The survival and growth strategies, community structure and functions of microbial decomposers vary with substrate stoichiometry, which profoundly influences substrate decomposition, turnover, and hence the carbon and nutrient cycles of terrestrial ecosystems. It is crucial to understand the relationships among microbial metabolism, community structure and ecosystem processes of terrestrial ecosystems and their responses and feedbacks to global changes. In this review, we first introduced the significance of microbial decomposers in the carbon, nitrogen, and phosphorus cycles of terrestrial ecosystems from perspectives of ecological stoichiometry and metabolic theories. Then we synthesized four potential mechanisms of microbial response and control on substrate stoichiometric variations, i.e., through (1) modifying microbial stoichiometry, (2) shifting microbial community structure, (3) producing extracellular enzymes to acquire limiting resources, and (4) changing microbial carbon, nitrogen, and phosphor use efficiencies. Finally, we proposed three research directions in this field: (1) to comprehensively explore various microbial mechanisms in response to changes in substrate stoichiometry and the relative importance of these mechanisms; (2) to examine influences of global changes on microbial-driven cycles of carbon, nitrogen, and phosphorus; and (3) to explore spatiotemporal changes in the strategies of microbial adaptation to changes in the substrate stoichiometry.
Keywords:ecological stoichiometry
;
soil microbes
;
enzymes
;
biogeochemical cycle
;
homeostasis
;
carbon use efficiency
;
nutrient use efficiency
ZHOUZheng-Hu, WANGChuan-Kuan. Responses and regulation mechanisms of microbial decomposers to substrate carbon, nitrogen, and phosphorus stoichiometry[J]. Chinese Journal of Plant Ecology, 2016, 40(6): 620-630 https://doi.org/10.17521/cjpe.2015.0449
Fig. 1 Mechanisms of microbial response and regulation on substrate stoichiometry. C:X, substrate carbon to nutrient (nitrogen and phosphor) ratios; BG, β-1,4-glucosidase; NAG, β-1,4-N-acetylglucosaminidase; AP, phosphatase, EEA, extracellular enzymatic activity; RAT, resource allocation theory; TER, threshold elemental ratio; GRH, growth rate hypothesis; CNR, consumer-driven nutrient recycling; OM, overflow metabolism; CUE, carbon use efficiency; NUE, nitrogen use efficiency; PUE, phosphor use efficiency; qMB, fraction of total substrate carbon in microbe; qCO2, microbial respiration rate per unit biomass carbon; Nmin, nitrogen mineralization; Pmin, phosphor mineralization. Size of the polygons in the dotted box indicates the strength of enzymatic activity; size of the ellipses indicates the amount of microbial biomass; and the width of the arrows indicates the magnitude of the carbon, nitrogen, and phosphor fluxes.
Is the C:N:P stoichiometry in soil and soil microbial biomass related to the landscape and land use in southern subtropical China? Global Biogeochemical Cycles, 26, GB4002.
Stoichiometric imbalances between terrestrial decomposer communities and their resources: Mechanisms and implications of microbial adaptations to their resources.
The elemental stoichiometry of aquatic and terrestrial ecosystems and its relationships with organismic lifestyle and ecosystem structure and function: A review and perspectives.
Forest succession on abandoned arable soils in European Russia-Impacts on microbial biomass, fungal-bacterial ratio, and basal CO2 respiration activity.
Reviews and syntheses: Soil resources and climate jointly drive variations in microbial
1)biomass carbon and nitrogen in China’s forest ecosystems.Biogeosciences, 12, 6751-6760.
[102]
ZhouZH, WangCK, ZhangQZ (2015).
The effect of land use change on soil carbon, nitrogen, and phosphorus contents and their stoichiometry in temperate sapling stands in northeastern China.
Acta Ecologica Sinica, 35, 6694-6702. (in Chinese with English abstract)[周正虎, 王传宽, 张全智 (2015).
Soil-carbon response to warming dependent on microbial physiology.
1
2010
... 土壤呼吸是陆地生态系统向大气圈排放的最大碳源, 其中以土壤微生物代谢和生长释放出的CO2为主的异养呼吸占总土壤呼吸一半以上(Högberg et al., 2001; Bond-Lamberty et al., 2004; Wang & Yang, 2007), 因而在全球变化和生态系统研究中备受关注(Wang et al., 2006).然而, 以往研究多关注土壤呼吸组分的划分以及与土壤温湿度、有机碳储量、植被覆盖、土壤微生物生物量、环境因子等之间的相关性(Högberg et al., 2001; Bond-Lamberty et al., 2004; Wang & Yang, 2007; Wang et al., 2013), 但可能由于对微生物生理代谢机制认识不足(Allison et al., 2010; Buchkowski et al., 2015; Steinauer et al., 2015), 未能阐明异养呼吸对复杂环境因子的响应机制(Bardgett et al., 2008).微生物驱动的异养呼吸与微生物生长所需能量投资密切相关, 关系到微生物在呼吸代谢和生物量生长之间的碳资源分配.因此, 微生物将底物碳转化为自身生物量碳的效率(即微生物碳利用效率, CUE; del Giorgio & Cole, 1998)与陆地生态系统碳循环直接相关(Manzoni et al., 2012; Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015). ...
Relating belowground microbial composition to the taxonomic phylogenetic and functional trait distributions of trees in a tropical forest.
1
2015
... 微生物作为陆地生态系统主要分解者, 在获取资源构建自身生物量的同时, 驱动着生态系统物质和能量的流通, 调控着碳和养分在土壤-植物-大气连续体(SPAC)之间的循环, 进而影响生态系统的结构与功能(Chapin et al., 2003; Leff et al., 2015).尽管土壤微生物生物量碳只占土壤有机碳库的1.92% (Zhou & Wang, 2015), 但其分解作用消耗了生物圈的绝大部分净初级生产力(Cebrian & Lartigue, 2004; 王兴昌和王传宽, 2015).微生物代谢释放的CO2占土壤呼吸总通量的一半以上(Wang & Yang, 2007), 其矿化和固氮作用维持了土壤的肥力(van der Heijen et al., 2008; Leff et al., 2015); 同时, 土壤微生物与植物的共生、寄生和养分需求关系也会影响地上植被生产力和生物多样性.因此, 探索陆地生态系统地上植被组成和功能特征与地下微生物的耦合关系成为目前生态学的研究热点(van der Heijen et al., 2008; Barberán et al., 2015; Moore et al., 2015); 认识土壤微生物对碳氮磷循环的调控机制也是全球变化背景下生物地球化学循环研究的重要内容(Sinsabaugh & Follstad Shah, 2011; Mooshammer et al., 2014; Zechmeister-Boltenstern et al., 2015). ...
Microbial contributions to climate change through carbon cycle feedbacks.
1
2008
... 土壤呼吸是陆地生态系统向大气圈排放的最大碳源, 其中以土壤微生物代谢和生长释放出的CO2为主的异养呼吸占总土壤呼吸一半以上(Högberg et al., 2001; Bond-Lamberty et al., 2004; Wang & Yang, 2007), 因而在全球变化和生态系统研究中备受关注(Wang et al., 2006).然而, 以往研究多关注土壤呼吸组分的划分以及与土壤温湿度、有机碳储量、植被覆盖、土壤微生物生物量、环境因子等之间的相关性(Högberg et al., 2001; Bond-Lamberty et al., 2004; Wang & Yang, 2007; Wang et al., 2013), 但可能由于对微生物生理代谢机制认识不足(Allison et al., 2010; Buchkowski et al., 2015; Steinauer et al., 2015), 未能阐明异养呼吸对复杂环境因子的响应机制(Bardgett et al., 2008).微生物驱动的异养呼吸与微生物生长所需能量投资密切相关, 关系到微生物在呼吸代谢和生物量生长之间的碳资源分配.因此, 微生物将底物碳转化为自身生物量碳的效率(即微生物碳利用效率, CUE; del Giorgio & Cole, 1998)与陆地生态系统碳循环直接相关(Manzoni et al., 2012; Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015). ...
Active microorganisms in soil: Critical review of estimation criteria and approaches.
A global relationship between the heterotrophic and autotrophic components of soil respiration.
2
2004
... 土壤呼吸是陆地生态系统向大气圈排放的最大碳源, 其中以土壤微生物代谢和生长释放出的CO2为主的异养呼吸占总土壤呼吸一半以上(Högberg et al., 2001; Bond-Lamberty et al., 2004; Wang & Yang, 2007), 因而在全球变化和生态系统研究中备受关注(Wang et al., 2006).然而, 以往研究多关注土壤呼吸组分的划分以及与土壤温湿度、有机碳储量、植被覆盖、土壤微生物生物量、环境因子等之间的相关性(Högberg et al., 2001; Bond-Lamberty et al., 2004; Wang & Yang, 2007; Wang et al., 2013), 但可能由于对微生物生理代谢机制认识不足(Allison et al., 2010; Buchkowski et al., 2015; Steinauer et al., 2015), 未能阐明异养呼吸对复杂环境因子的响应机制(Bardgett et al., 2008).微生物驱动的异养呼吸与微生物生长所需能量投资密切相关, 关系到微生物在呼吸代谢和生物量生长之间的碳资源分配.因此, 微生物将底物碳转化为自身生物量碳的效率(即微生物碳利用效率, CUE; del Giorgio & Cole, 1998)与陆地生态系统碳循环直接相关(Manzoni et al., 2012; Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015). ...
... ; Bond-Lamberty et al., 2004; Wang & Yang, 2007; Wang et al., 2013), 但可能由于对微生物生理代谢机制认识不足(Allison et al., 2010; Buchkowski et al., 2015; Steinauer et al., 2015), 未能阐明异养呼吸对复杂环境因子的响应机制(Bardgett et al., 2008).微生物驱动的异养呼吸与微生物生长所需能量投资密切相关, 关系到微生物在呼吸代谢和生物量生长之间的碳资源分配.因此, 微生物将底物碳转化为自身生物量碳的效率(即微生物碳利用效率, CUE; del Giorgio & Cole, 1998)与陆地生态系统碳循环直接相关(Manzoni et al., 2012; Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015). ...
Global biogeography of autotroph chemistry: Is insolation a driving force?
2013
Toward a metabolic theory of ecology.
5
2004
... 随着微生物群落鉴定技术的突破和完善、分子生物学和同位素示踪在土壤微生物研究中的应用, 以土壤微生物为核心的生态系统变化过程与机理研究正蓬勃发展.由于土壤微生物个体微小且数量庞大, 其系统发生、生物多样性及其与生物和非生物因素的相互作用远比动物和植物复杂, 因而微生物生态学研究面临着巨大挑战(Prosser et al., 2007; Jansson & Prosser, 2013).运用生态化学计量学理论(Sterner & Elser, 2002)和代谢理论(Brown et al., 2004)研究土壤微生物, 不但可以提升我们对土壤微生物生态过程和机理的认识, 而且有助于生态学理论的发展与完善(Sterner & Elser, 2002; Brown et al., 2004; Sinsabaugh & Follstad Shah, 2011; Manzoni et al., 2012; Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015).生态化学计量学是研究生物系统能量平衡和多重化学元素(主要是碳、氮、磷)平衡的科学(Sterner & Elser, 2002); 代谢是指有机体从环境中获取能量和物质资源, 以最优方式分配在存活、生长和繁殖上, 并将废物排泄回环境(Brown et al., 2004).这两者都结合了生物学、化学、物理学等基本原理, 可以跨越生物科学不同层次(从细胞新陈代谢到生态系统结构和养分循环)探索不同生态系统组分之间的联系与内在机理(Sterner & Elser, 2002; Brown et al., 2004; Elser, 2006). ...
... ; Brown et al., 2004; Sinsabaugh & Follstad Shah, 2011; Manzoni et al., 2012; Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015).生态化学计量学是研究生物系统能量平衡和多重化学元素(主要是碳、氮、磷)平衡的科学(Sterner & Elser, 2002); 代谢是指有机体从环境中获取能量和物质资源, 以最优方式分配在存活、生长和繁殖上, 并将废物排泄回环境(Brown et al., 2004).这两者都结合了生物学、化学、物理学等基本原理, 可以跨越生物科学不同层次(从细胞新陈代谢到生态系统结构和养分循环)探索不同生态系统组分之间的联系与内在机理(Sterner & Elser, 2002; Brown et al., 2004; Elser, 2006). ...
... ); 代谢是指有机体从环境中获取能量和物质资源, 以最优方式分配在存活、生长和繁殖上, 并将废物排泄回环境(Brown et al., 2004).这两者都结合了生物学、化学、物理学等基本原理, 可以跨越生物科学不同层次(从细胞新陈代谢到生态系统结构和养分循环)探索不同生态系统组分之间的联系与内在机理(Sterner & Elser, 2002; Brown et al., 2004; Elser, 2006). ...
... ; Brown et al., 2004; Elser, 2006). ...
... 地球上所有生物的生长和繁殖都需要25种相同的化学元素, 但这些元素的相对比例(及化学计量)在生物体之间存在巨大差异; 这一方面取决于生物有机体自身的结构限制和代谢需求(Sterner & Elser, 2002; Brown et al., 2004), 另一方面也受环境影响而表现出一定环境可塑性(Sardans et al., 2012; Yang et al., 2015). ...
Microbial stoichiometry overrides biomass as a regulator of soil carbon and nitrogen cycling.
6
2015
... 随着微生物群落鉴定技术的突破和完善、分子生物学和同位素示踪在土壤微生物研究中的应用, 以土壤微生物为核心的生态系统变化过程与机理研究正蓬勃发展.由于土壤微生物个体微小且数量庞大, 其系统发生、生物多样性及其与生物和非生物因素的相互作用远比动物和植物复杂, 因而微生物生态学研究面临着巨大挑战(Prosser et al., 2007; Jansson & Prosser, 2013).运用生态化学计量学理论(Sterner & Elser, 2002)和代谢理论(Brown et al., 2004)研究土壤微生物, 不但可以提升我们对土壤微生物生态过程和机理的认识, 而且有助于生态学理论的发展与完善(Sterner & Elser, 2002; Brown et al., 2004; Sinsabaugh & Follstad Shah, 2011; Manzoni et al., 2012; Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015).生态化学计量学是研究生物系统能量平衡和多重化学元素(主要是碳、氮、磷)平衡的科学(Sterner & Elser, 2002); 代谢是指有机体从环境中获取能量和物质资源, 以最优方式分配在存活、生长和繁殖上, 并将废物排泄回环境(Brown et al., 2004).这两者都结合了生物学、化学、物理学等基本原理, 可以跨越生物科学不同层次(从细胞新陈代谢到生态系统结构和养分循环)探索不同生态系统组分之间的联系与内在机理(Sterner & Elser, 2002; Brown et al., 2004; Elser, 2006). ...
... 土壤呼吸是陆地生态系统向大气圈排放的最大碳源, 其中以土壤微生物代谢和生长释放出的CO2为主的异养呼吸占总土壤呼吸一半以上(Högberg et al., 2001; Bond-Lamberty et al., 2004; Wang & Yang, 2007), 因而在全球变化和生态系统研究中备受关注(Wang et al., 2006).然而, 以往研究多关注土壤呼吸组分的划分以及与土壤温湿度、有机碳储量、植被覆盖、土壤微生物生物量、环境因子等之间的相关性(Högberg et al., 2001; Bond-Lamberty et al., 2004; Wang & Yang, 2007; Wang et al., 2013), 但可能由于对微生物生理代谢机制认识不足(Allison et al., 2010; Buchkowski et al., 2015; Steinauer et al., 2015), 未能阐明异养呼吸对复杂环境因子的响应机制(Bardgett et al., 2008).微生物驱动的异养呼吸与微生物生长所需能量投资密切相关, 关系到微生物在呼吸代谢和生物量生长之间的碳资源分配.因此, 微生物将底物碳转化为自身生物量碳的效率(即微生物碳利用效率, CUE; del Giorgio & Cole, 1998)与陆地生态系统碳循环直接相关(Manzoni et al., 2012; Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015). ...
... ; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015). ...
... 陆地生态系统中微生物的分解底物主要来源于植物的枯枝落叶、粗木质残体、根系凋落物和分泌物、土壤有机质等(Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015).在全球尺度上, 森林生态系统中植物叶片C:N:P为1212:28:1 (摩尔比, McGroddy et al., 2004).植物叶片在凋亡过程中的养分再吸收作用, 使得枯叶养分含量骤减, 从而使枯叶的C:N:P变为3007:45:1 (McGroddy et al., 2004; Yuan & Chen, 2009; Reed et al., 2012; Vergutz et al., 2012; Han et al., 2013).此外, 枯叶的C:N:P还会随生态系统、生物区系以及生活型的不同而发生显著变化(McGroddy et al., 2004).粗木质残体和枯立木的现存量占生态系统碎屑碳库的45% (Zhang & Wang, 2010); 粗木质残体占整个生态系统植物碎屑年输入量的10% (温带森林, Harmon et al., 1986)至30% (热带森林, Martinelli et al., 2000), 其C:N:P可高达14103:40:1 (Weedon et al., 2009).植物细根周转产生的凋落物是土壤碳和养分的重要来源, 其死根C:N:P的全球平均值为4184:43:1 (Yuan et al., 2011).土壤C:N:P最低(287:17:1, Xu et al., 2013), 且随着土壤深度增加而显著减小(Tian et al., 2010).相比之下, 土壤微生物生物量C:N:P比其分解底物低得多.例如: Cleveland和Liptzin (2007)整合全球数据时发现, 陆地生态系统土壤微生物生物量碳氮磷化学计量虽然存在类似于海洋浮游植物的“Redfield ratio” 在资源不受限制时浮游植物的C:N:P为106:16:1; Redfield, 1958), 即土壤微生物生物量C:N:P具有内稳性(homeostasis; Sterner & Elser, 2002), 但森林和草地的土壤微生物生物量C:N:P之间存在显著差异.近期Xu等(2013)进一步整合分析表明, 土壤微生物生物量C:N:P随生物区系不同有很大的变异性(从沙漠的31:4:1到天然湿地的131:14:1).面临底物和微生物化学计量的巨大变异性, 微生物主要通过调整自身化学计量特征、群落结构、胞外酶比例、元素利用效率等4种机制来响应和调节(图1). ...
... 微生物是陆地生态系统的主要分解者, 其生存和生长策略、群落结构和功能会随着底物化学计量特征而改变, 从而强烈地影响底物的分解速度和元素的周转速率, 进而影响生态系统的功能过程(Manzoni et al., 2010; Sinsabaugh & Shah, 2011; Mooshammer et al., 2012; Xu et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015; Zhou & Wang, 2015).深入理解微生物生理代谢活动和群落结构与陆地生态系统过程之间联系及其对全球变化的响应和反馈机理, 是生态学和全球变化生物学领域值得关注的重大科学命题, 其中亟待解决的主要科学问题有: ...
Patterns of herbivory and decomposition in aquatic and terrestrial ecosystems.
1
2004
... 微生物作为陆地生态系统主要分解者, 在获取资源构建自身生物量的同时, 驱动着生态系统物质和能量的流通, 调控着碳和养分在土壤-植物-大气连续体(SPAC)之间的循环, 进而影响生态系统的结构与功能(Chapin et al., 2003; Leff et al., 2015).尽管土壤微生物生物量碳只占土壤有机碳库的1.92% (Zhou & Wang, 2015), 但其分解作用消耗了生物圈的绝大部分净初级生产力(Cebrian & Lartigue, 2004; 王兴昌和王传宽, 2015).微生物代谢释放的CO2占土壤呼吸总通量的一半以上(Wang & Yang, 2007), 其矿化和固氮作用维持了土壤的肥力(van der Heijen et al., 2008; Leff et al., 2015); 同时, 土壤微生物与植物的共生、寄生和养分需求关系也会影响地上植被生产力和生物多样性.因此, 探索陆地生态系统地上植被组成和功能特征与地下微生物的耦合关系成为目前生态学的研究热点(van der Heijen et al., 2008; Barberán et al., 2015; Moore et al., 2015); 认识土壤微生物对碳氮磷循环的调控机制也是全球变化背景下生物地球化学循环研究的重要内容(Sinsabaugh & Follstad Shah, 2011; Mooshammer et al., 2014; Zechmeister-Boltenstern et al., 2015). ...
Nitrogen phosphorus and carbon mineralization in response to nutrient and lime additions in peatlands.
1
2003
... 微生物作为陆地生态系统主要分解者, 在获取资源构建自身生物量的同时, 驱动着生态系统物质和能量的流通, 调控着碳和养分在土壤-植物-大气连续体(SPAC)之间的循环, 进而影响生态系统的结构与功能(Chapin et al., 2003; Leff et al., 2015).尽管土壤微生物生物量碳只占土壤有机碳库的1.92% (Zhou & Wang, 2015), 但其分解作用消耗了生物圈的绝大部分净初级生产力(Cebrian & Lartigue, 2004; 王兴昌和王传宽, 2015).微生物代谢释放的CO2占土壤呼吸总通量的一半以上(Wang & Yang, 2007), 其矿化和固氮作用维持了土壤的肥力(van der Heijen et al., 2008; Leff et al., 2015); 同时, 土壤微生物与植物的共生、寄生和养分需求关系也会影响地上植被生产力和生物多样性.因此, 探索陆地生态系统地上植被组成和功能特征与地下微生物的耦合关系成为目前生态学的研究热点(van der Heijen et al., 2008; Barberán et al., 2015; Moore et al., 2015); 认识土壤微生物对碳氮磷循环的调控机制也是全球变化背景下生物地球化学循环研究的重要内容(Sinsabaugh & Follstad Shah, 2011; Mooshammer et al., 2014; Zechmeister-Boltenstern et al., 2015). ...
C:N:P stoichiometry in soil: Is there a “Redfield ratio” for the microbial biomass?
2007
Bacterial growth efficiency in natural aquatic systems.
1
1998
... 土壤呼吸是陆地生态系统向大气圈排放的最大碳源, 其中以土壤微生物代谢和生长释放出的CO2为主的异养呼吸占总土壤呼吸一半以上(Högberg et al., 2001; Bond-Lamberty et al., 2004; Wang & Yang, 2007), 因而在全球变化和生态系统研究中备受关注(Wang et al., 2006).然而, 以往研究多关注土壤呼吸组分的划分以及与土壤温湿度、有机碳储量、植被覆盖、土壤微生物生物量、环境因子等之间的相关性(Högberg et al., 2001; Bond-Lamberty et al., 2004; Wang & Yang, 2007; Wang et al., 2013), 但可能由于对微生物生理代谢机制认识不足(Allison et al., 2010; Buchkowski et al., 2015; Steinauer et al., 2015), 未能阐明异养呼吸对复杂环境因子的响应机制(Bardgett et al., 2008).微生物驱动的异养呼吸与微生物生长所需能量投资密切相关, 关系到微生物在呼吸代谢和生物量生长之间的碳资源分配.因此, 微生物将底物碳转化为自身生物量碳的效率(即微生物碳利用效率, CUE; del Giorgio & Cole, 1998)与陆地生态系统碳循环直接相关(Manzoni et al., 2012; Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015). ...
Carbon-nitrogen interactions during afforestation in central China.
Large-scale forest girdling shows that current photosynthesis drives soil respiration.
3
2001
... 土壤呼吸是陆地生态系统向大气圈排放的最大碳源, 其中以土壤微生物代谢和生长释放出的CO2为主的异养呼吸占总土壤呼吸一半以上(Högberg et al., 2001; Bond-Lamberty et al., 2004; Wang & Yang, 2007), 因而在全球变化和生态系统研究中备受关注(Wang et al., 2006).然而, 以往研究多关注土壤呼吸组分的划分以及与土壤温湿度、有机碳储量、植被覆盖、土壤微生物生物量、环境因子等之间的相关性(Högberg et al., 2001; Bond-Lamberty et al., 2004; Wang & Yang, 2007; Wang et al., 2013), 但可能由于对微生物生理代谢机制认识不足(Allison et al., 2010; Buchkowski et al., 2015; Steinauer et al., 2015), 未能阐明异养呼吸对复杂环境因子的响应机制(Bardgett et al., 2008).微生物驱动的异养呼吸与微生物生长所需能量投资密切相关, 关系到微生物在呼吸代谢和生物量生长之间的碳资源分配.因此, 微生物将底物碳转化为自身生物量碳的效率(即微生物碳利用效率, CUE; del Giorgio & Cole, 1998)与陆地生态系统碳循环直接相关(Manzoni et al., 2012; Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015). ...
... ).然而, 以往研究多关注土壤呼吸组分的划分以及与土壤温湿度、有机碳储量、植被覆盖、土壤微生物生物量、环境因子等之间的相关性(Högberg et al., 2001; Bond-Lamberty et al., 2004; Wang & Yang, 2007; Wang et al., 2013), 但可能由于对微生物生理代谢机制认识不足(Allison et al., 2010; Buchkowski et al., 2015; Steinauer et al., 2015), 未能阐明异养呼吸对复杂环境因子的响应机制(Bardgett et al., 2008).微生物驱动的异养呼吸与微生物生长所需能量投资密切相关, 关系到微生物在呼吸代谢和生物量生长之间的碳资源分配.因此, 微生物将底物碳转化为自身生物量碳的效率(即微生物碳利用效率, CUE; del Giorgio & Cole, 1998)与陆地生态系统碳循环直接相关(Manzoni et al., 2012; Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015). ...
... 陆地生态系统中微生物的分解底物主要来源于植物的枯枝落叶、粗木质残体、根系凋落物和分泌物、土壤有机质等(Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015).在全球尺度上, 森林生态系统中植物叶片C:N:P为1212:28:1 (摩尔比, McGroddy et al., 2004).植物叶片在凋亡过程中的养分再吸收作用, 使得枯叶养分含量骤减, 从而使枯叶的C:N:P变为3007:45:1 (McGroddy et al., 2004; Yuan & Chen, 2009; Reed et al., 2012; Vergutz et al., 2012; Han et al., 2013).此外, 枯叶的C:N:P还会随生态系统、生物区系以及生活型的不同而发生显著变化(McGroddy et al., 2004).粗木质残体和枯立木的现存量占生态系统碎屑碳库的45% (Zhang & Wang, 2010); 粗木质残体占整个生态系统植物碎屑年输入量的10% (温带森林, Harmon et al., 1986)至30% (热带森林, Martinelli et al., 2000), 其C:N:P可高达14103:40:1 (Weedon et al., 2009).植物细根周转产生的凋落物是土壤碳和养分的重要来源, 其死根C:N:P的全球平均值为4184:43:1 (Yuan et al., 2011).土壤C:N:P最低(287:17:1, Xu et al., 2013), 且随着土壤深度增加而显著减小(Tian et al., 2010).相比之下, 土壤微生物生物量C:N:P比其分解底物低得多.例如: Cleveland和Liptzin (2007)整合全球数据时发现, 陆地生态系统土壤微生物生物量碳氮磷化学计量虽然存在类似于海洋浮游植物的“Redfield ratio” 在资源不受限制时浮游植物的C:N:P为106:16:1; Redfield, 1958), 即土壤微生物生物量C:N:P具有内稳性(homeostasis; Sterner & Elser, 2002), 但森林和草地的土壤微生物生物量C:N:P之间存在显著差异.近期Xu等(2013)进一步整合分析表明, 土壤微生物生物量C:N:P随生物区系不同有很大的变异性(从沙漠的31:4:1到天然湿地的131:14:1).面临底物和微生物化学计量的巨大变异性, 微生物主要通过调整自身化学计量特征、群落结构、胞外酶比例、元素利用效率等4种机制来响应和调节(图1). ...
Metabolic quotient of the soil microflora in relation to plant succession.
1989
Phosphorus supply and cycling at long-term forest monitoring sites in Germany.
1
2009
... (3)探索微生物对底物化学计量变化适应对策的时空变化.Odum (1969)生态系统演替理论指出, 生态系统呼吸和生物量的比例会随着演替进程而减小.Insam和Haselwandter (1989)首次提出生态系统演替过程中微生物也有类似的变化趋势, 即单位微生物生物量的呼吸速率随着演替进程而减小.但Wardle和Ghani (1995)的整合研究发现大多数情况下单位微生物生物量的呼吸速率随着演替进程先减后增.此后相继出现大量研究, 但仍无统一结论和解释(Saviozzi et al., 2001; Goberna et al., 2006; Susyan et al., 2011; Lu et al., 2014; Raiesi & Beheshti, 2015).运用化学计量学理论可能有助于解释这些争议: 随着生态系统的演替进程, 一方面植物光合作用和微生物固氮作用将大气中的碳和氮素不断输入土壤, 从而导致土壤C:P和N:P的增加; 另一方面, 凋落物、地表枯落物层、土壤C:N随着演替进程基本保持不变(Yang & Luo, 2011; Yang et al., 2011; 周正虎等, 2015), 而单位微生物生物量的呼吸速率随着C:P的增加而增加(图1; Spohn & Chodak, 2015), 从而出现Wardle和Ghani (1995)的“先减后增”格局.另外, 普遍认为, 高纬度地区受氮限制, 而低纬度地区受磷限制; 生态系统演替初期受氮限制, 而后期受磷限制(Vitousek et al., 2010).然而, Spohn和Chodak (2015)在德国发现温带森林土壤C:P与单位微生物生物量的呼吸速率显著相关, 而与C:N不相关; 长期监测结果也显示, 该地区森林植物叶片的磷含量也在减小(Ilg et al., 2009).因此, 上述生态系统演替和全球变化(尤其是氮沉降)可能会导致生态系统的磷限制而突破磷会受纬度限制的传统认识(Vitousek et al., 2010). ...
Microbiology: The life beneath our feet.
2
2013
... 随着微生物群落鉴定技术的突破和完善、分子生物学和同位素示踪在土壤微生物研究中的应用, 以土壤微生物为核心的生态系统变化过程与机理研究正蓬勃发展.由于土壤微生物个体微小且数量庞大, 其系统发生、生物多样性及其与生物和非生物因素的相互作用远比动物和植物复杂, 因而微生物生态学研究面临着巨大挑战(Prosser et al., 2007; Jansson & Prosser, 2013).运用生态化学计量学理论(Sterner & Elser, 2002)和代谢理论(Brown et al., 2004)研究土壤微生物, 不但可以提升我们对土壤微生物生态过程和机理的认识, 而且有助于生态学理论的发展与完善(Sterner & Elser, 2002; Brown et al., 2004; Sinsabaugh & Follstad Shah, 2011; Manzoni et al., 2012; Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015).生态化学计量学是研究生物系统能量平衡和多重化学元素(主要是碳、氮、磷)平衡的科学(Sterner & Elser, 2002); 代谢是指有机体从环境中获取能量和物质资源, 以最优方式分配在存活、生长和繁殖上, 并将废物排泄回环境(Brown et al., 2004).这两者都结合了生物学、化学、物理学等基本原理, 可以跨越生物科学不同层次(从细胞新陈代谢到生态系统结构和养分循环)探索不同生态系统组分之间的联系与内在机理(Sterner & Elser, 2002; Brown et al., 2004; Elser, 2006). ...
... 氮和磷是陆地生态系统生产力的关键限制元素(Vitousek & Howarth, 1991; Vitousek et al., 2010).微生物分解者主要通过驱动如下过程来调节陆地生态系统氮循环: 通过共生固氮、自生固氮、联合固氮等将大气中分子氮还原为铵态氮; 优先吸收铵态氮后同化为有机氮; 通过胞外酶将资源中有机氮转化为无机氮的过程(即矿化作用); 通过自养硝化细菌的硝化作用将氨氧化为硝态氮; 通过反硝化细菌的反硝化作用将硝态氮还原为氮气或氧化亚氮重新进入大气圈(Jetten, 2008; Rick & Thomas, 2001).尽管微生物不能像固氮作用一样增加土壤磷总含量, 但土壤微生物的活动会显著改变土壤中磷的化学形式和比例, 从而在调节植物磷可利用性中扮演重要角色(van der Heijden et al., 2008).例如: 微生物分解有机质释放有效磷; 在磷限制的生态系统中, 微生物生物量磷占土壤总磷的比例可达到23.6% (天然湿地生态系统Xu et al., 2013); 又因为土壤微生物生物量的最大值出现在冬季, 使得在植被生长季中土壤微生物生物量减小时释放出磷, 及时增加土壤中磷的可利用性(Richardson & Simpson, 2011; Xu et al., 2013).另外, 土壤微生物的存在还会改变土壤的吸附平衡, 从而增加正磷酸盐向土壤溶液中的转移(Seeling & Zasoski, 1993).由此可见, 微生物分解者在陆地生态系统生物地球化学循环中起着重要的驱动和调节作用. ...
Microbial community dynamics alleviate stoichiometric constraints during litter decay.
3
2014
... 在微生物群落水平上, 自然选择会使那些能够充分获取并利用特定资源的种群在竞争中获胜(Moorhead & Sinsabaugh, 2006; Fierer et al., 2007; Strickland & Rousk, 2010; Martin et al., 2015; Zelezniak et al., 2015).因此, 底物的质量(高质量意味着低的碳与养分比(C:X), 低质量意味着高的C:X; Xu et al., 2014)的改变必然会改变微生物群落组成.目前很多学者认为, 微生物生物量C:N:P的可塑性是由微生物群落结构的变化导致的(Fanin et al., 2013; Kaiser et al., 2014; Mooshammer et al., 2014; Zechmeister-Boltenstern et al., 2015).生长和周转迅速的r对策微生物常具有较低的生物量C:X, 适应于高质量的底物; 相反, 生长和周转速度较慢的k对策微生物常具有较高的生物量C:X, 适应于低质量底物(Moorhead & Sinsabaugh, 2006; Fierer et al., 2007; Strickland & Rousk, 2010; Kaiser et al., 2014).然而, 由于微生物数量、群落组成以及代谢活动的复杂性, 微生物群落结构、生物量C:N:P与底物C:N:P之间的关系仍不明确.例如: 由于真菌的C:N比细菌高(Strickland & Rousk, 2010), 且真菌与细菌的比例以及底物C:N均随着土壤剖面而降低(Moore et al., 2010; Lee et al., 2013).Mooshammer等(2014)在分析微生物分解者对底物质量的适应机制时推测, 真菌与细菌比例应该随着底物C:N的增加而增加.Fanin等(2013)的确发现微生物生物量C:N与真菌与细菌比例存在显著的相关性, 但真菌与细菌比例与底物C:N:P并无相关性.然而, 同属于子囊菌门(Ascom- ycota)的刺盾炱纲(Chaetothyriomycetes)的真菌适应于低质量底物, 而锤舌菌纲(Leotiomycetes)和粪壳菌纲(Sordariomycetes)的真菌却适应于高质量的底物; 这表明, 即便是在真菌内部, 不同分类单元对底物质量的要求也不尽相同(Strickland et al., 2009; Sch- neider et al., 2012).因此, 微生物群落结构、生物量C:N:P与底物C:N:P之间普适性的关系还需验证. ...
... ; Kaiser et al., 2014).然而, 由于微生物数量、群落组成以及代谢活动的复杂性, 微生物群落结构、生物量C:N:P与底物C:N:P之间的关系仍不明确.例如: 由于真菌的C:N比细菌高(Strickland & Rousk, 2010), 且真菌与细菌的比例以及底物C:N均随着土壤剖面而降低(Moore et al., 2010; Lee et al., 2013).Mooshammer等(2014)在分析微生物分解者对底物质量的适应机制时推测, 真菌与细菌比例应该随着底物C:N的增加而增加.Fanin等(2013)的确发现微生物生物量C:N与真菌与细菌比例存在显著的相关性, 但真菌与细菌比例与底物C:N:P并无相关性.然而, 同属于子囊菌门(Ascom- ycota)的刺盾炱纲(Chaetothyriomycetes)的真菌适应于低质量底物, 而锤舌菌纲(Leotiomycetes)和粪壳菌纲(Sordariomycetes)的真菌却适应于高质量的底物; 这表明, 即便是在真菌内部, 不同分类单元对底物质量的要求也不尽相同(Strickland et al., 2009; Sch- neider et al., 2012).因此, 微生物群落结构、生物量C:N:P与底物C:N:P之间普适性的关系还需验证. ...
... (1)综合阐明微生物对底物化学计量变化响应的各种机制及其相对重要性.尽管已经提出上述4种微生物适应底物化学计量变化的可能机制, 但每种机制还需要进一步验证, 尤其需要通过综合性实验研究在不同时空尺度上各种机理的相对贡献程度.具体来说, 微生物生物量C:N:P的可塑性同时受底物化学计量、气候、土壤理化性质等众多因子影响, 因此微生物生物量C:N:P与底物C:N:P的关联性及其强度有待于量化.虽然近期大量研究关注底物C:N:P与微生物群落的相关性(Fanin et al., 2013; Kaiser et al., 2014), 但仍然没有普适性的结论.理论上, 微生物群落结构的变化必然导致微生物生物量C:N:P的变化, 但区分其贡献程度仍有难度.化学计量学通过简单、统一而又本质的元素计量特征将复杂的生态系统层次有机地联系起来,考虑微生物群落结构可能反而将问题复杂化.胞外酶在解释微生物对氮的获取和利用上仍具有很大的局限性.对微生物元素利用效率研究的不足主要体现在缺乏可靠的测定方法.目前主要通过添加同位素标记的特定基质进行测定, 但微生物同时也会利用未标记的土壤有机质, 因而测定结果具有不确定性(Sinsabaugh et al., 2013).至于回答各种机理在什么条件下起主导作用, 以及相对贡献程度等问题, 还需要设计综合实验. ...
Organic layer serves as a hotspot of microbial activity and abundance in Arctic tundra soils.
1
2013
... 在微生物群落水平上, 自然选择会使那些能够充分获取并利用特定资源的种群在竞争中获胜(Moorhead & Sinsabaugh, 2006; Fierer et al., 2007; Strickland & Rousk, 2010; Martin et al., 2015; Zelezniak et al., 2015).因此, 底物的质量(高质量意味着低的碳与养分比(C:X), 低质量意味着高的C:X; Xu et al., 2014)的改变必然会改变微生物群落组成.目前很多学者认为, 微生物生物量C:N:P的可塑性是由微生物群落结构的变化导致的(Fanin et al., 2013; Kaiser et al., 2014; Mooshammer et al., 2014; Zechmeister-Boltenstern et al., 2015).生长和周转迅速的r对策微生物常具有较低的生物量C:X, 适应于高质量的底物; 相反, 生长和周转速度较慢的k对策微生物常具有较高的生物量C:X, 适应于低质量底物(Moorhead & Sinsabaugh, 2006; Fierer et al., 2007; Strickland & Rousk, 2010; Kaiser et al., 2014).然而, 由于微生物数量、群落组成以及代谢活动的复杂性, 微生物群落结构、生物量C:N:P与底物C:N:P之间的关系仍不明确.例如: 由于真菌的C:N比细菌高(Strickland & Rousk, 2010), 且真菌与细菌的比例以及底物C:N均随着土壤剖面而降低(Moore et al., 2010; Lee et al., 2013).Mooshammer等(2014)在分析微生物分解者对底物质量的适应机制时推测, 真菌与细菌比例应该随着底物C:N的增加而增加.Fanin等(2013)的确发现微生物生物量C:N与真菌与细菌比例存在显著的相关性, 但真菌与细菌比例与底物C:N:P并无相关性.然而, 同属于子囊菌门(Ascom- ycota)的刺盾炱纲(Chaetothyriomycetes)的真菌适应于低质量底物, 而锤舌菌纲(Leotiomycetes)和粪壳菌纲(Sordariomycetes)的真菌却适应于高质量的底物; 这表明, 即便是在真菌内部, 不同分类单元对底物质量的要求也不尽相同(Strickland et al., 2009; Sch- neider et al., 2012).因此, 微生物群落结构、生物量C:N:P与底物C:N:P之间普适性的关系还需验证. ...
Consistent responses of soil microbial communities to elevated nutrient inputs in grasslands across the globe.
2
2015
... 微生物作为陆地生态系统主要分解者, 在获取资源构建自身生物量的同时, 驱动着生态系统物质和能量的流通, 调控着碳和养分在土壤-植物-大气连续体(SPAC)之间的循环, 进而影响生态系统的结构与功能(Chapin et al., 2003; Leff et al., 2015).尽管土壤微生物生物量碳只占土壤有机碳库的1.92% (Zhou & Wang, 2015), 但其分解作用消耗了生物圈的绝大部分净初级生产力(Cebrian & Lartigue, 2004; 王兴昌和王传宽, 2015).微生物代谢释放的CO2占土壤呼吸总通量的一半以上(Wang & Yang, 2007), 其矿化和固氮作用维持了土壤的肥力(van der Heijen et al., 2008; Leff et al., 2015); 同时, 土壤微生物与植物的共生、寄生和养分需求关系也会影响地上植被生产力和生物多样性.因此, 探索陆地生态系统地上植被组成和功能特征与地下微生物的耦合关系成为目前生态学的研究热点(van der Heijen et al., 2008; Barberán et al., 2015; Moore et al., 2015); 认识土壤微生物对碳氮磷循环的调控机制也是全球变化背景下生物地球化学循环研究的重要内容(Sinsabaugh & Follstad Shah, 2011; Mooshammer et al., 2014; Zechmeister-Boltenstern et al., 2015). ...
... ; Leff et al., 2015); 同时, 土壤微生物与植物的共生、寄生和养分需求关系也会影响地上植被生产力和生物多样性.因此, 探索陆地生态系统地上植被组成和功能特征与地下微生物的耦合关系成为目前生态学的研究热点(van der Heijen et al., 2008; Barberán et al., 2015; Moore et al., 2015); 认识土壤微生物对碳氮磷循环的调控机制也是全球变化背景下生物地球化学循环研究的重要内容(Sinsabaugh & Follstad Shah, 2011; Mooshammer et al., 2014; Zechmeister-Boltenstern et al., 2015). ...
Global patterns of soil microbial nitrogen and phosphorus stoichiometry in forest ecosystems.
Is the C:N:P stoichiometry in soil and soil microbial biomass related to the landscape and land use in southern subtropical China? Global Biogeochemical Cycles, 26, GB4002.
2012
Testing the growth rate vs. geochemical hypothesis for latitudinal variation in plant nutrients.
2007
Effect of vegetation types on chemical and biological properties of soils of karst ecosystems.
1
2014
... (3)探索微生物对底物化学计量变化适应对策的时空变化.Odum (1969)生态系统演替理论指出, 生态系统呼吸和生物量的比例会随着演替进程而减小.Insam和Haselwandter (1989)首次提出生态系统演替过程中微生物也有类似的变化趋势, 即单位微生物生物量的呼吸速率随着演替进程而减小.但Wardle和Ghani (1995)的整合研究发现大多数情况下单位微生物生物量的呼吸速率随着演替进程先减后增.此后相继出现大量研究, 但仍无统一结论和解释(Saviozzi et al., 2001; Goberna et al., 2006; Susyan et al., 2011; Lu et al., 2014; Raiesi & Beheshti, 2015).运用化学计量学理论可能有助于解释这些争议: 随着生态系统的演替进程, 一方面植物光合作用和微生物固氮作用将大气中的碳和氮素不断输入土壤, 从而导致土壤C:P和N:P的增加; 另一方面, 凋落物、地表枯落物层、土壤C:N随着演替进程基本保持不变(Yang & Luo, 2011; Yang et al., 2011; 周正虎等, 2015), 而单位微生物生物量的呼吸速率随着C:P的增加而增加(图1; Spohn & Chodak, 2015), 从而出现Wardle和Ghani (1995)的“先减后增”格局.另外, 普遍认为, 高纬度地区受氮限制, 而低纬度地区受磷限制; 生态系统演替初期受氮限制, 而后期受磷限制(Vitousek et al., 2010).然而, Spohn和Chodak (2015)在德国发现温带森林土壤C:P与单位微生物生物量的呼吸速率显著相关, 而与C:N不相关; 长期监测结果也显示, 该地区森林植物叶片的磷含量也在减小(Ilg et al., 2009).因此, 上述生态系统演替和全球变化(尤其是氮沉降)可能会导致生态系统的磷限制而突破磷会受纬度限制的传统认识(Vitousek et al., 2010). ...
Are bacteria more like plants or animals? Growth rate and resource dependence of bacterial C: N: P stoichiometry.
1
2003
... et al., 2015).此外, 尽管以往研究普遍认为微生物生物量C:N变异性很小(Makino et al., 2003), 然而Zhou和Wang (2015)最近整合中国森林土壤微生物生物量碳氮格局时发现, 土壤资源(土壤有机碳、全氮)和气候条件(年平均气温、年降水量)共同解释了土壤微生物生物量C:N的24.4%的变异性, 其中土壤资源和气候条件各占一半.由此可见, 针对底物化学计量的变异性, 微生物的适应对策之一是其个体生物量C:N:P也随之变化, 即非内稳性(non-homeos- tasis), 这被Sterner和Elser (2002)形象地描述为“You are what you eat”.然而, 微生物生物量C:N:P与底物C:N:P之间的关系仍没有定论.例如: Li等(2012)分析我国亚热带地区1069个土壤数据时报道, 微生物生物量C:N和C:P随着土壤C:N和C:P的增加而显著增加, 微生物生物量N:P却随着土壤N:P的增加而呈现减小的趋势.Zhou和Wang (2015)发现我国森林土壤微生物生物量C:N随着土壤C:N的增加反而减小, 与Cleveland和Liptzin (2007)的全球森林数据分析结果吻合.而Fanin等(2013)的凋落物分解实验却指出微生物生物量C:P和N:P均与凋落物浸出液C:P和N:P显著正相关.产生这种现象的可能原因是微生物化学计量比会随着研究区域气候条件的变化而变化; 大尺度上的全球整合分析夹杂了气候、土壤类型等因素的影响, 而小尺度或个体研究也许更能反映微生物化学计量比与底物化学计量比的相关性.因此, 微生物与底物化学计量之间关系的方向与强度还需要进一步验证. ...
Environmental and stoichiometric controls on microbial carbon-use efficiency in soils.
2
2012
... 随着微生物群落鉴定技术的突破和完善、分子生物学和同位素示踪在土壤微生物研究中的应用, 以土壤微生物为核心的生态系统变化过程与机理研究正蓬勃发展.由于土壤微生物个体微小且数量庞大, 其系统发生、生物多样性及其与生物和非生物因素的相互作用远比动物和植物复杂, 因而微生物生态学研究面临着巨大挑战(Prosser et al., 2007; Jansson & Prosser, 2013).运用生态化学计量学理论(Sterner & Elser, 2002)和代谢理论(Brown et al., 2004)研究土壤微生物, 不但可以提升我们对土壤微生物生态过程和机理的认识, 而且有助于生态学理论的发展与完善(Sterner & Elser, 2002; Brown et al., 2004; Sinsabaugh & Follstad Shah, 2011; Manzoni et al., 2012; Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015).生态化学计量学是研究生物系统能量平衡和多重化学元素(主要是碳、氮、磷)平衡的科学(Sterner & Elser, 2002); 代谢是指有机体从环境中获取能量和物质资源, 以最优方式分配在存活、生长和繁殖上, 并将废物排泄回环境(Brown et al., 2004).这两者都结合了生物学、化学、物理学等基本原理, 可以跨越生物科学不同层次(从细胞新陈代谢到生态系统结构和养分循环)探索不同生态系统组分之间的联系与内在机理(Sterner & Elser, 2002; Brown et al., 2004; Elser, 2006). ...
... 土壤呼吸是陆地生态系统向大气圈排放的最大碳源, 其中以土壤微生物代谢和生长释放出的CO2为主的异养呼吸占总土壤呼吸一半以上(Högberg et al., 2001; Bond-Lamberty et al., 2004; Wang & Yang, 2007), 因而在全球变化和生态系统研究中备受关注(Wang et al., 2006).然而, 以往研究多关注土壤呼吸组分的划分以及与土壤温湿度、有机碳储量、植被覆盖、土壤微生物生物量、环境因子等之间的相关性(Högberg et al., 2001; Bond-Lamberty et al., 2004; Wang & Yang, 2007; Wang et al., 2013), 但可能由于对微生物生理代谢机制认识不足(Allison et al., 2010; Buchkowski et al., 2015; Steinauer et al., 2015), 未能阐明异养呼吸对复杂环境因子的响应机制(Bardgett et al., 2008).微生物驱动的异养呼吸与微生物生长所需能量投资密切相关, 关系到微生物在呼吸代谢和生物量生长之间的碳资源分配.因此, 微生物将底物碳转化为自身生物量碳的效率(即微生物碳利用效率, CUE; del Giorgio & Cole, 1998)与陆地生态系统碳循环直接相关(Manzoni et al., 2012; Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015). ...
Stoichiometric controls on carbon, nitrogen, and phosphorus dynamics in decomposing litter.
... 微生物是陆地生态系统的主要分解者, 其生存和生长策略、群落结构和功能会随着底物化学计量特征而改变, 从而强烈地影响底物的分解速度和元素的周转速率, 进而影响生态系统的功能过程(Manzoni et al., 2010; Sinsabaugh & Shah, 2011; Mooshammer et al., 2012; Xu et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015; Zhou & Wang, 2015).深入理解微生物生理代谢活动和群落结构与陆地生态系统过程之间联系及其对全球变化的响应和反馈机理, 是生态学和全球变化生物学领域值得关注的重大科学命题, 其中亟待解决的主要科学问题有: ...
Stoichiometric imbalances between terrestrial decomposer communities and their resources: Mechanisms and implications of microbial adaptations to their resources.
6
2014
... 微生物作为陆地生态系统主要分解者, 在获取资源构建自身生物量的同时, 驱动着生态系统物质和能量的流通, 调控着碳和养分在土壤-植物-大气连续体(SPAC)之间的循环, 进而影响生态系统的结构与功能(Chapin et al., 2003; Leff et al., 2015).尽管土壤微生物生物量碳只占土壤有机碳库的1.92% (Zhou & Wang, 2015), 但其分解作用消耗了生物圈的绝大部分净初级生产力(Cebrian & Lartigue, 2004; 王兴昌和王传宽, 2015).微生物代谢释放的CO2占土壤呼吸总通量的一半以上(Wang & Yang, 2007), 其矿化和固氮作用维持了土壤的肥力(van der Heijen et al., 2008; Leff et al., 2015); 同时, 土壤微生物与植物的共生、寄生和养分需求关系也会影响地上植被生产力和生物多样性.因此, 探索陆地生态系统地上植被组成和功能特征与地下微生物的耦合关系成为目前生态学的研究热点(van der Heijen et al., 2008; Barberán et al., 2015; Moore et al., 2015); 认识土壤微生物对碳氮磷循环的调控机制也是全球变化背景下生物地球化学循环研究的重要内容(Sinsabaugh & Follstad Shah, 2011; Mooshammer et al., 2014; Zechmeister-Boltenstern et al., 2015). ...
... 随着微生物群落鉴定技术的突破和完善、分子生物学和同位素示踪在土壤微生物研究中的应用, 以土壤微生物为核心的生态系统变化过程与机理研究正蓬勃发展.由于土壤微生物个体微小且数量庞大, 其系统发生、生物多样性及其与生物和非生物因素的相互作用远比动物和植物复杂, 因而微生物生态学研究面临着巨大挑战(Prosser et al., 2007; Jansson & Prosser, 2013).运用生态化学计量学理论(Sterner & Elser, 2002)和代谢理论(Brown et al., 2004)研究土壤微生物, 不但可以提升我们对土壤微生物生态过程和机理的认识, 而且有助于生态学理论的发展与完善(Sterner & Elser, 2002; Brown et al., 2004; Sinsabaugh & Follstad Shah, 2011; Manzoni et al., 2012; Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015).生态化学计量学是研究生物系统能量平衡和多重化学元素(主要是碳、氮、磷)平衡的科学(Sterner & Elser, 2002); 代谢是指有机体从环境中获取能量和物质资源, 以最优方式分配在存活、生长和繁殖上, 并将废物排泄回环境(Brown et al., 2004).这两者都结合了生物学、化学、物理学等基本原理, 可以跨越生物科学不同层次(从细胞新陈代谢到生态系统结构和养分循环)探索不同生态系统组分之间的联系与内在机理(Sterner & Elser, 2002; Brown et al., 2004; Elser, 2006). ...
... 土壤呼吸是陆地生态系统向大气圈排放的最大碳源, 其中以土壤微生物代谢和生长释放出的CO2为主的异养呼吸占总土壤呼吸一半以上(Högberg et al., 2001; Bond-Lamberty et al., 2004; Wang & Yang, 2007), 因而在全球变化和生态系统研究中备受关注(Wang et al., 2006).然而, 以往研究多关注土壤呼吸组分的划分以及与土壤温湿度、有机碳储量、植被覆盖、土壤微生物生物量、环境因子等之间的相关性(Högberg et al., 2001; Bond-Lamberty et al., 2004; Wang & Yang, 2007; Wang et al., 2013), 但可能由于对微生物生理代谢机制认识不足(Allison et al., 2010; Buchkowski et al., 2015; Steinauer et al., 2015), 未能阐明异养呼吸对复杂环境因子的响应机制(Bardgett et al., 2008).微生物驱动的异养呼吸与微生物生长所需能量投资密切相关, 关系到微生物在呼吸代谢和生物量生长之间的碳资源分配.因此, 微生物将底物碳转化为自身生物量碳的效率(即微生物碳利用效率, CUE; del Giorgio & Cole, 1998)与陆地生态系统碳循环直接相关(Manzoni et al., 2012; Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015). ...
... 陆地生态系统中微生物的分解底物主要来源于植物的枯枝落叶、粗木质残体、根系凋落物和分泌物、土壤有机质等(Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015).在全球尺度上, 森林生态系统中植物叶片C:N:P为1212:28:1 (摩尔比, McGroddy et al., 2004).植物叶片在凋亡过程中的养分再吸收作用, 使得枯叶养分含量骤减, 从而使枯叶的C:N:P变为3007:45:1 (McGroddy et al., 2004; Yuan & Chen, 2009; Reed et al., 2012; Vergutz et al., 2012; Han et al., 2013).此外, 枯叶的C:N:P还会随生态系统、生物区系以及生活型的不同而发生显著变化(McGroddy et al., 2004).粗木质残体和枯立木的现存量占生态系统碎屑碳库的45% (Zhang & Wang, 2010); 粗木质残体占整个生态系统植物碎屑年输入量的10% (温带森林, Harmon et al., 1986)至30% (热带森林, Martinelli et al., 2000), 其C:N:P可高达14103:40:1 (Weedon et al., 2009).植物细根周转产生的凋落物是土壤碳和养分的重要来源, 其死根C:N:P的全球平均值为4184:43:1 (Yuan et al., 2011).土壤C:N:P最低(287:17:1, Xu et al., 2013), 且随着土壤深度增加而显著减小(Tian et al., 2010).相比之下, 土壤微生物生物量C:N:P比其分解底物低得多.例如: Cleveland和Liptzin (2007)整合全球数据时发现, 陆地生态系统土壤微生物生物量碳氮磷化学计量虽然存在类似于海洋浮游植物的“Redfield ratio” 在资源不受限制时浮游植物的C:N:P为106:16:1; Redfield, 1958), 即土壤微生物生物量C:N:P具有内稳性(homeostasis; Sterner & Elser, 2002), 但森林和草地的土壤微生物生物量C:N:P之间存在显著差异.近期Xu等(2013)进一步整合分析表明, 土壤微生物生物量C:N:P随生物区系不同有很大的变异性(从沙漠的31:4:1到天然湿地的131:14:1).面临底物和微生物化学计量的巨大变异性, 微生物主要通过调整自身化学计量特征、群落结构、胞外酶比例、元素利用效率等4种机制来响应和调节(图1). ...
... 在微生物群落水平上, 自然选择会使那些能够充分获取并利用特定资源的种群在竞争中获胜(Moorhead & Sinsabaugh, 2006; Fierer et al., 2007; Strickland & Rousk, 2010; Martin et al., 2015; Zelezniak et al., 2015).因此, 底物的质量(高质量意味着低的碳与养分比(C:X), 低质量意味着高的C:X; Xu et al., 2014)的改变必然会改变微生物群落组成.目前很多学者认为, 微生物生物量C:N:P的可塑性是由微生物群落结构的变化导致的(Fanin et al., 2013; Kaiser et al., 2014; Mooshammer et al., 2014; Zechmeister-Boltenstern et al., 2015).生长和周转迅速的r对策微生物常具有较低的生物量C:X, 适应于高质量的底物; 相反, 生长和周转速度较慢的k对策微生物常具有较高的生物量C:X, 适应于低质量底物(Moorhead & Sinsabaugh, 2006; Fierer et al., 2007; Strickland & Rousk, 2010; Kaiser et al., 2014).然而, 由于微生物数量、群落组成以及代谢活动的复杂性, 微生物群落结构、生物量C:N:P与底物C:N:P之间的关系仍不明确.例如: 由于真菌的C:N比细菌高(Strickland & Rousk, 2010), 且真菌与细菌的比例以及底物C:N均随着土壤剖面而降低(Moore et al., 2010; Lee et al., 2013).Mooshammer等(2014)在分析微生物分解者对底物质量的适应机制时推测, 真菌与细菌比例应该随着底物C:N的增加而增加.Fanin等(2013)的确发现微生物生物量C:N与真菌与细菌比例存在显著的相关性, 但真菌与细菌比例与底物C:N:P并无相关性.然而, 同属于子囊菌门(Ascom- ycota)的刺盾炱纲(Chaetothyriomycetes)的真菌适应于低质量底物, 而锤舌菌纲(Leotiomycetes)和粪壳菌纲(Sordariomycetes)的真菌却适应于高质量的底物; 这表明, 即便是在真菌内部, 不同分类单元对底物质量的要求也不尽相同(Strickland et al., 2009; Sch- neider et al., 2012).因此, 微生物群落结构、生物量C:N:P与底物C:N:P之间普适性的关系还需验证. ...
... 通常将生物对底物C:X所能承受的限度定义为元素比率阈值(threshold elemental ratio).在微生物研究中, 元素比率阈值建立在代谢理论和化学计量理论基础上, 整合了微生物生物量C:N:P、胞外酶化学计量关系(BG:NAG:AP), 以及元素利用效率对底物C:N:P的适应策略(Sinsabaugh & Follstad Shah, 2012).如果底物C:X超过这个阈值, 则微生物受养分限制, 使其碳利用效率随底物C:X的增加而减小、养分利用效率会增加(Mooshammer et al., 2014; Zechmeister-Boltenstern et al., 2015); 反之, 当底物C:X低于这个阈值时, 微生物受到能量(碳)限制, 使其碳利用效率随底物C:X的增加而增加, 而养分利用效率会降低(Sterner & Elser, 2002; Frost et al. 2006; Doi et al. 2010; Sinsabaugh & Follstad Shah, 2012). ...
The strategy of ecosystem development.
1969
The role of ecological theory in microbial ecology.
2
2007
... 随着微生物群落鉴定技术的突破和完善、分子生物学和同位素示踪在土壤微生物研究中的应用, 以土壤微生物为核心的生态系统变化过程与机理研究正蓬勃发展.由于土壤微生物个体微小且数量庞大, 其系统发生、生物多样性及其与生物和非生物因素的相互作用远比动物和植物复杂, 因而微生物生态学研究面临着巨大挑战(Prosser et al., 2007; Jansson & Prosser, 2013).运用生态化学计量学理论(Sterner & Elser, 2002)和代谢理论(Brown et al., 2004)研究土壤微生物, 不但可以提升我们对土壤微生物生态过程和机理的认识, 而且有助于生态学理论的发展与完善(Sterner & Elser, 2002; Brown et al., 2004; Sinsabaugh & Follstad Shah, 2011; Manzoni et al., 2012; Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015).生态化学计量学是研究生物系统能量平衡和多重化学元素(主要是碳、氮、磷)平衡的科学(Sterner & Elser, 2002); 代谢是指有机体从环境中获取能量和物质资源, 以最优方式分配在存活、生长和繁殖上, 并将废物排泄回环境(Brown et al., 2004).这两者都结合了生物学、化学、物理学等基本原理, 可以跨越生物科学不同层次(从细胞新陈代谢到生态系统结构和养分循环)探索不同生态系统组分之间的联系与内在机理(Sterner & Elser, 2002; Brown et al., 2004; Elser, 2006). ...
Microbiological indicators of soil quality and degradation following conversion of native forests to continuous croplands.
1
2015
... (3)探索微生物对底物化学计量变化适应对策的时空变化.Odum (1969)生态系统演替理论指出, 生态系统呼吸和生物量的比例会随着演替进程而减小.Insam和Haselwandter (1989)首次提出生态系统演替过程中微生物也有类似的变化趋势, 即单位微生物生物量的呼吸速率随着演替进程而减小.但Wardle和Ghani (1995)的整合研究发现大多数情况下单位微生物生物量的呼吸速率随着演替进程先减后增.此后相继出现大量研究, 但仍无统一结论和解释(Saviozzi et al., 2001; Goberna et al., 2006; Susyan et al., 2011; Lu et al., 2014; Raiesi & Beheshti, 2015).运用化学计量学理论可能有助于解释这些争议: 随着生态系统的演替进程, 一方面植物光合作用和微生物固氮作用将大气中的碳和氮素不断输入土壤, 从而导致土壤C:P和N:P的增加; 另一方面, 凋落物、地表枯落物层、土壤C:N随着演替进程基本保持不变(Yang & Luo, 2011; Yang et al., 2011; 周正虎等, 2015), 而单位微生物生物量的呼吸速率随着C:P的增加而增加(图1; Spohn & Chodak, 2015), 从而出现Wardle和Ghani (1995)的“先减后增”格局.另外, 普遍认为, 高纬度地区受氮限制, 而低纬度地区受磷限制; 生态系统演替初期受氮限制, 而后期受磷限制(Vitousek et al., 2010).然而, Spohn和Chodak (2015)在德国发现温带森林土壤C:P与单位微生物生物量的呼吸速率显著相关, 而与C:N不相关; 长期监测结果也显示, 该地区森林植物叶片的磷含量也在减小(Ilg et al., 2009).因此, 上述生态系统演替和全球变化(尤其是氮沉降)可能会导致生态系统的磷限制而突破磷会受纬度限制的传统认识(Vitousek et al., 2010). ...
The biological control of chemical factors in the environment.
1
1958
... 陆地生态系统中微生物的分解底物主要来源于植物的枯枝落叶、粗木质残体、根系凋落物和分泌物、土壤有机质等(Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015).在全球尺度上, 森林生态系统中植物叶片C:N:P为1212:28:1 (摩尔比, McGroddy et al., 2004).植物叶片在凋亡过程中的养分再吸收作用, 使得枯叶养分含量骤减, 从而使枯叶的C:N:P变为3007:45:1 (McGroddy et al., 2004; Yuan & Chen, 2009; Reed et al., 2012; Vergutz et al., 2012; Han et al., 2013).此外, 枯叶的C:N:P还会随生态系统、生物区系以及生活型的不同而发生显著变化(McGroddy et al., 2004).粗木质残体和枯立木的现存量占生态系统碎屑碳库的45% (Zhang & Wang, 2010); 粗木质残体占整个生态系统植物碎屑年输入量的10% (温带森林, Harmon et al., 1986)至30% (热带森林, Martinelli et al., 2000), 其C:N:P可高达14103:40:1 (Weedon et al., 2009).植物细根周转产生的凋落物是土壤碳和养分的重要来源, 其死根C:N:P的全球平均值为4184:43:1 (Yuan et al., 2011).土壤C:N:P最低(287:17:1, Xu et al., 2013), 且随着土壤深度增加而显著减小(Tian et al., 2010).相比之下, 土壤微生物生物量C:N:P比其分解底物低得多.例如: Cleveland和Liptzin (2007)整合全球数据时发现, 陆地生态系统土壤微生物生物量碳氮磷化学计量虽然存在类似于海洋浮游植物的“Redfield ratio” 在资源不受限制时浮游植物的C:N:P为106:16:1; Redfield, 1958), 即土壤微生物生物量C:N:P具有内稳性(homeostasis; Sterner & Elser, 2002), 但森林和草地的土壤微生物生物量C:N:P之间存在显著差异.近期Xu等(2013)进一步整合分析表明, 土壤微生物生物量C:N:P随生物区系不同有很大的变异性(从沙漠的31:4:1到天然湿地的131:14:1).面临底物和微生物化学计量的巨大变异性, 微生物主要通过调整自身化学计量特征、群落结构、胞外酶比例、元素利用效率等4种机制来响应和调节(图1). ...
Stoichiometric patterns in foliar nutrient resorption across multiple scales.
1
2012
... 陆地生态系统中微生物的分解底物主要来源于植物的枯枝落叶、粗木质残体、根系凋落物和分泌物、土壤有机质等(Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015).在全球尺度上, 森林生态系统中植物叶片C:N:P为1212:28:1 (摩尔比, McGroddy et al., 2004).植物叶片在凋亡过程中的养分再吸收作用, 使得枯叶养分含量骤减, 从而使枯叶的C:N:P变为3007:45:1 (McGroddy et al., 2004; Yuan & Chen, 2009; Reed et al., 2012; Vergutz et al., 2012; Han et al., 2013).此外, 枯叶的C:N:P还会随生态系统、生物区系以及生活型的不同而发生显著变化(McGroddy et al., 2004).粗木质残体和枯立木的现存量占生态系统碎屑碳库的45% (Zhang & Wang, 2010); 粗木质残体占整个生态系统植物碎屑年输入量的10% (温带森林, Harmon et al., 1986)至30% (热带森林, Martinelli et al., 2000), 其C:N:P可高达14103:40:1 (Weedon et al., 2009).植物细根周转产生的凋落物是土壤碳和养分的重要来源, 其死根C:N:P的全球平均值为4184:43:1 (Yuan et al., 2011).土壤C:N:P最低(287:17:1, Xu et al., 2013), 且随着土壤深度增加而显著减小(Tian et al., 2010).相比之下, 土壤微生物生物量C:N:P比其分解底物低得多.例如: Cleveland和Liptzin (2007)整合全球数据时发现, 陆地生态系统土壤微生物生物量碳氮磷化学计量虽然存在类似于海洋浮游植物的“Redfield ratio” 在资源不受限制时浮游植物的C:N:P为106:16:1; Redfield, 1958), 即土壤微生物生物量C:N:P具有内稳性(homeostasis; Sterner & Elser, 2002), 但森林和草地的土壤微生物生物量C:N:P之间存在显著差异.近期Xu等(2013)进一步整合分析表明, 土壤微生物生物量C:N:P随生物区系不同有很大的变异性(从沙漠的31:4:1到天然湿地的131:14:1).面临底物和微生物化学计量的巨大变异性, 微生物主要通过调整自身化学计量特征、群落结构、胞外酶比例、元素利用效率等4种机制来响应和调节(图1). ...
Soil microorganisms mediating phosphorus availability update on microbial phosphorus.
1
2011
... 氮和磷是陆地生态系统生产力的关键限制元素(Vitousek & Howarth, 1991; Vitousek et al., 2010).微生物分解者主要通过驱动如下过程来调节陆地生态系统氮循环: 通过共生固氮、自生固氮、联合固氮等将大气中分子氮还原为铵态氮; 优先吸收铵态氮后同化为有机氮; 通过胞外酶将资源中有机氮转化为无机氮的过程(即矿化作用); 通过自养硝化细菌的硝化作用将氨氧化为硝态氮; 通过反硝化细菌的反硝化作用将硝态氮还原为氮气或氧化亚氮重新进入大气圈(Jetten, 2008; Rick & Thomas, 2001).尽管微生物不能像固氮作用一样增加土壤磷总含量, 但土壤微生物的活动会显著改变土壤中磷的化学形式和比例, 从而在调节植物磷可利用性中扮演重要角色(van der Heijden et al., 2008).例如: 微生物分解有机质释放有效磷; 在磷限制的生态系统中, 微生物生物量磷占土壤总磷的比例可达到23.6% (天然湿地生态系统Xu et al., 2013); 又因为土壤微生物生物量的最大值出现在冬季, 使得在植被生长季中土壤微生物生物量减小时释放出磷, 及时增加土壤中磷的可利用性(Richardson & Simpson, 2011; Xu et al., 2013).另外, 土壤微生物的存在还会改变土壤的吸附平衡, 从而增加正磷酸盐向土壤溶液中的转移(Seeling & Zasoski, 1993).由此可见, 微生物分解者在陆地生态系统生物地球化学循环中起着重要的驱动和调节作用. ...
Microbial nitrogen cycles: Physiology genomics and applications.
1
2001
... 氮和磷是陆地生态系统生产力的关键限制元素(Vitousek & Howarth, 1991; Vitousek et al., 2010).微生物分解者主要通过驱动如下过程来调节陆地生态系统氮循环: 通过共生固氮、自生固氮、联合固氮等将大气中分子氮还原为铵态氮; 优先吸收铵态氮后同化为有机氮; 通过胞外酶将资源中有机氮转化为无机氮的过程(即矿化作用); 通过自养硝化细菌的硝化作用将氨氧化为硝态氮; 通过反硝化细菌的反硝化作用将硝态氮还原为氮气或氧化亚氮重新进入大气圈(Jetten, 2008; Rick & Thomas, 2001).尽管微生物不能像固氮作用一样增加土壤磷总含量, 但土壤微生物的活动会显著改变土壤中磷的化学形式和比例, 从而在调节植物磷可利用性中扮演重要角色(van der Heijden et al., 2008).例如: 微生物分解有机质释放有效磷; 在磷限制的生态系统中, 微生物生物量磷占土壤总磷的比例可达到23.6% (天然湿地生态系统Xu et al., 2013); 又因为土壤微生物生物量的最大值出现在冬季, 使得在植被生长季中土壤微生物生物量减小时释放出磷, 及时增加土壤中磷的可利用性(Richardson & Simpson, 2011; Xu et al., 2013).另外, 土壤微生物的存在还会改变土壤的吸附平衡, 从而增加正磷酸盐向土壤溶液中的转移(Seeling & Zasoski, 1993).由此可见, 微生物分解者在陆地生态系统生物地球化学循环中起着重要的驱动和调节作用. ...
The role of plants in the effects of global change on nutrient availability and stoichiometry in the plant-soil system.
3
2012
... 地球上所有生物的生长和繁殖都需要25种相同的化学元素, 但这些元素的相对比例(及化学计量)在生物体之间存在巨大差异; 这一方面取决于生物有机体自身的结构限制和代谢需求(Sterner & Elser, 2002; Brown et al., 2004), 另一方面也受环境影响而表现出一定环境可塑性(Sardans et al., 2012; Yang et al., 2015). ...
The elemental stoichiometry of aquatic and terrestrial ecosystems and its relationships with organismic lifestyle and ecosystem structure and function: A review and perspectives.
2012
A comparison of soil quality in adjacent cultivated, forest and native grassland soils.
1
2001
... (3)探索微生物对底物化学计量变化适应对策的时空变化.Odum (1969)生态系统演替理论指出, 生态系统呼吸和生物量的比例会随着演替进程而减小.Insam和Haselwandter (1989)首次提出生态系统演替过程中微生物也有类似的变化趋势, 即单位微生物生物量的呼吸速率随着演替进程而减小.但Wardle和Ghani (1995)的整合研究发现大多数情况下单位微生物生物量的呼吸速率随着演替进程先减后增.此后相继出现大量研究, 但仍无统一结论和解释(Saviozzi et al., 2001; Goberna et al., 2006; Susyan et al., 2011; Lu et al., 2014; Raiesi & Beheshti, 2015).运用化学计量学理论可能有助于解释这些争议: 随着生态系统的演替进程, 一方面植物光合作用和微生物固氮作用将大气中的碳和氮素不断输入土壤, 从而导致土壤C:P和N:P的增加; 另一方面, 凋落物、地表枯落物层、土壤C:N随着演替进程基本保持不变(Yang & Luo, 2011; Yang et al., 2011; 周正虎等, 2015), 而单位微生物生物量的呼吸速率随着C:P的增加而增加(图1; Spohn & Chodak, 2015), 从而出现Wardle和Ghani (1995)的“先减后增”格局.另外, 普遍认为, 高纬度地区受氮限制, 而低纬度地区受磷限制; 生态系统演替初期受氮限制, 而后期受磷限制(Vitousek et al., 2010).然而, Spohn和Chodak (2015)在德国发现温带森林土壤C:P与单位微生物生物量的呼吸速率显著相关, 而与C:N不相关; 长期监测结果也显示, 该地区森林植物叶片的磷含量也在减小(Ilg et al., 2009).因此, 上述生态系统演替和全球变化(尤其是氮沉降)可能会导致生态系统的磷限制而突破磷会受纬度限制的传统认识(Vitousek et al., 2010). ...
The implications of exoenzyme activity on microbial carbon and nitrogen limitation in soil: A theoretical model.
... ; Spohn & Chodak, 2015).相反.当微生物生长于高质量底物环境中时, 微生物氮磷矿化速率会随之增加; 这两者之间的相关性已被广泛证实(Manzoni et al., 2010; Moosha- mmer et al., 2012; Deng et al., 2014; Buchkowski et al., 2015).Zhou和Wang (2015)最近还发现, 微生物生物量碳在土壤总有机碳中所占的比例随土壤C:N的增加而显著减小, 但微生物生物量氮在土壤总氮中所占的比例却随土壤C:N的增加而显著增加, 这表明底物C:X还会影响微生物生物量大小以及土壤的元素化学计量. ...
... (3)探索微生物对底物化学计量变化适应对策的时空变化.Odum (1969)生态系统演替理论指出, 生态系统呼吸和生物量的比例会随着演替进程而减小.Insam和Haselwandter (1989)首次提出生态系统演替过程中微生物也有类似的变化趋势, 即单位微生物生物量的呼吸速率随着演替进程而减小.但Wardle和Ghani (1995)的整合研究发现大多数情况下单位微生物生物量的呼吸速率随着演替进程先减后增.此后相继出现大量研究, 但仍无统一结论和解释(Saviozzi et al., 2001; Goberna et al., 2006; Susyan et al., 2011; Lu et al., 2014; Raiesi & Beheshti, 2015).运用化学计量学理论可能有助于解释这些争议: 随着生态系统的演替进程, 一方面植物光合作用和微生物固氮作用将大气中的碳和氮素不断输入土壤, 从而导致土壤C:P和N:P的增加; 另一方面, 凋落物、地表枯落物层、土壤C:N随着演替进程基本保持不变(Yang & Luo, 2011; Yang et al., 2011; 周正虎等, 2015), 而单位微生物生物量的呼吸速率随着C:P的增加而增加(图1; Spohn & Chodak, 2015), 从而出现Wardle和Ghani (1995)的“先减后增”格局.另外, 普遍认为, 高纬度地区受氮限制, 而低纬度地区受磷限制; 生态系统演替初期受氮限制, 而后期受磷限制(Vitousek et al., 2010).然而, Spohn和Chodak (2015)在德国发现温带森林土壤C:P与单位微生物生物量的呼吸速率显著相关, 而与C:N不相关; 长期监测结果也显示, 该地区森林植物叶片的磷含量也在减小(Ilg et al., 2009).因此, 上述生态系统演替和全球变化(尤其是氮沉降)可能会导致生态系统的磷限制而突破磷会受纬度限制的传统认识(Vitousek et al., 2010). ...
Plant diversity effects on soil microbial functions and enzymes are stronger than warming in a grassland experiment.
1
2015
... 土壤呼吸是陆地生态系统向大气圈排放的最大碳源, 其中以土壤微生物代谢和生长释放出的CO2为主的异养呼吸占总土壤呼吸一半以上(Högberg et al., 2001; Bond-Lamberty et al., 2004; Wang & Yang, 2007), 因而在全球变化和生态系统研究中备受关注(Wang et al., 2006).然而, 以往研究多关注土壤呼吸组分的划分以及与土壤温湿度、有机碳储量、植被覆盖、土壤微生物生物量、环境因子等之间的相关性(Högberg et al., 2001; Bond-Lamberty et al., 2004; Wang & Yang, 2007; Wang et al., 2013), 但可能由于对微生物生理代谢机制认识不足(Allison et al., 2010; Buchkowski et al., 2015; Steinauer et al., 2015), 未能阐明异养呼吸对复杂环境因子的响应机制(Bardgett et al., 2008).微生物驱动的异养呼吸与微生物生长所需能量投资密切相关, 关系到微生物在呼吸代谢和生物量生长之间的碳资源分配.因此, 微生物将底物碳转化为自身生物量碳的效率(即微生物碳利用效率, CUE; del Giorgio & Cole, 1998)与陆地生态系统碳循环直接相关(Manzoni et al., 2012; Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015). ...
9
... 随着微生物群落鉴定技术的突破和完善、分子生物学和同位素示踪在土壤微生物研究中的应用, 以土壤微生物为核心的生态系统变化过程与机理研究正蓬勃发展.由于土壤微生物个体微小且数量庞大, 其系统发生、生物多样性及其与生物和非生物因素的相互作用远比动物和植物复杂, 因而微生物生态学研究面临着巨大挑战(Prosser et al., 2007; Jansson & Prosser, 2013).运用生态化学计量学理论(Sterner & Elser, 2002)和代谢理论(Brown et al., 2004)研究土壤微生物, 不但可以提升我们对土壤微生物生态过程和机理的认识, 而且有助于生态学理论的发展与完善(Sterner & Elser, 2002; Brown et al., 2004; Sinsabaugh & Follstad Shah, 2011; Manzoni et al., 2012; Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015).生态化学计量学是研究生物系统能量平衡和多重化学元素(主要是碳、氮、磷)平衡的科学(Sterner & Elser, 2002); 代谢是指有机体从环境中获取能量和物质资源, 以最优方式分配在存活、生长和繁殖上, 并将废物排泄回环境(Brown et al., 2004).这两者都结合了生物学、化学、物理学等基本原理, 可以跨越生物科学不同层次(从细胞新陈代谢到生态系统结构和养分循环)探索不同生态系统组分之间的联系与内在机理(Sterner & Elser, 2002; Brown et al., 2004; Elser, 2006). ...
... )研究土壤微生物, 不但可以提升我们对土壤微生物生态过程和机理的认识, 而且有助于生态学理论的发展与完善(Sterner & Elser, 2002; Brown et al., 2004; Sinsabaugh & Follstad Shah, 2011; Manzoni et al., 2012; Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015).生态化学计量学是研究生物系统能量平衡和多重化学元素(主要是碳、氮、磷)平衡的科学(Sterner & Elser, 2002); 代谢是指有机体从环境中获取能量和物质资源, 以最优方式分配在存活、生长和繁殖上, 并将废物排泄回环境(Brown et al., 2004).这两者都结合了生物学、化学、物理学等基本原理, 可以跨越生物科学不同层次(从细胞新陈代谢到生态系统结构和养分循环)探索不同生态系统组分之间的联系与内在机理(Sterner & Elser, 2002; Brown et al., 2004; Elser, 2006). ...
... ).生态化学计量学是研究生物系统能量平衡和多重化学元素(主要是碳、氮、磷)平衡的科学(Sterner & Elser, 2002); 代谢是指有机体从环境中获取能量和物质资源, 以最优方式分配在存活、生长和繁殖上, 并将废物排泄回环境(Brown et al., 2004).这两者都结合了生物学、化学、物理学等基本原理, 可以跨越生物科学不同层次(从细胞新陈代谢到生态系统结构和养分循环)探索不同生态系统组分之间的联系与内在机理(Sterner & Elser, 2002; Brown et al., 2004; Elser, 2006). ...
... ).这两者都结合了生物学、化学、物理学等基本原理, 可以跨越生物科学不同层次(从细胞新陈代谢到生态系统结构和养分循环)探索不同生态系统组分之间的联系与内在机理(Sterner & Elser, 2002; Brown et al., 2004; Elser, 2006). ...
... 地球上所有生物的生长和繁殖都需要25种相同的化学元素, 但这些元素的相对比例(及化学计量)在生物体之间存在巨大差异; 这一方面取决于生物有机体自身的结构限制和代谢需求(Sterner & Elser, 2002; Brown et al., 2004), 另一方面也受环境影响而表现出一定环境可塑性(Sardans et al., 2012; Yang et al., 2015). ...
... 陆地生态系统中微生物的分解底物主要来源于植物的枯枝落叶、粗木质残体、根系凋落物和分泌物、土壤有机质等(Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015).在全球尺度上, 森林生态系统中植物叶片C:N:P为1212:28:1 (摩尔比, McGroddy et al., 2004).植物叶片在凋亡过程中的养分再吸收作用, 使得枯叶养分含量骤减, 从而使枯叶的C:N:P变为3007:45:1 (McGroddy et al., 2004; Yuan & Chen, 2009; Reed et al., 2012; Vergutz et al., 2012; Han et al., 2013).此外, 枯叶的C:N:P还会随生态系统、生物区系以及生活型的不同而发生显著变化(McGroddy et al., 2004).粗木质残体和枯立木的现存量占生态系统碎屑碳库的45% (Zhang & Wang, 2010); 粗木质残体占整个生态系统植物碎屑年输入量的10% (温带森林, Harmon et al., 1986)至30% (热带森林, Martinelli et al., 2000), 其C:N:P可高达14103:40:1 (Weedon et al., 2009).植物细根周转产生的凋落物是土壤碳和养分的重要来源, 其死根C:N:P的全球平均值为4184:43:1 (Yuan et al., 2011).土壤C:N:P最低(287:17:1, Xu et al., 2013), 且随着土壤深度增加而显著减小(Tian et al., 2010).相比之下, 土壤微生物生物量C:N:P比其分解底物低得多.例如: Cleveland和Liptzin (2007)整合全球数据时发现, 陆地生态系统土壤微生物生物量碳氮磷化学计量虽然存在类似于海洋浮游植物的“Redfield ratio” 在资源不受限制时浮游植物的C:N:P为106:16:1; Redfield, 1958), 即土壤微生物生物量C:N:P具有内稳性(homeostasis; Sterner & Elser, 2002), 但森林和草地的土壤微生物生物量C:N:P之间存在显著差异.近期Xu等(2013)进一步整合分析表明, 土壤微生物生物量C:N:P随生物区系不同有很大的变异性(从沙漠的31:4:1到天然湿地的131:14:1).面临底物和微生物化学计量的巨大变异性, 微生物主要通过调整自身化学计量特征、群落结构、胞外酶比例、元素利用效率等4种机制来响应和调节(图1). ...
... ).微生物会保留自身组成元素维持和代谢需求的限制性元素, 排出多余的元素, 这一过程被称为微生物消费者驱动的养分循环(consumer- driven nutrient recycling; Sterner & Elser, 2002).例如: 当微生物生长在低质量底物环境中时, 微生物会通过呼吸释放出过量的碳来满足自身养分需求的代谢活动, 这一现象被称为溢出代谢或溢出呼吸(overflow metabolism/respiration; Hessen et al., 2013; Schimel & Weintraub, 2003; Sinsabaugh et al., 2013).最新研究表明, 单位微生物生物量碳呼吸速率会随着底物C:X的增加而显著增加, 一定程度上反映了微生物的溢出呼吸(Spohn, 2015; Spohn & Chodak, 2015).但微生物呼吸的增加是否伴随着更多能量投入到胞外酶的生产上, 尚需要进一步验证(Hartman & Richardson, 2013; Spohn, 2015; Spohn & Chodak, 2015).相反.当微生物生长于高质量底物环境中时, 微生物氮磷矿化速率会随之增加; 这两者之间的相关性已被广泛证实(Manzoni et al., 2010; Moosha- mmer et al., 2012; Deng et al., 2014; Buchkowski et al., 2015).Zhou和Wang (2015)最近还发现, 微生物生物量碳在土壤总有机碳中所占的比例随土壤C:N的增加而显著减小, 但微生物生物量氮在土壤总氮中所占的比例却随土壤C:N的增加而显著增加, 这表明底物C:X还会影响微生物生物量大小以及土壤的元素化学计量. ...
... 通常将生物对底物C:X所能承受的限度定义为元素比率阈值(threshold elemental ratio).在微生物研究中, 元素比率阈值建立在代谢理论和化学计量理论基础上, 整合了微生物生物量C:N:P、胞外酶化学计量关系(BG:NAG:AP), 以及元素利用效率对底物C:N:P的适应策略(Sinsabaugh & Follstad Shah, 2012).如果底物C:X超过这个阈值, 则微生物受养分限制, 使其碳利用效率随底物C:X的增加而减小、养分利用效率会增加(Mooshammer et al., 2014; Zechmeister-Boltenstern et al., 2015); 反之, 当底物C:X低于这个阈值时, 微生物受到能量(碳)限制, 使其碳利用效率随底物C:X的增加而增加, 而养分利用效率会降低(Sterner & Elser, 2002; Frost et al. 2006; Doi et al. 2010; Sinsabaugh & Follstad Shah, 2012). ...
Litter quality is in the eye of the beholder: Initial decomposition rates as a function of inoculum characteristics.
1
2009
... 在微生物群落水平上, 自然选择会使那些能够充分获取并利用特定资源的种群在竞争中获胜(Moorhead & Sinsabaugh, 2006; Fierer et al., 2007; Strickland & Rousk, 2010; Martin et al., 2015; Zelezniak et al., 2015).因此, 底物的质量(高质量意味着低的碳与养分比(C:X), 低质量意味着高的C:X; Xu et al., 2014)的改变必然会改变微生物群落组成.目前很多学者认为, 微生物生物量C:N:P的可塑性是由微生物群落结构的变化导致的(Fanin et al., 2013; Kaiser et al., 2014; Mooshammer et al., 2014; Zechmeister-Boltenstern et al., 2015).生长和周转迅速的r对策微生物常具有较低的生物量C:X, 适应于高质量的底物; 相反, 生长和周转速度较慢的k对策微生物常具有较高的生物量C:X, 适应于低质量底物(Moorhead & Sinsabaugh, 2006; Fierer et al., 2007; Strickland & Rousk, 2010; Kaiser et al., 2014).然而, 由于微生物数量、群落组成以及代谢活动的复杂性, 微生物群落结构、生物量C:N:P与底物C:N:P之间的关系仍不明确.例如: 由于真菌的C:N比细菌高(Strickland & Rousk, 2010), 且真菌与细菌的比例以及底物C:N均随着土壤剖面而降低(Moore et al., 2010; Lee et al., 2013).Mooshammer等(2014)在分析微生物分解者对底物质量的适应机制时推测, 真菌与细菌比例应该随着底物C:N的增加而增加.Fanin等(2013)的确发现微生物生物量C:N与真菌与细菌比例存在显著的相关性, 但真菌与细菌比例与底物C:N:P并无相关性.然而, 同属于子囊菌门(Ascom- ycota)的刺盾炱纲(Chaetothyriomycetes)的真菌适应于低质量底物, 而锤舌菌纲(Leotiomycetes)和粪壳菌纲(Sordariomycetes)的真菌却适应于高质量的底物; 这表明, 即便是在真菌内部, 不同分类单元对底物质量的要求也不尽相同(Strickland et al., 2009; Sch- neider et al., 2012).因此, 微生物群落结构、生物量C:N:P与底物C:N:P之间普适性的关系还需验证. ...
Considering fungal: Bacterial dominance in soils-methods, controls, and ecosystem implications.
3
2010
... 在微生物群落水平上, 自然选择会使那些能够充分获取并利用特定资源的种群在竞争中获胜(Moorhead & Sinsabaugh, 2006; Fierer et al., 2007; Strickland & Rousk, 2010; Martin et al., 2015; Zelezniak et al., 2015).因此, 底物的质量(高质量意味着低的碳与养分比(C:X), 低质量意味着高的C:X; Xu et al., 2014)的改变必然会改变微生物群落组成.目前很多学者认为, 微生物生物量C:N:P的可塑性是由微生物群落结构的变化导致的(Fanin et al., 2013; Kaiser et al., 2014; Mooshammer et al., 2014; Zechmeister-Boltenstern et al., 2015).生长和周转迅速的r对策微生物常具有较低的生物量C:X, 适应于高质量的底物; 相反, 生长和周转速度较慢的k对策微生物常具有较高的生物量C:X, 适应于低质量底物(Moorhead & Sinsabaugh, 2006; Fierer et al., 2007; Strickland & Rousk, 2010; Kaiser et al., 2014).然而, 由于微生物数量、群落组成以及代谢活动的复杂性, 微生物群落结构、生物量C:N:P与底物C:N:P之间的关系仍不明确.例如: 由于真菌的C:N比细菌高(Strickland & Rousk, 2010), 且真菌与细菌的比例以及底物C:N均随着土壤剖面而降低(Moore et al., 2010; Lee et al., 2013).Mooshammer等(2014)在分析微生物分解者对底物质量的适应机制时推测, 真菌与细菌比例应该随着底物C:N的增加而增加.Fanin等(2013)的确发现微生物生物量C:N与真菌与细菌比例存在显著的相关性, 但真菌与细菌比例与底物C:N:P并无相关性.然而, 同属于子囊菌门(Ascom- ycota)的刺盾炱纲(Chaetothyriomycetes)的真菌适应于低质量底物, 而锤舌菌纲(Leotiomycetes)和粪壳菌纲(Sordariomycetes)的真菌却适应于高质量的底物; 这表明, 即便是在真菌内部, 不同分类单元对底物质量的要求也不尽相同(Strickland et al., 2009; Sch- neider et al., 2012).因此, 微生物群落结构、生物量C:N:P与底物C:N:P之间普适性的关系还需验证. ...
... ; Strickland & Rousk, 2010; Kaiser et al., 2014).然而, 由于微生物数量、群落组成以及代谢活动的复杂性, 微生物群落结构、生物量C:N:P与底物C:N:P之间的关系仍不明确.例如: 由于真菌的C:N比细菌高(Strickland & Rousk, 2010), 且真菌与细菌的比例以及底物C:N均随着土壤剖面而降低(Moore et al., 2010; Lee et al., 2013).Mooshammer等(2014)在分析微生物分解者对底物质量的适应机制时推测, 真菌与细菌比例应该随着底物C:N的增加而增加.Fanin等(2013)的确发现微生物生物量C:N与真菌与细菌比例存在显著的相关性, 但真菌与细菌比例与底物C:N:P并无相关性.然而, 同属于子囊菌门(Ascom- ycota)的刺盾炱纲(Chaetothyriomycetes)的真菌适应于低质量底物, 而锤舌菌纲(Leotiomycetes)和粪壳菌纲(Sordariomycetes)的真菌却适应于高质量的底物; 这表明, 即便是在真菌内部, 不同分类单元对底物质量的要求也不尽相同(Strickland et al., 2009; Sch- neider et al., 2012).因此, 微生物群落结构、生物量C:N:P与底物C:N:P之间普适性的关系还需验证. ...
... ).然而, 由于微生物数量、群落组成以及代谢活动的复杂性, 微生物群落结构、生物量C:N:P与底物C:N:P之间的关系仍不明确.例如: 由于真菌的C:N比细菌高(Strickland & Rousk, 2010), 且真菌与细菌的比例以及底物C:N均随着土壤剖面而降低(Moore et al., 2010; Lee et al., 2013).Mooshammer等(2014)在分析微生物分解者对底物质量的适应机制时推测, 真菌与细菌比例应该随着底物C:N的增加而增加.Fanin等(2013)的确发现微生物生物量C:N与真菌与细菌比例存在显著的相关性, 但真菌与细菌比例与底物C:N:P并无相关性.然而, 同属于子囊菌门(Ascom- ycota)的刺盾炱纲(Chaetothyriomycetes)的真菌适应于低质量底物, 而锤舌菌纲(Leotiomycetes)和粪壳菌纲(Sordariomycetes)的真菌却适应于高质量的底物; 这表明, 即便是在真菌内部, 不同分类单元对底物质量的要求也不尽相同(Strickland et al., 2009; Sch- neider et al., 2012).因此, 微生物群落结构、生物量C:N:P与底物C:N:P之间普适性的关系还需验证. ...
Forest succession on abandoned arable soils in European Russia-Impacts on microbial biomass, fungal-bacterial ratio, and basal CO2 respiration activity.
1
2011
... (3)探索微生物对底物化学计量变化适应对策的时空变化.Odum (1969)生态系统演替理论指出, 生态系统呼吸和生物量的比例会随着演替进程而减小.Insam和Haselwandter (1989)首次提出生态系统演替过程中微生物也有类似的变化趋势, 即单位微生物生物量的呼吸速率随着演替进程而减小.但Wardle和Ghani (1995)的整合研究发现大多数情况下单位微生物生物量的呼吸速率随着演替进程先减后增.此后相继出现大量研究, 但仍无统一结论和解释(Saviozzi et al., 2001; Goberna et al., 2006; Susyan et al., 2011; Lu et al., 2014; Raiesi & Beheshti, 2015).运用化学计量学理论可能有助于解释这些争议: 随着生态系统的演替进程, 一方面植物光合作用和微生物固氮作用将大气中的碳和氮素不断输入土壤, 从而导致土壤C:P和N:P的增加; 另一方面, 凋落物、地表枯落物层、土壤C:N随着演替进程基本保持不变(Yang & Luo, 2011; Yang et al., 2011; 周正虎等, 2015), 而单位微生物生物量的呼吸速率随着C:P的增加而增加(图1; Spohn & Chodak, 2015), 从而出现Wardle和Ghani (1995)的“先减后增”格局.另外, 普遍认为, 高纬度地区受氮限制, 而低纬度地区受磷限制; 生态系统演替初期受氮限制, 而后期受磷限制(Vitousek et al., 2010).然而, Spohn和Chodak (2015)在德国发现温带森林土壤C:P与单位微生物生物量的呼吸速率显著相关, 而与C:N不相关; 长期监测结果也显示, 该地区森林植物叶片的磷含量也在减小(Ilg et al., 2009).因此, 上述生态系统演替和全球变化(尤其是氮沉降)可能会导致生态系统的磷限制而突破磷会受纬度限制的传统认识(Vitousek et al., 2010). ...
Pattern and variation of C:N:P ratios in China’s soils: A synthesis of observational data.
1
2010
... 陆地生态系统中微生物的分解底物主要来源于植物的枯枝落叶、粗木质残体、根系凋落物和分泌物、土壤有机质等(Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015).在全球尺度上, 森林生态系统中植物叶片C:N:P为1212:28:1 (摩尔比, McGroddy et al., 2004).植物叶片在凋亡过程中的养分再吸收作用, 使得枯叶养分含量骤减, 从而使枯叶的C:N:P变为3007:45:1 (McGroddy et al., 2004; Yuan & Chen, 2009; Reed et al., 2012; Vergutz et al., 2012; Han et al., 2013).此外, 枯叶的C:N:P还会随生态系统、生物区系以及生活型的不同而发生显著变化(McGroddy et al., 2004).粗木质残体和枯立木的现存量占生态系统碎屑碳库的45% (Zhang & Wang, 2010); 粗木质残体占整个生态系统植物碎屑年输入量的10% (温带森林, Harmon et al., 1986)至30% (热带森林, Martinelli et al., 2000), 其C:N:P可高达14103:40:1 (Weedon et al., 2009).植物细根周转产生的凋落物是土壤碳和养分的重要来源, 其死根C:N:P的全球平均值为4184:43:1 (Yuan et al., 2011).土壤C:N:P最低(287:17:1, Xu et al., 2013), 且随着土壤深度增加而显著减小(Tian et al., 2010).相比之下, 土壤微生物生物量C:N:P比其分解底物低得多.例如: Cleveland和Liptzin (2007)整合全球数据时发现, 陆地生态系统土壤微生物生物量碳氮磷化学计量虽然存在类似于海洋浮游植物的“Redfield ratio” 在资源不受限制时浮游植物的C:N:P为106:16:1; Redfield, 1958), 即土壤微生物生物量C:N:P具有内稳性(homeostasis; Sterner & Elser, 2002), 但森林和草地的土壤微生物生物量C:N:P之间存在显著差异.近期Xu等(2013)进一步整合分析表明, 土壤微生物生物量C:N:P随生物区系不同有很大的变异性(从沙漠的31:4:1到天然湿地的131:14:1).面临底物和微生物化学计量的巨大变异性, 微生物主要通过调整自身化学计量特征、群落结构、胞外酶比例、元素利用效率等4种机制来响应和调节(图1). ...
The unseen majority: Soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems.
3
2008
... 微生物作为陆地生态系统主要分解者, 在获取资源构建自身生物量的同时, 驱动着生态系统物质和能量的流通, 调控着碳和养分在土壤-植物-大气连续体(SPAC)之间的循环, 进而影响生态系统的结构与功能(Chapin et al., 2003; Leff et al., 2015).尽管土壤微生物生物量碳只占土壤有机碳库的1.92% (Zhou & Wang, 2015), 但其分解作用消耗了生物圈的绝大部分净初级生产力(Cebrian & Lartigue, 2004; 王兴昌和王传宽, 2015).微生物代谢释放的CO2占土壤呼吸总通量的一半以上(Wang & Yang, 2007), 其矿化和固氮作用维持了土壤的肥力(van der Heijen et al., 2008; Leff et al., 2015); 同时, 土壤微生物与植物的共生、寄生和养分需求关系也会影响地上植被生产力和生物多样性.因此, 探索陆地生态系统地上植被组成和功能特征与地下微生物的耦合关系成为目前生态学的研究热点(van der Heijen et al., 2008; Barberán et al., 2015; Moore et al., 2015); 认识土壤微生物对碳氮磷循环的调控机制也是全球变化背景下生物地球化学循环研究的重要内容(Sinsabaugh & Follstad Shah, 2011; Mooshammer et al., 2014; Zechmeister-Boltenstern et al., 2015). ...
... ); 同时, 土壤微生物与植物的共生、寄生和养分需求关系也会影响地上植被生产力和生物多样性.因此, 探索陆地生态系统地上植被组成和功能特征与地下微生物的耦合关系成为目前生态学的研究热点(van der Heijen et al., 2008; Barberán et al., 2015; Moore et al., 2015); 认识土壤微生物对碳氮磷循环的调控机制也是全球变化背景下生物地球化学循环研究的重要内容(Sinsabaugh & Follstad Shah, 2011; Mooshammer et al., 2014; Zechmeister-Boltenstern et al., 2015). ...
... 氮和磷是陆地生态系统生产力的关键限制元素(Vitousek & Howarth, 1991; Vitousek et al., 2010).微生物分解者主要通过驱动如下过程来调节陆地生态系统氮循环: 通过共生固氮、自生固氮、联合固氮等将大气中分子氮还原为铵态氮; 优先吸收铵态氮后同化为有机氮; 通过胞外酶将资源中有机氮转化为无机氮的过程(即矿化作用); 通过自养硝化细菌的硝化作用将氨氧化为硝态氮; 通过反硝化细菌的反硝化作用将硝态氮还原为氮气或氧化亚氮重新进入大气圈(Jetten, 2008; Rick & Thomas, 2001).尽管微生物不能像固氮作用一样增加土壤磷总含量, 但土壤微生物的活动会显著改变土壤中磷的化学形式和比例, 从而在调节植物磷可利用性中扮演重要角色(van der Heijden et al., 2008).例如: 微生物分解有机质释放有效磷; 在磷限制的生态系统中, 微生物生物量磷占土壤总磷的比例可达到23.6% (天然湿地生态系统Xu et al., 2013); 又因为土壤微生物生物量的最大值出现在冬季, 使得在植被生长季中土壤微生物生物量减小时释放出磷, 及时增加土壤中磷的可利用性(Richardson & Simpson, 2011; Xu et al., 2013).另外, 土壤微生物的存在还会改变土壤的吸附平衡, 从而增加正磷酸盐向土壤溶液中的转移(Seeling & Zasoski, 1993).由此可见, 微生物分解者在陆地生态系统生物地球化学循环中起着重要的驱动和调节作用. ...
Global resorption efficiencies and concentrations of carbon and nutrients in leaves of terrestrial plants.
1
2012
... 陆地生态系统中微生物的分解底物主要来源于植物的枯枝落叶、粗木质残体、根系凋落物和分泌物、土壤有机质等(Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015).在全球尺度上, 森林生态系统中植物叶片C:N:P为1212:28:1 (摩尔比, McGroddy et al., 2004).植物叶片在凋亡过程中的养分再吸收作用, 使得枯叶养分含量骤减, 从而使枯叶的C:N:P变为3007:45:1 (McGroddy et al., 2004; Yuan & Chen, 2009; Reed et al., 2012; Vergutz et al., 2012; Han et al., 2013).此外, 枯叶的C:N:P还会随生态系统、生物区系以及生活型的不同而发生显著变化(McGroddy et al., 2004).粗木质残体和枯立木的现存量占生态系统碎屑碳库的45% (Zhang & Wang, 2010); 粗木质残体占整个生态系统植物碎屑年输入量的10% (温带森林, Harmon et al., 1986)至30% (热带森林, Martinelli et al., 2000), 其C:N:P可高达14103:40:1 (Weedon et al., 2009).植物细根周转产生的凋落物是土壤碳和养分的重要来源, 其死根C:N:P的全球平均值为4184:43:1 (Yuan et al., 2011).土壤C:N:P最低(287:17:1, Xu et al., 2013), 且随着土壤深度增加而显著减小(Tian et al., 2010).相比之下, 土壤微生物生物量C:N:P比其分解底物低得多.例如: Cleveland和Liptzin (2007)整合全球数据时发现, 陆地生态系统土壤微生物生物量碳氮磷化学计量虽然存在类似于海洋浮游植物的“Redfield ratio” 在资源不受限制时浮游植物的C:N:P为106:16:1; Redfield, 1958), 即土壤微生物生物量C:N:P具有内稳性(homeostasis; Sterner & Elser, 2002), 但森林和草地的土壤微生物生物量C:N:P之间存在显著差异.近期Xu等(2013)进一步整合分析表明, 土壤微生物生物量C:N:P随生物区系不同有很大的变异性(从沙漠的31:4:1到天然湿地的131:14:1).面临底物和微生物化学计量的巨大变异性, 微生物主要通过调整自身化学计量特征、群落结构、胞外酶比例、元素利用效率等4种机制来响应和调节(图1). ...
Nitrogen limitation on land and in the sea: How can it occur?
1
1991
... 氮和磷是陆地生态系统生产力的关键限制元素(Vitousek & Howarth, 1991; Vitousek et al., 2010).微生物分解者主要通过驱动如下过程来调节陆地生态系统氮循环: 通过共生固氮、自生固氮、联合固氮等将大气中分子氮还原为铵态氮; 优先吸收铵态氮后同化为有机氮; 通过胞外酶将资源中有机氮转化为无机氮的过程(即矿化作用); 通过自养硝化细菌的硝化作用将氨氧化为硝态氮; 通过反硝化细菌的反硝化作用将硝态氮还原为氮气或氧化亚氮重新进入大气圈(Jetten, 2008; Rick & Thomas, 2001).尽管微生物不能像固氮作用一样增加土壤磷总含量, 但土壤微生物的活动会显著改变土壤中磷的化学形式和比例, 从而在调节植物磷可利用性中扮演重要角色(van der Heijden et al., 2008).例如: 微生物分解有机质释放有效磷; 在磷限制的生态系统中, 微生物生物量磷占土壤总磷的比例可达到23.6% (天然湿地生态系统Xu et al., 2013); 又因为土壤微生物生物量的最大值出现在冬季, 使得在植被生长季中土壤微生物生物量减小时释放出磷, 及时增加土壤中磷的可利用性(Richardson & Simpson, 2011; Xu et al., 2013).另外, 土壤微生物的存在还会改变土壤的吸附平衡, 从而增加正磷酸盐向土壤溶液中的转移(Seeling & Zasoski, 1993).由此可见, 微生物分解者在陆地生态系统生物地球化学循环中起着重要的驱动和调节作用. ...
Terrestrial phosphorus limitation: Mechanisms implications and nitrogen-phosphorus interactions.
3
2010
... 氮和磷是陆地生态系统生产力的关键限制元素(Vitousek & Howarth, 1991; Vitousek et al., 2010).微生物分解者主要通过驱动如下过程来调节陆地生态系统氮循环: 通过共生固氮、自生固氮、联合固氮等将大气中分子氮还原为铵态氮; 优先吸收铵态氮后同化为有机氮; 通过胞外酶将资源中有机氮转化为无机氮的过程(即矿化作用); 通过自养硝化细菌的硝化作用将氨氧化为硝态氮; 通过反硝化细菌的反硝化作用将硝态氮还原为氮气或氧化亚氮重新进入大气圈(Jetten, 2008; Rick & Thomas, 2001).尽管微生物不能像固氮作用一样增加土壤磷总含量, 但土壤微生物的活动会显著改变土壤中磷的化学形式和比例, 从而在调节植物磷可利用性中扮演重要角色(van der Heijden et al., 2008).例如: 微生物分解有机质释放有效磷; 在磷限制的生态系统中, 微生物生物量磷占土壤总磷的比例可达到23.6% (天然湿地生态系统Xu et al., 2013); 又因为土壤微生物生物量的最大值出现在冬季, 使得在植被生长季中土壤微生物生物量减小时释放出磷, 及时增加土壤中磷的可利用性(Richardson & Simpson, 2011; Xu et al., 2013).另外, 土壤微生物的存在还会改变土壤的吸附平衡, 从而增加正磷酸盐向土壤溶液中的转移(Seeling & Zasoski, 1993).由此可见, 微生物分解者在陆地生态系统生物地球化学循环中起着重要的驱动和调节作用. ...
... (3)探索微生物对底物化学计量变化适应对策的时空变化.Odum (1969)生态系统演替理论指出, 生态系统呼吸和生物量的比例会随着演替进程而减小.Insam和Haselwandter (1989)首次提出生态系统演替过程中微生物也有类似的变化趋势, 即单位微生物生物量的呼吸速率随着演替进程而减小.但Wardle和Ghani (1995)的整合研究发现大多数情况下单位微生物生物量的呼吸速率随着演替进程先减后增.此后相继出现大量研究, 但仍无统一结论和解释(Saviozzi et al., 2001; Goberna et al., 2006; Susyan et al., 2011; Lu et al., 2014; Raiesi & Beheshti, 2015).运用化学计量学理论可能有助于解释这些争议: 随着生态系统的演替进程, 一方面植物光合作用和微生物固氮作用将大气中的碳和氮素不断输入土壤, 从而导致土壤C:P和N:P的增加; 另一方面, 凋落物、地表枯落物层、土壤C:N随着演替进程基本保持不变(Yang & Luo, 2011; Yang et al., 2011; 周正虎等, 2015), 而单位微生物生物量的呼吸速率随着C:P的增加而增加(图1; Spohn & Chodak, 2015), 从而出现Wardle和Ghani (1995)的“先减后增”格局.另外, 普遍认为, 高纬度地区受氮限制, 而低纬度地区受磷限制; 生态系统演替初期受氮限制, 而后期受磷限制(Vitousek et al., 2010).然而, Spohn和Chodak (2015)在德国发现温带森林土壤C:P与单位微生物生物量的呼吸速率显著相关, 而与C:N不相关; 长期监测结果也显示, 该地区森林植物叶片的磷含量也在减小(Ilg et al., 2009).因此, 上述生态系统演替和全球变化(尤其是氮沉降)可能会导致生态系统的磷限制而突破磷会受纬度限制的传统认识(Vitousek et al., 2010). ...
... ).因此, 上述生态系统演替和全球变化(尤其是氮沉降)可能会导致生态系统的磷限制而突破磷会受纬度限制的传统认识(Vitousek et al., 2010). ...
Seasonality of soil CO2 efflux in a temperate forest: Biophysical effects of snowpack and spring freeze-thaw cycles.
1
2013
... 土壤呼吸是陆地生态系统向大气圈排放的最大碳源, 其中以土壤微生物代谢和生长释放出的CO2为主的异养呼吸占总土壤呼吸一半以上(Högberg et al., 2001; Bond-Lamberty et al., 2004; Wang & Yang, 2007), 因而在全球变化和生态系统研究中备受关注(Wang et al., 2006).然而, 以往研究多关注土壤呼吸组分的划分以及与土壤温湿度、有机碳储量、植被覆盖、土壤微生物生物量、环境因子等之间的相关性(Högberg et al., 2001; Bond-Lamberty et al., 2004; Wang & Yang, 2007; Wang et al., 2013), 但可能由于对微生物生理代谢机制认识不足(Allison et al., 2010; Buchkowski et al., 2015; Steinauer et al., 2015), 未能阐明异养呼吸对复杂环境因子的响应机制(Bardgett et al., 2008).微生物驱动的异养呼吸与微生物生长所需能量投资密切相关, 关系到微生物在呼吸代谢和生物量生长之间的碳资源分配.因此, 微生物将底物碳转化为自身生物量碳的效率(即微生物碳利用效率, CUE; del Giorgio & Cole, 1998)与陆地生态系统碳循环直接相关(Manzoni et al., 2012; Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015). ...
Rhizospheric and heterotrophic components of soil respiration in six Chinese temperate forests.
3
2007
... 微生物作为陆地生态系统主要分解者, 在获取资源构建自身生物量的同时, 驱动着生态系统物质和能量的流通, 调控着碳和养分在土壤-植物-大气连续体(SPAC)之间的循环, 进而影响生态系统的结构与功能(Chapin et al., 2003; Leff et al., 2015).尽管土壤微生物生物量碳只占土壤有机碳库的1.92% (Zhou & Wang, 2015), 但其分解作用消耗了生物圈的绝大部分净初级生产力(Cebrian & Lartigue, 2004; 王兴昌和王传宽, 2015).微生物代谢释放的CO2占土壤呼吸总通量的一半以上(Wang & Yang, 2007), 其矿化和固氮作用维持了土壤的肥力(van der Heijen et al., 2008; Leff et al., 2015); 同时, 土壤微生物与植物的共生、寄生和养分需求关系也会影响地上植被生产力和生物多样性.因此, 探索陆地生态系统地上植被组成和功能特征与地下微生物的耦合关系成为目前生态学的研究热点(van der Heijen et al., 2008; Barberán et al., 2015; Moore et al., 2015); 认识土壤微生物对碳氮磷循环的调控机制也是全球变化背景下生物地球化学循环研究的重要内容(Sinsabaugh & Follstad Shah, 2011; Mooshammer et al., 2014; Zechmeister-Boltenstern et al., 2015). ...
... 土壤呼吸是陆地生态系统向大气圈排放的最大碳源, 其中以土壤微生物代谢和生长释放出的CO2为主的异养呼吸占总土壤呼吸一半以上(Högberg et al., 2001; Bond-Lamberty et al., 2004; Wang & Yang, 2007), 因而在全球变化和生态系统研究中备受关注(Wang et al., 2006).然而, 以往研究多关注土壤呼吸组分的划分以及与土壤温湿度、有机碳储量、植被覆盖、土壤微生物生物量、环境因子等之间的相关性(Högberg et al., 2001; Bond-Lamberty et al., 2004; Wang & Yang, 2007; Wang et al., 2013), 但可能由于对微生物生理代谢机制认识不足(Allison et al., 2010; Buchkowski et al., 2015; Steinauer et al., 2015), 未能阐明异养呼吸对复杂环境因子的响应机制(Bardgett et al., 2008).微生物驱动的异养呼吸与微生物生长所需能量投资密切相关, 关系到微生物在呼吸代谢和生物量生长之间的碳资源分配.因此, 微生物将底物碳转化为自身生物量碳的效率(即微生物碳利用效率, CUE; del Giorgio & Cole, 1998)与陆地生态系统碳循环直接相关(Manzoni et al., 2012; Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015). ...
... ; Wang & Yang, 2007; Wang et al., 2013), 但可能由于对微生物生理代谢机制认识不足(Allison et al., 2010; Buchkowski et al., 2015; Steinauer et al., 2015), 未能阐明异养呼吸对复杂环境因子的响应机制(Bardgett et al., 2008).微生物驱动的异养呼吸与微生物生长所需能量投资密切相关, 关系到微生物在呼吸代谢和生物量生长之间的碳资源分配.因此, 微生物将底物碳转化为自身生物量碳的效率(即微生物碳利用效率, CUE; del Giorgio & Cole, 1998)与陆地生态系统碳循环直接相关(Manzoni et al., 2012; Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015). ...
Soil respiration in six temperate forests in China.
1
2006
... 土壤呼吸是陆地生态系统向大气圈排放的最大碳源, 其中以土壤微生物代谢和生长释放出的CO2为主的异养呼吸占总土壤呼吸一半以上(Högberg et al., 2001; Bond-Lamberty et al., 2004; Wang & Yang, 2007), 因而在全球变化和生态系统研究中备受关注(Wang et al., 2006).然而, 以往研究多关注土壤呼吸组分的划分以及与土壤温湿度、有机碳储量、植被覆盖、土壤微生物生物量、环境因子等之间的相关性(Högberg et al., 2001; Bond-Lamberty et al., 2004; Wang & Yang, 2007; Wang et al., 2013), 但可能由于对微生物生理代谢机制认识不足(Allison et al., 2010; Buchkowski et al., 2015; Steinauer et al., 2015), 未能阐明异养呼吸对复杂环境因子的响应机制(Bardgett et al., 2008).微生物驱动的异养呼吸与微生物生长所需能量投资密切相关, 关系到微生物在呼吸代谢和生物量生长之间的碳资源分配.因此, 微生物将底物碳转化为自身生物量碳的效率(即微生物碳利用效率, CUE; del Giorgio & Cole, 1998)与陆地生态系统碳循环直接相关(Manzoni et al., 2012; Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015). ...
... 陆地生态系统中微生物的分解底物主要来源于植物的枯枝落叶、粗木质残体、根系凋落物和分泌物、土壤有机质等(Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015).在全球尺度上, 森林生态系统中植物叶片C:N:P为1212:28:1 (摩尔比, McGroddy et al., 2004).植物叶片在凋亡过程中的养分再吸收作用, 使得枯叶养分含量骤减, 从而使枯叶的C:N:P变为3007:45:1 (McGroddy et al., 2004; Yuan & Chen, 2009; Reed et al., 2012; Vergutz et al., 2012; Han et al., 2013).此外, 枯叶的C:N:P还会随生态系统、生物区系以及生活型的不同而发生显著变化(McGroddy et al., 2004).粗木质残体和枯立木的现存量占生态系统碎屑碳库的45% (Zhang & Wang, 2010); 粗木质残体占整个生态系统植物碎屑年输入量的10% (温带森林, Harmon et al., 1986)至30% (热带森林, Martinelli et al., 2000), 其C:N:P可高达14103:40:1 (Weedon et al., 2009).植物细根周转产生的凋落物是土壤碳和养分的重要来源, 其死根C:N:P的全球平均值为4184:43:1 (Yuan et al., 2011).土壤C:N:P最低(287:17:1, Xu et al., 2013), 且随着土壤深度增加而显著减小(Tian et al., 2010).相比之下, 土壤微生物生物量C:N:P比其分解底物低得多.例如: Cleveland和Liptzin (2007)整合全球数据时发现, 陆地生态系统土壤微生物生物量碳氮磷化学计量虽然存在类似于海洋浮游植物的“Redfield ratio” 在资源不受限制时浮游植物的C:N:P为106:16:1; Redfield, 1958), 即土壤微生物生物量C:N:P具有内稳性(homeostasis; Sterner & Elser, 2002), 但森林和草地的土壤微生物生物量C:N:P之间存在显著差异.近期Xu等(2013)进一步整合分析表明, 土壤微生物生物量C:N:P随生物区系不同有很大的变异性(从沙漠的31:4:1到天然湿地的131:14:1).面临底物和微生物化学计量的巨大变异性, 微生物主要通过调整自身化学计量特征、群落结构、胞外酶比例、元素利用效率等4种机制来响应和调节(图1). ...
C:N:P stoichiometry of Artemisia species and close relatives across northern China: Unravelling effects of climate, soil and taxonomy.
1
2015
... 地球上所有生物的生长和繁殖都需要25种相同的化学元素, 但这些元素的相对比例(及化学计量)在生物体之间存在巨大差异; 这一方面取决于生物有机体自身的结构限制和代谢需求(Sterner & Elser, 2002; Brown et al., 2004), 另一方面也受环境影响而表现出一定环境可塑性(Sardans et al., 2012; Yang et al., 2015). ...
Carbon:nitrogen stoichiometry in forest ecosystems during stand development.
1
2011
... (3)探索微生物对底物化学计量变化适应对策的时空变化.Odum (1969)生态系统演替理论指出, 生态系统呼吸和生物量的比例会随着演替进程而减小.Insam和Haselwandter (1989)首次提出生态系统演替过程中微生物也有类似的变化趋势, 即单位微生物生物量的呼吸速率随着演替进程而减小.但Wardle和Ghani (1995)的整合研究发现大多数情况下单位微生物生物量的呼吸速率随着演替进程先减后增.此后相继出现大量研究, 但仍无统一结论和解释(Saviozzi et al., 2001; Goberna et al., 2006; Susyan et al., 2011; Lu et al., 2014; Raiesi & Beheshti, 2015).运用化学计量学理论可能有助于解释这些争议: 随着生态系统的演替进程, 一方面植物光合作用和微生物固氮作用将大气中的碳和氮素不断输入土壤, 从而导致土壤C:P和N:P的增加; 另一方面, 凋落物、地表枯落物层、土壤C:N随着演替进程基本保持不变(Yang & Luo, 2011; Yang et al., 2011; 周正虎等, 2015), 而单位微生物生物量的呼吸速率随着C:P的增加而增加(图1; Spohn & Chodak, 2015), 从而出现Wardle和Ghani (1995)的“先减后增”格局.另外, 普遍认为, 高纬度地区受氮限制, 而低纬度地区受磷限制; 生态系统演替初期受氮限制, 而后期受磷限制(Vitousek et al., 2010).然而, Spohn和Chodak (2015)在德国发现温带森林土壤C:P与单位微生物生物量的呼吸速率显著相关, 而与C:N不相关; 长期监测结果也显示, 该地区森林植物叶片的磷含量也在减小(Ilg et al., 2009).因此, 上述生态系统演替和全球变化(尤其是氮沉降)可能会导致生态系统的磷限制而突破磷会受纬度限制的传统认识(Vitousek et al., 2010). ...
Carbon and nitrogen dynamics during forest stand development: A global synthesis.
1
2011
... (3)探索微生物对底物化学计量变化适应对策的时空变化.Odum (1969)生态系统演替理论指出, 生态系统呼吸和生物量的比例会随着演替进程而减小.Insam和Haselwandter (1989)首次提出生态系统演替过程中微生物也有类似的变化趋势, 即单位微生物生物量的呼吸速率随着演替进程而减小.但Wardle和Ghani (1995)的整合研究发现大多数情况下单位微生物生物量的呼吸速率随着演替进程先减后增.此后相继出现大量研究, 但仍无统一结论和解释(Saviozzi et al., 2001; Goberna et al., 2006; Susyan et al., 2011; Lu et al., 2014; Raiesi & Beheshti, 2015).运用化学计量学理论可能有助于解释这些争议: 随着生态系统的演替进程, 一方面植物光合作用和微生物固氮作用将大气中的碳和氮素不断输入土壤, 从而导致土壤C:P和N:P的增加; 另一方面, 凋落物、地表枯落物层、土壤C:N随着演替进程基本保持不变(Yang & Luo, 2011; Yang et al., 2011; 周正虎等, 2015), 而单位微生物生物量的呼吸速率随着C:P的增加而增加(图1; Spohn & Chodak, 2015), 从而出现Wardle和Ghani (1995)的“先减后增”格局.另外, 普遍认为, 高纬度地区受氮限制, 而低纬度地区受磷限制; 生态系统演替初期受氮限制, 而后期受磷限制(Vitousek et al., 2010).然而, Spohn和Chodak (2015)在德国发现温带森林土壤C:P与单位微生物生物量的呼吸速率显著相关, 而与C:N不相关; 长期监测结果也显示, 该地区森林植物叶片的磷含量也在减小(Ilg et al., 2009).因此, 上述生态系统演替和全球变化(尤其是氮沉降)可能会导致生态系统的磷限制而突破磷会受纬度限制的传统认识(Vitousek et al., 2010). ...
Global-scale patterns of nutrient resorption associated with latitude, temperature and precipitation.
1
2009
... 陆地生态系统中微生物的分解底物主要来源于植物的枯枝落叶、粗木质残体、根系凋落物和分泌物、土壤有机质等(Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015).在全球尺度上, 森林生态系统中植物叶片C:N:P为1212:28:1 (摩尔比, McGroddy et al., 2004).植物叶片在凋亡过程中的养分再吸收作用, 使得枯叶养分含量骤减, 从而使枯叶的C:N:P变为3007:45:1 (McGroddy et al., 2004; Yuan & Chen, 2009; Reed et al., 2012; Vergutz et al., 2012; Han et al., 2013).此外, 枯叶的C:N:P还会随生态系统、生物区系以及生活型的不同而发生显著变化(McGroddy et al., 2004).粗木质残体和枯立木的现存量占生态系统碎屑碳库的45% (Zhang & Wang, 2010); 粗木质残体占整个生态系统植物碎屑年输入量的10% (温带森林, Harmon et al., 1986)至30% (热带森林, Martinelli et al., 2000), 其C:N:P可高达14103:40:1 (Weedon et al., 2009).植物细根周转产生的凋落物是土壤碳和养分的重要来源, 其死根C:N:P的全球平均值为4184:43:1 (Yuan et al., 2011).土壤C:N:P最低(287:17:1, Xu et al., 2013), 且随着土壤深度增加而显著减小(Tian et al., 2010).相比之下, 土壤微生物生物量C:N:P比其分解底物低得多.例如: Cleveland和Liptzin (2007)整合全球数据时发现, 陆地生态系统土壤微生物生物量碳氮磷化学计量虽然存在类似于海洋浮游植物的“Redfield ratio” 在资源不受限制时浮游植物的C:N:P为106:16:1; Redfield, 1958), 即土壤微生物生物量C:N:P具有内稳性(homeostasis; Sterner & Elser, 2002), 但森林和草地的土壤微生物生物量C:N:P之间存在显著差异.近期Xu等(2013)进一步整合分析表明, 土壤微生物生物量C:N:P随生物区系不同有很大的变异性(从沙漠的31:4:1到天然湿地的131:14:1).面临底物和微生物化学计量的巨大变异性, 微生物主要通过调整自身化学计量特征、群落结构、胞外酶比例、元素利用效率等4种机制来响应和调节(图1). ...
Global-scale latitudinal patterns of plant fine-root nitrogen and phosphorus.
1
2011
... 陆地生态系统中微生物的分解底物主要来源于植物的枯枝落叶、粗木质残体、根系凋落物和分泌物、土壤有机质等(Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015).在全球尺度上, 森林生态系统中植物叶片C:N:P为1212:28:1 (摩尔比, McGroddy et al., 2004).植物叶片在凋亡过程中的养分再吸收作用, 使得枯叶养分含量骤减, 从而使枯叶的C:N:P变为3007:45:1 (McGroddy et al., 2004; Yuan & Chen, 2009; Reed et al., 2012; Vergutz et al., 2012; Han et al., 2013).此外, 枯叶的C:N:P还会随生态系统、生物区系以及生活型的不同而发生显著变化(McGroddy et al., 2004).粗木质残体和枯立木的现存量占生态系统碎屑碳库的45% (Zhang & Wang, 2010); 粗木质残体占整个生态系统植物碎屑年输入量的10% (温带森林, Harmon et al., 1986)至30% (热带森林, Martinelli et al., 2000), 其C:N:P可高达14103:40:1 (Weedon et al., 2009).植物细根周转产生的凋落物是土壤碳和养分的重要来源, 其死根C:N:P的全球平均值为4184:43:1 (Yuan et al., 2011).土壤C:N:P最低(287:17:1, Xu et al., 2013), 且随着土壤深度增加而显著减小(Tian et al., 2010).相比之下, 土壤微生物生物量C:N:P比其分解底物低得多.例如: Cleveland和Liptzin (2007)整合全球数据时发现, 陆地生态系统土壤微生物生物量碳氮磷化学计量虽然存在类似于海洋浮游植物的“Redfield ratio” 在资源不受限制时浮游植物的C:N:P为106:16:1; Redfield, 1958), 即土壤微生物生物量C:N:P具有内稳性(homeostasis; Sterner & Elser, 2002), 但森林和草地的土壤微生物生物量C:N:P之间存在显著差异.近期Xu等(2013)进一步整合分析表明, 土壤微生物生物量C:N:P随生物区系不同有很大的变异性(从沙漠的31:4:1到天然湿地的131:14:1).面临底物和微生物化学计量的巨大变异性, 微生物主要通过调整自身化学计量特征、群落结构、胞外酶比例、元素利用效率等4种机制来响应和调节(图1). ...
The application of ecological stoichiometry to plant-microbial- soil organic matter transformations.
7
2015
... 微生物作为陆地生态系统主要分解者, 在获取资源构建自身生物量的同时, 驱动着生态系统物质和能量的流通, 调控着碳和养分在土壤-植物-大气连续体(SPAC)之间的循环, 进而影响生态系统的结构与功能(Chapin et al., 2003; Leff et al., 2015).尽管土壤微生物生物量碳只占土壤有机碳库的1.92% (Zhou & Wang, 2015), 但其分解作用消耗了生物圈的绝大部分净初级生产力(Cebrian & Lartigue, 2004; 王兴昌和王传宽, 2015).微生物代谢释放的CO2占土壤呼吸总通量的一半以上(Wang & Yang, 2007), 其矿化和固氮作用维持了土壤的肥力(van der Heijen et al., 2008; Leff et al., 2015); 同时, 土壤微生物与植物的共生、寄生和养分需求关系也会影响地上植被生产力和生物多样性.因此, 探索陆地生态系统地上植被组成和功能特征与地下微生物的耦合关系成为目前生态学的研究热点(van der Heijen et al., 2008; Barberán et al., 2015; Moore et al., 2015); 认识土壤微生物对碳氮磷循环的调控机制也是全球变化背景下生物地球化学循环研究的重要内容(Sinsabaugh & Follstad Shah, 2011; Mooshammer et al., 2014; Zechmeister-Boltenstern et al., 2015). ...
... 随着微生物群落鉴定技术的突破和完善、分子生物学和同位素示踪在土壤微生物研究中的应用, 以土壤微生物为核心的生态系统变化过程与机理研究正蓬勃发展.由于土壤微生物个体微小且数量庞大, 其系统发生、生物多样性及其与生物和非生物因素的相互作用远比动物和植物复杂, 因而微生物生态学研究面临着巨大挑战(Prosser et al., 2007; Jansson & Prosser, 2013).运用生态化学计量学理论(Sterner & Elser, 2002)和代谢理论(Brown et al., 2004)研究土壤微生物, 不但可以提升我们对土壤微生物生态过程和机理的认识, 而且有助于生态学理论的发展与完善(Sterner & Elser, 2002; Brown et al., 2004; Sinsabaugh & Follstad Shah, 2011; Manzoni et al., 2012; Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015).生态化学计量学是研究生物系统能量平衡和多重化学元素(主要是碳、氮、磷)平衡的科学(Sterner & Elser, 2002); 代谢是指有机体从环境中获取能量和物质资源, 以最优方式分配在存活、生长和繁殖上, 并将废物排泄回环境(Brown et al., 2004).这两者都结合了生物学、化学、物理学等基本原理, 可以跨越生物科学不同层次(从细胞新陈代谢到生态系统结构和养分循环)探索不同生态系统组分之间的联系与内在机理(Sterner & Elser, 2002; Brown et al., 2004; Elser, 2006). ...
... 土壤呼吸是陆地生态系统向大气圈排放的最大碳源, 其中以土壤微生物代谢和生长释放出的CO2为主的异养呼吸占总土壤呼吸一半以上(Högberg et al., 2001; Bond-Lamberty et al., 2004; Wang & Yang, 2007), 因而在全球变化和生态系统研究中备受关注(Wang et al., 2006).然而, 以往研究多关注土壤呼吸组分的划分以及与土壤温湿度、有机碳储量、植被覆盖、土壤微生物生物量、环境因子等之间的相关性(Högberg et al., 2001; Bond-Lamberty et al., 2004; Wang & Yang, 2007; Wang et al., 2013), 但可能由于对微生物生理代谢机制认识不足(Allison et al., 2010; Buchkowski et al., 2015; Steinauer et al., 2015), 未能阐明异养呼吸对复杂环境因子的响应机制(Bardgett et al., 2008).微生物驱动的异养呼吸与微生物生长所需能量投资密切相关, 关系到微生物在呼吸代谢和生物量生长之间的碳资源分配.因此, 微生物将底物碳转化为自身生物量碳的效率(即微生物碳利用效率, CUE; del Giorgio & Cole, 1998)与陆地生态系统碳循环直接相关(Manzoni et al., 2012; Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015). ...
... 陆地生态系统中微生物的分解底物主要来源于植物的枯枝落叶、粗木质残体、根系凋落物和分泌物、土壤有机质等(Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015).在全球尺度上, 森林生态系统中植物叶片C:N:P为1212:28:1 (摩尔比, McGroddy et al., 2004).植物叶片在凋亡过程中的养分再吸收作用, 使得枯叶养分含量骤减, 从而使枯叶的C:N:P变为3007:45:1 (McGroddy et al., 2004; Yuan & Chen, 2009; Reed et al., 2012; Vergutz et al., 2012; Han et al., 2013).此外, 枯叶的C:N:P还会随生态系统、生物区系以及生活型的不同而发生显著变化(McGroddy et al., 2004).粗木质残体和枯立木的现存量占生态系统碎屑碳库的45% (Zhang & Wang, 2010); 粗木质残体占整个生态系统植物碎屑年输入量的10% (温带森林, Harmon et al., 1986)至30% (热带森林, Martinelli et al., 2000), 其C:N:P可高达14103:40:1 (Weedon et al., 2009).植物细根周转产生的凋落物是土壤碳和养分的重要来源, 其死根C:N:P的全球平均值为4184:43:1 (Yuan et al., 2011).土壤C:N:P最低(287:17:1, Xu et al., 2013), 且随着土壤深度增加而显著减小(Tian et al., 2010).相比之下, 土壤微生物生物量C:N:P比其分解底物低得多.例如: Cleveland和Liptzin (2007)整合全球数据时发现, 陆地生态系统土壤微生物生物量碳氮磷化学计量虽然存在类似于海洋浮游植物的“Redfield ratio” 在资源不受限制时浮游植物的C:N:P为106:16:1; Redfield, 1958), 即土壤微生物生物量C:N:P具有内稳性(homeostasis; Sterner & Elser, 2002), 但森林和草地的土壤微生物生物量C:N:P之间存在显著差异.近期Xu等(2013)进一步整合分析表明, 土壤微生物生物量C:N:P随生物区系不同有很大的变异性(从沙漠的31:4:1到天然湿地的131:14:1).面临底物和微生物化学计量的巨大变异性, 微生物主要通过调整自身化学计量特征、群落结构、胞外酶比例、元素利用效率等4种机制来响应和调节(图1). ...
... 在微生物群落水平上, 自然选择会使那些能够充分获取并利用特定资源的种群在竞争中获胜(Moorhead & Sinsabaugh, 2006; Fierer et al., 2007; Strickland & Rousk, 2010; Martin et al., 2015; Zelezniak et al., 2015).因此, 底物的质量(高质量意味着低的碳与养分比(C:X), 低质量意味着高的C:X; Xu et al., 2014)的改变必然会改变微生物群落组成.目前很多学者认为, 微生物生物量C:N:P的可塑性是由微生物群落结构的变化导致的(Fanin et al., 2013; Kaiser et al., 2014; Mooshammer et al., 2014; Zechmeister-Boltenstern et al., 2015).生长和周转迅速的r对策微生物常具有较低的生物量C:X, 适应于高质量的底物; 相反, 生长和周转速度较慢的k对策微生物常具有较高的生物量C:X, 适应于低质量底物(Moorhead & Sinsabaugh, 2006; Fierer et al., 2007; Strickland & Rousk, 2010; Kaiser et al., 2014).然而, 由于微生物数量、群落组成以及代谢活动的复杂性, 微生物群落结构、生物量C:N:P与底物C:N:P之间的关系仍不明确.例如: 由于真菌的C:N比细菌高(Strickland & Rousk, 2010), 且真菌与细菌的比例以及底物C:N均随着土壤剖面而降低(Moore et al., 2010; Lee et al., 2013).Mooshammer等(2014)在分析微生物分解者对底物质量的适应机制时推测, 真菌与细菌比例应该随着底物C:N的增加而增加.Fanin等(2013)的确发现微生物生物量C:N与真菌与细菌比例存在显著的相关性, 但真菌与细菌比例与底物C:N:P并无相关性.然而, 同属于子囊菌门(Ascom- ycota)的刺盾炱纲(Chaetothyriomycetes)的真菌适应于低质量底物, 而锤舌菌纲(Leotiomycetes)和粪壳菌纲(Sordariomycetes)的真菌却适应于高质量的底物; 这表明, 即便是在真菌内部, 不同分类单元对底物质量的要求也不尽相同(Strickland et al., 2009; Sch- neider et al., 2012).因此, 微生物群落结构、生物量C:N:P与底物C:N:P之间普适性的关系还需验证. ...
... 通常将生物对底物C:X所能承受的限度定义为元素比率阈值(threshold elemental ratio).在微生物研究中, 元素比率阈值建立在代谢理论和化学计量理论基础上, 整合了微生物生物量C:N:P、胞外酶化学计量关系(BG:NAG:AP), 以及元素利用效率对底物C:N:P的适应策略(Sinsabaugh & Follstad Shah, 2012).如果底物C:X超过这个阈值, 则微生物受养分限制, 使其碳利用效率随底物C:X的增加而减小、养分利用效率会增加(Mooshammer et al., 2014; Zechmeister-Boltenstern et al., 2015); 反之, 当底物C:X低于这个阈值时, 微生物受到能量(碳)限制, 使其碳利用效率随底物C:X的增加而增加, 而养分利用效率会降低(Sterner & Elser, 2002; Frost et al. 2006; Doi et al. 2010; Sinsabaugh & Follstad Shah, 2012). ...
... 微生物是陆地生态系统的主要分解者, 其生存和生长策略、群落结构和功能会随着底物化学计量特征而改变, 从而强烈地影响底物的分解速度和元素的周转速率, 进而影响生态系统的功能过程(Manzoni et al., 2010; Sinsabaugh & Shah, 2011; Mooshammer et al., 2012; Xu et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015; Zhou & Wang, 2015).深入理解微生物生理代谢活动和群落结构与陆地生态系统过程之间联系及其对全球变化的响应和反馈机理, 是生态学和全球变化生物学领域值得关注的重大科学命题, 其中亟待解决的主要科学问题有: ...
Metabolic dependencies drive species co-occurrence in diverse microbial communities.
1
2015
... 在微生物群落水平上, 自然选择会使那些能够充分获取并利用特定资源的种群在竞争中获胜(Moorhead & Sinsabaugh, 2006; Fierer et al., 2007; Strickland & Rousk, 2010; Martin et al., 2015; Zelezniak et al., 2015).因此, 底物的质量(高质量意味着低的碳与养分比(C:X), 低质量意味着高的C:X; Xu et al., 2014)的改变必然会改变微生物群落组成.目前很多学者认为, 微生物生物量C:N:P的可塑性是由微生物群落结构的变化导致的(Fanin et al., 2013; Kaiser et al., 2014; Mooshammer et al., 2014; Zechmeister-Boltenstern et al., 2015).生长和周转迅速的r对策微生物常具有较低的生物量C:X, 适应于高质量的底物; 相反, 生长和周转速度较慢的k对策微生物常具有较高的生物量C:X, 适应于低质量底物(Moorhead & Sinsabaugh, 2006; Fierer et al., 2007; Strickland & Rousk, 2010; Kaiser et al., 2014).然而, 由于微生物数量、群落组成以及代谢活动的复杂性, 微生物群落结构、生物量C:N:P与底物C:N:P之间的关系仍不明确.例如: 由于真菌的C:N比细菌高(Strickland & Rousk, 2010), 且真菌与细菌的比例以及底物C:N均随着土壤剖面而降低(Moore et al., 2010; Lee et al., 2013).Mooshammer等(2014)在分析微生物分解者对底物质量的适应机制时推测, 真菌与细菌比例应该随着底物C:N的增加而增加.Fanin等(2013)的确发现微生物生物量C:N与真菌与细菌比例存在显著的相关性, 但真菌与细菌比例与底物C:N:P并无相关性.然而, 同属于子囊菌门(Ascom- ycota)的刺盾炱纲(Chaetothyriomycetes)的真菌适应于低质量底物, 而锤舌菌纲(Leotiomycetes)和粪壳菌纲(Sordariomycetes)的真菌却适应于高质量的底物; 这表明, 即便是在真菌内部, 不同分类单元对底物质量的要求也不尽相同(Strickland et al., 2009; Sch- neider et al., 2012).因此, 微生物群落结构、生物量C:N:P与底物C:N:P之间普适性的关系还需验证. ...
Carbon density and distribution of six Chinese temperate forests. Science China (
1
2010
... 陆地生态系统中微生物的分解底物主要来源于植物的枯枝落叶、粗木质残体、根系凋落物和分泌物、土壤有机质等(Mooshammer et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015).在全球尺度上, 森林生态系统中植物叶片C:N:P为1212:28:1 (摩尔比, McGroddy et al., 2004).植物叶片在凋亡过程中的养分再吸收作用, 使得枯叶养分含量骤减, 从而使枯叶的C:N:P变为3007:45:1 (McGroddy et al., 2004; Yuan & Chen, 2009; Reed et al., 2012; Vergutz et al., 2012; Han et al., 2013).此外, 枯叶的C:N:P还会随生态系统、生物区系以及生活型的不同而发生显著变化(McGroddy et al., 2004).粗木质残体和枯立木的现存量占生态系统碎屑碳库的45% (Zhang & Wang, 2010); 粗木质残体占整个生态系统植物碎屑年输入量的10% (温带森林, Harmon et al., 1986)至30% (热带森林, Martinelli et al., 2000), 其C:N:P可高达14103:40:1 (Weedon et al., 2009).植物细根周转产生的凋落物是土壤碳和养分的重要来源, 其死根C:N:P的全球平均值为4184:43:1 (Yuan et al., 2011).土壤C:N:P最低(287:17:1, Xu et al., 2013), 且随着土壤深度增加而显著减小(Tian et al., 2010).相比之下, 土壤微生物生物量C:N:P比其分解底物低得多.例如: Cleveland和Liptzin (2007)整合全球数据时发现, 陆地生态系统土壤微生物生物量碳氮磷化学计量虽然存在类似于海洋浮游植物的“Redfield ratio” 在资源不受限制时浮游植物的C:N:P为106:16:1; Redfield, 1958), 即土壤微生物生物量C:N:P具有内稳性(homeostasis; Sterner & Elser, 2002), 但森林和草地的土壤微生物生物量C:N:P之间存在显著差异.近期Xu等(2013)进一步整合分析表明, 土壤微生物生物量C:N:P随生物区系不同有很大的变异性(从沙漠的31:4:1到天然湿地的131:14:1).面临底物和微生物化学计量的巨大变异性, 微生物主要通过调整自身化学计量特征、群落结构、胞外酶比例、元素利用效率等4种机制来响应和调节(图1). ...
Reviews and syntheses: Soil resources and climate jointly drive variations in microbial
2015
土地利用变化对东北温带幼龄林土壤碳氮磷含量及其化学计量特征的影响
3
2015
... 微生物作为陆地生态系统主要分解者, 在获取资源构建自身生物量的同时, 驱动着生态系统物质和能量的流通, 调控着碳和养分在土壤-植物-大气连续体(SPAC)之间的循环, 进而影响生态系统的结构与功能(Chapin et al., 2003; Leff et al., 2015).尽管土壤微生物生物量碳只占土壤有机碳库的1.92% (Zhou & Wang, 2015), 但其分解作用消耗了生物圈的绝大部分净初级生产力(Cebrian & Lartigue, 2004; 王兴昌和王传宽, 2015).微生物代谢释放的CO2占土壤呼吸总通量的一半以上(Wang & Yang, 2007), 其矿化和固氮作用维持了土壤的肥力(van der Heijen et al., 2008; Leff et al., 2015); 同时, 土壤微生物与植物的共生、寄生和养分需求关系也会影响地上植被生产力和生物多样性.因此, 探索陆地生态系统地上植被组成和功能特征与地下微生物的耦合关系成为目前生态学的研究热点(van der Heijen et al., 2008; Barberán et al., 2015; Moore et al., 2015); 认识土壤微生物对碳氮磷循环的调控机制也是全球变化背景下生物地球化学循环研究的重要内容(Sinsabaugh & Follstad Shah, 2011; Mooshammer et al., 2014; Zechmeister-Boltenstern et al., 2015). ...
... 微生物是陆地生态系统的主要分解者, 其生存和生长策略、群落结构和功能会随着底物化学计量特征而改变, 从而强烈地影响底物的分解速度和元素的周转速率, 进而影响生态系统的功能过程(Manzoni et al., 2010; Sinsabaugh & Shah, 2011; Mooshammer et al., 2012; Xu et al., 2014; Buchkowski et al., 2015; Zechmeister-Boltenstern et al., 2015; Zhou & Wang, 2015).深入理解微生物生理代谢活动和群落结构与陆地生态系统过程之间联系及其对全球变化的响应和反馈机理, 是生态学和全球变化生物学领域值得关注的重大科学命题, 其中亟待解决的主要科学问题有: ...
... (3)探索微生物对底物化学计量变化适应对策的时空变化.Odum (1969)生态系统演替理论指出, 生态系统呼吸和生物量的比例会随着演替进程而减小.Insam和Haselwandter (1989)首次提出生态系统演替过程中微生物也有类似的变化趋势, 即单位微生物生物量的呼吸速率随着演替进程而减小.但Wardle和Ghani (1995)的整合研究发现大多数情况下单位微生物生物量的呼吸速率随着演替进程先减后增.此后相继出现大量研究, 但仍无统一结论和解释(Saviozzi et al., 2001; Goberna et al., 2006; Susyan et al., 2011; Lu et al., 2014; Raiesi & Beheshti, 2015).运用化学计量学理论可能有助于解释这些争议: 随着生态系统的演替进程, 一方面植物光合作用和微生物固氮作用将大气中的碳和氮素不断输入土壤, 从而导致土壤C:P和N:P的增加; 另一方面, 凋落物、地表枯落物层、土壤C:N随着演替进程基本保持不变(Yang & Luo, 2011; Yang et al., 2011; 周正虎等, 2015), 而单位微生物生物量的呼吸速率随着C:P的增加而增加(图1; Spohn & Chodak, 2015), 从而出现Wardle和Ghani (1995)的“先减后增”格局.另外, 普遍认为, 高纬度地区受氮限制, 而低纬度地区受磷限制; 生态系统演替初期受氮限制, 而后期受磷限制(Vitousek et al., 2010).然而, Spohn和Chodak (2015)在德国发现温带森林土壤C:P与单位微生物生物量的呼吸速率显著相关, 而与C:N不相关; 长期监测结果也显示, 该地区森林植物叶片的磷含量也在减小(Ilg et al., 2009).因此, 上述生态系统演替和全球变化(尤其是氮沉降)可能会导致生态系统的磷限制而突破磷会受纬度限制的传统认识(Vitousek et al., 2010). ...



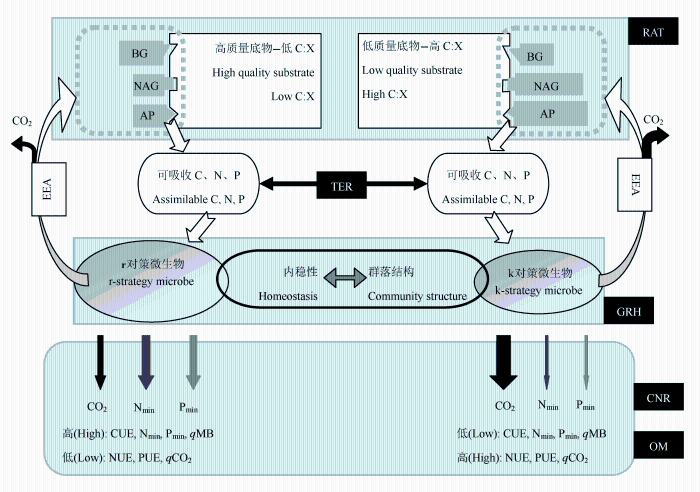
{kind=link}
{kind=link}