Aims Our objective was to investigate the responses of maize photosynthesis and growth to repeated drought.Methods Maize seedlings were exposed to different soil water deficit for three weeks, then rewatering for one week, and again to different water deficit for three weeks, to examine the effects of repeated drought on photosynthesis and growth.Important findings After the first water deficit treatments, under severe drought, plant height, total leaf area of individual plant, shoot and root biomass declined significantly, also transpiration rate (Tr), stomatal conductance (Gs), intercellular CO2 concentration (Ci), net photosynthetic rate (Pn), maximum net photosynthetic rate (Amax), but light compensation point and dark respiration rate increased significantly. Under medium drought, plant height, leaf area, and shoot biomass decreased significantly, but root biomass did not vary, hence, the ratio of roots to shoots (R/S) increased. Moreover, plants did not show significant differences in photosynthetic parameters. After rewatering, photosynthesis and growth rate of plants previously exposed to water deficit could recover to the levels of well-watered plants, but plant height and leaf area did not recover to the levels of the control. When maize were subjected to recurrent drought, plants pre-exposed to medium drought showed no significant difference in plant height, biomass, and photosynthetic parameters, but a significant decrease in leaf area, compared to plants only exposed to second medium drought. Plants pre-exposed to severe drought had significantly higher Tr, Gs, Ci, Pn, Amax, and, apparent quantum yield but significantly lower plant height, leaf area, and biomass than plants without previous exposure. These results indicated that the first severe drought significantly reduced photosynthetic capacity and maize growth, rewatering could recover photosynthesis and growth rate to the levels of well-watered plants, but could not eliminate the adverse influence of the first drought on growth. The first medium drought could stimulate the growth of maize root system and significantly increased R/S, which can enhance maize drought resistance to subsequent repeated drought, and maintain the total biomass in the control level; the first severe drought could enhance maize drought resistance to subsequent repeated drought in the aspect of photosynthesis, but could not compensate for the adverse effect of early drought on plant growth. Hence, in practice, drought hardening should be limited in the level of medium drought, and avoiding severe drought.
ZHAOWen-Sai, SUNYong-Lin, LIUXi-Ping. Effects of drought-rewatering-drought on photosynthesis and growth of maize[J]. Chinese Journal of Plant Ecology, 2016, 40(6): 594-603 https://doi.org/10.17521/ cjpe.2015.0345
试验地点设在陕西省杨凌西北农林科技大学旱区节水研究院内(108.07° E, 34.28° N, 海拔516 m)。该地区年平均气温12.9 ℃, 极端最高气温42 ℃, 极端最低气温-19.4 ℃, 年日照时数2 150 h, 年降水量632 mm, 年蒸发量1 500 mm, 属大陆性暖温带季风气候。
Fig. 1 Effects of first water treatments on diurnal variations of transpiration rate (Tr) (A), stomatal conductance (Gs) (B), intercellular CO2 concentration (Ci) (C), and net photosynthetic rate (Pn) (D) of maize leaves (mean ± SD).
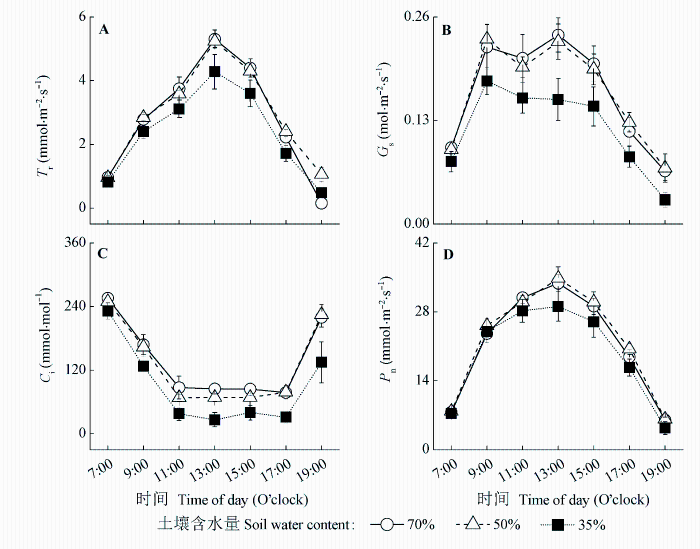
Fig. 2 Effects of rewatering on diurnal variations of transpiration rate (Tr) (A), stomatal conductance (Gs) (B), intercellular CO2 concentration (Ci) (C), and net photosynthetic rate (Pn) (D) of maize leaves (mean ± SD).
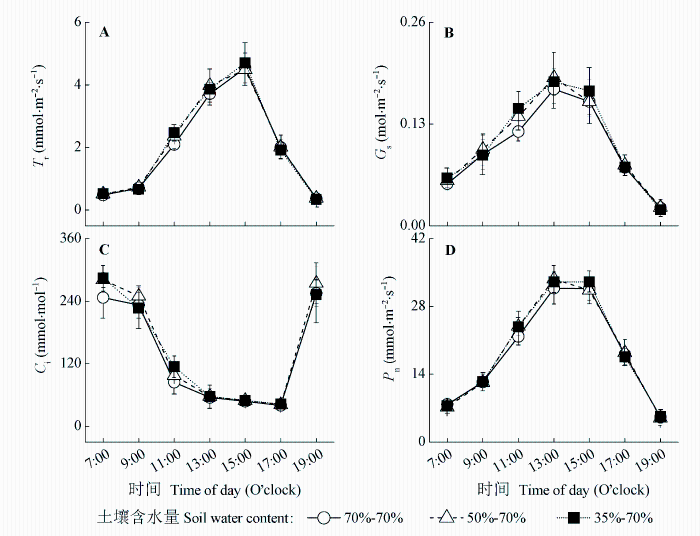
Fig. 3 Effects of the second water treatments on diurnal variations of transpiration rate (Tr) (A, B), stomatal conductance (Gs) (C, D), intercellular CO2 concentration (Ci) (E, F), and net photosynthetic rate (Pn) (G, H) of maize leaves (mean ± SD).
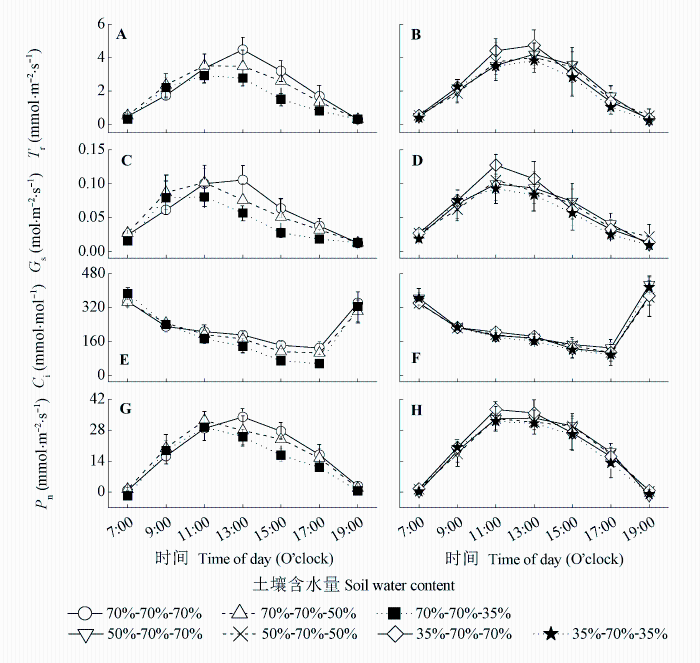
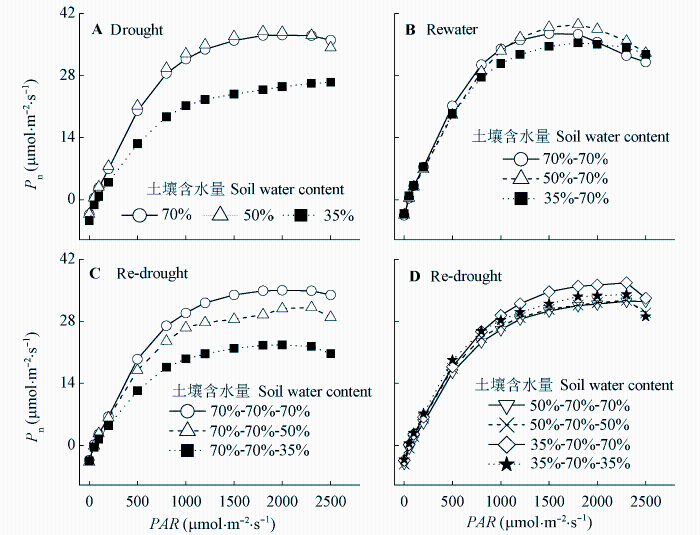
Fig. 4 Effects of drought-rewatering-drought treatments on response curves of net photosynthetic rate (Pn) to photosynthetically active radiation (PAR) in maize leaves.
Table 2 Effects of drought-rewatering-drought treatments on characteristic parameters of the response curves of net photosynthetic rate (Pn) to photosynthetically active radiation (PAR) in maize leaves (mean ± SD)
处理 Treatment
最大净光合速率 Amax (μmol·m-2·s-1)
光补偿点 LCP (μmol·m-2·s-1)
光饱和点 LSP (μmol·m-2·s-1)
暗呼吸速率 Rd (μmol·m-2·s-1)
表观量子效率 AQY (μmol·m-2·s-1)
第一次水分处理 First water treatment
70%
41.39 ± 0.88b
53.97 ± 15.17a
892.50 ± 74.93a
2.58 ± 0.20a
0.050 ± 0.002ab
50%
41.01 ± 3.54b
50.87 ± 6.60a
832.98 ± 65.23a
2.48 ± 0.68a
0.051 ± 0.005b
35%
33.43 ± 3.57a
95.09 ± 8.79b
906.90 ± 66.14a
3.92 ± 0.17b
0.044 ± 0.002a
复水 Rewatering
70%-70%
37.79 ± 4.55a
52.95 ± 7.99a
770.05 ± 95.33a
2.30 ± 0.41a
0.047 ± 0.002a
50%-70%
39.82 ± 4.64a
53.96 ± 3.16a
859.91 ± 104.84a
1.88 ± 0.38a
0.045 ± 0.003a
35%-70%
37.88 ± 5.05a
46.89 ± 1.32a
796.46 ± 97.11a
1.99 ± 0.33a
0.046 ± 0.004a
第二次水分处理 Second water treatments
70%-70%-70%
39.95 ± 2.03c
57.46 ± 13.82ab
893.05 ± 75.07ab
2.78 ± 0.84a
0.050 ± 0.003c
70%-70%-50%
35.11 ± 2.31b
61.78 ± 13.99ab
780.32 ± 37.03a
2.87 ± 0.99a
0.048 ± 0.005bc
70%-70%-35%
26.00 ± 0.95a
76.51 ± 14.81b
752.90 ± 105.37a
2.63 ± 0.32a
0.035 ± 0.004a
50%-70%-70%
39.64 ± 2.31c
77.17 ± 13.24b
972.55 ± 118.34b
3.50 ± 0.63a
0.048 ± 0.003bc
50%-70%-50%
37.56 ± 1.68bc
71.32 ± 11.07ab
815.10 ± 50.04ab
3.43 ± 0.65a
0.049 ± 0.001bc
35%-70%-70%
40.05 ± 0.76c
57.89 ± 0.31ab
901.57 ± 40.73ab
2.36 ± 0.36a
0.043 ± 0.006b
35%-70%-35%
36.65 ± 2.94bc
52.57 ± 8.62a
789.16 ± 143.48a
2.75 ± 0.32a
0.051 ± 0.004c
Different lowercase letters indicate significant differences between different treatments (p < 0.05). Amax, maximum net photosynthetic rate; AQY, apparent quantum yield; LCP, light compensation point; LSP, light saturation point; Rd, dark respiration.不同小写字母表示处理间差异显著(p < 0.05)。
Effects of rewatering after drought stress on photosynthesis and yield during flowering and boll-setting stage of cotton under-mulch-drip irrigation in Xinjiang.
Acta Agronomica Sinica, 34, 171-174. (in Chinese with English abstract)[罗宏海, 张亚黎, 张旺锋, 白慧东, 何在菊, 杜明伟, 张宏芝 (2008).
Involvement of antioxidant defense system in enhancement of drought re-sistance in tobacco (Nicotiana tabacum L.) plants through circular drought-hardening.
Plant Physiology Journal, 48, 705-713. (in Chinese with English abstract)[荣智媛, 张晓海, 杨双龙, 徐照丽, 李军营, 黄国宾, 赵静, 龚明 (2012).
Root water uptake and transport: Using physiological processes in global predictions.
1
2000
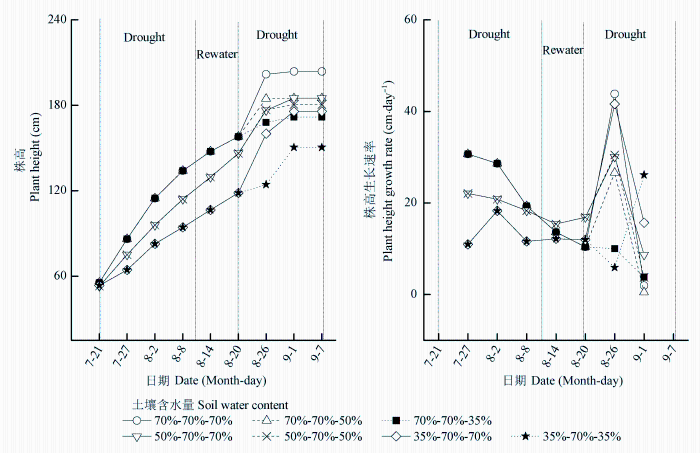
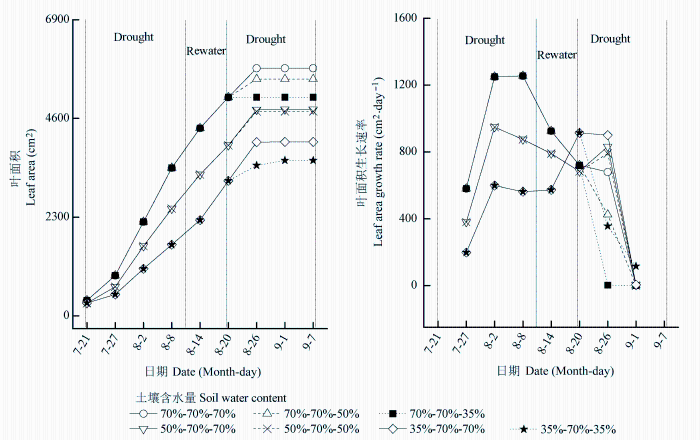
... (Tsuji et al., 2003; Yao et al., 2009; Shen et al., 2015).干旱胁迫对玉米的生长发育有抑制作用, 并能引起玉米最终产量的降低(张仁和等, 2011; 姜鹏等, 2013; Farhangfar et al., 2015).本研究的结果也显示, 第一次重度干旱显著降低玉米叶片的Tr、Gs、Ci、Pn、Amax, 并使LCP和Rd显著升高, 说明玉米通过减小气孔开度降低了蒸腾水分散失, 同时, 光合能力下降, 光合产物的消耗增加, 这不利于有机物质的积累.遭受中度和重度干旱处理的玉米植株的株高和叶面积均显著降低, 生物量的积累随干旱程度的加剧呈显著降低趋势, 而根冠比呈显著升高趋势, 表明水分亏缺处理下生物量优先向根分配, 有利于植物吸收土壤中的水分, 以更好地适应干旱环境(Jackson et al., 2000).虽然中度和重度干旱处理均引起根冠比的增加, 但不同的是, 中度干旱下, 地上部分生物量降低, 但根系生长没有受到影响, 且这种根冠比的增加一直延续到复水和二次干旱处理之后; 而重度干旱下, 地上部分和根系生物量均降低, 且根系生物量的降低幅度更大.干旱处理后, 复水能使经历过中度或重度干旱处理的玉米植株的光合能力和生长速率恢复到甚至超过正常水分处理植株的水平, 这与张林春等(2010)对玉米的研究结果是一致的, 但株高和叶面积依然低于正常水分处理的植株, 表明复水并不能完全弥补之前干旱对玉米生长造成的影响.第二次水分处理后, 与只遭受第二次中度或重度干旱处理的植株相比, 经历过第一次中度干旱处理的植株在遭受第二次中度干旱后, 其株高和生物量没有受到明显影响, 说明第一次中度干旱处理可以缓解第二次中度干旱对玉米生长的影响; 经历过第一次重度干旱处理的植株在遭受第二次重度干旱后, Tr、Gs、Ci、Pn、Amax、AQY和叶面积生长速率显著升高, 而株高、叶面积和生物量则显著降低, 说明第一次重度干旱处理在光合上能提高植株对二次干旱的抵抗能力, 但不能弥补前期干旱处理对生长产生的不利影响. ...
不同时期干旱对玉米生长发育及产量的影响
1
2013
... (Tsuji et al., 2003; Yao et al., 2009; Shen et al., 2015).干旱胁迫对玉米的生长发育有抑制作用, 并能引起玉米最终产量的降低(张仁和等, 2011; 姜鹏等, 2013; Farhangfar et al., 2015).本研究的结果也显示, 第一次重度干旱显著降低玉米叶片的Tr、Gs、Ci、Pn、Amax, 并使LCP和Rd显著升高, 说明玉米通过减小气孔开度降低了蒸腾水分散失, 同时, 光合能力下降, 光合产物的消耗增加, 这不利于有机物质的积累.遭受中度和重度干旱处理的玉米植株的株高和叶面积均显著降低, 生物量的积累随干旱程度的加剧呈显著降低趋势, 而根冠比呈显著升高趋势, 表明水分亏缺处理下生物量优先向根分配, 有利于植物吸收土壤中的水分, 以更好地适应干旱环境(Jackson et al., 2000).虽然中度和重度干旱处理均引起根冠比的增加, 但不同的是, 中度干旱下, 地上部分生物量降低, 但根系生长没有受到影响, 且这种根冠比的增加一直延续到复水和二次干旱处理之后; 而重度干旱下, 地上部分和根系生物量均降低, 且根系生物量的降低幅度更大.干旱处理后, 复水能使经历过中度或重度干旱处理的玉米植株的光合能力和生长速率恢复到甚至超过正常水分处理植株的水平, 这与张林春等(2010)对玉米的研究结果是一致的, 但株高和叶面积依然低于正常水分处理的植株, 表明复水并不能完全弥补之前干旱对玉米生长造成的影响.第二次水分处理后, 与只遭受第二次中度或重度干旱处理的植株相比, 经历过第一次中度干旱处理的植株在遭受第二次中度干旱后, 其株高和生物量没有受到明显影响, 说明第一次中度干旱处理可以缓解第二次中度干旱对玉米生长的影响; 经历过第一次重度干旱处理的植株在遭受第二次重度干旱后, Tr、Gs、Ci、Pn、Amax、AQY和叶面积生长速率显著升高, 而株高、叶面积和生物量则显著降低, 说明第一次重度干旱处理在光合上能提高植株对二次干旱的抵抗能力, 但不能弥补前期干旱处理对生长产生的不利影响. ...
苗期干旱及复水条件下不同花生品种的光合特性,
1
2014
... 当植物遭受干旱胁迫后, 体内的水分平衡被破坏, 并引起气孔关闭和光合速率下降.随着干旱的加剧, 植物体内会产生和积累大量的活性氧, 使得叶绿体结构被破坏, 光合作用的相关酶失活或变性, 并最终抑制植物的生长(Galmés et al., 2007; McDowell et al., 2008; Chaves et al., 2009).旱后复水能够使植物的生理功能得到恢复, 可在一定程度上弥补干旱对植物造成的伤害(山仑, 2003), 提高植物的光合速率并使其生长加速(Fortunati et al., 2008; 罗宏海等, 2008; Aidar et al., 2014; 厉广辉等, 2014).不过, 干旱后复水对植物生长上的补偿往往是有限的, 植株生长的恢复程度可能与复水前干旱的胁迫程度和持续时间有关(刘晓英等, 2001; Xu & Zhou, 2007; Xu et al., 2009).当经历过水分胁迫及复水的植物再次遇到干旱时, 植物对干旱适应性的增强可能与耐受基因的转录、渗透调节和抗氧化能力有关, 从而可在生理、生化和分子水平上具有更好地适应和抵抗干旱的能力(Ruiz-Sánchez et al., 2000; Villar-Salvador et al., 2004; Bruce et al., 2007; 徐芬芬等, 2009; Luo et al., 2011; Ding et al., 2012; 荣智媛等, 2012; 徐超华等, 2012).不过, 也有研究显示, 经历过干旱胁迫的植物再次遇到干旱时并没有提高自身抵御干旱的能力(Lloret et al., 2004; Zavalloni et al., 2008; Walter et al., 2011).因此, 在全球气候变化导致的干旱频率增加的背景下, 深入研究植物对多次干旱的响应机制, 在理论和实践上都具有重要 意义. ...
水分胁迫后复水对冬小麦叶面积的激发作用
1
2001
... 当植物遭受干旱胁迫后, 体内的水分平衡被破坏, 并引起气孔关闭和光合速率下降.随着干旱的加剧, 植物体内会产生和积累大量的活性氧, 使得叶绿体结构被破坏, 光合作用的相关酶失活或变性, 并最终抑制植物的生长(Galmés et al., 2007; McDowell et al., 2008; Chaves et al., 2009).旱后复水能够使植物的生理功能得到恢复, 可在一定程度上弥补干旱对植物造成的伤害(山仑, 2003), 提高植物的光合速率并使其生长加速(Fortunati et al., 2008; 罗宏海等, 2008; Aidar et al., 2014; 厉广辉等, 2014).不过, 干旱后复水对植物生长上的补偿往往是有限的, 植株生长的恢复程度可能与复水前干旱的胁迫程度和持续时间有关(刘晓英等, 2001; Xu & Zhou, 2007; Xu et al., 2009).当经历过水分胁迫及复水的植物再次遇到干旱时, 植物对干旱适应性的增强可能与耐受基因的转录、渗透调节和抗氧化能力有关, 从而可在生理、生化和分子水平上具有更好地适应和抵抗干旱的能力(Ruiz-Sánchez et al., 2000; Villar-Salvador et al., 2004; Bruce et al., 2007; 徐芬芬等, 2009; Luo et al., 2011; Ding et al., 2012; 荣智媛等, 2012; 徐超华等, 2012).不过, 也有研究显示, 经历过干旱胁迫的植物再次遇到干旱时并没有提高自身抵御干旱的能力(Lloret et al., 2004; Zavalloni et al., 2008; Walter et al., 2011).因此, 在全球气候变化导致的干旱频率增加的背景下, 深入研究植物对多次干旱的响应机制, 在理论和实践上都具有重要 意义. ...
Canopy recovery after drought dieback in holm-oak Mediterranean forests of Catalonia (NE Spain).
1
2004
... 当植物遭受干旱胁迫后, 体内的水分平衡被破坏, 并引起气孔关闭和光合速率下降.随着干旱的加剧, 植物体内会产生和积累大量的活性氧, 使得叶绿体结构被破坏, 光合作用的相关酶失活或变性, 并最终抑制植物的生长(Galmés et al., 2007; McDowell et al., 2008; Chaves et al., 2009).旱后复水能够使植物的生理功能得到恢复, 可在一定程度上弥补干旱对植物造成的伤害(山仑, 2003), 提高植物的光合速率并使其生长加速(Fortunati et al., 2008; 罗宏海等, 2008; Aidar et al., 2014; 厉广辉等, 2014).不过, 干旱后复水对植物生长上的补偿往往是有限的, 植株生长的恢复程度可能与复水前干旱的胁迫程度和持续时间有关(刘晓英等, 2001; Xu & Zhou, 2007; Xu et al., 2009).当经历过水分胁迫及复水的植物再次遇到干旱时, 植物对干旱适应性的增强可能与耐受基因的转录、渗透调节和抗氧化能力有关, 从而可在生理、生化和分子水平上具有更好地适应和抵抗干旱的能力(Ruiz-Sánchez et al., 2000; Villar-Salvador et al., 2004; Bruce et al., 2007; 徐芬芬等, 2009; Luo et al., 2011; Ding et al., 2012; 荣智媛等, 2012; 徐超华等, 2012).不过, 也有研究显示, 经历过干旱胁迫的植物再次遇到干旱时并没有提高自身抵御干旱的能力(Lloret et al., 2004; Zavalloni et al., 2008; Walter et al., 2011).因此, 在全球气候变化导致的干旱频率增加的背景下, 深入研究植物对多次干旱的响应机制, 在理论和实践上都具有重要 意义. ...
新疆滴灌棉花花铃期干旱复水对叶片光合特性及产量的影响
1
2008
... 当植物遭受干旱胁迫后, 体内的水分平衡被破坏, 并引起气孔关闭和光合速率下降.随着干旱的加剧, 植物体内会产生和积累大量的活性氧, 使得叶绿体结构被破坏, 光合作用的相关酶失活或变性, 并最终抑制植物的生长(Galmés et al., 2007; McDowell et al., 2008; Chaves et al., 2009).旱后复水能够使植物的生理功能得到恢复, 可在一定程度上弥补干旱对植物造成的伤害(山仑, 2003), 提高植物的光合速率并使其生长加速(Fortunati et al., 2008; 罗宏海等, 2008; Aidar et al., 2014; 厉广辉等, 2014).不过, 干旱后复水对植物生长上的补偿往往是有限的, 植株生长的恢复程度可能与复水前干旱的胁迫程度和持续时间有关(刘晓英等, 2001; Xu & Zhou, 2007; Xu et al., 2009).当经历过水分胁迫及复水的植物再次遇到干旱时, 植物对干旱适应性的增强可能与耐受基因的转录、渗透调节和抗氧化能力有关, 从而可在生理、生化和分子水平上具有更好地适应和抵抗干旱的能力(Ruiz-Sánchez et al., 2000; Villar-Salvador et al., 2004; Bruce et al., 2007; 徐芬芬等, 2009; Luo et al., 2011; Ding et al., 2012; 荣智媛等, 2012; 徐超华等, 2012).不过, 也有研究显示, 经历过干旱胁迫的植物再次遇到干旱时并没有提高自身抵御干旱的能力(Lloret et al., 2004; Zavalloni et al., 2008; Walter et al., 2011).因此, 在全球气候变化导致的干旱频率增加的背景下, 深入研究植物对多次干旱的响应机制, 在理论和实践上都具有重要 意义. ...
Physiological acclimation of two psammophytes to repeated soil drought and rewatering.
1
2011
... 当植物遭受干旱胁迫后, 体内的水分平衡被破坏, 并引起气孔关闭和光合速率下降.随着干旱的加剧, 植物体内会产生和积累大量的活性氧, 使得叶绿体结构被破坏, 光合作用的相关酶失活或变性, 并最终抑制植物的生长(Galmés et al., 2007; McDowell et al., 2008; Chaves et al., 2009).旱后复水能够使植物的生理功能得到恢复, 可在一定程度上弥补干旱对植物造成的伤害(山仑, 2003), 提高植物的光合速率并使其生长加速(Fortunati et al., 2008; 罗宏海等, 2008; Aidar et al., 2014; 厉广辉等, 2014).不过, 干旱后复水对植物生长上的补偿往往是有限的, 植株生长的恢复程度可能与复水前干旱的胁迫程度和持续时间有关(刘晓英等, 2001; Xu & Zhou, 2007; Xu et al., 2009).当经历过水分胁迫及复水的植物再次遇到干旱时, 植物对干旱适应性的增强可能与耐受基因的转录、渗透调节和抗氧化能力有关, 从而可在生理、生化和分子水平上具有更好地适应和抵抗干旱的能力(Ruiz-Sánchez et al., 2000; Villar-Salvador et al., 2004; Bruce et al., 2007; 徐芬芬等, 2009; Luo et al., 2011; Ding et al., 2012; 荣智媛等, 2012; 徐超华等, 2012).不过, 也有研究显示, 经历过干旱胁迫的植物再次遇到干旱时并没有提高自身抵御干旱的能力(Lloret et al., 2004; Zavalloni et al., 2008; Walter et al., 2011).因此, 在全球气候变化导致的干旱频率增加的背景下, 深入研究植物对多次干旱的响应机制, 在理论和实践上都具有重要 意义. ...
Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought?
1
2008
... 当植物遭受干旱胁迫后, 体内的水分平衡被破坏, 并引起气孔关闭和光合速率下降.随着干旱的加剧, 植物体内会产生和积累大量的活性氧, 使得叶绿体结构被破坏, 光合作用的相关酶失活或变性, 并最终抑制植物的生长(Galmés et al., 2007; McDowell et al., 2008; Chaves et al., 2009).旱后复水能够使植物的生理功能得到恢复, 可在一定程度上弥补干旱对植物造成的伤害(山仑, 2003), 提高植物的光合速率并使其生长加速(Fortunati et al., 2008; 罗宏海等, 2008; Aidar et al., 2014; 厉广辉等, 2014).不过, 干旱后复水对植物生长上的补偿往往是有限的, 植株生长的恢复程度可能与复水前干旱的胁迫程度和持续时间有关(刘晓英等, 2001; Xu & Zhou, 2007; Xu et al., 2009).当经历过水分胁迫及复水的植物再次遇到干旱时, 植物对干旱适应性的增强可能与耐受基因的转录、渗透调节和抗氧化能力有关, 从而可在生理、生化和分子水平上具有更好地适应和抵抗干旱的能力(Ruiz-Sánchez et al., 2000; Villar-Salvador et al., 2004; Bruce et al., 2007; 徐芬芬等, 2009; Luo et al., 2011; Ding et al., 2012; 荣智媛等, 2012; 徐超华等, 2012).不过, 也有研究显示, 经历过干旱胁迫的植物再次遇到干旱时并没有提高自身抵御干旱的能力(Lloret et al., 2004; Zavalloni et al., 2008; Walter et al., 2011).因此, 在全球气候变化导致的干旱频率增加的背景下, 深入研究植物对多次干旱的响应机制, 在理论和实践上都具有重要 意义. ...
抗氧化系统参与循环干旱锻炼提高烟草植株抗旱性的形成
1
2012
... 当植物遭受干旱胁迫后, 体内的水分平衡被破坏, 并引起气孔关闭和光合速率下降.随着干旱的加剧, 植物体内会产生和积累大量的活性氧, 使得叶绿体结构被破坏, 光合作用的相关酶失活或变性, 并最终抑制植物的生长(Galmés et al., 2007; McDowell et al., 2008; Chaves et al., 2009).旱后复水能够使植物的生理功能得到恢复, 可在一定程度上弥补干旱对植物造成的伤害(山仑, 2003), 提高植物的光合速率并使其生长加速(Fortunati et al., 2008; 罗宏海等, 2008; Aidar et al., 2014; 厉广辉等, 2014).不过, 干旱后复水对植物生长上的补偿往往是有限的, 植株生长的恢复程度可能与复水前干旱的胁迫程度和持续时间有关(刘晓英等, 2001; Xu & Zhou, 2007; Xu et al., 2009).当经历过水分胁迫及复水的植物再次遇到干旱时, 植物对干旱适应性的增强可能与耐受基因的转录、渗透调节和抗氧化能力有关, 从而可在生理、生化和分子水平上具有更好地适应和抵抗干旱的能力(Ruiz-Sánchez et al., 2000; Villar-Salvador et al., 2004; Bruce et al., 2007; 徐芬芬等, 2009; Luo et al., 2011; Ding et al., 2012; 荣智媛等, 2012; 徐超华等, 2012).不过, 也有研究显示, 经历过干旱胁迫的植物再次遇到干旱时并没有提高自身抵御干旱的能力(Lloret et al., 2004; Zavalloni et al., 2008; Walter et al., 2011).因此, 在全球气候变化导致的干旱频率增加的背景下, 深入研究植物对多次干旱的响应机制, 在理论和实践上都具有重要 意义. ...
Water stress preconditioning to improve drought resistance in young apricot plants.
1
2000
... 当植物遭受干旱胁迫后, 体内的水分平衡被破坏, 并引起气孔关闭和光合速率下降.随着干旱的加剧, 植物体内会产生和积累大量的活性氧, 使得叶绿体结构被破坏, 光合作用的相关酶失活或变性, 并最终抑制植物的生长(Galmés et al., 2007; McDowell et al., 2008; Chaves et al., 2009).旱后复水能够使植物的生理功能得到恢复, 可在一定程度上弥补干旱对植物造成的伤害(山仑, 2003), 提高植物的光合速率并使其生长加速(Fortunati et al., 2008; 罗宏海等, 2008; Aidar et al., 2014; 厉广辉等, 2014).不过, 干旱后复水对植物生长上的补偿往往是有限的, 植株生长的恢复程度可能与复水前干旱的胁迫程度和持续时间有关(刘晓英等, 2001; Xu & Zhou, 2007; Xu et al., 2009).当经历过水分胁迫及复水的植物再次遇到干旱时, 植物对干旱适应性的增强可能与耐受基因的转录、渗透调节和抗氧化能力有关, 从而可在生理、生化和分子水平上具有更好地适应和抵抗干旱的能力(Ruiz-Sánchez et al., 2000; Villar-Salvador et al., 2004; Bruce et al., 2007; 徐芬芬等, 2009; Luo et al., 2011; Ding et al., 2012; 荣智媛等, 2012; 徐超华等, 2012).不过, 也有研究显示, 经历过干旱胁迫的植物再次遇到干旱时并没有提高自身抵御干旱的能力(Lloret et al., 2004; Zavalloni et al., 2008; Walter et al., 2011).因此, 在全球气候变化导致的干旱频率增加的背景下, 深入研究植物对多次干旱的响应机制, 在理论和实践上都具有重要 意义. ...
节水农业与作物高效用水
1
2003
... 当植物遭受干旱胁迫后, 体内的水分平衡被破坏, 并引起气孔关闭和光合速率下降.随着干旱的加剧, 植物体内会产生和积累大量的活性氧, 使得叶绿体结构被破坏, 光合作用的相关酶失活或变性, 并最终抑制植物的生长(Galmés et al., 2007; McDowell et al., 2008; Chaves et al., 2009).旱后复水能够使植物的生理功能得到恢复, 可在一定程度上弥补干旱对植物造成的伤害(山仑, 2003), 提高植物的光合速率并使其生长加速(Fortunati et al., 2008; 罗宏海等, 2008; Aidar et al., 2014; 厉广辉等, 2014).不过, 干旱后复水对植物生长上的补偿往往是有限的, 植株生长的恢复程度可能与复水前干旱的胁迫程度和持续时间有关(刘晓英等, 2001; Xu & Zhou, 2007; Xu et al., 2009).当经历过水分胁迫及复水的植物再次遇到干旱时, 植物对干旱适应性的增强可能与耐受基因的转录、渗透调节和抗氧化能力有关, 从而可在生理、生化和分子水平上具有更好地适应和抵抗干旱的能力(Ruiz-Sánchez et al., 2000; Villar-Salvador et al., 2004; Bruce et al., 2007; 徐芬芬等, 2009; Luo et al., 2011; Ding et al., 2012; 荣智媛等, 2012; 徐超华等, 2012).不过, 也有研究显示, 经历过干旱胁迫的植物再次遇到干旱时并没有提高自身抵御干旱的能力(Lloret et al., 2004; Zavalloni et al., 2008; Walter et al., 2011).因此, 在全球气候变化导致的干旱频率增加的背景下, 深入研究植物对多次干旱的响应机制, 在理论和实践上都具有重要 意义. ...
Drought and UV-B radiation effect on photosynthesis and antioxidant parameters in soybean and maize.
1
2015
... (Tsuji et al., 2003; Yao et al., 2009; Shen et al., 2015).干旱胁迫对玉米的生长发育有抑制作用, 并能引起玉米最终产量的降低(张仁和等, 2011; 姜鹏等, 2013; Farhangfar et al., 2015).本研究的结果也显示, 第一次重度干旱显著降低玉米叶片的Tr、Gs、Ci、Pn、Amax, 并使LCP和Rd显著升高, 说明玉米通过减小气孔开度降低了蒸腾水分散失, 同时, 光合能力下降, 光合产物的消耗增加, 这不利于有机物质的积累.遭受中度和重度干旱处理的玉米植株的株高和叶面积均显著降低, 生物量的积累随干旱程度的加剧呈显著降低趋势, 而根冠比呈显著升高趋势, 表明水分亏缺处理下生物量优先向根分配, 有利于植物吸收土壤中的水分, 以更好地适应干旱环境(Jackson et al., 2000).虽然中度和重度干旱处理均引起根冠比的增加, 但不同的是, 中度干旱下, 地上部分生物量降低, 但根系生长没有受到影响, 且这种根冠比的增加一直延续到复水和二次干旱处理之后; 而重度干旱下, 地上部分和根系生物量均降低, 且根系生物量的降低幅度更大.干旱处理后, 复水能使经历过中度或重度干旱处理的玉米植株的光合能力和生长速率恢复到甚至超过正常水分处理植株的水平, 这与张林春等(2010)对玉米的研究结果是一致的, 但株高和叶面积依然低于正常水分处理的植株, 表明复水并不能完全弥补之前干旱对玉米生长造成的影响.第二次水分处理后, 与只遭受第二次中度或重度干旱处理的植株相比, 经历过第一次中度干旱处理的植株在遭受第二次中度干旱后, 其株高和生物量没有受到明显影响, 说明第一次中度干旱处理可以缓解第二次中度干旱对玉米生长的影响; 经历过第一次重度干旱处理的植株在遭受第二次重度干旱后, Tr、Gs、Ci、Pn、Amax、AQY和叶面积生长速率显著升高, 而株高、叶面积和生物量则显著降低, 说明第一次重度干旱处理在光合上能提高植株对二次干旱的抵抗能力, 但不能弥补前期干旱处理对生长产生的不利影响. ...
Growth and gas exchange of three sorghum cultivars under drought stress.
1
2003
... (Tsuji et al., 2003; Yao et al., 2009; Shen et al., 2015).干旱胁迫对玉米的生长发育有抑制作用, 并能引起玉米最终产量的降低(张仁和等, 2011; 姜鹏等, 2013; Farhangfar et al., 2015).本研究的结果也显示, 第一次重度干旱显著降低玉米叶片的Tr、Gs、Ci、Pn、Amax, 并使LCP和Rd显著升高, 说明玉米通过减小气孔开度降低了蒸腾水分散失, 同时, 光合能力下降, 光合产物的消耗增加, 这不利于有机物质的积累.遭受中度和重度干旱处理的玉米植株的株高和叶面积均显著降低, 生物量的积累随干旱程度的加剧呈显著降低趋势, 而根冠比呈显著升高趋势, 表明水分亏缺处理下生物量优先向根分配, 有利于植物吸收土壤中的水分, 以更好地适应干旱环境(Jackson et al., 2000).虽然中度和重度干旱处理均引起根冠比的增加, 但不同的是, 中度干旱下, 地上部分生物量降低, 但根系生长没有受到影响, 且这种根冠比的增加一直延续到复水和二次干旱处理之后; 而重度干旱下, 地上部分和根系生物量均降低, 且根系生物量的降低幅度更大.干旱处理后, 复水能使经历过中度或重度干旱处理的玉米植株的光合能力和生长速率恢复到甚至超过正常水分处理植株的水平, 这与张林春等(2010)对玉米的研究结果是一致的, 但株高和叶面积依然低于正常水分处理的植株, 表明复水并不能完全弥补之前干旱对玉米生长造成的影响.第二次水分处理后, 与只遭受第二次中度或重度干旱处理的植株相比, 经历过第一次中度干旱处理的植株在遭受第二次中度干旱后, 其株高和生物量没有受到明显影响, 说明第一次中度干旱处理可以缓解第二次中度干旱对玉米生长的影响; 经历过第一次重度干旱处理的植株在遭受第二次重度干旱后, Tr、Gs、Ci、Pn、Amax、AQY和叶面积生长速率显著升高, 而株高、叶面积和生物量则显著降低, 说明第一次重度干旱处理在光合上能提高植株对二次干旱的抵抗能力, 但不能弥补前期干旱处理对生长产生的不利影响. ...
Drought tolerance and transplanting performance of holm oak (Quercus ilex) seedlings after drought hardening in the nursery.
1
2004
... 当植物遭受干旱胁迫后, 体内的水分平衡被破坏, 并引起气孔关闭和光合速率下降.随着干旱的加剧, 植物体内会产生和积累大量的活性氧, 使得叶绿体结构被破坏, 光合作用的相关酶失活或变性, 并最终抑制植物的生长(Galmés et al., 2007; McDowell et al., 2008; Chaves et al., 2009).旱后复水能够使植物的生理功能得到恢复, 可在一定程度上弥补干旱对植物造成的伤害(山仑, 2003), 提高植物的光合速率并使其生长加速(Fortunati et al., 2008; 罗宏海等, 2008; Aidar et al., 2014; 厉广辉等, 2014).不过, 干旱后复水对植物生长上的补偿往往是有限的, 植株生长的恢复程度可能与复水前干旱的胁迫程度和持续时间有关(刘晓英等, 2001; Xu & Zhou, 2007; Xu et al., 2009).当经历过水分胁迫及复水的植物再次遇到干旱时, 植物对干旱适应性的增强可能与耐受基因的转录、渗透调节和抗氧化能力有关, 从而可在生理、生化和分子水平上具有更好地适应和抵抗干旱的能力(Ruiz-Sánchez et al., 2000; Villar-Salvador et al., 2004; Bruce et al., 2007; 徐芬芬等, 2009; Luo et al., 2011; Ding et al., 2012; 荣智媛等, 2012; 徐超华等, 2012).不过, 也有研究显示, 经历过干旱胁迫的植物再次遇到干旱时并没有提高自身抵御干旱的能力(Lloret et al., 2004; Zavalloni et al., 2008; Walter et al., 2011).因此, 在全球气候变化导致的干旱频率增加的背景下, 深入研究植物对多次干旱的响应机制, 在理论和实践上都具有重要 意义. ...
Do plants remember drought? Hints towards a drought-memory in grasses.
1
2011
... 当植物遭受干旱胁迫后, 体内的水分平衡被破坏, 并引起气孔关闭和光合速率下降.随着干旱的加剧, 植物体内会产生和积累大量的活性氧, 使得叶绿体结构被破坏, 光合作用的相关酶失活或变性, 并最终抑制植物的生长(Galmés et al., 2007; McDowell et al., 2008; Chaves et al., 2009).旱后复水能够使植物的生理功能得到恢复, 可在一定程度上弥补干旱对植物造成的伤害(山仑, 2003), 提高植物的光合速率并使其生长加速(Fortunati et al., 2008; 罗宏海等, 2008; Aidar et al., 2014; 厉广辉等, 2014).不过, 干旱后复水对植物生长上的补偿往往是有限的, 植株生长的恢复程度可能与复水前干旱的胁迫程度和持续时间有关(刘晓英等, 2001; Xu & Zhou, 2007; Xu et al., 2009).当经历过水分胁迫及复水的植物再次遇到干旱时, 植物对干旱适应性的增强可能与耐受基因的转录、渗透调节和抗氧化能力有关, 从而可在生理、生化和分子水平上具有更好地适应和抵抗干旱的能力(Ruiz-Sánchez et al., 2000; Villar-Salvador et al., 2004; Bruce et al., 2007; 徐芬芬等, 2009; Luo et al., 2011; Ding et al., 2012; 荣智媛等, 2012; 徐超华等, 2012).不过, 也有研究显示, 经历过干旱胁迫的植物再次遇到干旱时并没有提高自身抵御干旱的能力(Lloret et al., 2004; Zavalloni et al., 2008; Walter et al., 2011).因此, 在全球气候变化导致的干旱频率增加的背景下, 深入研究植物对多次干旱的响应机制, 在理论和实践上都具有重要 意义. ...
水分胁迫前的干旱锻炼对小麦光合生理特性的影响
2010
循环干旱锻炼对在干旱胁迫下烟草植株光合参数及叶绿素荧光参数的影响
1
2012
... 当植物遭受干旱胁迫后, 体内的水分平衡被破坏, 并引起气孔关闭和光合速率下降.随着干旱的加剧, 植物体内会产生和积累大量的活性氧, 使得叶绿体结构被破坏, 光合作用的相关酶失活或变性, 并最终抑制植物的生长(Galmés et al., 2007; McDowell et al., 2008; Chaves et al., 2009).旱后复水能够使植物的生理功能得到恢复, 可在一定程度上弥补干旱对植物造成的伤害(山仑, 2003), 提高植物的光合速率并使其生长加速(Fortunati et al., 2008; 罗宏海等, 2008; Aidar et al., 2014; 厉广辉等, 2014).不过, 干旱后复水对植物生长上的补偿往往是有限的, 植株生长的恢复程度可能与复水前干旱的胁迫程度和持续时间有关(刘晓英等, 2001; Xu & Zhou, 2007; Xu et al., 2009).当经历过水分胁迫及复水的植物再次遇到干旱时, 植物对干旱适应性的增强可能与耐受基因的转录、渗透调节和抗氧化能力有关, 从而可在生理、生化和分子水平上具有更好地适应和抵抗干旱的能力(Ruiz-Sánchez et al., 2000; Villar-Salvador et al., 2004; Bruce et al., 2007; 徐芬芬等, 2009; Luo et al., 2011; Ding et al., 2012; 荣智媛等, 2012; 徐超华等, 2012).不过, 也有研究显示, 经历过干旱胁迫的植物再次遇到干旱时并没有提高自身抵御干旱的能力(Lloret et al., 2004; Zavalloni et al., 2008; Walter et al., 2011).因此, 在全球气候变化导致的干旱频率增加的背景下, 深入研究植物对多次干旱的响应机制, 在理论和实践上都具有重要 意义. ...
干湿交替灌溉方式下水稻节水增产机理研究
2
2009
... 当植物遭受干旱胁迫后, 体内的水分平衡被破坏, 并引起气孔关闭和光合速率下降.随着干旱的加剧, 植物体内会产生和积累大量的活性氧, 使得叶绿体结构被破坏, 光合作用的相关酶失活或变性, 并最终抑制植物的生长(Galmés et al., 2007; McDowell et al., 2008; Chaves et al., 2009).旱后复水能够使植物的生理功能得到恢复, 可在一定程度上弥补干旱对植物造成的伤害(山仑, 2003), 提高植物的光合速率并使其生长加速(Fortunati et al., 2008; 罗宏海等, 2008; Aidar et al., 2014; 厉广辉等, 2014).不过, 干旱后复水对植物生长上的补偿往往是有限的, 植株生长的恢复程度可能与复水前干旱的胁迫程度和持续时间有关(刘晓英等, 2001; Xu & Zhou, 2007; Xu et al., 2009).当经历过水分胁迫及复水的植物再次遇到干旱时, 植物对干旱适应性的增强可能与耐受基因的转录、渗透调节和抗氧化能力有关, 从而可在生理、生化和分子水平上具有更好地适应和抵抗干旱的能力(Ruiz-Sánchez et al., 2000; Villar-Salvador et al., 2004; Bruce et al., 2007; 徐芬芬等, 2009; Luo et al., 2011; Ding et al., 2012; 荣智媛等, 2012; 徐超华等, 2012).不过, 也有研究显示, 经历过干旱胁迫的植物再次遇到干旱时并没有提高自身抵御干旱的能力(Lloret et al., 2004; Zavalloni et al., 2008; Walter et al., 2011).因此, 在全球气候变化导致的干旱频率增加的背景下, 深入研究植物对多次干旱的响应机制, 在理论和实践上都具有重要 意义. ...
... ; 徐芬芬等, 2009; Luo et al., 2011; Ding et al., 2012; 荣智媛等, 2012; 徐超华等, 2012).不过, 也有研究显示, 经历过干旱胁迫的植物再次遇到干旱时并没有提高自身抵御干旱的能力(Lloret et al., 2004; Zavalloni et al., 2008; Walter et al., 2011).因此, 在全球气候变化导致的干旱频率增加的背景下, 深入研究植物对多次干旱的响应机制, 在理论和实践上都具有重要 意义. ...
Photosynthetic recovery of a perennial grass Leymus chinensis after different periods of soil drought.
1
2007
... 当植物遭受干旱胁迫后, 体内的水分平衡被破坏, 并引起气孔关闭和光合速率下降.随着干旱的加剧, 植物体内会产生和积累大量的活性氧, 使得叶绿体结构被破坏, 光合作用的相关酶失活或变性, 并最终抑制植物的生长(Galmés et al., 2007; McDowell et al., 2008; Chaves et al., 2009).旱后复水能够使植物的生理功能得到恢复, 可在一定程度上弥补干旱对植物造成的伤害(山仑, 2003), 提高植物的光合速率并使其生长加速(Fortunati et al., 2008; 罗宏海等, 2008; Aidar et al., 2014; 厉广辉等, 2014).不过, 干旱后复水对植物生长上的补偿往往是有限的, 植株生长的恢复程度可能与复水前干旱的胁迫程度和持续时间有关(刘晓英等, 2001; Xu & Zhou, 2007; Xu et al., 2009).当经历过水分胁迫及复水的植物再次遇到干旱时, 植物对干旱适应性的增强可能与耐受基因的转录、渗透调节和抗氧化能力有关, 从而可在生理、生化和分子水平上具有更好地适应和抵抗干旱的能力(Ruiz-Sánchez et al., 2000; Villar-Salvador et al., 2004; Bruce et al., 2007; 徐芬芬等, 2009; Luo et al., 2011; Ding et al., 2012; 荣智媛等, 2012; 徐超华等, 2012).不过, 也有研究显示, 经历过干旱胁迫的植物再次遇到干旱时并没有提高自身抵御干旱的能力(Lloret et al., 2004; Zavalloni et al., 2008; Walter et al., 2011).因此, 在全球气候变化导致的干旱频率增加的背景下, 深入研究植物对多次干旱的响应机制, 在理论和实践上都具有重要 意义. ...
Are plant growth and photosynthesis limited by pre-drought following rewatering in grass?
2009
Effects of drought stress and selenium supply on growth and physiological characteristics of wheat seedlings.
1
2009
... (Tsuji et al., 2003; Yao et al., 2009; Shen et al., 2015).干旱胁迫对玉米的生长发育有抑制作用, 并能引起玉米最终产量的降低(张仁和等, 2011; 姜鹏等, 2013; Farhangfar et al., 2015).本研究的结果也显示, 第一次重度干旱显著降低玉米叶片的Tr、Gs、Ci、Pn、Amax, 并使LCP和Rd显著升高, 说明玉米通过减小气孔开度降低了蒸腾水分散失, 同时, 光合能力下降, 光合产物的消耗增加, 这不利于有机物质的积累.遭受中度和重度干旱处理的玉米植株的株高和叶面积均显著降低, 生物量的积累随干旱程度的加剧呈显著降低趋势, 而根冠比呈显著升高趋势, 表明水分亏缺处理下生物量优先向根分配, 有利于植物吸收土壤中的水分, 以更好地适应干旱环境(Jackson et al., 2000).虽然中度和重度干旱处理均引起根冠比的增加, 但不同的是, 中度干旱下, 地上部分生物量降低, 但根系生长没有受到影响, 且这种根冠比的增加一直延续到复水和二次干旱处理之后; 而重度干旱下, 地上部分和根系生物量均降低, 且根系生物量的降低幅度更大.干旱处理后, 复水能使经历过中度或重度干旱处理的玉米植株的光合能力和生长速率恢复到甚至超过正常水分处理植株的水平, 这与张林春等(2010)对玉米的研究结果是一致的, 但株高和叶面积依然低于正常水分处理的植株, 表明复水并不能完全弥补之前干旱对玉米生长造成的影响.第二次水分处理后, 与只遭受第二次中度或重度干旱处理的植株相比, 经历过第一次中度干旱处理的植株在遭受第二次中度干旱后, 其株高和生物量没有受到明显影响, 说明第一次中度干旱处理可以缓解第二次中度干旱对玉米生长的影响; 经历过第一次重度干旱处理的植株在遭受第二次重度干旱后, Tr、Gs、Ci、Pn、Amax、AQY和叶面积生长速率显著升高, 而株高、叶面积和生物量则显著降低, 说明第一次重度干旱处理在光合上能提高植株对二次干旱的抵抗能力, 但不能弥补前期干旱处理对生长产生的不利影响. ...
Does a warmer climate with frequent mild water shortages protect grassland communities against a prolonged drought?
1
2008
... 当植物遭受干旱胁迫后, 体内的水分平衡被破坏, 并引起气孔关闭和光合速率下降.随着干旱的加剧, 植物体内会产生和积累大量的活性氧, 使得叶绿体结构被破坏, 光合作用的相关酶失活或变性, 并最终抑制植物的生长(Galmés et al., 2007; McDowell et al., 2008; Chaves et al., 2009).旱后复水能够使植物的生理功能得到恢复, 可在一定程度上弥补干旱对植物造成的伤害(山仑, 2003), 提高植物的光合速率并使其生长加速(Fortunati et al., 2008; 罗宏海等, 2008; Aidar et al., 2014; 厉广辉等, 2014).不过, 干旱后复水对植物生长上的补偿往往是有限的, 植株生长的恢复程度可能与复水前干旱的胁迫程度和持续时间有关(刘晓英等, 2001; Xu & Zhou, 2007; Xu et al., 2009).当经历过水分胁迫及复水的植物再次遇到干旱时, 植物对干旱适应性的增强可能与耐受基因的转录、渗透调节和抗氧化能力有关, 从而可在生理、生化和分子水平上具有更好地适应和抵抗干旱的能力(Ruiz-Sánchez et al., 2000; Villar-Salvador et al., 2004; Bruce et al., 2007; 徐芬芬等, 2009; Luo et al., 2011; Ding et al., 2012; 荣智媛等, 2012; 徐超华等, 2012).不过, 也有研究显示, 经历过干旱胁迫的植物再次遇到干旱时并没有提高自身抵御干旱的能力(Lloret et al., 2004; Zavalloni et al., 2008; Walter et al., 2011).因此, 在全球气候变化导致的干旱频率增加的背景下, 深入研究植物对多次干旱的响应机制, 在理论和实践上都具有重要 意义. ...
干旱及复水对不同抗旱性玉米光合特性的影响
2010
干旱胁迫对玉米苗期植株生长和光合特性的影响
1
2011
... (Tsuji et al., 2003; Yao et al., 2009; Shen et al., 2015).干旱胁迫对玉米的生长发育有抑制作用, 并能引起玉米最终产量的降低(张仁和等, 2011; 姜鹏等, 2013; Farhangfar et al., 2015).本研究的结果也显示, 第一次重度干旱显著降低玉米叶片的Tr、Gs、Ci、Pn、Amax, 并使LCP和Rd显著升高, 说明玉米通过减小气孔开度降低了蒸腾水分散失, 同时, 光合能力下降, 光合产物的消耗增加, 这不利于有机物质的积累.遭受中度和重度干旱处理的玉米植株的株高和叶面积均显著降低, 生物量的积累随干旱程度的加剧呈显著降低趋势, 而根冠比呈显著升高趋势, 表明水分亏缺处理下生物量优先向根分配, 有利于植物吸收土壤中的水分, 以更好地适应干旱环境(Jackson et al., 2000).虽然中度和重度干旱处理均引起根冠比的增加, 但不同的是, 中度干旱下, 地上部分生物量降低, 但根系生长没有受到影响, 且这种根冠比的增加一直延续到复水和二次干旱处理之后; 而重度干旱下, 地上部分和根系生物量均降低, 且根系生物量的降低幅度更大.干旱处理后, 复水能使经历过中度或重度干旱处理的玉米植株的光合能力和生长速率恢复到甚至超过正常水分处理植株的水平, 这与张林春等(2010)对玉米的研究结果是一致的, 但株高和叶面积依然低于正常水分处理的植株, 表明复水并不能完全弥补之前干旱对玉米生长造成的影响.第二次水分处理后, 与只遭受第二次中度或重度干旱处理的植株相比, 经历过第一次中度干旱处理的植株在遭受第二次中度干旱后, 其株高和生物量没有受到明显影响, 说明第一次中度干旱处理可以缓解第二次中度干旱对玉米生长的影响; 经历过第一次重度干旱处理的植株在遭受第二次重度干旱后, Tr、Gs、Ci、Pn、Amax、AQY和叶面积生长速率显著升高, 而株高、叶面积和生物量则显著降低, 说明第一次重度干旱处理在光合上能提高植株对二次干旱的抵抗能力, 但不能弥补前期干旱处理对生长产生的不利影响. ...

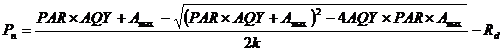


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}