Essential resources for plant growth are usually patchily distributed. During the process of propagation, interconnected ramet pairs of stoloniferous plant buffalograss (Buchloe dactyloides) may therefore experience contrasting resource supply such as light. Under heterogeneous light supply, anatomical structure of newly developed leaves is regulated by the light condition of mature leaves. However, little is known about whether leaf anatomical structure of clonal ramets is affected by the light environment of interconnected ramets in clonal plants under heterogeneous light supply.
Methods
Two light levels were set, with high light (natural sunlight) and low light (shade, 10% natural sunlight). Interconnected ramet pairs of buffalograss were exposed to homogeneous or heterogeneous light conditions.
Important findings Main vein diameter, bundle sheath cell number, leaf thickness and adaxial/abaxial mesophyll thickness of shaded ramets were remarkably decreased in spatially heterogeneous light environment; while no signifiacnt difference of these parameters was observed between ramets developed under homogeneous high light and low light conditions. Under heterogeneous light supply, adaxial/abaxial mesophyll thickness and abaxial stomatal size in unshaded elder daughter ramets (EDR) were remarkably increased, while adaxial/abaxial mesophyll thickness, stomatal density and size, leaf thickness, and bundle sheath cell number in unshaded younger daughter ramets (YDR) were reduced. Ramets under homogeneous high light conditions had higher stomatal density and larger stomata than those under homogeneous low light conditions. Adaxial/abaxial mesophyll thickness, stomatal density and size in buffalograss ramets were significantly affected by the light conditions of interconnected ramets.
Conclusions
Unshaded EDR benefit from their connection to shaded YDR, while unshaded YDR experience marked cost due to its connection to shaded EDR. The plastic decrease of shaded ramets under heterogeneous light may be associated with its reduced survival cost, and elevated survival rate under shading.
Keywords:buffalograss
;
light heterogeneity
;
anatomical structure
;
stomatal density
HAOChen-Song, WANGQing-Kai, SUNXiao-Ling. Effects of light heterogeneity on leaf anatomical structure in Buchloe dactyloides[J]. Chinese Journal of Plant Ecology, 2016, 40(3): 246-254 https://doi.org/10.17521/cjpe.2015.0377
Fig. 1 Diagrammatic presentation of a Buchloe dactyloides stolon segment consisting of mother ramet, elder daughter ramet and younger daughter ramet at the start of this experiment.
Table 1
表1
表1 姊妹分株接受的不同光处理
Table 1 The pattern of light treatments to elder daughter ramets (EDR) and younger daughter ramets (YDR)
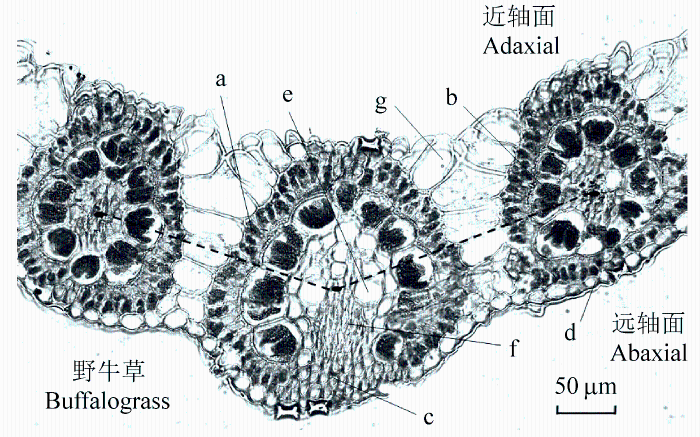
Fig. 2 Illustration of an image used to measure the adaxial (lower) and abaxial (upper) mesophyll tissues of Buchloe dactyloides in a cross-section light micrograph. The adaxial and abaxial mesophyll thickness were measured separately relative to the middle of the bundle sheath as shown by the dashed line, which generally represented the middle of the leaf (Jiang et al., 2011). a, mesophyll cells; b, bundle sheath cells; c, sclerenchyma; d, epidermal cell; e, xylem; f, phloem; g, motor cell.
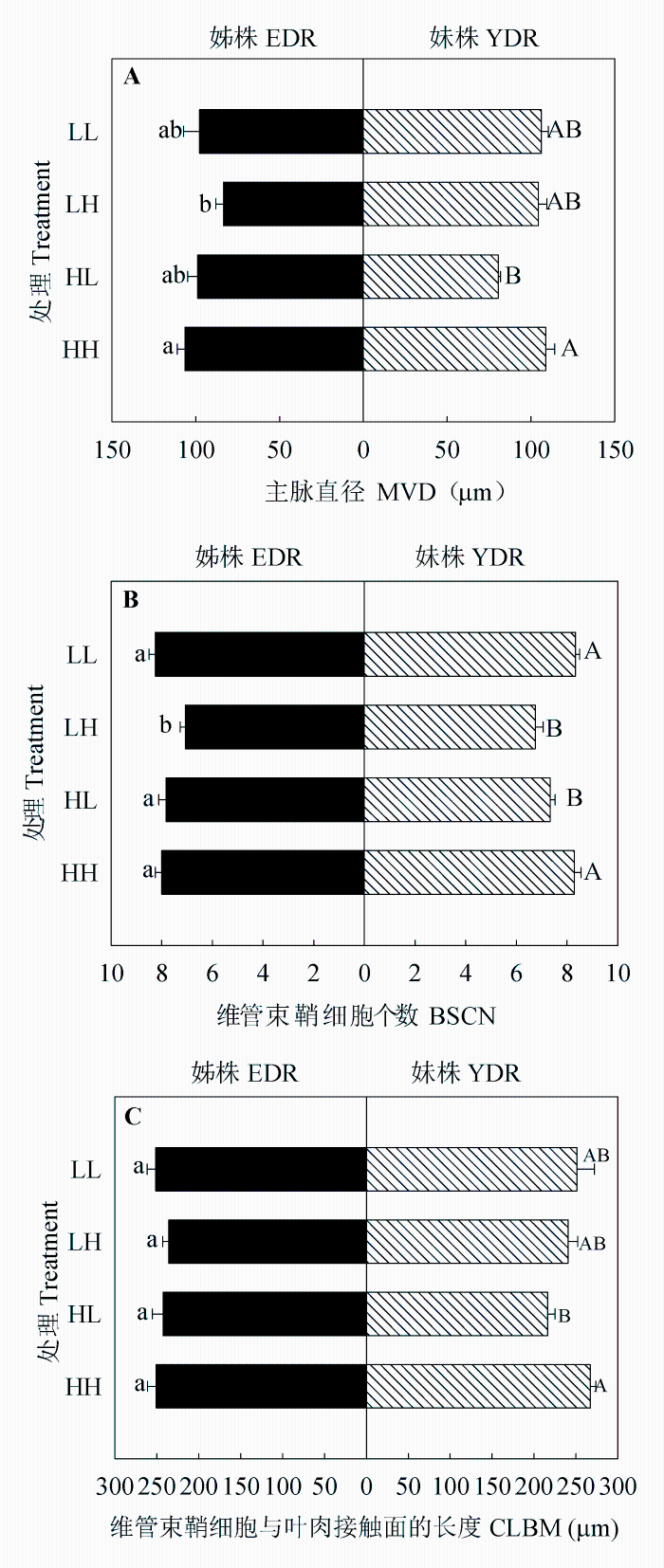
图3 同质和异质性光处理下野牛草姊株和妹株的主脉直径(A)、维管束鞘细胞个数(B)和维管束鞘细胞与叶肉接触面的长度(C) (平均值±标准误差, n = 7)。不同小写/大写字母表示处理间差异显著(p < 0.05)。HH、HL、LH、LL同表1。
Fig. 3 Main vein diameter (MVD) (A), bundle sheath cell number (BSCN) (B), and total contact length between bundle sheath and mesophyll cells (CLBM) (C) of Buchloe dactyloides elder daughter ramets (EDR) and younger daughter ramets (YDR) under homogeneous and heterogeneous light treatments (means ± SE, n = 7). Different letters indicate significant difference among treatments (p < 0.05). For HH, HL, LH and LL, see Table 1.
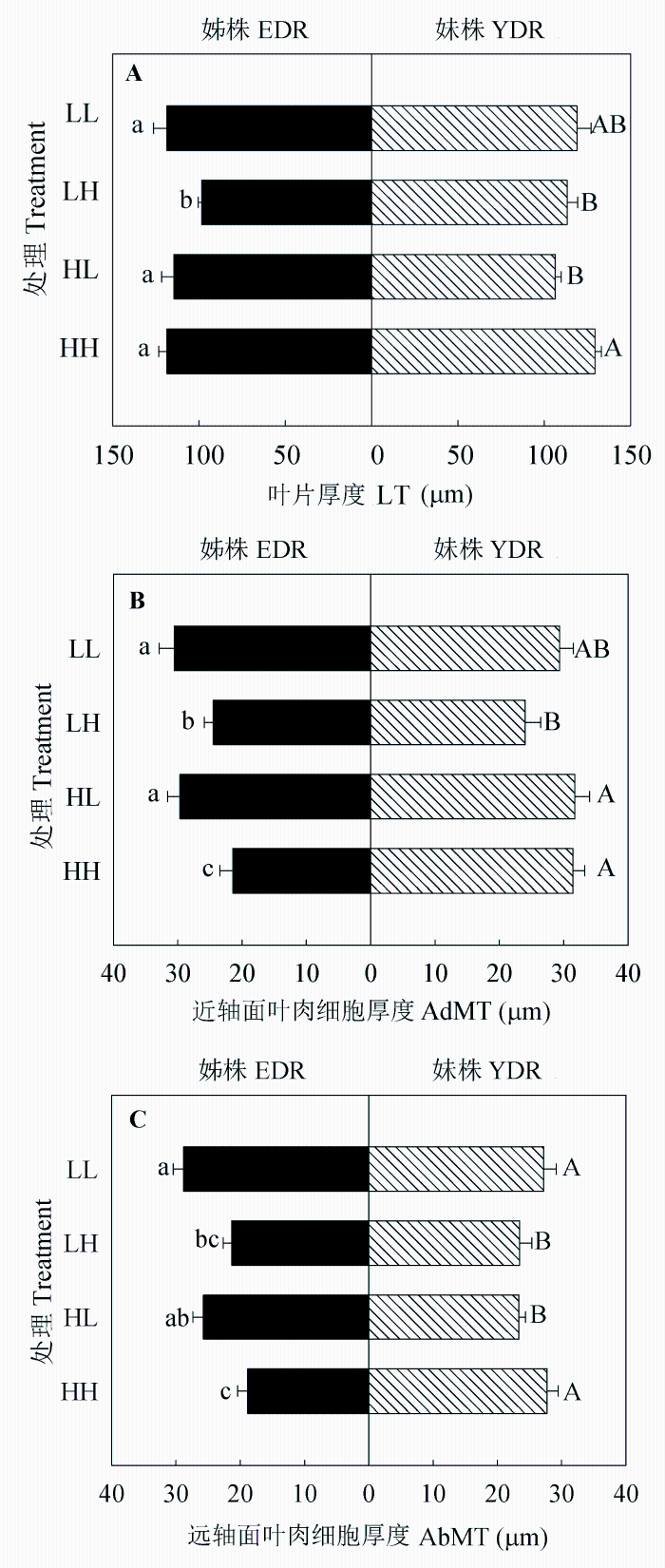
图4 同质和异质性光处理下野牛草姊株和妹株的叶片厚度(A)、近轴侧叶肉细胞的厚度(B)和远轴侧叶肉细胞的厚度(C) (平均值±标准误差, n = 7)。不同小写/大写字母表示处理间差异显著(p < 0.05)。HH、HL、LH、LL同表1。
Fig 4 Leaf thickness (LT) (A), adaxial mesophyll thickness (AdMT) (B), and abaxial mesophyll thickness (AbMT) (C) of Buchloe dactyloides elder daughter ramets (EDR) and younger daughter ramets (YDR) under homogeneous and heterogeneous light treatments (means ± SE, n = 7). Different letters indicate significant difference among treatments (p < 0.05). For HH, HL, LH and LL, see Table 1.
Table 2 F-values of three-way ANOVA for the effects of age (A), local condition (Lc), remote condition (Rc), and their interactions on leaf anatomical traits of Buchloe dactyloides
处理 Treatment
自由度 Degree of freedom
主脉直径 MVD
维管束鞘细胞个数 BSCN
维管束鞘细胞与叶肉接触面的长度 CLBM
叶片 厚度 LT
近轴侧叶肉细胞厚度 AdMT
远轴侧叶肉细胞厚度 AbMT
近轴侧 气孔密度 AdSD
远轴侧 气孔密度 AbSD
近轴侧 气孔大小 AdSZ
远轴侧 气孔大小 AbSZ
年龄 A
1, 48
0.754
0.388
0.017
1.168
4.19*
4.28*
0.368
0.116
33.925***
22.998***
分株自身所处光照 Lc
1, 48
10.667**
0.028
1.864
4.027
0.910
3.740
23.540***
31.030***
55.845***
85.684***
相连分株所处光照 Rc
1, 48
3.265
0.447
0.209
0.559
8.380**
17.550***
29.793***
35.030***
130.628***
98.520***
A × Lc
1, 48
0.018
2.495
0.988
0.008
0.110
1.630
0.828
0.485
37.874***
102.998***
Lc × Rc
1, 48
11.227**
28.096***
6.339*
10.221**
16.570***
20.700***
4.506*
1.486
2.041
0.112
A × Rc
1, 48
0.883
4.457*
0.001
1.380
12.780***
3.730
0.092
0.059
21.304***
81.604***
A × Lc × Rc
1, 48
0.285
2.592
1.299
0.079
1.390
3.490
0.368
0.021
0.592
66.281***
MVD, main vein diameter; BSCN, bundle sheath cell number; CLBM, total contact length between bundle sheath and mesophyll cells; LT, Leaf thickness; AdMT, adaxial mesophyll thickness; AbMT, abaxial mesophyll thickness; AdSD, adaxial stomatal density; AbSD, abaxial stomatal density; AdSZ, adaxial stomatal size; AbSZ, abaxial stomatal size. *, p < 0.05; **, p < 0.01; ***, p < 0.001; ns, p > 0.05.
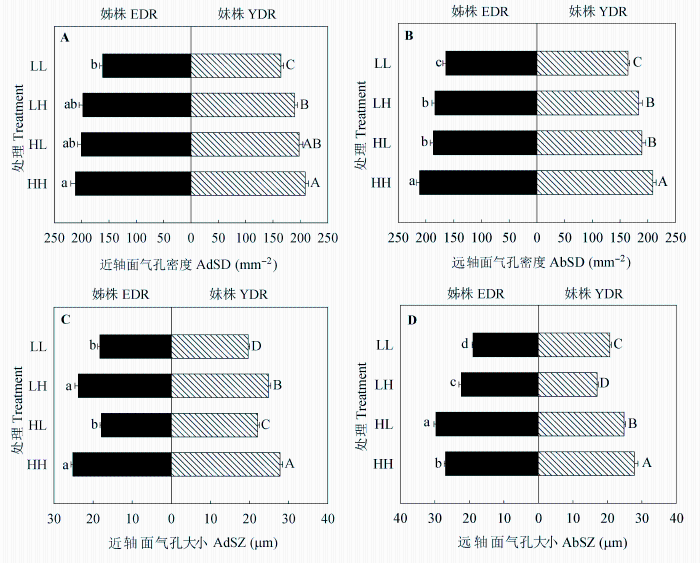
图5 同质和异质性光处理下野牛草姊株和妹株近轴侧和远轴侧的气孔密度(A和B)以及近轴侧和远轴侧的气孔大小(C和D) (平均值±标准误差, n = 7)。不同小写/大写字母表示处理间差异显著(p < 0.05)。HH、HL、LH、LL同表1。
Fig. 5 Adaxial stomatal density (AdSD) (A), abaxial stomatal density (AbSD) (B), adaxial stomatal size (AdSZ) (C) and abaxial stomatal size (AbSZ) (D) of Buchloe dactyloides elder daughter ramets (EDR) and younger daughter ramets (YDR) under homogeneous and heterogeneous light treatments (means ± SE, n = 7). Different letters indicate significant difference among treatments (p < 0.05). For HH, HL, LH and LL, see Table 1.
Acclimation of photosynthesis to irradiance and spectral quality in British plant species: Chlorophyll content, photosynthetic capacity and habitat preference.
Variations in the dorso- ventral organization of leaf structure and Kranz anatomy coordinate the control of photosynthesis and associated signalling at the whole leaf level in monocotyledonous species.
Clonal integration affects growth, photosynthetic efficiency and biomass allocation, but not the competitive ability, of the alien invasive Alternanthera philoxeroides under severe stress.
Clonal integration in Fragaria chiloensis differs between populations: Ramets from grassland are selfish.
1
1999
... 异质性环境条件是普遍存在的, 异质性光条件是具有发达匍匐茎的植物在蔓延过程中不可避免的环境限制因素.当克隆植物相互连接的克隆分株处于资源水平不同的小生境时, 相连分株之间可能会存在养分、水分或光合产物的克隆整合, 这种克隆整合通常被认为是植物适应自然条件的一种表现(Yu et al., 2004).目前对异质性光条件下克隆整合的研究, 绝大多数局限于对生物量(Stuefer et al., 1994; Alpert, 1999; Guo et al., 2011)、克隆形态(Magyar et al., 2007)的研究, 还有少数对光合参数(Wang et al., 2008; Li et al., 2011; 陶应时等, 2013)和激素含量(Alpert et al., 2002; Qian et al., 2010)的研究, 在异质性光条件下克隆植物叶片解剖结构的差异尚缺乏相关研究. ...
Hormonal modification of resource sharing in the clonal plant Fragaria chiloensis.
1
2002
... 异质性环境条件是普遍存在的, 异质性光条件是具有发达匍匐茎的植物在蔓延过程中不可避免的环境限制因素.当克隆植物相互连接的克隆分株处于资源水平不同的小生境时, 相连分株之间可能会存在养分、水分或光合产物的克隆整合, 这种克隆整合通常被认为是植物适应自然条件的一种表现(Yu et al., 2004).目前对异质性光条件下克隆整合的研究, 绝大多数局限于对生物量(Stuefer et al., 1994; Alpert, 1999; Guo et al., 2011)、克隆形态(Magyar et al., 2007)的研究, 还有少数对光合参数(Wang et al., 2008; Li et al., 2011; 陶应时等, 2013)和激素含量(Alpert et al., 2002; Qian et al., 2010)的研究, 在异质性光条件下克隆植物叶片解剖结构的差异尚缺乏相关研究. ...
The relative advantages of plasticity and fixity in different environments: When is it good for a plant to adjust?
Heterogeneous light supply affects growth and biomass allocation of the understory fern Diplopterygium glaucum at high patch contrast.
1
2011
... 异质性环境条件是普遍存在的, 异质性光条件是具有发达匍匐茎的植物在蔓延过程中不可避免的环境限制因素.当克隆植物相互连接的克隆分株处于资源水平不同的小生境时, 相连分株之间可能会存在养分、水分或光合产物的克隆整合, 这种克隆整合通常被认为是植物适应自然条件的一种表现(Yu et al., 2004).目前对异质性光条件下克隆整合的研究, 绝大多数局限于对生物量(Stuefer et al., 1994; Alpert, 1999; Guo et al., 2011)、克隆形态(Magyar et al., 2007)的研究, 还有少数对光合参数(Wang et al., 2008; Li et al., 2011; 陶应时等, 2013)和激素含量(Alpert et al., 2002; Qian et al., 2010)的研究, 在异质性光条件下克隆植物叶片解剖结构的差异尚缺乏相关研究. ...
Systemic signaling and acclimation in response to excess excitation energy in
1999
Influence of light availability on leaf structure and growth of two Eucalyptus globulus ssp. globulus provenances.
... Karpinski等(1999)发现将适应低光处理的拟南芥(Arabidopsis thaliana)部分重新置于高光处理下时, 没有经过高光处理的拟南芥叶片也会作出一个系统调整, 表现出对过多激发能和光氧化的响应.此后, 其他科学家陆续发现拟南芥、烟草(Nicotiana tabaccum)和高粱(Sorghum bicolor)新生叶的叶片厚度、近轴侧和远轴侧叶肉细胞厚度、近轴侧和远轴侧的气孔密度、维管束鞘细胞表面积和栅栏组织的解剖学特征均取决于成熟叶所处的光照条件, 而非自身所处的光照条件(Lake et al., 2001; Thomas et al., 2004; Jiang et al., 2011).换言之, 对于同一株植物来说, 由于遮阴的成熟叶与不遮阴的新生叶之间存在着信号联系, 致使未遮阴的新生叶表现出阴生植物的特点, 因此新生叶的叶片形态解剖结构受自身光照条件的影响不大. ...
... Illustration of an image used to measure the adaxial (lower) and abaxial (upper) mesophyll tissues of Buchloe dactyloides in a cross-section light micrograph. The adaxial and abaxial mesophyll thickness were measured separately relative to the middle of the bundle sheath as shown by the dashed line, which generally represented the middle of the leaf (Jiang et al., 2011). a, mesophyll cells; b, bundle sheath cells; c, sclerenchyma; d, epidermal cell; e, xylem; f, phloem; g, motor cell. ...
... 在异质光环境中, 未遮阴妹株近轴侧和远轴侧叶肉细胞的厚度、气孔密度和气孔大小、叶片厚度, 以及维管束鞘细胞个数均显著降低, 未遮阴妹株在异质性光处理下产生损耗的原因应该与胁迫信号的传导有关.Jiang等(2011)证明同为C4植物的高粱在成熟叶遮阴的条件下, 新生叶明显表现出阴生植物的特点, 如叶片上下表皮气孔密度变小, 叶片厚度和近轴侧叶肉细胞厚度减小, 维管束鞘细胞与叶肉的接触面积减少, 碳同化能力和气孔导度下降, 更易发生光抑制; 而对新生叶遮阴, 对成熟叶的这些指标则没有显著的影响.根据遮阴的成熟叶会给予未遮阴的新生叶胁迫信号的理论(Lake et al., 2001; Thomas et al., 2004; Jiang et al., 2011), 我们推断遮阴胁迫下的姊株(老株)应该是对妹株(幼株)发出了胁迫信号, 导致未遮阴的妹株做出了对遮阴胁迫的形态响应. ...
Effects of physiological integration on photosynthetic efficiency of Trifolium repens in response to heterogeneous UV-B radiation.
1
2011
... 异质性环境条件是普遍存在的, 异质性光条件是具有发达匍匐茎的植物在蔓延过程中不可避免的环境限制因素.当克隆植物相互连接的克隆分株处于资源水平不同的小生境时, 相连分株之间可能会存在养分、水分或光合产物的克隆整合, 这种克隆整合通常被认为是植物适应自然条件的一种表现(Yu et al., 2004).目前对异质性光条件下克隆整合的研究, 绝大多数局限于对生物量(Stuefer et al., 1994; Alpert, 1999; Guo et al., 2011)、克隆形态(Magyar et al., 2007)的研究, 还有少数对光合参数(Wang et al., 2008; Li et al., 2011; 陶应时等, 2013)和激素含量(Alpert et al., 2002; Qian et al., 2010)的研究, 在异质性光条件下克隆植物叶片解剖结构的差异尚缺乏相关研究. ...
Separating the contribution of the upper and lower mesophyll to photosynthesis in Zea mays L. leaves.
1989
Importance of plasticity and decision-making strategies for plant resource acquisition in spatio-temporally variable environments.
1
2007
... 异质性环境条件是普遍存在的, 异质性光条件是具有发达匍匐茎的植物在蔓延过程中不可避免的环境限制因素.当克隆植物相互连接的克隆分株处于资源水平不同的小生境时, 相连分株之间可能会存在养分、水分或光合产物的克隆整合, 这种克隆整合通常被认为是植物适应自然条件的一种表现(Yu et al., 2004).目前对异质性光条件下克隆整合的研究, 绝大多数局限于对生物量(Stuefer et al., 1994; Alpert, 1999; Guo et al., 2011)、克隆形态(Magyar et al., 2007)的研究, 还有少数对光合参数(Wang et al., 2008; Li et al., 2011; 陶应时等, 2013)和激素含量(Alpert et al., 2002; Qian et al., 2010)的研究, 在异质性光条件下克隆植物叶片解剖结构的差异尚缺乏相关研究. ...
Acclimation of photosynthesis to irradiance and spectral quality in British plant species: Chlorophyll content, photosynthetic capacity and habitat preference.
Inter-ramet photosynthate translocation in buffalograss under differential water defi- cit stress.
2
2010
... 异质性环境条件是普遍存在的, 异质性光条件是具有发达匍匐茎的植物在蔓延过程中不可避免的环境限制因素.当克隆植物相互连接的克隆分株处于资源水平不同的小生境时, 相连分株之间可能会存在养分、水分或光合产物的克隆整合, 这种克隆整合通常被认为是植物适应自然条件的一种表现(Yu et al., 2004).目前对异质性光条件下克隆整合的研究, 绝大多数局限于对生物量(Stuefer et al., 1994; Alpert, 1999; Guo et al., 2011)、克隆形态(Magyar et al., 2007)的研究, 还有少数对光合参数(Wang et al., 2008; Li et al., 2011; 陶应时等, 2013)和激素含量(Alpert et al., 2002; Qian et al., 2010)的研究, 在异质性光条件下克隆植物叶片解剖结构的差异尚缺乏相关研究. ...
Variations in the dorso- ventral organization of leaf structure and Kranz anatomy coordinate the control of photosynthesis and associated signalling at the whole leaf level in monocotyledonous species.
High benefits of clonal integration in two stoloniferous species, in response to heterogeneous light environments.
2
1994
... 异质性环境条件是普遍存在的, 异质性光条件是具有发达匍匐茎的植物在蔓延过程中不可避免的环境限制因素.当克隆植物相互连接的克隆分株处于资源水平不同的小生境时, 相连分株之间可能会存在养分、水分或光合产物的克隆整合, 这种克隆整合通常被认为是植物适应自然条件的一种表现(Yu et al., 2004).目前对异质性光条件下克隆整合的研究, 绝大多数局限于对生物量(Stuefer et al., 1994; Alpert, 1999; Guo et al., 2011)、克隆形态(Magyar et al., 2007)的研究, 还有少数对光合参数(Wang et al., 2008; Li et al., 2011; 陶应时等, 2013)和激素含量(Alpert et al., 2002; Qian et al., 2010)的研究, 在异质性光条件下克隆植物叶片解剖结构的差异尚缺乏相关研究. ...
... 异质性环境条件是普遍存在的, 异质性光条件是具有发达匍匐茎的植物在蔓延过程中不可避免的环境限制因素.当克隆植物相互连接的克隆分株处于资源水平不同的小生境时, 相连分株之间可能会存在养分、水分或光合产物的克隆整合, 这种克隆整合通常被认为是植物适应自然条件的一种表现(Yu et al., 2004).目前对异质性光条件下克隆整合的研究, 绝大多数局限于对生物量(Stuefer et al., 1994; Alpert, 1999; Guo et al., 2011)、克隆形态(Magyar et al., 2007)的研究, 还有少数对光合参数(Wang et al., 2008; Li et al., 2011; 陶应时等, 2013)和激素含量(Alpert et al., 2002; Qian et al., 2010)的研究, 在异质性光条件下克隆植物叶片解剖结构的差异尚缺乏相关研究. ...
Systemic irradiance signalling in tobacco.
2
2004
... Karpinski等(1999)发现将适应低光处理的拟南芥(Arabidopsis thaliana)部分重新置于高光处理下时, 没有经过高光处理的拟南芥叶片也会作出一个系统调整, 表现出对过多激发能和光氧化的响应.此后, 其他科学家陆续发现拟南芥、烟草(Nicotiana tabaccum)和高粱(Sorghum bicolor)新生叶的叶片厚度、近轴侧和远轴侧叶肉细胞厚度、近轴侧和远轴侧的气孔密度、维管束鞘细胞表面积和栅栏组织的解剖学特征均取决于成熟叶所处的光照条件, 而非自身所处的光照条件(Lake et al., 2001; Thomas et al., 2004; Jiang et al., 2011).换言之, 对于同一株植物来说, 由于遮阴的成熟叶与不遮阴的新生叶之间存在着信号联系, 致使未遮阴的新生叶表现出阴生植物的特点, 因此新生叶的叶片形态解剖结构受自身光照条件的影响不大. ...
... 在异质光环境中, 未遮阴妹株近轴侧和远轴侧叶肉细胞的厚度、气孔密度和气孔大小、叶片厚度, 以及维管束鞘细胞个数均显著降低, 未遮阴妹株在异质性光处理下产生损耗的原因应该与胁迫信号的传导有关.Jiang等(2011)证明同为C4植物的高粱在成熟叶遮阴的条件下, 新生叶明显表现出阴生植物的特点, 如叶片上下表皮气孔密度变小, 叶片厚度和近轴侧叶肉细胞厚度减小, 维管束鞘细胞与叶肉的接触面积减少, 碳同化能力和气孔导度下降, 更易发生光抑制; 而对新生叶遮阴, 对成熟叶的这些指标则没有显著的影响.根据遮阴的成熟叶会给予未遮阴的新生叶胁迫信号的理论(Lake et al., 2001; Thomas et al., 2004; Jiang et al., 2011), 我们推断遮阴胁迫下的姊株(老株)应该是对妹株(幼株)发出了胁迫信号, 导致未遮阴的妹株做出了对遮阴胁迫的形态响应. ...
Progress in the detection of costs of phenotypic plasticity in plants.
Clonal integration affects growth, photosynthetic efficiency and biomass allocation, but not the competitive ability, of the alien invasive Alternanthera philoxeroides under severe stress.
1
2008
... 异质性环境条件是普遍存在的, 异质性光条件是具有发达匍匐茎的植物在蔓延过程中不可避免的环境限制因素.当克隆植物相互连接的克隆分株处于资源水平不同的小生境时, 相连分株之间可能会存在养分、水分或光合产物的克隆整合, 这种克隆整合通常被认为是植物适应自然条件的一种表现(Yu et al., 2004).目前对异质性光条件下克隆整合的研究, 绝大多数局限于对生物量(Stuefer et al., 1994; Alpert, 1999; Guo et al., 2011)、克隆形态(Magyar et al., 2007)的研究, 还有少数对光合参数(Wang et al., 2008; Li et al., 2011; 陶应时等, 2013)和激素含量(Alpert et al., 2002; Qian et al., 2010)的研究, 在异质性光条件下克隆植物叶片解剖结构的差异尚缺乏相关研究. ...
Effect of light intensity and nutrient availability on clonal growth and clonal morphology of the stoloniferous herb
1
2003
... 所有试验处理的分株对被随机放置于温室内, 定期移位以减少位置因素造成的误差.异质性光处理之前, 保证所有分株形态和大小的一致性, 留3个叶片, 根系剪至3 cm长.实验过程中, 每个栽培槽每周浇30 mL的全能Hoagland营养液(4.373 g NH4 NO3·L-1, 2.063 g NaH2PO4·L-1和2.876 g KCl·L-1), 两个分株各浇15 mL, 保证植物对养分的基本需求(Yu & Dong, 2003).定期浇水, 保证植物不受到水分胁迫. ...
Clonal integration helps Psammochloa villosa survive sand burial in an inland dune.
1
2004
... 异质性环境条件是普遍存在的, 异质性光条件是具有发达匍匐茎的植物在蔓延过程中不可避免的环境限制因素.当克隆植物相互连接的克隆分株处于资源水平不同的小生境时, 相连分株之间可能会存在养分、水分或光合产物的克隆整合, 这种克隆整合通常被认为是植物适应自然条件的一种表现(Yu et al., 2004).目前对异质性光条件下克隆整合的研究, 绝大多数局限于对生物量(Stuefer et al., 1994; Alpert, 1999; Guo et al., 2011)、克隆形态(Magyar et al., 2007)的研究, 还有少数对光合参数(Wang et al., 2008; Li et al., 2011; 陶应时等, 2013)和激素含量(Alpert et al., 2002; Qian et al., 2010)的研究, 在异质性光条件下克隆植物叶片解剖结构的差异尚缺乏相关研究. ...




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}