碳(C)、氮(N)作为两种重要的生命元素, 其生物地球化学循环过程一直是生态学研究的重点, 二者在自然界都有两种稳定性同位素(12 C、13 C和14 N、15 N)。由于同位素分馏效应, 不同物质间C、N同位素自然丰度存在差异, 这为同位素技术在生态系统碳-氮循环过程中的应用提供了理论基础(Nadelhoffer & Fry, 1988 )。13 C同位素丰度最早被用于区分植物的光合途径(O’Leary, 1981 ), 其后又被发现与植物光合代谢和水分利用效率之间的关系(Farquhar et al ., 1982 ; Ehleringer et al ., 1986 ), 并被广泛应用于土壤碳循环和植物功能群的研究(Garten & Taylor, 1992 ; 于贵瑞等, 2005 )。在发生C3 -C4 植被转换的区域, 通过土壤稳定碳同位素比率(δ 13 C)在时间上的变化, 可以区分不同来源有机质的相对贡献, 同时也可反演植被覆盖的历史, 同样的方法也用于土壤不同有机组分周转过程的研究(Bernoux et al ., 1998 ; Ehleringer et al ., 2000 )。植物-土壤稳定氮同位素比率(δ 15 N)被认为可以用来评估生态系统的N通量, 且已被部分学者用作生态系统氮饱和状态的指示指标(Högberg, 1997 ; Pardo et al ., 2002 ; Kahmen et al ., 2008 ; Xu et al ., 2010 )。Templer等(2007)研究发现土壤δ 15 N值可以表征土壤的净矿化和硝化速率; Craine等(2009)认为植物-土壤δ 15 N高的地区土壤N有效性也更高, 植物根系对菌根真菌的依赖度更弱, 生态系统氮的饱和度更高。显然, 稳定同位素技术的应用为生态系统C、N元素循环的研究提供了一个新的窗口。但目前δ 13 C自然丰度法多应用于发生过C3 -C4 植被转换区域的碳周转研究(Galdo et al ., 2003 ; Liao et al ., 2006 ; Cheng et al ., 2013 ), 而在植被系统未发生C3 -C4 型转换的亚热带森林, 如何应用δ 13 C自然丰度法探究森林演替过程中土壤碳周转的变化, 仍需更多的探索。
尽管已有研究指出, 在地上部分生物量不再增加的情况下, 南亚热带成熟森林土壤依然具有持续积累有机碳的潜力(Zhou et al ., 2006a ), 但对于这种积累机制的解释仍存有很多疑问。诸多研究表明, 具有较高水分利用效率的植物往往氮素利用效率较低, 即在自然条件下, 植物对水分和氮素的利用效率存在补偿制约, 这种制约关系在草原和温带森林生态系统中都得到验证(Field et al ., 1983 ; Chen et al ., 2005 )。而在雨量充沛以及高氮沉降的南亚热带森林, 植物对水分和氮素利用效率的制约关系是否依然存在? 另外,该区域森林生态系统是否已达到“氮饱和”状态? 森林演替过程中土壤氮素有效性如何变化? 这些都是森林生态学研究亟待解决的关键问题。然而, 上述生态学过程和现象的变化在短时间内通过传统的研究手段难以被精确观测。δ 13 C、δ 15 N自然丰度法的应用或许能为我们提供更多有价值的信息。本文选取鼎湖山森林演替序列中3个代表林型——马尾松(Pinus massoniana )针叶林(简称松林)、针阔叶混交林(简称混交林)和季风常绿阔叶林(简称阔叶林), 分别代表演替水平的前、中、后期(Zhou et al ., 2006b ), 通过测定植物-土壤δ 13 C和δ 15 N, 分析叶片δ 13 C、δ 15 N值与其叶片元素含量的关系, 揭示植物-土壤δ 13 C、δ 15 N在演替水平和垂直方向上的变化特征, 试图阐明南亚热带森林演替过程中植物群落对资源的利用策略, 为生态系统碳氮元素循环过程机理的研究提供支持。
1 材料和方法
1.1 研究区域概况
鼎湖山生物圈保护区(23.16°-23.19° N, 112.51°-112.56° E)位于广东省中西部的肇庆市境内, 面积1155 hm2 , 最高峰鸡笼山海拔1000.3 m, 是中国第一个自然保护区。该区属南亚热带季风性气候, 地带性顶级群落类型为季风常绿阔叶林, 恢复演替序列为针阔叶混交林和马尾松针叶林。年降水量和蒸发量分别为1955 mm和1115 mm, 年平均相对湿度80%。干湿季明显, 4-9月为湿季, 降水量约占全年的80%, 11月至次年1月为旱季。年平均气温21.0 ℃, 最冷月(1月)和最热月(7月)平均气温分别为12.6 ℃和28.0 ℃。该区基岩为泥盆系的厚层砂岩、砂页岩, 土壤类型主要为赤红壤, 自然酸化严重, pH值介于4.1-4.9 (Zhou et al ., 2006b )。
鼎湖山阔叶林主要分布在庆云寺周围海拔100-490 m的山坡上, 已有400多年受保护的历史。群落结构复杂, 其中乔木有3层、灌木1层、草本1层, 此外还有较多的藤本植物和附生植物, 主要优势树种有木荷(Schima superba )、锥(Castanopsis chinensis )、黄果厚壳桂(Cryptocarya concinna )、云南银柴(Aporusa yunnanensis )、肖蒲桃(Acmena acuminatissima )、白颜树(Gironniera subaequalis )、橄榄(Canarium album )等(黄忠良等, 1998 )。土壤为水化赤红壤, 土层厚度50-80 cm。
混交林多分布于自然林的林缘, 海拔80-400 m之间, 目前正快速向阔叶林恢复演替。地上垂直结构大致可分为4层, 其中乔木2层、灌木1层、草本1层, 此外还有少量藤本和附生等层间植物, 主要优势树种有马尾松、木荷、锥、黄果厚壳桂、滇粤山胡椒(Lindera metcalfiana )等(欧阳旭等, 2014 )。土壤为赤红壤, 土层厚度40-70 cm。
松林主要分布于保护区外缘的丘陵和低山上, 受人为干扰而未能恢复演替到针阔叶混交林阶段, 但已有相当数量的阔叶小乔木、灌木及草本植物, 优势植物种类有马尾松、白楸(Mallotus paniculatus )、三桠苦(Melicope pteleifolia, 又叫Evodia lepta )、桃金娘(Rhodomyrtus tomentosa )、芒萁(Dicranopteris dicho- toma )等。土壤为粗骨赤红壤, 土层厚度30-50 cm。
1.2 研究方法
1.2.1 样品采集
2014年12月在鼎湖山3个代表性森林(松林、混交林、阔叶林)的永久样地(100 m × 100 m)进行采样。叶片样品的采集: 选取各样地植物群落的主要优势种共14种(其中马尾松为松林和混交林共有种, 木荷、锥、黄果厚壳桂为混交林和阔叶林共有种), 每个树种选择生长状况相近的个体3株, 用高枝剪摘取不同生长部位的叶片并混合均匀, 4个共有种, 在每个林分内取3个样品作为重复, 非共有种也取3个样品作为重复。凋落物样品的采集不分树种, 在各取土点附近随机采集样品并混合均匀, 每个样地取3个样品作为重复。土壤样品的采集: 每个样地按不同坡向、坡度随机选择8个采样点(每个采样点附近随机选取4-5个取土点), 用内径2.5 cm的土钻分4层(0-10 cm, 10-20 cm, 20-30 cm, 30-50 cm)取样, 同一土层混合均匀后装入密封袋保存(约300 g)。共采集叶片样品54个、凋落物样品9个、土壤样品96个。
1.2.2 样品处理和分析
叶片和凋落物样品带回实验室后用蒸馏水洗净, 65 ℃烘至恒质量后, 粉碎过100目筛。土壤样品带回实验室后, 用镊子剔除植物残体、根系和岩屑等异物, 过2 mm孔径(10目)筛, 取部分湿土样(约100 g)留作微生物生物量碳(MBC )的测定。其余样品于室温下自然风干, 按四分法取30-50 g土样, 去除细根后研细过100目筛, 加入过量1 mol·L-1 HCl于室温下处理24 h, 以去除碳酸盐组分, 用蒸馏水反复洗涤至中性, 过滤后65 ℃烘至恒质量。
采用稳定同位素比质谱仪(IsoPrime100, IsoPrime, Manchester, UK)和元素分析仪(Vario ISOTOPE CUBE, Elementar, Hanau, Germany)联用测定样品的δ 13 C、δ 15 N和有机碳(SOC )、全氮(TN )含量及碳氮比(C:N)。同位素比值采用千分比单位(‰), 用δ 表示, 根据国际标准公式计算:
δ (‰) = [(R sam /R std ) -1] × 1000
式中, R = 13 C/12 C或15 N/14 N, R sam 表示样品的相对丰度, R std 表示国际标准物的相对丰度。碳同位素国际标准物为Pee Dee Belemnite (PDB), 即美国南卡罗来纳州白垩系皮狄组地层中的美洲拟箭石(R PDB = 0.0112372, δ 13 CPDB = 0‰); 氮同位素国际标准物为标准大气中的N2 (R air = 0.0036765, δ 15 Nair = 0‰)。本次实验所得同位素比值δ 的测定精度为± 0.1‰, 由中国科学院华南植物园公共实验室提供。土壤微生物生物量碳的测定采用氯仿熏蒸-K2 SO4 浸提法(Vance et al ., 1987 )。土壤易氧化有机碳的测定采用高锰酸钾氧化法(Blair et al ., 1995 )。
1.3 数据处理
利用SAS统计软件对叶片和凋落物的各个指标在林型间的差异进行单因素方差分析, 对土壤各指标在林型和土层间的差异进行双因素方差分析。运用最小显著差异法(LSD )进行均值间的多重比较。采用直线回归分析参数间的相关性。运用SigmaPlot 10.0软件作图。
2 结果和分析
2.1 叶片稳定同位素丰度
2.1.1 共有优势种叶片δ 13 C、δ 15 N在林型间的比较
鼎湖山森林群落主要优势种叶片δ 13 C、δ 15 N值变化范围分别为-33.37‰ - -29.19‰和-4.78‰ - -2.29‰。同种植物在不同演替阶段叶片δ 13 C、δ 15 N存在差异。从表1 可以看出, 松林中马尾松叶片δ 13 C值较混交林高, δ 15 N却相反; 混交林和阔叶林的3个
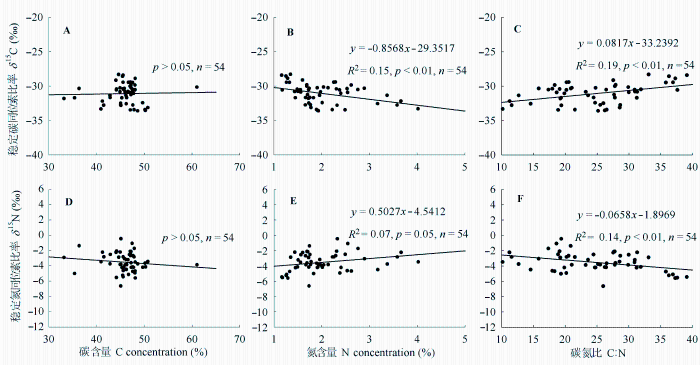
图1 叶片稳定碳(C)、氮(N)同位素比率与元素含量的关系。
Fig. 1 Correlations of foliar stable carbon isotope ratio (δ 13 C) and stable nitrogen isotope ratio (δ 15 N) with foliar elemental contents.
共有优势种木荷、锥、黄果厚壳桂中, 只有木荷叶片δ 15 N在两林型间表现出极显著差异(p < 0.01), 木荷叶片δ 13 C, 锥叶片δ 13 C、δ 15 N, 黄果厚壳桂 叶片δ 13 C、δ 15 N在不同林型间差异均不显著(p > 0.05)。
2.1.2 叶片δ 13 C、δ 15 N与元素含量的关系
由图1 可见, 鼎湖山优势树种叶片δ 13 C与叶片N含量极显著负相关(p < 0.01), 与叶片C:N极显著正相关(p < 0.01); 叶片δ 15 N与N含量正相关(p = 0.05), 与C:N极显著负相关(p < 0.01); 叶片C含量与δ 13 C、δ 15 N均无显著相关性(p > 0.05)。
2.2 凋落物δ 13 C、δ 15 N与碳氮比(C:N)在不同林型间的差异
凋落物δ 13 C值和C:N在不同林型间均存在显著差异(p < 0.05), 具体表现为松林>混交林>阔叶林, 沿着演替方向逐渐降低; 凋落物δ 15 N沿演替方向逐渐增大, 具体表现为松林<混交林<阔叶林(表2 )。
2.3 土壤δ 13 C、δ 15 N与碳氮含量在不同林型间的差异
2.3.1 土壤δ 13 C与土壤总有机碳含量(TOC )、易氧化有机碳含量(ROC )、微生物生物量碳含量(MBC )在不同林型间的差异
土壤δ 13 C值变化范围为-27.31‰ - -21.38‰, 不
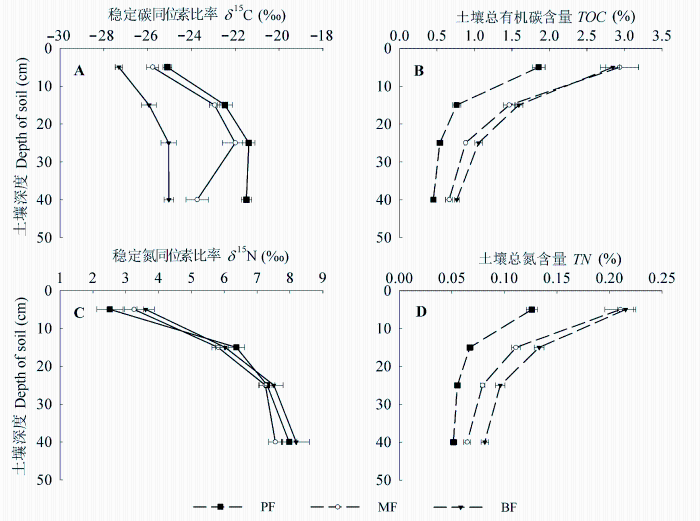
同林型和土层间差异极显著(p < 0.01)(表3 ), 各层土壤δ 13 C均表现为松林>混交林>阔叶林, 沿演替方向逐渐降低。由图2 可见, 土壤δ 13 C沿剖面深度逐渐增大, 至某一深度达到最大值, 松林和混交林在土壤剖面30 cm处δ 13 C值转而减小, 阔叶林土壤剖面20-30 cm和30-50 cm处δ 13 C变化不明显。
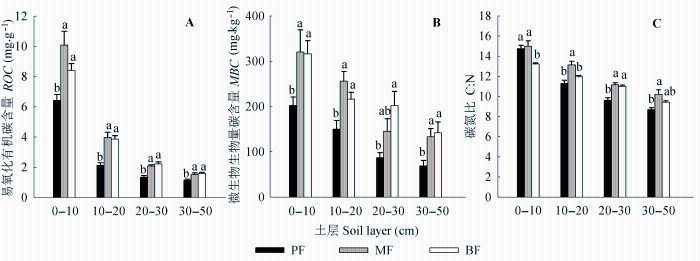
土壤各项C指标在林型和土层间均存在极显著差异(p < 0.01)(表3 ), 松林各层土壤TOC 和活性有机碳(ROC 和MBC )含量均显著低于混交林和阔叶林, 后两者间差异较小; 所有林型土壤TOC 含量沿剖面深度逐渐降低, 与δ 13 C值变化相反(图2 , 图3 )。
2.3.2 土壤δ 15 N与土壤TN和C:N在不同林型间的差异
土壤δ 15 N值变化范围为2.52‰-8.19‰, 表层土壤(0-10 cm) δ 15 N沿演替方向逐渐增大, 具体表现为松林<混交林<阔叶林, 而其他各层土壤δ 15 N值在林型间无显著差异(p > 0.05); 所有林型土壤δ 15 N均沿垂直剖面呈现增大的趋势。各层土壤TN 含量均表现为松林<混交林<阔叶林, 沿剖面深度逐渐降低, 与δ 15 N值变化相反(图2 )。土壤C:N沿剖面深度逐渐降低, 混交林各土层C:N均高于松林和阔叶林(图3C )。
3 讨论
3.1 叶片δ 13 C、δ 15 N特征
多数研究结果证实, C3 植物的碳同位素分馏效应与叶片胞间CO2 浓度和大气CO2 浓度的比值(C i /C a )具有明显的线性正相关关系(Farquhar et al ., 1982 ), 而C i /C a 受光合速率和气孔导度的影响, 因此, 叶片δ 13 C与植物光合和蒸腾等生理过程有着密切的联系, 可以作为估测植物长期水分利用效率(WUE )的重要指标(Ehleringer et al ., 1986 )。本文结果显示混交林中马尾松叶片δ 13 C显著低于松林, 这与演替过程中林内小气候的变化相关。随着演替的进行, 森林郁闭度增加, 林内光照水平减弱, 气温降低, 林内空气湿度和土壤含水量升高(刘效东等, 2014 ), 由此引起植物光合和蒸腾速率的改变, 植物WUE 下降, 叶片C i /C a 升高, δ 13 C值降低。这与Ehleringer等(1986)在鼎湖山的研究结果相吻合, 孙谷畴等(1993)对亚热带人工松林的研究也得到类似的结论。马尾松作为喜光性的耐旱先锋种, 在荫蔽潮湿的环境下优势度减弱, 逐渐被阔叶树种所替代, 群落结构逐步演化成针阔叶混交林。而混交林和阔叶林的3个共有树种叶片δ 13 C均没有表现出林型间的显著差异, 这或许可以间接证明处于演替中、后期的两林型间林分差异越来越小, 混交林林内气温、空气湿度、土壤含水量及植物生理功能逐渐趋同于阔叶林(监测数据, 未发表), 混交林正快速向演替顶级过渡(Zhou et al ., 2006b )。
通过对鼎湖山主要优势树种叶片δ 13 C与其元素含量的相关性分析, 我们发现叶片δ 13 C与N含量极
显著负相关, 与C:N极显著正相关, 与Tsialtas等(2001)的结果一致, 而与其他的一些结论相反(Cernusak et al ., 2013 ; Zhang et al ., 2015 ), 这可能与不同环境条件下土壤养分供应水平及物种在群落中的资源利用策略有关。理论上, 在“氮限制”的环境下, 叶片N含量的增加有利于提高核酮糖1,5-二磷酸羧化氧化酶(Rubisco)的数量和活性, 光合速率加快, C i /C a 降低, 叶片δ 13 C增大(O’Leary, 1988 )。而南亚热带地区氮沉降水平高, 生态系统趋于“氮饱和”状态, 植物的生长转而受到其他营养元素(如磷)的限制(Chen et al ., 2015 ); 鼎湖山降水充沛, 土壤含水量高, 更多的N吸收意味着更多水分的摄入, 从而导致叶片气孔导度增大, C i /C a 升高, δ 13 C降低。以上结果表明: 出于对湿润气候和高氮沉降水平的一种适应策略, 南亚热带森林群落中植物可能同时具有较低的水分利用效率和氮素利用效率(C:N), 并不一定存在相互制约的补偿机制。
除少数固氮植物外, 植物氮素主要来源于根系吸收土壤中的硝态氮和铵态氮, 再经过同化还原作用固定在有机质中, 这两个阶段都会产生氮同位素分馏, 其分馏效应主要取决于土壤N有效性以及植物对氮素的需求(Tcherkez & Hodges, 2008 )。许多研究已经在不同空间尺度上证明叶片δ 15 N与其N含量正相关(Hobbie et al ., 2000 ; Craine et al ., 2009 ), 本文的结果与之相吻合(图1E ), Craine等(2009)认为叶片N含量高的地区土壤N有效性高, 系统N循环开放度更高, 更多贫化15 N的气态N (如NH3 、N2 O和N2 等)被释放, 系统富集15 N; 同时植物对菌根真菌的依赖度下降, N吸收过程中同位素分馏效应减弱, 导致植物δ 15 N增大。表1 结果显示马尾松叶片δ 15 N值在混交林中较松林中显著增大, 可以解释为混交林土壤N有效性明显高于松林, 土壤中氮的净矿化和硝化速率更快(Templer et al ., 2007 )。而混交林和阔叶林的3个共有优势种, 除木荷外, 锥和黄果厚壳桂叶片δ 15 N值均没有表现出林型间的差异, 表明处于演替中期的混交林土壤N有效性越来越接近阔叶林土壤N有效性水平。
图2 土壤稳定碳同位素比率(A)、总有机碳含量TOC (B)、稳定氮同位素比率(C)、总氮含量TN (D)沿剖面变化特征(平均值±标准误差)。BF, 阔叶林; MF, 混交林; PF, 松林。
Fig. 2 Distribution characteristics of soil stable carbon isotope ratio (δ 13 C, A) and total organic carbon content (TOC, B) and stable nitrogen isotope ratio (δ 15 N, C) and total nitrogen content (TN, D) along soil profiles (mean ± SE ). BF, broad-leaved forest; MF, mixed forest; PF, pine forest.
图3 不同林型各土层易氧化有机碳含量(ROC, A)、微生物生物量碳含量(MBC, B)和碳氮比(C:N, C)的变化(平均值±标准误差)。不同字母表示同一土层不同林型间差异显著(p < 0.05)。PF, 松林; MF, 混交林; BF, 阔叶林。
Fig. 3 Change of readily oxidized organic carbon content (ROC, A) and microbial biomass carbon content (MBC, B) and the C to N ratio (C:N, C) at different soil layers under different forests (mean ± SE ). Different letters indicate significant differences among forests for the same soil layer at p < 0.05. BF, broad-leaved forest; MF, mixed forest; PF, pine forest.
3.2 土壤δ 13 C、δ 15 N沿演替方向的变化
本文结果显示各层土壤δ 13 C均沿演替方向(松林→混交林→阔叶林)逐渐降低, 我们认为可能的解释有两种: 地表凋落物的差异以及凋落物分解形成的有机碳(简称新碳)和土壤中原来存储的有机碳(简称老碳)的混合效应。森林土壤有机质主要来源于植物生物量的输入, Balesdent等(1993)对温带森林的研究发现土壤δ 13 C与凋落物δ 13 C呈正相关关系。本文中地表凋落物δ 13 C表现为松林>混交林>阔叶林, 沿演替方向显著降低(表2 ), 与土壤δ 13 C的变化一致。然而, 土壤并不能完全继承现存凋落物的δ 13 C, 如果不考虑凋落物在分解过程中的分馏效应, 土壤δ 13 C应是新碳和老碳混合作用的结果, 其值由同位素混合效应决定(Liao et al ., 2006 )。随着森林演替的进行, 群落结构和林内小气候均发生改变, 地表凋落物C:N降低(表2 ), 微生物数量和活性增加, 地表微环境更易于凋落物分解, 而凋落物分解速率在一定程度上决定着凋落物中碳的流向(以气体形态释放、以碎屑物残存、以可溶性有机碳形态下渗)。Huang等(2011)研究发现, 更快的分解将导致凋落物在分解过程中呼吸消耗的部分减少, 向地下转移的部分(碎屑物和可溶性有机碳)增加。同时, 新碳的快速输入又在一定程度上减缓了土壤中老碳的分解, 因为土壤微生物更倾向于优先分解富含有机质的新鲜碳源(Högberg & Ekblad, 1996 )。对比15年前后鼎湖山森林土壤δ 13 C值的变化, 建立同位素混合模型, 我们发现森林演替后期土壤有机质的平均驻留时间更长(未发表数据)。但目前我们还没有得到凋落物分解过程中转入地下部分的确切比例, 利用同位素技术更为精确地量化凋落物分解过程中碳的流向, 估算土壤不同有机组分的分解速率, 将有助于提供更多清晰的线索。
表层(0-10 cm)土壤δ 15 N沿演替方向呈现增大的规律, 与凋落物δ 15 N的变化一致, 表明土壤δ 15 N与凋落物的输入密切相关。Falkengren-Grerup等(2004)认为δ 15 N可以作为植物吸收NO3 - 的相对指标。Pardo等(2002)发现, 生态系统越接近“氮饱和”, 土壤和叶片的δ 15 N值越高。Craine等(2009)认为植物-土壤δ 15 N高的区域土壤N有效性也更高。也有研究显示表层土壤δ 15 N自然丰度可以作为指示氮循环速率的指标(Xu et al ., 2010 )。Templer等(2007)研究表明细根和土壤的δ 15 N与净矿化和硝化速率显著正相关。我们或许可以据此推测, 随着森林演替的进行, 土壤N有效性将增加, 生态系统“氮饱和”程度越来越高。深层土壤δ 15 N没有表现出不同林型间的显著差异, 这与土壤垂直剖面15 N的分馏效应和N元素的迁移有关(Nadelhoffer & Fry, 1988 )。
3.3 土壤δ 13 C、δ 15 N沿垂直剖面的变化
3个林型土壤δ 13 C均沿剖面深度不断增大, 与TOC 的变化规律相反(图2 ), 这与多数研究结果相吻合(Ehleringer et al ., 2000 ; Liao et al ., 2006 )。对于这种富集机制的解释主要有以下几种假说:
1)休斯效应。工业革命以来, 贫化13 C的化石燃料的燃烧使得大气中δ 13 CO2 值逐渐降低, 底层土壤有机质δ 13 C值与早期大气δ 13 CO2 值相对应, 高于表层土壤。
2)微生物对13 C贫化基质的优先选择。不同有机化合物(糖类、纤维素、木质素、脂类等)间δ 13 C值最大差异可达10.3‰ (Hobbie & Werner, 2004 )。微生物在呼吸代谢过程中更倾向于利用环境中贫化13 C的碳源, 从而产生同位素分馏效应, 导致残留的土壤有机质13 C富集, 越往底层土壤有机质分解越彻底, 土壤δ 13 C沿剖面深度不断增大(Nadelhoffer & Fry, 1988 ; Högberg & Ekblad, 1996 )。
3)同位素动力分馏。与重同位素相比, 轻同位素活性更高, 能够更快地在产物中富集, 微生物呼吸代谢过程中12 C同位素更容易以气体形式沿土壤孔隙扩散逸出, 致使残留的土壤有机质δ 13 C增大。
4)微生物残体的贡献。Ehleringe等(2000)指出土壤中的微生物和真菌相对于它们的分解底物而言δ 13 C值更大。其原因很可能是它们在代谢过程中利用了13 C更为富集的土壤空气作为羧化反应的碳源, 微生物残体和真菌的贡献可能会使土壤有机质δ 13 C趋向更大的值(Gleixner et al ., 1993 )。
但本文的结果显示, 3个林型土壤微生物含量均沿剖面深度下降(图3B ), 与δ 13 C沿深度的变化并不一致。综上所述, 影响土壤剖面δ 13 C值变化的因素很多, 这些变化极可能是多种因素交互作用的结果。值得注意的是, 本文结果显示松林和混交林中土壤δ 13 C在土壤剖面30 cm处转而减小, 这与植被更替过程中新、老碳的混合效应有关。也有学者指出, 这是由于土壤剖面不同更新周期组分的构成和有机质分解的阶段性所致(陈庆强等, 2002 )。
土壤δ 15 N在所有林型中也表现出沿剖面增大的趋势, 与土壤TN 呈现相反的变化规律(图2 ), 这与多数学者在森林和草地中的研究结果一致(Mariotti et al ., 1980 ; Tiessen et al ., 1984 ; Shearer & Kohl, 1986 )。引起15 N沿土壤剖面富集的因素很多, 包括菌根真菌对15 N的分馏效应、贫化15 N有机化合物被优先分解利用, 以及硝化和反硝化过程中土壤N的迁移和淋溶(Hobbie & Ouimette, 2009 )。土壤经硝化和反硝化作用, 贫化15 N的气态氮和硝态氮逐渐被释放出系统, 残留的土壤有机质15 N更加富集(Ledgard et al ., 1984 ); 沿剖面深度向下, 土壤粒径越来越小, N含量越来越低, 有机质分解越来越彻底, 土壤腐殖化程度越来越高, 15 N越来越富集(Liao et al ., 2006 ); 另一方面, 植物吸收贫化15 N的NO3 - 和NH4 + 离子后又以凋落物的形式回归到土壤中, 使得表层土壤贫化15 N; 14 N和15 N在垂直剖面不断发生这样的反向迁移, 使得土壤δ 15 N沿剖面深度不断增大。
4 结论与展望
通过对鼎湖山森林演替序列植物-土壤碳氮同位素特征的研究, 得出以下结论: 1)鼎湖山植物叶片、凋落物和土壤的δ 13 C值范围分别为-33.37‰ - -29.19‰、-30.43‰ - -28.84‰和-27.31‰ - -21.38‰, 相应的δ 15 N值范围分别为-4.78‰ - -2.29‰、-5.19‰ - -4.02‰和2.52‰-8.19‰; 2)鼎湖山主要优势树种叶片δ 13 C与其C:N呈极显著正相关关系, 表明南亚热带地区植物生长并不一定受水分利用和氮素利用的补偿制约; 3)鼎湖山森林土壤和凋落物δ 13 C沿演替方向逐渐降低, δ 13 C自然丰度法的应用有助于成熟森林土壤“碳汇”机制的阐释; 4)叶片δ 15 N与叶片N含量呈正相关关系, 表层土壤和凋落物δ 15 N沿演替方向逐渐增大, 植物-土壤δ 15 N值可作为评估土壤氮素有效性和生态系统“氮饱和”状态的潜在指标; 5)土壤δ 13 C、δ 15 N沿垂直剖面呈现增大的趋势, 表明随着剖面深度增加, 土壤有机质分解更加完全。
在此研究的基础上, 我们将继续深化以下两方面的工作: 1)利用同位素手段更为精确地量化凋落物分解过程中碳的流向, 估算土壤不同有机组分的分解速率; 2)通过追踪叶片和树轮的δ 15 N值探究南亚热带森林土壤N有效性的长期变化。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Balesdent J Girardin C Mariotti A 1993 ). Site-related δ13 C of tree leaves and soil organic matter in a temperate forest.
Ecology , 74 , 1713 -1721 .
[2]
Bernoux M Cerri CC Neill C Moraes JFL 1998 ). The use of stable carbon isotopes for estimating soil organic matter turnover rates.
Geoderma , 82 , 43 -58 .
[本文引用: 1]
[3]
Blair GJ Lefroy RDB Lisle L 1995 ). Soil carbon fractions based on their degree of oxidation and the development of a carbon management index for agricultural systems.
Aus- tralian Journal of Agricultural Research , 46 , 1459 -1466 .
[本文引用: 1]
[4]
Cernusak LA Ubierna N Winter K Holtum JAM Marshall JD Farquhar GD 2013 ). Environmental and physiological determinants of carbon isotope discrimination in terrestrial plants.
New Phytologist , 200 , 950 -965 .
[本文引用: 1]
[5]
Chen H Gurmesa GA Zhang W Zhu XM Zheng MH Mao QG Zhang T Mo JM 2015 ). Nitrogen saturation in humid tropical forests after 6 years of nitrogen and phosphorus addition: Hypothesis testing.
Functional Ecology , 17 (2 ), 59 -73 .
[本文引用: 1]
[6]
Chen QQ Shen CD Peng SL Yi WX Sun YM Li ZA Jiang MT 2002 ). Characteristics and controlling factors of soil organic matter turnover processes in the subtropical mountainous area, South China.
Acta Ecologica Sinica , 22 , 1446 -1454 . (in Chinese with English abstract)[陈庆强 , 沈承德 , 彭少麟 , 易惟熙 , 孙彦敏 , 李志安 , 姜漫涛 (2002 ). 华南亚热带山地土壤有机质更新特征及其影响因子
. 生态学报 , 22 , 1446 -1454 .]
[本文引用: 1]
[7]
Chen SP Bai YF Zhang LX Han XG 2005 ). Comparing physiological responses of two dominant grass species to nitrogen addition in Xilin River Basin of China.
Environmental and Experimental Botany , 53 , 65 -75 .
[本文引用: 1]
[8]
Cheng XL Yang YH Li M Dou XL Zhang QF 2013 ). The impact of agricultural land use changes on soil organic carbon dynamics in the Danjiangkou Reservior area of China.
Plant and Soil , 366 , 415 -424 .
[本文引用: 1]
[9]
Craine JM Elmore AJ Aidar MPM Bustamante M Dawson TE Hobbie EA 2009 ). Global patterns of foliar nitrogen isotopes and their relationships with climate, mycorrhizal fungi, foliar nutrient concentrations, and nitrogen availability.
New Phytologist , 183 , 980 -992 .
[本文引用: 1]
[10]
Ehleringer JR Buchmann N Flanagan LB 2000 ). Carbon isotope ratios in belowground carbon cycle processes.
Ecological Applications , 10 , 412 -422 .
[本文引用: 2]
[11]
Ehleringer JR Lin ZF Field CB Kuo CY 1986 ). Leaf carbon isotope ratio and mineral composition in subtropical plants along an irradiance cline.
Oecologia , 72 , 109 -114 .
[本文引用: 2]
[12]
Falkengren-Grerup U Michelsen A Olsson MO Quarmby C Sleep D 2004 ). Plant nitrate use in deciduous woodland: The relationship between leaf N, 15 N natural abundance of forbs and soil N mineralisationmineralization.
Soil Biology & Biochemistry , 36 , 1885 -1891 .
[13]
Farquhar GD O’Leary MH Berry JA 1982 ). On the relationship between carbon isotope discrimination and the inter- cellular carbon dioxide concentration in leaves.
Australian Journal of Plant Physiology , 9 , 121 -137 .
[本文引用: 2]
[14]
Field C Merino J Mooney HA 1983 ). Compromises between water-use efficiency and nitrogen-use efficiency in five species of California evergreens.
Oecologia , 60 , 384 -389 .
[本文引用: 1]
[15]
Galdo ID Six J Peressotti A Cotrufo MF 2003 ). Assessing the impact of land-use change on soil sequestration in agriculture soils by means of organic matter fraction and stable C isotopes.
Global Change Biology , 9 , 1204 -1213 .
[本文引用: 1]
[16]
Garten CT Taylor GE 1992 ). Foliar δ13 C within a temperate deciduous forest: Spatial, temporal, and species sources of variation.
Oecologia , 90 , 1 -7 .
[本文引用: 1]
[17]
Gleixner G Danier HJ Werner RA Schmidt HL 1993 ). Correlations between the 13 C content of primary and secondary plant products in different cell compartments and that in decomposing Basidiomycetes.
Plant Physiology , 102 , 1287 -1290 .
[本文引用: 1]
[18]
Hobbie EA Macko SA Williams M 2000 ). Correlation be- tween foliar δ15 N and nitrogen concentrations may indicate plant-mycorrhizal interactions.
Oecologia , 122 , 273 -283 .
[本文引用: 1]
[19]
Hobbie EA Ouimette AP 2009 ). Controls of nitrogen isotope patterns in soil profiles.
Biogeochemistry , 95 , 355 -371 .
[本文引用: 1]
[20]
Hobbie EA Werner RA 2004 ). Intramolecular, compound- specific, and bulk carbon isotope patterns in C3 and C4 plants: A review and synthesis.
New Phytologist , 161 , 371 -385 .
[本文引用: 1]
[21]
Högberg P 1997 ). 15 N natural abundance in soil-plant systems.
New Phytologist , 137 , 179 -203 .
[本文引用: 1]
[22]
Högberg P Ekblad A 1996 ). Substrate-induced respiration measured in situ in a C3 -plant ecosystem using additions of C4 -sucrose.
Soil Biology & Biochemistry , 28 , 1131 -1138 .
[本文引用: 2]
[23]
Huang YH Li YL Xiao Y Wenigmann KO Zhou GY Zhang DQ Wenigmann M Tang XL Liu JX 2011 ). Controls of litter quality on the carbon sink in soils through partitioning the products of decomposing litter in a forest succession series in South China.
Forest Ecology and Management , 261 , 1170 -1177 .
[24]
Huang ZL Kong GH Zhang QM Liu SZ 1998 ). Structure, species diversity and population dynamics of the lower subtropical evergreen broad-leaved forest in Dinghushan Biosphere Reserve.
Tropical and Subtropical Forest Ecosystem , 1 , 64 -75 . (in Chinese with English abstract)[黄忠良 , 孔国辉 , 张倩媚 , 刘世忠 (1998 ). 鼎湖山南亚热带常绿阔叶林结构、物种多样性及种群动态的研究
. 热带亚热带森林生态系统研究 , 1 , 64 -75 .]
[本文引用: 1]
[25]
Kahmen A Wanek W Buchmann N 2008 ). Foliar δ15 N values characterize soil N cycling and reflect nitrate or ammonium preference of plants along a temperate grassland gradient.
Oecologia , 156 , 861 -870 .
[本文引用: 1]
[26]
Ledgard SF Feney JR Simpson JR 1984 ). Variations in natural enrichment of 15 N in the profiles of some Australian pasture soils.
Australian Journal of Soil Research , 22 , 155 -164 .
[本文引用: 1]
[27]
Liao JD Boutton TW Jastrow JD 2006 ). Organic matter turnover in soil physical fractions following woody plant invasion of grassland: Evidence from natural 13 C and 15 N.
Soil Biology & Biochemistry , 38 , 3197 -3210 .
[本文引用: 4]
[28]
Liu XD Zhou GY Chen XZ Zhang DQ Zhang QM 2014 ). Forest microclimate change along with the succession and response to climate change in south subtropical region.
Acta Ecologica Sinica , 34 , 2755 -2764 . (in Chinese with English abstract)[刘效东 , 周国逸 , 陈修治 , 张德强 , 张倩媚 (2014 ). 南亚热带森林演替过程中小气候的改变及对气候变化的响应
. 生态学报 , 34 , 2755 -2764 .]
[本文引用: 1]
[29]
Mariotti A Pierre D Vedy JC Bruckert S Guillemot J 1980 ). The abundance of natural nitrogen 15 in the organic matter of soils along an altitudinal gradient.
Catena , 7 , 293 -300 .
[本文引用: 1]
[30]
Nadelhoffer KJ Fry B 1988 ). Controls on natural nitrogen-15 and carbon-13 abundances in forest soil organic matter.
Soil Science Society of America Journal , 52 , 1633 -1640 .
[本文引用: 3]
[31]
O’Leary MH 1981 ). Carbon isotope fractionation in plants.
Phytochemistry , 20 , 553 -567 .
[本文引用: 1]
[32]
O’Leary MH 1988 ). Carbon isotopes in photosynthesis.
Bioscience , 38 , 328 -336 .
[本文引用: 1]
[33]
Ouyang X Li YL Zhang QM 2014 ). Characteristics of microclimate in a mixed coniferous and broadleaf forest in Dinghushan Biosphere Reserve.
Chinese Journal of Ecology , 33 , 575 -582 . (in Chinese with English abstract)[欧阳旭 , 李跃林 , 张倩媚 (2014 ). 鼎湖山针阔叶混交林小气候调节效应
. 生态学杂志 , 33 , 575 -582 .]
[本文引用: 1]
[34]
Pardo LH Hemond HF Montoya JP Fahey TJ Siccama TG 2002 ). Response of the natural abundance of 15 N in forest soils and foliage to high nitrate loss following clear cutting.
Canadian Journal of Forest Research , 32 , 1126 -1136 .
[本文引用: 1]
[35]
Shearer G Kohl DH 1986 ). N2 -fixation in field settings: Estimations based on natural 15 N abundance.
Australian Journal of Plant Physiology , 13 , 699 -756 .
[本文引用: 1]
[36]
Sun GC Lin ZF Lin GZ Li SS 1993 ). 13 C/12 C ratio and water use efficiency of Pinus massoniana in subtropical artificial forest.
Chinese Journal of Applied Ecology , 4 , 325 -327 . (in Chinese with English abstract)[孙谷畴 , 林植芳 , 林桂珠 , 李双顺 (1993 ). 亚热带人工林松树13 C/12 C比率和水分利用效率
. 应用生态学报 , 4 , 325 -327 .]
[37]
Tcherkez G Hodges M 2008 ). How stable isotopes may help to elucidate primary nitrogen metabolism and its interaction with (photo) respiration in C3 leaves.
Journal of Experimental Botany , 59 , 941 -953 .
[本文引用: 1]
[38]
Templer PH Arthur MA Lovett GM Weathers KC 2007 ). Plant and soil natural abundance δ15 N: Indicators of relative rates of nitrogen cycling in temperate forest ecosystems.
Oecologia , 153 , 399 -406 .
[本文引用: 1]
[39]
Tiessen H Karamanos RE Stewart JWB Selles F 1984 ). Natural nitrogen-15 abundance as an indicator of soil organic matter transformations in native and cultivated soils.
Soil Science Society of America Journal , 48 , 312 -315 .
[本文引用: 1]
[40]
Tsialtas JT Handley LL Kassioumi MT Veresoglou DS Gagianas AA 2001 ). Interspecific variation in potential water use efficiency and its relation to plant species abundance in a water-limited grassland.
Functional Ecology , 15 , 605 -614 .
[41]
Vance ED Brookes PC Jenkinson DS 1987 ). An extraction method for measuring soil microbial biomass C.
Soil Biology & Biochemistry , 19 , 703 -707 .
[本文引用: 1]
[42]
Xu YQ He JC Cheng WX Xing XR Li LH 2010 ). Natural 15 N abundance in soils and plants in relation to N cycling in a rangeland in Inner Mongolia.
Journal of Plant Ecology , 3 , 201 -207 .
[本文引用: 2]
[43]
Yu GR Wang SQ Chen PQ Li QK 2005 ). Isotope tracer approaches in soil organic carbon cycle research.
Advances in Earth Science , 20 , 568 -577 . (in Chinese with English abstract)[于贵瑞 , 王绍强 , 陈泮勤 , 李庆康 (2005 ). 碳同位素技术在土壤碳循环研究中的应用
. 地球科学进展 , 20 , 568 -577 .]
[本文引用: 1]
[44]
Zhang J Gu L Bao F Cao Y Hao Y He J Li J Li Y Ren Y Wang F Wu R Yao B Zhao Y Lin G Wu B Lu Q Meng P 2015 ). Nitrogen control of 13 C enrichment in heterotrophic organs relative to leaves in a landscape- building desert plant species.
Biogeosciences , 12 , 15 -27 .
[本文引用: 1]
[45]
Zhou GY Liu SG Li ZA Zhang DQ Tang XL Zhou CY Yan JH Mo JM 2006 a). Old-growth forests can accumulate carbon in soils.
Science , 314 , 1417 .
[46]
Zhou GY Zhou CY Liu SG Tang XL Ouyang XJ Zhang DQ Liu SZ Liu JX Yan JH Wen DZ Xu GL Zhou CY Luo Y Guan LL Liu Y 2006 b). Belowground carbon balance and carbon accumulation rate in the successional serials of monsoon evergreen broad-leaved forest.
Science in China Serial D-Earth Science , 49 , 311 -321 .
Site-related δ13 C of tree leaves and soil organic matter in a temperate forest.
1993
The use of stable carbon isotopes for estimating soil organic matter turnover rates.
1
1998
... 碳(C)、氮(N)作为两种重要的生命元素, 其生物地球化学循环过程一直是生态学研究的重点, 二者在自然界都有两种稳定性同位素(12 C、13 C和14 N、15 N).由于同位素分馏效应, 不同物质间C、N同位素自然丰度存在差异, 这为同位素技术在生态系统碳-氮循环过程中的应用提供了理论基础(Nadelhoffer & Fry, 1988 ).13 C同位素丰度最早被用于区分植物的光合途径(O’Leary, 1981 ), 其后又被发现与植物光合代谢和水分利用效率之间的关系(Farquhar et al ., 1982 ; Ehleringer et al ., 1986 ), 并被广泛应用于土壤碳循环和植物功能群的研究(Garten & Taylor, 1992 ; 于贵瑞等, 2005 ).在发生C3 -C4 植被转换的区域, 通过土壤稳定碳同位素比率(δ 13 C)在时间上的变化, 可以区分不同来源有机质的相对贡献, 同时也可反演植被覆盖的历史, 同样的方法也用于土壤不同有机组分周转过程的研究(Bernoux et al ., 1998 ; Ehleringer et al ., 2000 ).植物-土壤稳定氮同位素比率(δ 15 N)被认为可以用来评估生态系统的N通量, 且已被部分学者用作生态系统氮饱和状态的指示指标(Högberg, 1997 ; Pardo et al ., 2002 ; Kahmen et al ., 2008 ; Xu et al ., 2010 ).Templer等(2007)研究发现土壤δ 15 N值可以表征土壤的净矿化和硝化速率; Craine等(2009)认为植物-土壤δ 15 N高的地区土壤N有效性也更高, 植物根系对菌根真菌的依赖度更弱, 生态系统氮的饱和度更高.显然, 稳定同位素技术的应用为生态系统C、N元素循环的研究提供了一个新的窗口.但目前δ 13 C自然丰度法多应用于发生过C3 -C4 植被转换区域的碳周转研究(Galdo et al ., 2003 ; Liao et al ., 2006 ; Cheng et al ., 2013 ), 而在植被系统未发生C3 -C4 型转换的亚热带森林, 如何应用δ 13 C自然丰度法探究森林演替过程中土壤碳周转的变化, 仍需更多的探索. ...
Soil carbon fractions based on their degree of oxidation and the development of a carbon management index for agricultural systems.
1
1995
... 式中, R = 13 C/12 C或15 N/14 N, R sam 表示样品的相对丰度, R std 表示国际标准物的相对丰度.碳同位素国际标准物为Pee Dee Belemnite (PDB), 即美国南卡罗来纳州白垩系皮狄组地层中的美洲拟箭石(R PDB = 0.0112372, δ 13 CPDB = 0‰); 氮同位素国际标准物为标准大气中的N2 (R air = 0.0036765, δ 15 Nair = 0‰).本次实验所得同位素比值δ 的测定精度为± 0.1‰, 由中国科学院华南植物园公共实验室提供.土壤微生物生物量碳的测定采用氯仿熏蒸-K2 SO4 浸提法(Vance et al ., 1987 ).土壤易氧化有机碳的测定采用高锰酸钾氧化法(Blair et al ., 1995 ). ...
Environmental and physiological determinants of carbon isotope discrimination in terrestrial plants.
1
2013
... 显著负相关, 与C:N极显著正相关, 与Tsialtas等(2001)的结果一致, 而与其他的一些结论相反(Cernusak et al ., 2013 ; Zhang et al ., 2015 ), 这可能与不同环境条件下土壤养分供应水平及物种在群落中的资源利用策略有关.理论上, 在“氮限制”的环境下, 叶片N含量的增加有利于提高核酮糖1,5-二磷酸羧化氧化酶(Rubisco)的数量和活性, 光合速率加快, C i /C a 降低, 叶片δ 13 C增大(O’Leary, 1988 ).而南亚热带地区氮沉降水平高, 生态系统趋于“氮饱和”状态, 植物的生长转而受到其他营养元素(如磷)的限制(Chen et al ., 2015 ); 鼎湖山降水充沛, 土壤含水量高, 更多的N吸收意味着更多水分的摄入, 从而导致叶片气孔导度增大, C i /C a 升高, δ 13 C降低.以上结果表明: 出于对湿润气候和高氮沉降水平的一种适应策略, 南亚热带森林群落中植物可能同时具有较低的水分利用效率和氮素利用效率(C:N), 并不一定存在相互制约的补偿机制. ...
Nitrogen saturation in humid tropical forests after 6 years of nitrogen and phosphorus addition: Hypothesis testing.
1
2015
... 显著负相关, 与C:N极显著正相关, 与Tsialtas等(2001)的结果一致, 而与其他的一些结论相反(Cernusak et al ., 2013 ; Zhang et al ., 2015 ), 这可能与不同环境条件下土壤养分供应水平及物种在群落中的资源利用策略有关.理论上, 在“氮限制”的环境下, 叶片N含量的增加有利于提高核酮糖1,5-二磷酸羧化氧化酶(Rubisco)的数量和活性, 光合速率加快, C i /C a 降低, 叶片δ 13 C增大(O’Leary, 1988 ).而南亚热带地区氮沉降水平高, 生态系统趋于“氮饱和”状态, 植物的生长转而受到其他营养元素(如磷)的限制(Chen et al ., 2015 ); 鼎湖山降水充沛, 土壤含水量高, 更多的N吸收意味着更多水分的摄入, 从而导致叶片气孔导度增大, C i /C a 升高, δ 13 C降低.以上结果表明: 出于对湿润气候和高氮沉降水平的一种适应策略, 南亚热带森林群落中植物可能同时具有较低的水分利用效率和氮素利用效率(C:N), 并不一定存在相互制约的补偿机制. ...
华南亚热带山地土壤有机质更新特征及其影响因子
1
2002
... 但本文的结果显示, 3个林型土壤微生物含量均沿剖面深度下降(图3B ), 与δ 13 C沿深度的变化并不一致.综上所述, 影响土壤剖面δ 13 C值变化的因素很多, 这些变化极可能是多种因素交互作用的结果.值得注意的是, 本文结果显示松林和混交林中土壤δ 13 C在土壤剖面30 cm处转而减小, 这与植被更替过程中新、老碳的混合效应有关.也有学者指出, 这是由于土壤剖面不同更新周期组分的构成和有机质分解的阶段性所致(陈庆强等, 2002 ). ...
Comparing physiological responses of two dominant grass species to nitrogen addition in Xilin River Basin of China.
1
2005
... 尽管已有研究指出, 在地上部分生物量不再增加的情况下, 南亚热带成熟森林土壤依然具有持续积累有机碳的潜力(Zhou et al ., 2006a ), 但对于这种积累机制的解释仍存有很多疑问.诸多研究表明, 具有较高水分利用效率的植物往往氮素利用效率较低, 即在自然条件下, 植物对水分和氮素的利用效率存在补偿制约, 这种制约关系在草原和温带森林生态系统中都得到验证(Field et al ., 1983 ; Chen et al ., 2005 ).而在雨量充沛以及高氮沉降的南亚热带森林, 植物对水分和氮素利用效率的制约关系是否依然存在? 另外,该区域森林生态系统是否已达到“氮饱和”状态? 森林演替过程中土壤氮素有效性如何变化? 这些都是森林生态学研究亟待解决的关键问题.然而, 上述生态学过程和现象的变化在短时间内通过传统的研究手段难以被精确观测.δ 13 C、δ 15 N自然丰度法的应用或许能为我们提供更多有价值的信息.本文选取鼎湖山森林演替序列中3个代表林型——马尾松(Pinus massoniana )针叶林(简称松林)、针阔叶混交林(简称混交林)和季风常绿阔叶林(简称阔叶林), 分别代表演替水平的前、中、后期(Zhou et al ., 2006b ), 通过测定植物-土壤δ 13 C和δ 15 N, 分析叶片δ 13 C、δ 15 N值与其叶片元素含量的关系, 揭示植物-土壤δ 13 C、δ 15 N在演替水平和垂直方向上的变化特征, 试图阐明南亚热带森林演替过程中植物群落对资源的利用策略, 为生态系统碳氮元素循环过程机理的研究提供支持. ...
The impact of agricultural land use changes on soil organic carbon dynamics in the Danjiangkou Reservior area of China.
1
2013
... 碳(C)、氮(N)作为两种重要的生命元素, 其生物地球化学循环过程一直是生态学研究的重点, 二者在自然界都有两种稳定性同位素(12 C、13 C和14 N、15 N).由于同位素分馏效应, 不同物质间C、N同位素自然丰度存在差异, 这为同位素技术在生态系统碳-氮循环过程中的应用提供了理论基础(Nadelhoffer & Fry, 1988 ).13 C同位素丰度最早被用于区分植物的光合途径(O’Leary, 1981 ), 其后又被发现与植物光合代谢和水分利用效率之间的关系(Farquhar et al ., 1982 ; Ehleringer et al ., 1986 ), 并被广泛应用于土壤碳循环和植物功能群的研究(Garten & Taylor, 1992 ; 于贵瑞等, 2005 ).在发生C3 -C4 植被转换的区域, 通过土壤稳定碳同位素比率(δ 13 C)在时间上的变化, 可以区分不同来源有机质的相对贡献, 同时也可反演植被覆盖的历史, 同样的方法也用于土壤不同有机组分周转过程的研究(Bernoux et al ., 1998 ; Ehleringer et al ., 2000 ).植物-土壤稳定氮同位素比率(δ 15 N)被认为可以用来评估生态系统的N通量, 且已被部分学者用作生态系统氮饱和状态的指示指标(Högberg, 1997 ; Pardo et al ., 2002 ; Kahmen et al ., 2008 ; Xu et al ., 2010 ).Templer等(2007)研究发现土壤δ 15 N值可以表征土壤的净矿化和硝化速率; Craine等(2009)认为植物-土壤δ 15 N高的地区土壤N有效性也更高, 植物根系对菌根真菌的依赖度更弱, 生态系统氮的饱和度更高.显然, 稳定同位素技术的应用为生态系统C、N元素循环的研究提供了一个新的窗口.但目前δ 13 C自然丰度法多应用于发生过C3 -C4 植被转换区域的碳周转研究(Galdo et al ., 2003 ; Liao et al ., 2006 ; Cheng et al ., 2013 ), 而在植被系统未发生C3 -C4 型转换的亚热带森林, 如何应用δ 13 C自然丰度法探究森林演替过程中土壤碳周转的变化, 仍需更多的探索. ...
Global patterns of foliar nitrogen isotopes and their relationships with climate, mycorrhizal fungi, foliar nutrient concentrations, and nitrogen availability.
1
2009
... 除少数固氮植物外, 植物氮素主要来源于根系吸收土壤中的硝态氮和铵态氮, 再经过同化还原作用固定在有机质中, 这两个阶段都会产生氮同位素分馏, 其分馏效应主要取决于土壤N有效性以及植物对氮素的需求(Tcherkez & Hodges, 2008 ).许多研究已经在不同空间尺度上证明叶片δ 15 N与其N含量正相关(Hobbie et al ., 2000 ; Craine et al ., 2009 ), 本文的结果与之相吻合(图1E ), Craine等(2009)认为叶片N含量高的地区土壤N有效性高, 系统N循环开放度更高, 更多贫化15 N的气态N (如NH3 、N2 O和N2 等)被释放, 系统富集15 N; 同时植物对菌根真菌的依赖度下降, N吸收过程中同位素分馏效应减弱, 导致植物δ 15 N增大.表1 结果显示马尾松叶片δ 15 N值在混交林中较松林中显著增大, 可以解释为混交林土壤N有效性明显高于松林, 土壤中氮的净矿化和硝化速率更快(Templer et al ., 2007 ).而混交林和阔叶林的3个共有优势种, 除木荷外, 锥和黄果厚壳桂叶片δ 15 N值均没有表现出林型间的差异, 表明处于演替中期的混交林土壤N有效性越来越接近阔叶林土壤N有效性水平. ...
Carbon isotope ratios in belowground carbon cycle processes.
2
2000
... 碳(C)、氮(N)作为两种重要的生命元素, 其生物地球化学循环过程一直是生态学研究的重点, 二者在自然界都有两种稳定性同位素(12 C、13 C和14 N、15 N).由于同位素分馏效应, 不同物质间C、N同位素自然丰度存在差异, 这为同位素技术在生态系统碳-氮循环过程中的应用提供了理论基础(Nadelhoffer & Fry, 1988 ).13 C同位素丰度最早被用于区分植物的光合途径(O’Leary, 1981 ), 其后又被发现与植物光合代谢和水分利用效率之间的关系(Farquhar et al ., 1982 ; Ehleringer et al ., 1986 ), 并被广泛应用于土壤碳循环和植物功能群的研究(Garten & Taylor, 1992 ; 于贵瑞等, 2005 ).在发生C3 -C4 植被转换的区域, 通过土壤稳定碳同位素比率(δ 13 C)在时间上的变化, 可以区分不同来源有机质的相对贡献, 同时也可反演植被覆盖的历史, 同样的方法也用于土壤不同有机组分周转过程的研究(Bernoux et al ., 1998 ; Ehleringer et al ., 2000 ).植物-土壤稳定氮同位素比率(δ 15 N)被认为可以用来评估生态系统的N通量, 且已被部分学者用作生态系统氮饱和状态的指示指标(Högberg, 1997 ; Pardo et al ., 2002 ; Kahmen et al ., 2008 ; Xu et al ., 2010 ).Templer等(2007)研究发现土壤δ 15 N值可以表征土壤的净矿化和硝化速率; Craine等(2009)认为植物-土壤δ 15 N高的地区土壤N有效性也更高, 植物根系对菌根真菌的依赖度更弱, 生态系统氮的饱和度更高.显然, 稳定同位素技术的应用为生态系统C、N元素循环的研究提供了一个新的窗口.但目前δ 13 C自然丰度法多应用于发生过C3 -C4 植被转换区域的碳周转研究(Galdo et al ., 2003 ; Liao et al ., 2006 ; Cheng et al ., 2013 ), 而在植被系统未发生C3 -C4 型转换的亚热带森林, 如何应用δ 13 C自然丰度法探究森林演替过程中土壤碳周转的变化, 仍需更多的探索. ...
... 3个林型土壤δ 13 C均沿剖面深度不断增大, 与TOC 的变化规律相反(图2 ), 这与多数研究结果相吻合(Ehleringer et al ., 2000 ; Liao et al ., 2006 ).对于这种富集机制的解释主要有以下几种假说: ...
Leaf carbon isotope ratio and mineral composition in subtropical plants along an irradiance cline.
2
1986
... 碳(C)、氮(N)作为两种重要的生命元素, 其生物地球化学循环过程一直是生态学研究的重点, 二者在自然界都有两种稳定性同位素(12 C、13 C和14 N、15 N).由于同位素分馏效应, 不同物质间C、N同位素自然丰度存在差异, 这为同位素技术在生态系统碳-氮循环过程中的应用提供了理论基础(Nadelhoffer & Fry, 1988 ).13 C同位素丰度最早被用于区分植物的光合途径(O’Leary, 1981 ), 其后又被发现与植物光合代谢和水分利用效率之间的关系(Farquhar et al ., 1982 ; Ehleringer et al ., 1986 ), 并被广泛应用于土壤碳循环和植物功能群的研究(Garten & Taylor, 1992 ; 于贵瑞等, 2005 ).在发生C3 -C4 植被转换的区域, 通过土壤稳定碳同位素比率(δ 13 C)在时间上的变化, 可以区分不同来源有机质的相对贡献, 同时也可反演植被覆盖的历史, 同样的方法也用于土壤不同有机组分周转过程的研究(Bernoux et al ., 1998 ; Ehleringer et al ., 2000 ).植物-土壤稳定氮同位素比率(δ 15 N)被认为可以用来评估生态系统的N通量, 且已被部分学者用作生态系统氮饱和状态的指示指标(Högberg, 1997 ; Pardo et al ., 2002 ; Kahmen et al ., 2008 ; Xu et al ., 2010 ).Templer等(2007)研究发现土壤δ 15 N值可以表征土壤的净矿化和硝化速率; Craine等(2009)认为植物-土壤δ 15 N高的地区土壤N有效性也更高, 植物根系对菌根真菌的依赖度更弱, 生态系统氮的饱和度更高.显然, 稳定同位素技术的应用为生态系统C、N元素循环的研究提供了一个新的窗口.但目前δ 13 C自然丰度法多应用于发生过C3 -C4 植被转换区域的碳周转研究(Galdo et al ., 2003 ; Liao et al ., 2006 ; Cheng et al ., 2013 ), 而在植被系统未发生C3 -C4 型转换的亚热带森林, 如何应用δ 13 C自然丰度法探究森林演替过程中土壤碳周转的变化, 仍需更多的探索. ...
... 多数研究结果证实, C3 植物的碳同位素分馏效应与叶片胞间CO2 浓度和大气CO2 浓度的比值(C i /C a )具有明显的线性正相关关系(Farquhar et al ., 1982 ), 而C i /C a 受光合速率和气孔导度的影响, 因此, 叶片δ 13 C与植物光合和蒸腾等生理过程有着密切的联系, 可以作为估测植物长期水分利用效率(WUE )的重要指标(Ehleringer et al ., 1986 ).本文结果显示混交林中马尾松叶片δ 13 C显著低于松林, 这与演替过程中林内小气候的变化相关.随着演替的进行, 森林郁闭度增加, 林内光照水平减弱, 气温降低, 林内空气湿度和土壤含水量升高(刘效东等, 2014 ), 由此引起植物光合和蒸腾速率的改变, 植物WUE 下降, 叶片C i /C a 升高, δ 13 C值降低.这与Ehleringer等(1986)在鼎湖山的研究结果相吻合, 孙谷畴等(1993)对亚热带人工松林的研究也得到类似的结论.马尾松作为喜光性的耐旱先锋种, 在荫蔽潮湿的环境下优势度减弱, 逐渐被阔叶树种所替代, 群落结构逐步演化成针阔叶混交林.而混交林和阔叶林的3个共有树种叶片δ 13 C均没有表现出林型间的显著差异, 这或许可以间接证明处于演替中、后期的两林型间林分差异越来越小, 混交林林内气温、空气湿度、土壤含水量及植物生理功能逐渐趋同于阔叶林(监测数据, 未发表), 混交林正快速向演替顶级过渡(Zhou et al ., 2006b ). ...
Plant nitrate use in deciduous woodland: The relationship between leaf N, 15 N natural abundance of forbs and soil N mineralisationmineralization.
2004
On the relationship between carbon isotope discrimination and the inter- cellular carbon dioxide concentration in leaves.
2
1982
... 碳(C)、氮(N)作为两种重要的生命元素, 其生物地球化学循环过程一直是生态学研究的重点, 二者在自然界都有两种稳定性同位素(12 C、13 C和14 N、15 N).由于同位素分馏效应, 不同物质间C、N同位素自然丰度存在差异, 这为同位素技术在生态系统碳-氮循环过程中的应用提供了理论基础(Nadelhoffer & Fry, 1988 ).13 C同位素丰度最早被用于区分植物的光合途径(O’Leary, 1981 ), 其后又被发现与植物光合代谢和水分利用效率之间的关系(Farquhar et al ., 1982 ; Ehleringer et al ., 1986 ), 并被广泛应用于土壤碳循环和植物功能群的研究(Garten & Taylor, 1992 ; 于贵瑞等, 2005 ).在发生C3 -C4 植被转换的区域, 通过土壤稳定碳同位素比率(δ 13 C)在时间上的变化, 可以区分不同来源有机质的相对贡献, 同时也可反演植被覆盖的历史, 同样的方法也用于土壤不同有机组分周转过程的研究(Bernoux et al ., 1998 ; Ehleringer et al ., 2000 ).植物-土壤稳定氮同位素比率(δ 15 N)被认为可以用来评估生态系统的N通量, 且已被部分学者用作生态系统氮饱和状态的指示指标(Högberg, 1997 ; Pardo et al ., 2002 ; Kahmen et al ., 2008 ; Xu et al ., 2010 ).Templer等(2007)研究发现土壤δ 15 N值可以表征土壤的净矿化和硝化速率; Craine等(2009)认为植物-土壤δ 15 N高的地区土壤N有效性也更高, 植物根系对菌根真菌的依赖度更弱, 生态系统氮的饱和度更高.显然, 稳定同位素技术的应用为生态系统C、N元素循环的研究提供了一个新的窗口.但目前δ 13 C自然丰度法多应用于发生过C3 -C4 植被转换区域的碳周转研究(Galdo et al ., 2003 ; Liao et al ., 2006 ; Cheng et al ., 2013 ), 而在植被系统未发生C3 -C4 型转换的亚热带森林, 如何应用δ 13 C自然丰度法探究森林演替过程中土壤碳周转的变化, 仍需更多的探索. ...
... 多数研究结果证实, C3 植物的碳同位素分馏效应与叶片胞间CO2 浓度和大气CO2 浓度的比值(C i /C a )具有明显的线性正相关关系(Farquhar et al ., 1982 ), 而C i /C a 受光合速率和气孔导度的影响, 因此, 叶片δ 13 C与植物光合和蒸腾等生理过程有着密切的联系, 可以作为估测植物长期水分利用效率(WUE )的重要指标(Ehleringer et al ., 1986 ).本文结果显示混交林中马尾松叶片δ 13 C显著低于松林, 这与演替过程中林内小气候的变化相关.随着演替的进行, 森林郁闭度增加, 林内光照水平减弱, 气温降低, 林内空气湿度和土壤含水量升高(刘效东等, 2014 ), 由此引起植物光合和蒸腾速率的改变, 植物WUE 下降, 叶片C i /C a 升高, δ 13 C值降低.这与Ehleringer等(1986)在鼎湖山的研究结果相吻合, 孙谷畴等(1993)对亚热带人工松林的研究也得到类似的结论.马尾松作为喜光性的耐旱先锋种, 在荫蔽潮湿的环境下优势度减弱, 逐渐被阔叶树种所替代, 群落结构逐步演化成针阔叶混交林.而混交林和阔叶林的3个共有树种叶片δ 13 C均没有表现出林型间的显著差异, 这或许可以间接证明处于演替中、后期的两林型间林分差异越来越小, 混交林林内气温、空气湿度、土壤含水量及植物生理功能逐渐趋同于阔叶林(监测数据, 未发表), 混交林正快速向演替顶级过渡(Zhou et al ., 2006b ). ...
Compromises between water-use efficiency and nitrogen-use efficiency in five species of California evergreens.
1
1983
... 尽管已有研究指出, 在地上部分生物量不再增加的情况下, 南亚热带成熟森林土壤依然具有持续积累有机碳的潜力(Zhou et al ., 2006a ), 但对于这种积累机制的解释仍存有很多疑问.诸多研究表明, 具有较高水分利用效率的植物往往氮素利用效率较低, 即在自然条件下, 植物对水分和氮素的利用效率存在补偿制约, 这种制约关系在草原和温带森林生态系统中都得到验证(Field et al ., 1983 ; Chen et al ., 2005 ).而在雨量充沛以及高氮沉降的南亚热带森林, 植物对水分和氮素利用效率的制约关系是否依然存在? 另外,该区域森林生态系统是否已达到“氮饱和”状态? 森林演替过程中土壤氮素有效性如何变化? 这些都是森林生态学研究亟待解决的关键问题.然而, 上述生态学过程和现象的变化在短时间内通过传统的研究手段难以被精确观测.δ 13 C、δ 15 N自然丰度法的应用或许能为我们提供更多有价值的信息.本文选取鼎湖山森林演替序列中3个代表林型——马尾松(Pinus massoniana )针叶林(简称松林)、针阔叶混交林(简称混交林)和季风常绿阔叶林(简称阔叶林), 分别代表演替水平的前、中、后期(Zhou et al ., 2006b ), 通过测定植物-土壤δ 13 C和δ 15 N, 分析叶片δ 13 C、δ 15 N值与其叶片元素含量的关系, 揭示植物-土壤δ 13 C、δ 15 N在演替水平和垂直方向上的变化特征, 试图阐明南亚热带森林演替过程中植物群落对资源的利用策略, 为生态系统碳氮元素循环过程机理的研究提供支持. ...
Assessing the impact of land-use change on soil sequestration in agriculture soils by means of organic matter fraction and stable C isotopes.
1
2003
... 碳(C)、氮(N)作为两种重要的生命元素, 其生物地球化学循环过程一直是生态学研究的重点, 二者在自然界都有两种稳定性同位素(12 C、13 C和14 N、15 N).由于同位素分馏效应, 不同物质间C、N同位素自然丰度存在差异, 这为同位素技术在生态系统碳-氮循环过程中的应用提供了理论基础(Nadelhoffer & Fry, 1988 ).13 C同位素丰度最早被用于区分植物的光合途径(O’Leary, 1981 ), 其后又被发现与植物光合代谢和水分利用效率之间的关系(Farquhar et al ., 1982 ; Ehleringer et al ., 1986 ), 并被广泛应用于土壤碳循环和植物功能群的研究(Garten & Taylor, 1992 ; 于贵瑞等, 2005 ).在发生C3 -C4 植被转换的区域, 通过土壤稳定碳同位素比率(δ 13 C)在时间上的变化, 可以区分不同来源有机质的相对贡献, 同时也可反演植被覆盖的历史, 同样的方法也用于土壤不同有机组分周转过程的研究(Bernoux et al ., 1998 ; Ehleringer et al ., 2000 ).植物-土壤稳定氮同位素比率(δ 15 N)被认为可以用来评估生态系统的N通量, 且已被部分学者用作生态系统氮饱和状态的指示指标(Högberg, 1997 ; Pardo et al ., 2002 ; Kahmen et al ., 2008 ; Xu et al ., 2010 ).Templer等(2007)研究发现土壤δ 15 N值可以表征土壤的净矿化和硝化速率; Craine等(2009)认为植物-土壤δ 15 N高的地区土壤N有效性也更高, 植物根系对菌根真菌的依赖度更弱, 生态系统氮的饱和度更高.显然, 稳定同位素技术的应用为生态系统C、N元素循环的研究提供了一个新的窗口.但目前δ 13 C自然丰度法多应用于发生过C3 -C4 植被转换区域的碳周转研究(Galdo et al ., 2003 ; Liao et al ., 2006 ; Cheng et al ., 2013 ), 而在植被系统未发生C3 -C4 型转换的亚热带森林, 如何应用δ 13 C自然丰度法探究森林演替过程中土壤碳周转的变化, 仍需更多的探索. ...
Foliar δ13 C within a temperate deciduous forest: Spatial, temporal, and species sources of variation.
1
1992
... 碳(C)、氮(N)作为两种重要的生命元素, 其生物地球化学循环过程一直是生态学研究的重点, 二者在自然界都有两种稳定性同位素(12 C、13 C和14 N、15 N).由于同位素分馏效应, 不同物质间C、N同位素自然丰度存在差异, 这为同位素技术在生态系统碳-氮循环过程中的应用提供了理论基础(Nadelhoffer & Fry, 1988 ).13 C同位素丰度最早被用于区分植物的光合途径(O’Leary, 1981 ), 其后又被发现与植物光合代谢和水分利用效率之间的关系(Farquhar et al ., 1982 ; Ehleringer et al ., 1986 ), 并被广泛应用于土壤碳循环和植物功能群的研究(Garten & Taylor, 1992 ; 于贵瑞等, 2005 ).在发生C3 -C4 植被转换的区域, 通过土壤稳定碳同位素比率(δ 13 C)在时间上的变化, 可以区分不同来源有机质的相对贡献, 同时也可反演植被覆盖的历史, 同样的方法也用于土壤不同有机组分周转过程的研究(Bernoux et al ., 1998 ; Ehleringer et al ., 2000 ).植物-土壤稳定氮同位素比率(δ 15 N)被认为可以用来评估生态系统的N通量, 且已被部分学者用作生态系统氮饱和状态的指示指标(Högberg, 1997 ; Pardo et al ., 2002 ; Kahmen et al ., 2008 ; Xu et al ., 2010 ).Templer等(2007)研究发现土壤δ 15 N值可以表征土壤的净矿化和硝化速率; Craine等(2009)认为植物-土壤δ 15 N高的地区土壤N有效性也更高, 植物根系对菌根真菌的依赖度更弱, 生态系统氮的饱和度更高.显然, 稳定同位素技术的应用为生态系统C、N元素循环的研究提供了一个新的窗口.但目前δ 13 C自然丰度法多应用于发生过C3 -C4 植被转换区域的碳周转研究(Galdo et al ., 2003 ; Liao et al ., 2006 ; Cheng et al ., 2013 ), 而在植被系统未发生C3 -C4 型转换的亚热带森林, 如何应用δ 13 C自然丰度法探究森林演替过程中土壤碳周转的变化, 仍需更多的探索. ...
Correlations between the 13 C content of primary and secondary plant products in different cell compartments and that in decomposing Basidiomycetes.
1
1993
... 4)微生物残体的贡献.Ehleringe等(2000)指出土壤中的微生物和真菌相对于它们的分解底物而言δ 13 C值更大.其原因很可能是它们在代谢过程中利用了13 C更为富集的土壤空气作为羧化反应的碳源, 微生物残体和真菌的贡献可能会使土壤有机质δ 13 C趋向更大的值(Gleixner et al ., 1993 ). ...
Correlation be- tween foliar δ15 N and nitrogen concentrations may indicate plant-mycorrhizal interactions.
1
2000
... 除少数固氮植物外, 植物氮素主要来源于根系吸收土壤中的硝态氮和铵态氮, 再经过同化还原作用固定在有机质中, 这两个阶段都会产生氮同位素分馏, 其分馏效应主要取决于土壤N有效性以及植物对氮素的需求(Tcherkez & Hodges, 2008 ).许多研究已经在不同空间尺度上证明叶片δ 15 N与其N含量正相关(Hobbie et al ., 2000 ; Craine et al ., 2009 ), 本文的结果与之相吻合(图1E ), Craine等(2009)认为叶片N含量高的地区土壤N有效性高, 系统N循环开放度更高, 更多贫化15 N的气态N (如NH3 、N2 O和N2 等)被释放, 系统富集15 N; 同时植物对菌根真菌的依赖度下降, N吸收过程中同位素分馏效应减弱, 导致植物δ 15 N增大.表1 结果显示马尾松叶片δ 15 N值在混交林中较松林中显著增大, 可以解释为混交林土壤N有效性明显高于松林, 土壤中氮的净矿化和硝化速率更快(Templer et al ., 2007 ).而混交林和阔叶林的3个共有优势种, 除木荷外, 锥和黄果厚壳桂叶片δ 15 N值均没有表现出林型间的差异, 表明处于演替中期的混交林土壤N有效性越来越接近阔叶林土壤N有效性水平. ...
Controls of nitrogen isotope patterns in soil profiles.
1
2009
... 土壤δ 15 N在所有林型中也表现出沿剖面增大的趋势, 与土壤TN 呈现相反的变化规律(图2 ), 这与多数学者在森林和草地中的研究结果一致(Mariotti et al ., 1980 ; Tiessen et al ., 1984 ; Shearer & Kohl, 1986 ).引起15 N沿土壤剖面富集的因素很多, 包括菌根真菌对15 N的分馏效应、贫化15 N有机化合物被优先分解利用, 以及硝化和反硝化过程中土壤N的迁移和淋溶(Hobbie & Ouimette, 2009 ).土壤经硝化和反硝化作用, 贫化15 N的气态氮和硝态氮逐渐被释放出系统, 残留的土壤有机质15 N更加富集(Ledgard et al ., 1984 ); 沿剖面深度向下, 土壤粒径越来越小, N含量越来越低, 有机质分解越来越彻底, 土壤腐殖化程度越来越高, 15 N越来越富集(Liao et al ., 2006 ); 另一方面, 植物吸收贫化15 N的NO3 - 和NH4 + 离子后又以凋落物的形式回归到土壤中, 使得表层土壤贫化15 N; 14 N和15 N在垂直剖面不断发生这样的反向迁移, 使得土壤δ 15 N沿剖面深度不断增大. ...
Intramolecular, compound- specific, and bulk carbon isotope patterns in C3 and C4 plants: A review and synthesis.
1
2004
... 2)微生物对13 C贫化基质的优先选择.不同有机化合物(糖类、纤维素、木质素、脂类等)间δ 13 C值最大差异可达10.3‰ (Hobbie & Werner, 2004 ).微生物在呼吸代谢过程中更倾向于利用环境中贫化13 C的碳源, 从而产生同位素分馏效应, 导致残留的土壤有机质13 C富集, 越往底层土壤有机质分解越彻底, 土壤δ 13 C沿剖面深度不断增大(Nadelhoffer & Fry, 1988 ; Högberg & Ekblad, 1996 ). ...
15 N natural abundance in soil-plant systems.
1
1997
... 碳(C)、氮(N)作为两种重要的生命元素, 其生物地球化学循环过程一直是生态学研究的重点, 二者在自然界都有两种稳定性同位素(12 C、13 C和14 N、15 N).由于同位素分馏效应, 不同物质间C、N同位素自然丰度存在差异, 这为同位素技术在生态系统碳-氮循环过程中的应用提供了理论基础(Nadelhoffer & Fry, 1988 ).13 C同位素丰度最早被用于区分植物的光合途径(O’Leary, 1981 ), 其后又被发现与植物光合代谢和水分利用效率之间的关系(Farquhar et al ., 1982 ; Ehleringer et al ., 1986 ), 并被广泛应用于土壤碳循环和植物功能群的研究(Garten & Taylor, 1992 ; 于贵瑞等, 2005 ).在发生C3 -C4 植被转换的区域, 通过土壤稳定碳同位素比率(δ 13 C)在时间上的变化, 可以区分不同来源有机质的相对贡献, 同时也可反演植被覆盖的历史, 同样的方法也用于土壤不同有机组分周转过程的研究(Bernoux et al ., 1998 ; Ehleringer et al ., 2000 ).植物-土壤稳定氮同位素比率(δ 15 N)被认为可以用来评估生态系统的N通量, 且已被部分学者用作生态系统氮饱和状态的指示指标(Högberg, 1997 ; Pardo et al ., 2002 ; Kahmen et al ., 2008 ; Xu et al ., 2010 ).Templer等(2007)研究发现土壤δ 15 N值可以表征土壤的净矿化和硝化速率; Craine等(2009)认为植物-土壤δ 15 N高的地区土壤N有效性也更高, 植物根系对菌根真菌的依赖度更弱, 生态系统氮的饱和度更高.显然, 稳定同位素技术的应用为生态系统C、N元素循环的研究提供了一个新的窗口.但目前δ 13 C自然丰度法多应用于发生过C3 -C4 植被转换区域的碳周转研究(Galdo et al ., 2003 ; Liao et al ., 2006 ; Cheng et al ., 2013 ), 而在植被系统未发生C3 -C4 型转换的亚热带森林, 如何应用δ 13 C自然丰度法探究森林演替过程中土壤碳周转的变化, 仍需更多的探索. ...
Substrate-induced respiration measured in situ in a C3 -plant ecosystem using additions of C4 -sucrose.
2
1996
... 本文结果显示各层土壤δ 13 C均沿演替方向(松林→混交林→阔叶林)逐渐降低, 我们认为可能的解释有两种: 地表凋落物的差异以及凋落物分解形成的有机碳(简称新碳)和土壤中原来存储的有机碳(简称老碳)的混合效应.森林土壤有机质主要来源于植物生物量的输入, Balesdent等(1993)对温带森林的研究发现土壤δ 13 C与凋落物δ 13 C呈正相关关系.本文中地表凋落物δ 13 C表现为松林>混交林>阔叶林, 沿演替方向显著降低(表2 ), 与土壤δ 13 C的变化一致.然而, 土壤并不能完全继承现存凋落物的δ 13 C, 如果不考虑凋落物在分解过程中的分馏效应, 土壤δ 13 C应是新碳和老碳混合作用的结果, 其值由同位素混合效应决定(Liao et al ., 2006 ).随着森林演替的进行, 群落结构和林内小气候均发生改变, 地表凋落物C:N降低(表2 ), 微生物数量和活性增加, 地表微环境更易于凋落物分解, 而凋落物分解速率在一定程度上决定着凋落物中碳的流向(以气体形态释放、以碎屑物残存、以可溶性有机碳形态下渗).Huang等(2011)研究发现, 更快的分解将导致凋落物在分解过程中呼吸消耗的部分减少, 向地下转移的部分(碎屑物和可溶性有机碳)增加.同时, 新碳的快速输入又在一定程度上减缓了土壤中老碳的分解, 因为土壤微生物更倾向于优先分解富含有机质的新鲜碳源(Högberg & Ekblad, 1996 ).对比15年前后鼎湖山森林土壤δ 13 C值的变化, 建立同位素混合模型, 我们发现森林演替后期土壤有机质的平均驻留时间更长(未发表数据).但目前我们还没有得到凋落物分解过程中转入地下部分的确切比例, 利用同位素技术更为精确地量化凋落物分解过程中碳的流向, 估算土壤不同有机组分的分解速率, 将有助于提供更多清晰的线索. ...
... 2)微生物对13 C贫化基质的优先选择.不同有机化合物(糖类、纤维素、木质素、脂类等)间δ 13 C值最大差异可达10.3‰ (Hobbie & Werner, 2004 ).微生物在呼吸代谢过程中更倾向于利用环境中贫化13 C的碳源, 从而产生同位素分馏效应, 导致残留的土壤有机质13 C富集, 越往底层土壤有机质分解越彻底, 土壤δ 13 C沿剖面深度不断增大(Nadelhoffer & Fry, 1988 ; Högberg & Ekblad, 1996 ). ...
Controls of litter quality on the carbon sink in soils through partitioning the products of decomposing litter in a forest succession series in South China.
2011
鼎湖山南亚热带常绿阔叶林结构、物种多样性及种群动态的研究
1
1998
... 鼎湖山阔叶林主要分布在庆云寺周围海拔100-490 m的山坡上, 已有400多年受保护的历史.群落结构复杂, 其中乔木有3层、灌木1层、草本1层, 此外还有较多的藤本植物和附生植物, 主要优势树种有木荷(Schima superba )、锥(Castanopsis chinensis )、黄果厚壳桂(Cryptocarya concinna )、云南银柴(Aporusa yunnanensis )、肖蒲桃(Acmena acuminatissima )、白颜树(Gironniera subaequalis )、橄榄(Canarium album )等(黄忠良等, 1998 ).土壤为水化赤红壤, 土层厚度50-80 cm. ...
Foliar δ15 N values characterize soil N cycling and reflect nitrate or ammonium preference of plants along a temperate grassland gradient.
1
2008
... 碳(C)、氮(N)作为两种重要的生命元素, 其生物地球化学循环过程一直是生态学研究的重点, 二者在自然界都有两种稳定性同位素(12 C、13 C和14 N、15 N).由于同位素分馏效应, 不同物质间C、N同位素自然丰度存在差异, 这为同位素技术在生态系统碳-氮循环过程中的应用提供了理论基础(Nadelhoffer & Fry, 1988 ).13 C同位素丰度最早被用于区分植物的光合途径(O’Leary, 1981 ), 其后又被发现与植物光合代谢和水分利用效率之间的关系(Farquhar et al ., 1982 ; Ehleringer et al ., 1986 ), 并被广泛应用于土壤碳循环和植物功能群的研究(Garten & Taylor, 1992 ; 于贵瑞等, 2005 ).在发生C3 -C4 植被转换的区域, 通过土壤稳定碳同位素比率(δ 13 C)在时间上的变化, 可以区分不同来源有机质的相对贡献, 同时也可反演植被覆盖的历史, 同样的方法也用于土壤不同有机组分周转过程的研究(Bernoux et al ., 1998 ; Ehleringer et al ., 2000 ).植物-土壤稳定氮同位素比率(δ 15 N)被认为可以用来评估生态系统的N通量, 且已被部分学者用作生态系统氮饱和状态的指示指标(Högberg, 1997 ; Pardo et al ., 2002 ; Kahmen et al ., 2008 ; Xu et al ., 2010 ).Templer等(2007)研究发现土壤δ 15 N值可以表征土壤的净矿化和硝化速率; Craine等(2009)认为植物-土壤δ 15 N高的地区土壤N有效性也更高, 植物根系对菌根真菌的依赖度更弱, 生态系统氮的饱和度更高.显然, 稳定同位素技术的应用为生态系统C、N元素循环的研究提供了一个新的窗口.但目前δ 13 C自然丰度法多应用于发生过C3 -C4 植被转换区域的碳周转研究(Galdo et al ., 2003 ; Liao et al ., 2006 ; Cheng et al ., 2013 ), 而在植被系统未发生C3 -C4 型转换的亚热带森林, 如何应用δ 13 C自然丰度法探究森林演替过程中土壤碳周转的变化, 仍需更多的探索. ...
Variations in natural enrichment of 15 N in the profiles of some Australian pasture soils.
1
1984
... 土壤δ 15 N在所有林型中也表现出沿剖面增大的趋势, 与土壤TN 呈现相反的变化规律(图2 ), 这与多数学者在森林和草地中的研究结果一致(Mariotti et al ., 1980 ; Tiessen et al ., 1984 ; Shearer & Kohl, 1986 ).引起15 N沿土壤剖面富集的因素很多, 包括菌根真菌对15 N的分馏效应、贫化15 N有机化合物被优先分解利用, 以及硝化和反硝化过程中土壤N的迁移和淋溶(Hobbie & Ouimette, 2009 ).土壤经硝化和反硝化作用, 贫化15 N的气态氮和硝态氮逐渐被释放出系统, 残留的土壤有机质15 N更加富集(Ledgard et al ., 1984 ); 沿剖面深度向下, 土壤粒径越来越小, N含量越来越低, 有机质分解越来越彻底, 土壤腐殖化程度越来越高, 15 N越来越富集(Liao et al ., 2006 ); 另一方面, 植物吸收贫化15 N的NO3 - 和NH4 + 离子后又以凋落物的形式回归到土壤中, 使得表层土壤贫化15 N; 14 N和15 N在垂直剖面不断发生这样的反向迁移, 使得土壤δ 15 N沿剖面深度不断增大. ...
Organic matter turnover in soil physical fractions following woody plant invasion of grassland: Evidence from natural 13 C and 15 N.
4
2006
... 碳(C)、氮(N)作为两种重要的生命元素, 其生物地球化学循环过程一直是生态学研究的重点, 二者在自然界都有两种稳定性同位素(12 C、13 C和14 N、15 N).由于同位素分馏效应, 不同物质间C、N同位素自然丰度存在差异, 这为同位素技术在生态系统碳-氮循环过程中的应用提供了理论基础(Nadelhoffer & Fry, 1988 ).13 C同位素丰度最早被用于区分植物的光合途径(O’Leary, 1981 ), 其后又被发现与植物光合代谢和水分利用效率之间的关系(Farquhar et al ., 1982 ; Ehleringer et al ., 1986 ), 并被广泛应用于土壤碳循环和植物功能群的研究(Garten & Taylor, 1992 ; 于贵瑞等, 2005 ).在发生C3 -C4 植被转换的区域, 通过土壤稳定碳同位素比率(δ 13 C)在时间上的变化, 可以区分不同来源有机质的相对贡献, 同时也可反演植被覆盖的历史, 同样的方法也用于土壤不同有机组分周转过程的研究(Bernoux et al ., 1998 ; Ehleringer et al ., 2000 ).植物-土壤稳定氮同位素比率(δ 15 N)被认为可以用来评估生态系统的N通量, 且已被部分学者用作生态系统氮饱和状态的指示指标(Högberg, 1997 ; Pardo et al ., 2002 ; Kahmen et al ., 2008 ; Xu et al ., 2010 ).Templer等(2007)研究发现土壤δ 15 N值可以表征土壤的净矿化和硝化速率; Craine等(2009)认为植物-土壤δ 15 N高的地区土壤N有效性也更高, 植物根系对菌根真菌的依赖度更弱, 生态系统氮的饱和度更高.显然, 稳定同位素技术的应用为生态系统C、N元素循环的研究提供了一个新的窗口.但目前δ 13 C自然丰度法多应用于发生过C3 -C4 植被转换区域的碳周转研究(Galdo et al ., 2003 ; Liao et al ., 2006 ; Cheng et al ., 2013 ), 而在植被系统未发生C3 -C4 型转换的亚热带森林, 如何应用δ 13 C自然丰度法探究森林演替过程中土壤碳周转的变化, 仍需更多的探索. ...
... 本文结果显示各层土壤δ 13 C均沿演替方向(松林→混交林→阔叶林)逐渐降低, 我们认为可能的解释有两种: 地表凋落物的差异以及凋落物分解形成的有机碳(简称新碳)和土壤中原来存储的有机碳(简称老碳)的混合效应.森林土壤有机质主要来源于植物生物量的输入, Balesdent等(1993)对温带森林的研究发现土壤δ 13 C与凋落物δ 13 C呈正相关关系.本文中地表凋落物δ 13 C表现为松林>混交林>阔叶林, 沿演替方向显著降低(表2 ), 与土壤δ 13 C的变化一致.然而, 土壤并不能完全继承现存凋落物的δ 13 C, 如果不考虑凋落物在分解过程中的分馏效应, 土壤δ 13 C应是新碳和老碳混合作用的结果, 其值由同位素混合效应决定(Liao et al ., 2006 ).随着森林演替的进行, 群落结构和林内小气候均发生改变, 地表凋落物C:N降低(表2 ), 微生物数量和活性增加, 地表微环境更易于凋落物分解, 而凋落物分解速率在一定程度上决定着凋落物中碳的流向(以气体形态释放、以碎屑物残存、以可溶性有机碳形态下渗).Huang等(2011)研究发现, 更快的分解将导致凋落物在分解过程中呼吸消耗的部分减少, 向地下转移的部分(碎屑物和可溶性有机碳)增加.同时, 新碳的快速输入又在一定程度上减缓了土壤中老碳的分解, 因为土壤微生物更倾向于优先分解富含有机质的新鲜碳源(Högberg & Ekblad, 1996 ).对比15年前后鼎湖山森林土壤δ 13 C值的变化, 建立同位素混合模型, 我们发现森林演替后期土壤有机质的平均驻留时间更长(未发表数据).但目前我们还没有得到凋落物分解过程中转入地下部分的确切比例, 利用同位素技术更为精确地量化凋落物分解过程中碳的流向, 估算土壤不同有机组分的分解速率, 将有助于提供更多清晰的线索. ...
... 3个林型土壤δ 13 C均沿剖面深度不断增大, 与TOC 的变化规律相反(图2 ), 这与多数研究结果相吻合(Ehleringer et al ., 2000 ; Liao et al ., 2006 ).对于这种富集机制的解释主要有以下几种假说: ...
... 土壤δ 15 N在所有林型中也表现出沿剖面增大的趋势, 与土壤TN 呈现相反的变化规律(图2 ), 这与多数学者在森林和草地中的研究结果一致(Mariotti et al ., 1980 ; Tiessen et al ., 1984 ; Shearer & Kohl, 1986 ).引起15 N沿土壤剖面富集的因素很多, 包括菌根真菌对15 N的分馏效应、贫化15 N有机化合物被优先分解利用, 以及硝化和反硝化过程中土壤N的迁移和淋溶(Hobbie & Ouimette, 2009 ).土壤经硝化和反硝化作用, 贫化15 N的气态氮和硝态氮逐渐被释放出系统, 残留的土壤有机质15 N更加富集(Ledgard et al ., 1984 ); 沿剖面深度向下, 土壤粒径越来越小, N含量越来越低, 有机质分解越来越彻底, 土壤腐殖化程度越来越高, 15 N越来越富集(Liao et al ., 2006 ); 另一方面, 植物吸收贫化15 N的NO3 - 和NH4 + 离子后又以凋落物的形式回归到土壤中, 使得表层土壤贫化15 N; 14 N和15 N在垂直剖面不断发生这样的反向迁移, 使得土壤δ 15 N沿剖面深度不断增大. ...
南亚热带森林演替过程中小气候的改变及对气候变化的响应
1
2014
... 多数研究结果证实, C3 植物的碳同位素分馏效应与叶片胞间CO2 浓度和大气CO2 浓度的比值(C i /C a )具有明显的线性正相关关系(Farquhar et al ., 1982 ), 而C i /C a 受光合速率和气孔导度的影响, 因此, 叶片δ 13 C与植物光合和蒸腾等生理过程有着密切的联系, 可以作为估测植物长期水分利用效率(WUE )的重要指标(Ehleringer et al ., 1986 ).本文结果显示混交林中马尾松叶片δ 13 C显著低于松林, 这与演替过程中林内小气候的变化相关.随着演替的进行, 森林郁闭度增加, 林内光照水平减弱, 气温降低, 林内空气湿度和土壤含水量升高(刘效东等, 2014 ), 由此引起植物光合和蒸腾速率的改变, 植物WUE 下降, 叶片C i /C a 升高, δ 13 C值降低.这与Ehleringer等(1986)在鼎湖山的研究结果相吻合, 孙谷畴等(1993)对亚热带人工松林的研究也得到类似的结论.马尾松作为喜光性的耐旱先锋种, 在荫蔽潮湿的环境下优势度减弱, 逐渐被阔叶树种所替代, 群落结构逐步演化成针阔叶混交林.而混交林和阔叶林的3个共有树种叶片δ 13 C均没有表现出林型间的显著差异, 这或许可以间接证明处于演替中、后期的两林型间林分差异越来越小, 混交林林内气温、空气湿度、土壤含水量及植物生理功能逐渐趋同于阔叶林(监测数据, 未发表), 混交林正快速向演替顶级过渡(Zhou et al ., 2006b ). ...
The abundance of natural nitrogen 15 in the organic matter of soils along an altitudinal gradient.
1
1980
... 土壤δ 15 N在所有林型中也表现出沿剖面增大的趋势, 与土壤TN 呈现相反的变化规律(图2 ), 这与多数学者在森林和草地中的研究结果一致(Mariotti et al ., 1980 ; Tiessen et al ., 1984 ; Shearer & Kohl, 1986 ).引起15 N沿土壤剖面富集的因素很多, 包括菌根真菌对15 N的分馏效应、贫化15 N有机化合物被优先分解利用, 以及硝化和反硝化过程中土壤N的迁移和淋溶(Hobbie & Ouimette, 2009 ).土壤经硝化和反硝化作用, 贫化15 N的气态氮和硝态氮逐渐被释放出系统, 残留的土壤有机质15 N更加富集(Ledgard et al ., 1984 ); 沿剖面深度向下, 土壤粒径越来越小, N含量越来越低, 有机质分解越来越彻底, 土壤腐殖化程度越来越高, 15 N越来越富集(Liao et al ., 2006 ); 另一方面, 植物吸收贫化15 N的NO3 - 和NH4 + 离子后又以凋落物的形式回归到土壤中, 使得表层土壤贫化15 N; 14 N和15 N在垂直剖面不断发生这样的反向迁移, 使得土壤δ 15 N沿剖面深度不断增大. ...
Controls on natural nitrogen-15 and carbon-13 abundances in forest soil organic matter.
3
1988
... 碳(C)、氮(N)作为两种重要的生命元素, 其生物地球化学循环过程一直是生态学研究的重点, 二者在自然界都有两种稳定性同位素(12 C、13 C和14 N、15 N).由于同位素分馏效应, 不同物质间C、N同位素自然丰度存在差异, 这为同位素技术在生态系统碳-氮循环过程中的应用提供了理论基础(Nadelhoffer & Fry, 1988 ).13 C同位素丰度最早被用于区分植物的光合途径(O’Leary, 1981 ), 其后又被发现与植物光合代谢和水分利用效率之间的关系(Farquhar et al ., 1982 ; Ehleringer et al ., 1986 ), 并被广泛应用于土壤碳循环和植物功能群的研究(Garten & Taylor, 1992 ; 于贵瑞等, 2005 ).在发生C3 -C4 植被转换的区域, 通过土壤稳定碳同位素比率(δ 13 C)在时间上的变化, 可以区分不同来源有机质的相对贡献, 同时也可反演植被覆盖的历史, 同样的方法也用于土壤不同有机组分周转过程的研究(Bernoux et al ., 1998 ; Ehleringer et al ., 2000 ).植物-土壤稳定氮同位素比率(δ 15 N)被认为可以用来评估生态系统的N通量, 且已被部分学者用作生态系统氮饱和状态的指示指标(Högberg, 1997 ; Pardo et al ., 2002 ; Kahmen et al ., 2008 ; Xu et al ., 2010 ).Templer等(2007)研究发现土壤δ 15 N值可以表征土壤的净矿化和硝化速率; Craine等(2009)认为植物-土壤δ 15 N高的地区土壤N有效性也更高, 植物根系对菌根真菌的依赖度更弱, 生态系统氮的饱和度更高.显然, 稳定同位素技术的应用为生态系统C、N元素循环的研究提供了一个新的窗口.但目前δ 13 C自然丰度法多应用于发生过C3 -C4 植被转换区域的碳周转研究(Galdo et al ., 2003 ; Liao et al ., 2006 ; Cheng et al ., 2013 ), 而在植被系统未发生C3 -C4 型转换的亚热带森林, 如何应用δ 13 C自然丰度法探究森林演替过程中土壤碳周转的变化, 仍需更多的探索. ...
... 表层(0-10 cm)土壤δ 15 N沿演替方向呈现增大的规律, 与凋落物δ 15 N的变化一致, 表明土壤δ 15 N与凋落物的输入密切相关.Falkengren-Grerup等(2004)认为δ 15 N可以作为植物吸收NO3 - 的相对指标.Pardo等(2002)发现, 生态系统越接近“氮饱和”, 土壤和叶片的δ 15 N值越高.Craine等(2009)认为植物-土壤δ 15 N高的区域土壤N有效性也更高.也有研究显示表层土壤δ 15 N自然丰度可以作为指示氮循环速率的指标(Xu et al ., 2010 ).Templer等(2007)研究表明细根和土壤的δ 15 N与净矿化和硝化速率显著正相关.我们或许可以据此推测, 随着森林演替的进行, 土壤N有效性将增加, 生态系统“氮饱和”程度越来越高.深层土壤δ 15 N没有表现出不同林型间的显著差异, 这与土壤垂直剖面15 N的分馏效应和N元素的迁移有关(Nadelhoffer & Fry, 1988 ). ...
... 2)微生物对13 C贫化基质的优先选择.不同有机化合物(糖类、纤维素、木质素、脂类等)间δ 13 C值最大差异可达10.3‰ (Hobbie & Werner, 2004 ).微生物在呼吸代谢过程中更倾向于利用环境中贫化13 C的碳源, 从而产生同位素分馏效应, 导致残留的土壤有机质13 C富集, 越往底层土壤有机质分解越彻底, 土壤δ 13 C沿剖面深度不断增大(Nadelhoffer & Fry, 1988 ; Högberg & Ekblad, 1996 ). ...
Carbon isotope fractionation in plants.
1
1981
... 碳(C)、氮(N)作为两种重要的生命元素, 其生物地球化学循环过程一直是生态学研究的重点, 二者在自然界都有两种稳定性同位素(12 C、13 C和14 N、15 N).由于同位素分馏效应, 不同物质间C、N同位素自然丰度存在差异, 这为同位素技术在生态系统碳-氮循环过程中的应用提供了理论基础(Nadelhoffer & Fry, 1988 ).13 C同位素丰度最早被用于区分植物的光合途径(O’Leary, 1981 ), 其后又被发现与植物光合代谢和水分利用效率之间的关系(Farquhar et al ., 1982 ; Ehleringer et al ., 1986 ), 并被广泛应用于土壤碳循环和植物功能群的研究(Garten & Taylor, 1992 ; 于贵瑞等, 2005 ).在发生C3 -C4 植被转换的区域, 通过土壤稳定碳同位素比率(δ 13 C)在时间上的变化, 可以区分不同来源有机质的相对贡献, 同时也可反演植被覆盖的历史, 同样的方法也用于土壤不同有机组分周转过程的研究(Bernoux et al ., 1998 ; Ehleringer et al ., 2000 ).植物-土壤稳定氮同位素比率(δ 15 N)被认为可以用来评估生态系统的N通量, 且已被部分学者用作生态系统氮饱和状态的指示指标(Högberg, 1997 ; Pardo et al ., 2002 ; Kahmen et al ., 2008 ; Xu et al ., 2010 ).Templer等(2007)研究发现土壤δ 15 N值可以表征土壤的净矿化和硝化速率; Craine等(2009)认为植物-土壤δ 15 N高的地区土壤N有效性也更高, 植物根系对菌根真菌的依赖度更弱, 生态系统氮的饱和度更高.显然, 稳定同位素技术的应用为生态系统C、N元素循环的研究提供了一个新的窗口.但目前δ 13 C自然丰度法多应用于发生过C3 -C4 植被转换区域的碳周转研究(Galdo et al ., 2003 ; Liao et al ., 2006 ; Cheng et al ., 2013 ), 而在植被系统未发生C3 -C4 型转换的亚热带森林, 如何应用δ 13 C自然丰度法探究森林演替过程中土壤碳周转的变化, 仍需更多的探索. ...
Carbon isotopes in photosynthesis.
1
1988
... 显著负相关, 与C:N极显著正相关, 与Tsialtas等(2001)的结果一致, 而与其他的一些结论相反(Cernusak et al ., 2013 ; Zhang et al ., 2015 ), 这可能与不同环境条件下土壤养分供应水平及物种在群落中的资源利用策略有关.理论上, 在“氮限制”的环境下, 叶片N含量的增加有利于提高核酮糖1,5-二磷酸羧化氧化酶(Rubisco)的数量和活性, 光合速率加快, C i /C a 降低, 叶片δ 13 C增大(O’Leary, 1988 ).而南亚热带地区氮沉降水平高, 生态系统趋于“氮饱和”状态, 植物的生长转而受到其他营养元素(如磷)的限制(Chen et al ., 2015 ); 鼎湖山降水充沛, 土壤含水量高, 更多的N吸收意味着更多水分的摄入, 从而导致叶片气孔导度增大, C i /C a 升高, δ 13 C降低.以上结果表明: 出于对湿润气候和高氮沉降水平的一种适应策略, 南亚热带森林群落中植物可能同时具有较低的水分利用效率和氮素利用效率(C:N), 并不一定存在相互制约的补偿机制. ...
鼎湖山针阔叶混交林小气候调节效应
1
2014
... 混交林多分布于自然林的林缘, 海拔80-400 m之间, 目前正快速向阔叶林恢复演替.地上垂直结构大致可分为4层, 其中乔木2层、灌木1层、草本1层, 此外还有少量藤本和附生等层间植物, 主要优势树种有马尾松、木荷、锥、黄果厚壳桂、滇粤山胡椒(Lindera metcalfiana )等(欧阳旭等, 2014 ).土壤为赤红壤, 土层厚度40-70 cm. ...
Response of the natural abundance of 15 N in forest soils and foliage to high nitrate loss following clear cutting.
1
2002
... 碳(C)、氮(N)作为两种重要的生命元素, 其生物地球化学循环过程一直是生态学研究的重点, 二者在自然界都有两种稳定性同位素(12 C、13 C和14 N、15 N).由于同位素分馏效应, 不同物质间C、N同位素自然丰度存在差异, 这为同位素技术在生态系统碳-氮循环过程中的应用提供了理论基础(Nadelhoffer & Fry, 1988 ).13 C同位素丰度最早被用于区分植物的光合途径(O’Leary, 1981 ), 其后又被发现与植物光合代谢和水分利用效率之间的关系(Farquhar et al ., 1982 ; Ehleringer et al ., 1986 ), 并被广泛应用于土壤碳循环和植物功能群的研究(Garten & Taylor, 1992 ; 于贵瑞等, 2005 ).在发生C3 -C4 植被转换的区域, 通过土壤稳定碳同位素比率(δ 13 C)在时间上的变化, 可以区分不同来源有机质的相对贡献, 同时也可反演植被覆盖的历史, 同样的方法也用于土壤不同有机组分周转过程的研究(Bernoux et al ., 1998 ; Ehleringer et al ., 2000 ).植物-土壤稳定氮同位素比率(δ 15 N)被认为可以用来评估生态系统的N通量, 且已被部分学者用作生态系统氮饱和状态的指示指标(Högberg, 1997 ; Pardo et al ., 2002 ; Kahmen et al ., 2008 ; Xu et al ., 2010 ).Templer等(2007)研究发现土壤δ 15 N值可以表征土壤的净矿化和硝化速率; Craine等(2009)认为植物-土壤δ 15 N高的地区土壤N有效性也更高, 植物根系对菌根真菌的依赖度更弱, 生态系统氮的饱和度更高.显然, 稳定同位素技术的应用为生态系统C、N元素循环的研究提供了一个新的窗口.但目前δ 13 C自然丰度法多应用于发生过C3 -C4 植被转换区域的碳周转研究(Galdo et al ., 2003 ; Liao et al ., 2006 ; Cheng et al ., 2013 ), 而在植被系统未发生C3 -C4 型转换的亚热带森林, 如何应用δ 13 C自然丰度法探究森林演替过程中土壤碳周转的变化, 仍需更多的探索. ...
N2 -fixation in field settings: Estimations based on natural 15 N abundance.
1
1986
... 土壤δ 15 N在所有林型中也表现出沿剖面增大的趋势, 与土壤TN 呈现相反的变化规律(图2 ), 这与多数学者在森林和草地中的研究结果一致(Mariotti et al ., 1980 ; Tiessen et al ., 1984 ; Shearer & Kohl, 1986 ).引起15 N沿土壤剖面富集的因素很多, 包括菌根真菌对15 N的分馏效应、贫化15 N有机化合物被优先分解利用, 以及硝化和反硝化过程中土壤N的迁移和淋溶(Hobbie & Ouimette, 2009 ).土壤经硝化和反硝化作用, 贫化15 N的气态氮和硝态氮逐渐被释放出系统, 残留的土壤有机质15 N更加富集(Ledgard et al ., 1984 ); 沿剖面深度向下, 土壤粒径越来越小, N含量越来越低, 有机质分解越来越彻底, 土壤腐殖化程度越来越高, 15 N越来越富集(Liao et al ., 2006 ); 另一方面, 植物吸收贫化15 N的NO3 - 和NH4 + 离子后又以凋落物的形式回归到土壤中, 使得表层土壤贫化15 N; 14 N和15 N在垂直剖面不断发生这样的反向迁移, 使得土壤δ 15 N沿剖面深度不断增大. ...
亚热带人工林松树13 C/12 C比率和水分利用效率
1993
How stable isotopes may help to elucidate primary nitrogen metabolism and its interaction with (photo) respiration in C3 leaves.
1
2008
... 除少数固氮植物外, 植物氮素主要来源于根系吸收土壤中的硝态氮和铵态氮, 再经过同化还原作用固定在有机质中, 这两个阶段都会产生氮同位素分馏, 其分馏效应主要取决于土壤N有效性以及植物对氮素的需求(Tcherkez & Hodges, 2008 ).许多研究已经在不同空间尺度上证明叶片δ 15 N与其N含量正相关(Hobbie et al ., 2000 ; Craine et al ., 2009 ), 本文的结果与之相吻合(图1E ), Craine等(2009)认为叶片N含量高的地区土壤N有效性高, 系统N循环开放度更高, 更多贫化15 N的气态N (如NH3 、N2 O和N2 等)被释放, 系统富集15 N; 同时植物对菌根真菌的依赖度下降, N吸收过程中同位素分馏效应减弱, 导致植物δ 15 N增大.表1 结果显示马尾松叶片δ 15 N值在混交林中较松林中显著增大, 可以解释为混交林土壤N有效性明显高于松林, 土壤中氮的净矿化和硝化速率更快(Templer et al ., 2007 ).而混交林和阔叶林的3个共有优势种, 除木荷外, 锥和黄果厚壳桂叶片δ 15 N值均没有表现出林型间的差异, 表明处于演替中期的混交林土壤N有效性越来越接近阔叶林土壤N有效性水平. ...
Plant and soil natural abundance δ15 N: Indicators of relative rates of nitrogen cycling in temperate forest ecosystems.
1
2007
... 除少数固氮植物外, 植物氮素主要来源于根系吸收土壤中的硝态氮和铵态氮, 再经过同化还原作用固定在有机质中, 这两个阶段都会产生氮同位素分馏, 其分馏效应主要取决于土壤N有效性以及植物对氮素的需求(Tcherkez & Hodges, 2008 ).许多研究已经在不同空间尺度上证明叶片δ 15 N与其N含量正相关(Hobbie et al ., 2000 ; Craine et al ., 2009 ), 本文的结果与之相吻合(图1E ), Craine等(2009)认为叶片N含量高的地区土壤N有效性高, 系统N循环开放度更高, 更多贫化15 N的气态N (如NH3 、N2 O和N2 等)被释放, 系统富集15 N; 同时植物对菌根真菌的依赖度下降, N吸收过程中同位素分馏效应减弱, 导致植物δ 15 N增大.表1 结果显示马尾松叶片δ 15 N值在混交林中较松林中显著增大, 可以解释为混交林土壤N有效性明显高于松林, 土壤中氮的净矿化和硝化速率更快(Templer et al ., 2007 ).而混交林和阔叶林的3个共有优势种, 除木荷外, 锥和黄果厚壳桂叶片δ 15 N值均没有表现出林型间的差异, 表明处于演替中期的混交林土壤N有效性越来越接近阔叶林土壤N有效性水平. ...
Natural nitrogen-15 abundance as an indicator of soil organic matter transformations in native and cultivated soils.
1
1984
... 土壤δ 15 N在所有林型中也表现出沿剖面增大的趋势, 与土壤TN 呈现相反的变化规律(图2 ), 这与多数学者在森林和草地中的研究结果一致(Mariotti et al ., 1980 ; Tiessen et al ., 1984 ; Shearer & Kohl, 1986 ).引起15 N沿土壤剖面富集的因素很多, 包括菌根真菌对15 N的分馏效应、贫化15 N有机化合物被优先分解利用, 以及硝化和反硝化过程中土壤N的迁移和淋溶(Hobbie & Ouimette, 2009 ).土壤经硝化和反硝化作用, 贫化15 N的气态氮和硝态氮逐渐被释放出系统, 残留的土壤有机质15 N更加富集(Ledgard et al ., 1984 ); 沿剖面深度向下, 土壤粒径越来越小, N含量越来越低, 有机质分解越来越彻底, 土壤腐殖化程度越来越高, 15 N越来越富集(Liao et al ., 2006 ); 另一方面, 植物吸收贫化15 N的NO3 - 和NH4 + 离子后又以凋落物的形式回归到土壤中, 使得表层土壤贫化15 N; 14 N和15 N在垂直剖面不断发生这样的反向迁移, 使得土壤δ 15 N沿剖面深度不断增大. ...
Interspecific variation in potential water use efficiency and its relation to plant species abundance in a water-limited grassland.
2001
An extraction method for measuring soil microbial biomass C.
1
1987
... 式中, R = 13 C/12 C或15 N/14 N, R sam 表示样品的相对丰度, R std 表示国际标准物的相对丰度.碳同位素国际标准物为Pee Dee Belemnite (PDB), 即美国南卡罗来纳州白垩系皮狄组地层中的美洲拟箭石(R PDB = 0.0112372, δ 13 CPDB = 0‰); 氮同位素国际标准物为标准大气中的N2 (R air = 0.0036765, δ 15 Nair = 0‰).本次实验所得同位素比值δ 的测定精度为± 0.1‰, 由中国科学院华南植物园公共实验室提供.土壤微生物生物量碳的测定采用氯仿熏蒸-K2 SO4 浸提法(Vance et al ., 1987 ).土壤易氧化有机碳的测定采用高锰酸钾氧化法(Blair et al ., 1995 ). ...
Natural 15 N abundance in soils and plants in relation to N cycling in a rangeland in Inner Mongolia.
2
2010
... 碳(C)、氮(N)作为两种重要的生命元素, 其生物地球化学循环过程一直是生态学研究的重点, 二者在自然界都有两种稳定性同位素(12 C、13 C和14 N、15 N).由于同位素分馏效应, 不同物质间C、N同位素自然丰度存在差异, 这为同位素技术在生态系统碳-氮循环过程中的应用提供了理论基础(Nadelhoffer & Fry, 1988 ).13 C同位素丰度最早被用于区分植物的光合途径(O’Leary, 1981 ), 其后又被发现与植物光合代谢和水分利用效率之间的关系(Farquhar et al ., 1982 ; Ehleringer et al ., 1986 ), 并被广泛应用于土壤碳循环和植物功能群的研究(Garten & Taylor, 1992 ; 于贵瑞等, 2005 ).在发生C3 -C4 植被转换的区域, 通过土壤稳定碳同位素比率(δ 13 C)在时间上的变化, 可以区分不同来源有机质的相对贡献, 同时也可反演植被覆盖的历史, 同样的方法也用于土壤不同有机组分周转过程的研究(Bernoux et al ., 1998 ; Ehleringer et al ., 2000 ).植物-土壤稳定氮同位素比率(δ 15 N)被认为可以用来评估生态系统的N通量, 且已被部分学者用作生态系统氮饱和状态的指示指标(Högberg, 1997 ; Pardo et al ., 2002 ; Kahmen et al ., 2008 ; Xu et al ., 2010 ).Templer等(2007)研究发现土壤δ 15 N值可以表征土壤的净矿化和硝化速率; Craine等(2009)认为植物-土壤δ 15 N高的地区土壤N有效性也更高, 植物根系对菌根真菌的依赖度更弱, 生态系统氮的饱和度更高.显然, 稳定同位素技术的应用为生态系统C、N元素循环的研究提供了一个新的窗口.但目前δ 13 C自然丰度法多应用于发生过C3 -C4 植被转换区域的碳周转研究(Galdo et al ., 2003 ; Liao et al ., 2006 ; Cheng et al ., 2013 ), 而在植被系统未发生C3 -C4 型转换的亚热带森林, 如何应用δ 13 C自然丰度法探究森林演替过程中土壤碳周转的变化, 仍需更多的探索. ...
... 表层(0-10 cm)土壤δ 15 N沿演替方向呈现增大的规律, 与凋落物δ 15 N的变化一致, 表明土壤δ 15 N与凋落物的输入密切相关.Falkengren-Grerup等(2004)认为δ 15 N可以作为植物吸收NO3 - 的相对指标.Pardo等(2002)发现, 生态系统越接近“氮饱和”, 土壤和叶片的δ 15 N值越高.Craine等(2009)认为植物-土壤δ 15 N高的区域土壤N有效性也更高.也有研究显示表层土壤δ 15 N自然丰度可以作为指示氮循环速率的指标(Xu et al ., 2010 ).Templer等(2007)研究表明细根和土壤的δ 15 N与净矿化和硝化速率显著正相关.我们或许可以据此推测, 随着森林演替的进行, 土壤N有效性将增加, 生态系统“氮饱和”程度越来越高.深层土壤δ 15 N没有表现出不同林型间的显著差异, 这与土壤垂直剖面15 N的分馏效应和N元素的迁移有关(Nadelhoffer & Fry, 1988 ). ...
碳同位素技术在土壤碳循环研究中的应用
1
2005
... 碳(C)、氮(N)作为两种重要的生命元素, 其生物地球化学循环过程一直是生态学研究的重点, 二者在自然界都有两种稳定性同位素(12 C、13 C和14 N、15 N).由于同位素分馏效应, 不同物质间C、N同位素自然丰度存在差异, 这为同位素技术在生态系统碳-氮循环过程中的应用提供了理论基础(Nadelhoffer & Fry, 1988 ).13 C同位素丰度最早被用于区分植物的光合途径(O’Leary, 1981 ), 其后又被发现与植物光合代谢和水分利用效率之间的关系(Farquhar et al ., 1982 ; Ehleringer et al ., 1986 ), 并被广泛应用于土壤碳循环和植物功能群的研究(Garten & Taylor, 1992 ; 于贵瑞等, 2005 ).在发生C3 -C4 植被转换的区域, 通过土壤稳定碳同位素比率(δ 13 C)在时间上的变化, 可以区分不同来源有机质的相对贡献, 同时也可反演植被覆盖的历史, 同样的方法也用于土壤不同有机组分周转过程的研究(Bernoux et al ., 1998 ; Ehleringer et al ., 2000 ).植物-土壤稳定氮同位素比率(δ 15 N)被认为可以用来评估生态系统的N通量, 且已被部分学者用作生态系统氮饱和状态的指示指标(Högberg, 1997 ; Pardo et al ., 2002 ; Kahmen et al ., 2008 ; Xu et al ., 2010 ).Templer等(2007)研究发现土壤δ 15 N值可以表征土壤的净矿化和硝化速率; Craine等(2009)认为植物-土壤δ 15 N高的地区土壤N有效性也更高, 植物根系对菌根真菌的依赖度更弱, 生态系统氮的饱和度更高.显然, 稳定同位素技术的应用为生态系统C、N元素循环的研究提供了一个新的窗口.但目前δ 13 C自然丰度法多应用于发生过C3 -C4 植被转换区域的碳周转研究(Galdo et al ., 2003 ; Liao et al ., 2006 ; Cheng et al ., 2013 ), 而在植被系统未发生C3 -C4 型转换的亚热带森林, 如何应用δ 13 C自然丰度法探究森林演替过程中土壤碳周转的变化, 仍需更多的探索. ...
Nitrogen control of 13 C enrichment in heterotrophic organs relative to leaves in a landscape- building desert plant species.
1
2015
... 显著负相关, 与C:N极显著正相关, 与Tsialtas等(2001)的结果一致, 而与其他的一些结论相反(Cernusak et al ., 2013 ; Zhang et al ., 2015 ), 这可能与不同环境条件下土壤养分供应水平及物种在群落中的资源利用策略有关.理论上, 在“氮限制”的环境下, 叶片N含量的增加有利于提高核酮糖1,5-二磷酸羧化氧化酶(Rubisco)的数量和活性, 光合速率加快, C i /C a 降低, 叶片δ 13 C增大(O’Leary, 1988 ).而南亚热带地区氮沉降水平高, 生态系统趋于“氮饱和”状态, 植物的生长转而受到其他营养元素(如磷)的限制(Chen et al ., 2015 ); 鼎湖山降水充沛, 土壤含水量高, 更多的N吸收意味着更多水分的摄入, 从而导致叶片气孔导度增大, C i /C a 升高, δ 13 C降低.以上结果表明: 出于对湿润气候和高氮沉降水平的一种适应策略, 南亚热带森林群落中植物可能同时具有较低的水分利用效率和氮素利用效率(C:N), 并不一定存在相互制约的补偿机制. ...
a). Old-growth forests can accumulate carbon in soils.
2006
b). Belowground carbon balance and carbon accumulation rate in the successional serials of monsoon evergreen broad-leaved forest.
2006



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}