版权声明: 2016 植物生态学报编辑部 本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。
基金资助:
展开
摘要
以分布在海南岛西部(霸王岭国家级自然保护区, 21个样方)、西南部(尖峰岭国家级自然保护区, 12个样方)和中部(黎母山省级自然保护区, 15个样方)的热带云雾林为研究对象, 研究α及β物种多样性、功能多样性、谱系多样性的变化, 为植物多样性的保护提供科学依据。结果表明: 尖峰岭群落树木个体多度、物种丰富度最大, 黎母山群落树木个体多度、物种丰富度最小; 黎母山群落间物种组成差异最大, 霸王岭群落间物种组成差异最小, 海南岛霸王岭(西部)、尖峰岭(西南部)和黎母山(中部) 3个林区热带云雾林物种多样性差异可能与空气温度和相对湿度有关。尖峰岭群落内功能丰富度、Rao’s二次熵最低, 功能均匀度最高, 群落间平均成对性状距离最小, 反映群落构建主要受环境筛影响; 霸王岭群落Rao’s二次熵最高, 功能均匀度最低, 群落间平均成对性状距离最大, 反映群落构建主要受限制相似性影响; 黎母山群落内功能丰富度最高, 群落间平均最近性状距离最大, 表明限制相似性在黎母山热带云雾林群落构建中的作用更重要。霸王岭群落内谱系多样性、物种间平均最近相邻谱系距离均较大, 反映物种间谱系关系趋于发散; 黎母山群落内谱系多样性、物种间平均成对谱系距离及群落间平均成对谱系距离均最小, 反映物种间谱系关系趋于聚集; 而尖峰岭群落内物种间平均成对谱系距离、群落间平均成对谱系距离最大, 但物种间平均最近相邻谱系距离最小, 反映物种间谱系关系既具有发散又具有聚集的共存格局。因此, 海南岛热带云雾林群落植物多样性变化格局与环境及物种间相互作用有关。
关键词:
Abstract
We explored α and β species diversity, functional diversity and phylogenetic diversity distribution patterns in three tropical cloud forests along environmental gradients in air temperature and precipitation.
We sampled plots in three tropical cloud forests which are located in the west (Bawangling, 21 plots, BWL), the southwest (Jianfengling, 12 plots, JFL), and the central of Hainan Island (Limushan, 15 plots, LMS). We collected species data and functional trait data including plant height, specific leaf area, chlorophyll content, leaf thickness and wood density. We assessed the differences within- and among-community species diversity, functional diversity and phylogenetic diversity in these three tropical cloud forests using the Kruskal-Wallis test.
The tropical cloud forests in JFL had the highest species abundance and richness whereas the lowest in LMS. However, the Bray-Curtis and Jaccard dissimilarity coefficients showed the opposite distribution patterns (i.e. the highest in LMS whereas the lowest in BWL). Distinct distribution patterns in species diversity across the three tropical cloud forests may be explained by the air temperature and relative humidity. The functional evenness (FEve) within communities was the highest while functional richness (FRic), Rao’s quadratic entropy (RaoQ) and the mean pairwise trait distance among communities were the lowest in JFL, indicating that habitat filtering plays an important role in community assembly. BWL had the highest RaoQ and mean pairwise trait distance among communities, and the lowest FEve, which demonstrated that limiting similarity would be prevalent in forest communities assembled. LMS had the highest FRic within communities and mean nearest trait distance among communities, leading to a limiting similarity influencing forest communities. BWL had the highest Faith phylogenetic diversity (PD) within communities and mean nearest phylogenetic distance among communities, reflecting an overdispersed pattern in phylogenetic structures. LMS had the lowest PD and mean pairwise phylogenetic distance within and among communities, suggesting that a clustered pattern in phylogenetic structures. The mean pairwise phylogenetic distance within and across communities were the highest in JFL while the mean nearest phylogenetic distance within communities was the lowest, indicating that phylogenetic clustering and overdispersion patterns co-occur in this forest. We conclude that both plant species interactions and environmental filtering determine the distribution patterns of plant species diversity, functional diversity and phylogenetic diversity both within and among three tropical cloud forests in Hainan Island.
Keywords:
植物多样性是植物与其他生物及环境相互作用形成的生态组合及与此有关的各种生态过程。它是植物长期进化的结果, 与生态系统功能关系紧密(范娟, 2015; Venail et al., 2015), 也是人类赖以生存的基础。对物种多样性、功能多样性和谱系多样性的综合研究有助于更好地理解植物多样性的变化规律(Butterfield et al., 2013; Spasojevic et al., 2014), 为植物多样性的保护提供科学依据。
物种多样性是植物多样性最直观的表现, α物种多样性是对群落内物种数量及分布均匀度的测量, 体现了群落内物种通过资源竞争而形成的共存关系。β物种多样性对解释生态系统功能、生物多样性保护及生态系统管理有重大意义(Legendre et al., 2005)。物种多样性的变化格局与海拔梯度(Swenson et al., 2011)、演替阶段(卜文圣, 2013)、森林类型(薛倩妮, 2015)、生境类型(范娟, 2015; Pereira et al., 2007)等密切相关。功能多样性通常指影响生态系统功能的物种或有机体性状的数值和范围(Mason & de Bello, 2013)。已有研究表明功能多样性比物种多样性能更好地预测生态系统过程及其功能(Cadotte et al., 2011), 且功能多样性指数有揭示群落形成过程的潜能(Mouchet et al., 2010)。α和β 功能多样性用于解释群落内和群落间物种功能相似性与物种分布的关系(Mason & de Bello, 2013), 揭示空间尺度和环境异质性对群落构建的各个生态过程的影响(Swenson, 2013)。例如Swenson等(2012)用α和β多样性分析温带和热带森林群落构建, 发现样方内及样方间功能性状都表现出聚集分布, 揭示了环境筛同时作用于群落内和群落间物种共存。谱系多样性强调系统进化多样性并能反映生物的亲缘关系特征, 能从进化角度研究群落物种组成现状和成因, 因而能解释生物多样性分布格局(Rosauer et al., 2009)。受物种库大小和环境异质性的影响, α谱系多样性和β谱系多样性分别提供了群落内和群落间物种亲缘关系信息, 揭示不同空间尺度和环境梯度下物种间谱系相似性与群落构建的关系(Butterfield et al., 2013)。
热带云雾林指潮湿热带地区经常被云雾掩盖的森林(Stadtmüller, 1987), 主要分布在热带美洲、热带非洲和热带亚洲地区海拔500-3900 m的山顶或山脊(Bubb et al., 2004)。海南岛的热带山顶矮林是典型的热带云雾林(龙文兴等, 2011b), 归属于阔叶林植被型亚纲山地常绿苔藓林植被型组(吴征镒, 1995)。热带云雾林内山风强烈、气温低、云雾多, 土壤含水量常处于饱和状态(龙文兴等, 2011b); 植物对一些环境因子(如温度和降水等)的变化十分敏感, 因而植物多样性可能有独特的分布格局。龙文兴等对海南霸王岭地区热带云雾林群落的环境特征(龙文兴等, 2011b)、结构特征(龙文兴等, 2011a)、植物功能性状分异规律(Long et al., 2011a, 2011b)及群落构建规律(Long et al., 2013; Long et al., 2015)做过初步研究。本研究以海南岛西部(霸王岭国家级自然保护区)、西南部(尖峰岭国家级自然保护区)和中部(黎母山省级自然保护区)的热带云雾林为研究对象, 基于群落物种数据、5类重要功能性状以及系统发育进化树, 研究α及β物种多样性、功能多样性和谱系多样性变化, 阐明海南岛不同林区热带云雾林植物多样性变化规律, 为森林植物多样性保护及管理提供借鉴。
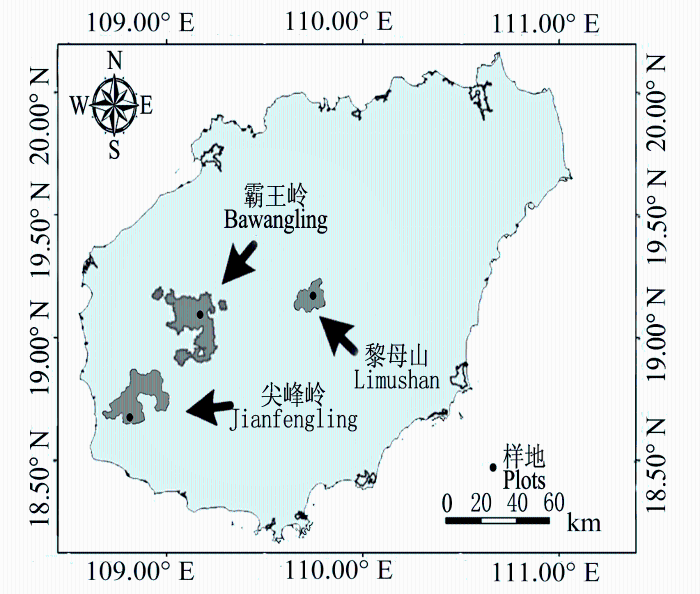
海南岛地处18.14°-20.02° N, 108.62°-111.05° E, 属于热带北缘。地形中部高四周低, 降雨东部多西部少, 南部温度比北部高; 西南和西部受西南季风影响, 干季更显著; 中部山区云雾较多、温度较低。海南岛热带云雾林主要分布在霸王岭、黎母山、五指山、尖峰岭等林区海拔1200 m以上的山顶(胡玉佳和李玉杏, 1992)。本研究主要以气象因子(如水分梯度和温度梯度)为环境因子轴, 分别在海南岛西部(霸王岭国家级自然保护区)、西南部(尖峰岭国家级自然保护区)和中部(黎母山省级自然保护区)的典型热带云雾林——热带山顶矮林分布区设置样地(图1), 样地概况如表1所示。
表1 样地概况
Table 1 Information of the study sites
| 研究样地 Study site | 海拔 Elevation (m) | 经度 Longitude (E) | 纬度 Latitude (N) | 坡度 Slope (°) | 样方数 No. of plots | 样方面积 Plot area (m2) | 优势种 Dominant species |
|---|---|---|---|---|---|---|---|
| 尖峰岭 Jianfengling | 1 187.17-1 397.19 | 108.87° | 18.72° | 10-65 | 12 | 4 800 | 罗浮锥 Castanopsis fabri, 丛花厚壳桂 Cryptocarya densiflora, 美丽新木姜子 Neolitsea pulchella, 黄叶树 Xanthophyllum hainanense |
| 霸王岭 Bawangling | 1 313.24-1 385.24 | 109.21° | 19.08° | 2-45 | 21 | 8 400 | 蚊母树 Distylium racemosum, 赤楠 Syzygium buxifolium, 九节 Psychotria asiatica, 黄杞 Engelhardia rox- burghiana |
| 黎母山 Limushan | 1 363.73-1 403.32 | 109.76° | 19.18° | 3-42 | 15 | 6 000 | 野茶 Camellia sinensis var. assamica, 岭南青冈 Cyclob- alanopsis championii, 罗浮锥 Castanopsis fabri, 细枝柃 Eurya loquaiana |
1.2.1 群落物种调查
2013-2014年, 用徕卡全站仪(Leica TPS1200+, Leica Geosystems, Heerbrugg, Switzerland)在霸王岭、尖峰岭和黎母山热带云雾林分布区分别设置21、12、15个20 m × 20 m的样方。样方间距离50 m以上, 用邻格法将每个400 m2的样方划分为16个5 m × 5 m的小样方。调查小样方内所有胸径≥1 cm植株的物种名、胸径和高度。根据《中国植物志》确定个体物种名, 现场无法确认的物种制成标本请专家鉴定。调查植株: 尖峰岭6879株, 235种; 霸王岭9714株, 117种; 黎母山3809株, 148种。
1.2.2 植物功能性状的选择和测定
功能性状的选择原则是: 反映植物重要生活史过程, 代表植物生态策略; 对环境因子变化响应迅速, 并对生物地球化学循环等生态功能产生影响; 容易测定但具有重要生态学意义。选取的5个功能性状包括2个主茎特征(木材密度(WD, g·cm-3)、植株高度(H, m))和3个叶片特征(叶片厚度(LTh, mm)、比叶面积(SLA, cm2·g-1)、叶绿素含量(Chl, SPAD))。
在每个20 m × 20 m的样方中, 测定所有胸径≥5 cm的植株(包括乔灌木和藤本)的功能性状。植株高度是用测高仪测量的树干基部到树冠的最长距离。每个植物个体采集2-3片成熟的、受太阳光直射的、健康的冠层叶片(Cornelissen et al., 2003), 叶片厚 度用数显游标卡尺(SF2000, Guilin, China)测量; 叶绿素含量用便携式叶绿素仪(SPAD-502 Plus,Konica Minolta Holdings, Tokyo, Japan)测量; 叶面积用叶面积仪(LI-COR 3100C Area Meter, LI-COR, Lincoln, USA)测定。然后将叶片装入信封, 放于80 ℃烘箱烘至恒定质量, 用电子天平(AR2140, Ohaus, Parsippany, USA)称量干质量。比叶面积是叶面积与干质量的比值。
在采集叶片样品的同时, 从每个植株上截取4根直径为2-3 cm、长约10 cm的枝条, 去皮后用排水法测量枝条体积。然后将枝条装入信封放于80 ℃烘箱烘至恒定质量, 称量其干质量, 根据枝条干质量与体积之比计算出木材密度。研究发现热带森林树木枝条密度与木材密度显著相关(Bu et al., 2014), 所以可用植物枝条密度表示木材密度, 这样可避免用生长锥钻取年轮条对树木的伤害。
1.3.1 物种多样性
用树木个体多度和物种丰富度表示20 m × 20 m样方的α物种多样性。采用改良后的Bray-Curtis相异性指数(Bray & Curtis, 1957)、Jaccard相异性指数(Jaccard, 1912)计算β物种多样性。二者的计算均基于数量数据, 具体计算由R 3.2.2 (https://cran.r-pro- ject.org) vegan软件包中的vegdist函数完成。
1.3.2 功能多样性
选择3个相互独立的功能多样性指数来计算α功能多样性(Mouchet et al., 2010): 功能丰富度(FRic)、功能均匀度(FEve)和Rao’s二次熵(RaoQ), 具体计算由R 3.2.2 FD软件包中的dbFD函数完成。
采用平均成对性状距离(Dpw)和平均最近相邻性状距离(Dnn)度量群落间β功能多样性, Dpw、Dnn分别由R 3.2.2 picante软件包中的comdist、comdistnt函数完成。
1.3.3 谱系树的建立及谱系多样性
Qian和Jin (2015)在Zanne等(2014)的谱系树基础上进一步更新、扩展建立了PhytoPhylo谱系树。在Qian和Jin (2015)的谱系树基础上, 基于样地物种数据, 用S. PhyloMaker函数建立了本文研究的谱系树, 结果会输出以不同方式添加物种的3种谱系树(e.g., Scenario 1, Scenario 2, Scenario 3)。我们选用第三种谱系树(Scenario 3), 该谱系树将PhytoPhylo中没有的物种以多分枝形态添加到其父母分类单元下, 用BLADJ算法为其分配枝长(Webb et al., 2008; Qian & Jin, 2015)。
用种间平均成对谱系距离(MPD)、平均最近相邻谱系距离(MNND)和Faith谱系多样性(PD)(Faith, 1992)表示α谱系多样性。PD是某一样地中分类单元谱系分支长度占谱系中所有分支长度之和的比例, 由Phylocom 4.2 (Webb et al., 2008)里的 pd 计算; MPD、MNND分别由R 3.2.2 picante软件包中的mpd、mntd函数完成。
β谱系多样性描述的是同一时间两个处于不同地理位置上的群落间的谱系距离, 可反映群落间亲缘关系的相异性(Graham & Fine, 2008)。采用平均成对谱系距离(Dpw)和平均最近相邻谱系距离(Dnn)度量群落间β谱系多样性。
1.3.4 基于功能性状的谱系信号检验
结合系统发育进化树, 用Blomberg等(2003)提出的基于Brownian motion进化模型的K值法来检测谱系信号的强弱, 分析功能性状与物种进化历史的相关性。K值是类群间性状差异偏离于随机过程的度量, 计算方法如下: K = 1表示功能性状沿谱系树随机变化, 符合布朗运动进化模型(Brownian motion model), K > 1表示功能性状表现出比按布朗运动模型方式进化更强的谱系信号, 而K < 1表示进化更弱的谱系信号。谱系信号的显著性可通过比较功能性状在整个谱系树中标准化独立差的方差观测值和零模型的随机化检验得到。具体计算由phytools包中的phylosig函数完成。
1.3.5 数据及其标准化处理
现有研究多用不同零模型对MPD、MNND、Dpw及Dnn进行标准化处理, 以排除群落间物种数不同所造成的影响, 但标准化的观察值会影响生境过滤和扩散限制过程对群落谱系结构的判断。比如随机化谱系树上物种的零模型虽然消除了不同物种数对MPD、MNND、Dpw及Dnn的影响, 但该零模型也限制了物种的空间扩散, 因此也消除了扩散限制对MPD、MNND、Dpw和Dnn的影响(Swenson et al., 2012)。因而本文不采用零模型对MPD、MNND、Dpw和Dnn标准化。功能多样性、谱系多样性的计算均采用多度加权。
群落的总体植物多样性分析用每个林区4800 m2样地面积的数据。热带云雾林群落内及群落间植物多样性比较用每个林区20 m × 20 m样方数据, 用Kruskal-Wallis检验比较尖峰岭、霸王岭、黎母山间的植物多样性差异; 该函数还可进行林区间的多重比较, 用Bonferroni法进行p值调整。所有数据统计分析及作图由R 3.2.2软件(R Development Core Team, 2015)完成。
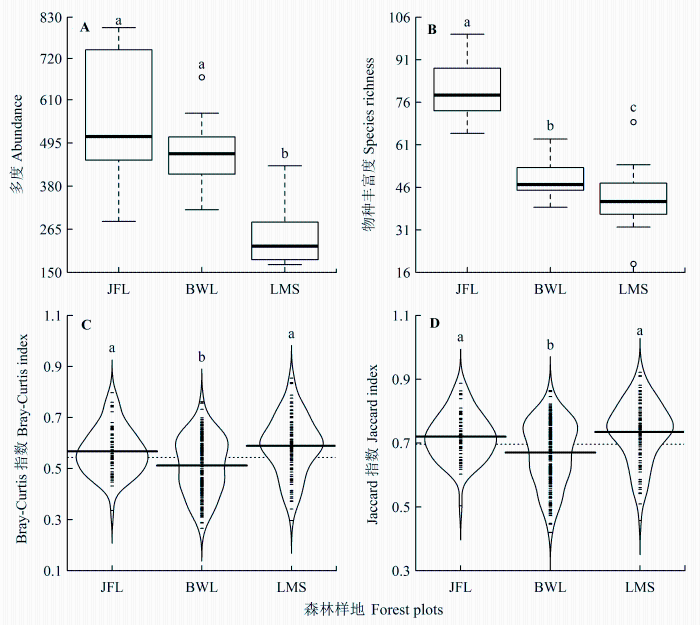
在4800 m2样方中, 尖峰岭、霸王岭、黎母山的多度分别为6879、5225和2951, 丰富度分别为235、99和136。20 m × 20 m样方中, 尖峰岭、霸王岭、黎母山热带云雾林树木个体多度、物种丰富度均有显著差异(p < 0.001)。尖峰岭的树木个体多度和丰富度最高, 黎母山的树木个体多度和丰富度最低(图2 A、2B)。
在4800 m2样方中, 尖峰岭与霸王岭、黎母山间的Bray-Curtis指数分别为0.73和0.77, Jaccard指数分别为0.84和0.87; 霸王岭与黎母山间的Bray-Curtis指数为0.88, Jaccard指数为0.94。20 m × 20 m样方中, 尖峰岭、霸王岭、黎母山热带云雾林Bray-Curtis相异性指数、Jaccard相异性指数均有显著差异(p < 0.001)。黎母山与尖峰岭的Bray-Curtis相异性指数、Jaccard相异性指数差异不显著, 但都显著高于霸王岭(图2 C、2D)。
图2 尖峰岭(JFL)、霸王岭(BWL)和黎母山(LMS)热带云雾林α (A、B)及β (C、D)物种多样性比较。图中不同小写字母表示样地间有显著差异(p < 0.05)。粗实线表示中位数, 虚线表示整体平均值。
Fig. 2 Comparisons in α (panel A and B) and β (panel C and D) species diversity among Jianfengling (JFL), Bawangling (BWL) and Limushan (LMS). Different low-case letters indicate significant differences among plots (p < 0.05). Heavy horizontal lines indicate median values and dashed horizontal lines indicate mean values.
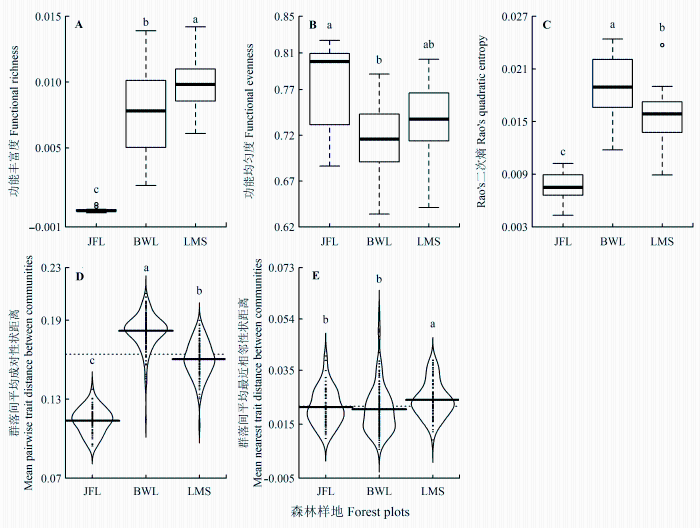
尖峰岭、霸王岭、黎母山热带云雾林总体群落FRic分别为0.00017、0.00004、0.00015; FEve分别为0.58、0.54、0.64; Rao’s二次熵分别为0.007、0.011、0.008。20 m × 20 m样方中, 尖峰岭、霸王岭、黎母山热带云雾林群落的FRic (p < 0.001)、FEve (p < 0.05)、Rao’s二次熵(p < 0.001)均差异显著。黎母山热带云雾林的FRic最高, 尖峰岭最低(图3A);尖峰岭热带云雾林的FEve最高, 霸王岭最低(图3 B); 霸王岭热带云雾林的Rao’s二次熵最大, 尖峰岭最小(图3C)。
图3 尖峰岭(JFL)、霸王岭(BWL)和黎母山(LMS)热带云雾林间α (A、B、C)及β (D、E)功能多样性比较。图中不同小写字母表示样地间有显著差异(p < 0.05)。粗实线表示中位数, 虚线表示整体平均值。
Fig. 3 Comparisons in α (A, B, C) and β (D, E) functional diversity am- ong Jianfengling (JFL), Bawangling (BWL) and Limushan (LMS). Different low-case letters indicate significant differences among plots (p < 0.05). Heavy horizontal lines indicate median values and dashed horizontal lines indicate mean values.
分析总体β功能多样性, 尖峰岭与霸王岭、黎母山的Dpw分别为0.14、0.13, Dnn分别为0.03和0.02; 霸王岭与黎母山Dpw为0.15, Dnn为0.04。20 m × 20 m样方中, 尖峰岭、霸王岭、黎母山热带云雾林群落间Dpw、Dnn均差异显著(p < 0.001)。霸王岭热带云雾林Dpw最大, 尖峰岭最小(图3D), 尖峰岭和霸王岭的Dnn无显著差异, 但都显著低于黎母山(图3E)。
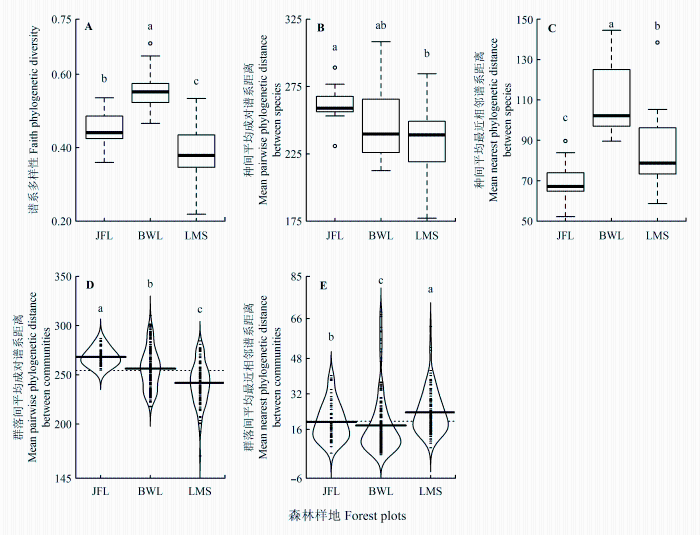
尖峰岭、霸王岭、黎母山热带云雾林总体PD为0.76、0.46、0.56, MPD为264.88、236.70、239.68, MNND为48.56、81.48、50.35。20 m × 20 m样方中, 尖峰岭、霸王岭、黎母山热带云雾林群落的PD (p < 0.001)、MPD (p < 0.05)、MNND (p < 0.001)均差异显著。霸王岭热带云雾林群落的PD最大, 黎母山最小(图4 A)。尖峰岭、黎母山均与霸王岭的MPD无显著差异, 尖峰岭显著高于黎母山(图4B)。霸王岭热带云雾林群落的MNND最大, 尖峰岭最小(图4C)。
图4 尖峰岭(JFL)、霸王岭(BWL)和黎母山(LMS)热带云雾林间α(A、B、C)及β(D、E)谱系多样性比较。图中不同小写字母表示样地间有显著差异(p < 0.05)。粗实线表示中位数,虚线表示整体平均值。
Fig. 4 Comparisons in α (A, B, C) and β (D, E) functional diversity among Jianfengling (JFL), Bawang- ling (BWL) and Limushan (LMS). Different low-case letters indicate significant differences among plots (p < 0.05). Heavy horizontal lines indicate median values and dashed horizontal lines indicate mean values.
尖峰岭与霸王岭、黎母山热带云雾林总体群落的Dpw分别为267.89、261.75, Dnn分别为33.34、20.35; 尖峰岭与黎母山热带云雾林总体Dpw为257.69, Dnn为47.69。20 m × 20 m样方中, 尖峰岭、霸王岭、黎母山热带云雾林群落间的Dpw、Dnn均存在显著差异(p < 0.001)。尖峰岭的Dpw最大, 黎母山最小(图4D)。黎母山Dnn最大, 霸王岭最小(图4E)。
霸王岭样地中只有叶绿素含量、叶厚度、木材密度检测到显著的谱系信号(p < 0.05; 表2), 黎母山样地中植株高度、比叶面积、叶绿素含量、叶厚度、木材密度5个功能性状均检测出显著的谱系信号(p < 0.05), 而尖峰岭样地中5个功能性状的谱系信号均不显著。
表2 霸王岭、尖峰岭和黎母山热带云雾林植物功能性状的谱系信号
Table 2 Phylogenetic signal in functional traits in tropical cloud forests in Bawangling, Jianfengling and Limushan
| 性状 Trait | 霸王岭 Bawangling | 黎母山 Limushan | 尖峰岭 Jianfengling | |||||
|---|---|---|---|---|---|---|---|---|
| K | p | K | p | K | p | |||
| 植株高度 Plant height (H) | 0.228 | 0.114 | 0.068 | 0.014 | 0.050 | 0.162 | ||
| 比叶面积 Specific leaf area (SLA) | 0.294 | 0.059 | 0.098 | 0.006 | 0.037 | 0.282 | ||
| 叶绿素含量 Chlorophyll content (Chl) | 0.223 | 0.042 | 0.225 | 0.001 | 0.053 | 0.073 | ||
| 叶厚度 Leaf thickness (LTh) | 0.255 | 0.048 | 0.141 | 0.001 | 0.044 | 0.151 | ||
| 木材密度 Wood density (WD) | 0.358 | 0.004 | 0.095 | 0.014 | 0.023 | 0.505 | ||
树木个体多度、物种丰富度整体变化格局是尖峰岭最大, 黎母山最小。这可能与3个样地的环境条件差异有关。由于地理位置差异, 尖峰岭(海南岛西南部)平均气温最高, 霸王岭(海南岛西部)次之, 黎母山最低(海南岛中部), 热带云雾林植物往往受空气低温胁迫影响(Long et al., 2011b), 因而尖峰岭热带云雾林群落温度胁迫可能最弱, 黎母山最强, 温度影响3个样地的物种多样性分布格局。3个林区热带云雾林群落中, 尖峰岭样地的平均海拔最低, 坡度最大(表1), 黎母山样地的平均海拔最大, 坡度最小。因而尖峰岭热带云雾林群落地形变化较大, 生境异质性强, 物种多样性最大。本文结果与Pereira 等(2007)的研究结果一致, 物种多样性随生境异质性的增加而增加。
本文研究群落β多样性时没有采用基于二元数据的多样性指数, 而是选择了考虑每一物种个体数量的多样性指数Bray-Curtis指数、Jaccard指数(马克平等, 1995)。基于数量数据的β多样性指数, 明确了常见种和偶见种对β多样性的贡献程度, 结果更加准确。三个林区间Bray-Curtis指数、Jaccard指数有显著差异。黎母山地处海南中部山区, 空气湿度相对较大, 热带云雾林中云雾降水较丰富, 样方间空气相对湿度差异较大, 因而样方间物种组成差异较大; 而尖峰岭和霸王岭地处海南西南部和西部地区(五指山雨影区), 又受到西南季风的影响, 空气湿度相对较小, 不同样方间空气湿度较为一致, 因而样方间物种组成差异较小。
功能丰富度指数主要揭示群落中有多少生态位空间被物种占据, 既取决于物种所占据的功能生态位, 也取决于功能特征值的范围(Mouchet et al., 2010); 功能丰富度低意味着群落内有生态空间未被有效利用, 以致生产力低下(范娟, 2015)。功能均匀度指数主要衡量群落内资源的利用效率, 该值高说明资源利用充分、均匀, 该值低则表明有资源过度利用, 而其他资源未被利用或利用得少(Schleuter et al., 2010)。Rao’s二次熵综合了功能丰富度和功能分散度信息, 该值高往往表明群落受到限制相似性的影响, 该值低则表明环境筛对群落的影响更大。尖峰岭热带云雾林群落功能丰富度、Rao’s二次熵最低, 而功能均匀度最高, 说明尖峰岭热带云雾林群落受环境筛的影响更大一些, 使群落中物种具有相近的功能性状(Lohbeck et al., 2012)。有研究认为, 随着物种丰富度的增加, 功能丰富度也增大(Mou- chet et al., 2010), 但尖峰岭热带云雾林群落物种丰富度最高, 功能丰富度最低, 这可能是因为海拔和坡度对功能丰富度有一定的影响(范娟, 2015), 尖峰岭样地海拔、坡度变化大, 生境异质性强, 有利于物种多样性的增加(Pereira et al., 2007), 但由于环境筛的限制作用, 使群落中具有相同适合度的物种稳定共存(Cornwell et al., 2006)。Rao’s二次熵在尖峰岭最小也间接验证了这一点。霸王岭热带云雾林群落Rao’s二次熵最高, 而功能均匀度最低, 说明限制相似性对霸王岭热带云雾林群落起作用, 共存的物种在资源利用策略上生态位高度分化(Mason et al., 2012)。黎母山热带云雾林群落物种丰富度最低, 但不同样方间的物种组成差异最大, 功能丰富度也最高, 说明该群落生态空间利用率高, 物种间生态位分化程度高, 限制相似性作用于群落共存(Mason et al., 2012)。
霸王岭热带云雾林的Dpw最大, 尖峰岭最小; 黎母山热带云雾林的Dnn最大, 霸王岭最小, 与Dpw呈现出不一样的变化格局。Dpw指的是两个群落不同物种或个体间的平均功能性状距离, 主要揭示不同群落物种在整个功能性状树上的聚集和发散程度, 而Dnn计算的是一个群落中的物种与另一群落中与其功能性状距离最近的物种间的平均功能性状距离, 着重说明不同群落物种在功能性状树的某一末端分枝上的局部聚集和发散程度(Webb et al., 2008)。尖峰岭热带云雾林群落Dpw最小, 这可能是由于生境过滤的作用, 使尖峰岭热带云雾林群落的功能性状呈现出趋同性。Swenson等(2012)曾提出生境过滤作用会使同一生境内群落间的功能性状差异较小, 即β功能多样性较小。霸王岭热带云雾林群落的Dpw最大, 而Dnn最小, 这可能与群落间物种多度分布存在较大差异有关(Swenson et al., 2012), 例如当群落1中有51个物种A、3个物种B, 而群落2中有4个物种A、48个物种B时, 这种物种多度格局将会使Dpw较大, 但并不存在Dnn差异。黎母山热带云雾林群落的Dnn最大, 可能是由于该群落样方间物种组成差异最大, 又受到限制相似性的影响。
各谱系多样性指数能反映不同的群落结构特征。MPD是群落中全部物种对间的平均进化距离, 可以很好地表示群落中物种的整体聚集程度(Webb, 2000), 是描述群落谱系多样性的基础指标。MNND是任一物种与群落中和自己亲缘关系最近的一个物种间的平均进化距离, 反映群落中物种的局部聚集程度。Dpw着重强调不同分类单元在整个谱系树上的聚集程度, 反映2个群落靠近谱系树根节点分枝的差异, 从而可以更好地揭示群落生境的差异。Dnn则反映不同分类单元在某一特定分枝上的局部聚集程度, 着重强调2个群落进化特异性上的差异, 揭示群落对资源利用策略的差异(Feng et al., 2012)。
本研究发现, 尖峰岭的MPD和Dpw最大, 反映群落中物种间亲缘关系较远, 群落谱系结构相对发散。其可能原因是尖峰岭热带云雾林群落海拔、坡度变化大, 生境异质性强, 受种间竞争作用的影响, 导致谱系结构相对发散, 物种倾向于采取差异化的资源利用策略(Devictor et al., 2010; Helmus & Ives, 2012; 王薇等, 2014); 但是尖峰岭热带云雾林群落物种间MNND较小, 反映了谱系结构相对聚集。其可能原因是尖峰岭热带云雾林群落虽然物种丰富度较高, 但由于环境筛的作用, 一些物种有较近的亲缘关系, 在谱系树上的进化枝长较小; 同时尖峰岭群落样方间生境差异大, 有利于样方内更相似的物种共存(Webb, 2000), 因此尖峰岭热带云雾林群落的谱系结构既有发散特征又有聚集特征。
霸王岭热带云雾林物种间MNND较大, 说明物种间谱系关系较为发散; 但PD最大, 与物种丰富度的变化格局并不完全一致, 说明物种丰富度大的群落其谱系多样性不一定大。其可能原因是霸王岭热带云雾林物种间竞争作用较强(Long et al., 2013), 生态位高度分化使物种间亲缘关系较远, 进化枝长较大, 群落谱系较发散(Spasojevic et al., 2014)。该特征也能通过群落间的Dnn在3个地方云雾林中最小的特征反映出来, 说明受生境异质性影响, 群落内部个体之间存在趋同的生态策略。
黎母山的PD、MPD及Dpw均是最小的, 可能由于黎母山热带云雾林群落海拔相对较高, 气温低、山风强烈、空气湿度相对较大, 环境相对恶劣, 在环境筛(如低温)的作用下, 群落谱系结构呈现出聚集的状态(Helmus & Ives, 2012)。黎母山的PD分布格局可能与其物种丰富度相关, 即较少物种组成群落的PD也较小, 而Dnn所反映的特征不明显。
黎母山热带云雾林植株高度、比叶面积、叶绿素含量、叶厚度、木材密度及霸王岭热带云雾林植物叶绿素含量、叶厚度、木材密度等均检测到显著的谱系信号, 这说明物种进化历史对两个林区热带云雾林植物的功能性状有影响。杨洁等(2014)在哀牢山亚热带中山湿性常绿阔叶林中检测到比叶面积、叶绿素含量、叶厚度等都具有显著谱系信号,我们的研究结果与之一致。但植物功能性状K值都小于1, 比布朗模型的预期值低(表2), 且K值都较小, 因而环境差异对功能性状的影响较大。
本研究中, β功能多样性与β谱系多样性的变化格局并不完全一致, 可能是因为3个热带云雾林群落功能性状的谱系信号都较弱(K < 1)。有研究者指出功能性状谱系信号较弱时, 生态位过程和中性过程共同作用于群落构建, 群落的谱系结构和功能性状会呈现出相反的变化格局(Swenson et al., 2012), 因此单独用谱系结构或功能性状来推断群落构建和物种共存的成因可能会导致错误的结论(Swen- son, 2013; 杨洁等; 2014)。部分功能性状并不能完全代表物种的实际生态位(Swenson, 2013), 本文仅选取了5个功能性状, 这可能也导致了谱系结构和功能性状的变化格局不完全一致。因此, 为准确推测不同生态学过程对群落构建的影响, 需同时考虑物种的谱系结构和重要功能性状信息。
总之, 海南岛3个林区物种多样性的空间变化格局与环境条件差异有关; 环境筛影响尖峰岭热带云雾林群落的功能多样性, 而限制相似性对霸王岭和黎母山热带云雾林群落的功能多样性影响更大; 霸王岭群落物种间谱系关系相对更为发散, 黎母山群落物种间谱系关系相对更为聚集, 尖峰岭则存在发散和聚集两种格局; 谱系结构和功能性状的变化格局不完全一致, 在以后的研究中需综合考虑谱系结构和功能性状对群落构建的影响。我们的研究表明海南岛热带云雾林群落植物多样性变化格局可能与环境及物种间相互作用有关。
致谢 本研究受到海南大学青年基金(qnjj1210)资助, 特此致谢。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
| [1] |
Testing for phylogenetic signal in comparative data: Behavioral traits are more labile. https://doi.org/10.1046/j.0306-5251.2001.01535.x URL PMID: 12778543 摘要
The primary rationale for the use of phylogenetically based statistical methods is that phylogenetic signal, the tendency for related species to resemble each other, is ubiquitous. Whether this assertion is true for a given trait in a given lineage is an empirical question, but general tools for detecting and quantifying phylogenetic signal are inadequately developed. We present new methods for continuous-valued characters that can be implemented with either phylogenetically independent contrasts or generalized least-squares models. First, a simple randomization procedure allows one to test the null hypothesis of no pattern of similarity among relatives. The test demonstrates correct Type I error rate at a nominal alpha = 0.05 and good power (0.8) for simulated datasets with 20 or more species. Second, we derive a descriptive statistic, K, which allows valid comparisons of the amount of phylogenetic signal across traits and trees. Third, we provide two biologically motivated branch-length transformations, one based on the Ornstein-Uhlenbeck (OU) model of stabilizing selection, the other based on a new model in which character evolution can accelerate or decelerate (ACDC) in rate (e.g., as may occur during or after an adaptive radiation). Maximum likelihood estimation of the OU (d) and ACDC (g) parameters can serve as tests for phylogenetic signal because an estimate of d or g near zero implies that a phylogeny with little hierarchical structure (a star) offers a good fit to the data. Transformations that improve the fit of a tree to comparative data will increase power to detect phylogenetic signal and may also be preferable for further comparative analyses, such as of correlated character evolution. Application of the methods to data from the literature revealed that, for trees with 20 or more species, 92% of traits exhibited significant phylogenetic signal (randomization test), including behavioral and ecological ones that are thought to be relatively evolutionarily malleable (e.g., highly adaptive) and/or subject to relatively strong environmental (nongenetic) effects or high levels of measurement error. Irrespective of sample size, most traits (but not body size, on average) showed less signal than expected given the topology, branch lengths, and a Brownian motion model of evolution (i.e., K was less than one), which may be attributed to adaptation and/or measurement error in the broad sense (including errors in estimates of phenotypes, branch lengths, and topology). Analysis of variance of log K for all 121 traits (from 35 trees) indicated that behavioral traits exhibit lower signal than body size, morphological, life-history, or physiological traits. In addition, physiological traits (corrected for body size) showed less signal than did body size itself. For trees with 20 or more species, the estimated OU (25% of traits) and/or ACDC (40%) transformation parameter differed significantly from both zero and unity, indicating that a hierarchical tree with less (or occasionally more) structure than the original better fit the data and so could be preferred for comparative analyses.
|
| [2] |
An ordination of the upland forest communities of southern Wisconsin. https://doi.org/10.2307/1942268 URL 摘要
Two aqueous suspensions, one containing crystals of uniform size and the other containing colloidal particles, were made with each of seven solid contact poisons (two DDT-analogues, DDT, rotenone, 2-bromomercurithiophen, dieldrin and endrin). The relative toxicity of each pair of suspensions was found in dipping or measured-drop tests on as many as possible of the species Oryzaephilus surinamennsis L., Tribolium castaneum Herbst and Tenebrio molitor L. In addition, some tests were made by injection of colloidal suspensions. In each test, insects were kept after treatment at two temperatures: 28 degrees C. and 11, 17 or 20 degrees C.; counts of kill were made after 24 hr. The tests measured three different kinds of temperature coefficient of insecticidal action. In the contact tests, the colloid was nearly always more toxic than the crystals. In all the tests, except those with dieldrin, the relative toxicity (colloid : crystals) was greater at the lower after-treatment temperature, i.e. the temperature coeficient of the relative toxicity was negative. But in the case of dieldrin, the coefficient was positive. The DDT-analogues, DDT, rotenone and endrin were more toxic at the lower after-treatment temperature, i.e. they had negative temperature coefficients of kill by contact action. In all these cases the temperature coefficient of kill by contact action was greater for colloid than for crystals. Dieldrin and 2-bromomercurithiophen had positive temperature coefficients of kill by contact action. With dieldrin, the temperature coefficient was greater for colloid than for crystals; but with 2-bromomercurithiophen, the reverse was true. The temperature coeficient of kill by injection was negative for DDT, but positive for dieldrin and endrin; the other poisons were not tested by injection. A possible explanation for the results of the contact tests is given in an Appendix. The explanation is based on a number of assumptions about the penetration of insect cuticle by solid poisons. These assumptions lead to the conclusion that the effect of temperature on the relative toxicity depends on the temperature coefficient of kill by internal action of the poison on the insect. This can be measured by injection tests. If it is negative or zero, the ratio of toxicities (colloid: crystals) by contact action, measured quite soon after treatment of the insects, will be greater at a low temperature after treatment than at a high temperature after treatment; but if the coefficient is positive, the effect of temperature on relative toxicity cannot be foretold. The experimental results seem to confirm the assumptions.
|
| [3] |
The Relationship Between Biodiversity and Ecosystem Functioning in Natural Tropical Forests of Hainan Island, China. PhD dissertation, 海南岛热带天然林生物多样性与生态系统功能关系的研究 .URL 摘要
全球生态系统正在遭遇剧烈的变化,从而加快了物种灭绝的速率。全 球生物多样性减少和丧失对生态系统功能的影响是当前生态学最为关注的领域之一。大量关于生物多样性与生态系统功能关系(BEF)的研究结果表明生物多样性 对生态系统功能具有显著的影响。目前,大多数有关生物多样性与生态系统功能关系(BEF)的研究主要是通过对草本植物的控制实验来进行的,由于其选择的物 种数量有限再加之控制的条件与野外群落的实际生态条件有较大差异,因此其结果与野外实际情况的吻合性还有待证实。基于野外实地的调查观察也还很少,尤其在 森林生态系统中更是这样。但基于...
|
| [4] |
Field observed relationship between biodiversity and ecosystem functioning during secondary succession in a tropical lowland rainforest.
|
| [5] |
Cloud Forest Agenda. UNEP-WCMC, Cambridge , |
| [6] |
Alpine cushion plants inhibit the loss of phylogenetic diversity in severe environments. https://doi.org/10.1111/ele.12070 URL PMID: 23346919 Magsci 摘要
Biotic interactions can shape phylogenetic community structure (PCS). However, we do not know how the asymmetric effects of foundation species on communities extend to effects on PCS. We assessed PCS of alpine plant communities around the world, both within cushion plant foundation species and adjacent open ground, and compared the effects of foundation species and climate on alpha (within-microsite), beta (between open and cushion) and gamma (open and cushion combined) PCS. In the open, alpha PCS shifted from highly related to distantly related with increasing potential productivity. However, we found no relationship between gamma PCS and climate, due to divergence in phylogenetic composition between cushion and open sub-communities in severe environments, as demonstrated by increasing phylo-beta diversity. Thus, foundation species functioned as micro-refugia by facilitating less stress-tolerant lineages in severe environments, erasing a global productivity phylogenetic diversity relationship that would go undetected without accounting for this important biotic interaction.
|
| [7] |
Beyond species: Functional diversity and the maintenance of ecological processes and services. https://doi.org/10.1111/j.1365-2664.2011.02048.x URL Magsci 摘要
1. The goal of conservation and restoration activities is to maintain biological diversity and the ecosystem services that this diversity provides. These activities traditionally focus on the measures of species diversity that include only information on the presence and abundance of species. Yet how diversity influences ecosystem function depends on the traits and niches filled by species.<br/>2. Biological diversity can be quantified in ways that account for functional and phenotypic differences. A number of such measures of functional diversity (FD) have been created, quantifying the distribution of traits in a community or the relative magnitude of species similarities and differences. We review FD measures and why they are intuitively useful for understanding ecological patterns and are important for management.<br/>3. In order for FD to be meaningful and worth measuring, it must be correlated with ecosystem function, and it should provide information above and beyond what species richness or diversity can explain. We review these two propositions, examining whether the strength of the correlation between FD and species richness varies across differing environmental gradients and whether FD offers greater explanatory power of ecosystem function than species richness.<br/>4. Previous research shows that the relationship between FD and richness is complex and context dependent. Different functional traits can show individual responses to different gradients, meaning that important changes in diversity can occur with minimal change in richness. Further, FD can explain variation in ecosystem function even when richness does not.<br/>5. Synthesis and applications. FD measures those aspects of diversity that potentially affect community assembly and function. Given this explanatory power, FD should be incorporated into conservation and restoration decision-making, especially for those efforts attempting to reconstruct or preserve healthy, functioning ecosystems.
|
| [8] |
A handbook of protocols for standardized and easy measurement of plant functional traits worldwide.
|
| [9] |
A trait-based test for habitat filtering: Convex hull volume. https://doi.org/10.1890/0012-9658(2006)87[1465:ATTFHF]2.0.CO;2 URL PMID: 16869422 摘要
Abstract Community assembly theory suggests that two processes affect the distribution of trait values within communities: competition and habitat filtering. Within a local community, competition leads to ecological differentiation of coexisting species, while habitat filtering reduces the spread of trait values, reflecting shared ecological tolerances. Many statistical tests for the effects of competition exist in the literature, but measures of habitat filtering are less well-developed. Here, we present convex hull volume, a construct from computational geometry, which provides an n-dimensional measure of the volume of trait space occupied by species in a community. Combined with ecological null models, this measure offers a useful test for habitat filtering. We use convex hull volume and a null model to analyze California woody-plant trait and community data. Our results show that observed plant communities occupy less trait space than expected from random assembly, a result consistent with habitat filtering.
|
| [10] |
Spatial mismatch and congruence between taxonomic phylogenetic and functional diversity: The need for integrative conservation strategies in a changing world. |
| [11] |
Conservation evaluation and phylogenetic diversity. https://doi.org/10.1016/0006-3207(92)91201-3 URL 摘要
Protecting biological diversity with limited resources may require placing conservation priorities on different taxa. A system of priorities that reflects the value of taxonomic diversity can be achieved by setting priorities such that the subset of taxa that is protected has maximum underlying feature diversity. Such feature diversity of taxon subsets is difficult to estimate directly, but can be predicted by the cladistic/phylogenetic relationships among the taxa. In this study, a simple measure of phylogenetic diversity is defined based on cladistic information. The measure of phylogenetic diversity, PD, is contrasted with a measure of taxic diversity recently developed by Vane-Wright et al. (Biol. Conserv. , 55 , 1991). In re-examining reserve-selection scenarios based on a phylogeny of bumble bees (Apidae), PD produces quite different priorities for species conservation, relative to taxic diversity. The potential application of PD at levels below that of the species is then illustrated using a mtDNA phylogeny for populations of crested newts Triturus cristatus . Calculation of PD for different population subsets shows that protection of populations at either of two extremes of the geographic range of the group can significantly increase the phylogenetic diversity that is protected.
|
| [12] |
Spatial and Temporal Pattern of Biodiversity and Its Correlation with Aboveground Biomass in Coniferous and Broadleaved Mixed Forests of Jiaohe, Jilin. PhD dissertation, 吉林蛟河针阔混交林生物多样性时空格局及其与地上部分生物量的关系 .URL 摘要
生物多样性对生态系统功能的影 响已成为科学界和公众关注的热点。本文以吉林蛟河林区人为采伐后处于不同发育阶段的针阔混交林为研究对象,通过群落调查、功能性状测定和DNA条形码技 术,探讨了物种多样性、功能多样性及谱系多样性随林分发育阶段(中龄林、近熟林、成熟林、老龄林)和生境类型(低谷、低坡、高谷、高坡、山顶)的时空格 局;研究了生物多样性与地上部分生物量之间的关系。主要研究结果如下:(1)从中龄林-近熟林-成熟林-老龄林的发育过程中,红松和紫椴等优势树种的优势 地位逐渐增强,先锋树种逐渐被耐阴树种所取代。四块样地中全部个体的径级结构均呈倒“J”型分布。(2)从中龄林到成熟林的发育过程中,林分地上部分生物 量逐渐增加,而成熟林和老龄林二者之间无显著差异(p0.05);30公顷老龄林样地中,从低谷至山顶地上部分生物量呈逐渐增大的趋势。地形因子对中龄 林、近熟林和成熟林地上部分生物量无显著影响(p0.05),而海拔和凹凸度对老龄林地上部分生物量影响显著(p0.05)。(3)不同发育阶段的针阔混 交林植物功能性状之间差异显著。中龄林和近熟林地上部分生物量与比叶面积、叶片全氮含量无显著相关性,而成熟林和老龄林地上部分生物量与比叶面积、叶片全 氮含量均呈显著负相关(p0.05);老龄林样地中,低坡和山顶生境的地上部分生物量与比叶面积、叶片全氮含量均呈显著负相关(p0.05)。(4)物种 丰富度和Shannon-wiener指数随林分发育均呈增加的趋势,而Pielou's均匀度指数在不同发育阶段的林分之间无显著差异(p0.05): 从中龄林到老龄林的发育过程中,功能丰富度、功能均匀度、Rao's二次熵均呈现先下降后上升的趋势,净谱系亲缘关系指数(NRI)和最近分类单元指数 (NTI)呈现逐渐减小的趋势,而谱系多样性指数PD则呈现逐渐增大的趋势;老龄林样地中,从低谷至山顶群落的NRI呈现降低趋势,而NTI呈现先上升后 下降的趋势。物种丰富度、功能丰富度与谱系多样性两两之间均呈显著正相关(p0.05)。(5)老龄林样地中,物种丰富度、功能丰富度、谱系多样性均与地 上部分生物量呈显著正相关(p0.05);中龄林和老龄林功能丰富度分别解释了地上部分生物量5%和12%的变异;除近熟林谱系多样性与地上部分生物量无 显著相关性外,其他三个发育阶段的林分谱系多样性均与地上部分生物量呈显著正相关(p0.05)。老龄林样地中,低谷和低坡生境中的物种丰富度和功能丰富 度均与地上部分生物量呈显著正相关且物种丰富度解释了更多的变异。总体上,地形因子对不同发育阶段的林分地上部分生物量、功能性状和生物多样性均有显著影 响,且地形因子对老龄林的影响较大:功能性状与地上部分生物量存在一定的关联,尤其是比叶面积和叶片全氮含量,且这种关联性在成熟林和老龄林,以及在老龄 林的低谷和山顶生境中达到显著水平。生物多样性是地上部分生物量的主要驱动力之一。
|
| [13] |
Comparison of phylobetadiversity indices based on community data from Gutianshan forest plot. https://doi.org/10.1007/s11434-011-4869-1 URL 摘要
Phylobetadiversity incorporates phylogenetic information and beta diversity,and can account for the ecological similarities between communities with a phylogenetic perspective.Although different phylobetadiversity indices reflect differences in different characteristics between communities,the results of different phylobetadiversity indices are not comparable.In this study we examined phylobetadiversity indices for a 24-hm 2 plot in the Gutianshan National Nature Reserve.It was found the abundanceweighted D pw was almost identical to Rao's D of Rao's quadratic entropy.PhyloSor had a similar ecological meaning and algorithm to UniFrac.Although Dnn was different in definition from UniFrac and PhyloSor,they were all strongly correlated.The effect of species abundance on phylobetadiversity was not significant when scales were relatively small,but was significant at larger scales.These contrasts likely resulted from reductions in evenness in communities as scales increased.P ST and Rao's H better reflected the distance-decay changes caused by spatial and habitat variation than other indices at larger scales,whereas AW-D nn and D nn better reflected these changes at small scales.
|
| [14] |
Phylogenetic beta diversity linking ecological and evolutionary processes across space in time. |
| [15] |
Phylogenetic diversity-area curves. |
| [16] |
|
| [17] |
The distribution of the flora in the alpine zone. |
| [18] |
Analyzing β diversity: Partitioning the spatial variation of community composition data. |
| [19] |
Functional diversity changes during tropical forest succession. https://doi.org/10.1016/j.ppees.2011.10.002 URL 摘要
Functional diversity (FD) ‘those components of biodiversity that influence how an ecosystem operates or functions’ is a promising tool to assess the effect of biodiversity loss on ecosystem functioning. FD has received ample theoretical attention, but empirical studies are limited. We evaluate changes in species richness and FD during tropical secondary forest succession after shifting cultivation in Mexico. We also test whether species richness is a good predictor of FD. FD was calculated based on a combination of nine functional traits, and based on two individual traits important for primary production (specific leaf area) and carbon sequestration (wood density). Stand basal area was a good predictor of successional changes in diversity and FD, in contrast to fallow age. Incidence-based FD indices increased logarithmically with stand basal area, but FD weighted by species’ importance values lacked pattern with succession. Species richness and diversity are strong predictors of FD when all traits were considered; linear relationships indicate that all species are equally functionally complementary, suggesting there is little functional redundancy. In contrast, when FD was calculated for individual traits and weighted for abundances, species richness may underestimate FD.
|
| [20] |
Community assembly in a tropical cloud forest related to specific leaf area and maximum species height. https://doi.org/10.1111/jvs.12256 URL 摘要
Abstract Question We tested whether co-existing tree species in tropical dwarf forests were deterministically assembled along gradients of air temperature, relative humidity and light availability, according to two important functional traits, specific leaf area (SLA) and maximum species height ( H max ). Location Tropical montane cloud forest, Bawangling Nature Reserve, Hainan Island, south China. Methods Null model analyses were used in conjunction with trait and species composition data collected to test our hypotheses at four plot sizes (25m 2 , 100m 2 , 400m 2 and 900m 2 ), addressing whether the consistent importance of variation in SLA and H max extends to these unique forests, as well as theoretical predictions concerning how patterns change with plot size. Results Low SLA species were significantly over-represented within forest communities for the two largest plot sizes, and taller-growing tree species were over-represented across all four plot sizes. Plot-level analyses indicated that low SLA species were associated with lower temperatures. Conclusions Our results show that tropical dwarf forests are deterministically assembled with respect to these two traits, and are consistent with other studies indicating that SLA responds to abiotic filters. Co-existing tree species were not significantly divergent for these two traits, indicating that variation in these two traits among trees does not contribute to niche differences (i.e. limiting similarity) and therefore co-existence within forest plots. Finally, our study demonstrates that patterns of community assembly change with plot size; however, trait convergence did not increase with plot size as previously predicted.
|
| [21] |
a). Air temperature and soil phosphorus availability correlate with trait differences between two types of tropical cloud forests. https://doi.org/10.1016/j.flora.2011.05.007 URL Magsci 摘要
Tropical cloud forests are characterized by lower air temperature and high frequency of fog condensation at canopy level, as compared with forests at lower altitudes. Few studies have been conducted to understand differences of plant functional traits in relation to the environment in this kind of forest. In this paper, we explored the community-level differences of specific leaf area (SLA) and height of plants in relation to major environmental conditions between two adjacent tropical cloud forests on Hainan Island, South China. We measured the two functional traits for all individual plants within twenty-nine and thirty-two 10 m x 10 m plots located in a low altitude tropical montane evergreen forest (TMEF) and a high altitude tropical dwarf forest (TDF), respectively.<br/>The results showed that both mean SLA and height decreased from TMEF to TDF, while phenotypic plasticity for the two functional traits increased from TMEF to TDF. Correlation analysis and multiple regression analysis showed that the mean SLA and its plasticity were significantly correlated with both air temperature and soil phosphorus. The mean height was only significantly correlated with air temperature, but its plasticity was significantly correlated with both air temperature and soil phosphorus.<br/>Our results suggest that plants in dwarf tropical cloud forests have decreased SLA and height, correlated with less favorable soil and atmospheric conditions, with a higher plasticity of these traits, as compared with the tropical montane evergreen forest. Community-level differences in SLA and plant height thus can be taken as indicators characterizing plant distribution to different types of tropical cloud forests. (C) 2011 Elsevier GmbH. All rights reserved.
|
| [22] |
a). Community characteristics of tropical montane evergreen forest and tropical montane dwarf forest in Bawangling National Nature Reserve on Hainan Island, South China. a). 海南岛霸王岭热带山地常绿林和热带山顶矮林群落特征 .https://doi.org/10.3724/SP.J.1003.2011.09309 URL Magsci 摘要
热带山地常绿林和热带山顶矮林均属于热带云雾林。为了揭示其群落结构和物种多样性特征, 在海南岛霸王岭热带山地常绿林和热带山顶矮林分别设置8个和10个2,500 m<sup>2</sup>样方, 调查所有DBH≥1 cm的乔木、灌木和藤本植株。结果显示: (1)热带山地常绿林幼树(1 cm≤DBH<5 cm)和小树(5 cm≤DBH<10 cm)的平均密度显著小于热带山顶矮林, 而成年树(DBH≥10 cm)平均密度无显著差异; 前者小树和成年树的平均胸径显著大于后者, 而幼树平均胸径显著小于后者; 前者所有径级植株的平均高度显著大于后者; (2)两森林类型优势种不同, 但优势科和优势属相似, Sørensen物种相似性指数为0.71; (3)逻辑斯蒂模型是拟合两森林类型种–面积关系的最优模型; 热带山地常绿林物种丰富度观测值及根据刀切法和抽样法估计的物种丰富度值都比热带山顶矮林高。综合分析表明, 热带山地常绿林和热带山顶矮林群落植株密度、胸径、高度和物种多样性差异较大。
|
| [23] |
Effects of competition and facilitation on species assemblage in two types of tropical cloud forest. https://doi.org/10.1371/journal.pone.0060252 URL 摘要
Competition and facilitation between tree individuals are two kinds of non-random processes influencing the structure and functioning of forest communities, but how these two plant-plant interactions change along gradient of resources or environments remains very much a matter of debate. We developed a null model to test the size-distance regression, and assessed the effects of competition and facilitation (including interspecific interactions, intraspecific interactions and overall species interactions) on each adult tree species assemblage [diameter at breast height (dbh) 鈮5 cm] across two types of tropical cloud forest with different environmental and resource regimes. The null model test revealed that 17% to 27% tree species had positive dbh-distance correlations while 11% to 19% tree species showed negative dbh-distance correlations within these two forest types, indicating that both competition and facilitation processes existed during the community assembly. The importance of competition for heterospecific species, and the intensity of competition for both heterospecific and overall species increased from high to low resources for all the shared species spanning the two forests. The importance of facilitation for conspecific and overall species, as well as that the intensity of facilitation for both heterospecific and conspecific species increased with increasing low air temperature stress for all the shared species spanning the two forests. Our results show that both competition and facilitation processes simultaneously affect parts of species assemblage in the tropical cloud forests. Moreover, the fact that nearly 50% species assemblage is not detected with our approaches suggest that tree species in these tropical forest systems are assembled with multiple ecological processes, and that there is a need to explore the processes other than the two biotic interactions in further researches.
|
| [24] |
b). Environmental characteristics of tropical cloud forests in the rainy season in Bawangling National Nature Reserve on Hainan Island, South China. b). 海南岛霸王岭热带云雾林雨季的环境特征 .https://doi.org/10.3724/SP.J.1258.2011.00137 URL Magsci 摘要
<p>以海南岛霸王岭热带云雾林中热带山地常绿林和热带山顶矮林群落为对象, 分析了热带云雾林雨季光照、空气湿度、空气温度、土壤和地形特征, 为分析生物多样性、生态系统功能及其对气候变化的反应等生态学过程奠定基础。结果表明: 热带山地常绿林和热带山顶矮林在一天中的光合有效辐射呈单峰曲线变化, 热带山地常绿林各时段的光合有效辐射显著低于热带山顶矮林; 5–10月两群落类型日平均空气温度分别为(21.76 ± 2.44) ℃和(19.33 ± 1.03) ℃, 且随时间变化呈单峰曲线, 热带山地常绿林日平均空气温度显著高于热带山顶矮林; 5–10月两群落类型日平均空气相对湿度分别为(88.44 ± 2.90)% 和(97.71 ± 0.80)%, 且随时间变化呈倒“S”型曲线, 热带山地常绿林各月日平均空气相对湿度显著小于热带山顶矮林; 与热带山顶矮林相比, 热带山地常绿林的土壤全氮、全磷、速效氮、有机质、pH和土壤厚度显著大, 而全钾和有效磷含量显著低; 热带山地常绿林的坡度、岩石裸露比例和海拔高度显著小于热带山顶矮林, 地形因子与其他生态因子显著相关; 主成分和相关性分析表明: 空气温度、有效磷、全钾、全氮及地形因子对热带云雾林植被分布有重要影响。</p>
|
| [25] |
b). Within- and among-species variation in specific leaf area drive community assembly in a tropical cloud forest. https://doi.org/10.1007/s00442-011-2050-9 URL PMID: 21695546 Magsci 摘要
Specific leaf area (SLA) is a key functional trait reflecting the trade-off between resource capture and conservation, and has been identified as playing an important role in plant community assembly. Mechanistic models of community assembly state that the assemblage of species in a local community is controlled by environment filters operating on functional traits. We measured within- and among-species variation of SLA, and environmental conditions in a tropical cloud forest to explore how variation in this functional trait contributes to community assembly. SLA variation at the species level was also decomposed into alpha (within assemblage variation), and beta (across assemblage variation) values. SLA decreased with increasing solar irradiance (approximated using plant height) within the three study sites, and differed among the three sites both for within- and among-species comparisons. Mean plot SLA, accounting for both within and among species across the three sites, increased significantly in relation to air temperature but not local photosynthetic photon flux density and soil total phosphorus. Alpha SLA decreased with increasing solar irradiance within the three sites and beta SLA differed among the three sites. Our results clearly demonstrate that light and air temperature are key environmental factors involved in organizing plant species within and among communities in tropical cloud forests. The strong relationship between both intra- and interspecific variation in SLA and environmental conditions strongly confirms the role of trait variation in the assembly of plant species in tropical cloud forest communities via environment filtering related to light availability and air temperature.
|
| [26] |
Biological community diversity measure II. β diversity measure method. 生物群落多样性的测度方法II. β多样性的测度方法 . |
| [27] |
Functional diversity: A tool for answering challenging ecological questions. https://doi.org/10.2307/23467164 URL 摘要
Abstract Functional trait diversity provides a powerful means of addressing ecology's persistent questions, through its dual role as an indicator of mechanisms driving differences in species composition between communities and as a predictor of ecosystem-level processes. Functional traits provide a means of testing mechanisms behind species turnover between communities because environmental heterogeneity, competition and disturbance influence species fitness via their traits. Functional traits also provide a link between species and multiple ecosystem-level processes, such as primary productivity, nutrient fluxes and resilience, since species influence these processes via their traits. This special issue demonstrates that functional diversity offers a practical means of investigating ecology's persistent questions.
|
| [28] |
Changes in coexistence mechanisms along a long-term soil chronosequence revealed by functional trait diversity. |
| [29] |
Functional diversity measures: An overview of their redundancy and their ability to discriminate community assembly rules. https://doi.org/10.1111/j.1365-2435.2010.01695.x URL 摘要
Summary Top of page Summary Introduction Materials and methods Results Discussion Conclusion Acknowledgements References 1. 68Indices quantifying the functional aspect of biodiversity are essential in understanding relationships between biodiversity, ecosystem functioning and environmental constraints. Many indices of functional diversity have been published but we lack consensus about what indices quantify, how redundant they are and which ones are recommended. 2. 68This study aims to build a typology of functional diversity indices from artificial data sets encompassing various community structures (different assembly rules, various species richness levels) and to identify a set of independent indices able to discriminate community assembly rules. 3. 68Our results confirm that indices can be divided into three main categories, each of these corresponding to one aspect of functional diversity: functional richness, functional evenness and functional divergence. Most published indices are highly correlated and quantify functional richness while quadratic entropy ( Q ) represents a mix between functional richness and functional divergence. Conversely, two indices (FEve and FDiv respectively quantifying functional evenness and functional divergence) are rather independent to all the others. The power analysis revealed that some indices efficiently detect assembly rules while others performed poorly. 4. 68To accurately assess functional diversity and establish its relationships with ecosystem functioning and environmental constraints, we recommend investigating each functional component separately with the appropriate index. Guidelines are provided to help choosing appropriate indices given the issue being investigated. 5. 68This study demonstrates that functional diversity indices have the potential to reveal the processes that structure biological communities. Combined with complementary methods (phylogenetic and taxonomic diversity), the multifaceted framework of functional diversity will help improve our understanding of how biodiversity interacts with ecosystem processes and environmental constraints.
|
| [30] |
Environmental heterogeneity and disturbance by humans control much of the tree species diversity of Atlantic montane forest fragments in SE Brazil. https://doi.org/10.1007/s10531-006-9063-4 URL Magsci 摘要
The effects of human impact and environmental heterogeneity on the tree species diversity were assessed in 20 fragments of tropical montane seasonal forest in southeastern Brazil. Previous surveys of the tree community, soils and topography of the fragments provided the bulk of the data. The diversity parameters used were the means of species richness, Shannon diversity (<i>H</i>′), and Pielou evenness (<i>J</i>′) obtained from “bootstrap” sub-samplings of 1,000 trees. Morphometric variables obtained for the fragments included total, edge, and inner areas. Investigation forms were used to survey the history of human interventions and prepare an impacts matrix containing scores assigned to assess the extent, severity and duration of selected impacts. Scores for overall environmental impacts were obtained from the ordination scores produced by a multivariate analysis of the impacts matrix. A multivariate analysis of the standard deviations of soil variables was used to identify the variable which contributed most to soil heterogeneity. The same procedure was repeated for the variables related to topography and ground-water regime. The three species diversity parameters were related to the proportions of edges, the overall impacts scores, and the standard deviations of two selected soil and topographic variables. The species diversity in the fragments increased with increasing heterogeneity of both soil chemical properties and topographic features, and decreased with increasing proportion of forest edges. The evenness component of species diversity also increased with increasing severity of overall environmental impacts. This probably occurred because the 20 fragments did not include highly disturbed forests in the range and the intermediate disturbance effect on species diversity was therefore detected.
|
| [31] |
An updated megaphylogeny of plants, a tool for generating plant phylogenies and an analysis of phylogenetic community structure. https://doi.org/10.1093/jpe/rtv047 URL 摘要
1 Research and Collections Center, Illinois State Museum, 1011 East Ash Street, Springfield, IL 62703, USA2 College of Life Sciences, Zhejiang University, Hangzhou 310058, China *Correspondence address. Research and Collections Center, Illinois State Museum, 1011 East Ash Street, Springfield, IL 62703, USA. Tel:聽+1 217 782 2621; Fax: +1 217 785 2857; E-mail: hqian@museum.state.il.us
|
| [32] |
R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing ,URL 摘要
CiteSeerX - Scientific documents that cite the following paper: R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing
|
| [33] |
Phylogenetic endemism: A new approach for identifying geographical concentrations of evolutionary history. https://doi.org/10.1111/j.1365-294X.2009.04311.x URL PMID: 19754516 摘要
We present a new, broadly applicable measure of the spatial restriction of phylogenetic diversity, termed(PE). PE combines the widely used phylogenetic diversity and weighted endemism measures to identify areas where substantial components of phylogenetic diversity are restricted. Such areas are likely to be of considerable importance for conservation. PE has a number of desirable properties not combined in previous approaches. It assesses endemism consistently, independent of taxonomic status or level, and independent of previously defined political or biological regions. The results can be directly compared between areas because they are based on equivalent spatial units. PE builds on previous phylogenetic analyses of endemism, but provides a more general solution for mapping endemism of lineages. We illustrate the broad applicability of PE using examples of Australian organisms having contrasting life histories: pea-flowered shrubs of the genus (Fabaceae) and the Australian species of the Australo-Papuan tree frog radiation within the family Hylidae.
|
| [34] |
A user’s guide to functional diversity indices. |
| [35] |
Functional diversity supports the physiological tolerance hypothesis for plant species richness along climatic gradients. https://doi.org/10.1111/1365-2745.12204 URL Magsci 摘要
<ol id="jec12204-list-0001" class="numbered"><li>The physiological tolerance hypothesis proposes that plant species richness is highest in warm and/or wet climates because a wider range of functional strategies can persist under such conditions. Functional diversity metrics, combined with statistical modelling, offer new ways to test whether diversity–environment relationships are consistent with this hypothesis.</li><li>In a classic study by R.H. Whittaker (<em>Ecological Monographs</em>, 1960), herb species richness declined from mesic (cool, moist, northerly) slopes to xeric (hot, dry, southerly) slopes. Building on this data set, we measured four plant functional traits (plant height, specific leaf area, leaf water content and foliar C:N) and used them to calculate three functional diversity metrics (functional richness, evenness and dispersion). We then used a structural equation model to ask whether ‘functional diversity’ (modelled as the joint responses of richness, evenness and dispersion) could explain the observed relationship of topographic climate gradients to species richness. We then repeated our model examining the functional diversity of each of the four traits individually.</li><li>Consistent with the physiological tolerance hypothesis, we found that functional diversity was higher in more favourable climatic conditions (mesic slopes) and that multivariate functional diversity mediated the relationship of the topographic climate gradient to plant species richness. We found similar patterns for models focusing on individual trait functional diversity of leaf water content and foliar C:N.</li><li><em>Synthesis</em>. Our results provide trait-based support for the physiological tolerance hypothesis, suggesting that benign climates support more species because they allow for a wider range of functional strategies.</li></ol>
|
| [36] |
Cloud Forest in the Humid Tropics: A Bibliographic Review. United Nations University, Tokyo ,
|
| [37] |
The assembly of tropical tree communities—The advances and shortcomings of phylogenetic and functional trait analyses. https://doi.org/10.1111/j.1600-0587.2012.00121.x URL Magsci 摘要
Tropical tree communities present one of the most challenging systems for studying the processes underlying community assembly. Most community assembly hypotheses consider the relative importance of the ecological similarity of co-occurring species. Quantifying this similarity is a daunting and potentially impossible task in species-rich assemblages. During the past decade tropical tree ecologists have increasingly utilized phylogenetic trees and functional traits to estimate the ecological similarity of species in order to test mechanistic community assembly hypotheses. A large amount of work has resulted with many important advances having been made along the way. That said, there are still many outstanding challenges facing those utilizing phylogenetic and functional trait approaches to study community assembly. Here I review the conceptual background, major advances and major remaining challenges in phylogenetic- and trait-based approaches to community ecology with a specific focus on tropical trees. I argue that both approaches tremendously improve our understanding of tropical tree community ecology, but neither approach has fully reached its potential thus far.
|
| [38] |
Deterministic tropical tree community turnover: Evidence from patterns of functional β diversity along an elevational gradient. |
| [39] |
Phylogenetic and functional alpha and beta diversity in temperate and tropical |
| [40] |
|
| [41] |
Species richness, but not phylogenetic diversity, influences community biomass production and temporal stability in a re-examination of 16 grassland biodiversity studies. https://doi.org/10.1111/1365-2435.12432 URL 摘要
Summary Top of page Summary Introduction Materials and methods Results Discussion Data accessibility References Supporting Information Hundreds of experiments have now manipulated species richness (SR) of various groups of organisms and examined how this aspect of biological diversity influences ecosystem functioning. Ecologists have recently expanded this field to look at whether phylogenetic diversity (PD) among species, often quantified as the sum of branch lengths on a molecular phylogeny leading to all species in a community, also predicts ecological function. Some have hypothesized that phylogenetic divergence should be a superior predictor of ecological function than SR because evolutionary relatedness represents the degree of ecological and functional differentiation among species. But studies to date have provided mixed support for this hypothesis. Here, we reanalyse data from 16 experiments that have manipulated plant SR in grassland ecosystems and examined the impact on above-ground biomass production over multiple time points. Using a new molecular phylogeny of the plant species used in these experiments, we quantified how the PD of plants impacts average community biomass production as well as the stability of community biomass production through time. Using four complementary analyses, we show that, after statistically controlling for variation in SR, PD (the sum of branches in a molecular phylogenetic tree connecting all species in a community) is neither related to mean community biomass nor to the temporal stability of biomass. These results run counter to past claims. However, after controlling for SR, PD was positively related to variation in community biomass over time due to an increase in the variances of individual species, but this relationship was not strong enough to influence community stability. In contrast to the non-significant relationships between PD, biomass and stability, our analyses show that SR per se tends to increase the mean biomass production of plant communities, after controlling for PD. The relationship between SR and temporal variation in community biomass was either positive, non-significant or negative depending on which analysis was used. However, the increases in community biomass with SR, independently of PD, always led to increased stability. These results suggest that PD is no better as a predictor of ecosystem functioning than SR. Synthesis . Our study on grasslands offers a cautionary tale when trying to relate PD to ecosystem functioning suggesting that there may be ecologically important trait and functional variation among species that is not explained by phylogenetic relatedness. Our results fail to support the hypothesis that the conservation of evolutionarily distinct species would be more effective than the conservation of SR as a way to maintain productive and stable communities under changing environmental conditions.
|
| [42] |
Effects of negative density dependence and habitat filtering on temporal variation in phylogenetic community structure of seedlings in a mid-subtropical forest. Chinese Science Bulletin ( 负密度制约和生境过滤对古田山幼苗系统发育多样性时间变化的影响 .https://doi.org/10.1360/972013-1141 URL Magsci 摘要
<p>负密度制约和生境过滤是解释森林群落物种共存和生物多样性维持机制的重要假说.已有大量研究表明,负密度制约通过优势物种的种内竞争及近缘种之间的种间竞争、个体间病菌传播等过程为非优势物种及非亲缘种提供生存空间,从而使局域群落的系统发育α多样性随时间的推移而增加;由于生境过滤的作用,特定的生境选择亲缘关系相近的物种,因而随着时间的推移,同一生境内群落间的系统发育β多样性会降低.前人的研究主要集中于利用负密度制约和生境过滤来解释物种多样性的空间变化格局,而利用这2个机制来解释在时间序列上群落系统发育多样性格局变化的研究还很少.本文以浙江古田山24hm<sup>2</sup>亚热带常绿阔叶林永久性样地中的幼苗(胸径<1cm)为研究对象,利用2006和2007年的幼苗动态监测数据,分析位于不同生境的342个幼苗样方内的系统发育α多样性和样方间的系统发育β多样性随时间的格局变化.结果表明,随着时间的变化,位于同一生境的样方幼苗系统发育α多样性显著增加;而幼苗系统发育β多样性在同一生境内显著降低.因此,在亚热带森林群落中,负密度制约效应和生境过滤共同维持幼苗群落的物种共存.</p>
|
| [43] |
Exploring the phylogenetic structure of ecological communities: An example for rain forest trees. |
| [44] |
Phylocom: software for the analysis of phylogenetic community structure and character evolution. https://doi.org/10.1093/bioinformatics/btn358 URL PMID: 18678590 摘要
Motivation: The increasing availability of phylogenetic and trait data for communities of co-occurring species has created a need for software that integrates ecological and evolutionary analyses.Capabilities: Phylocom calculates numerous metrics of phylogenetic community structure and trait similarity within communities. Hypothesis testing is implemented using several null models. Within the same framework, it measures phylogenetic signal and correlated evolution for species traits. A range of utility functions allow community and phylogenetic data manipulation, tree and trait generation, and integration into scientific workflows.
|
| [45] |
|
| [46] |
Functional Diversity Research of Forest Communities in Qiliyu, Shanxi. PhD dissertation, Shanxi Normal University, 山西七里峪森林群落功能多样性研究 .URL 摘要
近年来基于物种功能特征的功能 多样性研究方法在生态学领域倍受重视。本文在查阅资料的基础上,介绍了功能多样性的概念、功能性状的选择与测定、6种计算方法以及应注意的问题。最后在野 外样方调查的基础上,通过选取种子的扩散方式、生长型、植物的高度、盖度、叶干物质含量、比叶面积、叶面积、叶宽、叶长、叶厚度、开花期11个功能性状, 运用R2.15.2软件计算出功能性状距离、功能性状平均距离、功能体积、功能均匀度指数、功能分散指数、Rao二次熵指数6个功能多样性指数,结合物种 丰富度、香浓-威纳指数、辛普森指数、物种均匀度,对山西七里峪森林群落的物种组成、功能多样性指数的特点、功能多样性与物种多样性的关系、群落间功能多 样性和功能性状的差异进行了全面研究,旨在为山西七里峪森林群落的保护及管理提供科学依据。结果表明: (1)七里峪植物森林群落共有植物204种,隶属51科152属。其中,裸子植物有2科3属3种,其余为被子植物,其中双子叶植物包括44科127属 175种,单子叶植物5科22属26种。 (2)杨桦林的功能丰富度指数普遍较高,野核桃林和辽东栎林居中,针叶林最低。2个功能离散度指数在7个群落间排序一致。 (3)3个功能丰富度指数均与物种丰富度、香农-威纳指数在0.01水平呈现显著正相关关系;功能均匀度与香农-威纳指数、物种均匀度指数在0.05水平 显著正相关;Rao二次熵指数在0.05水平与香农-威纳指数、物种均匀度显著正相关,功能分散指数在0.01水平与香农-威纳指数、物种均匀度显著正相 关。 (4)功能性状距离与功能性状平均距离、功能体积在0.01水平显著正相关;功能性状平均距离与功能性状距离、功能体积在0.01水平显著正相关:功能体 积与功能性状距离与功能性状平均距离在0.01水平显著正相关;功能分散指数、Rao二次熵指数在0.01水平显著正相关。 (5)7个群落间物种多样性没有显著性差异,功能均匀度、功能离散度在群落间在0.01水平有显著性差异。7个群落间的的差异很大程度上是由于常见物种的 功能性状差异引起的。 (6)对6种功能多样性指数主分量分析可知,功能多样性指数可分为3类。第1主成分主要反映了功能丰富度;第2主成分主要反映了功能离散度;第3主成分主 要反映了功能均匀度。 (7)7个群落间叶干物质含量、比叶面积、叶面积、叶长、叶宽有显著性差异,能体现群落间的差异。
|
| [47] |
Phylogenetic and functional alpha and beta diversity in mid-mountain humid evergreen broad-leaved forest. Chinese Science Bulletin ( 中山湿性常绿阔叶林系统发育和功能性状的α及β多样性 .https://doi.org/10.1360/N972014-00401 URL Magsci 摘要
<p>群落构建机制一直是群落生态学研究的核心问题. 群落系统发育和功能性状的α 及β 多样性为探讨生态位过程和中性过程在局域群落构建中的相对作用提供了新的视角. 本研究以哀牢山6 hm<sup>2</sup> 中山湿性常绿阔叶林动态监测样地为研究平台,基于DNA条形码构建的群落系统发育树和树种8 个关键功能性状,结合地形数据,分析不同生活史阶段树种在不同空间尺度的系统发育和功能性状α 及β 多样性. 研究表明:(1) 各生活史阶段树种在不同空间尺度上均表现出非随机的系统发育和功能性状结构,系统发育和功能性状聚集程度随空间尺度的增大而增大;(2) 在所有空间尺度上,系统发育和功能性状的周转都为非随机,并且环境距离对系统发育和功能性状β 多样性的解释程度均高于空间距离. 研究结果揭示了环境过滤在中山湿性常绿阔叶林群落构建中起主导作用.</p>
|
| [48] |
Three keys to the radiation of angiosperms into freezing environments. https://doi.org/10.1038/nature14371 URL PMID: 24362564 摘要
Early flowering plants are thought to have been woody species restricted to warm habitats. This lineage has since radiated into almost every climate, with manifold growth forms. As angiosperms spread and climate changed, they evolved mechanisms to cope with episodic freezing. To explore the evolution of traits underpinning the ability to persist in freezing conditions, we assembled a large species-level database of growth habit (woody or herbaceous; 49,064 species), as well as leaf phenology (evergreen or deciduous), diameter of hydraulic conduits (that is, xylem vessels and tracheids) and climate occupancies (exposure to freezing). To model the evolution of species' traits and climate occupancies, we combined these data with an unparalleled dated molecular phylogeny (32,223 species) for land plants. Here we show that woody clades successfully moved into freezing-prone environments by either possessing transport networks of small safe conduits and/or shutting down hydraulic function by dropping leaves during freezing. Herbaceous species largely avoided freezing periods by senescing cheaply constructed aboveground tissue. Growth habit has long been considered labile, but we find that growth habit was less labile than climate occupancy. Additionally, freezing environments were largely filled by lineages that had already become herbs or, when remaining woody, already had small conduits (that is, the trait evolved before the climate occupancy). By contrast, most deciduous woody lineages had an evolutionary shift to seasonally shedding their leaves only after exposure to freezing (that is, the climate occupancy evolved before the trait). For angiosperms to inhabit novel cold environments they had to gain new structural and functional trait solutions; our results suggest that many of these solutions were probably acquired before their foray into the cold.
|

/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}