水淹胁迫是影响植物生长、生存的主要胁迫。每年在世界各地都会发生不同程度和不同时间的水淹胁迫(Candan & Tarhan, 2012 ), 尤其是水库建设中形成的消落带, 其水淹胁迫发生频率更高, 程度更加严重。消落带每年周期性地遭受不同程度的水淹胁迫, 大部分陆生植物因无法忍受长时间、高强度的反季节水淹逐渐死亡(樊大勇等, 2015 ), 造成消落带植物群落向结构简单化、生物多样性急剧减少的方向发展(Liu & Willison, 2013 )。水淹使土壤中气体交换量减少, 导致根系缺氧, 功能紊乱。同时土壤微生物与根系争夺O2 , 使根系通过线粒体氧化磷酸化途径供能更加困难(Sauter, 2013 ), 进而影响根系的一切耗能活动(水分输送、养分吸收、植物体的糖类代谢以及光合作用和活性氧代谢等)。植物在缺氧条件下会通过增加乙醇脱氢酶(ADH)、乳酸脱氢酶(LDH)以及丙酮酸脱羧酶(PDC)活性启动无氧呼吸, 以应对缺氧环境(Zhou et al ., 2012 )。
糖类是植物的能量来源, 植物体内的糖类主要包括结构性碳水化合物(主要用于植物形态建成)和非结构性碳水化合物(参与生命代谢), 非结构性碳水化合物(可溶性糖、淀粉等)的代谢在很大程度上决定了植物的生长发育及对环境的适应能力(施美芬等, 2010 )。可溶性糖参与植物体内的糖酵解过程, 为植物提供能量, 淀粉则是主要的能量储存形式。水淹对糖含量的影响较为复杂, 主要取决于植物对水淹的适应策略。植物适应水淹的策略主要包括逃避型和忍耐型。逃避型植物具发达的通气组织, 通过消耗糖类促进茎生长以接近水面, 从而增强植物体内部的气体交换, 获得更多O2 ; 而忍耐型植物通过抑制生长来储存能量(Akman et al ., 2012 )。目前对于植物适应水淹的策略研究大多集中在草本, 对木本植物的研究较少, 且木本植物适应水淹的策略现在还不明确。
中山杉(Taxodium ‘Zhongshansha’)是由江苏省中国科学院植物研究所通过落羽杉(Taxodium distichum )、墨西哥落羽杉(Taxodium mucronatum )以及池杉(Taxodium ascendens )种间杂交培育出的优良品种(华建峰等, 2011b ), 主要生长在包括三大湖(滇池、巢湖、太湖)以及三峡库区消落带(重庆)在内的长江中下游地区(刘艳, 2013 )。中山杉耐淹、耐碱、树形优美(King & Arnold, 2012 ), 其耐淹性尤为突出, 可以忍受较长时间的水淹胁迫(Oren et al ., 2001 )。对‘中山杉406’进行为期3个月的水淹试验后出现皮孔肥大、长出气生根等适应水淹的特征, 水淹并没有影响其光合特性(华建峰等, 2011a , 2011b )。但是中山杉水下呼吸机制以及糖类转化的研究鲜有报道。殷云龙等(2014)在重庆万州消落带内栽植两年生‘中山杉118’ ((落羽杉×墨西哥落羽杉)×墨西哥落羽杉)进行造林实验, 发现中山杉在非生长季经受122天、最大没顶深度12 m的水淹胁迫后, 仍有90%的保存率。植物的耐淹能力以及适应策略与水淹季节密切相关, 生长季植物新陈代谢速率较快, 需氧量更大(Crawford, 2003 ), 植物对生长季水淹的敏感度更大(Jackson & Ram, 2003 )。栽植在三峡库区万州段消落带内的中山杉表现出的极强耐淹性, 是其本身的遗传特性还是与消落带非生长季水淹的特殊环境有关, 中山杉又是通过何种策略适应不同季节的长期水淹等仍是需要探究的科学问题, 为此本文通过测定中山杉幼苗经受生长季不同程度水淹胁迫后叶片和根系的呼吸及糖类代谢, 从能量消耗的角度探索中山杉的耐淹机理, 以及中山杉幼苗对生长季水淹胁迫的适应策略, 为湿地生态修复和消落带生物治理中植物材料的选择提供依据。
1 材料和方法
1.1 试验材料与试验设计
供试材料为一年生‘中山杉118’幼苗, 平均株高44 cm, 平均地径0.6 cm, 栽植容器为高18 cm、直径15 cm的圆柱形花盆, 基质为营养土(土:肥料体积比为3:1), 试验环境为室温25-30 ℃, 利用植物生长灯(红光:蓝光:白光= 2:2:1)补光, 光周期为13/11 h (light/dark), 光强为200 μmol·m-2 ·s-1 。水淹容器为400 L的长方体白色不透光塑料水箱(980 mm × 760 mm × 660 mm)。水为自来水, 水温在20 ℃左右。
于2014年8月8日, 筛选生长健康、大小相近的24盆中山杉幼苗进行不同程度的水淹处理: 对照(不做处理, 正常生长, 每周浇一次水)、水浸(土壤水淹, 植株地上部分与空气接触)、浅淹(植物大部分浸于水中, 但水不没顶)、深淹(植株整体位于水中, 植株顶部距水面10 cm左右), 每组6个重复, 每5天换一次水。于2014年11月8日, 对中山杉幼苗根系及叶片相关指标进行测定。
1.2 酶活性测定
1 g样本+ 9 mL PBS (0.01 mol·L-1 , pH 7.2-7.4)缓冲液, 冰水浴研磨, 离心管采集样本研磨液, 5000 r·min-1 , 离心3 min, 取上清液测定。ADH、PDC、LDH酶活性均采用美国RB公司生产的ELISA测试盒测定, 每组6个重复。
1.3 淀粉和可溶性糖含量测定
称取1 g粉碎过100目筛的烘干样品, 置于15 mL刻度试管中, 加入7 mL 80%的乙醇, 在80 ℃水浴中提取30 min, 取出离心(3000 r·min-1 ) 5 min, 收集上清液。重复提取3次, 收集3次上清液合并于烧杯, 置于85 ℃恒温水浴, 使乙醇蒸发至2-3 mL, 转移至50 mL容量瓶中, 以蒸馏水定容, 供可溶性糖含量测定。
向离心后剩余的沉淀物中加3 mL蒸馏水, 搅拌均匀, 沸水浴糊化15 min, 冷却后加2 mL冷的9.2 mol·L-1 的高氯酸, 不时搅拌, 提取15 min后加蒸馏水至10 mL, 混匀, 离心10 min, 上清液倾入50 mL容量瓶。再向沉淀中加入2 mL 4.6 mol·L-1 高氯酸, 搅拌提取15 min后加水至10 mL, 混匀后离心10 min, 收集上清液于容量瓶。然后用水洗沉淀1-2次, 离心, 合并离心液于50 mL容量瓶用蒸馏水定容供淀粉含量测定。淀粉和可溶性糖含量采用蒽酮法测定(Yemm & Willis, 1954 )。
1.4 根系活力测定
根系活力采用TTC法测定。
1.5 生物量测定
叶片与根系鲜质量采用电子天平称量, 根长、根表面积以及根系分形维数采用万深(LA-S)根系分析仪(杭州万深检测科技有限公司, 杭州)测量。
1.6 数据分析
用Excel进行数据整理, 用SPSS进行数据分析, 用Origin 8.0作图。
2 结果和分析
2.1 水淹胁迫对中山杉幼苗生长的影响
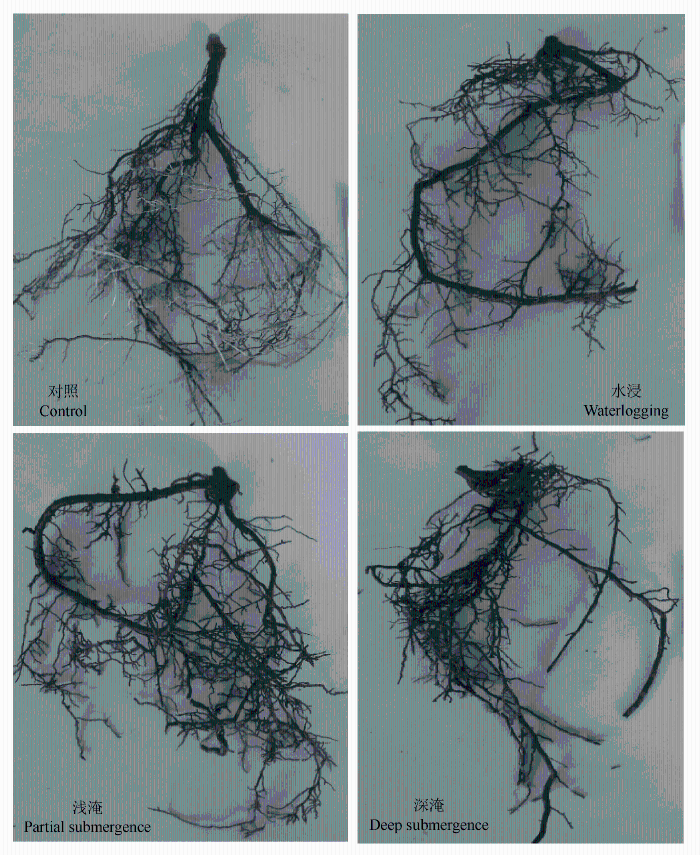
长期水淹显著抑制了中山杉地下部分的生长(表1 ), 对照根系鲜质量明显高于水淹处理的根系, 水浸与浅淹处理间无明显差异, 但深淹处理的根系鲜质量明显降低。根长随水淹深度的增加显著降低(p < 0.05)。不同水淹深度对根质量与根表面积的影响不显著(p > 0.05), 但深淹处理的中山杉根表面积最小。根系分形维数是表征根系在空间分布的指标, 经受不同水淹胁迫的中山杉根系分形维数明显降低, 但不同水淹处理的组间差异不显著(p > 0.05)。
经受不同水淹胁迫后的中山杉根茎处长有许多气生根, 且茎基部膨大。3种不同水淹胁迫处理的中山杉幼苗根系颜色呈黑色, 而对照的根系较浅, 这可能是因为水淹引起根系外壁木质化以及软木脂沉积物所致。另外经受水淹胁迫后中山杉根系复杂程度明显下降(图1 )。
图1 不同水淹处理中山杉气生根及茎基部变化。
Fig. 1 The changes of aerial root and basal part of stem in Taxodium ‘Zhongshansha 118’ under flooding treatments.
水淹胁迫同样抑制了中山杉地上部分的生长(表2 )。与对照相比不同水淹胁迫处理的株高和地径较小, 但不同水淹处理组间差异不显著(p > 0.05)。长期水淹胁迫后中山杉地上部分鲜质量随水淹深度的增加而显著降低(p < 0.05)。浅淹、深淹处理的幼苗根质量/地上部分鲜质量的比值显著大于对照及水浸处理(p < 0.05)。
2.2 水淹胁迫对中山杉根系活力的影响
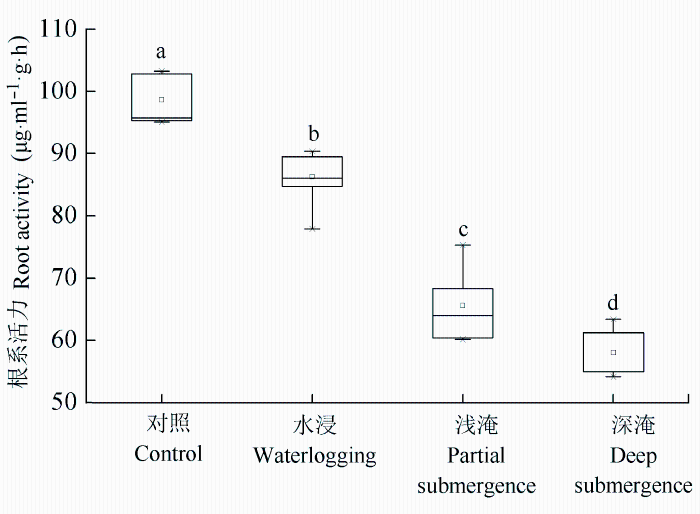
水淹胁迫使中山杉根系活力显著降低(p < 0.05) (图2 ), 但仍保持在50 μg·mL-1 ·g-1 ·h-1 以上。与对照相比, 水浸、浅淹与深淹处理的中山杉根系活力分别下降了12.57%、33.53%和41.18%。表明水淹胁迫降低了根系的合成与吸收能力, 影响了根系功能。
图2 长期水淹胁迫对中山杉幼苗根系活力的影响(平均值±标准偏差)。不同小写字母表示差异显著 (p < 0.05)。
Fig. 2 Effect on root activity of Taxodium ‘Zhongshansha 118’ during long-term flooding (mean ± SD ). Different lowercase letters indicate significant difference (p < 0.05).
2.3 水淹胁迫对中山杉无氧呼吸酶活性的影响
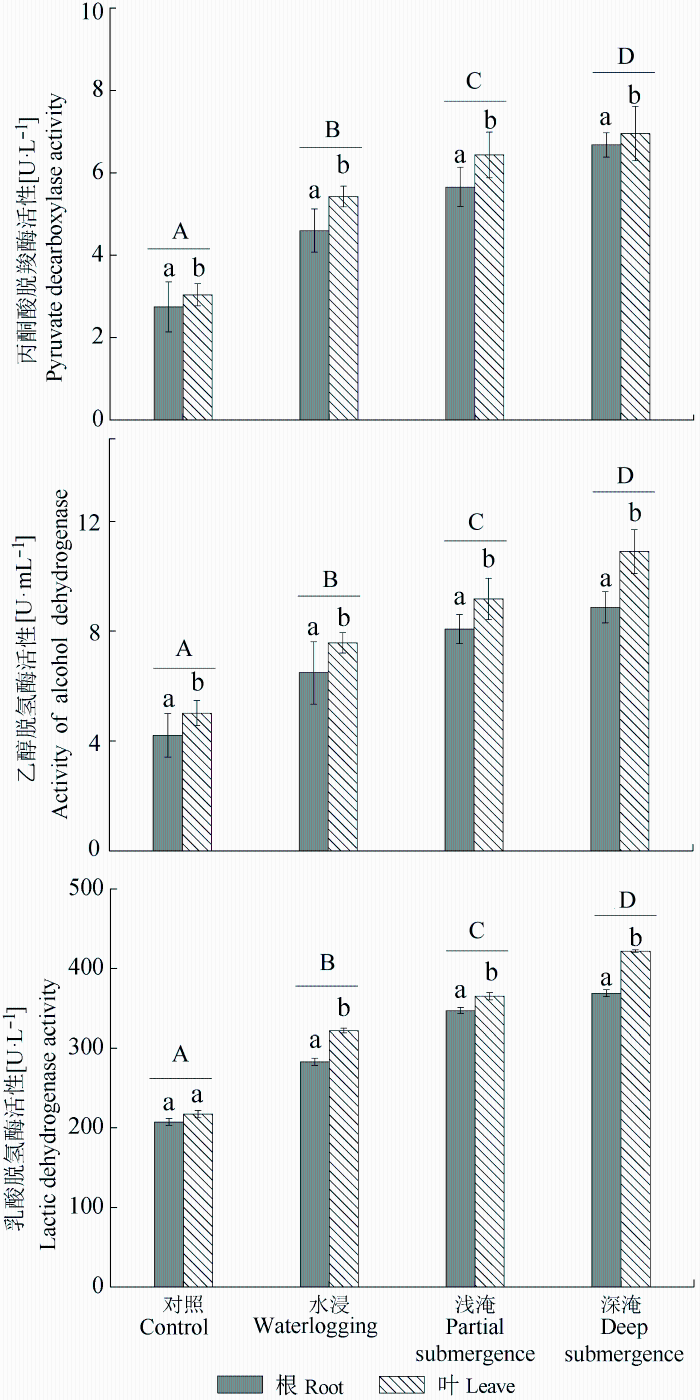
经长期水淹胁迫后中山杉根系和叶片中的丙酮酸脱羧酶活性(PDC)均随水淹深度的增加而显著增加(p < 0.05), 同时4种不同处理的叶片中PDC活性高于根系中PDC活性(图3 )。经长期水淹胁迫后中山杉根系和叶片中乙醇脱氢酶活性(ADH)和乳酸脱氢酶活性(LDH)与PDC活性具有相似的规律(图3 )。根系和叶片中ADH活性均高于对应的LDH活性, 这表明处于长期水淹胁迫时中山杉根系和叶片呼吸由线粒体有氧呼吸逐渐向以酒精发酵为主的无氧呼吸 转变。
深淹处理组的叶片与根系PDC活性的差值最小, 而叶片与根系ADH活性差值最大, 叶片与根系LDH活性的差值与ADH具有相似的规律, 这可能是由于3种酶的功能不同, 也说明深淹胁迫下中山杉叶片中乙醇代谢与乳酸代谢速率远高于根系。
图3 长期水淹胁迫对中山杉幼苗无氧呼吸酶活性的影响(平均值±标准偏差)。不同大写字母表示组间差异显著(p < 0.05), 不同小写字母表示组内差异显著(p < 0.05)。
Fig. 3 Effect on anaerobic respiration enzymes of Taxodium ‘Zhongshansha 118’ during long-term flooding (mean ± SD ). Capital letters indicate significant difference among groups (p < 0.05) and lowercase letters indicate significant difference in intra-group (p < 0.05).
2.4 水淹胁迫对中山杉淀粉、可溶性糖含量的影响
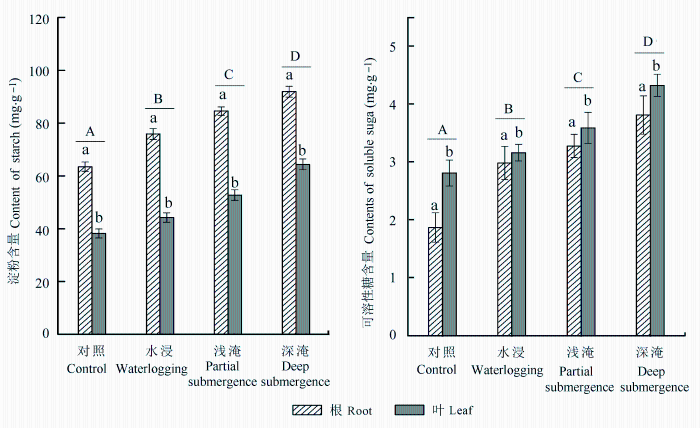
经过93天水淹胁迫的中山杉根系和叶片淀粉、可溶性糖含量均随水淹深度的增加显著增加(p <0.05)。4种不同处理的根系淀粉含量均高叶片, 而根系可溶性糖含量均低于叶片(图4 )。
图4 长期水淹胁迫对中山杉幼苗可溶性糖与淀粉含量的影响(平均值±标准偏差)。不同大写字母表示组间差异显著(p < 0.05), 不同小写字母表示组内差异显著(p < 0.05)。
Fig. 4 Effect on soluble sugar and starch of Taxodium ‘Zhongshansha 118’ during long-term flooding (mean ± SD ). Capital letters indicate significant difference among groups (p < 0.05) and lowercase letters indicate significant difference in intra-group (p < 0.05).
与对照组相比, 水浸、浅淹、深淹3种处理的中山杉根系可溶性糖含量分别增加了59.6%、75.3%和103.9%, 淀粉含量分别增加了19.5%、33.1%和44.7%, 根系可溶性糖的增长率高于淀粉的增长率。与对照组相比, 水浸、浅淹、深淹3种处理的中山杉叶片可溶性糖含量分别增加了12.51%、27.74%和53.90%, 淀粉含量分别增加了15.66%、37.89%和68.30%, 叶片淀粉的增长率均高于可溶性糖含量的增长率。
叶片与根系的总糖含量(淀粉+可溶性糖)同样随水淹深度的增加显著增加(p < 0.05)。叶片总糖含量明显低于根系总糖含量。根系淀粉含量/(根系+叶片)的淀粉含量比值随水淹深度的增加呈现减小的趋势, 但除了深淹处理(58%), 其他3个处理的比值均在60%以上。而根系可溶性糖含量/(根系+叶片) 可溶性糖含量的比值则呈现相反的趋势, 水淹处理的比值均高于对照处理(表3 )。
3 讨论
植物的耐淹性与PDC、ADH、LDH活性以及乙醇、酒精的积累密切相关(Cuiying et al ., 2010 ), ADH与LDH对于植物适应缺氧胁迫至关重要(Botondi et al ., 2012 )。在缺氧环境下植物从线粒体有氧呼吸转化为以酒精发酵和糖酵解为主的无氧呼吸。丙酮酸作为由无氧呼吸酶催化的乙醇或乳酸发酵的第一产物, PDC是乙醇代谢开始的关键因子。ADH活性与糖酵解途径的运行密切相关(Johnson et al ., 1994 ), 且ADH与乙烯的产生密切相关, 乙烯可以促进植物通气组织的形成, 提高根系对O2 的利用效率(Sairam et al ., 2008 )。对于水淹后ADH是否促进了中山杉根系乙烯的产生, 还有待进一步研究。与水淹后的Kosteletzkya virginica 幼苗根系相似(Zhou et al ., 2012 ), 水淹93天后, 中山杉幼苗根系和叶片的3种无氧呼吸酶活性均随水淹深度的增加显著增加(p < 0.05), 说明经受水淹处理后的幼苗叶片和根系呼吸方式均由以线粒体呼吸为主的有氧呼吸向以酒精发酵为主的无氧呼吸转变。但有氧呼吸仍旧存在, 因为经受水淹的中山杉在根茎结合部长出了细长的气生根, 且茎基部膨大, 这有助于增强体内的气体交换, 获得一定量的O2 (de Oliveira & Joly, 2009 )。同时由图1 看出中山杉根系颜色随水淹深度增加而加深, 表明水淹引起中山杉根系外壁木质化, 将根系与外界复杂的水淹环境隔离, 对根系起到保护作用, 同时防止根系内部O2 的损失(Sauter, 2013 )。与根系相比叶片所遭受的水淹胁迫程度较轻, 水浸处理的幼苗叶片全部出露水面, 浅淹处理的叶片大部分出露于水面之上, 深淹处理的叶片全部在水中, 但不同水淹处理的叶片中3种无氧呼吸酶活性(ADH、PDC、LDH)均高于根系, 间接证明了中山杉幼苗叶片对于缺氧环境更加敏感, 即使叶片没有处于水淹环境, 无氧呼吸同样也会加强。这是因为叶片是碳同化器官, 呼吸代谢相比根系要强, 而且根系产生的脱落酸(ABA)向上运输至叶片, 导致叶片气孔关闭, 进氧阻力加大, 从而使叶片无氧呼吸代谢加强。中山杉根系和叶片的ADH活性均高于LDH, 表明中山杉根系与叶片的呼吸途径均以酒精发酵为主, 乳酸发酵其次。这是因为酒精发酵和乳酸发酵均受细胞质pH值的影响, 乳酸发酵造成的细胞质酸化降低了细胞质pH值, 从而逐渐抑制乳酸发酵, 促进酒精发酵(Rivoal & Hanson, 1994 )。长时间水淹会改变土壤中的微生物菌丛, 水淹条件利于厌氧微生物的生存发展, 而厌氧微生物的代谢活动会产生对植物有害的毒性物质(如亚硝酸盐), 几乎没有植物能够适应这样的土壤条件(Johnson et al ., 1994 ), 对于经受水淹胁迫的中山杉根系与微生物菌丛间的关系, 以及根系如何适应含有毒性物质的土壤环境, 还有待进一步的研究。
无氧呼吸供能(每mol葡萄糖产生2 mol ATP)远低于三羧酸循环(每mol葡萄糖产生38 mol ATP), 且水淹胁迫使中山杉根系活力显著下降(p < 0.05), 根系向上输送水分、养分以及矿物质元素的能力显著降低, 因此经受水淹胁迫的中山杉生长明显受到抑制, 这与华建峰等(2011b)以及Visser等(2000)的研究结果相似。中山杉幼苗根质量/地上部分鲜质量的比值随水淹深度的增加呈现增加的趋势(p < 0.05), 这与池杉具有相似的规律, 由此说明中山杉将更多的生物量分配到了根系, 从而有助于提高其耐淹性(唐罗忠等, 2008 )。
糖类代谢对于植物适应水淹胁迫至关重要, 水淹前后的碳水化合物储量与植物耐淹性密切相关(施美芬等, 2010 ; Ferner et al ., 2012 ), 对水淹敏感的Fagus sylvatica 在经受水淹处理后, 根系可溶性糖和淀粉含量均下降, 而相对耐淹的夏栎(Quercus robur )的根系可溶性糖和淀粉含量则没有发生变化(Ferner et al ., 2012 ), 水淹使紫苜蓿(Medicago sativa ) 根系(Castonguay et al ., 1993 )和黄麻(Corchorus capsularis )根系(Parvin & Karmoker, 2013 )叶片的糖类含量增加。经受93天水淹胁迫的中山杉并没有表现出碳供应不足的现象, 水淹后的中山杉叶片与根系可溶性糖和淀粉含量均随水淹深度的增加显著增加(p < 0.05), 这是因为长期水淹使中山杉根系及叶片无氧呼吸加强, 有氧呼吸减弱, 植株对于糖类的利用效率降低, 因此出现非结构性糖的积累。由于中山杉生长受到抑制, 且出现淀粉及可溶性糖的积累, 所以推断中山杉对于生长季长期水淹的适应策略为忍耐型。很多草本植物在经受生长季短期水淹时, 表现出逃避型的适应策略(施美芬等, 2010 ; Voesenek & Bailey-Serres, 2013 ), 但是对于乔木来说, 其生长周期比草本长, 且生长速率比草本慢, 同时, 需要消耗大量糖类储备的逃避型适应策略对于植物种退水后的恢复生长并没有益处, 反而会使植物出现碳饥饿而威胁其生存, 因此以茎的快速伸长为主要特征的逃避型适应策略在一定程度上并不适合乔木树种, 而忍耐型策略则更加合适, 抑制生长造成的糖类积累为退水后的恢复生长提供了能量储备, 提高了退水后植物的保存率(施美芬等, 2010 )。这也是中山杉耐淹的重要特征。4种不同处理的中山杉根系和叶片淀粉含量均远高于可溶性糖含量, 说明淀粉的储备对于植物适应水淹胁迫发挥着重要的作用。
低氧环境下非结构性碳水化合物分配到不同器官的模式可能会影响植物的耐淹性(Angelov et al ., 1996 ; 王彪等, 2015 )。植物根系中高淀粉含量是植物耐淹的重要特征(Gravatt & Kirby, 1998 ), 中山杉根系淀粉含量远高于叶片, 虽然根系淀粉含量占总糖含量的百分比随水淹深度的增加呈下降趋势, 但是水淹对其没有显著影响(p > 0.05), 占比均在58%以上, 这是中山杉具有极强耐淹性的重要原因, 同时也表明水淹胁迫并没有严重抑制光合产物向根系的运输。由于根系通常为碳储藏器官, 叶片为碳同化器官, 所以中山杉根系可溶性糖含量明显低于叶片。可溶性糖可以提高组织细胞液渗透压(Molina- Montenegro et al ., 2012 ), 从而有助于叶片通过被动吸水的方式补充水分, 以弥补根系长期处于缺氧环境而无法正常向地上部分输送水分, 这也是中山杉叶片可溶性糖含量高于根系的一个原因。
植物的耐淹性及对水淹的生理响应与水淹季节密切相关(Crawford, 2003 ), 本研究初步从呼吸代谢以及物质消耗的角度论证了经受生长季93天水淹后的中山杉幼苗极强耐淹性的原因, 但是由于三峡库区消落带具有“冬水夏陆”的特点, 栽植在消落带内的植物主要遭受非生长季水淹胁迫, 中山杉同样在非生长季表现出了极强的耐淹性(殷云龙等, 2014 ; 张艳婷等, 2015 ), 对于中山杉非生长季的耐淹机理还有待进一步研究。同时, 经受水淹的植物种在退水后的恢复能力也是决定其耐淹能力的重要指标(Liu et al ., 2015 ), 中山杉在退水后的恢复能力同样需要进一步研究。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Akman M Bhikharie AV McLean EH Boonman A Visser EJ Schranz ME 2012 ). Wait or escape? Contrasting submergence tolerance strategies of Rorippa amphibia, Rorippa sylvestris and their hybrid.
Annals of botany , 109 , 1263 -1276 .
[本文引用: 1]
[2]
Angelov MN Sung S-JS Doong RL Harms WR Kormanik PP Clanton C Black J 1996 ). Long- and short-term flooding effects on survival and sink—Source relationships of swamp-adapted tree species.
Tree Physiology , 16 , 477 -484 .
[本文引用: 1]
[3]
Botondi R Russo V Mencarelli F 2012 ). Anaerobic metabolism during short and long term storage of kiwifruit.
Postharvest Biology and Technology , 64 , 83 -90 .
[本文引用: 1]
[4]
Candan N Tarhan L 2012 ). Tolerance or sensitivity responses of Mentha pulegium to osmotic and waterlogging stress in terms of antioxidant defense systems and membrane lipid peroxidation.
Environmental and Experimental Botany , 75 , 83 -88 .
[本文引用: 1]
[5]
Castonguay Y Nadeau P Simard RR 1993 ). Effects of flooding on carbohydrate and ABA levels in roots and shoots of alfalfa.
Plant, Cell & Environment , 16 , 695 -702 .
[本文引用: 1]
[6]
Crawford RM 2003 ). Seasonal differences in plant responses to flooding and anoxia.
Canadian Journal of Botany , 81 , 1224 -1246 .
[本文引用: 2]
[7]
Cuiying L Tuanhui B Fengwang M Mingyu H 2010 ). Hypoxia tolerance and adaptation of anaerobic respiration to hypoxia stress in two Malus species.
Scientia Horticulturae , 124 , 274 -279 .
[本文引用: 1]
[8]
de Oliveira VC Joly CA 2009 ). Flooding tolerance of Calophyllum brasiliense Camb. (Clusiaceae): Morphological, physiological and growth responses.
Trees , 24 , 185 -193 .
[本文引用: 1]
[9]
Fan DY Xiong GM Zhang AY Liu X Xie ZQ Li ZJ 2015 ). Effect of water-lever regulation on species selection for ecological restoration practice in the water-level fluctuation zone of Three Gorges Reservoir.
Chinese Journal of Plant Ecology , 39 , 416 -432 . (in Chinese with English abstract)[樊大勇 , 熊高明 , 张爱英 , 刘曦 , 谢宗强 , 李兆佳 (2015 ). 三峡库区水位调度对消落带生态修复中物种筛选实践的影响
. 植物生态学报 , 39 , 416 -432 .]
[本文引用: 1]
[10]
Ferner E Rennenberg H Kreuzwieser J 2012 ). Effect of flooding on C metabolism of flood-tolerant (Quercus robur) and non-tolerant (Fagus sylvatica) tree species.
Tree Physiology , 32 , 135 -145 .
[本文引用: 2]
[11]
Gravatt DA Kirby CJ 1998 ). Patterns of photosynthesis and starch allocation in seedlings of four bottomland hardwood tree species subjected to flooding.
Tree Physiology , 18 , 411 -417 .
[本文引用: 1]
[12]
Hua JF Hu LJ Du LJ Xu JH Yin YL 2011 a). Effects of soil water conditions on photosynthetic characteristics of Taxodium ‘Zhongshanshan 406’.
Ecology and Environmental Sciences , 20 (8-9 ), 1221 -1225 . (in Chinese with English abstract)[华建峰 , 胡李娟 , 杜丽娟 , 徐建华 , 殷云龙 (2011 ). 水分条件对中山杉406光合特性的影响
. 生态环境学报 , 20 (8-9 ), 1221 -1225 .]
[13]
Hua JF Yin YL Zhou DQ Yu CG Xu JH 2011 b). Effects of soil water conditions on growth and physiology of Taxodium ‘Zhongshanshan 406’.
Journal of Ecology and Rural Environment , 27 , 50 -54 . (in Chinese with English abstract)[华建峰 , 殷云龙 , 周冬琴 , 於朝广 , 徐建华 (2011 ). 不同水分条件对中山杉406生长与生理的影响
. 生态与农村环境学报 , 27 , 50 -54 .]
[14]
Jackson MB Ram PC 2003 ). Physiological and molecular basis of susceptibility and tolerance of rice plants to complete submergence.
Annals of botany , 91 , 227 -241 .
[本文引用: 1]
[15]
Johnson JR Cobb BC Drew MC 1994 ). Hypoxic induction of anoxia tolerance in roots of Adh1 null Zea mays L.
Plant Physiology , 105 , 61 -67 .
[本文引用: 2]
[16]
King AR Arnold MA 2012 ). Developmental stage and growth regulator concentration differentially affect vegetative propagation of select clones of Taxodium Rich.
Hortscience , 47 , 238 -248 .
[本文引用: 1]
[17]
Liu Y 2013 ). Cultivation and management of Ascendens mucronatum in middle and lower Yangtze River.
Modern Horticulture , (10 ), 22 -24 . (in Chinese with English abstract)[刘艳 (2013 ). 论长江中下游中山杉的栽培与管理
. 现代园艺 , (10 ), 22 -24 .]
[本文引用: 1]
[18]
Liu Y Willison JH 2013 ). Prospects for cultivating white mulberry (Morus alba) in the drawdown zone of the Three Gorges Reservoir, China.
Environmental Science and Pollution Research International , 20 , 7142 -7151 .
[本文引用: 1]
[19]
Liu Z Cheng R Xiao W Guo Q Wang Y Wang N Wang Y 2015 ). Leaf gas exchange, chlorophyll fluorescence, non-structural carbohydrate content and growth responses of Distylium chinense during complete submergence and subaerial re-emergence.
Aquatic Botany , 124 , 70 -77 .
[本文引用: 1]
[20]
Molina-Montenegro MA Gallardo-Cerda J Flores TSM Atala C 2012 ). The trade-off between cold resistance and growth determines the Nothofagus pumilio treeline.
Plant Ecology , 213 , 133 -142 .
[本文引用: 1]
[21]
Oren R Sperry JS Ewers BE Pataki DE Phillips N Megonigal JP 2001 ). Sensitivity of mean canopy stomatal conductance to vapor pressure deficit in a flooded Taxodium distichum L. forest: Hydraulic and non-hydraulic effects.
Oecologia , 126 , 21 -29 .
[本文引用: 1]
[22]
Parvin D Karmoker JL 2013 ). Effects of waterlogging on ion accumulation and sugar, protein and proline contents in Corchorus capsularis.
Bangladesh Journal of Botany , 42 , 55 -63 .
[本文引用: 1]
[23]
Rivoal J Hanson AD 1994 ). Metabolic control of anaerobic glycolysis—Overexpression of lactate dehydrogenase in transgenic tomato root supports the Davies-Roberts hypothesis and points to a critical role for lactate secretion.
Plant Physiology , 106 , 1179 -1185 .
[本文引用: 1]
[24]
Sairam , Kumutha D Ezhilmathi K Deshmukh PS Srivastava GC 2008 ). Physiology and biochemistry of waterlogging tolerance in plants.
Biologla Plantarum , 52 , 401 -412 .
[25]
Sauter M 2013 ). Root responses to flooding.
Current Opinion in Plant Biology , 16 , 282 -286 .
[本文引用: 2]
[26]
Shi MF Zeng B Shen JH Lei ST Zhu Z Liu JH 2010 ). A review of the correlation of flooding adaptability and carbohydrates in plants.
Chinese Journal of Plant Ecology , 34 , 855 -866 . (in Chinese with English abstract)[施美芬 , 曾波 , 申建红 , 类淑桐 , 朱智 , 刘建辉 (2010 ). 植物水淹适应与碳水化合物的相关性
. 植物生态学报 , 34 , 855 -866 .]
[本文引用: 4]
[27]
Tang LZ Huang BL Haibara K Toda H 2008 ) Ecologicaladaptation mechanisms of roots to flooded soil and respiration characteristics of knee roots of Taxodium ascendens
. Journal of Plant Ecology (Chinese Version) , 32 , 1258 -1267 . (in Chinese with English abstract)[唐罗忠 , 黄宝龙 , 生原喜久雄 , 户田浩人 (2008 ). 高水位条件下池杉根系的生态适应机制和膝根的呼吸特性
. 植物生态学报 , 32 , 1258 -1267 .]
[本文引用: 1]
[28]
Visser E Bogemann GM Steeg H Pierik R Blom C 2000 ). Flooding tolerance of Carex species in relation to field distribution and aerenchyma formation.
New Phytologist , 148 , 93 -103 .
[29]
Voesenek LA Bailey-Serres J 2013 ). Flooding tolerance: O2 sensing and survival strategies.
Current Opinion in Plant Biology , 16 , 647 -653 .
[本文引用: 1]
[30]
Wang B Jiang Y Wang MC Dong MY Zhang YP 2015 ). Variations of non-structural carbohydrate concentration of Picea meyeri at different elevations of Luya Mountain, China.
Chinese Journal of Plant Ecology , 39 , 746 -752 . (in Chinese with English abstract)[王彪 , 江源 , 王明昌 , 董满宇 , 章异平 (2015 ). 芦芽山不同海拔白杄非结构性碳水化合物含量动态
. 植物生态学报 , 39 , 746 -752 .]
[本文引用: 1]
[31]
Yemm BEW Willis AJ 1954 ). The estimation of carbohydrates in plant extracts by anthrone.
Biochemical Journal , 57 , 508 -514 .
[本文引用: 1]
[32]
Yin YL Yu CG Hua JF Huan JJ Han LW Qi BY Ren P Wu XH Qi XC 2014 ). A trial on the silviculture of Taxodium hybrid ‘Zhonshanshan118’ planted in the hydro-fluctuation belt of the Three Gorges Reservoir within the Wanzhou district area of Chongqing City.
China Forestry Science and Technology , 28 (2 ), 110 -114 . (in Chinese with English abstract)[殷云龙 , 於朝广 , 华建峰 , 环姣姣 , 韩路弯 , 亓白岩 , 任凭 , 吴晓洪 , 祁小川 (2014 ). 重庆万州三峡库区消落带中山杉造林实验
. 林业科技开发 , 28 (2 ), 110 -114 .]
[本文引用: 1]
[33]
Zhang YT Zhang JJ Wu XH Wang JX Duan FP 2015 ). Flooding tolerance of Taxodium hybrid ‘Zhongshanshan’ along the hydro-fluctuation belt of the Three Gorges Reservoir.
Science of Soil and Water Conservation ,13 (2 ), 56 -62 . (in Chinese with English abstract)[张艳婷 , 张建军 , 吴晓洪 , 王建修 , 段丰沛 (2015 ). 长江三峡库区消落带中山杉耐淹试验
. 中国水土保持科学 , 13 (2 ), 56 -62 .]
[本文引用: 1]
[34]
Zhou J Tian X Qiao L Qin P 2012 ). Respiratory enzyme activity and regulation of respiration pathway in seashore mallow (Kosteletzkya virginica) seedlings under waterlogging conditions.
Australian Journal of Crop Science , 6 , 756 -762 .
[本文引用: 2]
Wait or escape? Contrasting submergence tolerance strategies of Rorippa amphibia, Rorippa sylvestris and their hybrid.
1
2012
... 糖类是植物的能量来源, 植物体内的糖类主要包括结构性碳水化合物(主要用于植物形态建成)和非结构性碳水化合物(参与生命代谢), 非结构性碳水化合物(可溶性糖、淀粉等)的代谢在很大程度上决定了植物的生长发育及对环境的适应能力(施美芬等, 2010 ).可溶性糖参与植物体内的糖酵解过程, 为植物提供能量, 淀粉则是主要的能量储存形式.水淹对糖含量的影响较为复杂, 主要取决于植物对水淹的适应策略.植物适应水淹的策略主要包括逃避型和忍耐型.逃避型植物具发达的通气组织, 通过消耗糖类促进茎生长以接近水面, 从而增强植物体内部的气体交换, 获得更多O2 ; 而忍耐型植物通过抑制生长来储存能量(Akman et al ., 2012 ).目前对于植物适应水淹的策略研究大多集中在草本, 对木本植物的研究较少, 且木本植物适应水淹的策略现在还不明确. ...
Long- and short-term flooding effects on survival and sink—Source relationships of swamp-adapted tree species.
1
1996
... 低氧环境下非结构性碳水化合物分配到不同器官的模式可能会影响植物的耐淹性(Angelov et al ., 1996 ; 王彪等, 2015 ).植物根系中高淀粉含量是植物耐淹的重要特征(Gravatt & Kirby, 1998 ), 中山杉根系淀粉含量远高于叶片, 虽然根系淀粉含量占总糖含量的百分比随水淹深度的增加呈下降趋势, 但是水淹对其没有显著影响(p > 0.05), 占比均在58%以上, 这是中山杉具有极强耐淹性的重要原因, 同时也表明水淹胁迫并没有严重抑制光合产物向根系的运输.由于根系通常为碳储藏器官, 叶片为碳同化器官, 所以中山杉根系可溶性糖含量明显低于叶片.可溶性糖可以提高组织细胞液渗透压(Molina- Montenegro et al ., 2012 ), 从而有助于叶片通过被动吸水的方式补充水分, 以弥补根系长期处于缺氧环境而无法正常向地上部分输送水分, 这也是中山杉叶片可溶性糖含量高于根系的一个原因. ...
Anaerobic metabolism during short and long term storage of kiwifruit.
1
2012
... 植物的耐淹性与PDC、ADH、LDH活性以及乙醇、酒精的积累密切相关(Cuiying et al ., 2010 ), ADH与LDH对于植物适应缺氧胁迫至关重要(Botondi et al ., 2012 ).在缺氧环境下植物从线粒体有氧呼吸转化为以酒精发酵和糖酵解为主的无氧呼吸.丙酮酸作为由无氧呼吸酶催化的乙醇或乳酸发酵的第一产物, PDC是乙醇代谢开始的关键因子.ADH活性与糖酵解途径的运行密切相关(Johnson et al ., 1994 ), 且ADH与乙烯的产生密切相关, 乙烯可以促进植物通气组织的形成, 提高根系对O2 的利用效率(Sairam et al ., 2008 ).对于水淹后ADH是否促进了中山杉根系乙烯的产生, 还有待进一步研究.与水淹后的Kosteletzkya virginica 幼苗根系相似(Zhou et al ., 2012 ), 水淹93天后, 中山杉幼苗根系和叶片的3种无氧呼吸酶活性均随水淹深度的增加显著增加(p < 0.05), 说明经受水淹处理后的幼苗叶片和根系呼吸方式均由以线粒体呼吸为主的有氧呼吸向以酒精发酵为主的无氧呼吸转变.但有氧呼吸仍旧存在, 因为经受水淹的中山杉在根茎结合部长出了细长的气生根, 且茎基部膨大, 这有助于增强体内的气体交换, 获得一定量的O2 (de Oliveira & Joly, 2009 ).同时由图1 看出中山杉根系颜色随水淹深度增加而加深, 表明水淹引起中山杉根系外壁木质化, 将根系与外界复杂的水淹环境隔离, 对根系起到保护作用, 同时防止根系内部O2 的损失(Sauter, 2013 ).与根系相比叶片所遭受的水淹胁迫程度较轻, 水浸处理的幼苗叶片全部出露水面, 浅淹处理的叶片大部分出露于水面之上, 深淹处理的叶片全部在水中, 但不同水淹处理的叶片中3种无氧呼吸酶活性(ADH、PDC、LDH)均高于根系, 间接证明了中山杉幼苗叶片对于缺氧环境更加敏感, 即使叶片没有处于水淹环境, 无氧呼吸同样也会加强.这是因为叶片是碳同化器官, 呼吸代谢相比根系要强, 而且根系产生的脱落酸(ABA)向上运输至叶片, 导致叶片气孔关闭, 进氧阻力加大, 从而使叶片无氧呼吸代谢加强.中山杉根系和叶片的ADH活性均高于LDH, 表明中山杉根系与叶片的呼吸途径均以酒精发酵为主, 乳酸发酵其次.这是因为酒精发酵和乳酸发酵均受细胞质pH值的影响, 乳酸发酵造成的细胞质酸化降低了细胞质pH值, 从而逐渐抑制乳酸发酵, 促进酒精发酵(Rivoal & Hanson, 1994 ).长时间水淹会改变土壤中的微生物菌丛, 水淹条件利于厌氧微生物的生存发展, 而厌氧微生物的代谢活动会产生对植物有害的毒性物质(如亚硝酸盐), 几乎没有植物能够适应这样的土壤条件(Johnson et al ., 1994 ), 对于经受水淹胁迫的中山杉根系与微生物菌丛间的关系, 以及根系如何适应含有毒性物质的土壤环境, 还有待进一步的研究. ...
Tolerance or sensitivity responses of Mentha pulegium to osmotic and waterlogging stress in terms of antioxidant defense systems and membrane lipid peroxidation.
1
2012
... 水淹胁迫是影响植物生长、生存的主要胁迫.每年在世界各地都会发生不同程度和不同时间的水淹胁迫(Candan & Tarhan, 2012 ), 尤其是水库建设中形成的消落带, 其水淹胁迫发生频率更高, 程度更加严重.消落带每年周期性地遭受不同程度的水淹胁迫, 大部分陆生植物因无法忍受长时间、高强度的反季节水淹逐渐死亡(樊大勇等, 2015 ), 造成消落带植物群落向结构简单化、生物多样性急剧减少的方向发展(Liu & Willison, 2013 ).水淹使土壤中气体交换量减少, 导致根系缺氧, 功能紊乱.同时土壤微生物与根系争夺O2 , 使根系通过线粒体氧化磷酸化途径供能更加困难(Sauter, 2013 ), 进而影响根系的一切耗能活动(水分输送、养分吸收、植物体的糖类代谢以及光合作用和活性氧代谢等).植物在缺氧条件下会通过增加乙醇脱氢酶(ADH)、乳酸脱氢酶(LDH)以及丙酮酸脱羧酶(PDC)活性启动无氧呼吸, 以应对缺氧环境(Zhou et al ., 2012 ). ...
Effects of flooding on carbohydrate and ABA levels in roots and shoots of alfalfa.
1
1993
... 糖类代谢对于植物适应水淹胁迫至关重要, 水淹前后的碳水化合物储量与植物耐淹性密切相关(施美芬等, 2010 ; Ferner et al ., 2012 ), 对水淹敏感的Fagus sylvatica 在经受水淹处理后, 根系可溶性糖和淀粉含量均下降, 而相对耐淹的夏栎(Quercus robur )的根系可溶性糖和淀粉含量则没有发生变化(Ferner et al ., 2012 ), 水淹使紫苜蓿(Medicago sativa ) 根系(Castonguay et al ., 1993 )和黄麻(Corchorus capsularis )根系(Parvin & Karmoker, 2013 )叶片的糖类含量增加.经受93天水淹胁迫的中山杉并没有表现出碳供应不足的现象, 水淹后的中山杉叶片与根系可溶性糖和淀粉含量均随水淹深度的增加显著增加(p < 0.05), 这是因为长期水淹使中山杉根系及叶片无氧呼吸加强, 有氧呼吸减弱, 植株对于糖类的利用效率降低, 因此出现非结构性糖的积累.由于中山杉生长受到抑制, 且出现淀粉及可溶性糖的积累, 所以推断中山杉对于生长季长期水淹的适应策略为忍耐型.很多草本植物在经受生长季短期水淹时, 表现出逃避型的适应策略(施美芬等, 2010 ; Voesenek & Bailey-Serres, 2013 ), 但是对于乔木来说, 其生长周期比草本长, 且生长速率比草本慢, 同时, 需要消耗大量糖类储备的逃避型适应策略对于植物种退水后的恢复生长并没有益处, 反而会使植物出现碳饥饿而威胁其生存, 因此以茎的快速伸长为主要特征的逃避型适应策略在一定程度上并不适合乔木树种, 而忍耐型策略则更加合适, 抑制生长造成的糖类积累为退水后的恢复生长提供了能量储备, 提高了退水后植物的保存率(施美芬等, 2010 ).这也是中山杉耐淹的重要特征.4种不同处理的中山杉根系和叶片淀粉含量均远高于可溶性糖含量, 说明淀粉的储备对于植物适应水淹胁迫发挥着重要的作用. ...
Seasonal differences in plant responses to flooding and anoxia.
2
2003
... 中山杉(Taxodium ‘Zhongshansha’)是由江苏省中国科学院植物研究所通过落羽杉(Taxodium distichum )、墨西哥落羽杉(Taxodium mucronatum )以及池杉(Taxodium ascendens )种间杂交培育出的优良品种(华建峰等, 2011b ), 主要生长在包括三大湖(滇池、巢湖、太湖)以及三峡库区消落带(重庆)在内的长江中下游地区(刘艳, 2013 ).中山杉耐淹、耐碱、树形优美(King & Arnold, 2012 ), 其耐淹性尤为突出, 可以忍受较长时间的水淹胁迫(Oren et al ., 2001 ).对‘中山杉406’进行为期3个月的水淹试验后出现皮孔肥大、长出气生根等适应水淹的特征, 水淹并没有影响其光合特性(华建峰等, 2011a , 2011b ).但是中山杉水下呼吸机制以及糖类转化的研究鲜有报道.殷云龙等(2014)在重庆万州消落带内栽植两年生‘中山杉118’ ((落羽杉×墨西哥落羽杉)×墨西哥落羽杉)进行造林实验, 发现中山杉在非生长季经受122天、最大没顶深度12 m的水淹胁迫后, 仍有90%的保存率.植物的耐淹能力以及适应策略与水淹季节密切相关, 生长季植物新陈代谢速率较快, 需氧量更大(Crawford, 2003 ), 植物对生长季水淹的敏感度更大(Jackson & Ram, 2003 ).栽植在三峡库区万州段消落带内的中山杉表现出的极强耐淹性, 是其本身的遗传特性还是与消落带非生长季水淹的特殊环境有关, 中山杉又是通过何种策略适应不同季节的长期水淹等仍是需要探究的科学问题, 为此本文通过测定中山杉幼苗经受生长季不同程度水淹胁迫后叶片和根系的呼吸及糖类代谢, 从能量消耗的角度探索中山杉的耐淹机理, 以及中山杉幼苗对生长季水淹胁迫的适应策略, 为湿地生态修复和消落带生物治理中植物材料的选择提供依据. ...
... 植物的耐淹性及对水淹的生理响应与水淹季节密切相关(Crawford, 2003 ), 本研究初步从呼吸代谢以及物质消耗的角度论证了经受生长季93天水淹后的中山杉幼苗极强耐淹性的原因, 但是由于三峡库区消落带具有“冬水夏陆”的特点, 栽植在消落带内的植物主要遭受非生长季水淹胁迫, 中山杉同样在非生长季表现出了极强的耐淹性(殷云龙等, 2014 ; 张艳婷等, 2015 ), 对于中山杉非生长季的耐淹机理还有待进一步研究.同时, 经受水淹的植物种在退水后的恢复能力也是决定其耐淹能力的重要指标(Liu et al ., 2015 ), 中山杉在退水后的恢复能力同样需要进一步研究. ...
Hypoxia tolerance and adaptation of anaerobic respiration to hypoxia stress in two Malus species.
1
2010
... 植物的耐淹性与PDC、ADH、LDH活性以及乙醇、酒精的积累密切相关(Cuiying et al ., 2010 ), ADH与LDH对于植物适应缺氧胁迫至关重要(Botondi et al ., 2012 ).在缺氧环境下植物从线粒体有氧呼吸转化为以酒精发酵和糖酵解为主的无氧呼吸.丙酮酸作为由无氧呼吸酶催化的乙醇或乳酸发酵的第一产物, PDC是乙醇代谢开始的关键因子.ADH活性与糖酵解途径的运行密切相关(Johnson et al ., 1994 ), 且ADH与乙烯的产生密切相关, 乙烯可以促进植物通气组织的形成, 提高根系对O2 的利用效率(Sairam et al ., 2008 ).对于水淹后ADH是否促进了中山杉根系乙烯的产生, 还有待进一步研究.与水淹后的Kosteletzkya virginica 幼苗根系相似(Zhou et al ., 2012 ), 水淹93天后, 中山杉幼苗根系和叶片的3种无氧呼吸酶活性均随水淹深度的增加显著增加(p < 0.05), 说明经受水淹处理后的幼苗叶片和根系呼吸方式均由以线粒体呼吸为主的有氧呼吸向以酒精发酵为主的无氧呼吸转变.但有氧呼吸仍旧存在, 因为经受水淹的中山杉在根茎结合部长出了细长的气生根, 且茎基部膨大, 这有助于增强体内的气体交换, 获得一定量的O2 (de Oliveira & Joly, 2009 ).同时由图1 看出中山杉根系颜色随水淹深度增加而加深, 表明水淹引起中山杉根系外壁木质化, 将根系与外界复杂的水淹环境隔离, 对根系起到保护作用, 同时防止根系内部O2 的损失(Sauter, 2013 ).与根系相比叶片所遭受的水淹胁迫程度较轻, 水浸处理的幼苗叶片全部出露水面, 浅淹处理的叶片大部分出露于水面之上, 深淹处理的叶片全部在水中, 但不同水淹处理的叶片中3种无氧呼吸酶活性(ADH、PDC、LDH)均高于根系, 间接证明了中山杉幼苗叶片对于缺氧环境更加敏感, 即使叶片没有处于水淹环境, 无氧呼吸同样也会加强.这是因为叶片是碳同化器官, 呼吸代谢相比根系要强, 而且根系产生的脱落酸(ABA)向上运输至叶片, 导致叶片气孔关闭, 进氧阻力加大, 从而使叶片无氧呼吸代谢加强.中山杉根系和叶片的ADH活性均高于LDH, 表明中山杉根系与叶片的呼吸途径均以酒精发酵为主, 乳酸发酵其次.这是因为酒精发酵和乳酸发酵均受细胞质pH值的影响, 乳酸发酵造成的细胞质酸化降低了细胞质pH值, 从而逐渐抑制乳酸发酵, 促进酒精发酵(Rivoal & Hanson, 1994 ).长时间水淹会改变土壤中的微生物菌丛, 水淹条件利于厌氧微生物的生存发展, 而厌氧微生物的代谢活动会产生对植物有害的毒性物质(如亚硝酸盐), 几乎没有植物能够适应这样的土壤条件(Johnson et al ., 1994 ), 对于经受水淹胁迫的中山杉根系与微生物菌丛间的关系, 以及根系如何适应含有毒性物质的土壤环境, 还有待进一步的研究. ...
Flooding tolerance of Calophyllum brasiliense Camb. (Clusiaceae): Morphological, physiological and growth responses.
1
2009
... 植物的耐淹性与PDC、ADH、LDH活性以及乙醇、酒精的积累密切相关(Cuiying et al ., 2010 ), ADH与LDH对于植物适应缺氧胁迫至关重要(Botondi et al ., 2012 ).在缺氧环境下植物从线粒体有氧呼吸转化为以酒精发酵和糖酵解为主的无氧呼吸.丙酮酸作为由无氧呼吸酶催化的乙醇或乳酸发酵的第一产物, PDC是乙醇代谢开始的关键因子.ADH活性与糖酵解途径的运行密切相关(Johnson et al ., 1994 ), 且ADH与乙烯的产生密切相关, 乙烯可以促进植物通气组织的形成, 提高根系对O2 的利用效率(Sairam et al ., 2008 ).对于水淹后ADH是否促进了中山杉根系乙烯的产生, 还有待进一步研究.与水淹后的Kosteletzkya virginica 幼苗根系相似(Zhou et al ., 2012 ), 水淹93天后, 中山杉幼苗根系和叶片的3种无氧呼吸酶活性均随水淹深度的增加显著增加(p < 0.05), 说明经受水淹处理后的幼苗叶片和根系呼吸方式均由以线粒体呼吸为主的有氧呼吸向以酒精发酵为主的无氧呼吸转变.但有氧呼吸仍旧存在, 因为经受水淹的中山杉在根茎结合部长出了细长的气生根, 且茎基部膨大, 这有助于增强体内的气体交换, 获得一定量的O2 (de Oliveira & Joly, 2009 ).同时由图1 看出中山杉根系颜色随水淹深度增加而加深, 表明水淹引起中山杉根系外壁木质化, 将根系与外界复杂的水淹环境隔离, 对根系起到保护作用, 同时防止根系内部O2 的损失(Sauter, 2013 ).与根系相比叶片所遭受的水淹胁迫程度较轻, 水浸处理的幼苗叶片全部出露水面, 浅淹处理的叶片大部分出露于水面之上, 深淹处理的叶片全部在水中, 但不同水淹处理的叶片中3种无氧呼吸酶活性(ADH、PDC、LDH)均高于根系, 间接证明了中山杉幼苗叶片对于缺氧环境更加敏感, 即使叶片没有处于水淹环境, 无氧呼吸同样也会加强.这是因为叶片是碳同化器官, 呼吸代谢相比根系要强, 而且根系产生的脱落酸(ABA)向上运输至叶片, 导致叶片气孔关闭, 进氧阻力加大, 从而使叶片无氧呼吸代谢加强.中山杉根系和叶片的ADH活性均高于LDH, 表明中山杉根系与叶片的呼吸途径均以酒精发酵为主, 乳酸发酵其次.这是因为酒精发酵和乳酸发酵均受细胞质pH值的影响, 乳酸发酵造成的细胞质酸化降低了细胞质pH值, 从而逐渐抑制乳酸发酵, 促进酒精发酵(Rivoal & Hanson, 1994 ).长时间水淹会改变土壤中的微生物菌丛, 水淹条件利于厌氧微生物的生存发展, 而厌氧微生物的代谢活动会产生对植物有害的毒性物质(如亚硝酸盐), 几乎没有植物能够适应这样的土壤条件(Johnson et al ., 1994 ), 对于经受水淹胁迫的中山杉根系与微生物菌丛间的关系, 以及根系如何适应含有毒性物质的土壤环境, 还有待进一步的研究. ...
三峡库区水位调度对消落带生态修复中物种筛选实践的影响
1
2015
... 水淹胁迫是影响植物生长、生存的主要胁迫.每年在世界各地都会发生不同程度和不同时间的水淹胁迫(Candan & Tarhan, 2012 ), 尤其是水库建设中形成的消落带, 其水淹胁迫发生频率更高, 程度更加严重.消落带每年周期性地遭受不同程度的水淹胁迫, 大部分陆生植物因无法忍受长时间、高强度的反季节水淹逐渐死亡(樊大勇等, 2015 ), 造成消落带植物群落向结构简单化、生物多样性急剧减少的方向发展(Liu & Willison, 2013 ).水淹使土壤中气体交换量减少, 导致根系缺氧, 功能紊乱.同时土壤微生物与根系争夺O2 , 使根系通过线粒体氧化磷酸化途径供能更加困难(Sauter, 2013 ), 进而影响根系的一切耗能活动(水分输送、养分吸收、植物体的糖类代谢以及光合作用和活性氧代谢等).植物在缺氧条件下会通过增加乙醇脱氢酶(ADH)、乳酸脱氢酶(LDH)以及丙酮酸脱羧酶(PDC)活性启动无氧呼吸, 以应对缺氧环境(Zhou et al ., 2012 ). ...
Effect of flooding on C metabolism of flood-tolerant (Quercus robur) and non-tolerant (Fagus sylvatica) tree species.
2
2012
... 糖类代谢对于植物适应水淹胁迫至关重要, 水淹前后的碳水化合物储量与植物耐淹性密切相关(施美芬等, 2010 ; Ferner et al ., 2012 ), 对水淹敏感的Fagus sylvatica 在经受水淹处理后, 根系可溶性糖和淀粉含量均下降, 而相对耐淹的夏栎(Quercus robur )的根系可溶性糖和淀粉含量则没有发生变化(Ferner et al ., 2012 ), 水淹使紫苜蓿(Medicago sativa ) 根系(Castonguay et al ., 1993 )和黄麻(Corchorus capsularis )根系(Parvin & Karmoker, 2013 )叶片的糖类含量增加.经受93天水淹胁迫的中山杉并没有表现出碳供应不足的现象, 水淹后的中山杉叶片与根系可溶性糖和淀粉含量均随水淹深度的增加显著增加(p < 0.05), 这是因为长期水淹使中山杉根系及叶片无氧呼吸加强, 有氧呼吸减弱, 植株对于糖类的利用效率降低, 因此出现非结构性糖的积累.由于中山杉生长受到抑制, 且出现淀粉及可溶性糖的积累, 所以推断中山杉对于生长季长期水淹的适应策略为忍耐型.很多草本植物在经受生长季短期水淹时, 表现出逃避型的适应策略(施美芬等, 2010 ; Voesenek & Bailey-Serres, 2013 ), 但是对于乔木来说, 其生长周期比草本长, 且生长速率比草本慢, 同时, 需要消耗大量糖类储备的逃避型适应策略对于植物种退水后的恢复生长并没有益处, 反而会使植物出现碳饥饿而威胁其生存, 因此以茎的快速伸长为主要特征的逃避型适应策略在一定程度上并不适合乔木树种, 而忍耐型策略则更加合适, 抑制生长造成的糖类积累为退水后的恢复生长提供了能量储备, 提高了退水后植物的保存率(施美芬等, 2010 ).这也是中山杉耐淹的重要特征.4种不同处理的中山杉根系和叶片淀粉含量均远高于可溶性糖含量, 说明淀粉的储备对于植物适应水淹胁迫发挥着重要的作用. ...
... )的根系可溶性糖和淀粉含量则没有发生变化(Ferner et al ., 2012 ), 水淹使紫苜蓿(Medicago sativa ) 根系(Castonguay et al ., 1993 )和黄麻(Corchorus capsularis )根系(Parvin & Karmoker, 2013 )叶片的糖类含量增加.经受93天水淹胁迫的中山杉并没有表现出碳供应不足的现象, 水淹后的中山杉叶片与根系可溶性糖和淀粉含量均随水淹深度的增加显著增加(p < 0.05), 这是因为长期水淹使中山杉根系及叶片无氧呼吸加强, 有氧呼吸减弱, 植株对于糖类的利用效率降低, 因此出现非结构性糖的积累.由于中山杉生长受到抑制, 且出现淀粉及可溶性糖的积累, 所以推断中山杉对于生长季长期水淹的适应策略为忍耐型.很多草本植物在经受生长季短期水淹时, 表现出逃避型的适应策略(施美芬等, 2010 ; Voesenek & Bailey-Serres, 2013 ), 但是对于乔木来说, 其生长周期比草本长, 且生长速率比草本慢, 同时, 需要消耗大量糖类储备的逃避型适应策略对于植物种退水后的恢复生长并没有益处, 反而会使植物出现碳饥饿而威胁其生存, 因此以茎的快速伸长为主要特征的逃避型适应策略在一定程度上并不适合乔木树种, 而忍耐型策略则更加合适, 抑制生长造成的糖类积累为退水后的恢复生长提供了能量储备, 提高了退水后植物的保存率(施美芬等, 2010 ).这也是中山杉耐淹的重要特征.4种不同处理的中山杉根系和叶片淀粉含量均远高于可溶性糖含量, 说明淀粉的储备对于植物适应水淹胁迫发挥着重要的作用. ...
Patterns of photosynthesis and starch allocation in seedlings of four bottomland hardwood tree species subjected to flooding.
1
1998
... 低氧环境下非结构性碳水化合物分配到不同器官的模式可能会影响植物的耐淹性(Angelov et al ., 1996 ; 王彪等, 2015 ).植物根系中高淀粉含量是植物耐淹的重要特征(Gravatt & Kirby, 1998 ), 中山杉根系淀粉含量远高于叶片, 虽然根系淀粉含量占总糖含量的百分比随水淹深度的增加呈下降趋势, 但是水淹对其没有显著影响(p > 0.05), 占比均在58%以上, 这是中山杉具有极强耐淹性的重要原因, 同时也表明水淹胁迫并没有严重抑制光合产物向根系的运输.由于根系通常为碳储藏器官, 叶片为碳同化器官, 所以中山杉根系可溶性糖含量明显低于叶片.可溶性糖可以提高组织细胞液渗透压(Molina- Montenegro et al ., 2012 ), 从而有助于叶片通过被动吸水的方式补充水分, 以弥补根系长期处于缺氧环境而无法正常向地上部分输送水分, 这也是中山杉叶片可溶性糖含量高于根系的一个原因. ...
不同水分条件对中山杉406生长与生理的影响
2011
Physiological and molecular basis of susceptibility and tolerance of rice plants to complete submergence.
1
2003
... 中山杉(Taxodium ‘Zhongshansha’)是由江苏省中国科学院植物研究所通过落羽杉(Taxodium distichum )、墨西哥落羽杉(Taxodium mucronatum )以及池杉(Taxodium ascendens )种间杂交培育出的优良品种(华建峰等, 2011b ), 主要生长在包括三大湖(滇池、巢湖、太湖)以及三峡库区消落带(重庆)在内的长江中下游地区(刘艳, 2013 ).中山杉耐淹、耐碱、树形优美(King & Arnold, 2012 ), 其耐淹性尤为突出, 可以忍受较长时间的水淹胁迫(Oren et al ., 2001 ).对‘中山杉406’进行为期3个月的水淹试验后出现皮孔肥大、长出气生根等适应水淹的特征, 水淹并没有影响其光合特性(华建峰等, 2011a , 2011b ).但是中山杉水下呼吸机制以及糖类转化的研究鲜有报道.殷云龙等(2014)在重庆万州消落带内栽植两年生‘中山杉118’ ((落羽杉×墨西哥落羽杉)×墨西哥落羽杉)进行造林实验, 发现中山杉在非生长季经受122天、最大没顶深度12 m的水淹胁迫后, 仍有90%的保存率.植物的耐淹能力以及适应策略与水淹季节密切相关, 生长季植物新陈代谢速率较快, 需氧量更大(Crawford, 2003 ), 植物对生长季水淹的敏感度更大(Jackson & Ram, 2003 ).栽植在三峡库区万州段消落带内的中山杉表现出的极强耐淹性, 是其本身的遗传特性还是与消落带非生长季水淹的特殊环境有关, 中山杉又是通过何种策略适应不同季节的长期水淹等仍是需要探究的科学问题, 为此本文通过测定中山杉幼苗经受生长季不同程度水淹胁迫后叶片和根系的呼吸及糖类代谢, 从能量消耗的角度探索中山杉的耐淹机理, 以及中山杉幼苗对生长季水淹胁迫的适应策略, 为湿地生态修复和消落带生物治理中植物材料的选择提供依据. ...
Hypoxic induction of anoxia tolerance in roots of Adh1 null Zea mays L.
2
1994
... 植物的耐淹性与PDC、ADH、LDH活性以及乙醇、酒精的积累密切相关(Cuiying et al ., 2010 ), ADH与LDH对于植物适应缺氧胁迫至关重要(Botondi et al ., 2012 ).在缺氧环境下植物从线粒体有氧呼吸转化为以酒精发酵和糖酵解为主的无氧呼吸.丙酮酸作为由无氧呼吸酶催化的乙醇或乳酸发酵的第一产物, PDC是乙醇代谢开始的关键因子.ADH活性与糖酵解途径的运行密切相关(Johnson et al ., 1994 ), 且ADH与乙烯的产生密切相关, 乙烯可以促进植物通气组织的形成, 提高根系对O2 的利用效率(Sairam et al ., 2008 ).对于水淹后ADH是否促进了中山杉根系乙烯的产生, 还有待进一步研究.与水淹后的Kosteletzkya virginica 幼苗根系相似(Zhou et al ., 2012 ), 水淹93天后, 中山杉幼苗根系和叶片的3种无氧呼吸酶活性均随水淹深度的增加显著增加(p < 0.05), 说明经受水淹处理后的幼苗叶片和根系呼吸方式均由以线粒体呼吸为主的有氧呼吸向以酒精发酵为主的无氧呼吸转变.但有氧呼吸仍旧存在, 因为经受水淹的中山杉在根茎结合部长出了细长的气生根, 且茎基部膨大, 这有助于增强体内的气体交换, 获得一定量的O2 (de Oliveira & Joly, 2009 ).同时由图1 看出中山杉根系颜色随水淹深度增加而加深, 表明水淹引起中山杉根系外壁木质化, 将根系与外界复杂的水淹环境隔离, 对根系起到保护作用, 同时防止根系内部O2 的损失(Sauter, 2013 ).与根系相比叶片所遭受的水淹胁迫程度较轻, 水浸处理的幼苗叶片全部出露水面, 浅淹处理的叶片大部分出露于水面之上, 深淹处理的叶片全部在水中, 但不同水淹处理的叶片中3种无氧呼吸酶活性(ADH、PDC、LDH)均高于根系, 间接证明了中山杉幼苗叶片对于缺氧环境更加敏感, 即使叶片没有处于水淹环境, 无氧呼吸同样也会加强.这是因为叶片是碳同化器官, 呼吸代谢相比根系要强, 而且根系产生的脱落酸(ABA)向上运输至叶片, 导致叶片气孔关闭, 进氧阻力加大, 从而使叶片无氧呼吸代谢加强.中山杉根系和叶片的ADH活性均高于LDH, 表明中山杉根系与叶片的呼吸途径均以酒精发酵为主, 乳酸发酵其次.这是因为酒精发酵和乳酸发酵均受细胞质pH值的影响, 乳酸发酵造成的细胞质酸化降低了细胞质pH值, 从而逐渐抑制乳酸发酵, 促进酒精发酵(Rivoal & Hanson, 1994 ).长时间水淹会改变土壤中的微生物菌丛, 水淹条件利于厌氧微生物的生存发展, 而厌氧微生物的代谢活动会产生对植物有害的毒性物质(如亚硝酸盐), 几乎没有植物能够适应这样的土壤条件(Johnson et al ., 1994 ), 对于经受水淹胁迫的中山杉根系与微生物菌丛间的关系, 以及根系如何适应含有毒性物质的土壤环境, 还有待进一步的研究. ...
... ).长时间水淹会改变土壤中的微生物菌丛, 水淹条件利于厌氧微生物的生存发展, 而厌氧微生物的代谢活动会产生对植物有害的毒性物质(如亚硝酸盐), 几乎没有植物能够适应这样的土壤条件(Johnson et al ., 1994 ), 对于经受水淹胁迫的中山杉根系与微生物菌丛间的关系, 以及根系如何适应含有毒性物质的土壤环境, 还有待进一步的研究. ...
Developmental stage and growth regulator concentration differentially affect vegetative propagation of select clones of Taxodium Rich.
1
2012
... 中山杉(Taxodium ‘Zhongshansha’)是由江苏省中国科学院植物研究所通过落羽杉(Taxodium distichum )、墨西哥落羽杉(Taxodium mucronatum )以及池杉(Taxodium ascendens )种间杂交培育出的优良品种(华建峰等, 2011b ), 主要生长在包括三大湖(滇池、巢湖、太湖)以及三峡库区消落带(重庆)在内的长江中下游地区(刘艳, 2013 ).中山杉耐淹、耐碱、树形优美(King & Arnold, 2012 ), 其耐淹性尤为突出, 可以忍受较长时间的水淹胁迫(Oren et al ., 2001 ).对‘中山杉406’进行为期3个月的水淹试验后出现皮孔肥大、长出气生根等适应水淹的特征, 水淹并没有影响其光合特性(华建峰等, 2011a , 2011b ).但是中山杉水下呼吸机制以及糖类转化的研究鲜有报道.殷云龙等(2014)在重庆万州消落带内栽植两年生‘中山杉118’ ((落羽杉×墨西哥落羽杉)×墨西哥落羽杉)进行造林实验, 发现中山杉在非生长季经受122天、最大没顶深度12 m的水淹胁迫后, 仍有90%的保存率.植物的耐淹能力以及适应策略与水淹季节密切相关, 生长季植物新陈代谢速率较快, 需氧量更大(Crawford, 2003 ), 植物对生长季水淹的敏感度更大(Jackson & Ram, 2003 ).栽植在三峡库区万州段消落带内的中山杉表现出的极强耐淹性, 是其本身的遗传特性还是与消落带非生长季水淹的特殊环境有关, 中山杉又是通过何种策略适应不同季节的长期水淹等仍是需要探究的科学问题, 为此本文通过测定中山杉幼苗经受生长季不同程度水淹胁迫后叶片和根系的呼吸及糖类代谢, 从能量消耗的角度探索中山杉的耐淹机理, 以及中山杉幼苗对生长季水淹胁迫的适应策略, 为湿地生态修复和消落带生物治理中植物材料的选择提供依据. ...
论长江中下游中山杉的栽培与管理
1
2013
... 中山杉(Taxodium ‘Zhongshansha’)是由江苏省中国科学院植物研究所通过落羽杉(Taxodium distichum )、墨西哥落羽杉(Taxodium mucronatum )以及池杉(Taxodium ascendens )种间杂交培育出的优良品种(华建峰等, 2011b ), 主要生长在包括三大湖(滇池、巢湖、太湖)以及三峡库区消落带(重庆)在内的长江中下游地区(刘艳, 2013 ).中山杉耐淹、耐碱、树形优美(King & Arnold, 2012 ), 其耐淹性尤为突出, 可以忍受较长时间的水淹胁迫(Oren et al ., 2001 ).对‘中山杉406’进行为期3个月的水淹试验后出现皮孔肥大、长出气生根等适应水淹的特征, 水淹并没有影响其光合特性(华建峰等, 2011a , 2011b ).但是中山杉水下呼吸机制以及糖类转化的研究鲜有报道.殷云龙等(2014)在重庆万州消落带内栽植两年生‘中山杉118’ ((落羽杉×墨西哥落羽杉)×墨西哥落羽杉)进行造林实验, 发现中山杉在非生长季经受122天、最大没顶深度12 m的水淹胁迫后, 仍有90%的保存率.植物的耐淹能力以及适应策略与水淹季节密切相关, 生长季植物新陈代谢速率较快, 需氧量更大(Crawford, 2003 ), 植物对生长季水淹的敏感度更大(Jackson & Ram, 2003 ).栽植在三峡库区万州段消落带内的中山杉表现出的极强耐淹性, 是其本身的遗传特性还是与消落带非生长季水淹的特殊环境有关, 中山杉又是通过何种策略适应不同季节的长期水淹等仍是需要探究的科学问题, 为此本文通过测定中山杉幼苗经受生长季不同程度水淹胁迫后叶片和根系的呼吸及糖类代谢, 从能量消耗的角度探索中山杉的耐淹机理, 以及中山杉幼苗对生长季水淹胁迫的适应策略, 为湿地生态修复和消落带生物治理中植物材料的选择提供依据. ...
Prospects for cultivating white mulberry (Morus alba) in the drawdown zone of the Three Gorges Reservoir, China.
1
2013
... 水淹胁迫是影响植物生长、生存的主要胁迫.每年在世界各地都会发生不同程度和不同时间的水淹胁迫(Candan & Tarhan, 2012 ), 尤其是水库建设中形成的消落带, 其水淹胁迫发生频率更高, 程度更加严重.消落带每年周期性地遭受不同程度的水淹胁迫, 大部分陆生植物因无法忍受长时间、高强度的反季节水淹逐渐死亡(樊大勇等, 2015 ), 造成消落带植物群落向结构简单化、生物多样性急剧减少的方向发展(Liu & Willison, 2013 ).水淹使土壤中气体交换量减少, 导致根系缺氧, 功能紊乱.同时土壤微生物与根系争夺O2 , 使根系通过线粒体氧化磷酸化途径供能更加困难(Sauter, 2013 ), 进而影响根系的一切耗能活动(水分输送、养分吸收、植物体的糖类代谢以及光合作用和活性氧代谢等).植物在缺氧条件下会通过增加乙醇脱氢酶(ADH)、乳酸脱氢酶(LDH)以及丙酮酸脱羧酶(PDC)活性启动无氧呼吸, 以应对缺氧环境(Zhou et al ., 2012 ). ...
Leaf gas exchange, chlorophyll fluorescence, non-structural carbohydrate content and growth responses of Distylium chinense during complete submergence and subaerial re-emergence.
1
2015
... 植物的耐淹性及对水淹的生理响应与水淹季节密切相关(Crawford, 2003 ), 本研究初步从呼吸代谢以及物质消耗的角度论证了经受生长季93天水淹后的中山杉幼苗极强耐淹性的原因, 但是由于三峡库区消落带具有“冬水夏陆”的特点, 栽植在消落带内的植物主要遭受非生长季水淹胁迫, 中山杉同样在非生长季表现出了极强的耐淹性(殷云龙等, 2014 ; 张艳婷等, 2015 ), 对于中山杉非生长季的耐淹机理还有待进一步研究.同时, 经受水淹的植物种在退水后的恢复能力也是决定其耐淹能力的重要指标(Liu et al ., 2015 ), 中山杉在退水后的恢复能力同样需要进一步研究. ...
The trade-off between cold resistance and growth determines the Nothofagus pumilio treeline.
1
2012
... 低氧环境下非结构性碳水化合物分配到不同器官的模式可能会影响植物的耐淹性(Angelov et al ., 1996 ; 王彪等, 2015 ).植物根系中高淀粉含量是植物耐淹的重要特征(Gravatt & Kirby, 1998 ), 中山杉根系淀粉含量远高于叶片, 虽然根系淀粉含量占总糖含量的百分比随水淹深度的增加呈下降趋势, 但是水淹对其没有显著影响(p > 0.05), 占比均在58%以上, 这是中山杉具有极强耐淹性的重要原因, 同时也表明水淹胁迫并没有严重抑制光合产物向根系的运输.由于根系通常为碳储藏器官, 叶片为碳同化器官, 所以中山杉根系可溶性糖含量明显低于叶片.可溶性糖可以提高组织细胞液渗透压(Molina- Montenegro et al ., 2012 ), 从而有助于叶片通过被动吸水的方式补充水分, 以弥补根系长期处于缺氧环境而无法正常向地上部分输送水分, 这也是中山杉叶片可溶性糖含量高于根系的一个原因. ...
Sensitivity of mean canopy stomatal conductance to vapor pressure deficit in a flooded Taxodium distichum L. forest: Hydraulic and non-hydraulic effects.
1
2001
... 中山杉(Taxodium ‘Zhongshansha’)是由江苏省中国科学院植物研究所通过落羽杉(Taxodium distichum )、墨西哥落羽杉(Taxodium mucronatum )以及池杉(Taxodium ascendens )种间杂交培育出的优良品种(华建峰等, 2011b ), 主要生长在包括三大湖(滇池、巢湖、太湖)以及三峡库区消落带(重庆)在内的长江中下游地区(刘艳, 2013 ).中山杉耐淹、耐碱、树形优美(King & Arnold, 2012 ), 其耐淹性尤为突出, 可以忍受较长时间的水淹胁迫(Oren et al ., 2001 ).对‘中山杉406’进行为期3个月的水淹试验后出现皮孔肥大、长出气生根等适应水淹的特征, 水淹并没有影响其光合特性(华建峰等, 2011a , 2011b ).但是中山杉水下呼吸机制以及糖类转化的研究鲜有报道.殷云龙等(2014)在重庆万州消落带内栽植两年生‘中山杉118’ ((落羽杉×墨西哥落羽杉)×墨西哥落羽杉)进行造林实验, 发现中山杉在非生长季经受122天、最大没顶深度12 m的水淹胁迫后, 仍有90%的保存率.植物的耐淹能力以及适应策略与水淹季节密切相关, 生长季植物新陈代谢速率较快, 需氧量更大(Crawford, 2003 ), 植物对生长季水淹的敏感度更大(Jackson & Ram, 2003 ).栽植在三峡库区万州段消落带内的中山杉表现出的极强耐淹性, 是其本身的遗传特性还是与消落带非生长季水淹的特殊环境有关, 中山杉又是通过何种策略适应不同季节的长期水淹等仍是需要探究的科学问题, 为此本文通过测定中山杉幼苗经受生长季不同程度水淹胁迫后叶片和根系的呼吸及糖类代谢, 从能量消耗的角度探索中山杉的耐淹机理, 以及中山杉幼苗对生长季水淹胁迫的适应策略, 为湿地生态修复和消落带生物治理中植物材料的选择提供依据. ...
Effects of waterlogging on ion accumulation and sugar, protein and proline contents in Corchorus capsularis.
1
2013
... 糖类代谢对于植物适应水淹胁迫至关重要, 水淹前后的碳水化合物储量与植物耐淹性密切相关(施美芬等, 2010 ; Ferner et al ., 2012 ), 对水淹敏感的Fagus sylvatica 在经受水淹处理后, 根系可溶性糖和淀粉含量均下降, 而相对耐淹的夏栎(Quercus robur )的根系可溶性糖和淀粉含量则没有发生变化(Ferner et al ., 2012 ), 水淹使紫苜蓿(Medicago sativa ) 根系(Castonguay et al ., 1993 )和黄麻(Corchorus capsularis )根系(Parvin & Karmoker, 2013 )叶片的糖类含量增加.经受93天水淹胁迫的中山杉并没有表现出碳供应不足的现象, 水淹后的中山杉叶片与根系可溶性糖和淀粉含量均随水淹深度的增加显著增加(p < 0.05), 这是因为长期水淹使中山杉根系及叶片无氧呼吸加强, 有氧呼吸减弱, 植株对于糖类的利用效率降低, 因此出现非结构性糖的积累.由于中山杉生长受到抑制, 且出现淀粉及可溶性糖的积累, 所以推断中山杉对于生长季长期水淹的适应策略为忍耐型.很多草本植物在经受生长季短期水淹时, 表现出逃避型的适应策略(施美芬等, 2010 ; Voesenek & Bailey-Serres, 2013 ), 但是对于乔木来说, 其生长周期比草本长, 且生长速率比草本慢, 同时, 需要消耗大量糖类储备的逃避型适应策略对于植物种退水后的恢复生长并没有益处, 反而会使植物出现碳饥饿而威胁其生存, 因此以茎的快速伸长为主要特征的逃避型适应策略在一定程度上并不适合乔木树种, 而忍耐型策略则更加合适, 抑制生长造成的糖类积累为退水后的恢复生长提供了能量储备, 提高了退水后植物的保存率(施美芬等, 2010 ).这也是中山杉耐淹的重要特征.4种不同处理的中山杉根系和叶片淀粉含量均远高于可溶性糖含量, 说明淀粉的储备对于植物适应水淹胁迫发挥着重要的作用. ...
Metabolic control of anaerobic glycolysis—Overexpression of lactate dehydrogenase in transgenic tomato root supports the Davies-Roberts hypothesis and points to a critical role for lactate secretion.
1
1994
... 植物的耐淹性与PDC、ADH、LDH活性以及乙醇、酒精的积累密切相关(Cuiying et al ., 2010 ), ADH与LDH对于植物适应缺氧胁迫至关重要(Botondi et al ., 2012 ).在缺氧环境下植物从线粒体有氧呼吸转化为以酒精发酵和糖酵解为主的无氧呼吸.丙酮酸作为由无氧呼吸酶催化的乙醇或乳酸发酵的第一产物, PDC是乙醇代谢开始的关键因子.ADH活性与糖酵解途径的运行密切相关(Johnson et al ., 1994 ), 且ADH与乙烯的产生密切相关, 乙烯可以促进植物通气组织的形成, 提高根系对O2 的利用效率(Sairam et al ., 2008 ).对于水淹后ADH是否促进了中山杉根系乙烯的产生, 还有待进一步研究.与水淹后的Kosteletzkya virginica 幼苗根系相似(Zhou et al ., 2012 ), 水淹93天后, 中山杉幼苗根系和叶片的3种无氧呼吸酶活性均随水淹深度的增加显著增加(p < 0.05), 说明经受水淹处理后的幼苗叶片和根系呼吸方式均由以线粒体呼吸为主的有氧呼吸向以酒精发酵为主的无氧呼吸转变.但有氧呼吸仍旧存在, 因为经受水淹的中山杉在根茎结合部长出了细长的气生根, 且茎基部膨大, 这有助于增强体内的气体交换, 获得一定量的O2 (de Oliveira & Joly, 2009 ).同时由图1 看出中山杉根系颜色随水淹深度增加而加深, 表明水淹引起中山杉根系外壁木质化, 将根系与外界复杂的水淹环境隔离, 对根系起到保护作用, 同时防止根系内部O2 的损失(Sauter, 2013 ).与根系相比叶片所遭受的水淹胁迫程度较轻, 水浸处理的幼苗叶片全部出露水面, 浅淹处理的叶片大部分出露于水面之上, 深淹处理的叶片全部在水中, 但不同水淹处理的叶片中3种无氧呼吸酶活性(ADH、PDC、LDH)均高于根系, 间接证明了中山杉幼苗叶片对于缺氧环境更加敏感, 即使叶片没有处于水淹环境, 无氧呼吸同样也会加强.这是因为叶片是碳同化器官, 呼吸代谢相比根系要强, 而且根系产生的脱落酸(ABA)向上运输至叶片, 导致叶片气孔关闭, 进氧阻力加大, 从而使叶片无氧呼吸代谢加强.中山杉根系和叶片的ADH活性均高于LDH, 表明中山杉根系与叶片的呼吸途径均以酒精发酵为主, 乳酸发酵其次.这是因为酒精发酵和乳酸发酵均受细胞质pH值的影响, 乳酸发酵造成的细胞质酸化降低了细胞质pH值, 从而逐渐抑制乳酸发酵, 促进酒精发酵(Rivoal & Hanson, 1994 ).长时间水淹会改变土壤中的微生物菌丛, 水淹条件利于厌氧微生物的生存发展, 而厌氧微生物的代谢活动会产生对植物有害的毒性物质(如亚硝酸盐), 几乎没有植物能够适应这样的土壤条件(Johnson et al ., 1994 ), 对于经受水淹胁迫的中山杉根系与微生物菌丛间的关系, 以及根系如何适应含有毒性物质的土壤环境, 还有待进一步的研究. ...
Physiology and biochemistry of waterlogging tolerance in plants.
2008
Root responses to flooding.
2
2013
... 水淹胁迫是影响植物生长、生存的主要胁迫.每年在世界各地都会发生不同程度和不同时间的水淹胁迫(Candan & Tarhan, 2012 ), 尤其是水库建设中形成的消落带, 其水淹胁迫发生频率更高, 程度更加严重.消落带每年周期性地遭受不同程度的水淹胁迫, 大部分陆生植物因无法忍受长时间、高强度的反季节水淹逐渐死亡(樊大勇等, 2015 ), 造成消落带植物群落向结构简单化、生物多样性急剧减少的方向发展(Liu & Willison, 2013 ).水淹使土壤中气体交换量减少, 导致根系缺氧, 功能紊乱.同时土壤微生物与根系争夺O2 , 使根系通过线粒体氧化磷酸化途径供能更加困难(Sauter, 2013 ), 进而影响根系的一切耗能活动(水分输送、养分吸收、植物体的糖类代谢以及光合作用和活性氧代谢等).植物在缺氧条件下会通过增加乙醇脱氢酶(ADH)、乳酸脱氢酶(LDH)以及丙酮酸脱羧酶(PDC)活性启动无氧呼吸, 以应对缺氧环境(Zhou et al ., 2012 ). ...
... 植物的耐淹性与PDC、ADH、LDH活性以及乙醇、酒精的积累密切相关(Cuiying et al ., 2010 ), ADH与LDH对于植物适应缺氧胁迫至关重要(Botondi et al ., 2012 ).在缺氧环境下植物从线粒体有氧呼吸转化为以酒精发酵和糖酵解为主的无氧呼吸.丙酮酸作为由无氧呼吸酶催化的乙醇或乳酸发酵的第一产物, PDC是乙醇代谢开始的关键因子.ADH活性与糖酵解途径的运行密切相关(Johnson et al ., 1994 ), 且ADH与乙烯的产生密切相关, 乙烯可以促进植物通气组织的形成, 提高根系对O2 的利用效率(Sairam et al ., 2008 ).对于水淹后ADH是否促进了中山杉根系乙烯的产生, 还有待进一步研究.与水淹后的Kosteletzkya virginica 幼苗根系相似(Zhou et al ., 2012 ), 水淹93天后, 中山杉幼苗根系和叶片的3种无氧呼吸酶活性均随水淹深度的增加显著增加(p < 0.05), 说明经受水淹处理后的幼苗叶片和根系呼吸方式均由以线粒体呼吸为主的有氧呼吸向以酒精发酵为主的无氧呼吸转变.但有氧呼吸仍旧存在, 因为经受水淹的中山杉在根茎结合部长出了细长的气生根, 且茎基部膨大, 这有助于增强体内的气体交换, 获得一定量的O2 (de Oliveira & Joly, 2009 ).同时由图1 看出中山杉根系颜色随水淹深度增加而加深, 表明水淹引起中山杉根系外壁木质化, 将根系与外界复杂的水淹环境隔离, 对根系起到保护作用, 同时防止根系内部O2 的损失(Sauter, 2013 ).与根系相比叶片所遭受的水淹胁迫程度较轻, 水浸处理的幼苗叶片全部出露水面, 浅淹处理的叶片大部分出露于水面之上, 深淹处理的叶片全部在水中, 但不同水淹处理的叶片中3种无氧呼吸酶活性(ADH、PDC、LDH)均高于根系, 间接证明了中山杉幼苗叶片对于缺氧环境更加敏感, 即使叶片没有处于水淹环境, 无氧呼吸同样也会加强.这是因为叶片是碳同化器官, 呼吸代谢相比根系要强, 而且根系产生的脱落酸(ABA)向上运输至叶片, 导致叶片气孔关闭, 进氧阻力加大, 从而使叶片无氧呼吸代谢加强.中山杉根系和叶片的ADH活性均高于LDH, 表明中山杉根系与叶片的呼吸途径均以酒精发酵为主, 乳酸发酵其次.这是因为酒精发酵和乳酸发酵均受细胞质pH值的影响, 乳酸发酵造成的细胞质酸化降低了细胞质pH值, 从而逐渐抑制乳酸发酵, 促进酒精发酵(Rivoal & Hanson, 1994 ).长时间水淹会改变土壤中的微生物菌丛, 水淹条件利于厌氧微生物的生存发展, 而厌氧微生物的代谢活动会产生对植物有害的毒性物质(如亚硝酸盐), 几乎没有植物能够适应这样的土壤条件(Johnson et al ., 1994 ), 对于经受水淹胁迫的中山杉根系与微生物菌丛间的关系, 以及根系如何适应含有毒性物质的土壤环境, 还有待进一步的研究. ...
植物水淹适应与碳水化合物的相关性
4
2010
... 糖类是植物的能量来源, 植物体内的糖类主要包括结构性碳水化合物(主要用于植物形态建成)和非结构性碳水化合物(参与生命代谢), 非结构性碳水化合物(可溶性糖、淀粉等)的代谢在很大程度上决定了植物的生长发育及对环境的适应能力(施美芬等, 2010 ).可溶性糖参与植物体内的糖酵解过程, 为植物提供能量, 淀粉则是主要的能量储存形式.水淹对糖含量的影响较为复杂, 主要取决于植物对水淹的适应策略.植物适应水淹的策略主要包括逃避型和忍耐型.逃避型植物具发达的通气组织, 通过消耗糖类促进茎生长以接近水面, 从而增强植物体内部的气体交换, 获得更多O2 ; 而忍耐型植物通过抑制生长来储存能量(Akman et al ., 2012 ).目前对于植物适应水淹的策略研究大多集中在草本, 对木本植物的研究较少, 且木本植物适应水淹的策略现在还不明确. ...
... 糖类代谢对于植物适应水淹胁迫至关重要, 水淹前后的碳水化合物储量与植物耐淹性密切相关(施美芬等, 2010 ; Ferner et al ., 2012 ), 对水淹敏感的Fagus sylvatica 在经受水淹处理后, 根系可溶性糖和淀粉含量均下降, 而相对耐淹的夏栎(Quercus robur )的根系可溶性糖和淀粉含量则没有发生变化(Ferner et al ., 2012 ), 水淹使紫苜蓿(Medicago sativa ) 根系(Castonguay et al ., 1993 )和黄麻(Corchorus capsularis )根系(Parvin & Karmoker, 2013 )叶片的糖类含量增加.经受93天水淹胁迫的中山杉并没有表现出碳供应不足的现象, 水淹后的中山杉叶片与根系可溶性糖和淀粉含量均随水淹深度的增加显著增加(p < 0.05), 这是因为长期水淹使中山杉根系及叶片无氧呼吸加强, 有氧呼吸减弱, 植株对于糖类的利用效率降低, 因此出现非结构性糖的积累.由于中山杉生长受到抑制, 且出现淀粉及可溶性糖的积累, 所以推断中山杉对于生长季长期水淹的适应策略为忍耐型.很多草本植物在经受生长季短期水淹时, 表现出逃避型的适应策略(施美芬等, 2010 ; Voesenek & Bailey-Serres, 2013 ), 但是对于乔木来说, 其生长周期比草本长, 且生长速率比草本慢, 同时, 需要消耗大量糖类储备的逃避型适应策略对于植物种退水后的恢复生长并没有益处, 反而会使植物出现碳饥饿而威胁其生存, 因此以茎的快速伸长为主要特征的逃避型适应策略在一定程度上并不适合乔木树种, 而忍耐型策略则更加合适, 抑制生长造成的糖类积累为退水后的恢复生长提供了能量储备, 提高了退水后植物的保存率(施美芬等, 2010 ).这也是中山杉耐淹的重要特征.4种不同处理的中山杉根系和叶片淀粉含量均远高于可溶性糖含量, 说明淀粉的储备对于植物适应水淹胁迫发挥着重要的作用. ...
... < 0.05), 这是因为长期水淹使中山杉根系及叶片无氧呼吸加强, 有氧呼吸减弱, 植株对于糖类的利用效率降低, 因此出现非结构性糖的积累.由于中山杉生长受到抑制, 且出现淀粉及可溶性糖的积累, 所以推断中山杉对于生长季长期水淹的适应策略为忍耐型.很多草本植物在经受生长季短期水淹时, 表现出逃避型的适应策略(施美芬等, 2010 ; Voesenek & Bailey-Serres, 2013 ), 但是对于乔木来说, 其生长周期比草本长, 且生长速率比草本慢, 同时, 需要消耗大量糖类储备的逃避型适应策略对于植物种退水后的恢复生长并没有益处, 反而会使植物出现碳饥饿而威胁其生存, 因此以茎的快速伸长为主要特征的逃避型适应策略在一定程度上并不适合乔木树种, 而忍耐型策略则更加合适, 抑制生长造成的糖类积累为退水后的恢复生长提供了能量储备, 提高了退水后植物的保存率(施美芬等, 2010 ).这也是中山杉耐淹的重要特征.4种不同处理的中山杉根系和叶片淀粉含量均远高于可溶性糖含量, 说明淀粉的储备对于植物适应水淹胁迫发挥着重要的作用. ...
... ), 但是对于乔木来说, 其生长周期比草本长, 且生长速率比草本慢, 同时, 需要消耗大量糖类储备的逃避型适应策略对于植物种退水后的恢复生长并没有益处, 反而会使植物出现碳饥饿而威胁其生存, 因此以茎的快速伸长为主要特征的逃避型适应策略在一定程度上并不适合乔木树种, 而忍耐型策略则更加合适, 抑制生长造成的糖类积累为退水后的恢复生长提供了能量储备, 提高了退水后植物的保存率(施美芬等, 2010 ).这也是中山杉耐淹的重要特征.4种不同处理的中山杉根系和叶片淀粉含量均远高于可溶性糖含量, 说明淀粉的储备对于植物适应水淹胁迫发挥着重要的作用. ...
高水位条件下池杉根系的生态适应机制和膝根的呼吸特性
1
2008
... 无氧呼吸供能(每mol葡萄糖产生2 mol ATP)远低于三羧酸循环(每mol葡萄糖产生38 mol ATP), 且水淹胁迫使中山杉根系活力显著下降(p < 0.05), 根系向上输送水分、养分以及矿物质元素的能力显著降低, 因此经受水淹胁迫的中山杉生长明显受到抑制, 这与华建峰等(2011b)以及Visser等(2000)的研究结果相似.中山杉幼苗根质量/地上部分鲜质量的比值随水淹深度的增加呈现增加的趋势(p < 0.05), 这与池杉具有相似的规律, 由此说明中山杉将更多的生物量分配到了根系, 从而有助于提高其耐淹性(唐罗忠等, 2008 ). ...
Flooding tolerance of Carex species in relation to field distribution and aerenchyma formation.
2000
Flooding tolerance: O2 sensing and survival strategies.
1
2013
... 糖类代谢对于植物适应水淹胁迫至关重要, 水淹前后的碳水化合物储量与植物耐淹性密切相关(施美芬等, 2010 ; Ferner et al ., 2012 ), 对水淹敏感的Fagus sylvatica 在经受水淹处理后, 根系可溶性糖和淀粉含量均下降, 而相对耐淹的夏栎(Quercus robur )的根系可溶性糖和淀粉含量则没有发生变化(Ferner et al ., 2012 ), 水淹使紫苜蓿(Medicago sativa ) 根系(Castonguay et al ., 1993 )和黄麻(Corchorus capsularis )根系(Parvin & Karmoker, 2013 )叶片的糖类含量增加.经受93天水淹胁迫的中山杉并没有表现出碳供应不足的现象, 水淹后的中山杉叶片与根系可溶性糖和淀粉含量均随水淹深度的增加显著增加(p < 0.05), 这是因为长期水淹使中山杉根系及叶片无氧呼吸加强, 有氧呼吸减弱, 植株对于糖类的利用效率降低, 因此出现非结构性糖的积累.由于中山杉生长受到抑制, 且出现淀粉及可溶性糖的积累, 所以推断中山杉对于生长季长期水淹的适应策略为忍耐型.很多草本植物在经受生长季短期水淹时, 表现出逃避型的适应策略(施美芬等, 2010 ; Voesenek & Bailey-Serres, 2013 ), 但是对于乔木来说, 其生长周期比草本长, 且生长速率比草本慢, 同时, 需要消耗大量糖类储备的逃避型适应策略对于植物种退水后的恢复生长并没有益处, 反而会使植物出现碳饥饿而威胁其生存, 因此以茎的快速伸长为主要特征的逃避型适应策略在一定程度上并不适合乔木树种, 而忍耐型策略则更加合适, 抑制生长造成的糖类积累为退水后的恢复生长提供了能量储备, 提高了退水后植物的保存率(施美芬等, 2010 ).这也是中山杉耐淹的重要特征.4种不同处理的中山杉根系和叶片淀粉含量均远高于可溶性糖含量, 说明淀粉的储备对于植物适应水淹胁迫发挥着重要的作用. ...
芦芽山不同海拔白杄非结构性碳水化合物含量动态
1
2015
... 低氧环境下非结构性碳水化合物分配到不同器官的模式可能会影响植物的耐淹性(Angelov et al ., 1996 ; 王彪等, 2015 ).植物根系中高淀粉含量是植物耐淹的重要特征(Gravatt & Kirby, 1998 ), 中山杉根系淀粉含量远高于叶片, 虽然根系淀粉含量占总糖含量的百分比随水淹深度的增加呈下降趋势, 但是水淹对其没有显著影响(p > 0.05), 占比均在58%以上, 这是中山杉具有极强耐淹性的重要原因, 同时也表明水淹胁迫并没有严重抑制光合产物向根系的运输.由于根系通常为碳储藏器官, 叶片为碳同化器官, 所以中山杉根系可溶性糖含量明显低于叶片.可溶性糖可以提高组织细胞液渗透压(Molina- Montenegro et al ., 2012 ), 从而有助于叶片通过被动吸水的方式补充水分, 以弥补根系长期处于缺氧环境而无法正常向地上部分输送水分, 这也是中山杉叶片可溶性糖含量高于根系的一个原因. ...
The estimation of carbohydrates in plant extracts by anthrone.
1
1954
... 向离心后剩余的沉淀物中加3 mL蒸馏水, 搅拌均匀, 沸水浴糊化15 min, 冷却后加2 mL冷的9.2 mol·L-1 的高氯酸, 不时搅拌, 提取15 min后加蒸馏水至10 mL, 混匀, 离心10 min, 上清液倾入50 mL容量瓶.再向沉淀中加入2 mL 4.6 mol·L-1 高氯酸, 搅拌提取15 min后加水至10 mL, 混匀后离心10 min, 收集上清液于容量瓶.然后用水洗沉淀1-2次, 离心, 合并离心液于50 mL容量瓶用蒸馏水定容供淀粉含量测定.淀粉和可溶性糖含量采用蒽酮法测定(Yemm & Willis, 1954 ). ...
重庆万州三峡库区消落带中山杉造林实验
1
2014
... 植物的耐淹性及对水淹的生理响应与水淹季节密切相关(Crawford, 2003 ), 本研究初步从呼吸代谢以及物质消耗的角度论证了经受生长季93天水淹后的中山杉幼苗极强耐淹性的原因, 但是由于三峡库区消落带具有“冬水夏陆”的特点, 栽植在消落带内的植物主要遭受非生长季水淹胁迫, 中山杉同样在非生长季表现出了极强的耐淹性(殷云龙等, 2014 ; 张艳婷等, 2015 ), 对于中山杉非生长季的耐淹机理还有待进一步研究.同时, 经受水淹的植物种在退水后的恢复能力也是决定其耐淹能力的重要指标(Liu et al ., 2015 ), 中山杉在退水后的恢复能力同样需要进一步研究. ...
长江三峡库区消落带中山杉耐淹试验
1
2015
... 植物的耐淹性及对水淹的生理响应与水淹季节密切相关(Crawford, 2003 ), 本研究初步从呼吸代谢以及物质消耗的角度论证了经受生长季93天水淹后的中山杉幼苗极强耐淹性的原因, 但是由于三峡库区消落带具有“冬水夏陆”的特点, 栽植在消落带内的植物主要遭受非生长季水淹胁迫, 中山杉同样在非生长季表现出了极强的耐淹性(殷云龙等, 2014 ; 张艳婷等, 2015 ), 对于中山杉非生长季的耐淹机理还有待进一步研究.同时, 经受水淹的植物种在退水后的恢复能力也是决定其耐淹能力的重要指标(Liu et al ., 2015 ), 中山杉在退水后的恢复能力同样需要进一步研究. ...
Respiratory enzyme activity and regulation of respiration pathway in seashore mallow (Kosteletzkya virginica) seedlings under waterlogging conditions.
2
2012
... 水淹胁迫是影响植物生长、生存的主要胁迫.每年在世界各地都会发生不同程度和不同时间的水淹胁迫(Candan & Tarhan, 2012 ), 尤其是水库建设中形成的消落带, 其水淹胁迫发生频率更高, 程度更加严重.消落带每年周期性地遭受不同程度的水淹胁迫, 大部分陆生植物因无法忍受长时间、高强度的反季节水淹逐渐死亡(樊大勇等, 2015 ), 造成消落带植物群落向结构简单化、生物多样性急剧减少的方向发展(Liu & Willison, 2013 ).水淹使土壤中气体交换量减少, 导致根系缺氧, 功能紊乱.同时土壤微生物与根系争夺O2 , 使根系通过线粒体氧化磷酸化途径供能更加困难(Sauter, 2013 ), 进而影响根系的一切耗能活动(水分输送、养分吸收、植物体的糖类代谢以及光合作用和活性氧代谢等).植物在缺氧条件下会通过增加乙醇脱氢酶(ADH)、乳酸脱氢酶(LDH)以及丙酮酸脱羧酶(PDC)活性启动无氧呼吸, 以应对缺氧环境(Zhou et al ., 2012 ). ...
... 植物的耐淹性与PDC、ADH、LDH活性以及乙醇、酒精的积累密切相关(Cuiying et al ., 2010 ), ADH与LDH对于植物适应缺氧胁迫至关重要(Botondi et al ., 2012 ).在缺氧环境下植物从线粒体有氧呼吸转化为以酒精发酵和糖酵解为主的无氧呼吸.丙酮酸作为由无氧呼吸酶催化的乙醇或乳酸发酵的第一产物, PDC是乙醇代谢开始的关键因子.ADH活性与糖酵解途径的运行密切相关(Johnson et al ., 1994 ), 且ADH与乙烯的产生密切相关, 乙烯可以促进植物通气组织的形成, 提高根系对O2 的利用效率(Sairam et al ., 2008 ).对于水淹后ADH是否促进了中山杉根系乙烯的产生, 还有待进一步研究.与水淹后的Kosteletzkya virginica 幼苗根系相似(Zhou et al ., 2012 ), 水淹93天后, 中山杉幼苗根系和叶片的3种无氧呼吸酶活性均随水淹深度的增加显著增加(p < 0.05), 说明经受水淹处理后的幼苗叶片和根系呼吸方式均由以线粒体呼吸为主的有氧呼吸向以酒精发酵为主的无氧呼吸转变.但有氧呼吸仍旧存在, 因为经受水淹的中山杉在根茎结合部长出了细长的气生根, 且茎基部膨大, 这有助于增强体内的气体交换, 获得一定量的O2 (de Oliveira & Joly, 2009 ).同时由图1 看出中山杉根系颜色随水淹深度增加而加深, 表明水淹引起中山杉根系外壁木质化, 将根系与外界复杂的水淹环境隔离, 对根系起到保护作用, 同时防止根系内部O2 的损失(Sauter, 2013 ).与根系相比叶片所遭受的水淹胁迫程度较轻, 水浸处理的幼苗叶片全部出露水面, 浅淹处理的叶片大部分出露于水面之上, 深淹处理的叶片全部在水中, 但不同水淹处理的叶片中3种无氧呼吸酶活性(ADH、PDC、LDH)均高于根系, 间接证明了中山杉幼苗叶片对于缺氧环境更加敏感, 即使叶片没有处于水淹环境, 无氧呼吸同样也会加强.这是因为叶片是碳同化器官, 呼吸代谢相比根系要强, 而且根系产生的脱落酸(ABA)向上运输至叶片, 导致叶片气孔关闭, 进氧阻力加大, 从而使叶片无氧呼吸代谢加强.中山杉根系和叶片的ADH活性均高于LDH, 表明中山杉根系与叶片的呼吸途径均以酒精发酵为主, 乳酸发酵其次.这是因为酒精发酵和乳酸发酵均受细胞质pH值的影响, 乳酸发酵造成的细胞质酸化降低了细胞质pH值, 从而逐渐抑制乳酸发酵, 促进酒精发酵(Rivoal & Hanson, 1994 ).长时间水淹会改变土壤中的微生物菌丛, 水淹条件利于厌氧微生物的生存发展, 而厌氧微生物的代谢活动会产生对植物有害的毒性物质(如亚硝酸盐), 几乎没有植物能够适应这样的土壤条件(Johnson et al ., 1994 ), 对于经受水淹胁迫的中山杉根系与微生物菌丛间的关系, 以及根系如何适应含有毒性物质的土壤环境, 还有待进一步的研究. ...
 , 王建修
, 王建修

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}