北京林业大学森林资源与生态系统过程北京市重点实验室, 北京林业大学林学院, 北京 100083
版权声明: 2016 植物生态学报编辑部 本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。
基金资助:
展开
摘要
黑龙江黑河为我国红松(Pinus koraiensis)分布的北界, 在研究红松林的生长、演替、分布, 及其对气候变化的响应上有独特的意义。该文通过研究胜山保护区内阔叶红松林的演替系列(软阔叶林、硬阔叶林、阔叶红松近熟林和成熟林4个阶段), 分析了树木径向生长与气候变化的关系在不同演替阶段的差异。结果表明: 从演替早期的软阔叶林到晚期的红松成熟林, 年表统计特征表明树木径向生长对气候波动的敏感性逐步降低。对年轮-气候关系的分析结果也表明气候对树木径向生长的影响随着演替的进展呈现规律性的变化。上年6月和12月的气温与红松成熟林的径向生长显著正相关, 表现出明显的“滞后效应”。红松成熟林的轮宽指数与当年6月气温显著负相关, 而与当年6月降水量显著正相关, 反映出生长季水分对红松生长的限制。上述这些限制作用均随着演替的进展而增强, 但在演替的早期影响不显著。相反, 上年6月降水量与软阔叶林的生长显著负相关, 但该限制作用在演替的中晚期消失。这些差异反映出随着演替的进展, 优势树种对水分的需求逐步提高。滑动相关分析表明研究区近几十年明显的气候干暖化趋势对各林型的生长兼具有利和不利的影响。不同因素综合作用下, 软、硬阔叶林阶段生长尚未产生清晰的长期变化趋势。但由于水分对红松林生长的限制作用增强, 红松林生长明显下降。今后气候进一步干暖化可能对红松林的生长、恢复演替和分布有不利影响。
关键词:
Abstract
This research examined the different response of tree growth to climate change at the early, middle, late and matured successional stages of Korean pine (Pinus koraiensis) and broadleaf mixed forest.
This research used dendroecological methods to analyze radial growth at different successional stages of Korean pine forests in response to climatic change in the Shengshan Nature Reserve of Heilongjiang.
Chronology statistics indicated that the sensitivity of radial growth to inter-annual climate variability decreased from early to later successional stage. Meanwhile, the influence of some climate indices to radial growth also changed during the successional process. Radial growth of matured forest was positively related to mean monthly temperature in June and December of previous year, revealing an obvious “lag effect” of temperature. The ring-width of matured forest had a negative correlation with mean monthly temperature but a positive correlation with monthly precipitation in June of current year, reflecting the limiting role of growing season water availability. However, these limiting effects gradually disappeared towards the earlier stages of forest succession. On the contrary, the growth of early successional forest was negatively correlated to precipitation in June of previous year, and this effect disappeared towards the later successional stages. These differences revealed increased demand of water by dominant species from early to later successional stages. A moving correlation analysis showed that the increased warming and drying climate in the research area had both positive and negative influences on radial growth of each forest type. Earlier successional forests did not show clear long-term growth responses, but Korean pine forests had decreased radial growth in the past decades due to reduced water availability. Climate warming and drying in the future may impose negative impact on the growth, succession and distribution of Korean pine forests.
Keywords:
全球气候变暖会影响陆地生态系统的结构、功能和分布, 特别是在中高纬度地区(Hu et al., 2002)。IPCC (2007)报告指出, 高纬度地区气候变化的幅度比低纬度地区大。根据大气环流模型(GCMs)预测, 我国大陆平均气温增幅可达5-6 ℃, 东北地区作为我国纬度最高的森林分布区, 增幅可能更大(王冀等, 2008)。
气候变化对森林的一个直接影响是作用于树木的生长过程(Kramer et al., 2000), 使树木呈现出相应的径向生长变化(Goldblum & Rigg, 2005), 但径向生长对气候变化的响应因树种、环境因素和地域的差异而有明显不同(Leal et al., 2007)。气候变化的长期影响可能会改变物种的分布范围(Walther, 2003), 进而改变植物群落的分布和物种组成。树木年轮能够准确记载树木生长过程中经历的气候和环境的变化过程(Fritts, 1976), 因此成为研究树木生长、群落动态对气候变化响应的一种重要手段(Wang et al., 2006b; 常锦峰等, 2009)。
目前有关树轮气候学的研究, 由于针叶树种的早晚材区别明显, 轮印清晰更便于分析, 更多地选取针叶树种作为研究对象, 如通过云杉(Picea asperata)、落叶松(Larix gmelinii)、红松(Pinus koraiensis)等针叶树种来研究年轮与气候的关系(Leal et al., 2007; 陈力等, 2011; 曾令兵等, 2012); 也有少数研究用阔叶树进行年轮气候分析, 如在长白山林线附近研究气候因子对岳桦(Betula ermanii)的生长限制(Yu et al., 2007), 在东北南部研究蒙古栎(Quercus mongolica)生长对气候变化的响应(李腾等, 2014)等。在混交林中开展的工作则相对较少(Lebourgeois et al., 2013)。
红松针阔混交林是我国东北东部的地带性森林植被, 在东北亚森林中占有重要的地位(周以良, 1997)。但是, 目前阔叶红松林被严重破坏, 形成了大量处于不同演替阶段的次生林。研究不同演替阶段树木生长对气候变化的响应, 对于气候变化背景下促进阔叶红松林的恢复演替有着重要的意义。但是, 迄今为止还很少有人对阔叶红松林的不同演替阶段进行年轮-气候关系的比较研究。高露双等(2013)曾对长白山阔叶红松林中共存的红松和山杨(Populus davidiana)进行了比较研究, 但并没有包括阔叶红松林各演替阶段的不同林型。丘阳等(2014)在长白山的研究虽然包括了3个演替阶段, 但只针对红松种群的生产力进行了分析, 而没有对每个演替阶段的优势种进行比较。这些研究对于理解红松林恢复演替对气候变化的响应有重要意义, 直接针对不同演替阶段的比较研究无疑是必要的。
目前关于阔叶红松林及其次生林的树木年轮研究多集中在长白山地区, 仅有少量文献涉及其他区域, 这不利于全面地理解阔叶红松林的恢复演替与气候变化的关系。黑龙江黑河市的胜山保护区地处小兴安岭西北坡, 是温带、寒温带的过渡地带。这种生态交错区由于许多物种处于寒冷胁迫的临界状态, 生态系统对气候变化十分敏感(国庆喜等, 2001; Wang et al., 2006b)。同时, 该地区是我国红松分布的北界, 对于研究阔叶红松林的生长、分布、恢复演替对气候变化的响应都有着重要的意义。
本文以胜山阔叶红松林不同演替阶段的4种林型(软阔叶林(早期)、硬阔叶林(中期)、阔叶红松近熟林(中晚期)和红松成熟林(晚期))为研究对象, 通过研究树木径向生长与气候因子的关系, 探讨气候变化对不同演替阶段树木生长影响的差异。这对于预测未来气候变化背景下森林的演替以及红松林的分布格局具有重要的参考价值。
研究地点位于黑龙江省黑河市境内的胜山国家级自然保护区(49.42°-49.67° N, 126.42°-127.03° E)。保护区地处小兴安岭西北坡, 毗邻大兴安岭林区, 为大小兴安岭的交错过渡地带。该地区属于温带和寒温带气候的过渡区, 气候寒冷, 年平均气温-2 ℃, 最低气温-40 ℃, 最高气温36 ℃, 年降水量550-620 mm, 主要集中在夏季6-8月, 占全年降水量的65.7%。这里海拔不高, 一般为500 m左右, 没有明显的植被垂直分布。其植被呈现出大、小兴安岭植物区系交错、过渡的特点。代表林型是以红松为主的针阔混交林, 主要阔叶树包括紫椴(Tilia amurensis)、白桦(Betula platyphylla)、蒙古栎、裂叶榆(Ulmus laciniata)、色木槭(又称五角枫, Acer pictum subsp. mono)、大青杨(Populus ussuriensis)等, 同时还伴生一些欧亚大陆针叶林中的寒温性树种, 如红皮云杉(Picea koraiensis)、鱼鳞云杉(Picea jezoensis var. microsperma)、臭冷杉(Abies nephrolepis)等。研究区内地带性土壤为暗棕壤, 非地带性土壤有草甸土、沼泽土、泥炭土等(吴鹏, 2015)。
本研究在胜山自然保护区选择阔叶红松林演替序列上的4种林型, 即软阔叶林、硬阔叶林、阔叶红松近熟林和成熟林。其中软阔叶林的主要树种为白桦, 硬阔叶林的主要树种为蒙古栎, 红松近熟林和成熟林的主要树种包括红松、红皮云杉。为了避免局域环境差异的影响, 不同林型尽量选择立地条件相似的林分设置样方。样方坡向均为北坡, 海拔范围在460-630 m, 坡度在3°-18°之间, 立地条件基本相似。各林型基本情况见表1。每个林型设置1个样方, 样方面积50 m × 40 m, 由10 m × 10 m的样格组成。对样方中的乔木进行调查, 记录物种组成, 对每木测定胸径和树高。
在每个林型的样方内, 随机选择50株达到林冠层且健康生长的优势树种的个体。用生长锥在胸径处(距地面1.3 m)沿东北、西南方向各钻取一根树芯, 样本于2013年夏季采集。采集的年轮样本按照标准的处理程序进行干燥、固定和打磨(Stokes, 1996), 直至能在显微镜下清楚地分辨树木年轮的早材和晚材的分界线。然后通过LINTAB6年轮测量仪(Rintech, Heidelberg, Germany)和TSAP软件进行年轮宽度的测定, 量测的精度是0.001 mm。采用骨架图法、COFECHA程序对年轮序列进行交叉定年(Holmes, 1983), 对数据进行检验、修正错误并剔除异常序列及与主序列相关性差的样芯, 直至最终满足要求。最后利用ARSTAN程序对经过交叉定年的数据进一步处理(Cook, 1985), 采用负指数函数模拟树木的生长趋势, 以去除树木自身遗传因素产生的生长趋势和树木之间竞争导致的抑制和释放等生长趋势, 得到标准年表和差值年表, 年表中的轮宽指数为实际年轮宽度与去除生长趋势后的轮宽值的比值(Fritts, 1976)。经比较, 差值年表的统计特征明显好于标准年表, 其优点在于保留了更多的高频振荡信息, 因此最终采用差值年表进行树木生长和气候关系的分析。
胜山保护区地处偏远地区, 附近没有气象台站。因此本文气候资料采用CRU TS 3.22高分辨率的月平均气温和月降水量数据, 空间分辨率是0.5° × 0.5°。该数据在国内外树轮年代学研究中得到了广泛的应用(Liang et al., 2008)。闻新宇等(2006)指出CRU资料在区域尺度上与全国典型分区的气温变率一致, 降水量的变化与160个气象站点的观测吻合, 有较高的可信度。
表1 各演替阶段样方的林分特征
Table 1 Stand characteristics of the plots for different succession stages
| 演替阶段 Successional stage | 优势种 Dominant species | 胸径平均值 Mean DBH (cm) | 树高平均值 Mean tree height (m) | 最大胸径 Max DBH (cm) | 最大树高 Max tree height (m) | 林分密度 Stem density (tree·hm-2) |
|---|---|---|---|---|---|---|
| 早期 Early | 白桦 Betula platyphylla | 13.2 | 13.3 | 29.8 | 22.6 | 953 |
| 中期 Middle | 蒙古栎 Quercus mongolica | 11.1 | 8.9 | 43.8 | 24.3 | 1 576 |
| 中晚期 Mid-late | 红松 Pinus koraiensis, 红皮云杉 Picea koraiensis | 11.9 | 12.0 | 51.8 | 28.9 | 1 723 |
| 晚期 Late | 红松 Pinus koraiensis, 红皮云杉 Picea koraiensis | 16.6 | 13.1 | 67.7 | 28.6 | 1 180 |
气候数据的时间段为1960-2012年。考虑到上一年气候对当年树木生长可能会产生影响, 即产生“滞后效应” (李江风, 2000), 选取上年6月到当年9月平均气温和月降水量与轮宽指数进行相关性分析、滑动相关分析, 其中滑动分析选择的滑动区间为32年, 从1961年开始每32年计算一个相关系数, 然后逐年向后滑动, 分析了1961-1992年至1981-2012年的轮宽指数与气候因子的相关关系, 以研究树木径向生长对气候变化的响应。另外, 还分析了其他气候指标, 如各季节的平均气温和降水量等, 由于结果相似, 故不列出。数据分析采用DendroClim 2002程序完成。
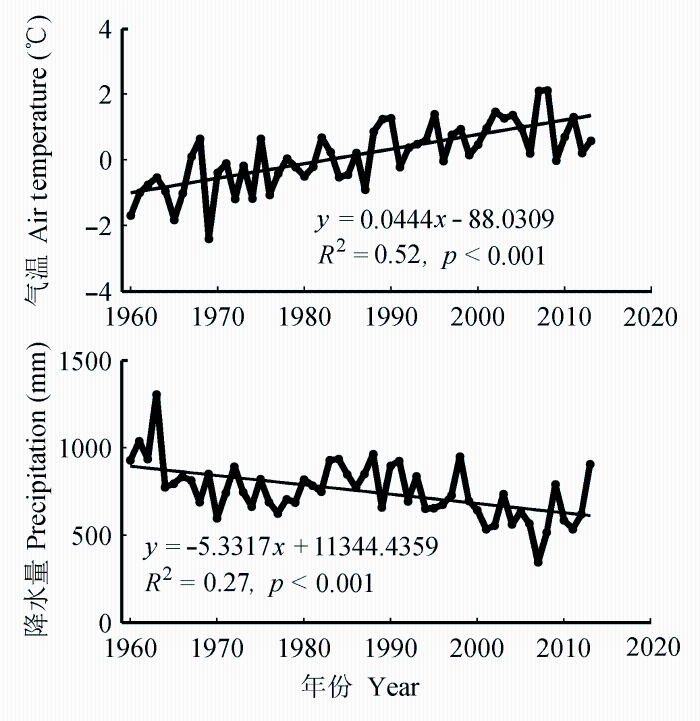
胜山地区的气候记录表明, 自1960年以来, 研究区的年平均气温以每年0.04 ℃ (R2 = 0.52)的速度递增, 年降水量以每年5.33 mm (R2 = 0.27)的速度递减, 即该地区的气候近几十年来表现出明显的干暖化趋势。
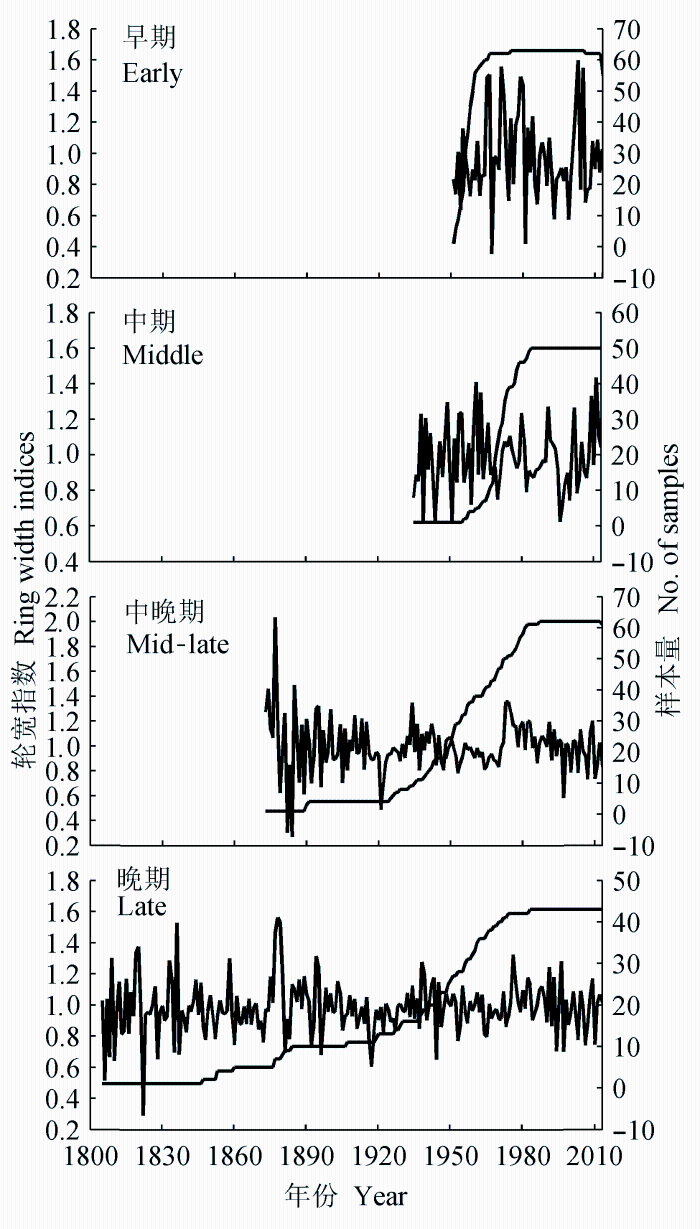
由胜山自然保护区各林型的差值年表(图2)可以看出, 阔叶红松近熟林和成熟林的轮宽指数的波动趋势较为一致, 但与阔叶林的波动并不一致。而软阔叶林和硬阔叶林的波动趋势也有明显的差异。各演替阶段年表之间的相关分析结果(表2)进一步表明, 演替阶段相差越远, 年表之间的相关性越低。这些结果说明径向生长对气候变化的响应在不同的演替阶段有明显的差异。在近几十年的共同区间内, 阔叶林年表的波动幅度明显大于红松林, 说明演替前期阔叶林阶段的树木生长对气候波动更为敏感。
表2 1960-2012年4个演替阶段差值年表间的相关系数
Table 2 Correlations among the residual chronologies for four successional stages during the period from 1960 to 2012
| 早期 Early | 中期 Middle | 中晚期 Mid-late | |
|---|---|---|---|
| 中期 Middle | 0.186 | ||
| 中晚期 Mid-late | 0.074 | 0.261 | |
| 晚期 Late | 0.048 | 0.242 | 0.569** |
年表的统计参数也证实了这一点。由表3可知: 从演替早期的软阔叶林到演替晚期的红松成熟林, 平均敏感度和标准偏差总体上下降, 说明树木生长对气候波动的敏感性降低; 第一主成分所占的方差量、信噪比以及样本的总体代表性也均随着演替的进展而呈现总体下降的趋势, 表明演替早期的林分生长受环境因子波动的限制作用更强。
总体代表性能够反映所建年表代表整个群体的程度。表3中4种林型的差值年表的总体代表性均超过了阈值0.85, 这说明年表的可信度较高, 适于进行年轮-气候关系的分析(Cook & Kairiukstis, 1990)。
表3 不同演替阶段的年表统计参数
Table 3 Summary statistics for residual chronology of different successional stages
| 演替阶段 Successional stage | 早期 Early | 中期 Middle | 中晚期 Mid-late | 晚期 Late |
|---|---|---|---|---|
| 共同区间 Common interval time span of year | 1957-2013 | 1969-2013 | 1951-2013 | 1949-2013 |
| 样本数(树芯/株数) Number of cores/trees | 63/48 | 50/41 | 63/48 | 43/33 |
| 平均敏感度 Mean sensitivity | 0.289 | 0.236 | 0.248 | 0.178 |
| 标准偏差 Standard deviation | 0.280 | 0.188 | 0.216 | 0.177 |
| 一阶自相关系数 Autocorrelation order 1 | 0.165 | -0.244 | -0.166 | 0.188 |
| R1样本间平均相关系数 Mean correlations among all radii | 0.473 | 0.251 | 0.341 | 0.299 |
| R2不同树木之间平均相关系数 Mean correlations between trees | 0.729 | 0.402 | 0.689 | 0.700 |
| R3同一树木不同样本之间平均相关系数 Mean correlations between trees and within trees | 0.470 | 0.250 | 0.336 | 0.293 |
| 信噪比 Signal-to-noise ratio | 26.043 | 6.380 | 13.459 | 10.682 |
| 样本总体代表性 Express population signal | 0.963 | 0.864 | 0.931 | 0.914 |
| 第一主成分所占方差量 PCA1 (%) | 0.501 | 0.308 | 0.377 | 0.347 |
不同林型年表的轮宽指数与不同月份气候指标的相关关系见表4, 可以看出树木生长与气候因子的关系在不同的演替阶段存在明显的差异。
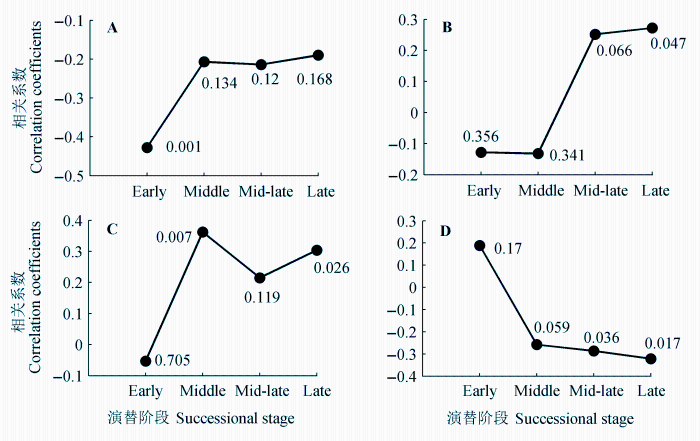
气候对树木生长的影响表现出明显的“滞后效应” (李江风, 2000), 但上年气候对生长的影响随着演替阶段的不同而有明显的差异。比如, 上年6月的气温与红松成熟林的轮宽指数显著正相关, 但与前几个演替阶段径向生长的相关性均不显著(表4)。上年6月降水量与软阔叶林的轮宽指数显著负相关(r = -0.43), 但这种关系在后几个演替阶段较弱(r > -0.22)且不显著(图3A)。
图1 研究区年平均气温和年降水量的变化。
Fig. 1 Changes in mean annual temperature and precipitation in the study area.
上年12月的气温与红松成熟林的轮宽指数显著正相关(p < 0.05), 但与其他几个林型的相关关系并不显著, 且相关系数从演替晚期的0.27逐步下降到演替早期的-0.13 (表4; 图3B), 这也表明了气候对径向生长的影响随着演替的进展有规律性的变化。
图2 不同演替阶段的轮宽指数(差值年表)和样本量。
Fig. 2 The residual chronology and number of samples for different successional stages.
从当年气候来看, 当年6月的气温和红松近熟林、成熟林的轮宽指数显著负相关, 而在两个阔叶林型中相关关系不显著。同样, 这种相关性也表现出随着演替进展的规律性变化: 相关系数从演替早期的0.19逐步下降到演替晚期的-0.32 (图3D)。这实际上反映了红松林相对于演替早期树种对生长季降水的较高需求。这一点从当年6月降水量和轮宽指数的关系也可得到印证(图3C)。该相关关系在硬阔叶林到红松成熟林的演替阶段较高(r = 0.22-0.36), 而在软阔叶林中很低(r = -0.05), 也反映出随着演替的进展优势树种对水分需求提高的总体趋势。
图3 不同演替阶段轮宽指数与单月气候因子的相关系数(标注的数字为p值)。A, 上年6月降水。B, 上年12月平均气温。C, 当年6月降水。D, 当年6月平均气温。Early, 早期; Middle, 中期; Mid-late, 中晚期; Late, 晚期。
Fig. 3 Correlation coefficient between ring-width indices and monthly climate parameters of different successional stage (the number associated with each coefficient was p value). A, Precipitation in June of the previous year. B, Mean air temperature in December of the previous year. C, Precipitation in June of the current year. D, Mean air temperature in June of the current year.
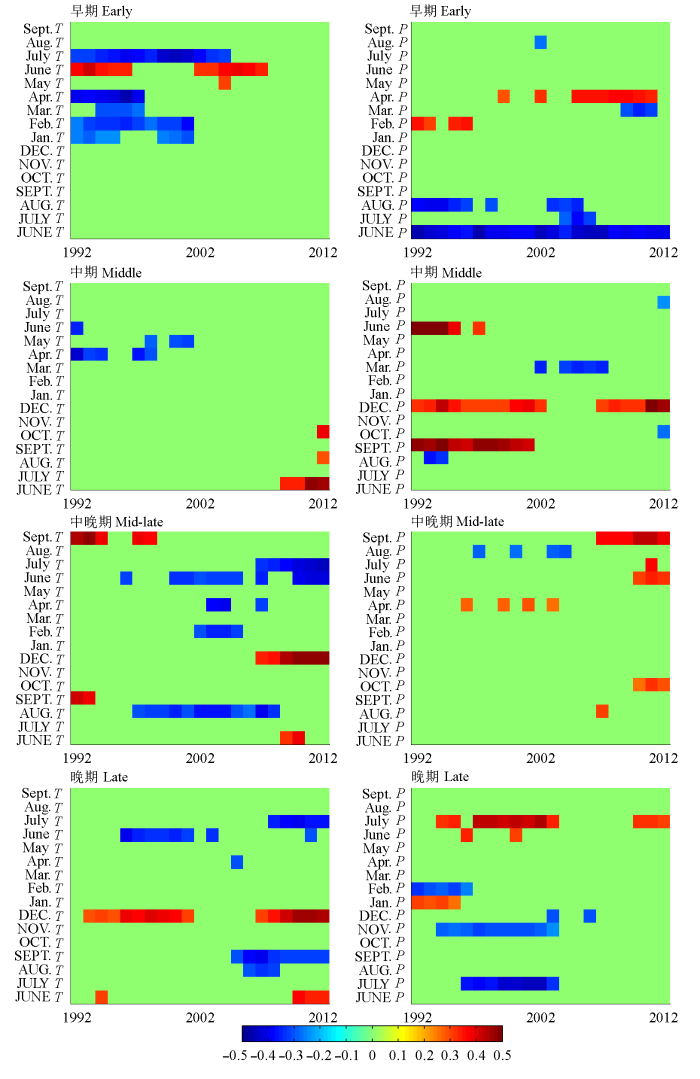
采用窗口为32年的滑动相关来分析各演替阶段年表与气候因子的动态关系。结果表明, 对于软阔叶林, 在滑动区间内(1992-2012年)上年6月的降水量与径向生长的负相关是一种稳定的限制作用。当年4月降水量与树木生长的正相关关系增强, 当年1-4月及6-7月气温与生长的显著相关性则都在近些年消失。
对于硬阔叶林, 上年12月的降水量与生长的正相关虽有所波动, 但总体上表现为稳定的限制因素。上年9月降水量与生长的正相关则在2002年后 消失。
对于红松林, 当年生长季水分对生长的限制作用有增强趋势, 表现为近熟、成熟林的生长与6、7月气温的负相关, 以及与6、7月降水量的正相关都总体上增强。上年12月气温与生长的正相关在近熟、成熟林中也有增强的趋势。
在各林型中, 都还有一些其他的显著关系, 由于相关性显著的年份较少或相关性存在起伏波动, 难以判断为明确的变化趋势, 也不能排除为偶然的相关(常锦峰等, 2009; 曾令兵等, 2012), 暂不予讨论。
总体来看, 除一些气候因子对径向生长表现出稳定的限制作用之外, 对红松林而言, 当年生长季水分、上年冬季气温对其径向生长的正作用都有增强趋势, 而对于演替早期的软阔叶林, 当年春季(4月)降水量与其径向生长的正相关也有增强的趋势。
表4 研究区不同演替阶段差值年表轮宽指数与单月气候因子的相关性
Table 4 Correlations between ring-width indices and monthly climatic indices for different successional stages
| 月平均气温 Monthly mean air temperature | 月降水量 Monthly precipitation | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 早期 Early | 中期 Middle | 中晚期 Mid-late | 晚期 Late | 早期 Early | 中期 Middle | 中晚期 Mid-late | 晚期 Late | ||
| p6 | 0.175 | 0.246 | 0.107 | 0.276* | -0.428** | -0.207 | -0.214 | -0.190 | |
| p7 | -0.023 | -0.059 | 0.088 | 0.068 | -0.112 | 0.028 | -0.077 | -0.155 | |
| p8 | -0.074 | 0.069 | -0.232 | -0.120 | -0.266 | -0.037 | -0.009 | 0.032 | |
| p9 | 0.060 | 0.058 | 0.021 | -0.151 | -0.022 | 0.147 | 0.091 | 0.114 | |
| p10 | -0.147 | 0.139 | -0.061 | 0.131 | -0.199 | -0.102 | 0.068 | -0.025 | |
| p11 | 0.102 | 0.011 | 0.021 | 0.089 | -0.088 | 0.048 | 0.211 | -0.133 | |
| p12 | -0.128 | -0.132 | 0.252’ | 0.272* | 0.110 | 0.353** | -0.171 | -0.086 | |
| c1 | -0.129 | 0.010 | -0.052 | -0.048 | 0.149 | 0.023 | -0.110 | 0.065 | |
| c2 | -0.224 | -0.033 | -0.021 | 0.015 | 0.160 | -0.007 | 0.031 | -0.139 | |
| c3 | -0.013 | -0.025 | 0.128 | 0.052 | -0.130 | -0.154 | -0.113 | -0.019 | |
| c4 | -0.184 | -0.275* | 0.053 | 0.051 | 0.208 | 0.037 | 0.182 | 0.045 | |
| c5 | 0.027 | -0.114 | -0.103 | -0.065 | -0.065 | 0.054 | -0.051 | 0.018 | |
| c6 | 0.189 | -0.258’ | -0.286* | -0.322* | -0.053 | 0.362** | 0.215 | 0.304* | |
| c7 | -0.318* | -0.106 | -0.115 | -0.158 | 0.067 | 0.100 | 0.132 | 0.228 | |
| c8 | -0.028 | -0.020 | -0.015 | -0.004 | -0.019 | 0.079 | -0.174 | 0.079 | |
| c9 | 0.021 | 0.001 | 0.092 | -0.069 | 0.059 | 0.117 | 0.180 | 0.021 | |
图4 不同演替阶段年表与气候因子的滑动相关关系分析。T, 月平均气温; P, 月降水量。T或P前的大写字母(如JUNE)为上年各月, 首字母大写的(如June)为当年各月。绿色表示年表与气候因子的相关关系不显著。
Fig. 4 Correlations with moving intervals between ring-width indices and monthly climatic indices for different successional stages. T, monthly mean air temperature; P, monthly precipitation. Capital letters before T or P (e.g. JUNE) denote the climatic indices for the previous year, and the first letter capitalized (e.g. June) is for the current year. Green color indicates non-significant correlations.
本文通过对阔叶红松林不同演替阶段的径向生长和气候因子关系的分析发现气温和降水量对树木径向生长的影响随着演替的进展存在着规律性的变化。本文研究结果与以往在东北地区的研究既有一致的地方, 也有一些明显的差异。
前人对长白山中低海拔红松林的研究表明, 红松的径向生长与上年冬季(12月)月平均气温显著负相关(高露双等, 2013), 本研究的结果与之相反, 这可能是由于研究地点的不同所导致的。很多研究都指出, 不同气候条件下, 同一树种的生长和某一气候指标的关系会有很大的差异(常锦峰等, 2009; 曾令兵等, 2012)。胜山自然保护区位于红松分布的北纬上限, 冬季的气温远低于长白山。在本文研究中, 上年冬季气温和年轮生长的正相关关系应当反映了高纬度地区冬季低温对红松生长的限制作用。比如, 在寒冷地区冬季的气温较高可保持树木代谢活动的正常, 有利于常绿针叶树进行光合作用, 从而为下年树木的生长积累能量, 加快树木下一生长季的径向生长(Fritts, 1976; Gutiérrez, 1991)。这种冬季气温对树木径向生长的正效应在软阔叶林、硬阔叶林阶段完全不显著, 可能与其优势种桦树、蒙古栎等在冬季不能进行光合作用有关。相反, 较高的冬季气温还会增加这些落叶阔叶树种因呼吸作用而消耗的有机物质, 可能影响来年树木生长。
对长白山中低海拔红松林的研究表明, 红松的轮宽指数与当年生长季的气温负相关(高露双等, 2013), 本研究的结果与之一致。树木径向生长与生长季的气温负相关主要是由于气温上升加快了土壤的水分蒸发, 并促进植物的蒸腾作用, 进而导致水分有效性降低, 实际上体现了水分的限制作用(曾令兵等, 2012)。对长白山暗针叶林和红松林过渡带的研究则表明, 红松径向生长和生长季气温正相关(高露双等, 2011)。有研究表明, 随着海拔升高, 热量减少而水分条件改善(包括降水量的增加和蒸发量的减少), 降水量对树木生长的限制作用逐步减弱乃至消失(常锦峰等, 2009)。因此上述研究结果的差异说明, 红松对水分条件的要求是比较高的, 其生长首先受到水分的限制, 生长季气温升高对其生长的正效应只有在没有水分亏缺的情况下(如海拔分布上限)才能实现, 这一点在本文与高露双等(2011)的研究结果比较中表现得更为明显。本研究地区处于红松纬度分布的上限, 从热量水平来说和高露双等(2011)的研究相似, 都处在红松分布的热量下限(Wang et al., 2006b)。但由于胜山是红松的分布北界, 即红松只能分布在低海拔(周以良, 1997), 水分条件不如长白山的暗针叶林和红松林过渡带, 因此和长白山低海拔红松林(高露双等, 2013)一样, 也表现出明显的生长季水分限制。森林或树种的海拔和纬度分布上限在气候变化尤其是树轮年代学的研究中受到了广泛的关注(Holtmeier & Broll, 2005; Wang et al., 2006a), 但关于同一树种在海拔和纬度分布上限的系统比较研究还很有限。上述分析表明, 这种比较研究对揭示树种对气候变化的响应可能具有重要的意义, 今后需要加强这方面的研究。
此外, 其他的研究证据也支持红松对水分的这种生理需求。如陈列等(2013)对长白山典型红松林的研究发现红松的径向生长与当年生长季的降水正相关。吴玉莲等(2014)利用Biome-BGC模型模拟长白山阔叶红松林净初级生产力对气候变化的响应, 模拟结果表明红松净初级生产力与生长季的降水量正相关、与生长季初期的气温负相关, 从另一个方面也印证了上述通过树木年轮得到的结果。
相对于针叶树和高山林线(王晓春等, 2004; Yu et al., 2007; 常锦峰等, 2009; 高露双等, 2011), 在东北关于红松林演替系列的树轮比较研究还较少。本文的研究结果表明, 径向生长对气候变化的响应随着演替的进展存在规律性的变化(图3), 而这种变化与演替前期阔叶树与红松对水热条件的生理需求不同有着密切关系。比如, 本文的结果表明软阔叶林与上年生长季的降水显著负相关且关系稳定(图4), 这与高露双等(2013)对长白山山杨的研究结果一致。阔叶红松林演替的先锋树种山杨、白桦等属阳性树种, 喜光耐寒, 但不耐湿, 生长季过多的降水会改变土壤的水环境, 从而可能抑制下一年树木的生长(Armstrong et al., 1994)。而随着演替的进展, 林分优势种逐渐过渡为中生、耐阴树种, 因此上年6月的降水与径向生长的负相关关系在后几个演替阶段就不再显著(图3A)。又如, 当年6月气温对径向生长的负作用从演替早期到演替晚期逐渐增强, 而当年6月降水对径向生长的正效应则呈相反的趋势(图3C、3D), 这与随着演替的进展优势树种对水分的需求逐步提高的事实是一致的。
关于东北地区硬阔叶树的树轮研究目前还十分有限。李腾等(2014)在东北南部(辽宁千山)的研究表明, 蒙古栎的年轮宽度与当年春、夏的降水正相关, 与气温负相关, 和本研究在胜山得到的结果(表4) 相似。此外, 吴玉莲等(2014)的Biome-BGC模拟结果表明, 长白山硬阔叶树净初级生产力也与生长季降水正相关。辽宁千山已超出了阔叶红松林分布的南界(周以良, 1997), 综合上述研究结果可以推测, 在整个红松林分布区, 当年生长季水分对硬阔叶树径向生长的强限制作用可能是普遍现象。从图3可以看出, 硬阔叶林与气候因子的关系总体上与两种红松林更为相似, 而与软阔叶林差距较大, 这与硬阔叶林的主要树种也是红松的伴生树种的事实也是吻合的。不过, 由于类似的研究还很少, 需要进一步开展红松林不同演替阶段的比较研究, 对这些问题进行系统的研究。
本研究区的气候近几十年来表现出明显的干暖化现象(图1), 这种气候变化趋势可能对红松林的生长、演替乃至分布产生影响。本文结果表明, 生长季降水对于分布区北缘的红松林生长是一种关键的限制作用(图3C、3D)。该限制作用在近十几年来有增强的趋势(图4), 也与气候干暖化的趋势一致。对长白山中低海拔红松林的研究(高露双等, 2013)也发现, 红松径向生长由于气候变暖, 与生长季气温的负相关关系在增强。长白山近几十年来降水量下降的幅度较弱, 因此, 高露双等(2013)的研究说明仅生长季气温上升导致的水分有效性降低就足以对红松生长产生明显抑制作用, 进一步反映了红松生长对生长季水分较高的需求。胜山地区降水量减少的趋势十分明显(图1), 可以预测, 气候变化对胜山红松林生长的不利影响较长白山将更为严重。
气候变暖对胜山红松林也表现出一定的促进作用: 上年冬季气温与生长正相关(图3), 且这种促进作用在两种红松林中都表现出增强的趋势(图4)。因此气候变暖可能存在有利于红松林生长的方面。不过, 从图2来看, 两种红松林的轮宽指数在20世纪70年代以来均表现出明显的下降趋势(轮宽和年份的相关系数在近熟林、成熟林分别为-0.47、-0.31), 说明生长季水分的抑制作用影响更大, 其综合效应表现为近几十年来的气候变化对分布区北缘红松林的生长不利。近年来, 不少气候-年轮学研究也都表明, 气候变暖导致的水分不足可能导致高纬度森林生长下降, 而不是如早期所认为的气候变暖促进生长(Barber et al., 2000)。
本文结果表明上年6月的降水与软阔叶林生长的负相关关系具有稳定性(图3A, 图4), 上年6月的降水是关键的限制作用, 进一步反映了白桦等先锋树种不耐湿的生理特征。未来降水进一步减少是否会导致该限制作用消失, 从而有利于软阔叶林的生长还有待进一步研究。滑动相关表明软阔叶林的生长与气温的显著相关性消失, 而与当年4月降水的正相关关系有增强趋势, 与研究区气候干暖化的趋势一致。从轮宽指数近几十年没有明显上升或下降的趋势(图2)来看, 气候变化的正、负效应同时作用, 对软阔叶林生长尚未产生明显的长期影响。对于硬阔叶林, 上年12月的降水与生长的正相关为稳定的限制作用, 而其余限制作用各有增强或消失的趋势(图4)。从年表来看, 各种因素综合作用下, 近几十年轮宽指数总体平稳, 但在1995年后有明显的增长。这种增长的趋势是否指示着气候变化的长期影响, 还有待进一步研究。
综合来看, 近几十年的气候干暖化对红松林各演替阶段的径向生长影响复杂, 正负效应都存在。对于演替前期的软阔叶林、硬阔叶林, 气候变化还没有导致明显的长期生长变化趋势。但对于红松林, 生长季水分的主导限制作用已导致生长的下降。可以预测, 今后的气候干暖化是不利于东北地区红松林的生长和恢复演替的。这有可能导致红松林分布区向南方降水量较高的区域退缩。高露双等(2011, 2013)在长白山的研究也认为气候变暖可能导致红松在低海拔的适宜分布区缩小, 但可能向较湿润的高海拔扩展。如果这些趋势在今后的研究中得到证实, 则意味着气候变暖后东北红松的适宜分布区将大幅缩小, 对阔叶红松林的恢复是十分不利的。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
| [1] |
Mechanisms of flood tolerance in plants. https://doi.org/10.1111/j.1438-8677.1994.tb00756.x URL [本文引用: 1] 摘要
First page of article
|
| [2] |
Reduced growth of Alaskan white spruce in the twentieth century from temperature-induced drought stress. |
| [3] |
Alpine timberline dynamics in relation to climatic variability in the northern Daxing’an Mountains. 大兴安岭北部大白山高山林线动态与气候变化的关系 .
高山林线植被对气候变化十分敏感,已成为全球变化研究的热点.研究了大兴安岭北部大白山高山林线的树木生长和群落更新动态及其与气候变化、火干扰等因素的关系.结果表明,林线树木的生长对气候变化十分敏感,但其敏感性随着海拔的降低而减弱;在高海拔,林线树木的径向生长与上年生长季后期(8月)降水负相关,而与上年初秋(9月)温度正相关,这限制作用随着海拔的降低而逐渐消失;与此相反,低海拔树木生长与当年冬末春初(3月)的温度负相关,但随着海拔上升这种限制作用消失.分析结果还表明,本地区的群落更新主要受火干扰驱动,而与气候变化没有显著关系.不同树种在火灾后更新的时间存在差异,这种差异又因海拔的不同而异,反映出不同树种的更新策略及对环境变化适应能力的差异.大兴安岭北部的高山林线在树木生长对气候变化的敏感性、与气候因子的关系及群落更新动态等方面均与干旱区林线有明显的差异,这些差异与气候条件、树种、更新驱动力等方面的不同有密切的关系.<dt><strong><t>Abstract:</t></strong></dt><dd>Alpine timberline is very sensitive to zlimatic change,and has become a focus in global change research.In this study,we developed tree-ring chronologies and age structures for timberline forests in the northern Daxing'all Mountains,to examine timberline dynamics in relation to climatic variability.Our results showed that:1)The statistical characters of the chronologies suggested that the sensitivity of tree growth to environmental changes increased with increasing altitude.2)The correlations between tree-ring width and climatic records showed remarkable altitudinal differences.Tree radial growth was mainly limited by previous August precipitation and previous September temperature at high-elevation sites,however,the importance of these climatic factors decreased with decreasing altitude.In contrast,tree growth was mainly limited by current March temperature at low-elevation site.3) The age structures for the timberline forests suggested that fire disturbance,instead of climatic factors,was the major driver for tree recruitment in the study region.The recruitment patterns differed with tree species and elevation,suggesting the differences in regeneration strategy and cold tolerance for different tree species.4)The timberline dynamic in the study region showed clear differences from that of arid regions in several respects,which may be related to differences in climatic conditions,tree species and recruitment drives.
|
| [4] |
Characteristics of tree-ring chronology of Pinus koraiensis and its relationship with climate factors on the northern slope of Changbai Mountain. 长白山北坡不同林型内红松年表特征及其与气候因子的关系 .https://doi.org/10.5846/stxb201209181309 URL Magsci 摘要
运用树木年轮学的基本原理和方法,选取了长白山北坡保存完好的典型性植被阔叶红松林,探讨了杨桦红松林和椴树红松林内建群种红松(<em>Pinus koraiensis</em>)径向生长对气候要素的响应。结果表明,长白山北坡红松的径向生长对降水较为敏感,杨桦红松林和椴树红松林中,红松年轮宽度均与当年7月以及上一年9月的降水呈显著正相关关系。不同林型内红松的生长与气候因子的关系也有差异。椴树红松的年轮宽度还与上年7月的降水显著负相关,与当年3、4月份的平均气温呈显著正相关。而杨桦红松林内红松年轮宽度和平均温度没有显著的相关关系。特征年分析进一步验证了响应函数相关分析的结果,即当年生长季以及上年生长季末的降水充足促进了红松的径向生长;椴树红松林中,初春温度的升高有利于红松的生长。
|
| [5] |
Analysis of the tree-ring width chronologies of Pinus koraiensis and Larix olgensis on Changbai Mountains, Northeast China. 长白山红松和落叶松树轮宽度年表特征 .
利用采自长白山红松和落叶松分布上限和下限的树轮样本,分别建立树轮宽度差值年表。结果表明:下限红松年表的平均敏感度、树轮宽度指数的年际变率、信噪比等年表特征值比上限年表的高,表明下限红松生长对环境的敏感性大于上限;落叶松年表特征值在不同海拔表现与红松相反。进一步分析发现分布上限和下限的红松生长与气候因子之间的关系不同。下限红松生长与当年气温和上年降水显著相关,上限红松生长与当年气温显著相关,下限落叶松与上年气温显著相关,上限落叶松生长与当年气温和降水显著相关,降水随着海拔的升高而增加,过多的雨水伴随气温下降,进而限制落叶松生长,这表明落叶松生长的重要限制因子是气温,气温对下限落叶松生长产生滞后效应。
|
| [6] |
A Time Series Analysis Approach to Tree-ring Standardization. PhD dissertation, The University of Arizona, Tucson , |
| [7] |
Methods of Dendrochronology: Applications in the Environmental Sciences. Springer Science & Business Media, Berlin , |
| [8] |
Tree rings and climate. |
| [9] |
Response of Pinus koraiensis and Picea jezoensis var. komarovii to climate in the transition zone of Changbai Mountain, China. 长白山过渡带红松和鱼鳞云杉径向生长对气候因子的响应 .https://doi.org/10.3724/SP.J.1258.2011.00027 URL Magsci [本文引用: 2] 摘要
<p>利用树轮生态学方法, 研究了长白山阔叶红松林和暗针叶林过渡带优势树种红松(<em>Pinus koraiensis</em>)和鱼鳞云杉(<em>Picea jezoensis</em> var. <em>komarovii</em>)的生长特征及其与气候因子的关系, 以期揭示气候响应关系的种间差异性。结果表明, 红松和鱼鳞云杉年平均径向生长量与生理年龄显著相关, 红松先于鱼鳞云杉达到最大年生长量, 且红松年平均生长速率显著高于鱼鳞云杉(<em>p</em> < 0.001); 红松和鱼鳞云杉对气候的响应存在差异, 红松与7月份的月平均温度和降水显著正相关, 而鱼鳞云杉与5月平均温度显著正相关, 与5月降水显著负相关。响应面分析进一步证实, 红松生长主要与生长季温度和降水相关, 而生长季初期的降水是限制鱼鳞云杉生长的主要原因。全球变暖有利于红松径向生长, 红松种群有向高海拔上升的可能。</p>
|
| [10] |
Growth response of two coexisting species to climate change in broadleaved Korean pine forests in Changbai Mountain, northeastern China. 长白山阔叶红松林共存树种径向生长对气候变化的响应 .
利用树木年轮学方法,通过研究长白山阔叶红松林建群树种红松和先锋树种山杨生长特征及其对气候因子的响应,以期揭示不同物种对气候因子的响应差异.结果表 明:红松和山杨对气候因子响应关系存在差异.红松主要受上年冬季温度和降水的负作用,山杨主要受到生长季末降水的限制作用.在极端情况下,上一年冬季的高 温和较多降水以及当年生长季高温是导致红松形成窄轮的主要原因,而上一年生长季的较多降水和当年春季高温是影响山杨生长的主要原因.因此,树木生长—气候 因子的关系具有一定的物种特性.此外,还发现随着温度上升和降水格局的改变,春季降水对红松生长的正效应以及月最低温度对红松和山杨的负效应将逐渐突显, 这可能是导致红松适宜区面积缩小的主要因素.
|
| [11] |
Tree growth response to climate change at the deciduous boreal forest ecotone, Ontario, Canada. |
| [12] |
The scientific significance of the forest vegetation ecotone between Daxing’an and Xiaoxing’an Mountains to global climate change study. 大小兴安岭森林植被交错区生态系统在全球气候变化研究中的科学意义 .https://doi.org/10.3969/j.issn.1000-5382.2001.05.001 URL [本文引用: 1] 摘要
对大小兴安岭森林植被交错区的 地理位置、地形地貌、气候、土壤类型、植被类型以及生态脆弱性等进行了论述。在交错区内 ,随着气候从温带向寒温带逐渐过渡 ,土壤、植被等表现出明显的梯度变化 ,该地区又是我国近百年来温度增加最明显的区域 ,导致冻土等环境条件的显著变化 ,为研究气候变暖对森林生态系统结构、功能的影响提供了良好的场所 ,在全球气候变化研究中占有重要地位
|
| [13] |
Climate-tree-growth relationships for Pinus uncinata Ram. in the Spanish pre-Pyrenees. |
| [14] |
Computer-assisted quality control in tree- ring dating and measurement. |
| [15] |
Sensitivity and response of northern hemisphere altitudinal and polar treelines to environmental change at landscape and local scales. https://doi.org/10.1111/j.1466-822X.2005.00168.x URL [本文引用: 1] 摘要
The sensitivity and response of northern hemisphere altitudinal and polar treelines to environmental change are increasingly discussed in terms of climate change, often forgetting that climate is only one aspect of environmental variation. As treeline heterogeneity increases from global to regional and smaller scales, assessment of treeline sensitivity at the landscape and local scales requires a more complex approach than at the global scale. The time scale (short-, medium-, long-term) also plays an important role when considering treeline sensitivity. The sensitivity of the treeline to a changing environment varies among different types of treeline. Treelines controlled mainly by orographic influences are not very susceptible to the effects of warming climates. Greatest sensitivity can be expected in anthropogenic treelines after the cessation of human activity. However, tree invasion into former forested areas above the anthropogenic forest limit is controlled by site conditions, and in particular, by microclimates and soils. Apart from changes in tree physiognomy, the spontaneous advance of young growth of forest-forming tree species into present treeless areas within the treeline ecotone and beyond the tree limit is considered to be the best indicator of treeline sensitivity to environmental change. The sensitivity of climatic treelines to climate warming varies both in the local and regional topographical conditions. Furthermore, treeline history and its after-effects also play an important role. The sensitivity of treelines to changes in given factors (e.g. winter snow pack, soil moisture, temperature, evaporation, etc.) may vary among areas with differing climatic characteristics. In general, forest will not advance in a closed front but will follow sites that became more favourable to tree establishment under the changed climatic conditions.
|
| [16] |
Response of tundra ecosystem in southwestern Alaska to Younger Dryas climatic oscillation. |
| [17] |
|
| [18] |
The importance of phenology for the evaluation of impact of climate change on growth of boreal, temperate and Mediterranean forests ecosystems: An overview. https://doi.org/10.1007/s004840000066 Magsci [本文引用: 2] 摘要
<a name="Abs1"></a>An overview is presented of the phenological models relevant for boreal coniferous, temperate-zone deciduous and Mediterranean coniferous forest ecosystems. The phenology of the boreal forests is mainly driven by temperature, affecting the timing of the start of the growing season and thereby its duration, and the level of frost hardiness and thereby the reduction of foliage area and photosynthetic capacity by severe frost events. The phenology of temperate-zone forests is also mainly driven by temperature. Since temperate-zone forests are mostly mixed-species deciduous forests, differences in phenological response may affect competition between tree species. The phenology of Mediterranean coniferous forests is mainly driven by water availability, affecting the development of leaf area, rather than the timing of phenological events. These phenological models were subsequently coupled to the process-based forest model FORGRO to evaluate the effect of different climate change scenarios on growth. The results indicate that the phenology of each of the forest types significantly affects the growth response to a given climate change scenario. The absolute responses presented in this study should, however, be used with caution as there are still uncertainties in the phenological models, the growth models, the parameter values obtained and the climate change scenarios used. Future research should attempt to reduce these uncertainties. It is recommended that phenological models that describe the mechanisms by which seasonality in climatic drivers affects the phenological aspects of trees should be developed and carefully tested. Only by using such models may we make an assessment of the impact of climate change on the functioning and productivity of different forest ecosystems.
|
| [19] |
Tree-ring growth variability in the Austrian Alps: The influence of site, altitude, tree species and climate. https://doi.org/10.1080/03009480701267063 URL [本文引用: 1] 摘要
We present an extensive new network of ring-width chronologies, comprising data from 100 sites within the Austrian Eastern Alps, made up of multiple tree species. Principal components analysis and cluster analysis were used to identify five separate high-frequency tree-growth signals from these data. Tree-growth variability at these sites is explained by site altitude and species differences that moderate the effects of the annually varying climatic forcing on tree growth. Recently developed, two-century long gridded climate data sets for the Greater Alpine Region were used to assess the relationship between climate and tree growth. Tree growth at low altitudes is controlled mainly by spring-summer moisture availability. At high altitudes precipitation is no longer a limiting factor and growth is mostly determined by summer temperature. In the intermediate altitudinal range, we did not find any direct relationships with specific climatic variables. High-altitude chronologies suitable for reconstructing past temperatures and low-altitude chronologies suitable for reconstructing past precipitation were identified.
|
| [20] |
Mixed stands reduce Abies alba tree-ring sensitivity to summer drought in the Vosges mountains, western Europe. https://doi.org/10.1016/j.foreco.2013.04.003 URL Magsci 摘要
In most dendroecological studies, climate-tree growth relationships are established for trees growing on pure stands. However, response to climate may be affected by inter-species interactions and local constraints, which beg the question of the effect of mixture on tree growth response under various ecological conditions. To assess these effects, climate-tree growth relationships of pure Abies alba stands were compared to those of three different mixtures: A. alba with Fagus sylvatica, with Picea abies and with both species. 151 stands (456 A. alba trees) were sampled in the Vosges mountains in north-eastern France under three contrasted climates, from low altitude and dry conditions (mean precipitation in July <85 mm and altitude <600 m) to high altitude and humid conditions (P July >115 mm and alt. >900 m). We sampled adult trees and homogeneous stand conditions to clearly assess differences in sensitivity to climate. Climate-tree growth relationships were evaluated from 12 A. alba chronologies (four mixtures x three climatic conditions) through pointer years and response function analyses. Late previous summer conditions and current summer soil water deficit and temperature played a major role on A. alba growth. Results showed greater sensitivity to temperature at high elevation, and to summer drought at low altitude and under dry conditions. Mixture allowed maintaining a higher level of A. alba growth during extreme climatic events and reduced A. alba response to summer drought especially under the driest contexts. Different facilitation processes may explain mixture effects such as changes in rooting depth, water input by stemflow and rainfall interception. This differentiated functioning of mixed forests highlights their importance for adapting forest management to climate change. (c) 2013 Elsevier B.V. All rights reserved.
|
| [21] |
|
| [22] |
Tree-ring growth responses of Mongolian oak (Quercus mongolica) to climate change in southern Northeast: A case study in Qianshan Mountains. 东北南部蒙古栎径向生长对气候变化的响应——以千山为例 .
<p>蒙古栎是东北森林中最重要的阔叶树种之一.本研究利用树木年代学方法研究中国东北南部千山地区蒙古栎的径向变化,结合1951—2010年的温度和降水等气象数据,利用相关函数分析了树木生长与气候变化的关系,揭示蒙古栎径向生长对气候响应规律.结果表明:研究区4—7月的降水量与蒙古栎年轮宽度呈显著正相关,是限制该地区蒙古栎径向生长的主要限制因子;5月极端最高温度与蒙古栎年轮宽度呈显著负相关,也是影响蒙古栎生长的关键因素.研究期间,蒙古栎年轮宽度与4月降水量的相关显著且稳定,自20世纪80年代开始蒙古栎径向生长对夏季温度的响应敏感性逐渐减弱,对温度的响应表现出从响应夏季温度向响应春季温度的转变.</p>
|
| [23] |
Tree-ring based summer temperature reconstruction for the source region of the Yangtze River on the Tibetan Plateau. https://doi.org/10.1016/j.gloplacha.2007.10.008 URL 摘要
Understanding of past climatic variability over the Tibetan Plateau is still limited because of the lack of long-term climatic records. Here we reconstruct the mean summer (June-August) minimum temperature for the past 379 years based on tree-ring data in the source region of the Yangtze River. This reconstruction successfully captures recent abrupt climatic changes and agrees in general with other temperature reconstructions for the Tibetan Plateau on a decadal timescale. The cold and warm periods coincide with documented glacier advances and retreats on the east and southeast Tibetan Plateau. The interval 1816-22 is among the coldest periods in the reconstruction and may be related to the influence of the Tambora eruption in Indonesia in 1815. Comparisons with other paleoclimatic proxies imply a high degree of confidence for our reconstruction and its indicative power for a large-scale climate variability on the Tibetan Plateau.
|
| [24] |
Effect of climate change on net primary productivity of Korean pine (Pinus koraiensis) at different successional stages of broad-leaved Korean pine forest. 气候变化对阔叶红松林不同演替阶段红松种群生产力的影响 .
<p>本文以长白山地区阔叶红松林不同演替阶段(次生杨桦林、次生针阔混交林、原始红松林)内红松种群作为研究对象,采用树轮学与相对生长式相结合,获取红松种群净初级生产力(NPP)连年生长(1921—2006年)数据以及相对增长率的年际变化数据,建立红松种群NPP与年际和季节性气候因子的关系,分析不同气候时期长白山阔叶红松林不同演替阶段内红松种群NPP年际变化特征及其对气候变化的响应差异.结果表明: 研究期间,不同演替阶段红松种群NPP与气候因子响应关系存在差异.随着温度上升,次生杨桦林红松种群NPP与上年生长季和当年生长季低温由显著负相关关系转变为显著正相关关系;次生针阔混交林红松种群NPP由与当年春季最低温度的正相关关系转变为与上年和当年生长季温度的显著正相关关系,气候因素对次生针阔混交林红松种群NPP影响的滞后效应增强;原始红松林红松种群NPP与温度的相关性减弱,与上年生长季降水量的正相关关系增强.研究区气候变化表现为低温和平均温度显著上升,而最高温度和降水没有明显变化.气候变化有利于提高演替初级阶段次生杨桦林和演替中级阶段次生针阔混交林内红松种群生产力,尤其是次生针阔混交林,而对演替顶极阶段红松种群NPP影响不明显.</p>
|
| [25] |
|
| [26] |
Plants in a warmer world. https://doi.org/10.1078/1433-8319-00076 URL [本文引用: 1] 摘要
The multitude of recently published studies providing evidence for the ecological impacts of climate change on many different continents strongly suggests that the last 30 years of warmer temperatures have had a substantial influence on both seasonal patterns, and altitudinal and poleward shifts in vegetation. Common features of change, but also some discrepancies in the response of plants to climate change, are discussed, as well as implications for biodiversity, higher level impacts on community structure and trophic interactions, and some ecosystem consequences.
|
| [27] |
Multi-model ensemble prediction of extreme temperature indices in China. 21世纪中国极端气温指数变化情况预估 .
利用政府间气候变化委员会资料中心的模式预测结果(IPCC-AR4),分析了在高(SRES A2)、中(SRES A1B)、低(SRES B1)3种不同的排放情景下中国未来21 世纪极端气温变化特征。研究结果表明:随着全球变暖,中国区域极端气温指数的变化是一致的增加(减少)趋势。其中霜冻日数和温度年较差呈减少趋势,生长季指数、热浪指数和暖夜指数是增加趋势。在高排放(<i>A</i>2)情景下,中国地区各极端气温指数增加(减少)趋势最明显,低排放(<i>B</i>1)情景下极端气温指数变化趋势最小。在2075年前,极端气温指数在<i>A</i>1<i>B</i>情景下的变化幅度要大于<i>A</i>2情景下的变化,2075年之后,却与之相反。在各极端气温指数中,热浪指数(<i>HWDI</i>)和暖夜指数(<i>TN</i>90)上升趋势最为明显,其次是霜冻日数(<i>FD</i>)和生长季指数, 气温年较差(<i>ETR</i>)变化最小。在空间分布的特征上,极端气温指数基本上保持一致的增加或减少的变化趋势(温度年较差除外),变化基本上是由北向南变化率逐渐增大,其中西北的极端指数变化率高于东北,只有暖夜指数的变化情况有所不同,在西南和华南地区出现了变化的大值区。
|
| [28] |
a). Treeline dynamics in relation to climatic variability in the central Tianshan Mountains, northwestern China. https://doi.org/10.1111/j.1466-822X.2006.00233.x URL [本文引用: 1] 摘要
ABSTRACT Aim68 Climate variability may be an important mediating agent of ecosystem dynamics in cold, arid regions such as the central Tianshan Mountains, north-western China. Tree-ring chronologies and the age structure of a Schrenk spruce () forest were developed to examine treeline dynamics in recent decades in relation to climatic variability. Of particular interest was whether tree-ring growth and population recruitment patterns responded similarly to climate warming. Location68 The study was conducted in eight stands that ranged from 250002m to 275002m a.s.l. near the treeline in the Tianchi Nature Reserve (43°45′6143°59′02N, 88°00′6188°20′02E) in the central Xinjiang Uygur Autonomous Region, northwestern China. Methods68 Tree-ring cores were collected and used to develop tree-ring chronologies. The age of sampled trees was determined from basal cores sampled as close as possible to the ground. Population age structure and recruitment information were obtained using an age–d.b.h. (diameter at breast height) regression from the sampled cores and the02d.b.h. measured on all trees in the plots. Ring-width chronologies and tree age structure were both used to investigate the relationship between treeline dynamics and climate change. Results68 Comparisons with the climatic records showed that both the radial growth of trees and tree recruitment were influenced positively by temperature and precipitation in the cold high treeline zone, but the patterns of their responses differed. The annual variation in tree rings could be explained largely by the average monthly minimum temperatures during February and August of the current year and by the monthly precipitation of the previous August and January, which had a significant and positive effect on tree radial growth. recruitment was influenced mainly by consecutive years of high minimum summer temperatures and high precipitation during spring. Over the last several decades, the treeline did not show an obvious upward shift and new recruitment was rare. Some trees had established at the treeline at least 20002years ago. Recruitment increased until the early 20th century (1910s) but then decreased with poor recruitment over the past several decades (1950–2000). Main conclusions68 There were strong associations between climatic change and ring-width patterns, and with recruitments in Schrenk spruce. Average minimum temperatures in February and August, and total precipitation in the previous August and January, had a positive effect on tree-ring width, and several consecutive years of high minimum summer temperature and spring precipitation was a main factor favouring the establishment of following germination within the treeline ecotone. Both dendroclimatology and recruitment analysis were useful and compatible to understand and reconstruct treeline dynamics in the central Tianshan Mountains.
|
| [29] |
The effect of climate warming on the structure characteristic of the timberline in Laotudingzi Mountain. 气候变暖对老秃顶子林线结构特征的影响 .
运用样带样方法和年轮气候学方法对大海林地区的气候因子和样地数据进行了分析。结果表明 ,近 30 a来老秃顶子地区气候变暖明显 ,尤其是冬季增温最明显 ,月份增温中以 2月份最大 ;寒冷时期 (12月份、翌年 1和 2月份 )和温暖时期 (6~ 9月份 )的温度都有增加 ,但是寒冷时期温度的增加幅度较大 ;冬季与夏季温差稍有减少 ,但积温有所增加 ,整年的热量正在增加。全球变暖导致的大海林地区增温对老秃顶子林线结构特征产生了很大的影响 ,由样地调查和分析可知 ,全球变暖导致林线中上部幼苗、幼树的更新和存活增多 ,森林密度加大 ,树木平均年龄降低 ,年龄结构呈倒 J字型 ,并且多呈聚集分布 ;而在林线的下部 ,幼苗更新很少 ,主要以中龄林存在 ,并且多呈零散分布形式。通过年轮分析得出 ,气候变暖导致林线树木径生长和高生长增加 ,而且增加的趋势和近 30 a来温度的变化基本一致。通过对年轮指数与气候因子的相关性分析 ,表明林线树木年轮指数与温度的相关性较强 ,而与降水的相关性较弱 ,并且年轮指数与温暖时期温度和积温呈正相关 ,而与寒冷时期温度和年平均温度呈负相关 ,表明温暖时期温度和积温控制着林线的海拔高度 ,而寒冷时期的温度和年平均温度主要对林线树种类型起着决定性的作用。从敏感度分析看出,林线中上部对气候变化敏感,而非最上部;在不同树种中比较敏感度时,发现岳桦的平均敏感度要比云冷杉好,这也主要是与岳桦所处的位置有关,并且岳桦中也以中等年龄的树木对气候变化最为敏感。
|
| [30] |
b). Climatic control on forests and tree species distribution in the forest region of Northeast China. https://doi.org/10.1111/j.1744-7909.2006.00294.x URL |
| [31] |
An overview of China climate change over the 20th century using UK UEA/CRU high resolution grid data. 英国CRU高分辨率格点资料揭示的20世纪中国气候变化 .https://doi.org/10.3878/j.issn.1006-9895.2006.05.18 URL Magsci [本文引用: 1] 摘要
中国覆盖比较完整的台站观测始于1951年,1951年之前虽然有一些观测记录,但是残缺不全.所以要建立更长的气候序列就要吸收代用资料,但是代用资料可能与气候要素仅有一定程度的相关,不可能一一对应,因此应用代用资料重建的气候序列有一定的不确定性.英国East Anglia大学的Climatic Research Unit(简称CRU)通过整合已有的若干个知名数据库,重建了一套覆盖完整、高分辨率、且无缺测的月平均地表气候要素数据集,时间范围覆盖1901~2003年,空间为0.5°×0.5°经纬网格覆盖所有陆地.这套资料和中国已有的气候数据相比具有如下优点: 第一,中国西部20世纪前半期非常缺少观测,CRU资料尽管包含插值带来的误差,经比较仍可作为有一定信度的参考; 第二,中国现有的百年温度序列只是年或季分辨率,而CRU资料达到月分辨率; 第三,建立这个序列仅使用观测结果,做统计内插,不包括代用资料所带来的不确定性.因此,CRU的序列与用代用资料补充得到的序列在资料方面有较大不同,比较这两个序列,不仅可以进一步确认中国气候变化的特征,也可以彼此校正.结果表明:(1)CRU资料反映的全国年平均温度年际变化和考虑代用资料重建的序列吻合得很好,相关系数达到0.84;(2)区域尺度上,两者在10个典型分区的气温变率也相当一致,相关整体保持在0.8左右,仅新疆西南部和西藏西北部两者差异较大;(3)CRU资料揭示的中国年总降水量在1951~2000年的变化与160站观测吻合,相关系数达到0.93;(4)CRU资料的中国东部四季降水量和重建资料十分一致,秋季一致性最好,相关0.93;(5)CRU资料和重建的序列比较一致地表现出中国温度和降水年代际变化的主要特征,其给出的20世纪20年代中国大旱和20世纪40年代中国高温的空间分布与作者过去的结论相一致.这表明,作者过去重建的中国气候序列有比较大的可靠性,而CRU资料也提供了新的信息,特别是在20世纪前半期和中国西部.
|
| [32] |
Litter Fall Production and Decomposition in Relation to Climate, Biological and Local Environments Factors in Forests of Northeast China. Master degree dissertation, Beijing Forestry University, Beijing. (in Chinese)东北地区森林凋落物产量、分解速率及其与气候、生物因素、局域环境因子的关系 .
|
| [33] |
Response of broad-leaved Korean pine forest productivity of Mt. Changbai to climate change: An analysis based on BIOME-BGC Modeling. 长白山阔叶红松林净初级生产力对气候变化的响应: 基于BIOME-BGC模型的分析 . |
| [34] |
Dendroclimatic analysis of Betula ermanii forests at their upper limit of distribution in Changbai Mountain, Northeast China. |
| [35] |
Alpine timberline ecotone tree growth in relation to climatic variability for picea crassifolia forests in the middle Qilian Mountains, northwestern China. 祁连山中段青海云杉高山林线交错区树轮宽度与气候变化的关系 .URL 摘要
通过对祁连山中段青海云杉高山林线交错区进行取样,研究了交错区内4个不同海拔树木生长与气候变化的关系。结果表明:在林线交错区内,树木生长对气候变化的敏感性不高,年表统计特征沿海拔并没有一致的变化趋势。气候对树木生长的影响表现出非常明显的“生长滞后效应”,所有样地的轮宽指数与上一年10月的降水显著正相关,多数样地的轮宽指数与上一年7、8月的温度显著负相关,后者体现了有效水分供应的限制作用。同时,多数样地的轮宽指数与当年2月降水显著负相关。整个林线交错区的树轮生长主要受生长季之前(上一生长季和秋冬季)的气候指标的影响,而与当年生长季气候没有明显的联系。本文和相关研究发现的林线树木生长对气候变化并不敏感的现象,与高山林线主要受生长季温度限制的传统认识明显不同。
|
| [36] |
|

/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}