邱曦
Qiu Xi
通讯作者:
收稿日期: 2015-10-13
修回日期: 2016-01-18
网络出版日期: 2016-04-11
版权声明: 2016 植物生态学报编辑部 本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。
基金资助:
展开
摘要
研究侵蚀土壤有机质矿化及其温度敏感性(Q10)对深入认识水土流失地区土壤有机碳动态变化具有重要意义。该文以福建省长汀县河田镇严重侵蚀区的裸露红壤为研究对象, 通过测定不同培养温度(10 ℃、20 ℃和30 ℃)下的土壤有机碳矿化速率、培养过程中微生物生物量碳(MBC)和可溶性有机碳(DOC)含量的变化, 探讨了温度对严重侵蚀红壤有机碳矿化特征的影响及其Q10。结果表明: 温度对严重侵蚀红壤有机碳矿化具有显著影响, 温度越高土壤有机碳矿化速率和矿化率越高; 培养过程中土壤有机碳累积矿化量与MBC显著正相关, 与DOC极显著负相关, 说明微生物生物量和可利用碳含量显著影响土壤有机碳的矿化。尽管严重侵蚀红壤有机碳含量仅为1.54 g·kg-1, 但培养180天的土壤有机碳的累积矿化率高达22.2%-33.3%, 表明侵蚀红壤有机碳容易被矿化。严重侵蚀红壤在10-20 ℃时的Q10值为1.41, 20-30 ℃时Q10值下降到1.06, 土壤有机碳质量低是导致Q10值较低的重要原因, 而严重侵蚀区的红壤长期裸露使微生物对高温产生适应性是高温时Q10值接近1的重要原因。因此, 在未来气候变暖的趋势下, 恢复植被覆盖对减少严重侵蚀红壤有机碳矿化损失具有重要意义。
关键词:
Abstract
Keywords:
土壤碳库约占陆地生态系统碳(C)储量的67%, 是陆地生态系统中最大的碳库(约1500 Pg C) (Jenkinson et al., 1991; Hashimoto et al., 2004; Lal, 2004)。土壤碳矿化是陆地生态系统中碳循环的重要过程, 对维持土壤碳库动态平衡起着重要作用(刘绍辉和方精云, 1997)。温度作为土壤碳矿化的关键调控因子备受关注。根据联合国政府间气候变化专门委员会(IPCC)第五次报告, 从1880年到2012年, 地表和海平面的平均温度上升了0.85 ℃ (IPCC, 2013)。由于温室效应, 即使人们以后不再排放温室气体, 地表温度也将持续上升。因此, 温度升高如何影响土壤碳矿化已成为全球变化和碳循环研究的焦点问题, 目前国内外有关森林、草地和水田的土壤碳矿化及其温度敏感性的报道(艾丽等, 2007; 王清奎等, 2007; Chatterjee et al., 2008; 王若梦等, 2013) 非常多。
我国是全球水土流失最严重的国家, 水土流失总面积为484.74万km2, 占国土总面积的51.1% (李智广, 2009)。福建长汀县是我国南方花岗岩地区最典型的水土流失区, 从20世纪80年代起, 各级主管部门相继采取许多治理措施, 水土流失得到很好的控制, 大面积以马尾松(Pinus massoniana)为主的植被得到不同程度的恢复(谢锦升等, 2008)。然而, 仍存在大面积的林下裸露地, 造成严重的林下水土流失(何圣嘉等, 2011), 而且在阳光直射下土壤温度显著提高。因此, 研究温度如何影响裸露地土壤有机碳的矿化过程及其温度敏感性, 对深入了解严重侵蚀红壤有机碳动态过程及其如何响应气候变暖具有重要意义。
本研究以福建省西部汀江上游的长汀县河田镇未经治理的严重侵蚀红壤为研究对象, 测定了土壤在不同培养温度(10 ℃、20 ℃和30 ℃)下的有机碳矿化速率和温度敏感性(Q10), 并观测了不同培养阶段土壤的可溶性有机碳(DOC)和微生物生物量碳(MBC), 以及微生物代谢熵的变化特征, 旨在阐明温度对严重侵蚀红壤有机碳矿化的影响程度及其控制因子, 为揭示严重侵蚀红壤有机碳动态过程提供科学依据。
试验地位于福建省长汀县河田镇(25.55°- 25.80° N, 116.30°-116.52° E), 海拔300-500 m。该地区属中亚热带季风气候区, 年平均气温17.5-18.8 ℃, 年降水量1700 mm, 年蒸发量1403 mm。平均日照时间1924.6 h, 平均无霜期260天, ≥10 ℃积温4100-4650 ℃。地带性植被为常绿阔叶林, 但已基本被破坏殆尽, 现有植被主要以人工种植的马尾松和次生林为主, 树种结构单一, 生长较差。该研究区土壤为中粗粒花岗岩发育的红壤, 抗侵蚀能力弱, 属强度土壤侵蚀区, 土壤A层流失殆尽, B层出露, 立地条件极差。2003年土壤侵蚀遥感资料显示, 河田镇水土流失面积为13586.83 hm2, 占全镇总土地面积的46.68%。
2014年5月上旬, 选取立地条件基本一致的未治理马尾松“小老头林”地, 设置3个20 m × 20 m的标准样方进行调查。马尾松密度1225株·hm-2, 平均树高1.91 m, 平均胸径2.8 cm, 林下植被主要为芒萁(Dicranopteris pedata), 盖度仅10%左右。按“S”随机采集裸地表层0-10 cm土壤, 以12点组成一个土样, 重复3次。将采集的土样除去植物残体和石砾, 每个样地土样分别混匀分成两份, 一份土样过2 mm筛后置于4 ℃冰箱保鲜备用。另一份土样一部分用于测定土壤质量含水量, 另一部分风干后过0.149 mm筛, 用于测定土壤总有机碳、氮。采样的同时, 采用环刀法测定土壤饱和持水量(鲁如坤, 1999)。
根据测定的土壤含水量, 称取相当于50 g干土质量的鲜土装入500 mL特制的培养瓶中, 并调节土壤含水量至饱和持水量的60%, 之后用扎孔的保鲜膜封住(既可保持通风又可防止土壤水分损失过快), 在20 ℃恒温箱内预培养两周, 使土壤内部环境趋于稳定。预培养结束后, 将土壤样品分别置于10 ℃、20 ℃、30 ℃恒温培养箱中。每个温度梯度设置23个培养瓶, 其中5个用于长期测定土壤碳矿化速率, 18个用于定期破坏性取样。培养期间每5天采用称重法调节土壤含水量至60%饱和持水量, 同样用扎孔保鲜膜封住。在培养第7、14、21、30、60、90、120、150和180天抽气取样, 测定CO2浓度和计算矿化速率。每次测定时, 先用标准压缩空气通入培养瓶中洗气30 s, 使培养瓶内的气体稳定后用瓶盖密封, 用注射器从培养瓶橡胶口抽取10 mL气体, 注入密闭气袋, 然后将培养瓶放回培养箱, 2 h后采用同样的方法取气, 用气相色谱-质谱联用仪(GC-2014, Shimadzu, Kyoto, Japan)测定土壤培养第0、2 h的CO2浓度, 并计算出CO2排放速率和CO2累积排放量。在培养的第30、90、180天每次随机取出6个破坏性取样的土壤样品, 用于测定DOC和MBC等指标。
土壤有机碳、全氮含量用碳氮元素分析仪(Vario MAX, Elementar Analysensysteme GmbH, Hanau, Germany)测定。测定土壤pH值时, 风干土水比为1:2.5。土壤机械组成用土壤粒径分析系统(SEDIMAT 4-12, UGT, Berlin, Germany)测定。浸提得到的DOC及氯仿-熏蒸浸提法得到的MBC用总有机碳分析仪(TOC-VCPH, Shimadzu, Kyoto, Japan)测定。土壤微生物熵为微生物生物量碳含量与土壤有机碳的比值(用百分比表示)。代谢熵为呼吸速率与微生物生物量碳含量的比值(单位为h-1)。
1.4.1 土壤矿化速率计算
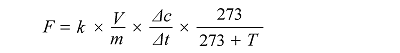
式中: F为气体排放速率(mg·kg-1·h-1); k是气体转化为标准单位的系数(kg·m-3); V为培养瓶内总气体体积(m3); m为干土质量(kg); Δc为CO2浓度差(μmol·mol-1; Δt为间隔时间(h); T为培养温度(℃)(李杰等, 2014)。
1.4.2 土壤碳矿化Q10值计算
本研究采用Conant等(2008)采用的Q10值计算方法, 计算公式为:
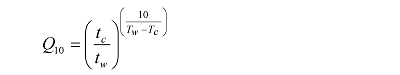
式中tc和tw分别是当土壤有机碳的矿化率相同时, 较低培养温度(Tc)和相对更高培养温度(Tw)持续的时间。用这种计算方法获得的Q10值避免了底物供给差异造成的低估(Luo et al., 2001)。
1.4.3 数据处理方法
采用方差分析方法比较不同温度培养下的矿化速率、累积矿化量、Q10、活性碳含量、微生物熵和代谢熵等指标的差异, 观测值如果出现高于平均值2倍标准差的数据, 在分析过程中将被剔除; 采用相关分析方法分析土壤有机碳累积矿化量与MBC、DOC含量的关系。所有数据均用Excel 2013进行处理, 用SPSS 19. 0进行差异显著性检验和相关分析。采用Origin 9. 0画图软件绘制图形。
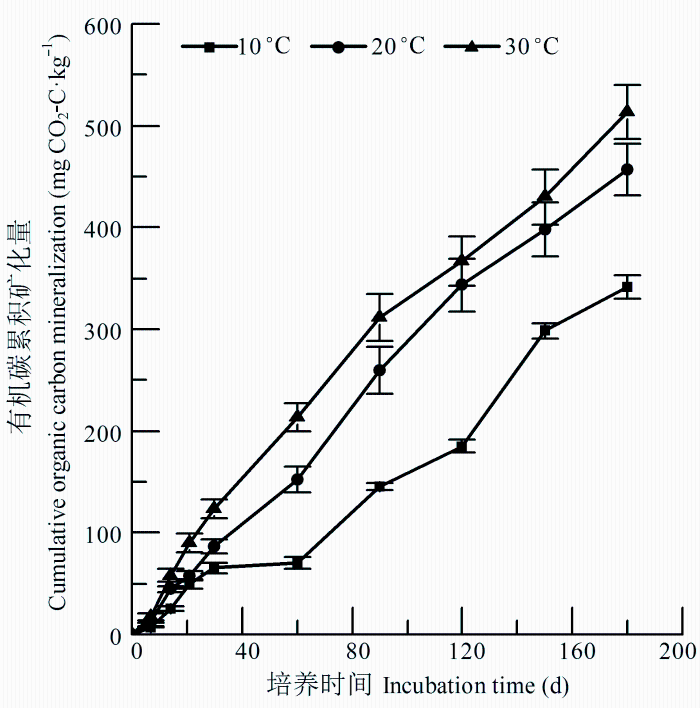
严重侵蚀红壤在不同温度(10、20、30 ℃)下, 培养前30天矿化速率较快, 之后矿化速率相对变慢(图1)。经过180天培养, 10、20和30 ℃有机碳累积矿化量的平均值分别为341.21、468.53、541.19 mg·kg-1, 矿化率分别为22.2%、29.7%和33.3%, 并且不同温度间都有显著差异(p < 0.05)。
图1 不同温度下严重侵蚀红壤有机碳累积矿化量(平均值±标准偏差)。
Fig. 1 Cumulative mineralization of soil organic carbon under different temperatures in severely eroded red soil (mean ± SD).
Q10是表达土壤有机碳矿化速率与温度之间关系的较好参数, 多数研究用Q10值反映有机碳矿化速率对温度的敏感性。严重侵蚀红壤碳矿化在10-20 ℃和20-30 ℃的Q10值分别为1.41 ± 0.06和1.06 ± 0.10, 前者显著高于后者(p < 0.05)。
3个培养温度下, 土壤MBC均随着培养时间增加而增加(图2A), 且培养30天后土壤MBC随温度增加呈减少趋势。培养180天后, 10、20和30 ℃培养的MBC平均值分别为19.21、17.57和16.65 mg·kg-1。在整个培养过程中, 不同温度下土壤DOC含量无显著性差异(p > 0.05), 但均随着培养时间延长呈减少趋势(p < 0.05, 图2B), 说明温度并没有显著改变土壤DOC含量, 培养结束时, 10、20和30 ℃土壤DOC的含量分别比初始时(3.05 mg·kg-1)降低了80.9%、79.9%和63.8%。
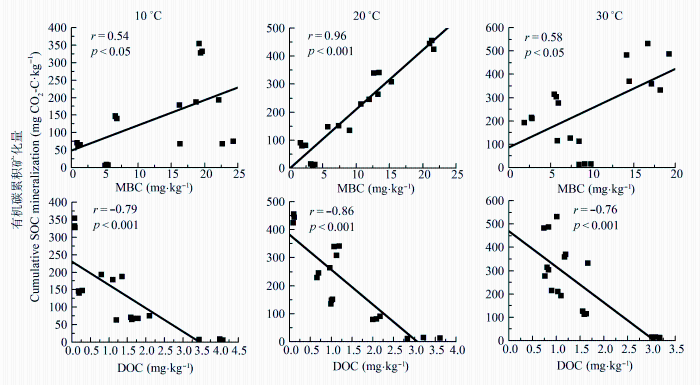
由图3可以看出, 土壤有机碳累积矿化量与MBC显著正相关(p < 0.05), 与DOC极显著负相关 (p < 0.001)。这表明严重侵蚀红壤的MBC、DOC含量能够显著影响土壤有机碳的矿化; 在培养过程中DOC作为碳源被微生物所利用而逐渐减少。
图3 严重侵蚀红壤在不同温度下有机碳累积矿化量与微生物生物量碳(MBC)、可溶性有机碳(DOC)的关系。
Fig. 3 The relationship cumulative soil organic carbon (SOC) mineralization and microbial biomass carbon (MBC) and dissolved organic carbon (DOC) in different incubation temperature of severely eroded red soil.
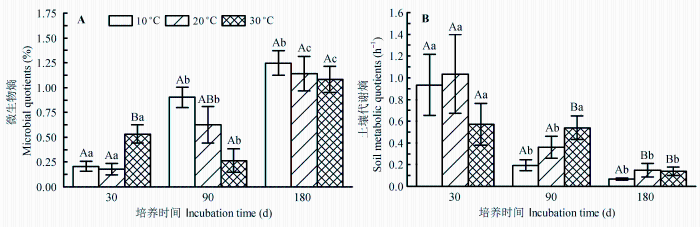
土壤微生物熵为MBC含量与土壤有机碳的比值, 反映土壤微生物对有机碳的利用效率。从图4A可见, 3个温度下微生物熵均随着培养时间显著升高(p < 0.05), 其中第180天时10、20和30 ℃的平均值分别为1.25%、1.04%和1.08%, 分别是第30天时的6.05倍、6.41倍和2.03倍。培养初期微生物熵小, 说明微生物对土壤有机碳的利用率低; 培养后期较初期显著升高, 说明随着培养的进行土壤微生物有效提高了对有机碳的利用率。但第180天时不同温度间并没有显著差异(p > 0.05), 这是由于后期MBC含量差异不显著导致的。
代谢熵为呼吸速率与MBC含量的比值, 代谢熵值小, 意味着微生物呼吸消耗碳的比例较小, 建造微生物细胞碳的比例相对较大, 对碳源的利用效率高。从代谢熵随时间的变化(图4B)可见, 10、20和30 ℃时, 代谢熵都随着培养的进行而显著降低(p < 0.05), 其中培养第180天时10、20和30 ℃下的代谢熵平均值分别为0.07、0.14和0.13 h-1, 分别比培养第30天时低92.7%、85.7%和75.8%; 进一步说明与培养前期相比, 培养后期微生物对土壤碳的利用率提高了。从温度间的差异可以看出, 培养第30天时不同温度间代谢熵没显著差异(p > 0.05), 但是培养第90天和180天时10 ℃下的代谢熵显著低于20 ℃和30 ℃下的代谢熵(p < 0.05)。
图4 不同温度培养过程中微生物熵和土壤代谢熵随时间的变化(平均值±标准偏差)。不同大写字母表示同一时间不同温度间差异显著; 不同小写字母表示同一温度不同时间差异显著(p < 0.05)。
Fig. 4 Change of microbial quotients and soil metabolic quotients in different incubation temperatures during incubation period (mean ± SD). Different capital letters indicate significant differences between temperature at the same time. Different lowercase letters indicate significant differences between time at the same temperature at p < 0.05.
土壤有机碳矿化是土壤微生物活动的结果, 而微生物的作用在很大程度上依赖于土壤中养分的供应。本研究中CO2产生速率呈现出培养前期快、后期慢的趋势, 与大多数研究结果(Balser & Wixon, 2009; 王若梦等, 2013; 李杰等, 2014)类似。在培养前期, 土壤中有较多的有机物质为微生物提供养分, 使微生物活动强烈, 土壤有机碳矿化速率很快达到最高峰, 累积矿化量大, 随着培养时间的延长, 培养后期土壤中主要为难分解的复杂的纤维素、木质素等有机碳, 微生物活动相对缓和(林杉等, 2014), 累积矿化量小。在培养过程中MBC和DOC与累积矿化量为显著线性关系(图3), 可溶性有机碳作为碳源被微生物所利用而逐渐减少(图2B), 这些过程说明土壤中活性有机碳是有机碳矿化的重要碳源。
温度是影响有机碳矿化的关键因素, 随着温度的变化, 其他因素的作用也会发生相应的改变。本研究中经过180天的培养, 10、20和30 ℃下的有机碳平均矿化率为22.2%、29.7%和33.3%, 并且温度间有显著性差异, 表明随着温度升高土壤有机碳矿化比例呈递增趋势, 这与大多数研究的结果(王若梦等, 2013; 李杰等, 2014; 林杉等, 2014)一致。然而, 本研究中侵蚀红壤的有机碳含量很低(表1), 而土壤碳矿化率与其他研究相比却高得多(表2), 这表明严重侵蚀红壤有机碳易被分解, 矿化潜力较高。土壤碳的数量和质量以及微生物对碳的利用效率是影响有机碳矿化的重要因子。本研究的土壤黏粒含量较低(表1)。有研究表明, 黏粒含量与土壤有机碳周转时间关系较密切, 黏粒含量低的土壤有机碳周转快, 有机质易被微生物降解(陈国潮等, 2002)。严重侵蚀红壤团聚体稳定性低(谢锦升等, 2008), 而团聚体保护性差的土壤有机碳的矿化率也相对较高(李忠佩等, 2004; 朱书法等, 2005)。本研究的微生物代谢熵为0.07-1.04 h-1, 明显高于其他研究(Grego & Kennedy, 1989; 陈璟和杨宁, 2013), 如陈璟和杨宁(2013)对衡阳紫色土丘陵坡地不同恢复阶段土壤的研究结果表明其微生物代谢熵为0.039-0.076 h-1, 这是严重侵蚀红壤矿化率高的重要原因。
表1 土壤基本理化性质(平均值±标准偏差)
Table 1 Physical and chemical characteristics of soil samples (mean ± SD)
| 实验地 Sample plot | 有机碳 Organic C (g·kg-1) | 全氮 Total N (g·kg-1) | 可溶性有机碳 Dissolved organic carbon (mg·kg-1) | 微生物生物量碳 Microbial biomass carbon (mg·kg-1) | 砂粒 Sand (%) | 粉粒 Silt (%) | 黏粒 Clay (%) | 酸碱度 pH value | 土壤含水量 Soil moisture (%) | 饱和持水量 Saturation moisture capacity (%) |
|---|---|---|---|---|---|---|---|---|---|---|
| 长汀侵蚀地Changting’s eroded land | 1.54 ± 0.06 | 0.35 ± 0.03 | 3.05 ± 0.22 | 19.59 ± 0.96 | 43.24 ± 3.00 | 37.85 ± 2.00 | 18.91 ± 3.00 | 4.74 ± 0.20 | 15.00 ± 0.25 | 43.00 ± 1.33 |
表2 不同研究区土壤有机碳矿化率的比较
Table 2 Mineralization rate respired in different study area
| 地点 Site | 土壤类型 Soil type | 土地利用类型 Land use type | 培养时间 Incubation time (d) | 有机碳含量 Organic C (g·kg-1) | 矿化率 Mineralization rate (%) | 文献来源 Literature resources |
|---|---|---|---|---|---|---|
| 福建长汀 Changting, Fujian | 红壤 Red soil | 侵蚀地 Eroded land | 7 | 1.54 | 0.80 (20 ℃) | 本研究 This study |
| 21 | 1.54 | 3.76 (20 ℃) | ||||
| 30 | 1.54 | 5.61 (20 ℃) | ||||
| 60 | 1.54 | 9.87 (20 ℃) | ||||
| 90 | 1.54 | 16.84 (20 ℃) | ||||
| 120 | 1.54 | 22.30 (20 ℃) | ||||
| 180 | 1.54 | 29.68 (20 ℃) | ||||
| 江苏宜兴 Yixing, Jiangsu | 棕红壤 Brown-red soil | 裸地 Bare land | 20 | 9.23 | 4.12 (28 ℃) | Li et al., 2007 |
| 杉木人工林 Cunninghamia lanceolata plantation | 20 | 9.49 | 2.49 (28 ℃) | |||
| 浙江 Zhejiang | 红壤 Red soil | 裸地 Bare land | 365 | 2.21 | 4.98 (25 ℃) | Xie & Zhang, 2014 |
| 裸地 Bare land | 365 | 4.36 | 2.98 (25 ℃) | |||
| 裸地 Bare land | 365 | 13.78 | 1.74 (25 ℃) | |||
| 林地 Forest land | 365 | 29.17 | 3.71 (25 ℃) | |||
| 林地 Forest land | 365 | 35.32 | 3.91 (25 ℃) | |||
| 福建泉头 Quantou, Fujian | 红壤 Red soil | 果园 Garden land | 85 | 13.21 | 3.91 (25 ℃) | Wang et al., 2012 |
| 湖南会同 Huitong, Hunan | 红壤 Red soil | 杉木人工林 Cunninghamia lanceolata plantation | 21 | 17.00 | 0.09 (9 ℃) | Wang et al., 2007 |
| 常绿阔叶林 Evergreen broad-leaved forest | 21 | 31.90 | 0.12 (9 ℃) | |||
| 福建三明 Sanming, Fujian | 红壤 Red soil | 杉木人工林 Cunninghamia lanceolata plantation | 28 | 19.23 | 0.92 (28 ℃) | Shen et al., 2012 |
| 江西吉泰 Jitai, Jiangxi | 红壤 Red soil | 马尾松林 Pinus massoniana forest | 45 | 9.57 | 1.14 (25 ℃) | Xue et al., 2014 |
| 福建武夷山 Mt. Wuyi, Fujian | 红壤 Red soil | 常绿阔叶林 Evergreen broad-leaved forest | 110 | 44.77 | Zhou et al., 2008 | |
| 江西千烟洲 Qianyanzhou, Jiangxi | 红壤 Red soil | 柑橘园 Citrus orchard | 7 | 11.50 | 0.05 (5 ℃) | Li et al., 2014 |
| 湿地马尾松人工林 Wetlands Pinus massoniana plantation | 7 | 10.12 | 0.06 (5 ℃) |
Q10表示温度变化对土壤有机碳矿化速率的影响。许多的土壤室内培养试验结果表明, 在土壤水分不受限制的情况下, 土壤碳矿化的温度敏感性随着温度的增加而下降。本研究中10-20 ℃和20-30 ℃下的Q10值分别为1.41和1.06, Q10值随温度增加而下降, 与前人的多数研究结果一致。Niklińska等(1999)沿欧洲大陆气候样带取7个欧洲赤松(Pinus sylvestris)林下土壤腐殖质样品, 在持水量(WHC)为50%、5-25 ℃下进行14周室内培养, 发现样带北部土壤在10-15 ℃时, 土壤呼吸的Q10超过5, 但在25 ℃附近时, Q10约为1。Leifeld和Fuhrer (2005)对采自农田和草地的土壤样品在不同温度下连续培养发现, 在15-25 ℃时, Q10值为5.2; 而在25-30 ℃时, Q10值为2.8。土壤微生物作为有机碳矿化的动力, 是决定Q10的内在因子, 同时对温度变化的响应十分敏感。本研究中, 随着培养时间的增加, 微生物熵逐渐升高, 表明微生物对有机碳的同化效率逐渐提高, 这可能是微生物群落中真菌逐渐增加而细菌逐渐减少的缘故。Zhang等(2005)的研究表明: 真菌比细菌的同化效率更高, 土壤中真菌数量增加, 而且温度升高也会提高真菌的生物量, 促进惰性有机碳组分的分解。土壤微生物对碳的利用效率的变化也能影响有机碳矿化(Reichstein et al., 2000)。本研究中, 培养90天后, 土壤微生物的代谢熵随着温度升高而增加, 这说明一定时间后有机碳矿化对温度的响应更敏感; 因为温度升高会抑制不适应高温的细菌生长, 而促进真菌生长, 提高真菌对惰性组分的分解利用, 这一结果亦证实了我们上述的推测, 也就是说, 培养初期细菌利用的土壤有机碳的活性组分(Zogg et al., 1997)随着培养的进行逐渐减少(图2B); 然而微生物生物量是增加的, 这可能是由真菌生物量的提高引起的。Zhang等(2005)对北美高草草原人工升温2 ℃土壤进行磷脂脂肪酸分析发现: 升温使真菌优势度增加, 细菌优势度降低, 微生物群落结构发生转变, 降低了土壤异养呼吸Q10。因此, 微生物群落组成和对碳利用效率的改变可能是Q10随温度升高而降低的主要原因。
周涛等(2009)的研究表明: 中国由北到南的黑棕壤、棕壤、褐土、黄壤、红壤、赤红壤和砖红壤的Q10值分别为1.61、1.54、1.63、2.1、1.87、2.21和1.78。而本研究的Q10值为1.41和1.06, 低于大多数研究结果(表3), 特别是20-30 ℃时的Q10值接近于1。呼吸底物有效性是决定Q10的重要因子(同小娟等, 2005), 研究表明呼吸底物质量越高, Q10值越高(Melillo et al., 2002; Fang et al., 2005)。Macdonald等(1999)对美国密歇根州硬木林灰土的培养研究发现: 底物质量较高的表层土壤呼吸的Q10高于底物质量较低的底层土壤呼吸的Q10。然而, 土壤微生物对温度的适应性对Q10具有重要影响, 本研究中的严重侵蚀红壤长期处于裸露状态, 温度变幅大, 如夏季地表温度高达40 ℃以上, 土壤微生物对高温的适应可能降低土壤碳矿化的温度敏感性。Balser和Wixon (2009)的研究表明土壤微生物群落的适应能力与土壤有机质的质量和呼吸底物有效性无关。Lange和Green (2005)的研究表明真菌的呼吸对土壤温度的季节性变化有明显的适应性, 如果土壤微生物对高温环境产生适应性, 那么在较高温度时会出现较低的Q10 (Luo et al., 2001)。因此, 土壤微生物的适应性也是严重侵蚀红壤Q10值在较高温度时显著低于较低温度时的一个原因。
表3 不同研究区土壤温度敏感性(Q10)的比较
Table 3 Temperature sensitivity (Q10) contrast of soil in different study area
| 气候类型 Climate type | 地点 Site | 土壤类型 Soil type | 土地利用类型 Land use type | 有机碳含量 Organic C (g·kg-1) | Q10 | 文献来源 Literature resources |
|---|---|---|---|---|---|---|
| 亚热带季风气候Subtropical monsoon climate | 福建长汀 Changting, Fujian | 红壤 Red soil | 侵蚀地 Eroded land | 1.54 | 1.06-1.41 | 本研究 This study |
| 亚热带季风气候 Subtropical monsoon climate | 福建泉头 Quantou, Fujian | 红壤 Red soil | 果园 Garden land | 13.21 | 1.49 | Wang et al., 2012 |
| 亚热带季风气候 Subtropical monsoon climate | 江西千烟洲 Qianyanzhou, Jiangxi | 红壤 Red soil | 柑橘园 Citrus orchard | 11.50 | 3.40 | Li et al., 2014 |
| 湿地马尾松人工林 Wetlands Pinus massoniana plantation | 10.12 | 2.20 | ||||
| 亚热带季风气候 Subtropical monsoon climate | 广东鼎湖 Dinghu, Guangdong | 赤红壤 Lateritic red soil | 针阔叶混交林 Mixed coniferous and broad-leaved forest | 26.80 | 2.03 | Deng et al., 2007 |
| 亚热带季风气候 Subtropical monsoon climate | 福建南平 Nanping, Fujian | 红壤 Red soil | 杉木人工林 Cunninghamia lanceolata plantation | 22.60 | 1.40 | Wei & Ma, 2006 |
| 马尾松人工林 Pinus massoniana plantation | 20.90 | 1.39 | ||||
| 亚热带季风气候 Subtropical monsoon climate | 四川盆地 Sichuan Basin | 红壤 Red soil | 林地 Forestland 裸地 Bare land | 19.26 11.98 | 1.25-3.25 1.54-3.68 | Wang et al., 2007 |
| 亚热带季风气候 Subtropical monsoon climate | 湖南 Hunan | 水稻土 Paddy soil | 水稻田 Paddy field | 19.20 | 2.11-2.78 | Ren et al., 2007 |
| 干燥亚湿润气候 Dry sub-humid climate | 内蒙古科尔沁沙地 Horqin sandy lands, Nei Mongol | 风沙土 Aeolian sandy soil | 裸地 Bare land | 3.36 | 1.55-1.97 | Wang et al., 2008 |
| 高原气候 Plateau climate | 青藏高原 Qinghai-Xizang Plateau | 草甸土 Meadow soil | 封育草地 Enclosure grassland 自由放牧草地 Grazing grassland | 19.04 | 1.46-1.99 | Xu et al., 2013 |
| 10.30 | 1.41-2.11 | |||||
| 温带季风气候Temperate monsoon climate | 吉林长白山 Changbai Mountain, Jilin | 草甸土 Meadow soil | 原生林 Original forest | 8.06 | 1.23-1.61 | Wang et al., 2013 |
| 次生林 Secondary forest | 4.03 | 1.23-1.61 |
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
| [1] |
The mineralization of alpine meadow soil organic carbon and factors influencing it in the Qilian Mountain. 祁连山中部高山草甸土壤有机碳矿化及其影响因素研究 . |
| [2] |
Investigating biological control over soil carbon temperature sensitivity. |
| [3] |
Timber harvesting alters soil carbon mineralization and microbial community structure in coniferous forests. |
| [4] |
Turnover of microbial biomass C in red soils and its significance in soil fertility evaluation. 红壤微生物生物量C周转及其研究 . |
| [5] |
Changes in SBR and qCO2 at different re-vegetation stages on sloping land with purple soils in Hengyang of Hunan Province, south-central China. 衡阳紫色土丘陵坡地不同恢复阶段土壤基础呼吸及代谢熵的变化 . |
| [6] |
Sensitivity of organic matter decomposition to warming varies with its quality.
|
| [7] |
Contributions of litter-fall to soil respiration and its affacting factors in southern subtropical forests of China. 南亚热带森林凋落物对土壤呼吸的贡献及其影响因素 .
|
| [8] |
Similar response of labile and resistant soil organic matter pools to changes in temperature. |
| [9] |
Effect of ammonitrate and stabilized farm yard manure on microbial biomass and metabolic quotient of soil under Zea mays. |
| [10] |
Soil respiration and soil CO2 concentration in a tropical forest, Thailand. |
| [11] |
Status, causes and prevention of soil and water loss in Pinus massoniana woodland in hilly red soil region of southern. 南方红壤丘陵区马尾松林下水土流失现状、成因及防治 . |
| [13] |
Model estimates of CO2 emissions from soil in response to global warming. |
| [14] |
Soil carbon sequestration impacts on global climate change and food security.
|
| [15] |
Lichens show that fungi can acclimate their respiration to seasonal changes in temperature.
|
| [16] |
The temperature response of CO2 production from bulk soils and soil fractions is related to soil organic matter quality. |
| [17] |
Impacts of land-use types on soil C mineralization and temperature sensitivity of forests in Qianyanzhou, Jiangxi Province, China. 土地利用类型对千烟洲森林土壤碳矿化及其温度敏感性的影响 . |
| [18] |
Current condition and dynamic changes of soil erosion in China. 中国水土流失现状与动态变化 .
|
| [19] |
Changes in transformation of soil organic carbon and functional diversity of soil microbial community under different land use patterns. 不同利用方式下土壤有机碳转化及微生物群落功能多样性变化 . |
| [20] |
Dynamics of soluble organic carbon and its relation to mineralization of soil organic carbon. 可溶性有机碳的含量动态及其与土壤有机碳矿化的关系 . |
| [21] |
Characteristics of soil organic carbon mineralization at different temperatures in paddy soils under long-term fertilization. 不同培养温度下长期施肥水稻土的有机碳矿化特征 . |
| [22] |
Effect factors of soil respiration and the temperature's effects on soil respiration in the global scale. 土壤呼吸的影响因素及全球尺度下温度的影响 .
|
| [23] |
|
| [24] |
Acclimatization of soil respiration to warming in a tall grass prairie.
|
| [25] |
Soil warming and carbon loss from a lake states spodosol. |
| [26] |
Soil warming and carbon-cycle feedbacks to the climate system.
|
| [27] |
Effect of temperature on humus respiration rate and nitrogen mineralization: Implications for global climate change. |
| [28] |
Temperature dependence of carbon mineralization: Conclusions from a long-term incubation of subalpine soil samples .
|
| [29] |
Effects of temperature on organic carbon mineralization in paddy soils with different clay content. 温度对不同黏粒含量稻田土壤有机碳矿化的影响 .
|
| [30] |
Effects of elevated nitrogen deposition on soil organic carbon mineralization and soil enzyme activities in a Chinese fir plantation. 氮沉降对杉木人工林土壤有机碳矿化和土壤酶活性的影响 . |
| [31] |
The responses of soil respiration and nitrogen mineralization to global warming in terrestrial ecosystems. 陆地生态系统土壤呼吸、氮矿化对气候变暖的响应 .
|
| [32] |
Impact of changes in vegetation types on soil C mineralization and associated temperature sensitivity in the Changbai Mountain forests of China. 植被类型变化对长白山森林土壤碳矿化及其温度敏感性的影响 .
|
| [33] |
Effects of temperature and green manure treatments on organic carbon mineralization in orchard soil. 不同温度及化肥绿肥施用比例对果园土壤有机碳矿化的影响 .
|
| [34] |
Effects of environmental factors on soil organic carbon mineralization in a Pinus sylvestris var. mongolica plantation. 不同环境因子对樟子松人工林土壤有机碳矿化的影响 . |
| [35] |
Soil carbon mineralization potential and its effect on soil active organic carbon in evergreen broad-leaved forest and Chinese fir plantation. 常绿阔叶林与杉木林的土壤碳矿化潜力及其对土壤活性有机碳的影响 . |
| [36] |
Effect of enclosure on soil C mineralization and priming effect in Stipa grandis grassland of Inner Mongolia. 围封对内蒙古大针茅草地土壤碳矿化及其激发效应的影响 .
|
| [37] |
Soil respiration and its sensitivity to temperature under different land use conditions. 不同土地利用方式下土壤呼吸及其温度敏感性 .
|
| [38] |
Dynamics of soil respiration in three major plantations in mid-subtropical zone. Journal of Fujian Agriculture and Forestry University (Natural Science Edition) ,中亚热带3种主要人工林的土壤呼吸动态 .
|
| [39] |
Influence of biochar application on mineralization and fractions of organic carbon in red soils with different land use. 施用生物质炭对红壤有机碳矿化及其组分的影响 . |
| [40] |
Effects of vegetation restoration on water stability and organic carbon distribution in aggregates of degraded red soil in subtropics of China. 植被恢复对退化红壤团聚体稳定性及碳分布的影响 .
|
| [41] |
Soil C mineralization and temperature sensitivity in alpine grasslands of the Qinghai-Xizang Plateau. 青藏高原高寒草地土壤碳矿化及其温度敏感性 .
|
| [42] |
Effect of land-use type on soil carbon mineralization and its priming effect on red soils in the mid-subtropics of China. 土地利用方式对中亚热带红壤碳矿化及其激发效应的影响 .
|
| [43] |
Soil microbial responses to experimental warming and clipping in a tall grass prairie.
|
| [44] |
The temperature sensitivity of soil respiration spatial pattern of inversion in China. 中国土壤呼吸温度敏感性空间格局的反演 .
|
| [45] |
Mineralization rates of soil organic carbon along an elevation gradient in Wuyi Mountain of Southeast China. 武夷山不同海拔高度土壤有机碳矿化速率的比较 . |
| [46] |
Use of δ13C method in studying soil organic matter. δ13C方法在土壤有机质研究中的应用 . |
| [47] |
Compositional and functional shifts in microbial communities due to soil warming.
|

/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}