多环芳烃(polycyclic aromatic hydrocarbons, PAHs)是环境中普遍存在的持久性有机污染物, 多数具有致癌、致畸或致突变效应(Chen & Liao, 2006 )。因其性质稳定、水溶性差, 易于被土壤颗粒吸附, 土壤中PAHs含量逐年上升(Tao et al ., 2004 ; Li et al ., 2006 ), 严重威胁着土壤环境质量及农产品质量安全; 土壤中PAHs的迁移转化行为及其环境效应已成为环境修复领域的研究热点(Mueller & Shann, 2006 ; Lakshmanan et al ., 2014 )。
植物的存在能加快土壤中PAHs的去除(Joner & Leyval, 2003 ; Kim et al ., 2004 ; Gao et al ., 2006 ), 根系分泌物(root exudates)营造的根际微生态特征则是影响PAHs快速消解的重要原因(Sumia et al ., 2013 ; Tejeda-Agredano et al ., 2013 )。植物修复过程中, 根系分泌物在改善土壤属性、提高污染物的生物可利用性、缓解环境胁迫等方面具有重要作用(Bertin & Yang, 2003 ; Phillipsa et al ., 2012 ; Shahzad et al ., 2015 )。一方面, 根系分泌的某些胞外酶能够直接参与PAHs的降解过程; 另一方面, 分泌物中的可溶性糖、低分子量有机酸、氨基酸等物质, 为根际微生物提供充足的营养, 增加根际微生物的数量、提高微生物活性, 进而加速PAHs降解(Susarla et al ., 2002 ; Xie et al ., 2012 ; Lakshmanan et al ., 2014 )。同时, 根系分泌物还会直接影响PAHs的固定和活化, 进而影响PAHs在土壤-植物系统中的迁移转化行为、促进PAHs去除(Marschner et al ., 2004 ; Abhilash et al ., 2012 ); PAHs的环数越多, 根际效益越明显(Sun et al ., 2010 )。不同植物间, 因根系分泌物的组成成分及其释放特征常存在一定差异, 对根际微生物的区系结构、代谢活性的影响也不一样, 进而表现出不同的修复潜力(Kirk et al ., 2005 ; Luo et al ., 2006 ; Hunter et al ., 2014 )。因此, 探讨PAHs胁迫下不同修复潜力植物间根系分泌物中主要组分的释放特征及其种间差异, 对于研究植物修复的生物学机制、揭示不同植物间修复潜力的种间差异具有重要意义。鉴于此, 本研究以对PAHs污染土壤具有一定修复潜力的羊茅属(Festuca )植物为试验对象, 以苯并[α]芘(BaP)为PAHs代表物, 借助根际袋土培法, 对比研究PAHs胁迫下不同修复潜力的5种羊茅属植物在不同生长期时根系分泌物中几种低分子量有机物的释放特征, 为植物修复理论的研究与实践提供依据。
1 材料和方法
1.1 试验材料
1.1.1 PAHs代表物
BaP是PAHs类污染物中分布最广、致癌性最强的烃类物质, 其来源广泛、稳定性强, 且与其他PAHs含量有一定相关性。本研究选择BaP (Aldrich, 纯度> 97%)为PAHs代表物。
1.1.2 供试植物
供试植物为5种羊茅属植物: 苇状羊茅(F. arundinacea, Fa)、草原羊茅(F. chelungkiangnica, Fc)、毛稃羊茅(F. rubra subsp. arctica, Fr)、贫芒羊茅(F. sinomutica, Fm)、细芒羊茅(F. stapfii, Fs), 种子购于四川省草原研究所。种子经3% H2 O2 溶液消毒20 min、蒸馏水冲洗干净后于烧杯中浸泡吸胀24 h, 置于培养皿中恒温(35 ℃)催芽15 h后, 播种于根际袋。
1.1.3 供试土壤
供试土壤为中性紫色土, 采自成都大学试验农场表层土(5-20 cm, 44.6 pg·L-1 检出限下未检测到BaP)。土壤理化性质: pH 7.19, 有机质含量2.14 × 104 mg·kg-1 , 阳离子交换量(CEC) 20.43 cmol·kg-1 , 速效N、P、K分别为31.62、24.74、94.81 mg·kg-1 ; 砂粒、粉粒和黏粒的质量分数分别为71.27%、9.59%和19.14%。土壤风干后过2 mm筛, 备用。
1.2 试验方法
土培试验采用根际袋试验装置在智能温室内进行(许超等, 2010 ), 涉及3个处理、5个水平(5个BaP胁迫水平下的5种植物、5个生长期), 重复3次。具体如下:
按照150 mg·kg-1 CO(NH2 )2 、50 mg·kg-1 Ca(H2 PO4 )·2H2 O和100 mg·kg-1 K2 SO4 比例补充底肥后, 取适量土壤过2 mm筛, 加入BaP丙酮溶液; 待丙酮挥发后, 用未污染土壤不断稀释、搅拌, 过2 mm筛、混匀, 结合农田土壤中PAHs的背景值, 获得5个污染水平。50%田间持水量(WHC)、室温状态下平衡7天(Pan et al ., 2008 ; Wei & Pan, 2010 )后, 测得土样中BaP的初始含量分别为: 10.25 mg·kg-1 (T1 )、20.37 mg·kg-1 (T2 )、40.45 mg·kg-1 (T3 )、80.24 mg·kg-1 (T4 )和161.74 mg·kg-1 (T5 )。取污染土样2 kg装入20 cm × 15 cm的塑料盆钵中, 其中的400 g装在300目尼龙网袋中; 尼龙网袋放在盆中央, 四周及底部均为同质土样, 保持袋内外土壤高度相同。每种污染水平分装75盆, 50% WHC、室温状态下平衡5天(Pan et al ., 2008 ; Wei & Pan, 2010 ); 选取籽粒饱满、大小均匀的催芽后种子单播于袋内土壤中, 每种植物播种15盆, 每盆10颗, 置于温室中; 另设一组没有播种的污染土样作为对照, 每个处理重复3次。试验期间, 日间室温为25 ℃、夜间为18 ℃, 自然光照条件; 每2天随机交换盆钵在温室中的位置, WHC维持在50% (称重补水法)。播种日记为0天, 在植物生长30、40、50、60和70天时, 分别对不同胁迫强度的供试植物、土样破坏性取样: 连同根际袋小心取出植物幼苗、尽可能不伤及其根系, 轻轻抖落根际土壤后收集其根系分泌物; 混匀根际土壤、非根际土壤后, 四分法取样, 装袋、冷冻干燥, 过 1 mm筛后低温(-20 ℃)保存、分析BaP残留量。
1.3 根系分泌物收集
取出根际袋, 用自来水将根洗净、去离子水冲洗3遍后, 浸于盛有适量Milli-Q超纯水的烧杯中(锡纸包裹杯壁, 确保根部避光; 液面高度以漫过根部为宜), 自然光照下收集4 h, 用少量高纯水冲洗根系3次, 冲洗液与收集液合并、慢速滤过0.45 μm微孔滤膜(Tu et al ., 2004 ; 许超等, 2010 )。将滤液定容至50 mL, -20 ℃以下储存, 待分析。取10 mL滤液真空冷冻干燥至干, 加1 mL Milli-Q超纯水溶解, -20 ℃以下储存, 用于分析低分子量有机酸; 取20 mL滤液真空冷冻干燥至干, 加2 mL Milli-Q超纯水溶解, -20 ℃以下储存, 用于分析氨基酸(许超等, 2010 )。
1.4 分析方法
1.4.1 可溶性糖测定
使用蒽酮比色法。可溶性糖含量以每kg根(干质量)分泌的可溶性糖计量, 单位为mg·kg-1 。
1.4.2 低分子量有机酸测定
使用高效液相色谱(HPLC)测定。检测条件: Agilent 1100色谱仪(Agilent Technologies Palo Alto, USA), XB-C18 色谱柱、紫外检测器(λ = 210 nm); 流动相为0.025 mol·L-1 的H3 PO4 -KH2 PO4 缓冲液(pH值2.3), 流速为0.7 mL·min-1 , 进样量为20 μL, 柱温30 ℃。所用样品3个重复。低分子量有机酸释放量以每kg根(干质量)分泌的有机酸总量计量, 单位为mg·kg-1 。
1.4.3 氨基酸测定
使用日立L-8900型氨基酸自动分析仪(Hitachi, Tokyo, Japan)测定。检测条件: 离子交换柱规格2.6 mm × 150 mm, 交换树型号: NO 2619 (52051), 柱温53 ℃, 泵流速0.225 mL·min-1 , 进样量50 μL, 分析时间72 min。在检测波长λ = 440 nm条件下, 检出限为2.2 × 10-9 mol, 保留时间重现性变异系数(CV ) 1%以下、峰面积重现性CV 1.5%以下。所用样品3个重复。氨基酸量以每kg根(干质量)分泌的有机酸总量计量, 单位为mg·kg-1 。
1.4.4 BaP测定
参照Pan等(2008) 的方法, 略作改进: 取2.0 g土样于离心管内, 加入2.0 g无水Na2 SO4 , 混匀; 加入10 mL二氯甲烷(CH2 Cl2 ), 40 ℃下超声萃取1 h; 4000 r·min-1 下离心。取3 mL上清液过Fisher Pasteur玻璃管硅胶柱, 1:1二氯甲烷和正己烷洗脱, 40 ℃下浓缩至干, 用甲醇定容到2 mL, 过0.22 µm微孔滤膜后进行HPLC (Waters 600, Waters China, Hong Kong, China)分析。检测条件: DAD检测器, ¢4.6 × 250 mm C18 反相色谱柱; 甲醇:水(V/V, 83:17)为流动相, λ = 290 nm。在此条件下, BaP检测限为44.6 pg·L-1 , 加标回收率为94.36% (n = 7, 相对标准差(RSD ) < 5.94%))。
1.5 数据分析
数据经Microsoft Excel 2003处理后, 借助SPSS 11.5软件最小显著差异(LSD )法分析不同处理间差异显著性。
2 结果和分析
2.1 BaP胁迫对待试植物生物量的影响
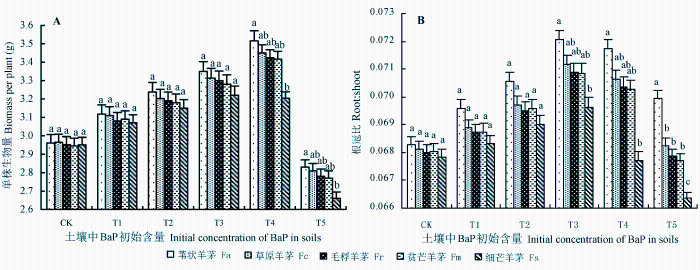
图1 为BaP胁迫下5种羊茅属植物在50天时的生长状况。可以看出, 轻度(T1 -T2 )或中度(T3 -T4 )胁迫刺激了植物生长, 5种植物生物量(单株干质量)变化呈现T4 ≥ T3 ≥ T2 ≥ T1 ≥ CK的趋势, T4 水平时达到峰值, 且T4 > T1 、T4 > CK (p < 0.05); 细芒羊茅的生物量变化虽然呈现T4 ≥ T3 ≥ T2 ≥ T1 ≥ CK的趋势, 但各胁迫水平间差异不显著(p > 0.05)。重度(T5 )胁迫抑制了植物生长, 其生物量明显低于其他胁迫水平(p < 0.05)。植物的根冠比也呈现与生物量类似的变化, 但细芒羊茅的峰值(0.0695 ± 0.0006)出现在T3 水平, 之后则逐渐降低。相同胁迫强度下, 除T4 、T5 水平的细芒羊茅外, 5种羊茅属植物的生物量间无明显差异(p > 0.05)。总体上, BaP胁迫对细芒羊茅的生长影响较大: 相同胁迫强度下, 其生物量、根冠比都低于其他植物, 胁迫强度越大、趋势越明显; T3 -T5 水平时, 差异达到显著水平(p < 0.05)。
试验期间, BaP胁迫下5种羊茅属植物在其他时段的生长状态与50天时类似。总体上, 待试植物除生物量略有变化外, 无其他表观受害症状(叶脉周围出现褐色或黄褐色坏死斑点, 叶缘褪绿、叶尖干枯, 叶片下垂; 叶色发黄、根毛减少), 均显示出较强的BaP污染耐受能力。
图1 苯并[α]芘(BaP)胁迫对5种羊茅属植物生物量、根冠比的影响(平均值±标准误差)。CK、T1 、T2 、T3 、T4 、T5 , BaP的初始含量分别为0、10.25、20.37、40.45、80.24和161.74 mg·kg-1 。不同小写字母表示相同胁迫条件下差异显著(p < 0.05)。
Fig. 1 Biomass and root: shoot ratio of plants in five Festuca species growing in soils contaminated with different concentrations of benzo [α] pyrene (BaP) (mean ± SE ). CK, T1 , T2 , T3 , T4 , and T5 designate the treatments with initial concentrations of BaP at 0, 10.25, 20.37, 40.45, 80.24 and 161.74 mg·kg-1 , respectively. Fa, F. arundinacea ; Fc, F. chelungkiangnica ; Fr, F. rubra subsp. arctica ; Fm, F. sinomutica ; Fs, F. stapfii. Different lowercase letters indicate significant differences (p < 0.05) under the same stress conditions.
2.2 待试植物对BaP污染土壤修复潜力的种间差异
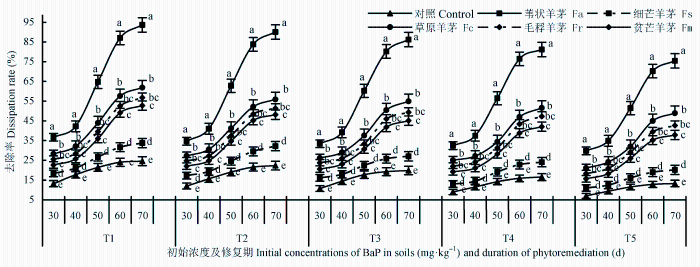
图2 为不同生长时期(30-70天)内5种羊茅属植物对BaP污染土壤(10.25-161.74 mg·kg-1 )修复效果的种间差异。可以看出, 植物的存在促进了土壤中BaP的去除: 相同污染水平下, 种植植物土壤中BaP的去除率远高于对照组(n = 5, p < 0.05), 修复期越长, 差异越显著; 植物的种类不同, 对土壤中BaP的去除效果也不一样。其间, 苇状羊茅表现出较强的修复潜力, 草原羊茅、毛稃羊茅、贫芒羊茅次之, 细芒羊茅最差。相同污染条件下, 5种羊茅属植物对BaP污染土壤的修复潜力依次为: 苇状羊茅>草原羊茅≥毛稃羊茅≥贫芒羊茅>细芒羊茅, 但草原羊茅与毛稃羊茅间、毛稃羊茅和贫芒羊茅间差异不甚显著(p > 0.05)。
图2 待试植物对BaP污染土壤修复潜力的种间差异(平均值±标准误差)。T1 、T2 、T3 、T4 、T5 , BaP的初始含量分别为10.25、20.37、40.45、80.24和161.74 mg·kg-1 。数据源右侧的不同小写字母表示相同胁迫条件下差异显著(p < 0.05)。
Fig. 2 Differences in phytoremediation potentials of benzo [α] pyrene (BaP)-contaminated soils among five Festuca species (mean ± SE ). T1 , T2 , T3 , T4 , and T5 designate the treatments with initial concentrations of BaP at 10.25, 20.37, 40.45, 80.24 and 161.74 mg·kg-1 , respectively. Fa, F. arundinacea ; Fc, F. chelungkiangnica ; Fr, F. rubra subsp. arctica ; Fm, F. sinomutica ; Fs, F. stapfii. Different lowercase letters next to the right of data points indicate significant differences (p < 0.05) under the same stress conditions.
不同修复期(生长期)内, 植物的修复潜力也不一样。修复前期(30-40天)、后期(60-70天)均低于修复中期(40-60天)(p < 0.05), 呈现明显出“S”形变化曲线, 曲线拐点出现在修复期的第50天前后。
2.3 待试植物根系分泌物中几种低分子量组分的释放特征
2.3.1 可溶性糖的释放特征
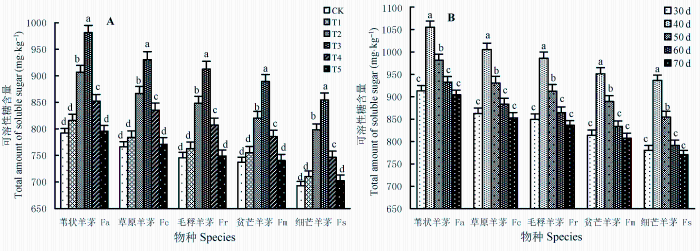
图3 显示, BaP胁迫不同程度地促进了根系分泌物中可溶性糖的分泌, 并随着胁迫强度增大(图3 A)、胁迫期延长(图3 B)呈现“先升后降”变化。相同胁迫期(图3 A: 50天)内, 各处理组根系可溶性糖的分泌量均高于对照组(CK), T3 胁迫水平下达到峰值; T2 、T3 、T4 处理与CK组间差异显著(p < 0.05), 但T1 、T5 处理与CK组间差异不显著(p > 0.05)。相同胁迫水平(图3 B: T3 )下, 不同胁迫期内根系可溶性糖分泌量也不一样, 40天时最多, 且明显多于其他时期(p < 0.05), 但在30、60和70天时可溶性糖释放量差异不大(p > 0.05)。可见, BaP胁迫能促进植物根系分泌更多的可溶性糖, 在一定胁迫强度、胁迫期内, 可溶性糖分泌量随胁迫强度的增强而增多; 超越此范围, 分泌量开始减少。
图3 苯并[α]芘(BaP)的胁迫强度(A)、T3 处理下胁迫期(B)对5种羊茅属植物根系分泌物中可溶性糖含量的影响(平均值±标准误差)。CK、T1 、T2 、T3 、T4 、T5 , BaP的初始含量分别为0、10.25、20.37、40.45、80.24和161.74 mg·kg-1 。不同小写字母表示相同胁迫条件下差异显著(p < 0.05)。
Fig. 3 The effects of benzo [α] pyrene (BaP) stress level (A) and stress stage under T3 treatment (B) on the total amount of soluble sugars in root exudates of five Festuca species (mean ± SE ) . CK, T1 , T2 , T3 , T4 , and T5 designate the treatments with initial concentrations of BaP at 0, 10.25, 20.37, 40.45, 80.24 and 161.74 mg·kg-1 , respectively. Fa, F. arundinacea ; Fc, F. chelungkiangnica ; Fr, F. rubra subsp. arctica ; Fm, F. sinomutica ; Fs, F. stapfii. Different lowercase letters indicate significant differences (p < 0.05) under the same stress conditions.
2.3.2 低分子量有机酸的释放特征
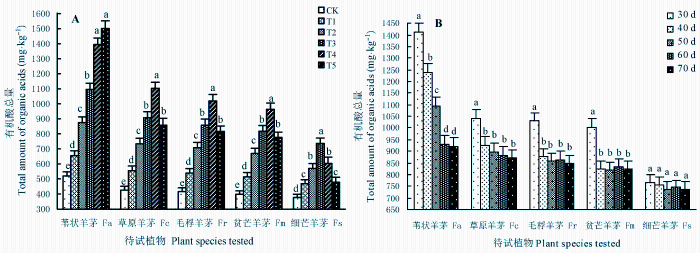
5种羊茅属植物根系分泌物中, 草酸(83.76% ±0.05%)、乙酸(6.34% ± 0.03%)、乳酸(4.95% ± 0.03%)和苹果酸(2.29% ± 0.02%)的质量分数之和超过了低分子量有机酸总量的97.34%。苇状羊茅的根系分泌物中检测出微量的顺丁烯二酸(0.27% ± 0.01%)、反丁烯二酸(0.18% ± 0.01%), 草原羊茅、毛稃羊茅、贫芒羊茅的根系分泌物中检测出反顺丁烯二酸(0.43% ± 0.02%), 细芒羊茅的根系分泌物中仅检测出微量顺丁烯二酸(0.16% ± 0.01%); 不同污染水平下同种植物根系分泌物中低分子量有机酸的组成成分基本不变。图4 为待试植物根系在不同胁迫期内释放的低分子量有机酸总量的变化趋势。
图4A 显示, 相同胁迫期(50天)内, 苇状羊茅根系分泌的有机酸总高于相同污染水平下的其他植物, 污染水平越高、有机酸分泌量越多, 呈现T5 ≥ T4 > T3 > T2 > T1 > CK趋势; 除T4 与T5 间外, 各污染水平间差异显著(p < 0.05)。草原羊茅、毛稃羊茅、贫芒羊茅根系分泌的有机酸总量以T4 水平最高, 呈现T4 > T3 ≥ T5 > T2 > T1 > CK趋势; 除T3 与T5 间外, 各污染水平间差异显著(p < 0.05)。细芒羊茅以T3 水平最高, 呈现T3 > T4 ≥ T2 > T5 ≥ T1 > CK趋势, 但T4 与T2 、T5 与T1 间差异不显著(p > 0.05)。
图4 苯并[α]芘(BaP)的胁迫强度(A)、T3 处理下胁迫期(B)对5种羊茅属植物根系分泌物中有机酸总量的影响(平均值±标准误差)。CK、T1 、T2 、T3 、T4 、T5 , BaP的初始含量分别为0、10.25、20.37、40.45、80.24和161.74 mg·kg-1 。不同小写字母表示相同胁迫条件下差异显著(p < 0.05)。
Fig. 4 The effects of benzo [α] pyrene (BaP) stress level (A) and stress stage under T3 treatment (B) on the total amount of organic acids in root exudates of five Festuca species (mean ± SE ) . CK, T1 , T2 , T3 , T4 , and T5 designate the treatments with initial concentrations of BaP at 0, 10.25, 20.37, 40.45, 80.24 and 161.74 mg·kg-1 , respectively. Fa, F. arundinacea ; Fc, F. chelungkiangnica ; Fr, F. rubra subsp. arctica ; Fm, F. sinomutica ; Fs, F. stapfii. Different lowercase letters indicate significant differences (p < 0.05) under the same stress conditions.
图4B 显示, 相同胁迫水平(T3 )下, 植物根系在不同胁迫期释放的有机酸总量也不一样。修复潜力最强的苇状羊茅在30天时释放量最多, 呈现30天> 40天> 50天> 60天≥ 70天趋势; 除第60、70天外, 其余各时期间差异显著(p < 0.05)。修复潜力较强的草原羊茅、毛稃羊茅、贫芒羊茅在30天时的释放量最多(p < 0.05), 其余各时期间差异不大(p > 0.05); 修复潜力最差的细芒羊茅在各个时期的释放量无明显差异(p > 0.05)。
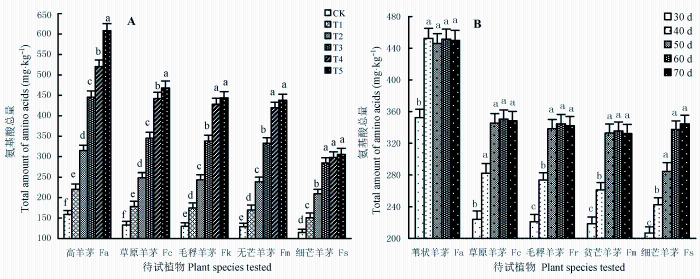
2.3.3 氨基酸的释放特征
除色氨酸外, 其他19种常见氨基酸均能在待试植物的根系分泌物中检测到。总体上, BaP胁迫促进了氨基酸的分泌, 胁迫强度越大、分泌量越多。相同胁迫期(50天; 图5A )内, 苇状羊茅根系的分泌量总高于相同污染水平下的其他植物(p < 0.05), 不同污染水平间差异显著(p < 0.05); 草原羊茅、毛稃羊茅、贫芒羊茅根系的分泌量呈现T5 ≥ T4 > T3 > T2 > T1 > CK趋势, T5 、T4 间差异不显著(p > 0.05); 细芒羊茅根系的分泌量虽随着污染水平的升高而增多, 但T5 、T4 、T3 间差异不显著(p > 0.05)。相同胁迫强度(T3 ; 图5B )下, 苇状羊茅根系的分泌量总多于其他植物(p < 0.05), 30天时的分泌量明显低于其他各时期(p < 0.05); 自40天起趋于稳定, 各时期释放量无显著差异(p > 0.05)。草原羊茅、毛稃羊茅、贫芒羊茅根系的释放量呈现30天<40天<50天≤60天≤70天趋势, 30、40、50天间差异显著(p < 0.05); 自50天起趋于稳定, 各时期释放量差异不大(p > 0.05)。细芒羊茅根系的释放量虽随胁迫时期的延长而增多, 但60、70天时的差异不显著(p > 0.05)。
图5 苯并[α]芘(BaP)的胁迫强度(A)、T3 处理下胁迫期(B)对5种羊茅属植物根系分泌物中氨基酸总量的影响(平均值±标准误差)。CK、T1 、T2 、T3 、T4 、T5 , BaP的初始含量分别为0、10.25、20.37、40.45、80.24和161.74 mg·kg-1 。不同小写字母表示相同胁迫条件下差异显著(p < 0.05)。
Fig. 5 The effects of benzo [α] pyrene (BaP) stress level (A) and stress stage under T3 treatment (B) on the total amount of amino acids in root exudates of five Festuca species (mean ± SE ) . CK, T1 , T2 , T3 , T4 , and T5 designate the treatments with initial concentrations of BaP at 0, 10.25, 20.37, 40.45, 80.24 and 161.74 mg·kg-1 , respectively. Fa, F. arundinacea ; Fc, F. chelungkiangnica ; Fr, F. rubra subsp. arctica ; Fm, F. sinomutica ; Fs, F. stapfii. Different lowercase letters indicate significant differences (p < 0.05) under the same stress conditions.
试验期间, BaP胁迫对根系分泌物中氨基酸的种类影响不大, 但对分泌量的影响较大, 且影响程度大小不一。相比之下, 苏氨酸、丝氨酸、甘氨酸、丙氨酸、脯氨酸和羟脯氨酸的释放特征较为突出。
与CK组相比, BaP胁迫促进了植物根系对苏氨酸、丝氨酸、甘氨酸、丙氨酸的释放活动。试验浓度(T1 -T5 )范围内, 其分泌量随着胁迫强度的增大而剧增, 不同胁迫水平间差异显著(p < 0.05); 脯氨酸、羟脯氨酸和天冬氨酸在苇状羊茅根系也呈现类似的释放特征, 但在细芒羊茅根系的释放量却忽高忽低、多少不一(表1 )。其间, 草原羊茅根系分泌的脯氨酸和天冬氨酸、毛稃羊茅根系分泌的脯氨酸和羟脯氨酸、贫芒羊茅根系分泌的天冬氨酸均随着BaP胁迫强度的增大而剧增, 不同水平间差异显著
(p < 0.05); 但草原羊茅根系分泌的羟脯氨酸、毛稃羊茅根系分泌的天冬氨酸, 以及贫芒羊茅根系分泌的脯氨酸、羟脯氨酸虽随着胁迫强度的增大而增多, 但不同水平间差异不显著(p > 0.05) (表1 )。
3 讨论
植物根际对PAHs的降解效果与修复植物的种类有关。Yi和Crowley (2007)研究43种植物根部组织对PAHs的降解作用发现, 有些植物对PAHs降解有明显的促进作用, 有些则无作用或有抑制作用。根系分泌物所营造的根际环境及其所呈现的根际效应是植物响应外界胁迫的重要途径, 也是影响修复效果的关键因素(Gao et al ., 2010 ; 吴林坤等, 2014 )。相同胁迫条件下, 根系分泌物的释放特征及其主要组分的种间差异主要处决于植物自身基因型的特异性及其生理状态的个体差异(Zhu et al ., 2009 ; D’Orazio et al ., 2013 )。本研究以亲缘关系较近的5种羊茅属植物为实验对象, 其基因型差异相对较小, 在一定程度上能增加研究结果的可比性。尽管待试植物在无污染(CK)、轻度或中度污染(T1 -T4 )时的生物量, 以及被植物积累的BaP量(0.08%-0.21%, 平均(0.15 ± 0.02)%差异甚微, 但它们对BaP污染土壤的修复潜力却存在显著的种间差异。可见, 植物修复过程中, 即使是遗传关系很近的同属植物间, 修复潜力的种间差异仍然存在。
植物生长指标的变化是植物受到不同程度胁迫伤害的直观表现症状(王海翠等, 2013 )。本试验中, 低度(T1 -T2 )或中度(T3 -T4 ) BaP胁迫刺激了植物生长, T4 水平时生物量出现峰值, 重度(T5 )胁迫则抑制了植物生长。通常, 多数植物对PAHs类环境胁迫具有一定的耐受域值, 而这种域值与PAHs的理化性质、植物种类和环境状况(Howsam et al ., 2000 ; Line et al ., 2003 ; 田晓雪等, 2008 )以及PAHs进入细胞后的累积或转化机制等因素有关(刘玉和周璐璐, 2011 ; 刘静等, 2015 )。一定域值内, 植物生长受到的抑制影响并不显著, 甚至能刺激植物生长; 超过这一域值, 植物生长就会受到抑制。Maliszewska-Kordy- bach和Smreczak (2000)报道, 由芴、蒽、芘和䓛4种PAHs组成的混合物, 在浓度不超过10 mg·kg-1 时促进了小麦(Triticum aestivum )、玉米(Zea mays )及燕麦(Avena sativa )的生长, 高浓度时抑制生长; 刘泓等(2008)研究了不同浓度的荧蒽培养基对拟南芥(Arabidopsis thaliana )生长的影响, 发现不同浓度的荧蒽胁迫30天后拟南芥生长量明显受到不同程度的抑制。尽管这些研究都表明, 高浓度PAHs胁迫抑制植物的生长, 但抑制作用机制尚不明确; 低浓度的PAHs对植物生长的促进作用, 有学者推测是因为PAHs具有与植物生长激素类似的作用(陈世军等, 2012 ), 但也仅仅局限于推测, 还没有被试验和实践证实。
在土壤中, 根系分泌物与根际微生物间的相互作用是一个十分重要的过程: 根系通过分泌各种次生代谢物质对根际微生物的种类、数量和分布产生影响, 对根际微生物群落结构有选择塑造作用, 进而影响PAHs的降解(Abhilash et al ., 2012 ; Edwards et al ., 2015 )。本研究显示, 胁迫环境中植物根系可溶性糖、低分子量有机酸、氨基酸等组分的释放特征与植物自身的修复潜力有关: 修复潜力较强的苇状羊茅所分泌的可溶性糖、低分子量有机酸、氨基酸总多于其他几种植物; 修复潜力越强, 其分泌量、组成成分的变化幅度越大。相同胁迫条件下, 修复潜力较强的植物对BaP的胁迫响应总表现出更强的环境适应性、生理活性上的可塑性, 修复潜力越强, 趋势越明显(图3 -5)。可以推测, 根系分泌物的释放特征势必影响根际微生物群落结构, 并产生选择性塑造作用, 但这种释放特征是植物为减轻BaP毒害而进行的自我生理调节还是BaP毒害破坏了根系正常的生理活动而造成细胞内有机物质的渗漏尚待进一步研究。
环境胁迫可能会影响根系分泌物的种类和数量(沈宏等, 2002 ; Wu et al ., 2014 )。本试验在修复潜力较强植物的根系分泌物中检测到一种低分子量有机酸——反丁烯二酸, 它可能与植物对BaP类有害物质的应激反应有关: 植物在应激反应过程中所产生的破坏性氧自由基(reactive oxygen species, ROS)容易与细胞内的生物大分子发生反应, 对线粒体、核糖体、溶酶体、过氧化物体、微粒体等细胞器以及核酸、细胞骨架蛋白等生物大分子的结构和功能产生损伤或毒害作用(Ramos & Garcia, 2007 ), 反丁烯二酸类抗氧化助剂对活性氧自由基较敏感, 能及时捕获并清除胁迫过程中产生的各种活性氧自由基、缓解环境胁迫的损害。但为什么仅在修复潜力极强的苇状羊茅、极差的细芒羊茅的根系分泌物中检测到顺丁烯二酸, 尚待进一步研究。
根系分泌物的释放特征是植物根系胁迫反应时的集中表现(Jennifer et al ., 2015 )。BaP胁迫促进了5种羊茅属植物对苏氨酸、丝氨酸、甘氨酸、丙氨酸的释放, 许超等(2010)、谢晓梅等(2011)等在研究芘胁迫对玉米、黑麦草根系分泌物的影响时也得出类似结论。本试验中, 根系分泌物中脯氨酸、羟脯氨酸和天冬氨酸等组分对BaP的胁迫响应更近乎以加和效应(additivity)甚至协同效应(synergism)形式参与植物的修复过程, 参与应激反应的组分越多, 修复潜力越强: 修复潜力最强的苇状羊茅根系以上3种组分的释放量均随胁迫强度的增强而剧增; 修复潜力次强的草原羊茅、毛稃羊茅根系上述组分中某2种的释放量随胁迫强度的增强而剧增; 修复潜力次差的贫芒羊茅根系仅上述组分中某1种的释放量随胁迫强度的增强而剧增; 修复潜力最差的细芒羊茅根系上述组分的释放量与BaP胁迫强度无明显相关性。但该响应模式是否具有普遍性, 尚待进一步验证。
4 结论
(1)试验浓度范围内, 待试植物对BaP胁迫具有较强的耐受能力, 其修复潜力依次为: 苇状羊茅> 草原羊茅≥毛稃羊茅≥贫芒羊茅>细芒羊茅, 但草原羊茅与毛稃羊茅、毛稃羊茅与贫芒羊茅间差异不显著(p > 0.05)。
(2) BaP胁迫促进了植物根系对可溶性糖的分泌, 呈现“先升后降”波动: 一定胁迫强度、胁迫时期内, 单位根质量分泌的可溶性糖随胁迫强度的增大而增加; 超过此范围, 其分泌量逐渐降低。
(3) BaP胁迫促进了根系对低分子量有机酸的释放。修复潜力越强, 其高峰值出现的胁迫强度越大、不同胁迫期内的释放量差异也越显著; 胁迫期越长, 释放量越低。其中, 草酸、乙酸、乳酸和苹果酸是主要组分, 其质量分数超过了有机酸总量的97.34%, 在修复潜力较强的植物根系的分泌物中还检测到反丁烯二酸。
(4) BaP胁迫对根系分泌物中氨基酸的种类影响不大, 但对分泌量影响较大。苏氨酸、丝氨酸、甘氨酸、丙氨酸的释放量随着胁迫强度的增加而剧增; 脯氨酸、羟脯氨酸和天冬氨酸则近乎以加和效应甚至协同效应的形式参与植物对BaP胁迫的应激反应, 参与响应的组分越多, 表现出的修复潜力越强。
总体上, BaP胁迫环境中, 5种羊茅属植物根系的几种低分子量有机物的释放特征与植物的修复潜力有关, 修复潜力越强, 释放量越多且成分越复杂, 并表现出更强的环境适应性及生理可塑性。
基金项目 国家自然科学基金(31300439)和四川省教育厅自然科学基金(16ZA0385)。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Abhilash PC Powell JR Singh HB Singh BK 2012 ). Plant- microbe interactions: Novel applications for exploitation in multipurpose remediation technologies.
Cell , 30 , 416 -420 .
[本文引用: 2]
[2]
Bertin C Yang X 2003 ). The role of root exudates and allelochemicals in the rhizosphere.
The Plant Soil , 256 , 67 -83 .
[本文引用: 1]
[3]
Chen SC Liao CM 2006 ). Health risk assessment on human exposed to environmental polycyclic aromatic hydrocar- bons pollution sources.
Science of the Total Environment , 366 , 112 -123 .
[本文引用: 1]
[4]
Chen SJ Zhu XL Feng XZ Huang LQ Mei YQ 2012 ). Effect of polycyclic aromatic hydrocarbons (PAHs) on plant.
Bulletin of Biology , 45 (2 ), 9 -11 . (in Chinese)[陈世军 , 祝贤凌 , 冯秀珍 , 黄烈琴 , 梅运群 (2012 ). 多环芳烃对植物的影响
. 生物学通报 , 45 (2 ), 9 -11 .]
[本文引用: 1]
[5]
D’Orazio V Ghanem A Senesi N 2013 ). Phytoremediation of pyrene contaminated soils by different plant species.
Clean-Soil, Air, Water , 41 , 377 -382 .
[本文引用: 1]
[6]
Edwards J Johnson C Santos-Medellín C Lurie E Podishetty NK Bhatnagar S Jonathan AE Sundaresan V 2015 ). Structure, variation, and assembly of the root-associated microbiomes of rice.
Proceedings of the National Academy of Sciences of the United States of America , 112 , 911 -920 .
[本文引用: 1]
[7]
Gao YZ Ling WT Wong MH 2006 ). Plant-accelerated dissipation of phenanthrene and pyrene from water in the presence of a nonionic-surfactant.
Chemosphere , 63 , 1560 -1567 .
[本文引用: 1]
[8]
Gao YZ Ren LL Ling WT Gong SS Sun BQ Zhang Y 2010 ). Desorption of phenanthrene and pyrene in soils by root exudates.
Bioresource Technology , 101 , 1159 -1165 .
[本文引用: 1]
[9]
Howsam M Jones KC Ineson P 2000 ). PAHs associated with the leaves of three decidous tree species. I–Concentration and profile.
Environmental Pollution , 108 , 413 -424 .
[本文引用: 1]
[10]
Hunter PJ Teakle GR Bending GD 2014 ). Root traits and microbial community interactions in relation to phosphorus availability and acquisition, with particular reference to Brassica.
Frontiers Plant Science , 2 , 352 -361 .
[本文引用: 1]
[11]
Jennifer AK Nina MG Frank G 2015 ). Plant-soil interactions in metal contaminated soils.
Soil Biology & Biochemistry , 80 , 232 -236 .
[本文引用: 1]
[12]
Joner EJ Leyval C 2003 ). Rhizosphere gradients of polycyclic aromatic hydrocarbon (PAH) dissipation in two industrial soils and the impact of arbuscular mycorrhiza.
Environm- ental Science & Technology , 37 , 2371 -2375 .
[本文引用: 1]
[13]
Kim YB Park KY Chung Y Buchanan BB 2004 ). Phytorem- ediation of anthracene contaminated soils by different plant species.
Journal of Plant Biology , 47 , 174 -178 .
[本文引用: 1]
[14]
Kirk JL Klironomos JN Lee H Trevors JT 2005 ). The effects of perennial ryegrass and alfalfa on microbial abundance and diversity in petroleum contaminated soil.
Environmental Pollution , 133 , 455 -465 .
[本文引用: 1]
[15]
Lakshmanan V Selvaraj G Bais HP 2014 ). Functional soil microbiome: Belowground solutions to an aboveground problem.
Plant Physiology , 166 , 689 -700 .
[本文引用: 2]
[16]
Li XH Ma LL Liu XF Fu S Cheng HX Xu XB 2006 ). Polycyclic aromatic hydrocarbon in urban soil from Beijing, China.
Journal of Environmental Sciences , 18 , 944 -950 .
[本文引用: 1]
[17]
Line ES Paul HK Torben N Christian K Jørgen S 2003 ). Toxicity of eight polycyclic aromatic compounds to red clover (Trifolium pratense), ryegrass (Lolium perenne) and mustard (Sinapsis alba).
Chemosphere , 53 , 993 -1003 .
[本文引用: 1]
[18]
Liu H Ye YB Cui B Zheng LM Huang YH Wang ZH 2008 ). Responses of Arabidopsis thaliana to oxidative stress induced by polycyclic aromatic hydrocarbon fluoranthene.
Chinese Journal of Applied Ecology , 19 , 413 -418 . (in Chinese with English abstract)[刘泓 , 叶媛蓓 , 崔波 , 郑荔敏 , 黄炎和 , 王宗华 (2008 ). 多环芳烃荧蒽诱导拟南芥氧化胁迫
. 应用生态学报 , 19 , 413 -418 .]
[19]
Liu J Zhou ML Zhang N Chen GP Zhao RR Gao X Shi FC 2015 ). Effects of polycyclic aromatic hydrocarbons (phenanthrene and pyrene) on the growth and physiological cheracteristics of
Spartina atterniflora. Acta Scientialiu Universitatis Nankaiensis , 48 (1 ), 14 -20 . (in Chinese with English abstract)[刘静 , 周美利 , 张楠 , 陈国平 , 赵瑞瑞 , 高鑫 , 石福臣 (2015 ). 多环芳烃菲和芘对互花米草生长和生理特征的影响
. 南开大学学报(自然科学版) , 48 (1 ), 14 -20 .]
[本文引用: 1]
[20]
Liu Y Zhou LL 2011 ). Comparative studies on absorption of benzo (a) pyrene (BaP) in city atmosphere by leaves of 8 street plants. Urban Environment & Urban Ecology
, 24 (4 ), 5 -8 . (in Chinese with English abstract)[刘玉 , 周璐璐 (2011 ). 8种植物叶片对城市大气苯并(a)芘(BaP)的吸收比较
. 城市环境与城市生态 , 24 (4 ), 5 -8 .]
[本文引用: 1]
[21]
Luo L Zhang SZ Shan XQ Zhu YG 2006 ). Oxalate and root exudates enhance the desorption of p,p′-DDT from soils.
Chemosphere , 63 , 1273 -1279 .
[本文引用: 1]
[22]
Maliszewska-Kordybach B Smreczak B 2000 ). Ecotoxico- logical activity of soils polluted with polycyclic aromatic hydrocarbons (PAHs)—Effect on plants.
Environmental Technology , 21 , 1099 -1110 .
[23]
Marschner P Crowley D Yang CH 2004 ). Development of specific rhizosphere bacterial communities in relation to plant species, nutrition and soil type.
The Plant Soil , 261 , 199 -208 .
[本文引用: 1]
[24]
Mueller KE Shann JR 2006 ). PAH dissipation in spiked soil: Impacts of bioavailability, microbial activity, and trees.
Chemosphere , 64 , 1006 -1014 .
[本文引用: 1]
[25]
Pan SW Wei SQ Yuan X Cao SX 2008 ). The removal and remediation of phenanthrene and pyrene in soil by mixed cropping of alfalfa and rape.
Agricultural Sciences in China , 7 , 1355 -1364 .
[本文引用: 3]
[26]
Phillipsa LA Greerb CW Farrella RE Germidaa JJ 2012 ). Plant root exudates impact the hydrocarbon degradation potential of a weathered-hydrocarbon contaminated soil.
Applied Soil Ecology , 52 , 56 -64 .
[本文引用: 1]
[27]
Ramos R Garcia E 2007 ). Induction of mixed-function oxygenase system and antioxidant enzymes in the coral Montastraea faveolata on acute exposure to benzo (a) pyrene.
Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology , 144 , 348 -355 .
[本文引用: 1]
[28]
Shahzad T Chenu C Genet P Barot S Perveen N Mougin C Fontaine S 2015 ). Contribution of exudates, arbuscular mycorrhizal fungi and litter depositions to the rhizosphere priming effect induced by grassland species.
Soil Biology & Biochemistry , 80 , 146 -155 .
[本文引用: 1]
[29]
Shen H Yan XL Zheng SL Wang XR 2002 ). Exudation and accumulation of citric acid in common bean in response to Al toxicity stress.
Chinese Journal of Applied Ecology , 13 , 307 -310 . (in Chinese with English abstract)[沈宏 , 严小龙 , 郑少玲 , 王秀荣 (2002 ). 铝毒胁迫诱导菜豆柠檬酸的分泌与累积
. 应用生态学报 , 13 , 307 -310 .]
[本文引用: 1]
[30]
Sumia K Muhammad A Samina I Qaiser K 2013 ). Plant- bacteria partnerships for the remediation of hydrocarbon contaminated soils.
Chemosphere , 90 , 1317 -1332 .
[本文引用: 1]
[31]
Sun TR Cang L Wang QY Zhou DM Cheng JM Xu H 2010 ). Roles of abiotic losses, microbes, plant roots, and root exudateson phytoremediation of PAHs in a barren soil.
Journal of Hazardous Materials , 176 , 919 -925 .
[本文引用: 1]
[32]
Susarla S Medina VF McCutcheon SC 2002 ). Phytoremedia- tion: An ecological solution to organic chemical contamination.
Ecological Engineering , 18 , 647 -658 .
[本文引用: 1]
[33]
Tao S Cui YH Xu FL Li BG Cao J Liu WX Schmitt G Wang XJ Shen WR Qing BP Sun R 2004 ). Polycyclic aromatic hydrocarbons (PAHs) in agricultural soil and vegetable from Tianjin.
Science of the Total Environment , 320 , 11 -24 .
[本文引用: 1]
[34]
Tejeda-Agredano MC Gallego S Vila J Grifoll M Ortega-Calvo JJ Cantos M 2013 ). Influence of the sunflower rhizosphere on the biodegradation of PAHs in soil.
Soil Biology & Biochemistry , 57 , 2065 -2076 .
[本文引用: 1]
[35]
Tian XX Zhou GY Peng PA 2008 ). Concentrations and influence factors of polycyclic aromatic hydrocarbons in leaves of dominant species in the Pearl River Delta, South China.
Environmental Science , 29 , 849 –854 . (in Chinese with English abstract)[田晓雪 , 周国逸 , 彭平安 (2008). 珠江三角洲地区主要树种叶片多环芳烃含量特征及影响因素分析
. 环境科学 , 2008 , 29 , 849 -854 .]
[本文引用: 1]
[36]
Tu SX Ma L Thomas L 2004 ). Root exudates and arsenic accumulation in arsenic hyperaccumulating Pteris vittata and non-hyperaccumulating Nephrolepis exaltata.
Plant and Soil , 258 , 9 -19 .
[本文引用: 1]
[37]
Wang HC Hu LL Li M Chen WF Wang Y Zhou JJ 2013 ). Growth effects and accumulations of polycyclic aromatic hydrocarbons (PAHs) in rape.
Chinese Journal of Plant Ecology , 37 , 1123 -1131 . (in Chinese with English abstract)[王海翠 , 胡林林 , 李敏 , 陈为峰 , 王莹 , 周佳佳 (2013 ). 多环芳烃(PAHs)对油菜生长的影响及其积累效应
. 植物生态学报 , 37 , 1123 -1131 .]
[本文引用: 1]
[38]
Wei SQ Pan SW 2010 ). Phytoremediation for soils contami- nated by phenanthrene and pyrene with multiple plant species.
Journal of Soils and Sediments , 10 , 886 -894 .
[本文引用: 2]
[39]
Wu LK Lin XM Lin WX 2014 ). Advances and perspective in research on plant-soil-microbe interactions mediated by root exudates.
Chinese Journal of Plant Ecology , 38 , 298 -310 . (in Chinese with English abstract)[吴林坤 , 林向民 , 林文雄 (2014 ). 根系分泌物介导下植物-土壤-微生物互作关系研究进展与展望
. 植物生态学报 , 38 , 298 -310 .]
[本文引用: 2]
[40]
Xie XM Liao M Yang J 2011 ).Effects of pyrene on low molecule weight organic compounds in the root exudates of ryegrass (Lolium perenne L).
Acta Ecologica Sinica , 31 , 7564 -7570 . (in Chinese with English abstract)[谢晓梅 , 廖敏 , 杨静 (2011 ). 芘对黑麦草根系几种低分子量有机分泌物的影响
. 生态学报 , 31 , 7564 -7570 .]
[41]
Xie XM Liao M Yang J Chai JJ Fang S Wang RH 2012 ). Influence of root-exudates concentration on pyrene degradation and soil microbial characteristics in pyrene contaminated soil.
Chemosphere , 88 , 1190 -1195 .
[本文引用: 1]
[42]
Xu C Lin XF Xia BC 2010 ). Response of root exudates of maize seedlings (Zea mays L.) to pyrene contamination.
Acta Ecologica Sinica , 30 , 3280 -3288 . (in Chinese with English abstract)[许超 , 林小方 , 夏北成 (2010 ). 玉米幼苗根系分泌物对芘污染的响应
. 生态学报 , 30 , 3280 -3288 .]
[本文引用: 3]
[43]
Yi H Crowley DE 2007 ). Biostimulation of PAH degradation with plants containing high concentrations of linoleic acid.
Environmental Science & Technology , 41 , 4382 -4388 .
[44]
Zhu YH Zhang SZ Huang HL Wen B 2009 ). Effects of maize root exudates and organic acids on the desorption of phenanthrene from soils.
Journal of Environmental Sci- ences , 21 , 920 -926 .
[本文引用: 1]
Plant- microbe interactions: Novel applications for exploitation in multipurpose remediation technologies.
2
2012
... 植物的存在能加快土壤中PAHs的去除(Joner & Leyval, 2003 ; Kim et al ., 2004 ; Gao et al ., 2006 ), 根系分泌物(root exudates)营造的根际微生态特征则是影响PAHs快速消解的重要原因(Sumia et al ., 2013 ; Tejeda-Agredano et al ., 2013 ).植物修复过程中, 根系分泌物在改善土壤属性、提高污染物的生物可利用性、缓解环境胁迫等方面具有重要作用(Bertin & Yang, 2003 ; Phillipsa et al ., 2012 ; Shahzad et al ., 2015 ).一方面, 根系分泌的某些胞外酶能够直接参与PAHs的降解过程; 另一方面, 分泌物中的可溶性糖、低分子量有机酸、氨基酸等物质, 为根际微生物提供充足的营养, 增加根际微生物的数量、提高微生物活性, 进而加速PAHs降解(Susarla et al ., 2002 ; Xie et al ., 2012 ; Lakshmanan et al ., 2014 ).同时, 根系分泌物还会直接影响PAHs的固定和活化, 进而影响PAHs在土壤-植物系统中的迁移转化行为、促进PAHs去除(Marschner et al ., 2004 ; Abhilash et al ., 2012 ); PAHs的环数越多, 根际效益越明显(Sun et al ., 2010 ).不同植物间, 因根系分泌物的组成成分及其释放特征常存在一定差异, 对根际微生物的区系结构、代谢活性的影响也不一样, 进而表现出不同的修复潜力(Kirk et al ., 2005 ; Luo et al ., 2006 ; Hunter et al ., 2014 ).因此, 探讨PAHs胁迫下不同修复潜力植物间根系分泌物中主要组分的释放特征及其种间差异, 对于研究植物修复的生物学机制、揭示不同植物间修复潜力的种间差异具有重要意义.鉴于此, 本研究以对PAHs污染土壤具有一定修复潜力的羊茅属(Festuca )植物为试验对象, 以苯并[α]芘(BaP)为PAHs代表物, 借助根际袋土培法, 对比研究PAHs胁迫下不同修复潜力的5种羊茅属植物在不同生长期时根系分泌物中几种低分子量有机物的释放特征, 为植物修复理论的研究与实践提供依据. ...
... 在土壤中, 根系分泌物与根际微生物间的相互作用是一个十分重要的过程: 根系通过分泌各种次生代谢物质对根际微生物的种类、数量和分布产生影响, 对根际微生物群落结构有选择塑造作用, 进而影响PAHs的降解(Abhilash et al ., 2012 ; Edwards et al ., 2015 ).本研究显示, 胁迫环境中植物根系可溶性糖、低分子量有机酸、氨基酸等组分的释放特征与植物自身的修复潜力有关: 修复潜力较强的苇状羊茅所分泌的可溶性糖、低分子量有机酸、氨基酸总多于其他几种植物; 修复潜力越强, 其分泌量、组成成分的变化幅度越大.相同胁迫条件下, 修复潜力较强的植物对BaP的胁迫响应总表现出更强的环境适应性、生理活性上的可塑性, 修复潜力越强, 趋势越明显(图3 -5).可以推测, 根系分泌物的释放特征势必影响根际微生物群落结构, 并产生选择性塑造作用, 但这种释放特征是植物为减轻BaP毒害而进行的自我生理调节还是BaP毒害破坏了根系正常的生理活动而造成细胞内有机物质的渗漏尚待进一步研究. ...
The role of root exudates and allelochemicals in the rhizosphere.
1
2003
... 植物的存在能加快土壤中PAHs的去除(Joner & Leyval, 2003 ; Kim et al ., 2004 ; Gao et al ., 2006 ), 根系分泌物(root exudates)营造的根际微生态特征则是影响PAHs快速消解的重要原因(Sumia et al ., 2013 ; Tejeda-Agredano et al ., 2013 ).植物修复过程中, 根系分泌物在改善土壤属性、提高污染物的生物可利用性、缓解环境胁迫等方面具有重要作用(Bertin & Yang, 2003 ; Phillipsa et al ., 2012 ; Shahzad et al ., 2015 ).一方面, 根系分泌的某些胞外酶能够直接参与PAHs的降解过程; 另一方面, 分泌物中的可溶性糖、低分子量有机酸、氨基酸等物质, 为根际微生物提供充足的营养, 增加根际微生物的数量、提高微生物活性, 进而加速PAHs降解(Susarla et al ., 2002 ; Xie et al ., 2012 ; Lakshmanan et al ., 2014 ).同时, 根系分泌物还会直接影响PAHs的固定和活化, 进而影响PAHs在土壤-植物系统中的迁移转化行为、促进PAHs去除(Marschner et al ., 2004 ; Abhilash et al ., 2012 ); PAHs的环数越多, 根际效益越明显(Sun et al ., 2010 ).不同植物间, 因根系分泌物的组成成分及其释放特征常存在一定差异, 对根际微生物的区系结构、代谢活性的影响也不一样, 进而表现出不同的修复潜力(Kirk et al ., 2005 ; Luo et al ., 2006 ; Hunter et al ., 2014 ).因此, 探讨PAHs胁迫下不同修复潜力植物间根系分泌物中主要组分的释放特征及其种间差异, 对于研究植物修复的生物学机制、揭示不同植物间修复潜力的种间差异具有重要意义.鉴于此, 本研究以对PAHs污染土壤具有一定修复潜力的羊茅属(Festuca )植物为试验对象, 以苯并[α]芘(BaP)为PAHs代表物, 借助根际袋土培法, 对比研究PAHs胁迫下不同修复潜力的5种羊茅属植物在不同生长期时根系分泌物中几种低分子量有机物的释放特征, 为植物修复理论的研究与实践提供依据. ...
Health risk assessment on human exposed to environmental polycyclic aromatic hydrocar- bons pollution sources.
1
2006
... 多环芳烃(polycyclic aromatic hydrocarbons, PAHs)是环境中普遍存在的持久性有机污染物, 多数具有致癌、致畸或致突变效应(Chen & Liao, 2006 ).因其性质稳定、水溶性差, 易于被土壤颗粒吸附, 土壤中PAHs含量逐年上升(Tao et al ., 2004 ; Li et al ., 2006 ), 严重威胁着土壤环境质量及农产品质量安全; 土壤中PAHs的迁移转化行为及其环境效应已成为环境修复领域的研究热点(Mueller & Shann, 2006 ; Lakshmanan et al ., 2014 ). ...
多环芳烃对植物的影响
1
2012
... 植物生长指标的变化是植物受到不同程度胁迫伤害的直观表现症状(王海翠等, 2013 ).本试验中, 低度(T1 -T2 )或中度(T3 -T4 ) BaP胁迫刺激了植物生长, T4 水平时生物量出现峰值, 重度(T5 )胁迫则抑制了植物生长.通常, 多数植物对PAHs类环境胁迫具有一定的耐受域值, 而这种域值与PAHs的理化性质、植物种类和环境状况(Howsam et al ., 2000 ; Line et al ., 2003 ; 田晓雪等, 2008 )以及PAHs进入细胞后的累积或转化机制等因素有关(刘玉和周璐璐, 2011 ; 刘静等, 2015 ).一定域值内, 植物生长受到的抑制影响并不显著, 甚至能刺激植物生长; 超过这一域值, 植物生长就会受到抑制.Maliszewska-Kordy- bach和Smreczak (2000)报道, 由芴、蒽、芘和䓛4种PAHs组成的混合物, 在浓度不超过10 mg·kg-1 时促进了小麦(Triticum aestivum )、玉米(Zea mays )及燕麦(Avena sativa )的生长, 高浓度时抑制生长; 刘泓等(2008)研究了不同浓度的荧蒽培养基对拟南芥(Arabidopsis thaliana )生长的影响, 发现不同浓度的荧蒽胁迫30天后拟南芥生长量明显受到不同程度的抑制.尽管这些研究都表明, 高浓度PAHs胁迫抑制植物的生长, 但抑制作用机制尚不明确; 低浓度的PAHs对植物生长的促进作用, 有学者推测是因为PAHs具有与植物生长激素类似的作用(陈世军等, 2012 ), 但也仅仅局限于推测, 还没有被试验和实践证实. ...
Phytoremediation of pyrene contaminated soils by different plant species.
1
2013
... 植物根际对PAHs的降解效果与修复植物的种类有关.Yi和Crowley (2007)研究43种植物根部组织对PAHs的降解作用发现, 有些植物对PAHs降解有明显的促进作用, 有些则无作用或有抑制作用.根系分泌物所营造的根际环境及其所呈现的根际效应是植物响应外界胁迫的重要途径, 也是影响修复效果的关键因素(Gao et al ., 2010 ; 吴林坤等, 2014 ).相同胁迫条件下, 根系分泌物的释放特征及其主要组分的种间差异主要处决于植物自身基因型的特异性及其生理状态的个体差异(Zhu et al ., 2009 ; D’Orazio et al ., 2013 ).本研究以亲缘关系较近的5种羊茅属植物为实验对象, 其基因型差异相对较小, 在一定程度上能增加研究结果的可比性.尽管待试植物在无污染(CK)、轻度或中度污染(T1 -T4 )时的生物量, 以及被植物积累的BaP量(0.08%-0.21%, 平均(0.15 ± 0.02)%差异甚微, 但它们对BaP污染土壤的修复潜力却存在显著的种间差异.可见, 植物修复过程中, 即使是遗传关系很近的同属植物间, 修复潜力的种间差异仍然存在. ...
Structure, variation, and assembly of the root-associated microbiomes of rice.
1
2015
... 在土壤中, 根系分泌物与根际微生物间的相互作用是一个十分重要的过程: 根系通过分泌各种次生代谢物质对根际微生物的种类、数量和分布产生影响, 对根际微生物群落结构有选择塑造作用, 进而影响PAHs的降解(Abhilash et al ., 2012 ; Edwards et al ., 2015 ).本研究显示, 胁迫环境中植物根系可溶性糖、低分子量有机酸、氨基酸等组分的释放特征与植物自身的修复潜力有关: 修复潜力较强的苇状羊茅所分泌的可溶性糖、低分子量有机酸、氨基酸总多于其他几种植物; 修复潜力越强, 其分泌量、组成成分的变化幅度越大.相同胁迫条件下, 修复潜力较强的植物对BaP的胁迫响应总表现出更强的环境适应性、生理活性上的可塑性, 修复潜力越强, 趋势越明显(图3 -5).可以推测, 根系分泌物的释放特征势必影响根际微生物群落结构, 并产生选择性塑造作用, 但这种释放特征是植物为减轻BaP毒害而进行的自我生理调节还是BaP毒害破坏了根系正常的生理活动而造成细胞内有机物质的渗漏尚待进一步研究. ...
Plant-accelerated dissipation of phenanthrene and pyrene from water in the presence of a nonionic-surfactant.
1
2006
... 植物的存在能加快土壤中PAHs的去除(Joner & Leyval, 2003 ; Kim et al ., 2004 ; Gao et al ., 2006 ), 根系分泌物(root exudates)营造的根际微生态特征则是影响PAHs快速消解的重要原因(Sumia et al ., 2013 ; Tejeda-Agredano et al ., 2013 ).植物修复过程中, 根系分泌物在改善土壤属性、提高污染物的生物可利用性、缓解环境胁迫等方面具有重要作用(Bertin & Yang, 2003 ; Phillipsa et al ., 2012 ; Shahzad et al ., 2015 ).一方面, 根系分泌的某些胞外酶能够直接参与PAHs的降解过程; 另一方面, 分泌物中的可溶性糖、低分子量有机酸、氨基酸等物质, 为根际微生物提供充足的营养, 增加根际微生物的数量、提高微生物活性, 进而加速PAHs降解(Susarla et al ., 2002 ; Xie et al ., 2012 ; Lakshmanan et al ., 2014 ).同时, 根系分泌物还会直接影响PAHs的固定和活化, 进而影响PAHs在土壤-植物系统中的迁移转化行为、促进PAHs去除(Marschner et al ., 2004 ; Abhilash et al ., 2012 ); PAHs的环数越多, 根际效益越明显(Sun et al ., 2010 ).不同植物间, 因根系分泌物的组成成分及其释放特征常存在一定差异, 对根际微生物的区系结构、代谢活性的影响也不一样, 进而表现出不同的修复潜力(Kirk et al ., 2005 ; Luo et al ., 2006 ; Hunter et al ., 2014 ).因此, 探讨PAHs胁迫下不同修复潜力植物间根系分泌物中主要组分的释放特征及其种间差异, 对于研究植物修复的生物学机制、揭示不同植物间修复潜力的种间差异具有重要意义.鉴于此, 本研究以对PAHs污染土壤具有一定修复潜力的羊茅属(Festuca )植物为试验对象, 以苯并[α]芘(BaP)为PAHs代表物, 借助根际袋土培法, 对比研究PAHs胁迫下不同修复潜力的5种羊茅属植物在不同生长期时根系分泌物中几种低分子量有机物的释放特征, 为植物修复理论的研究与实践提供依据. ...
Desorption of phenanthrene and pyrene in soils by root exudates.
1
2010
... 植物根际对PAHs的降解效果与修复植物的种类有关.Yi和Crowley (2007)研究43种植物根部组织对PAHs的降解作用发现, 有些植物对PAHs降解有明显的促进作用, 有些则无作用或有抑制作用.根系分泌物所营造的根际环境及其所呈现的根际效应是植物响应外界胁迫的重要途径, 也是影响修复效果的关键因素(Gao et al ., 2010 ; 吴林坤等, 2014 ).相同胁迫条件下, 根系分泌物的释放特征及其主要组分的种间差异主要处决于植物自身基因型的特异性及其生理状态的个体差异(Zhu et al ., 2009 ; D’Orazio et al ., 2013 ).本研究以亲缘关系较近的5种羊茅属植物为实验对象, 其基因型差异相对较小, 在一定程度上能增加研究结果的可比性.尽管待试植物在无污染(CK)、轻度或中度污染(T1 -T4 )时的生物量, 以及被植物积累的BaP量(0.08%-0.21%, 平均(0.15 ± 0.02)%差异甚微, 但它们对BaP污染土壤的修复潜力却存在显著的种间差异.可见, 植物修复过程中, 即使是遗传关系很近的同属植物间, 修复潜力的种间差异仍然存在. ...
PAHs associated with the leaves of three decidous tree species. I–Concentration and profile.
1
2000
... 植物生长指标的变化是植物受到不同程度胁迫伤害的直观表现症状(王海翠等, 2013 ).本试验中, 低度(T1 -T2 )或中度(T3 -T4 ) BaP胁迫刺激了植物生长, T4 水平时生物量出现峰值, 重度(T5 )胁迫则抑制了植物生长.通常, 多数植物对PAHs类环境胁迫具有一定的耐受域值, 而这种域值与PAHs的理化性质、植物种类和环境状况(Howsam et al ., 2000 ; Line et al ., 2003 ; 田晓雪等, 2008 )以及PAHs进入细胞后的累积或转化机制等因素有关(刘玉和周璐璐, 2011 ; 刘静等, 2015 ).一定域值内, 植物生长受到的抑制影响并不显著, 甚至能刺激植物生长; 超过这一域值, 植物生长就会受到抑制.Maliszewska-Kordy- bach和Smreczak (2000)报道, 由芴、蒽、芘和䓛4种PAHs组成的混合物, 在浓度不超过10 mg·kg-1 时促进了小麦(Triticum aestivum )、玉米(Zea mays )及燕麦(Avena sativa )的生长, 高浓度时抑制生长; 刘泓等(2008)研究了不同浓度的荧蒽培养基对拟南芥(Arabidopsis thaliana )生长的影响, 发现不同浓度的荧蒽胁迫30天后拟南芥生长量明显受到不同程度的抑制.尽管这些研究都表明, 高浓度PAHs胁迫抑制植物的生长, 但抑制作用机制尚不明确; 低浓度的PAHs对植物生长的促进作用, 有学者推测是因为PAHs具有与植物生长激素类似的作用(陈世军等, 2012 ), 但也仅仅局限于推测, 还没有被试验和实践证实. ...
Root traits and microbial community interactions in relation to phosphorus availability and acquisition, with particular reference to Brassica.
1
2014
... 植物的存在能加快土壤中PAHs的去除(Joner & Leyval, 2003 ; Kim et al ., 2004 ; Gao et al ., 2006 ), 根系分泌物(root exudates)营造的根际微生态特征则是影响PAHs快速消解的重要原因(Sumia et al ., 2013 ; Tejeda-Agredano et al ., 2013 ).植物修复过程中, 根系分泌物在改善土壤属性、提高污染物的生物可利用性、缓解环境胁迫等方面具有重要作用(Bertin & Yang, 2003 ; Phillipsa et al ., 2012 ; Shahzad et al ., 2015 ).一方面, 根系分泌的某些胞外酶能够直接参与PAHs的降解过程; 另一方面, 分泌物中的可溶性糖、低分子量有机酸、氨基酸等物质, 为根际微生物提供充足的营养, 增加根际微生物的数量、提高微生物活性, 进而加速PAHs降解(Susarla et al ., 2002 ; Xie et al ., 2012 ; Lakshmanan et al ., 2014 ).同时, 根系分泌物还会直接影响PAHs的固定和活化, 进而影响PAHs在土壤-植物系统中的迁移转化行为、促进PAHs去除(Marschner et al ., 2004 ; Abhilash et al ., 2012 ); PAHs的环数越多, 根际效益越明显(Sun et al ., 2010 ).不同植物间, 因根系分泌物的组成成分及其释放特征常存在一定差异, 对根际微生物的区系结构、代谢活性的影响也不一样, 进而表现出不同的修复潜力(Kirk et al ., 2005 ; Luo et al ., 2006 ; Hunter et al ., 2014 ).因此, 探讨PAHs胁迫下不同修复潜力植物间根系分泌物中主要组分的释放特征及其种间差异, 对于研究植物修复的生物学机制、揭示不同植物间修复潜力的种间差异具有重要意义.鉴于此, 本研究以对PAHs污染土壤具有一定修复潜力的羊茅属(Festuca )植物为试验对象, 以苯并[α]芘(BaP)为PAHs代表物, 借助根际袋土培法, 对比研究PAHs胁迫下不同修复潜力的5种羊茅属植物在不同生长期时根系分泌物中几种低分子量有机物的释放特征, 为植物修复理论的研究与实践提供依据. ...
Plant-soil interactions in metal contaminated soils.
1
2015
... 根系分泌物的释放特征是植物根系胁迫反应时的集中表现(Jennifer et al ., 2015 ).BaP胁迫促进了5种羊茅属植物对苏氨酸、丝氨酸、甘氨酸、丙氨酸的释放, 许超等(2010)、谢晓梅等(2011)等在研究芘胁迫对玉米、黑麦草根系分泌物的影响时也得出类似结论.本试验中, 根系分泌物中脯氨酸、羟脯氨酸和天冬氨酸等组分对BaP的胁迫响应更近乎以加和效应(additivity)甚至协同效应(synergism)形式参与植物的修复过程, 参与应激反应的组分越多, 修复潜力越强: 修复潜力最强的苇状羊茅根系以上3种组分的释放量均随胁迫强度的增强而剧增; 修复潜力次强的草原羊茅、毛稃羊茅根系上述组分中某2种的释放量随胁迫强度的增强而剧增; 修复潜力次差的贫芒羊茅根系仅上述组分中某1种的释放量随胁迫强度的增强而剧增; 修复潜力最差的细芒羊茅根系上述组分的释放量与BaP胁迫强度无明显相关性.但该响应模式是否具有普遍性, 尚待进一步验证. ...
Rhizosphere gradients of polycyclic aromatic hydrocarbon (PAH) dissipation in two industrial soils and the impact of arbuscular mycorrhiza.
1
2003
... 植物的存在能加快土壤中PAHs的去除(Joner & Leyval, 2003 ; Kim et al ., 2004 ; Gao et al ., 2006 ), 根系分泌物(root exudates)营造的根际微生态特征则是影响PAHs快速消解的重要原因(Sumia et al ., 2013 ; Tejeda-Agredano et al ., 2013 ).植物修复过程中, 根系分泌物在改善土壤属性、提高污染物的生物可利用性、缓解环境胁迫等方面具有重要作用(Bertin & Yang, 2003 ; Phillipsa et al ., 2012 ; Shahzad et al ., 2015 ).一方面, 根系分泌的某些胞外酶能够直接参与PAHs的降解过程; 另一方面, 分泌物中的可溶性糖、低分子量有机酸、氨基酸等物质, 为根际微生物提供充足的营养, 增加根际微生物的数量、提高微生物活性, 进而加速PAHs降解(Susarla et al ., 2002 ; Xie et al ., 2012 ; Lakshmanan et al ., 2014 ).同时, 根系分泌物还会直接影响PAHs的固定和活化, 进而影响PAHs在土壤-植物系统中的迁移转化行为、促进PAHs去除(Marschner et al ., 2004 ; Abhilash et al ., 2012 ); PAHs的环数越多, 根际效益越明显(Sun et al ., 2010 ).不同植物间, 因根系分泌物的组成成分及其释放特征常存在一定差异, 对根际微生物的区系结构、代谢活性的影响也不一样, 进而表现出不同的修复潜力(Kirk et al ., 2005 ; Luo et al ., 2006 ; Hunter et al ., 2014 ).因此, 探讨PAHs胁迫下不同修复潜力植物间根系分泌物中主要组分的释放特征及其种间差异, 对于研究植物修复的生物学机制、揭示不同植物间修复潜力的种间差异具有重要意义.鉴于此, 本研究以对PAHs污染土壤具有一定修复潜力的羊茅属(Festuca )植物为试验对象, 以苯并[α]芘(BaP)为PAHs代表物, 借助根际袋土培法, 对比研究PAHs胁迫下不同修复潜力的5种羊茅属植物在不同生长期时根系分泌物中几种低分子量有机物的释放特征, 为植物修复理论的研究与实践提供依据. ...
Phytorem- ediation of anthracene contaminated soils by different plant species.
1
2004
... 植物的存在能加快土壤中PAHs的去除(Joner & Leyval, 2003 ; Kim et al ., 2004 ; Gao et al ., 2006 ), 根系分泌物(root exudates)营造的根际微生态特征则是影响PAHs快速消解的重要原因(Sumia et al ., 2013 ; Tejeda-Agredano et al ., 2013 ).植物修复过程中, 根系分泌物在改善土壤属性、提高污染物的生物可利用性、缓解环境胁迫等方面具有重要作用(Bertin & Yang, 2003 ; Phillipsa et al ., 2012 ; Shahzad et al ., 2015 ).一方面, 根系分泌的某些胞外酶能够直接参与PAHs的降解过程; 另一方面, 分泌物中的可溶性糖、低分子量有机酸、氨基酸等物质, 为根际微生物提供充足的营养, 增加根际微生物的数量、提高微生物活性, 进而加速PAHs降解(Susarla et al ., 2002 ; Xie et al ., 2012 ; Lakshmanan et al ., 2014 ).同时, 根系分泌物还会直接影响PAHs的固定和活化, 进而影响PAHs在土壤-植物系统中的迁移转化行为、促进PAHs去除(Marschner et al ., 2004 ; Abhilash et al ., 2012 ); PAHs的环数越多, 根际效益越明显(Sun et al ., 2010 ).不同植物间, 因根系分泌物的组成成分及其释放特征常存在一定差异, 对根际微生物的区系结构、代谢活性的影响也不一样, 进而表现出不同的修复潜力(Kirk et al ., 2005 ; Luo et al ., 2006 ; Hunter et al ., 2014 ).因此, 探讨PAHs胁迫下不同修复潜力植物间根系分泌物中主要组分的释放特征及其种间差异, 对于研究植物修复的生物学机制、揭示不同植物间修复潜力的种间差异具有重要意义.鉴于此, 本研究以对PAHs污染土壤具有一定修复潜力的羊茅属(Festuca )植物为试验对象, 以苯并[α]芘(BaP)为PAHs代表物, 借助根际袋土培法, 对比研究PAHs胁迫下不同修复潜力的5种羊茅属植物在不同生长期时根系分泌物中几种低分子量有机物的释放特征, 为植物修复理论的研究与实践提供依据. ...
The effects of perennial ryegrass and alfalfa on microbial abundance and diversity in petroleum contaminated soil.
1
2005
... 植物的存在能加快土壤中PAHs的去除(Joner & Leyval, 2003 ; Kim et al ., 2004 ; Gao et al ., 2006 ), 根系分泌物(root exudates)营造的根际微生态特征则是影响PAHs快速消解的重要原因(Sumia et al ., 2013 ; Tejeda-Agredano et al ., 2013 ).植物修复过程中, 根系分泌物在改善土壤属性、提高污染物的生物可利用性、缓解环境胁迫等方面具有重要作用(Bertin & Yang, 2003 ; Phillipsa et al ., 2012 ; Shahzad et al ., 2015 ).一方面, 根系分泌的某些胞外酶能够直接参与PAHs的降解过程; 另一方面, 分泌物中的可溶性糖、低分子量有机酸、氨基酸等物质, 为根际微生物提供充足的营养, 增加根际微生物的数量、提高微生物活性, 进而加速PAHs降解(Susarla et al ., 2002 ; Xie et al ., 2012 ; Lakshmanan et al ., 2014 ).同时, 根系分泌物还会直接影响PAHs的固定和活化, 进而影响PAHs在土壤-植物系统中的迁移转化行为、促进PAHs去除(Marschner et al ., 2004 ; Abhilash et al ., 2012 ); PAHs的环数越多, 根际效益越明显(Sun et al ., 2010 ).不同植物间, 因根系分泌物的组成成分及其释放特征常存在一定差异, 对根际微生物的区系结构、代谢活性的影响也不一样, 进而表现出不同的修复潜力(Kirk et al ., 2005 ; Luo et al ., 2006 ; Hunter et al ., 2014 ).因此, 探讨PAHs胁迫下不同修复潜力植物间根系分泌物中主要组分的释放特征及其种间差异, 对于研究植物修复的生物学机制、揭示不同植物间修复潜力的种间差异具有重要意义.鉴于此, 本研究以对PAHs污染土壤具有一定修复潜力的羊茅属(Festuca )植物为试验对象, 以苯并[α]芘(BaP)为PAHs代表物, 借助根际袋土培法, 对比研究PAHs胁迫下不同修复潜力的5种羊茅属植物在不同生长期时根系分泌物中几种低分子量有机物的释放特征, 为植物修复理论的研究与实践提供依据. ...
Functional soil microbiome: Belowground solutions to an aboveground problem.
2
2014
... 多环芳烃(polycyclic aromatic hydrocarbons, PAHs)是环境中普遍存在的持久性有机污染物, 多数具有致癌、致畸或致突变效应(Chen & Liao, 2006 ).因其性质稳定、水溶性差, 易于被土壤颗粒吸附, 土壤中PAHs含量逐年上升(Tao et al ., 2004 ; Li et al ., 2006 ), 严重威胁着土壤环境质量及农产品质量安全; 土壤中PAHs的迁移转化行为及其环境效应已成为环境修复领域的研究热点(Mueller & Shann, 2006 ; Lakshmanan et al ., 2014 ). ...
... 植物的存在能加快土壤中PAHs的去除(Joner & Leyval, 2003 ; Kim et al ., 2004 ; Gao et al ., 2006 ), 根系分泌物(root exudates)营造的根际微生态特征则是影响PAHs快速消解的重要原因(Sumia et al ., 2013 ; Tejeda-Agredano et al ., 2013 ).植物修复过程中, 根系分泌物在改善土壤属性、提高污染物的生物可利用性、缓解环境胁迫等方面具有重要作用(Bertin & Yang, 2003 ; Phillipsa et al ., 2012 ; Shahzad et al ., 2015 ).一方面, 根系分泌的某些胞外酶能够直接参与PAHs的降解过程; 另一方面, 分泌物中的可溶性糖、低分子量有机酸、氨基酸等物质, 为根际微生物提供充足的营养, 增加根际微生物的数量、提高微生物活性, 进而加速PAHs降解(Susarla et al ., 2002 ; Xie et al ., 2012 ; Lakshmanan et al ., 2014 ).同时, 根系分泌物还会直接影响PAHs的固定和活化, 进而影响PAHs在土壤-植物系统中的迁移转化行为、促进PAHs去除(Marschner et al ., 2004 ; Abhilash et al ., 2012 ); PAHs的环数越多, 根际效益越明显(Sun et al ., 2010 ).不同植物间, 因根系分泌物的组成成分及其释放特征常存在一定差异, 对根际微生物的区系结构、代谢活性的影响也不一样, 进而表现出不同的修复潜力(Kirk et al ., 2005 ; Luo et al ., 2006 ; Hunter et al ., 2014 ).因此, 探讨PAHs胁迫下不同修复潜力植物间根系分泌物中主要组分的释放特征及其种间差异, 对于研究植物修复的生物学机制、揭示不同植物间修复潜力的种间差异具有重要意义.鉴于此, 本研究以对PAHs污染土壤具有一定修复潜力的羊茅属(Festuca )植物为试验对象, 以苯并[α]芘(BaP)为PAHs代表物, 借助根际袋土培法, 对比研究PAHs胁迫下不同修复潜力的5种羊茅属植物在不同生长期时根系分泌物中几种低分子量有机物的释放特征, 为植物修复理论的研究与实践提供依据. ...
Polycyclic aromatic hydrocarbon in urban soil from Beijing, China.
1
2006
... 多环芳烃(polycyclic aromatic hydrocarbons, PAHs)是环境中普遍存在的持久性有机污染物, 多数具有致癌、致畸或致突变效应(Chen & Liao, 2006 ).因其性质稳定、水溶性差, 易于被土壤颗粒吸附, 土壤中PAHs含量逐年上升(Tao et al ., 2004 ; Li et al ., 2006 ), 严重威胁着土壤环境质量及农产品质量安全; 土壤中PAHs的迁移转化行为及其环境效应已成为环境修复领域的研究热点(Mueller & Shann, 2006 ; Lakshmanan et al ., 2014 ). ...
Toxicity of eight polycyclic aromatic compounds to red clover (Trifolium pratense), ryegrass (Lolium perenne) and mustard (Sinapsis alba).
1
2003
... 植物生长指标的变化是植物受到不同程度胁迫伤害的直观表现症状(王海翠等, 2013 ).本试验中, 低度(T1 -T2 )或中度(T3 -T4 ) BaP胁迫刺激了植物生长, T4 水平时生物量出现峰值, 重度(T5 )胁迫则抑制了植物生长.通常, 多数植物对PAHs类环境胁迫具有一定的耐受域值, 而这种域值与PAHs的理化性质、植物种类和环境状况(Howsam et al ., 2000 ; Line et al ., 2003 ; 田晓雪等, 2008 )以及PAHs进入细胞后的累积或转化机制等因素有关(刘玉和周璐璐, 2011 ; 刘静等, 2015 ).一定域值内, 植物生长受到的抑制影响并不显著, 甚至能刺激植物生长; 超过这一域值, 植物生长就会受到抑制.Maliszewska-Kordy- bach和Smreczak (2000)报道, 由芴、蒽、芘和䓛4种PAHs组成的混合物, 在浓度不超过10 mg·kg-1 时促进了小麦(Triticum aestivum )、玉米(Zea mays )及燕麦(Avena sativa )的生长, 高浓度时抑制生长; 刘泓等(2008)研究了不同浓度的荧蒽培养基对拟南芥(Arabidopsis thaliana )生长的影响, 发现不同浓度的荧蒽胁迫30天后拟南芥生长量明显受到不同程度的抑制.尽管这些研究都表明, 高浓度PAHs胁迫抑制植物的生长, 但抑制作用机制尚不明确; 低浓度的PAHs对植物生长的促进作用, 有学者推测是因为PAHs具有与植物生长激素类似的作用(陈世军等, 2012 ), 但也仅仅局限于推测, 还没有被试验和实践证实. ...
多环芳烃菲和芘对互花米草生长和生理特征的影响
1
2015
... 植物生长指标的变化是植物受到不同程度胁迫伤害的直观表现症状(王海翠等, 2013 ).本试验中, 低度(T1 -T2 )或中度(T3 -T4 ) BaP胁迫刺激了植物生长, T4 水平时生物量出现峰值, 重度(T5 )胁迫则抑制了植物生长.通常, 多数植物对PAHs类环境胁迫具有一定的耐受域值, 而这种域值与PAHs的理化性质、植物种类和环境状况(Howsam et al ., 2000 ; Line et al ., 2003 ; 田晓雪等, 2008 )以及PAHs进入细胞后的累积或转化机制等因素有关(刘玉和周璐璐, 2011 ; 刘静等, 2015 ).一定域值内, 植物生长受到的抑制影响并不显著, 甚至能刺激植物生长; 超过这一域值, 植物生长就会受到抑制.Maliszewska-Kordy- bach和Smreczak (2000)报道, 由芴、蒽、芘和䓛4种PAHs组成的混合物, 在浓度不超过10 mg·kg-1 时促进了小麦(Triticum aestivum )、玉米(Zea mays )及燕麦(Avena sativa )的生长, 高浓度时抑制生长; 刘泓等(2008)研究了不同浓度的荧蒽培养基对拟南芥(Arabidopsis thaliana )生长的影响, 发现不同浓度的荧蒽胁迫30天后拟南芥生长量明显受到不同程度的抑制.尽管这些研究都表明, 高浓度PAHs胁迫抑制植物的生长, 但抑制作用机制尚不明确; 低浓度的PAHs对植物生长的促进作用, 有学者推测是因为PAHs具有与植物生长激素类似的作用(陈世军等, 2012 ), 但也仅仅局限于推测, 还没有被试验和实践证实. ...
8种植物叶片对城市大气苯并(a)芘(BaP)的吸收比较
1
2011
... 植物生长指标的变化是植物受到不同程度胁迫伤害的直观表现症状(王海翠等, 2013 ).本试验中, 低度(T1 -T2 )或中度(T3 -T4 ) BaP胁迫刺激了植物生长, T4 水平时生物量出现峰值, 重度(T5 )胁迫则抑制了植物生长.通常, 多数植物对PAHs类环境胁迫具有一定的耐受域值, 而这种域值与PAHs的理化性质、植物种类和环境状况(Howsam et al ., 2000 ; Line et al ., 2003 ; 田晓雪等, 2008 )以及PAHs进入细胞后的累积或转化机制等因素有关(刘玉和周璐璐, 2011 ; 刘静等, 2015 ).一定域值内, 植物生长受到的抑制影响并不显著, 甚至能刺激植物生长; 超过这一域值, 植物生长就会受到抑制.Maliszewska-Kordy- bach和Smreczak (2000)报道, 由芴、蒽、芘和䓛4种PAHs组成的混合物, 在浓度不超过10 mg·kg-1 时促进了小麦(Triticum aestivum )、玉米(Zea mays )及燕麦(Avena sativa )的生长, 高浓度时抑制生长; 刘泓等(2008)研究了不同浓度的荧蒽培养基对拟南芥(Arabidopsis thaliana )生长的影响, 发现不同浓度的荧蒽胁迫30天后拟南芥生长量明显受到不同程度的抑制.尽管这些研究都表明, 高浓度PAHs胁迫抑制植物的生长, 但抑制作用机制尚不明确; 低浓度的PAHs对植物生长的促进作用, 有学者推测是因为PAHs具有与植物生长激素类似的作用(陈世军等, 2012 ), 但也仅仅局限于推测, 还没有被试验和实践证实. ...
Oxalate and root exudates enhance the desorption of p,p′-DDT from soils.
1
2006
... 植物的存在能加快土壤中PAHs的去除(Joner & Leyval, 2003 ; Kim et al ., 2004 ; Gao et al ., 2006 ), 根系分泌物(root exudates)营造的根际微生态特征则是影响PAHs快速消解的重要原因(Sumia et al ., 2013 ; Tejeda-Agredano et al ., 2013 ).植物修复过程中, 根系分泌物在改善土壤属性、提高污染物的生物可利用性、缓解环境胁迫等方面具有重要作用(Bertin & Yang, 2003 ; Phillipsa et al ., 2012 ; Shahzad et al ., 2015 ).一方面, 根系分泌的某些胞外酶能够直接参与PAHs的降解过程; 另一方面, 分泌物中的可溶性糖、低分子量有机酸、氨基酸等物质, 为根际微生物提供充足的营养, 增加根际微生物的数量、提高微生物活性, 进而加速PAHs降解(Susarla et al ., 2002 ; Xie et al ., 2012 ; Lakshmanan et al ., 2014 ).同时, 根系分泌物还会直接影响PAHs的固定和活化, 进而影响PAHs在土壤-植物系统中的迁移转化行为、促进PAHs去除(Marschner et al ., 2004 ; Abhilash et al ., 2012 ); PAHs的环数越多, 根际效益越明显(Sun et al ., 2010 ).不同植物间, 因根系分泌物的组成成分及其释放特征常存在一定差异, 对根际微生物的区系结构、代谢活性的影响也不一样, 进而表现出不同的修复潜力(Kirk et al ., 2005 ; Luo et al ., 2006 ; Hunter et al ., 2014 ).因此, 探讨PAHs胁迫下不同修复潜力植物间根系分泌物中主要组分的释放特征及其种间差异, 对于研究植物修复的生物学机制、揭示不同植物间修复潜力的种间差异具有重要意义.鉴于此, 本研究以对PAHs污染土壤具有一定修复潜力的羊茅属(Festuca )植物为试验对象, 以苯并[α]芘(BaP)为PAHs代表物, 借助根际袋土培法, 对比研究PAHs胁迫下不同修复潜力的5种羊茅属植物在不同生长期时根系分泌物中几种低分子量有机物的释放特征, 为植物修复理论的研究与实践提供依据. ...
Ecotoxico- logical activity of soils polluted with polycyclic aromatic hydrocarbons (PAHs)—Effect on plants.
2000
Development of specific rhizosphere bacterial communities in relation to plant species, nutrition and soil type.
1
2004
... 植物的存在能加快土壤中PAHs的去除(Joner & Leyval, 2003 ; Kim et al ., 2004 ; Gao et al ., 2006 ), 根系分泌物(root exudates)营造的根际微生态特征则是影响PAHs快速消解的重要原因(Sumia et al ., 2013 ; Tejeda-Agredano et al ., 2013 ).植物修复过程中, 根系分泌物在改善土壤属性、提高污染物的生物可利用性、缓解环境胁迫等方面具有重要作用(Bertin & Yang, 2003 ; Phillipsa et al ., 2012 ; Shahzad et al ., 2015 ).一方面, 根系分泌的某些胞外酶能够直接参与PAHs的降解过程; 另一方面, 分泌物中的可溶性糖、低分子量有机酸、氨基酸等物质, 为根际微生物提供充足的营养, 增加根际微生物的数量、提高微生物活性, 进而加速PAHs降解(Susarla et al ., 2002 ; Xie et al ., 2012 ; Lakshmanan et al ., 2014 ).同时, 根系分泌物还会直接影响PAHs的固定和活化, 进而影响PAHs在土壤-植物系统中的迁移转化行为、促进PAHs去除(Marschner et al ., 2004 ; Abhilash et al ., 2012 ); PAHs的环数越多, 根际效益越明显(Sun et al ., 2010 ).不同植物间, 因根系分泌物的组成成分及其释放特征常存在一定差异, 对根际微生物的区系结构、代谢活性的影响也不一样, 进而表现出不同的修复潜力(Kirk et al ., 2005 ; Luo et al ., 2006 ; Hunter et al ., 2014 ).因此, 探讨PAHs胁迫下不同修复潜力植物间根系分泌物中主要组分的释放特征及其种间差异, 对于研究植物修复的生物学机制、揭示不同植物间修复潜力的种间差异具有重要意义.鉴于此, 本研究以对PAHs污染土壤具有一定修复潜力的羊茅属(Festuca )植物为试验对象, 以苯并[α]芘(BaP)为PAHs代表物, 借助根际袋土培法, 对比研究PAHs胁迫下不同修复潜力的5种羊茅属植物在不同生长期时根系分泌物中几种低分子量有机物的释放特征, 为植物修复理论的研究与实践提供依据. ...
PAH dissipation in spiked soil: Impacts of bioavailability, microbial activity, and trees.
1
2006
... 多环芳烃(polycyclic aromatic hydrocarbons, PAHs)是环境中普遍存在的持久性有机污染物, 多数具有致癌、致畸或致突变效应(Chen & Liao, 2006 ).因其性质稳定、水溶性差, 易于被土壤颗粒吸附, 土壤中PAHs含量逐年上升(Tao et al ., 2004 ; Li et al ., 2006 ), 严重威胁着土壤环境质量及农产品质量安全; 土壤中PAHs的迁移转化行为及其环境效应已成为环境修复领域的研究热点(Mueller & Shann, 2006 ; Lakshmanan et al ., 2014 ). ...
The removal and remediation of phenanthrene and pyrene in soil by mixed cropping of alfalfa and rape.
3
2008
... 按照150 mg·kg-1 CO(NH2 )2 、50 mg·kg-1 Ca(H2 PO4 )·2H2 O和100 mg·kg-1 K2 SO4 比例补充底肥后, 取适量土壤过2 mm筛, 加入BaP丙酮溶液; 待丙酮挥发后, 用未污染土壤不断稀释、搅拌, 过2 mm筛、混匀, 结合农田土壤中PAHs的背景值, 获得5个污染水平.50%田间持水量(WHC)、室温状态下平衡7天(Pan et al ., 2008 ; Wei & Pan, 2010 )后, 测得土样中BaP的初始含量分别为: 10.25 mg·kg-1 (T1 )、20.37 mg·kg-1 (T2 )、40.45 mg·kg-1 (T3 )、80.24 mg·kg-1 (T4 )和161.74 mg·kg-1 (T5 ).取污染土样2 kg装入20 cm × 15 cm的塑料盆钵中, 其中的400 g装在300目尼龙网袋中; 尼龙网袋放在盆中央, 四周及底部均为同质土样, 保持袋内外土壤高度相同.每种污染水平分装75盆, 50% WHC、室温状态下平衡5天(Pan et al ., 2008 ; Wei & Pan, 2010 ); 选取籽粒饱满、大小均匀的催芽后种子单播于袋内土壤中, 每种植物播种15盆, 每盆10颗, 置于温室中; 另设一组没有播种的污染土样作为对照, 每个处理重复3次.试验期间, 日间室温为25 ℃、夜间为18 ℃, 自然光照条件; 每2天随机交换盆钵在温室中的位置, WHC维持在50% (称重补水法).播种日记为0天, 在植物生长30、40、50、60和70天时, 分别对不同胁迫强度的供试植物、土样破坏性取样: 连同根际袋小心取出植物幼苗、尽可能不伤及其根系, 轻轻抖落根际土壤后收集其根系分泌物; 混匀根际土壤、非根际土壤后, 四分法取样, 装袋、冷冻干燥, 过 1 mm筛后低温(-20 ℃)保存、分析BaP残留量. ...
... ).取污染土样2 kg装入20 cm × 15 cm的塑料盆钵中, 其中的400 g装在300目尼龙网袋中; 尼龙网袋放在盆中央, 四周及底部均为同质土样, 保持袋内外土壤高度相同.每种污染水平分装75盆, 50% WHC、室温状态下平衡5天(Pan et al ., 2008 ; Wei & Pan, 2010 ); 选取籽粒饱满、大小均匀的催芽后种子单播于袋内土壤中, 每种植物播种15盆, 每盆10颗, 置于温室中; 另设一组没有播种的污染土样作为对照, 每个处理重复3次.试验期间, 日间室温为25 ℃、夜间为18 ℃, 自然光照条件; 每2天随机交换盆钵在温室中的位置, WHC维持在50% (称重补水法).播种日记为0天, 在植物生长30、40、50、60和70天时, 分别对不同胁迫强度的供试植物、土样破坏性取样: 连同根际袋小心取出植物幼苗、尽可能不伤及其根系, 轻轻抖落根际土壤后收集其根系分泌物; 混匀根际土壤、非根际土壤后, 四分法取样, 装袋、冷冻干燥, 过 1 mm筛后低温(-20 ℃)保存、分析BaP残留量. ...
... 参照Pan等(2008) 的方法, 略作改进: 取2.0 g土样于离心管内, 加入2.0 g无水Na2 SO4 , 混匀; 加入10 mL二氯甲烷(CH2 Cl2 ), 40 ℃下超声萃取1 h; 4000 r·min-1 下离心.取3 mL上清液过Fisher Pasteur玻璃管硅胶柱, 1:1二氯甲烷和正己烷洗脱, 40 ℃下浓缩至干, 用甲醇定容到2 mL, 过0.22 µm微孔滤膜后进行HPLC (Waters 600, Waters China, Hong Kong, China)分析.检测条件: DAD检测器, ¢4.6 × 250 mm C18 反相色谱柱; 甲醇:水(V/V, 83:17)为流动相, λ = 290 nm.在此条件下, BaP检测限为44.6 pg·L-1 , 加标回收率为94.36% (n = 7, 相对标准差(RSD ) < 5.94%)). ...
Plant root exudates impact the hydrocarbon degradation potential of a weathered-hydrocarbon contaminated soil.
1
2012
... 植物的存在能加快土壤中PAHs的去除(Joner & Leyval, 2003 ; Kim et al ., 2004 ; Gao et al ., 2006 ), 根系分泌物(root exudates)营造的根际微生态特征则是影响PAHs快速消解的重要原因(Sumia et al ., 2013 ; Tejeda-Agredano et al ., 2013 ).植物修复过程中, 根系分泌物在改善土壤属性、提高污染物的生物可利用性、缓解环境胁迫等方面具有重要作用(Bertin & Yang, 2003 ; Phillipsa et al ., 2012 ; Shahzad et al ., 2015 ).一方面, 根系分泌的某些胞外酶能够直接参与PAHs的降解过程; 另一方面, 分泌物中的可溶性糖、低分子量有机酸、氨基酸等物质, 为根际微生物提供充足的营养, 增加根际微生物的数量、提高微生物活性, 进而加速PAHs降解(Susarla et al ., 2002 ; Xie et al ., 2012 ; Lakshmanan et al ., 2014 ).同时, 根系分泌物还会直接影响PAHs的固定和活化, 进而影响PAHs在土壤-植物系统中的迁移转化行为、促进PAHs去除(Marschner et al ., 2004 ; Abhilash et al ., 2012 ); PAHs的环数越多, 根际效益越明显(Sun et al ., 2010 ).不同植物间, 因根系分泌物的组成成分及其释放特征常存在一定差异, 对根际微生物的区系结构、代谢活性的影响也不一样, 进而表现出不同的修复潜力(Kirk et al ., 2005 ; Luo et al ., 2006 ; Hunter et al ., 2014 ).因此, 探讨PAHs胁迫下不同修复潜力植物间根系分泌物中主要组分的释放特征及其种间差异, 对于研究植物修复的生物学机制、揭示不同植物间修复潜力的种间差异具有重要意义.鉴于此, 本研究以对PAHs污染土壤具有一定修复潜力的羊茅属(Festuca )植物为试验对象, 以苯并[α]芘(BaP)为PAHs代表物, 借助根际袋土培法, 对比研究PAHs胁迫下不同修复潜力的5种羊茅属植物在不同生长期时根系分泌物中几种低分子量有机物的释放特征, 为植物修复理论的研究与实践提供依据. ...
Induction of mixed-function oxygenase system and antioxidant enzymes in the coral Montastraea faveolata on acute exposure to benzo (a) pyrene.
1
2007
... 环境胁迫可能会影响根系分泌物的种类和数量(沈宏等, 2002 ; Wu et al ., 2014 ).本试验在修复潜力较强植物的根系分泌物中检测到一种低分子量有机酸——反丁烯二酸, 它可能与植物对BaP类有害物质的应激反应有关: 植物在应激反应过程中所产生的破坏性氧自由基(reactive oxygen species, ROS)容易与细胞内的生物大分子发生反应, 对线粒体、核糖体、溶酶体、过氧化物体、微粒体等细胞器以及核酸、细胞骨架蛋白等生物大分子的结构和功能产生损伤或毒害作用(Ramos & Garcia, 2007 ), 反丁烯二酸类抗氧化助剂对活性氧自由基较敏感, 能及时捕获并清除胁迫过程中产生的各种活性氧自由基、缓解环境胁迫的损害.但为什么仅在修复潜力极强的苇状羊茅、极差的细芒羊茅的根系分泌物中检测到顺丁烯二酸, 尚待进一步研究. ...
Contribution of exudates, arbuscular mycorrhizal fungi and litter depositions to the rhizosphere priming effect induced by grassland species.
1
2015
... 植物的存在能加快土壤中PAHs的去除(Joner & Leyval, 2003 ; Kim et al ., 2004 ; Gao et al ., 2006 ), 根系分泌物(root exudates)营造的根际微生态特征则是影响PAHs快速消解的重要原因(Sumia et al ., 2013 ; Tejeda-Agredano et al ., 2013 ).植物修复过程中, 根系分泌物在改善土壤属性、提高污染物的生物可利用性、缓解环境胁迫等方面具有重要作用(Bertin & Yang, 2003 ; Phillipsa et al ., 2012 ; Shahzad et al ., 2015 ).一方面, 根系分泌的某些胞外酶能够直接参与PAHs的降解过程; 另一方面, 分泌物中的可溶性糖、低分子量有机酸、氨基酸等物质, 为根际微生物提供充足的营养, 增加根际微生物的数量、提高微生物活性, 进而加速PAHs降解(Susarla et al ., 2002 ; Xie et al ., 2012 ; Lakshmanan et al ., 2014 ).同时, 根系分泌物还会直接影响PAHs的固定和活化, 进而影响PAHs在土壤-植物系统中的迁移转化行为、促进PAHs去除(Marschner et al ., 2004 ; Abhilash et al ., 2012 ); PAHs的环数越多, 根际效益越明显(Sun et al ., 2010 ).不同植物间, 因根系分泌物的组成成分及其释放特征常存在一定差异, 对根际微生物的区系结构、代谢活性的影响也不一样, 进而表现出不同的修复潜力(Kirk et al ., 2005 ; Luo et al ., 2006 ; Hunter et al ., 2014 ).因此, 探讨PAHs胁迫下不同修复潜力植物间根系分泌物中主要组分的释放特征及其种间差异, 对于研究植物修复的生物学机制、揭示不同植物间修复潜力的种间差异具有重要意义.鉴于此, 本研究以对PAHs污染土壤具有一定修复潜力的羊茅属(Festuca )植物为试验对象, 以苯并[α]芘(BaP)为PAHs代表物, 借助根际袋土培法, 对比研究PAHs胁迫下不同修复潜力的5种羊茅属植物在不同生长期时根系分泌物中几种低分子量有机物的释放特征, 为植物修复理论的研究与实践提供依据. ...
铝毒胁迫诱导菜豆柠檬酸的分泌与累积
1
2002
... 环境胁迫可能会影响根系分泌物的种类和数量(沈宏等, 2002 ; Wu et al ., 2014 ).本试验在修复潜力较强植物的根系分泌物中检测到一种低分子量有机酸——反丁烯二酸, 它可能与植物对BaP类有害物质的应激反应有关: 植物在应激反应过程中所产生的破坏性氧自由基(reactive oxygen species, ROS)容易与细胞内的生物大分子发生反应, 对线粒体、核糖体、溶酶体、过氧化物体、微粒体等细胞器以及核酸、细胞骨架蛋白等生物大分子的结构和功能产生损伤或毒害作用(Ramos & Garcia, 2007 ), 反丁烯二酸类抗氧化助剂对活性氧自由基较敏感, 能及时捕获并清除胁迫过程中产生的各种活性氧自由基、缓解环境胁迫的损害.但为什么仅在修复潜力极强的苇状羊茅、极差的细芒羊茅的根系分泌物中检测到顺丁烯二酸, 尚待进一步研究. ...
Plant- bacteria partnerships for the remediation of hydrocarbon contaminated soils.
1
2013
... 植物的存在能加快土壤中PAHs的去除(Joner & Leyval, 2003 ; Kim et al ., 2004 ; Gao et al ., 2006 ), 根系分泌物(root exudates)营造的根际微生态特征则是影响PAHs快速消解的重要原因(Sumia et al ., 2013 ; Tejeda-Agredano et al ., 2013 ).植物修复过程中, 根系分泌物在改善土壤属性、提高污染物的生物可利用性、缓解环境胁迫等方面具有重要作用(Bertin & Yang, 2003 ; Phillipsa et al ., 2012 ; Shahzad et al ., 2015 ).一方面, 根系分泌的某些胞外酶能够直接参与PAHs的降解过程; 另一方面, 分泌物中的可溶性糖、低分子量有机酸、氨基酸等物质, 为根际微生物提供充足的营养, 增加根际微生物的数量、提高微生物活性, 进而加速PAHs降解(Susarla et al ., 2002 ; Xie et al ., 2012 ; Lakshmanan et al ., 2014 ).同时, 根系分泌物还会直接影响PAHs的固定和活化, 进而影响PAHs在土壤-植物系统中的迁移转化行为、促进PAHs去除(Marschner et al ., 2004 ; Abhilash et al ., 2012 ); PAHs的环数越多, 根际效益越明显(Sun et al ., 2010 ).不同植物间, 因根系分泌物的组成成分及其释放特征常存在一定差异, 对根际微生物的区系结构、代谢活性的影响也不一样, 进而表现出不同的修复潜力(Kirk et al ., 2005 ; Luo et al ., 2006 ; Hunter et al ., 2014 ).因此, 探讨PAHs胁迫下不同修复潜力植物间根系分泌物中主要组分的释放特征及其种间差异, 对于研究植物修复的生物学机制、揭示不同植物间修复潜力的种间差异具有重要意义.鉴于此, 本研究以对PAHs污染土壤具有一定修复潜力的羊茅属(Festuca )植物为试验对象, 以苯并[α]芘(BaP)为PAHs代表物, 借助根际袋土培法, 对比研究PAHs胁迫下不同修复潜力的5种羊茅属植物在不同生长期时根系分泌物中几种低分子量有机物的释放特征, 为植物修复理论的研究与实践提供依据. ...
Roles of abiotic losses, microbes, plant roots, and root exudateson phytoremediation of PAHs in a barren soil.
1
2010
... 植物的存在能加快土壤中PAHs的去除(Joner & Leyval, 2003 ; Kim et al ., 2004 ; Gao et al ., 2006 ), 根系分泌物(root exudates)营造的根际微生态特征则是影响PAHs快速消解的重要原因(Sumia et al ., 2013 ; Tejeda-Agredano et al ., 2013 ).植物修复过程中, 根系分泌物在改善土壤属性、提高污染物的生物可利用性、缓解环境胁迫等方面具有重要作用(Bertin & Yang, 2003 ; Phillipsa et al ., 2012 ; Shahzad et al ., 2015 ).一方面, 根系分泌的某些胞外酶能够直接参与PAHs的降解过程; 另一方面, 分泌物中的可溶性糖、低分子量有机酸、氨基酸等物质, 为根际微生物提供充足的营养, 增加根际微生物的数量、提高微生物活性, 进而加速PAHs降解(Susarla et al ., 2002 ; Xie et al ., 2012 ; Lakshmanan et al ., 2014 ).同时, 根系分泌物还会直接影响PAHs的固定和活化, 进而影响PAHs在土壤-植物系统中的迁移转化行为、促进PAHs去除(Marschner et al ., 2004 ; Abhilash et al ., 2012 ); PAHs的环数越多, 根际效益越明显(Sun et al ., 2010 ).不同植物间, 因根系分泌物的组成成分及其释放特征常存在一定差异, 对根际微生物的区系结构、代谢活性的影响也不一样, 进而表现出不同的修复潜力(Kirk et al ., 2005 ; Luo et al ., 2006 ; Hunter et al ., 2014 ).因此, 探讨PAHs胁迫下不同修复潜力植物间根系分泌物中主要组分的释放特征及其种间差异, 对于研究植物修复的生物学机制、揭示不同植物间修复潜力的种间差异具有重要意义.鉴于此, 本研究以对PAHs污染土壤具有一定修复潜力的羊茅属(Festuca )植物为试验对象, 以苯并[α]芘(BaP)为PAHs代表物, 借助根际袋土培法, 对比研究PAHs胁迫下不同修复潜力的5种羊茅属植物在不同生长期时根系分泌物中几种低分子量有机物的释放特征, 为植物修复理论的研究与实践提供依据. ...
Phytoremedia- tion: An ecological solution to organic chemical contamination.
1
2002
... 植物的存在能加快土壤中PAHs的去除(Joner & Leyval, 2003 ; Kim et al ., 2004 ; Gao et al ., 2006 ), 根系分泌物(root exudates)营造的根际微生态特征则是影响PAHs快速消解的重要原因(Sumia et al ., 2013 ; Tejeda-Agredano et al ., 2013 ).植物修复过程中, 根系分泌物在改善土壤属性、提高污染物的生物可利用性、缓解环境胁迫等方面具有重要作用(Bertin & Yang, 2003 ; Phillipsa et al ., 2012 ; Shahzad et al ., 2015 ).一方面, 根系分泌的某些胞外酶能够直接参与PAHs的降解过程; 另一方面, 分泌物中的可溶性糖、低分子量有机酸、氨基酸等物质, 为根际微生物提供充足的营养, 增加根际微生物的数量、提高微生物活性, 进而加速PAHs降解(Susarla et al ., 2002 ; Xie et al ., 2012 ; Lakshmanan et al ., 2014 ).同时, 根系分泌物还会直接影响PAHs的固定和活化, 进而影响PAHs在土壤-植物系统中的迁移转化行为、促进PAHs去除(Marschner et al ., 2004 ; Abhilash et al ., 2012 ); PAHs的环数越多, 根际效益越明显(Sun et al ., 2010 ).不同植物间, 因根系分泌物的组成成分及其释放特征常存在一定差异, 对根际微生物的区系结构、代谢活性的影响也不一样, 进而表现出不同的修复潜力(Kirk et al ., 2005 ; Luo et al ., 2006 ; Hunter et al ., 2014 ).因此, 探讨PAHs胁迫下不同修复潜力植物间根系分泌物中主要组分的释放特征及其种间差异, 对于研究植物修复的生物学机制、揭示不同植物间修复潜力的种间差异具有重要意义.鉴于此, 本研究以对PAHs污染土壤具有一定修复潜力的羊茅属(Festuca )植物为试验对象, 以苯并[α]芘(BaP)为PAHs代表物, 借助根际袋土培法, 对比研究PAHs胁迫下不同修复潜力的5种羊茅属植物在不同生长期时根系分泌物中几种低分子量有机物的释放特征, 为植物修复理论的研究与实践提供依据. ...
Polycyclic aromatic hydrocarbons (PAHs) in agricultural soil and vegetable from Tianjin.
1
2004
... 多环芳烃(polycyclic aromatic hydrocarbons, PAHs)是环境中普遍存在的持久性有机污染物, 多数具有致癌、致畸或致突变效应(Chen & Liao, 2006 ).因其性质稳定、水溶性差, 易于被土壤颗粒吸附, 土壤中PAHs含量逐年上升(Tao et al ., 2004 ; Li et al ., 2006 ), 严重威胁着土壤环境质量及农产品质量安全; 土壤中PAHs的迁移转化行为及其环境效应已成为环境修复领域的研究热点(Mueller & Shann, 2006 ; Lakshmanan et al ., 2014 ). ...
Influence of the sunflower rhizosphere on the biodegradation of PAHs in soil.
1
2013
... 植物的存在能加快土壤中PAHs的去除(Joner & Leyval, 2003 ; Kim et al ., 2004 ; Gao et al ., 2006 ), 根系分泌物(root exudates)营造的根际微生态特征则是影响PAHs快速消解的重要原因(Sumia et al ., 2013 ; Tejeda-Agredano et al ., 2013 ).植物修复过程中, 根系分泌物在改善土壤属性、提高污染物的生物可利用性、缓解环境胁迫等方面具有重要作用(Bertin & Yang, 2003 ; Phillipsa et al ., 2012 ; Shahzad et al ., 2015 ).一方面, 根系分泌的某些胞外酶能够直接参与PAHs的降解过程; 另一方面, 分泌物中的可溶性糖、低分子量有机酸、氨基酸等物质, 为根际微生物提供充足的营养, 增加根际微生物的数量、提高微生物活性, 进而加速PAHs降解(Susarla et al ., 2002 ; Xie et al ., 2012 ; Lakshmanan et al ., 2014 ).同时, 根系分泌物还会直接影响PAHs的固定和活化, 进而影响PAHs在土壤-植物系统中的迁移转化行为、促进PAHs去除(Marschner et al ., 2004 ; Abhilash et al ., 2012 ); PAHs的环数越多, 根际效益越明显(Sun et al ., 2010 ).不同植物间, 因根系分泌物的组成成分及其释放特征常存在一定差异, 对根际微生物的区系结构、代谢活性的影响也不一样, 进而表现出不同的修复潜力(Kirk et al ., 2005 ; Luo et al ., 2006 ; Hunter et al ., 2014 ).因此, 探讨PAHs胁迫下不同修复潜力植物间根系分泌物中主要组分的释放特征及其种间差异, 对于研究植物修复的生物学机制、揭示不同植物间修复潜力的种间差异具有重要意义.鉴于此, 本研究以对PAHs污染土壤具有一定修复潜力的羊茅属(Festuca )植物为试验对象, 以苯并[α]芘(BaP)为PAHs代表物, 借助根际袋土培法, 对比研究PAHs胁迫下不同修复潜力的5种羊茅属植物在不同生长期时根系分泌物中几种低分子量有机物的释放特征, 为植物修复理论的研究与实践提供依据. ...
2008). 珠江三角洲地区主要树种叶片多环芳烃含量特征及影响因素分析
1
2008
... 植物生长指标的变化是植物受到不同程度胁迫伤害的直观表现症状(王海翠等, 2013 ).本试验中, 低度(T1 -T2 )或中度(T3 -T4 ) BaP胁迫刺激了植物生长, T4 水平时生物量出现峰值, 重度(T5 )胁迫则抑制了植物生长.通常, 多数植物对PAHs类环境胁迫具有一定的耐受域值, 而这种域值与PAHs的理化性质、植物种类和环境状况(Howsam et al ., 2000 ; Line et al ., 2003 ; 田晓雪等, 2008 )以及PAHs进入细胞后的累积或转化机制等因素有关(刘玉和周璐璐, 2011 ; 刘静等, 2015 ).一定域值内, 植物生长受到的抑制影响并不显著, 甚至能刺激植物生长; 超过这一域值, 植物生长就会受到抑制.Maliszewska-Kordy- bach和Smreczak (2000)报道, 由芴、蒽、芘和䓛4种PAHs组成的混合物, 在浓度不超过10 mg·kg-1 时促进了小麦(Triticum aestivum )、玉米(Zea mays )及燕麦(Avena sativa )的生长, 高浓度时抑制生长; 刘泓等(2008)研究了不同浓度的荧蒽培养基对拟南芥(Arabidopsis thaliana )生长的影响, 发现不同浓度的荧蒽胁迫30天后拟南芥生长量明显受到不同程度的抑制.尽管这些研究都表明, 高浓度PAHs胁迫抑制植物的生长, 但抑制作用机制尚不明确; 低浓度的PAHs对植物生长的促进作用, 有学者推测是因为PAHs具有与植物生长激素类似的作用(陈世军等, 2012 ), 但也仅仅局限于推测, 还没有被试验和实践证实. ...
Root exudates and arsenic accumulation in arsenic hyperaccumulating Pteris vittata and non-hyperaccumulating Nephrolepis exaltata.
1
2004
... 取出根际袋, 用自来水将根洗净、去离子水冲洗3遍后, 浸于盛有适量Milli-Q超纯水的烧杯中(锡纸包裹杯壁, 确保根部避光; 液面高度以漫过根部为宜), 自然光照下收集4 h, 用少量高纯水冲洗根系3次, 冲洗液与收集液合并、慢速滤过0.45 μm微孔滤膜(Tu et al ., 2004 ; 许超等, 2010 ).将滤液定容至50 mL, -20 ℃以下储存, 待分析.取10 mL滤液真空冷冻干燥至干, 加1 mL Milli-Q超纯水溶解, -20 ℃以下储存, 用于分析低分子量有机酸; 取20 mL滤液真空冷冻干燥至干, 加2 mL Milli-Q超纯水溶解, -20 ℃以下储存, 用于分析氨基酸(许超等, 2010 ). ...
多环芳烃(PAHs)对油菜生长的影响及其积累效应
1
2013
... 植物生长指标的变化是植物受到不同程度胁迫伤害的直观表现症状(王海翠等, 2013 ).本试验中, 低度(T1 -T2 )或中度(T3 -T4 ) BaP胁迫刺激了植物生长, T4 水平时生物量出现峰值, 重度(T5 )胁迫则抑制了植物生长.通常, 多数植物对PAHs类环境胁迫具有一定的耐受域值, 而这种域值与PAHs的理化性质、植物种类和环境状况(Howsam et al ., 2000 ; Line et al ., 2003 ; 田晓雪等, 2008 )以及PAHs进入细胞后的累积或转化机制等因素有关(刘玉和周璐璐, 2011 ; 刘静等, 2015 ).一定域值内, 植物生长受到的抑制影响并不显著, 甚至能刺激植物生长; 超过这一域值, 植物生长就会受到抑制.Maliszewska-Kordy- bach和Smreczak (2000)报道, 由芴、蒽、芘和䓛4种PAHs组成的混合物, 在浓度不超过10 mg·kg-1 时促进了小麦(Triticum aestivum )、玉米(Zea mays )及燕麦(Avena sativa )的生长, 高浓度时抑制生长; 刘泓等(2008)研究了不同浓度的荧蒽培养基对拟南芥(Arabidopsis thaliana )生长的影响, 发现不同浓度的荧蒽胁迫30天后拟南芥生长量明显受到不同程度的抑制.尽管这些研究都表明, 高浓度PAHs胁迫抑制植物的生长, 但抑制作用机制尚不明确; 低浓度的PAHs对植物生长的促进作用, 有学者推测是因为PAHs具有与植物生长激素类似的作用(陈世军等, 2012 ), 但也仅仅局限于推测, 还没有被试验和实践证实. ...
Phytoremediation for soils contami- nated by phenanthrene and pyrene with multiple plant species.
2
2010
... 按照150 mg·kg-1 CO(NH2 )2 、50 mg·kg-1 Ca(H2 PO4 )·2H2 O和100 mg·kg-1 K2 SO4 比例补充底肥后, 取适量土壤过2 mm筛, 加入BaP丙酮溶液; 待丙酮挥发后, 用未污染土壤不断稀释、搅拌, 过2 mm筛、混匀, 结合农田土壤中PAHs的背景值, 获得5个污染水平.50%田间持水量(WHC)、室温状态下平衡7天(Pan et al ., 2008 ; Wei & Pan, 2010 )后, 测得土样中BaP的初始含量分别为: 10.25 mg·kg-1 (T1 )、20.37 mg·kg-1 (T2 )、40.45 mg·kg-1 (T3 )、80.24 mg·kg-1 (T4 )和161.74 mg·kg-1 (T5 ).取污染土样2 kg装入20 cm × 15 cm的塑料盆钵中, 其中的400 g装在300目尼龙网袋中; 尼龙网袋放在盆中央, 四周及底部均为同质土样, 保持袋内外土壤高度相同.每种污染水平分装75盆, 50% WHC、室温状态下平衡5天(Pan et al ., 2008 ; Wei & Pan, 2010 ); 选取籽粒饱满、大小均匀的催芽后种子单播于袋内土壤中, 每种植物播种15盆, 每盆10颗, 置于温室中; 另设一组没有播种的污染土样作为对照, 每个处理重复3次.试验期间, 日间室温为25 ℃、夜间为18 ℃, 自然光照条件; 每2天随机交换盆钵在温室中的位置, WHC维持在50% (称重补水法).播种日记为0天, 在植物生长30、40、50、60和70天时, 分别对不同胁迫强度的供试植物、土样破坏性取样: 连同根际袋小心取出植物幼苗、尽可能不伤及其根系, 轻轻抖落根际土壤后收集其根系分泌物; 混匀根际土壤、非根际土壤后, 四分法取样, 装袋、冷冻干燥, 过 1 mm筛后低温(-20 ℃)保存、分析BaP残留量. ...
... ; Wei & Pan, 2010 ); 选取籽粒饱满、大小均匀的催芽后种子单播于袋内土壤中, 每种植物播种15盆, 每盆10颗, 置于温室中; 另设一组没有播种的污染土样作为对照, 每个处理重复3次.试验期间, 日间室温为25 ℃、夜间为18 ℃, 自然光照条件; 每2天随机交换盆钵在温室中的位置, WHC维持在50% (称重补水法).播种日记为0天, 在植物生长30、40、50、60和70天时, 分别对不同胁迫强度的供试植物、土样破坏性取样: 连同根际袋小心取出植物幼苗、尽可能不伤及其根系, 轻轻抖落根际土壤后收集其根系分泌物; 混匀根际土壤、非根际土壤后, 四分法取样, 装袋、冷冻干燥, 过 1 mm筛后低温(-20 ℃)保存、分析BaP残留量. ...
根系分泌物介导下植物-土壤-微生物互作关系研究进展与展望
2
2014
... 植物根际对PAHs的降解效果与修复植物的种类有关.Yi和Crowley (2007)研究43种植物根部组织对PAHs的降解作用发现, 有些植物对PAHs降解有明显的促进作用, 有些则无作用或有抑制作用.根系分泌物所营造的根际环境及其所呈现的根际效应是植物响应外界胁迫的重要途径, 也是影响修复效果的关键因素(Gao et al ., 2010 ; 吴林坤等, 2014 ).相同胁迫条件下, 根系分泌物的释放特征及其主要组分的种间差异主要处决于植物自身基因型的特异性及其生理状态的个体差异(Zhu et al ., 2009 ; D’Orazio et al ., 2013 ).本研究以亲缘关系较近的5种羊茅属植物为实验对象, 其基因型差异相对较小, 在一定程度上能增加研究结果的可比性.尽管待试植物在无污染(CK)、轻度或中度污染(T1 -T4 )时的生物量, 以及被植物积累的BaP量(0.08%-0.21%, 平均(0.15 ± 0.02)%差异甚微, 但它们对BaP污染土壤的修复潜力却存在显著的种间差异.可见, 植物修复过程中, 即使是遗传关系很近的同属植物间, 修复潜力的种间差异仍然存在. ...
... 环境胁迫可能会影响根系分泌物的种类和数量(沈宏等, 2002 ; Wu et al ., 2014 ).本试验在修复潜力较强植物的根系分泌物中检测到一种低分子量有机酸——反丁烯二酸, 它可能与植物对BaP类有害物质的应激反应有关: 植物在应激反应过程中所产生的破坏性氧自由基(reactive oxygen species, ROS)容易与细胞内的生物大分子发生反应, 对线粒体、核糖体、溶酶体、过氧化物体、微粒体等细胞器以及核酸、细胞骨架蛋白等生物大分子的结构和功能产生损伤或毒害作用(Ramos & Garcia, 2007 ), 反丁烯二酸类抗氧化助剂对活性氧自由基较敏感, 能及时捕获并清除胁迫过程中产生的各种活性氧自由基、缓解环境胁迫的损害.但为什么仅在修复潜力极强的苇状羊茅、极差的细芒羊茅的根系分泌物中检测到顺丁烯二酸, 尚待进一步研究. ...
芘对黑麦草根系几种低分子量有机分泌物的影响
2011
Influence of root-exudates concentration on pyrene degradation and soil microbial characteristics in pyrene contaminated soil.
1
2012
... 植物的存在能加快土壤中PAHs的去除(Joner & Leyval, 2003 ; Kim et al ., 2004 ; Gao et al ., 2006 ), 根系分泌物(root exudates)营造的根际微生态特征则是影响PAHs快速消解的重要原因(Sumia et al ., 2013 ; Tejeda-Agredano et al ., 2013 ).植物修复过程中, 根系分泌物在改善土壤属性、提高污染物的生物可利用性、缓解环境胁迫等方面具有重要作用(Bertin & Yang, 2003 ; Phillipsa et al ., 2012 ; Shahzad et al ., 2015 ).一方面, 根系分泌的某些胞外酶能够直接参与PAHs的降解过程; 另一方面, 分泌物中的可溶性糖、低分子量有机酸、氨基酸等物质, 为根际微生物提供充足的营养, 增加根际微生物的数量、提高微生物活性, 进而加速PAHs降解(Susarla et al ., 2002 ; Xie et al ., 2012 ; Lakshmanan et al ., 2014 ).同时, 根系分泌物还会直接影响PAHs的固定和活化, 进而影响PAHs在土壤-植物系统中的迁移转化行为、促进PAHs去除(Marschner et al ., 2004 ; Abhilash et al ., 2012 ); PAHs的环数越多, 根际效益越明显(Sun et al ., 2010 ).不同植物间, 因根系分泌物的组成成分及其释放特征常存在一定差异, 对根际微生物的区系结构、代谢活性的影响也不一样, 进而表现出不同的修复潜力(Kirk et al ., 2005 ; Luo et al ., 2006 ; Hunter et al ., 2014 ).因此, 探讨PAHs胁迫下不同修复潜力植物间根系分泌物中主要组分的释放特征及其种间差异, 对于研究植物修复的生物学机制、揭示不同植物间修复潜力的种间差异具有重要意义.鉴于此, 本研究以对PAHs污染土壤具有一定修复潜力的羊茅属(Festuca )植物为试验对象, 以苯并[α]芘(BaP)为PAHs代表物, 借助根际袋土培法, 对比研究PAHs胁迫下不同修复潜力的5种羊茅属植物在不同生长期时根系分泌物中几种低分子量有机物的释放特征, 为植物修复理论的研究与实践提供依据. ...
玉米幼苗根系分泌物对芘污染的响应
3
2010
... 土培试验采用根际袋试验装置在智能温室内进行(许超等, 2010 ), 涉及3个处理、5个水平(5个BaP胁迫水平下的5种植物、5个生长期), 重复3次.具体如下: ...
... 取出根际袋, 用自来水将根洗净、去离子水冲洗3遍后, 浸于盛有适量Milli-Q超纯水的烧杯中(锡纸包裹杯壁, 确保根部避光; 液面高度以漫过根部为宜), 自然光照下收集4 h, 用少量高纯水冲洗根系3次, 冲洗液与收集液合并、慢速滤过0.45 μm微孔滤膜(Tu et al ., 2004 ; 许超等, 2010 ).将滤液定容至50 mL, -20 ℃以下储存, 待分析.取10 mL滤液真空冷冻干燥至干, 加1 mL Milli-Q超纯水溶解, -20 ℃以下储存, 用于分析低分子量有机酸; 取20 mL滤液真空冷冻干燥至干, 加2 mL Milli-Q超纯水溶解, -20 ℃以下储存, 用于分析氨基酸(许超等, 2010 ). ...
... ).将滤液定容至50 mL, -20 ℃以下储存, 待分析.取10 mL滤液真空冷冻干燥至干, 加1 mL Milli-Q超纯水溶解, -20 ℃以下储存, 用于分析低分子量有机酸; 取20 mL滤液真空冷冻干燥至干, 加2 mL Milli-Q超纯水溶解, -20 ℃以下储存, 用于分析氨基酸(许超等, 2010 ). ...
Biostimulation of PAH degradation with plants containing high concentrations of linoleic acid.
2007
Effects of maize root exudates and organic acids on the desorption of phenanthrene from soils.
1
2009
... 植物根际对PAHs的降解效果与修复植物的种类有关.Yi和Crowley (2007)研究43种植物根部组织对PAHs的降解作用发现, 有些植物对PAHs降解有明显的促进作用, 有些则无作用或有抑制作用.根系分泌物所营造的根际环境及其所呈现的根际效应是植物响应外界胁迫的重要途径, 也是影响修复效果的关键因素(Gao et al ., 2010 ; 吴林坤等, 2014 ).相同胁迫条件下, 根系分泌物的释放特征及其主要组分的种间差异主要处决于植物自身基因型的特异性及其生理状态的个体差异(Zhu et al ., 2009 ; D’Orazio et al ., 2013 ).本研究以亲缘关系较近的5种羊茅属植物为实验对象, 其基因型差异相对较小, 在一定程度上能增加研究结果的可比性.尽管待试植物在无污染(CK)、轻度或中度污染(T1 -T4 )时的生物量, 以及被植物积累的BaP量(0.08%-0.21%, 平均(0.15 ± 0.02)%差异甚微, 但它们对BaP污染土壤的修复潜力却存在显著的种间差异.可见, 植物修复过程中, 即使是遗传关系很近的同属植物间, 修复潜力的种间差异仍然存在. ...
 , 袁馨
, 袁馨

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}