高度增加, 分枝角度减小, 将有限的小枝资源较多的用于机械支撑器官以增强其直立生长的能力, 捕获更多的光照资源, 能够降低邻体的遮挡和群落遮蔽程度; 相对而言III级霸王植株占据群落高度优势(表1 ), 株丛内部分枝数量多, 光照下透量减少, 叶片自遮阴严重, 阻止了小枝在冠内生长, III级霸王进一步降低分枝角度, 试图增加枝长度向高空发展以降低小枝在植冠内的遮挡, 因而II和III级霸王选择生长较长的枝条可能更有利.2) I级霸王小枝的枝横截面积小, 输水阻力大, 在不利环境下小枝更容易出现栓塞(Mcculloh & Sperry, 2005 ), 往往导致传输效率降低, 只能满足较小叶片的营养和水分传输需求, 同时较小的叶面积有助于植物组织规避水分亏缺的风险(Woodward et al., 2004 ); II级和III级霸王植株高度不断增大, 使木质部水分输送的距离增加, 增加枝横截面积(表1 ), 提高植株输水效率, 有助于增强植株向叶片运输水分能力, 因此, 荒漠草地中随着植物大小等级的增加, 植株可能增大枝横截面积以满足水分供给需求.3)研究区常受风力作用, 由风力引起的弯曲力矩与叶片面积和叶柄长度的立方成正比(Niklas, 1999 ), 小枝除了供给叶片水分和养分外, 还需提供支撑由于风等外界因素对叶片的拖曳的力, 这要求枝条需额外的资源配置, 叶片越大, 受到风等外力作用就越大, 枝条相应的投入也越高, 因而较大等级的霸王枝长度较长.值得注意的是, 随着大小等级的增加, 霸王枝长度逐渐增加, 叶面积也逐渐增大, 主要是由于自然选择的压力促使植物器官间沿最优化的方向进化(Enquist & Niklas, 2002 ), 大叶能够平铺排布, 有效地捕获来自垂直角度的光和大部分的光照(Horn, 1971 ), 为满足植株光合作用和水分运输的需求, 霸王植株叶面积与枝的输导面积相对应, 使得枝条所支撑的叶面积与枝条导管横截面积必须成比例. ...
Plant ecological strategies: Some leading dimensions of variation between species.
1
2002
... 植物的枝与叶性状之间始终存在着维持几何大小和重力平衡的需求, 二者间的大小匹配, 只有满足机械力学和水分动力学的要求, 才能保持枝叶机械支撑的平衡以及水分运输和蒸腾的协调(Westoby et al., 2002 ).I级霸王枝长度与叶面积表现出近等速生长关系, II级和III级霸王枝长度增加的速度大于叶面积增加的速度, 即大个体植株生长的大叶导致更高比例的枝长度投入, 霸王枝长度与叶面积间的关系具有个体大小差异性的特点.可能原因如下: 1)光照是影响植物生存和繁衍的关键性生态因子, 光照强度制约着植物的光合作用和有机物产量(Li et al., 2014 ).结合草地群落学调查、表1 和表2 资料认为, 研究区草地群落的高度和密度均较高, I级霸王位于群落中下层, 较II级和III级霸王接受的光合有效辐射降低(表2 ), 光照资源竞争能力弱, 而且群落中其他物种的邻体干扰使之遭受了较强烈的遮阴, 降低了植物光合作用能力.为了更多地捕获群落中下层的光斑, 在枝条分形方式上, I级霸王倾向于增大小枝与其支撑枝的分枝角度进行横向拓展(表1 ), 试图增加小枝空间分布范围和枝叶规模, 增强枝叶空间拓展能力和叶片光合作用, 因而I级霸王枝长度与叶面积近等速生长.II级霸王植株高度增加, 分枝角度减小, 将有限的小枝资源较多的用于机械支撑器官以增强其直立生长的能力, 捕获更多的光照资源, 能够降低邻体的遮挡和群落遮蔽程度; 相对而言III级霸王植株占据群落高度优势(表1 ), 株丛内部分枝数量多, 光照下透量减少, 叶片自遮阴严重, 阻止了小枝在冠内生长, III级霸王进一步降低分枝角度, 试图增加枝长度向高空发展以降低小枝在植冠内的遮挡, 因而II和III级霸王选择生长较长的枝条可能更有利.2) I级霸王小枝的枝横截面积小, 输水阻力大, 在不利环境下小枝更容易出现栓塞(Mcculloh & Sperry, 2005 ), 往往导致传输效率降低, 只能满足较小叶片的营养和水分传输需求, 同时较小的叶面积有助于植物组织规避水分亏缺的风险(Woodward et al., 2004 ); II级和III级霸王植株高度不断增大, 使木质部水分输送的距离增加, 增加枝横截面积(表1 ), 提高植株输水效率, 有助于增强植株向叶片运输水分能力, 因此, 荒漠草地中随着植物大小等级的增加, 植株可能增大枝横截面积以满足水分供给需求.3)研究区常受风力作用, 由风力引起的弯曲力矩与叶片面积和叶柄长度的立方成正比(Niklas, 1999 ), 小枝除了供给叶片水分和养分外, 还需提供支撑由于风等外界因素对叶片的拖曳的力, 这要求枝条需额外的资源配置, 叶片越大, 受到风等外力作用就越大, 枝条相应的投入也越高, 因而较大等级的霸王枝长度较长.值得注意的是, 随着大小等级的增加, 霸王枝长度逐渐增加, 叶面积也逐渐增大, 主要是由于自然选择的压力促使植物器官间沿最优化的方向进化(Enquist & Niklas, 2002 ), 大叶能够平铺排布, 有效地捕获来自垂直角度的光和大部分的光照(Horn, 1971 ), 为满足植株光合作用和水分运输的需求, 霸王植株叶面积与枝的输导面积相对应, 使得枝条所支撑的叶面积与枝条导管横截面积必须成比例. ...
Global climate and the distribution of plant biomes.
1
2004
... 植物的形态可塑性是适应异质环境的一个重要特征, 使植物能够把资源最佳地分配到不同的器官, 以提高物种生存适合度(李钰等, 2013a ).研究发现: 不同大小等级霸王的枝长度与叶数量呈正的异速生长关系, 回归方程具有共同的SMA斜率且显著小于1 (p<0.05) (图2 ), 表明枝长度的增加速度大于叶片数的增加速度, 这符合“长的当年生小枝比短的当年生小枝具有更小的叶数量与枝长度的比值, 即单位枝长度的叶片数更少”的结论(Yagi, 2004 ).不同大小等级枝长度与叶数量y轴截距的变化则表明霸王投入的枝长度一定时, 大个体植株的叶数量较少, 这种差异性可能归因于以下3个方面: (1)霸王叶片对生或簇生, 均匀地排列在枝条上(周向睿等, 2006 ), 彼此之间相互作用、相互关联, 有利于小枝与叶片内部物质传输和转移, 使得这些小枝与其上着生叶片的生长具有密切的关系.结合表3 中叶面积的变化趋势, 推测可能是随着霸王个体大小等级的增加, 其枝条长度逐渐增大, 叶片数与叶面积均增加, 过多和过大的叶片都会造成上部叶片对下部叶片光照的遮挡, 使植株下部光合有效辐射降低, 产生低效率的光合作用, 同时随着分枝角度的逐渐减小, 枝条之间相互拥挤程度加剧, 造成叶片之间相互遮光严重, 因而较大等级霸王植株叶数量减小以提高植株内部光照资源的获取效率.(2)可能是叶片“大小-数量”的权衡对枝长度与叶片数关系产生影响.Kleiman和Aarssen (2007)以及杨冬梅等(2012)在小枝内部“大小-数量”权衡关系的研究中发现, 叶片“大小-数量”间呈等速负相关关系, 在单位枝条大小上着生的叶片面积越大数量越少, 反之亦然.随着个体大小等级的增大, 霸王增加叶面积的投资意味着植株将通过减少叶数量的方式来弥补大叶构建和扩展增加的成本, 试图使内部的自我荫蔽程度最小化(Niklas, 1999 ).因此, 霸王叶数量与枝长度在y轴截距的差异也可能是叶片大小变异影响的结果.(3)在进化过程中为提高光合收益, 植物应该增大获取限制性资源的器官分配, 而减少获取非限制资源的器官分配(Pickup et al., 2005 ; Niinemets et al., 2006 ).为了保证其生存和发展的光合产物需求, I级霸王生长大量小叶可以提高叶片配置的机动性来接收透射光, 增加植株的光合收益效率; II和III级霸王随着植株高度的增加, 接受的太阳辐射能较多, 若仍生长大量的大叶有可能导致呼吸和蒸腾成本更高, 不利于其光拦截能力和光合碳获取能力最大化(Wright et al., 2002 ), 因而较大等级的霸王选择生长少量的大叶可能更有利. ...
Convergence towards higher leaf mass per area in dry and nutrient-poor habitats has different consequences for leaf life span.
1
2002
... 在不同环境下, 植物种群的发育特性和物质分配策略是植物局部特化的形态特征对资源异质性的响应(Kleyer & Minden, 2015 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ; 史元春等, 2015 ), 在植物的整个生长过程中, 功能性状是植物与环境相互作用的结果, 如何调节小枝内各构件的生长及关系, 是植物生存策略和生境适合度调整的重要内容(肖遥等, 2014 ).在植物各器官中, 枝条和叶片是重要的输导和物质生产器官, 在生物力学及功能上具有密切联系.枝条具有导水功能和机械支撑作用(Alméras et al., 2004 ), 其大小往往关系到植物的空间拓展策略, 决定叶片的物质投资方式; 叶片大小和多少直接影响着植株的发育模式, 以及植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ), 枝、叶关系不仅是植物个体发育过程的表现(Brouat et al., 1998 ), 也体现了植物对特定生境的适应性(Violle et al., 2007 ).植物的形态特征和枝、叶功能之间的平衡受年龄和个体大小制约(Heuret et al., 2006 ; Barthélémy & Caraglio, 2007 ), 不同大小株丛在群落中占据的资源生态位存在差异(姚婧等, 2013 ), 其受群落郁闭度和干扰程度不同, 不同层次枝叶接受的有效太阳辐射也不同, 从而影响植物的空间分布格局及光照、温度等环境条件(程栋梁等, 2011 ), 往往导致不同个体大小植株分枝构型出现明显的分化.为了提高植物构型构件的生境适应性和自身的生存适合度, 不同大小等级植株可能需要适时调整其分枝角度, 增加植株所接受的光照强度, 促使植株枝-叶性状生长及其相互关系进行相应的调整(林勇明等, 2007 ), 使种群内每个植株可获得性资源的数量最大化, 构建与生境协同进化的冠层构型(de Kroons & Hutchings, 1995 ; 武高林等, 2010 ), 体现了植物适应特定环境的构型 塑造. ...
三种高寒植物幼苗生物量分配及性状特征对光照和养分的响应. 生态学报,
1
2010
... 在不同环境下, 植物种群的发育特性和物质分配策略是植物局部特化的形态特征对资源异质性的响应(Kleyer & Minden, 2015 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ; 史元春等, 2015 ), 在植物的整个生长过程中, 功能性状是植物与环境相互作用的结果, 如何调节小枝内各构件的生长及关系, 是植物生存策略和生境适合度调整的重要内容(肖遥等, 2014 ).在植物各器官中, 枝条和叶片是重要的输导和物质生产器官, 在生物力学及功能上具有密切联系.枝条具有导水功能和机械支撑作用(Alméras et al., 2004 ), 其大小往往关系到植物的空间拓展策略, 决定叶片的物质投资方式; 叶片大小和多少直接影响着植株的发育模式, 以及植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ), 枝、叶关系不仅是植物个体发育过程的表现(Brouat et al., 1998 ), 也体现了植物对特定生境的适应性(Violle et al., 2007 ).植物的形态特征和枝、叶功能之间的平衡受年龄和个体大小制约(Heuret et al., 2006 ; Barthélémy & Caraglio, 2007 ), 不同大小株丛在群落中占据的资源生态位存在差异(姚婧等, 2013 ), 其受群落郁闭度和干扰程度不同, 不同层次枝叶接受的有效太阳辐射也不同, 从而影响植物的空间分布格局及光照、温度等环境条件(程栋梁等, 2011 ), 往往导致不同个体大小植株分枝构型出现明显的分化.为了提高植物构型构件的生境适应性和自身的生存适合度, 不同大小等级植株可能需要适时调整其分枝角度, 增加植株所接受的光照强度, 促使植株枝-叶性状生长及其相互关系进行相应的调整(林勇明等, 2007 ), 使种群内每个植株可获得性资源的数量最大化, 构建与生境协同进化的冠层构型(de Kroons & Hutchings, 1995 ; 武高林等, 2010 ), 体现了植物适应特定环境的构型 塑造. ...
古尔班通古特沙漠4种荒漠草本植物不同生长期的生物量分配与叶片化学计量特征
1
2014
... 蒺藜科霸王属霸王(Zygophyllum xanthoxylum)是分布于我国西北部干旱荒漠区的强旱生小灌木, 当年生枝条数量多、叶量大、生长快, 具有抗旱、速生、高产、耐沙埋、耐贫瘠、抗风蚀的能力, 具有较好的饲用价值和适口性, 是荒漠灌丛植被的主要优势种和建群种(Chen et al., 2015 ).目前, 许多学者已对植物个体大小构型差异与竞争环境的关系(Aarssen, 2015 )、不同发育阶段分枝格局与叶片生长变化规律(许强等, 2013 )、不同个体大小茎-叶资源分配策略(李钰等, 2013b )等方面进行了大量研究, 解释了植物适应特定环境的生长特性与物质分配规律.霸王在不同环境下的表型特征差异也得到了关注(南江等, 2014 ), 有关霸王枝、叶性状关系的个体大小差异及其对不同光照强度生长特性的研究还不够深入, 影响了植物枝叶功能性状对不同生境适应机制的认识.因此, 本研究从植株个体大小的角度研究了荒漠草地霸王的枝叶构型特征及其光环境响应策略, 试图揭示: (1)不同大小等级霸王小枝内枝大小、叶面积和叶数量资源配置模式; (2)这种资源配置模式对霸王枝长度与叶大小、枝长度和叶数量间异速生长关系的影响. ...
梭梭不同生长阶段的枝系构型特征
1
2013
... 植物的形态可塑性是适应异质环境的一个重要特征, 使植物能够把资源最佳地分配到不同的器官, 以提高物种生存适合度(李钰等, 2013a ).研究发现: 不同大小等级霸王的枝长度与叶数量呈正的异速生长关系, 回归方程具有共同的SMA斜率且显著小于1 (p<0.05) (图2 ), 表明枝长度的增加速度大于叶片数的增加速度, 这符合“长的当年生小枝比短的当年生小枝具有更小的叶数量与枝长度的比值, 即单位枝长度的叶片数更少”的结论(Yagi, 2004 ).不同大小等级枝长度与叶数量y轴截距的变化则表明霸王投入的枝长度一定时, 大个体植株的叶数量较少, 这种差异性可能归因于以下3个方面: (1)霸王叶片对生或簇生, 均匀地排列在枝条上(周向睿等, 2006 ), 彼此之间相互作用、相互关联, 有利于小枝与叶片内部物质传输和转移, 使得这些小枝与其上着生叶片的生长具有密切的关系.结合表3 中叶面积的变化趋势, 推测可能是随着霸王个体大小等级的增加, 其枝条长度逐渐增大, 叶片数与叶面积均增加, 过多和过大的叶片都会造成上部叶片对下部叶片光照的遮挡, 使植株下部光合有效辐射降低, 产生低效率的光合作用, 同时随着分枝角度的逐渐减小, 枝条之间相互拥挤程度加剧, 造成叶片之间相互遮光严重, 因而较大等级霸王植株叶数量减小以提高植株内部光照资源的获取效率.(2)可能是叶片“大小-数量”的权衡对枝长度与叶片数关系产生影响.Kleiman和Aarssen (2007)以及杨冬梅等(2012)在小枝内部“大小-数量”权衡关系的研究中发现, 叶片“大小-数量”间呈等速负相关关系, 在单位枝条大小上着生的叶片面积越大数量越少, 反之亦然.随着个体大小等级的增大, 霸王增加叶面积的投资意味着植株将通过减少叶数量的方式来弥补大叶构建和扩展增加的成本, 试图使内部的自我荫蔽程度最小化(Niklas, 1999 ).因此, 霸王叶数量与枝长度在y轴截距的差异也可能是叶片大小变异影响的结果.(3)在进化过程中为提高光合收益, 植物应该增大获取限制性资源的器官分配, 而减少获取非限制资源的器官分配(Pickup et al., 2005 ; Niinemets et al., 2006 ).为了保证其生存和发展的光合产物需求, I级霸王生长大量小叶可以提高叶片配置的机动性来接收透射光, 增加植株的光合收益效率; II和III级霸王随着植株高度的增加, 接受的太阳辐射能较多, 若仍生长大量的大叶有可能导致呼吸和蒸腾成本更高, 不利于其光拦截能力和光合碳获取能力最大化(Wright et al., 2002 ), 因而较大等级的霸王选择生长少量的大叶可能更有利. ...
Within-tree variations in shoot differentiation patterns of 10 tall tree species in a Japanese cool temperate forest.
2004
清凉峰不同海拔木本植物小枝内叶大小-数量权衡关系,
1
2012
... 植株的体积(植株个体地面高度×长冠幅×短冠幅)是反映其形体大小的重要指标(杨洪晓等, 2006 ).通过Excel 2007软件将植株个数设为纵坐标, 植株体积立方根设为横坐标, 生成一条正态曲线, 依据曲线走向将霸王划分为3个大小等级: I级(d ≤ 60 cm); II级(60 cm < d ≤ 120 cm); III级(d > 120 cm), 其中d为植株体积的立方根.经统计分析, 采集的117株霸王中有I级39株、II级44株、III级34株. ...
毛乌素沙地油蒿种群点格局分析
1
2006
... 在不同环境下, 植物种群的发育特性和物质分配策略是植物局部特化的形态特征对资源异质性的响应(Kleyer & Minden, 2015 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ; 史元春等, 2015 ), 在植物的整个生长过程中, 功能性状是植物与环境相互作用的结果, 如何调节小枝内各构件的生长及关系, 是植物生存策略和生境适合度调整的重要内容(肖遥等, 2014 ).在植物各器官中, 枝条和叶片是重要的输导和物质生产器官, 在生物力学及功能上具有密切联系.枝条具有导水功能和机械支撑作用(Alméras et al., 2004 ), 其大小往往关系到植物的空间拓展策略, 决定叶片的物质投资方式; 叶片大小和多少直接影响着植株的发育模式, 以及植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ), 枝、叶关系不仅是植物个体发育过程的表现(Brouat et al., 1998 ), 也体现了植物对特定生境的适应性(Violle et al., 2007 ).植物的形态特征和枝、叶功能之间的平衡受年龄和个体大小制约(Heuret et al., 2006 ; Barthélémy & Caraglio, 2007 ), 不同大小株丛在群落中占据的资源生态位存在差异(姚婧等, 2013 ), 其受群落郁闭度和干扰程度不同, 不同层次枝叶接受的有效太阳辐射也不同, 从而影响植物的空间分布格局及光照、温度等环境条件(程栋梁等, 2011 ), 往往导致不同个体大小植株分枝构型出现明显的分化.为了提高植物构型构件的生境适应性和自身的生存适合度, 不同大小等级植株可能需要适时调整其分枝角度, 增加植株所接受的光照强度, 促使植株枝-叶性状生长及其相互关系进行相应的调整(林勇明等, 2007 ), 使种群内每个植株可获得性资源的数量最大化, 构建与生境协同进化的冠层构型(de Kroons & Hutchings, 1995 ; 武高林等, 2010 ), 体现了植物适应特定环境的构型 塑造. ...
东灵山不同林型五角枫叶性状异速生长关系随发育阶段的变化
1
2013
... 植物的形态可塑性是适应异质环境的一个重要特征, 使植物能够把资源最佳地分配到不同的器官, 以提高物种生存适合度(李钰等, 2013a ).研究发现: 不同大小等级霸王的枝长度与叶数量呈正的异速生长关系, 回归方程具有共同的SMA斜率且显著小于1 (p<0.05) (图2 ), 表明枝长度的增加速度大于叶片数的增加速度, 这符合“长的当年生小枝比短的当年生小枝具有更小的叶数量与枝长度的比值, 即单位枝长度的叶片数更少”的结论(Yagi, 2004 ).不同大小等级枝长度与叶数量y轴截距的变化则表明霸王投入的枝长度一定时, 大个体植株的叶数量较少, 这种差异性可能归因于以下3个方面: (1)霸王叶片对生或簇生, 均匀地排列在枝条上(周向睿等, 2006 ), 彼此之间相互作用、相互关联, 有利于小枝与叶片内部物质传输和转移, 使得这些小枝与其上着生叶片的生长具有密切的关系.结合表3 中叶面积的变化趋势, 推测可能是随着霸王个体大小等级的增加, 其枝条长度逐渐增大, 叶片数与叶面积均增加, 过多和过大的叶片都会造成上部叶片对下部叶片光照的遮挡, 使植株下部光合有效辐射降低, 产生低效率的光合作用, 同时随着分枝角度的逐渐减小, 枝条之间相互拥挤程度加剧, 造成叶片之间相互遮光严重, 因而较大等级霸王植株叶数量减小以提高植株内部光照资源的获取效率.(2)可能是叶片“大小-数量”的权衡对枝长度与叶片数关系产生影响.Kleiman和Aarssen (2007)以及杨冬梅等(2012)在小枝内部“大小-数量”权衡关系的研究中发现, 叶片“大小-数量”间呈等速负相关关系, 在单位枝条大小上着生的叶片面积越大数量越少, 反之亦然.随着个体大小等级的增大, 霸王增加叶面积的投资意味着植株将通过减少叶数量的方式来弥补大叶构建和扩展增加的成本, 试图使内部的自我荫蔽程度最小化(Niklas, 1999 ).因此, 霸王叶数量与枝长度在y轴截距的差异也可能是叶片大小变异影响的结果.(3)在进化过程中为提高光合收益, 植物应该增大获取限制性资源的器官分配, 而减少获取非限制资源的器官分配(Pickup et al., 2005 ; Niinemets et al., 2006 ).为了保证其生存和发展的光合产物需求, I级霸王生长大量小叶可以提高叶片配置的机动性来接收透射光, 增加植株的光合收益效率; II和III级霸王随着植株高度的增加, 接受的太阳辐射能较多, 若仍生长大量的大叶有可能导致呼吸和蒸腾成本更高, 不利于其光拦截能力和光合碳获取能力最大化(Wright et al., 2002 ), 因而较大等级的霸王选择生长少量的大叶可能更有利. ...
 , 宋清华, 史元春, 王继伟, 陈静
, 宋清华, 史元春, 王继伟, 陈静


{kind=link}
{kind=link}
{kind=link}
{kind=link}