1南京信息工程大学应用气象学院生态气象环境研究中心, 南京 210044 2福建农林大学林学院, 福州 3500023CGCEO/Geography, Michigan State University, East Lansing, MI 48823, USA
Optimal stomatal behavior theory for simulating stomatal conductance
FAN Jia-Zhi1, WANG Dan1, *, , HU Ya-Lin2, JING Pan-Pan1, WANG Peng-Peng1, CHEN Jiquan1, 3, *,
1International Center for Ecology, Meteorology and Environment, School of Applied Meteorology, Nanjing University of Information Science & Technology, Nanjing 210044, China2Forestry College of Fujian Agriculture and Forestry University, Fuzhou 350002, China3CGCEO/Geography, Michigan State University, East Lansing, MI 48823, USA
Among the most critical processes in simulating terrestrial ecosystem performance is the regulatory role of stomata in carbon and water cycles. Compared with field measurements, the changes in stomatal slope caused by the biophysical environment provide a simple but effective synthetic framework for studying climate-related carbon and water cycling, due to its sensitivity to CO2, vapor pressure deficit, and photosynthesis. It is also crucial in understanding the effects of climate change on photosynthesis and water use efficiency. Endeavored by numerous scholastic efforts, stomatal conductance models have been improved based on experimental, semi-experimental, and mechanical processes. However, the underlying biological mechanisms and the dynamics of key parameters in these models remain unexplored, especially regarding the changes in stomatal slope. By improving the understanding of the stomata’s regulatory role, we reduced the uncertainty of stomatal conductance simulation. We then synthesized the recent developments and lessons in optimal stomatal behavior theory to simulate stomatal conductance and included an introduction to widely used stomatal conductance models and parameters, the main factors influencing stomatal slopes, and applications of the mechanical stomatal conductance models in different ecosystems. Based on our literature review, we proposed that future research is needed on the optimal stomatal behavior theory and its applications in simulating stomatal conductance.
Keywords:stomatal slope
;
carbon water coupling
;
optimal stomatal behavior theory
;
stomatal conductance models
;
marginal water use efficiency
Ball-Berry模型的特点是可以把复杂的环境因子影响综合到光合作用、湿度和CO2浓度3个要素中。它简化了气孔导度与环境因子的关系, 但是它把光合作用速率作为模型的输入参数处理, 因而无法应用这个模型直接从环境因子计算出气孔导度。Collatz等(1991)耦合了该气孔模型与光合作用生化模型, 建立联立方程组, 以便同时解出光合速率和气孔导度。Ball-Berry气孔导度模型同Farquhar等(1980)光合作用模型结合(FvCB模型), 可以有效地模拟叶片尺度光合作用和气孔导度。FvCB模型被广泛应用到大量陆地生态系统模型中, 并结合气候模式来预测全球变化对植被和大气之间碳、水循环的影响(Sellers et al., 1997; Damour et al., 2010)。因此这两个模型的准确性, 将直接影响大量生态系统模型、区域模型乃至地球系统模型预测的可靠性。
此后多人对Ball-Berry模型进行了优化、改进和应用, 如加入叶片水势的作用等(Sellers et al., 1992; Kim & Lieth, 2003; Tuzet et al., 2003; Yu et al., 2004), 但其本质上依然是半经验模型。在对Ball-Berry模型的众多改进中, Yu等(2004)的模型较为突出。此模型结合了Jarvis模型和Ball-Berry模型的优点, 采用了更便于计算的模型参数, 基于气孔与光合作用的联系及光合作用与光照之间的联系, 推导出了双曲线式的气孔导度模型, 使得直接从气象参数计算气孔导度成为可能。模型简洁、实用, 对于实测数据符合较好。
气孔导度斜率是Ball-Berry模型中一个非常重要的参数, 它很可能随植物种类、叶片养分状态等变化。Ball-Berry模型假定气孔导度斜率是一个常数, 这一点已经被后来的研究否定, 对于单个物种而言, 气孔导度斜率在一段生长期内保持稳定, 但对于不同物种, 气孔导度斜率会有所差异, 并且该斜率值与气孔导度的数值有显著相关关系(Yu et al., 2004)。在过去大约30年间, 生态系统模型通常都假定一个常数来代表所有C3植物的m值或者通过手动调整来拟合观测数据(m = 6, Wang et al., 2013b)。考虑到影响气孔导度的其他叶片性状在不同物种间存在着很大的变异性, 假定C3植物的m值是一个常数是不切实际的。气孔导度模拟的微小误差, 上升到生态系统或者全球尺度, 都有可能造成极大的谬误。通过对模型参数敏感性分析和由参数导致的模型不确定性的方差分解分析可知: 在14个被考虑的模型参数中, 气孔导度斜率对于模型不确定性(杨树(Populus spp.)净初级地上生产力(ANPP))的贡献位列第二, 仅次于生长呼吸(Dietze et al., 2014)。虽然Dietze等(2014)的研究只分析了一个陆地生态系统模型(Ecosystem Demography 2)的模拟结果, 但由于很多模型都利用相似的光合作用和气孔导度模型, 因此可以推测相似的结果可能也会发生在其他生态系统模型的模拟中, 而当采用这些模型预测蒸腾作用和水通量时, 气孔导度斜率对模型不确定性的贡献将会更大。
目前盛行的生态系统模型多是通过减小m值来模拟干旱对气孔导度和光合作用的影响(Wang et al., 2013b, 2015); 或者通过减小光合作用模型中的Vcmax来间接地减小气孔导度和光合作用来实现干旱条件下的生长模拟(Krinner et al., 2005)。没有叶片尺度上气孔导度模拟的准确性, 陆地生态系统模型将无法准确地预测冠层、生态系统乃至区域尺度上碳、水循环的变化(Damour et al., 2010)。成功模拟气孔行为的机理模型将为理解碳水耦合的适应过程以及陆地植被模型对植被结构、功能和演替的准确把握提供一个有效的框架。
2 最优气孔行为理论和气孔导度机理模型
最优气孔行为理论认为气孔的最优化行为就是在某一时间段内, 最大化光合碳固定的同时最小化蒸腾作用, 也就是说, 对于一定的水分消耗, 最大化光合碳固定, 或者也可以说, 对于一定的碳固定(A)而言, 最小化蒸腾作用(E), 即最大化A - λ × E或者最小化E - 1/λ × A (Cowan & Farquhar, 1977)。其中λ是临界水分利用效率(marginal water use efficiency), 也就是植物损耗单位水的碳生产量(∂A/∂E, mol CO2·
大多数地球系统模型或者植被模型都采用经验或者半经验模型来模拟气孔导度(Sellers et al., 1992, 1997; Bonan, 1995; Wang et al., 2013b, 2015)。虽然对于经验或半经验模型来说, 可以利用实验数据矫正参数, 但是在现实中, 并非如此。如上文中所提到的, 多数模型都只是区分了C3和C4植物的气孔导度模型参数m值(Krinner et al., 2005; Oleson et al., 2013)。 De Kauw等(2015)首次将气孔导度机理模型(Medlyn et al., 2011)耦合到陆地表面模型CABLE (Community Atmosphere Biosphere Land Exchange)中, 研究结果显示: 相比于气孔导度经验模型, 耦合气孔导度机理模型导致在常绿针叶林、苔原和C4草原区域, 年蒸腾量减少了30%。Kala等(2014)进一步耦合气孔导度机理模型、CABLE和ACCESS (Australian Community Climate and Earth Systems Simulator)模型, 研究结果显示在北半球的夏季, 新的模型预测的温带森林潜热通量每天减少0.5-1.0 mm, 直接导致每天最高和最低温度升高了1 ℃, 而极端高温升高了1.5 ℃。地表的能量平衡也将进一步影响净初级生产力和陆地碳平衡。由此可见, 深入研究气孔导度斜率的变化规律并提高气孔导度预测的准确性, 可大大减小生态系统模型对碳水通量预测的不确定性, 这对区域尺度生态系统碳水平衡的研究至关重要。然而, 由于对气孔导度斜率变化规律的理解还十分有限, 气孔导度机理模型在植被和地球系统模型中的应用还需要加强。
Decoupling the influence of leaf and root hydraulic conductances on stomatal conductance and its sensitivity to vapour pressure deficit as soil dries in a drained loblolly pine plantation.
EamusD, TaylorDT, Macinnis-NgCM, ShanahanS, de SilvaL (2008).
Comparing model predictions and experimental data for the response of stomatal conductance and guard cell turgor to manipulations of cuticular conductance, leaf-to-air vapour pressure difference and temperature: Feedback mechanisms are able to account for all observations.
Intercomparison of sugar maple (Acer saccharum Marsh.) stand transpiration responses to environmental conditions from the Western Great Lakes Region of the United States.
Agricultural and Forest Meteorology, 148, 231-246.
Modelling stomatal conductance of field-grown sunflower under varying soil water content and leaf environment: Comparison of three models of stomatal response to leaf environment and coupling with an abscisic acid-based model of stomatal response to soil drying.
IPCC (Intergovernmental Panel on Climate Change) (2013). Contribution of working group 1 to the fifth assessment report of the intergovernmental panel on climate change. In: Stocker TF, Qin D, Plattner GK, Tignor M, Allen SK, Boschung J, Nauels A, Xia Y, Bex V, Midgley PM eds. Climate Change in 2013: The Physical Science Basis. Cambridge University Press, Cambridge, UK.
Improved understanding of drought controls on seasonal variation in mediterranean forest canopy CO2 and water fluxes through combined in situ measurements and ecosystem modelling.
A simple dynamic model of photosynthesis in oak leaves: Coupling leaf conductance and photosynthetic carbon fixation by a variable intracellular CO2 pool.
On the complementary relationship between marginal nitrogen and water-use efficiencies among Pinus taeda leaves grown under ambient and CO2-enriched environments.
How should we model plant responses to drought? An analysis of stomatal and non-stomatal responses to water stress. A
gricultural and Forest Meteorology, 182, 204-214.
Adjustments in hydraulic architecture of Pinus palustris maintain similar stomatal conductance in xeric and mesic habitats.
2006
Stomatal sensitivity to vapor pressure deficit and its relationship to hydraulic conductance in Pinus palustris.
1
2004
... 以往研究显示叶片水势(Oren et al., 1999)、叶片、枝条、根部和整株植物水力传导(Salleo et al., 2000; Addington et al., 2004; Bunce, 2006; Domec et al., 2009)、叶片相对含水量(Ewers et al., 2005)等会影响气孔导度对饱和水汽压亏缺的响应.而由于树龄和树高都会影响水力传导和气孔导度, 进而都会影响气孔导度斜率(Schäfer et al., 2000; Franks, 2004; Ewers et al., 2005; Buckley & Roberts, 2006; Ambrose et al., 2010; Woodruff et al., 2010).木材密度和气孔导度斜率之间也存在着显著的负相关关系(Héroult et al., 2013; Lin et al., 2015).植物结构属性, 如根叶比、根深等以及水力传导结构属性也会影响气孔导度斜率(Litvak et al., 2012; McCulloh & Woodruff, 2012).如与散孔材木相比, 环孔材木的气孔对环境因素变化的反应更加敏感(Bush et al., 2008) , 但是在Litvak等(2012)的研究中并未发现木材结构影响气孔导度斜率的现象.而随着测量高度的增高, 气孔导度对饱和水汽压亏缺响应的敏感性也会降低.叶肉导度和Vcmax也通过影响光合作用影响植物的水分利用效率(Wang et al., 2013a).Flexas等(2013)分析了44种不同物种水分利用效率的规律, 发现水分利用效率和叶肉导度与气孔导度的比值成正比.这个关系表明水分利用效率只有在固碳能力的提高高于水分丢失增加(气孔导度升高)的情况下, 才可以升高.可以预测, 同样的关系也应该在气孔导度斜率及Vcmax和气孔导度的比值间存在.临界氮利用效率也被证明和临界水分利用效率有互补的关系(Palmroth et al., 2013). ...
Effects of height on treetop transpiration and stomatal conductance in coast redwood (Sequoia sempervirens).
1
2010
... 以往研究显示叶片水势(Oren et al., 1999)、叶片、枝条、根部和整株植物水力传导(Salleo et al., 2000; Addington et al., 2004; Bunce, 2006; Domec et al., 2009)、叶片相对含水量(Ewers et al., 2005)等会影响气孔导度对饱和水汽压亏缺的响应.而由于树龄和树高都会影响水力传导和气孔导度, 进而都会影响气孔导度斜率(Schäfer et al., 2000; Franks, 2004; Ewers et al., 2005; Buckley & Roberts, 2006; Ambrose et al., 2010; Woodruff et al., 2010).木材密度和气孔导度斜率之间也存在着显著的负相关关系(Héroult et al., 2013; Lin et al., 2015).植物结构属性, 如根叶比、根深等以及水力传导结构属性也会影响气孔导度斜率(Litvak et al., 2012; McCulloh & Woodruff, 2012).如与散孔材木相比, 环孔材木的气孔对环境因素变化的反应更加敏感(Bush et al., 2008) , 但是在Litvak等(2012)的研究中并未发现木材结构影响气孔导度斜率的现象.而随着测量高度的增高, 气孔导度对饱和水汽压亏缺响应的敏感性也会降低.叶肉导度和Vcmax也通过影响光合作用影响植物的水分利用效率(Wang et al., 2013a).Flexas等(2013)分析了44种不同物种水分利用效率的规律, 发现水分利用效率和叶肉导度与气孔导度的比值成正比.这个关系表明水分利用效率只有在固碳能力的提高高于水分丢失增加(气孔导度升高)的情况下, 才可以升高.可以预测, 同样的关系也应该在气孔导度斜率及Vcmax和气孔导度的比值间存在.临界氮利用效率也被证明和临界水分利用效率有互补的关系(Palmroth et al., 2013). ...
A model predicting stomatal conductance and its contribution to the control of photosynthesis under different environmental conditions.
2
1987
... 以碳积累为代表的光合作用和以水分消耗为代表的蒸腾作用是所有植物功能模型的基础, 准确地模拟光合作用和蒸腾作用也是生态系统模型、植被模型、地球系统模型能够有效模拟碳、水和能量循环的必要条件(Wang et al., 2013b, 2015, 2016).由于以Farquhar等(1980)的光合作用模型为代表的机理模型的广泛应用, 通过对物种生物属性和环境参数(如光合最大羧化速率(Vcmax)、CO2浓度、温度和光照)的测量, 可以有效地模拟光合作用(Wullschleger, 1993; Kattge et al., 2009).相对于光合作用的机理模型, 蒸腾作用通常都是由经验或者半经验模型模拟出的气孔导度值来计算.过去40年里, 气孔导度的模拟工作大量开展.概括来讲, 对气孔导度的模拟有以下几种方式: (1)经验模型.大多通过气孔导度与多个环境因素如空气湿度(Lange et al., 1971)、水汽压亏缺(VPD)(Monteith, 1995)、叶片温度(Jarvis, 1976; Jones, 1992)和臭氧(Uddling et al., 2009)等的统计关系建立模型; (2)半经验模型.基于植物生理特性的假设, 但是仍然和经验模型结合, 如基于气孔导度和光合作用(Ball et al., 1987; Leuning, 1990, 1995; Misson et al., 2004b)、脱落酸(ABA) (Tardieu & Davies, 1993; Guteschick & Simonneau, 2002)、水分传导(Sperry et al., 1998; Oren et al., 1999; Tuzet et al., 2003)和膨压调节(Dewar, 2002; Buckley et al., 2003)等的关系建立的模型, 其参数不具有生物学属性; (3)机理模型.基于植物生理特性的假设, 其参数具有生物学特性, 并具有可以应用在各种环境条件下的潜力(Dewar, 2002; Medlyn et al., 2011; Prentice et al., 2014).很多文献已经综述过气孔导度的经验与半经验模型, 本文将侧重阐述气孔导度机理模型的研究. ...
The growth of soybean under free air [CO2] enrichment (FACE) stimulates photosynthesis while decreasing in vivo Rubisco capacity.
1
2005
... 虽然光合作用、气孔导度的调查数据在文献中大量存在, 但是由于实验设计的不同以及缺少饱和水汽压亏缺等环境参数的信息, 很少有实验数据能够满足对气孔导度斜率进行模拟计算的要求(Zhou et al., 2013).以往大多数关于气孔导度斜率的研究都是在温室里的短期试验, 在自然条件下的研究还很少(Medlyn et al., 2011).对于增高的CO2浓度下气孔导度对环境因子的适应性变化的研究结果也不一致.在高CO2浓度的未来环境下, 植物会降低气孔导度和提高光合速率(Long et al., 2004; Bernacchi et al., 2005).有学者在培养箱中升高CO2浓度对大豆(Glycine max)和棉花(Gossypium hirsutum)进行培养研究得出, 气孔导度斜率会因CO2浓度的增加而降低(Harley et al., 1992; Bunce, 2004).但是Leakey等(2006)在FACE (free-air CO2 enrichment)实验样地对大豆进行了生长和生理参数测量, 他们的实验结果却表明气孔导度斜率不会因外界CO2浓度的升高而改变, 并且指出前人所得的气孔导度斜率降低的结论可能是增高的CO2浓度对气孔导度带来的瞬时改变的结果.Medlyn等(2001)也测定了生长在高CO2浓度下6种不同树种的生理参数, 发现CO2浓度升高并没有改变气孔导度斜率.这一结果也在Katul等(2009)的研究中得到证实, Katul等(2009)也发现CO2浓度升高并没有改变Ball-Berry模型中的光合作用与气孔导度的线性关系的斜率. ...
Improved temperature response functions for models of Rubisco-limited photosynthesis.
1
2001
... 对于外界环境的变化, 气孔导度斜率也会做出不同的响应.不同植物物种的水分利用策略不同, 在同样的湿度环境下气孔导度斜率会有差异, 同样的物种在不同水分环境下气孔导度斜率也会不同(Zhou et al., 2013).Lin等(2015)对超过1万条光合测量数据综合分析后发现, 在植物生长适宜的外界温湿度下, 随着温湿度升高, 气孔导度斜率都会增加.对于气孔导度斜率随温度升高而增加的原因, 有两种解释: 一是气孔导度斜率与CO2补偿点浓度相关(Medlyn et al., 2011), 而这一浓度会随温度升高而增加(Bernacchi et al., 2001); 二是水分子黏性会随温度升高而降低, 这就降低了运输水分的消耗, 从而增高了气孔导度斜率(Prentice et al., 2014).Héroult等(2013)报道了产地不同的4种桉属植物在相同环境条件下, 气孔导度斜率随着自然产地不同而不同, 自然生长在湿润地区的植物气孔导度斜率比在半湿润地区的植物高.在湿润环境生长的长叶松(Pinus palustris)气孔关闭对增加的饱和水汽压亏缺的反应比在干旱环境下更加敏感(Addingtong et al., 2006). ...
The Joint UK Land Environment Simulator (JULES), model description-Part 1: Energy and water fluxes.
1
2011
... 在很多生态系统模型中, 土壤干旱胁迫对气孔导度影响的模拟通常是通过改变水分胁迫与光合作用/气孔导度之间的作用关系来实现的(Sala & Tenhunen, 1996; Verhoef & Allen, 2000).但也有模型是通过改变水分胁迫与光合作用或者光合作用参数Vcmax和最大电子传输速率(Jmax)的关系, 模拟水分胁迫下光合作用的改变进而模拟气孔导度的改变(Sellers et al., 1996; Ronda et al., 2001; Best et al., 2011).虽然在轻度干旱情况下, 上述几种做法可以模拟出水分胁迫对叶片尺度碳水通量的影响, 但在严重干旱情况下无法同时准确模拟光合作用和蒸腾作用(Verhoef & Allen, 2000; Reichstein et al., 2002; Keenan et al., 2009).Egea等(2011)提议, 在干旱条件下可以通过先限制叶肉导度, 进而限制气孔导度和光合作用参数的方案来提高光合作用和气孔导度模拟的准确性.Zhou等(2013)的研究结果也表明模拟干旱状况下的气孔导度, 必须同时考虑干旱对气孔和非气孔因素(如Vcmax)的综合影响.由于对干旱状况下气孔导度预测的不准确性, 现有的模型一般都倾向于高估光合作用和蒸腾作用对干旱的响应(Dietze, 2014).与以前从气孔(物理过程)或者非气孔(生化过程)的角度出发研究干旱对光合作用和气孔导度的影响不同, 气孔导度机理模型中的气孔导度斜率在干旱环境下的变化规律包含了干旱对气孔(生物物理因素)和非气孔因素, 包括最大光合效率和临界水分利用效率的综合影响, 为研究全球变化下陆地生态系统碳水耦合的适应性提供了一个简明且综合的框架.Manzoni等(2011)计算了50种植物的临界水分利用效率, 发现在忽略表皮水分丢失的情况下, 植物临界水分利用效率在轻微干旱的情况下增高, 而在严重干旱的情况下下降; 如果不忽略表皮水分丢失, 植物临界水分利用效率会随着水分胁迫的增加而提高; 而不同的植物功能型和不同的气候条件下, 反应也会不同.总体说来, 土壤水分较少时, 植物的水分利用效率会增高, 此时气孔导度斜率值也会降低(Lin et al., 2015).在环境水分条件与气孔导度斜率的变化规律之间还有很多值得讨论的地方, 我国学者对于水分利用的研究主要集中在单叶水分利用效率(付士磊等, 2006; 杨建伟等, 2006; 全先奎和王传宽, 2015)、气孔导度时空变异的调查(司建华等, 2008)、气孔导度对水分环境的响应(鱼腾飞等, 2012; 袁国富等, 2012)、单叶或者幼苗蒸腾速率(宋炳煜等, 2004; 岳广阳等, 2009)、树干液流密度(赵平等, 2006)、土壤水分定位监测(尹春梅等, 2008), 以及涡度协相关法测量生态系统水分利用效率(Zhou et al., 2013)等方面.土壤干旱如何影响不同功能型和物种的气孔导度斜率和临界水分利用效率, 还有待于进一步的研究. ...
Land-atmosphere CO2 exchange simulated by a land surface process model coupled to an atmospheric general circulation model.
1
1995
... 大多数地球系统模型或者植被模型都采用经验或者半经验模型来模拟气孔导度(Sellers et al., 1992, 1997; Bonan, 1995; Wang et al., 2013b, 2015).虽然对于经验或半经验模型来说, 可以利用实验数据矫正参数, 但是在现实中, 并非如此.如上文中所提到的, 多数模型都只是区分了C3和C4植物的气孔导度模型参数m值(Krinner et al., 2005; Oleson et al., 2013). De Kauw等(2015)首次将气孔导度机理模型(Medlyn et al., 2011)耦合到陆地表面模型CABLE (Community Atmosphere Biosphere Land Exchange)中, 研究结果显示: 相比于气孔导度经验模型, 耦合气孔导度机理模型导致在常绿针叶林、苔原和C4草原区域, 年蒸腾量减少了30%.Kala等(2014)进一步耦合气孔导度机理模型、CABLE和ACCESS (Australian Community Climate and Earth Systems Simulator)模型, 研究结果显示在北半球的夏季, 新的模型预测的温带森林潜热通量每天减少0.5-1.0 mm, 直接导致每天最高和最低温度升高了1 ℃, 而极端高温升高了1.5 ℃.地表的能量平衡也将进一步影响净初级生产力和陆地碳平衡.由此可见, 深入研究气孔导度斜率的变化规律并提高气孔导度预测的准确性, 可大大减小生态系统模型对碳水通量预测的不确定性, 这对区域尺度生态系统碳水平衡的研究至关重要.然而, 由于对气孔导度斜率变化规律的理解还十分有限, 气孔导度机理模型在植被和地球系统模型中的应用还需要加强. ...
A hydromechanical and biochemical model of stomatal conductance.
1
2003
... 以碳积累为代表的光合作用和以水分消耗为代表的蒸腾作用是所有植物功能模型的基础, 准确地模拟光合作用和蒸腾作用也是生态系统模型、植被模型、地球系统模型能够有效模拟碳、水和能量循环的必要条件(Wang et al., 2013b, 2015, 2016).由于以Farquhar等(1980)的光合作用模型为代表的机理模型的广泛应用, 通过对物种生物属性和环境参数(如光合最大羧化速率(Vcmax)、CO2浓度、温度和光照)的测量, 可以有效地模拟光合作用(Wullschleger, 1993; Kattge et al., 2009).相对于光合作用的机理模型, 蒸腾作用通常都是由经验或者半经验模型模拟出的气孔导度值来计算.过去40年里, 气孔导度的模拟工作大量开展.概括来讲, 对气孔导度的模拟有以下几种方式: (1)经验模型.大多通过气孔导度与多个环境因素如空气湿度(Lange et al., 1971)、水汽压亏缺(VPD)(Monteith, 1995)、叶片温度(Jarvis, 1976; Jones, 1992)和臭氧(Uddling et al., 2009)等的统计关系建立模型; (2)半经验模型.基于植物生理特性的假设, 但是仍然和经验模型结合, 如基于气孔导度和光合作用(Ball et al., 1987; Leuning, 1990, 1995; Misson et al., 2004b)、脱落酸(ABA) (Tardieu & Davies, 1993; Guteschick & Simonneau, 2002)、水分传导(Sperry et al., 1998; Oren et al., 1999; Tuzet et al., 2003)和膨压调节(Dewar, 2002; Buckley et al., 2003)等的关系建立的模型, 其参数不具有生物学属性; (3)机理模型.基于植物生理特性的假设, 其参数具有生物学特性, 并具有可以应用在各种环境条件下的潜力(Dewar, 2002; Medlyn et al., 2011; Prentice et al., 2014).很多文献已经综述过气孔导度的经验与半经验模型, 本文将侧重阐述气孔导度机理模型的研究. ...
How should leaf area, sapwood area and stomatal conductance vary with tree height to maximize growth?
1
2006
... 以往研究显示叶片水势(Oren et al., 1999)、叶片、枝条、根部和整株植物水力传导(Salleo et al., 2000; Addington et al., 2004; Bunce, 2006; Domec et al., 2009)、叶片相对含水量(Ewers et al., 2005)等会影响气孔导度对饱和水汽压亏缺的响应.而由于树龄和树高都会影响水力传导和气孔导度, 进而都会影响气孔导度斜率(Schäfer et al., 2000; Franks, 2004; Ewers et al., 2005; Buckley & Roberts, 2006; Ambrose et al., 2010; Woodruff et al., 2010).木材密度和气孔导度斜率之间也存在着显著的负相关关系(Héroult et al., 2013; Lin et al., 2015).植物结构属性, 如根叶比、根深等以及水力传导结构属性也会影响气孔导度斜率(Litvak et al., 2012; McCulloh & Woodruff, 2012).如与散孔材木相比, 环孔材木的气孔对环境因素变化的反应更加敏感(Bush et al., 2008) , 但是在Litvak等(2012)的研究中并未发现木材结构影响气孔导度斜率的现象.而随着测量高度的增高, 气孔导度对饱和水汽压亏缺响应的敏感性也会降低.叶肉导度和Vcmax也通过影响光合作用影响植物的水分利用效率(Wang et al., 2013a).Flexas等(2013)分析了44种不同物种水分利用效率的规律, 发现水分利用效率和叶肉导度与气孔导度的比值成正比.这个关系表明水分利用效率只有在固碳能力的提高高于水分丢失增加(气孔导度升高)的情况下, 才可以升高.可以预测, 同样的关系也应该在气孔导度斜率及Vcmax和气孔导度的比值间存在.临界氮利用效率也被证明和临界水分利用效率有互补的关系(Palmroth et al., 2013). ...
Carbon dioxide effects on stomatal responses to the environment and water use by crops under field conditions.
1
2004
... 虽然光合作用、气孔导度的调查数据在文献中大量存在, 但是由于实验设计的不同以及缺少饱和水汽压亏缺等环境参数的信息, 很少有实验数据能够满足对气孔导度斜率进行模拟计算的要求(Zhou et al., 2013).以往大多数关于气孔导度斜率的研究都是在温室里的短期试验, 在自然条件下的研究还很少(Medlyn et al., 2011).对于增高的CO2浓度下气孔导度对环境因子的适应性变化的研究结果也不一致.在高CO2浓度的未来环境下, 植物会降低气孔导度和提高光合速率(Long et al., 2004; Bernacchi et al., 2005).有学者在培养箱中升高CO2浓度对大豆(Glycine max)和棉花(Gossypium hirsutum)进行培养研究得出, 气孔导度斜率会因CO2浓度的增加而降低(Harley et al., 1992; Bunce, 2004).但是Leakey等(2006)在FACE (free-air CO2 enrichment)实验样地对大豆进行了生长和生理参数测量, 他们的实验结果却表明气孔导度斜率不会因外界CO2浓度的升高而改变, 并且指出前人所得的气孔导度斜率降低的结论可能是增高的CO2浓度对气孔导度带来的瞬时改变的结果.Medlyn等(2001)也测定了生长在高CO2浓度下6种不同树种的生理参数, 发现CO2浓度升高并没有改变气孔导度斜率.这一结果也在Katul等(2009)的研究中得到证实, Katul等(2009)也发现CO2浓度升高并没有改变Ball-Berry模型中的光合作用与气孔导度的线性关系的斜率. ...
How do leaf hydraulics limit stomatal conductance at high water vapour pressure deficits?
1
2006
... 以往研究显示叶片水势(Oren et al., 1999)、叶片、枝条、根部和整株植物水力传导(Salleo et al., 2000; Addington et al., 2004; Bunce, 2006; Domec et al., 2009)、叶片相对含水量(Ewers et al., 2005)等会影响气孔导度对饱和水汽压亏缺的响应.而由于树龄和树高都会影响水力传导和气孔导度, 进而都会影响气孔导度斜率(Schäfer et al., 2000; Franks, 2004; Ewers et al., 2005; Buckley & Roberts, 2006; Ambrose et al., 2010; Woodruff et al., 2010).木材密度和气孔导度斜率之间也存在着显著的负相关关系(Héroult et al., 2013; Lin et al., 2015).植物结构属性, 如根叶比、根深等以及水力传导结构属性也会影响气孔导度斜率(Litvak et al., 2012; McCulloh & Woodruff, 2012).如与散孔材木相比, 环孔材木的气孔对环境因素变化的反应更加敏感(Bush et al., 2008) , 但是在Litvak等(2012)的研究中并未发现木材结构影响气孔导度斜率的现象.而随着测量高度的增高, 气孔导度对饱和水汽压亏缺响应的敏感性也会降低.叶肉导度和Vcmax也通过影响光合作用影响植物的水分利用效率(Wang et al., 2013a).Flexas等(2013)分析了44种不同物种水分利用效率的规律, 发现水分利用效率和叶肉导度与气孔导度的比值成正比.这个关系表明水分利用效率只有在固碳能力的提高高于水分丢失增加(气孔导度升高)的情况下, 才可以升高.可以预测, 同样的关系也应该在气孔导度斜率及Vcmax和气孔导度的比值间存在.临界氮利用效率也被证明和临界水分利用效率有互补的关系(Palmroth et al., 2013). ...
Wood anatomy constrains stomatal responses to atmospheric vapor pressure deficit in irrigated, urban trees.
1
2008
... 以往研究显示叶片水势(Oren et al., 1999)、叶片、枝条、根部和整株植物水力传导(Salleo et al., 2000; Addington et al., 2004; Bunce, 2006; Domec et al., 2009)、叶片相对含水量(Ewers et al., 2005)等会影响气孔导度对饱和水汽压亏缺的响应.而由于树龄和树高都会影响水力传导和气孔导度, 进而都会影响气孔导度斜率(Schäfer et al., 2000; Franks, 2004; Ewers et al., 2005; Buckley & Roberts, 2006; Ambrose et al., 2010; Woodruff et al., 2010).木材密度和气孔导度斜率之间也存在着显著的负相关关系(Héroult et al., 2013; Lin et al., 2015).植物结构属性, 如根叶比、根深等以及水力传导结构属性也会影响气孔导度斜率(Litvak et al., 2012; McCulloh & Woodruff, 2012).如与散孔材木相比, 环孔材木的气孔对环境因素变化的反应更加敏感(Bush et al., 2008) , 但是在Litvak等(2012)的研究中并未发现木材结构影响气孔导度斜率的现象.而随着测量高度的增高, 气孔导度对饱和水汽压亏缺响应的敏感性也会降低.叶肉导度和Vcmax也通过影响光合作用影响植物的水分利用效率(Wang et al., 2013a).Flexas等(2013)分析了44种不同物种水分利用效率的规律, 发现水分利用效率和叶肉导度与气孔导度的比值成正比.这个关系表明水分利用效率只有在固碳能力的提高高于水分丢失增加(气孔导度升高)的情况下, 才可以升高.可以预测, 同样的关系也应该在气孔导度斜率及Vcmax和气孔导度的比值间存在.临界氮利用效率也被证明和临界水分利用效率有互补的关系(Palmroth et al., 2013). ...
Physiological and environmental regulation of stomatal conductance, photosynthesis and transpiration: A model that includes a laminar boundary layer.
1991
Stomatal function in relation to leaf metabolism and environment.
1
1977
... 最优气孔行为理论认为气孔的最优化行为就是在某一时间段内, 最大化光合碳固定的同时最小化蒸腾作用, 也就是说, 对于一定的水分消耗, 最大化光合碳固定, 或者也可以说, 对于一定的碳固定(A)而言, 最小化蒸腾作用(E), 即最大化A - λ × E或者最小化E - 1/λ × A (Cowan & Farquhar, 1977).其中λ是临界水分利用效率(marginal water use efficiency), 也就是植物损耗单位水的碳生产量(∂A/∂E, mol CO2· ...
An overview of models of stomatal conductance at the leaf level.
3
2010
... Ball-Berry模型的特点是可以把复杂的环境因子影响综合到光合作用、湿度和CO2浓度3个要素中.它简化了气孔导度与环境因子的关系, 但是它把光合作用速率作为模型的输入参数处理, 因而无法应用这个模型直接从环境因子计算出气孔导度.Collatz等(1991)耦合了该气孔模型与光合作用生化模型, 建立联立方程组, 以便同时解出光合速率和气孔导度.Ball-Berry气孔导度模型同Farquhar等(1980)光合作用模型结合(FvCB模型), 可以有效地模拟叶片尺度光合作用和气孔导度.FvCB模型被广泛应用到大量陆地生态系统模型中, 并结合气候模式来预测全球变化对植被和大气之间碳、水循环的影响(Sellers et al., 1997; Damour et al., 2010).因此这两个模型的准确性, 将直接影响大量生态系统模型、区域模型乃至地球系统模型预测的可靠性. ...
... 目前盛行的生态系统模型多是通过减小m值来模拟干旱对气孔导度和光合作用的影响(Wang et al., 2013b, 2015); 或者通过减小光合作用模型中的Vcmax来间接地减小气孔导度和光合作用来实现干旱条件下的生长模拟(Krinner et al., 2005).没有叶片尺度上气孔导度模拟的准确性, 陆地生态系统模型将无法准确地预测冠层、生态系统乃至区域尺度上碳、水循环的变化(Damour et al., 2010).成功模拟气孔行为的机理模型将为理解碳水耦合的适应过程以及陆地植被模型对植被结构、功能和演替的准确把握提供一个有效的框架. ...
... 依据最优气孔导度理论并借鉴Ball-Berry的半经验模型发展的气孔导度机理模型(推导过程见Medlyn et al., 2011)(方程3), 可以将模型中的斜率参数与广受肯定的最优气孔理论中的临界水分利用效率(λ = ∂A/∂E, mol CO2·mol-1 H2O)(Damour et al., 2010; Medlyn et al., 2011)联系在一起(方程4), 使得模型参数具有了重要的生物学含义. ...
Stomatal control by chemical signalling and the exploitation of this mechanism to increase water use efficiency in agriculture.
A test of an optimal stomatal conductance scheme within the CABLE land surface model.
2015
The Ball-Berry-Leuning and Tardieu-Davies stomatal models: Synthesis and extension within a spatially aggregated picture of guard cell function.
2
2002
... 以碳积累为代表的光合作用和以水分消耗为代表的蒸腾作用是所有植物功能模型的基础, 准确地模拟光合作用和蒸腾作用也是生态系统模型、植被模型、地球系统模型能够有效模拟碳、水和能量循环的必要条件(Wang et al., 2013b, 2015, 2016).由于以Farquhar等(1980)的光合作用模型为代表的机理模型的广泛应用, 通过对物种生物属性和环境参数(如光合最大羧化速率(Vcmax)、CO2浓度、温度和光照)的测量, 可以有效地模拟光合作用(Wullschleger, 1993; Kattge et al., 2009).相对于光合作用的机理模型, 蒸腾作用通常都是由经验或者半经验模型模拟出的气孔导度值来计算.过去40年里, 气孔导度的模拟工作大量开展.概括来讲, 对气孔导度的模拟有以下几种方式: (1)经验模型.大多通过气孔导度与多个环境因素如空气湿度(Lange et al., 1971)、水汽压亏缺(VPD)(Monteith, 1995)、叶片温度(Jarvis, 1976; Jones, 1992)和臭氧(Uddling et al., 2009)等的统计关系建立模型; (2)半经验模型.基于植物生理特性的假设, 但是仍然和经验模型结合, 如基于气孔导度和光合作用(Ball et al., 1987; Leuning, 1990, 1995; Misson et al., 2004b)、脱落酸(ABA) (Tardieu & Davies, 1993; Guteschick & Simonneau, 2002)、水分传导(Sperry et al., 1998; Oren et al., 1999; Tuzet et al., 2003)和膨压调节(Dewar, 2002; Buckley et al., 2003)等的关系建立的模型, 其参数不具有生物学属性; (3)机理模型.基于植物生理特性的假设, 其参数具有生物学特性, 并具有可以应用在各种环境条件下的潜力(Dewar, 2002; Medlyn et al., 2011; Prentice et al., 2014).很多文献已经综述过气孔导度的经验与半经验模型, 本文将侧重阐述气孔导度机理模型的研究. ...
... )等的关系建立的模型, 其参数不具有生物学属性; (3)机理模型.基于植物生理特性的假设, 其参数具有生物学特性, 并具有可以应用在各种环境条件下的潜力(Dewar, 2002; Medlyn et al., 2011; Prentice et al., 2014).很多文献已经综述过气孔导度的经验与半经验模型, 本文将侧重阐述气孔导度机理模型的研究. ...
Gaps in knowledge and data driving uncertainty in models of photosynthesis.
2
2014
... 气孔导度斜率是Ball-Berry模型中一个非常重要的参数, 它很可能随植物种类、叶片养分状态等变化.Ball-Berry模型假定气孔导度斜率是一个常数, 这一点已经被后来的研究否定, 对于单个物种而言, 气孔导度斜率在一段生长期内保持稳定, 但对于不同物种, 气孔导度斜率会有所差异, 并且该斜率值与气孔导度的数值有显著相关关系(Yu et al., 2004).在过去大约30年间, 生态系统模型通常都假定一个常数来代表所有C3植物的m值或者通过手动调整来拟合观测数据(m = 6, Wang et al., 2013b).考虑到影响气孔导度的其他叶片性状在不同物种间存在着很大的变异性, 假定C3植物的m值是一个常数是不切实际的.气孔导度模拟的微小误差, 上升到生态系统或者全球尺度, 都有可能造成极大的谬误.通过对模型参数敏感性分析和由参数导致的模型不确定性的方差分解分析可知: 在14个被考虑的模型参数中, 气孔导度斜率对于模型不确定性(杨树(Populus spp.)净初级地上生产力(ANPP))的贡献位列第二, 仅次于生长呼吸(Dietze et al., 2014).虽然Dietze等(2014)的研究只分析了一个陆地生态系统模型(Ecosystem Demography 2)的模拟结果, 但由于很多模型都利用相似的光合作用和气孔导度模型, 因此可以推测相似的结果可能也会发生在其他生态系统模型的模拟中, 而当采用这些模型预测蒸腾作用和水通量时, 气孔导度斜率对模型不确定性的贡献将会更大. ...
... 在很多生态系统模型中, 土壤干旱胁迫对气孔导度影响的模拟通常是通过改变水分胁迫与光合作用/气孔导度之间的作用关系来实现的(Sala & Tenhunen, 1996; Verhoef & Allen, 2000).但也有模型是通过改变水分胁迫与光合作用或者光合作用参数Vcmax和最大电子传输速率(Jmax)的关系, 模拟水分胁迫下光合作用的改变进而模拟气孔导度的改变(Sellers et al., 1996; Ronda et al., 2001; Best et al., 2011).虽然在轻度干旱情况下, 上述几种做法可以模拟出水分胁迫对叶片尺度碳水通量的影响, 但在严重干旱情况下无法同时准确模拟光合作用和蒸腾作用(Verhoef & Allen, 2000; Reichstein et al., 2002; Keenan et al., 2009).Egea等(2011)提议, 在干旱条件下可以通过先限制叶肉导度, 进而限制气孔导度和光合作用参数的方案来提高光合作用和气孔导度模拟的准确性.Zhou等(2013)的研究结果也表明模拟干旱状况下的气孔导度, 必须同时考虑干旱对气孔和非气孔因素(如Vcmax)的综合影响.由于对干旱状况下气孔导度预测的不准确性, 现有的模型一般都倾向于高估光合作用和蒸腾作用对干旱的响应(Dietze, 2014).与以前从气孔(物理过程)或者非气孔(生化过程)的角度出发研究干旱对光合作用和气孔导度的影响不同, 气孔导度机理模型中的气孔导度斜率在干旱环境下的变化规律包含了干旱对气孔(生物物理因素)和非气孔因素, 包括最大光合效率和临界水分利用效率的综合影响, 为研究全球变化下陆地生态系统碳水耦合的适应性提供了一个简明且综合的框架.Manzoni等(2011)计算了50种植物的临界水分利用效率, 发现在忽略表皮水分丢失的情况下, 植物临界水分利用效率在轻微干旱的情况下增高, 而在严重干旱的情况下下降; 如果不忽略表皮水分丢失, 植物临界水分利用效率会随着水分胁迫的增加而提高; 而不同的植物功能型和不同的气候条件下, 反应也会不同.总体说来, 土壤水分较少时, 植物的水分利用效率会增高, 此时气孔导度斜率值也会降低(Lin et al., 2015).在环境水分条件与气孔导度斜率的变化规律之间还有很多值得讨论的地方, 我国学者对于水分利用的研究主要集中在单叶水分利用效率(付士磊等, 2006; 杨建伟等, 2006; 全先奎和王传宽, 2015)、气孔导度时空变异的调查(司建华等, 2008)、气孔导度对水分环境的响应(鱼腾飞等, 2012; 袁国富等, 2012)、单叶或者幼苗蒸腾速率(宋炳煜等, 2004; 岳广阳等, 2009)、树干液流密度(赵平等, 2006)、土壤水分定位监测(尹春梅等, 2008), 以及涡度协相关法测量生态系统水分利用效率(Zhou et al., 2013)等方面.土壤干旱如何影响不同功能型和物种的气孔导度斜率和临界水分利用效率, 还有待于进一步的研究. ...
A quantitative assessment of a terrestrial biosphere model’s data needs across North American biomes.
2014
Decoupling the influence of leaf and root hydraulic conductances on stomatal conductance and its sensitivity to vapour pressure deficit as soil dries in a drained loblolly pine plantation.
1
2009
... 以往研究显示叶片水势(Oren et al., 1999)、叶片、枝条、根部和整株植物水力传导(Salleo et al., 2000; Addington et al., 2004; Bunce, 2006; Domec et al., 2009)、叶片相对含水量(Ewers et al., 2005)等会影响气孔导度对饱和水汽压亏缺的响应.而由于树龄和树高都会影响水力传导和气孔导度, 进而都会影响气孔导度斜率(Schäfer et al., 2000; Franks, 2004; Ewers et al., 2005; Buckley & Roberts, 2006; Ambrose et al., 2010; Woodruff et al., 2010).木材密度和气孔导度斜率之间也存在着显著的负相关关系(Héroult et al., 2013; Lin et al., 2015).植物结构属性, 如根叶比、根深等以及水力传导结构属性也会影响气孔导度斜率(Litvak et al., 2012; McCulloh & Woodruff, 2012).如与散孔材木相比, 环孔材木的气孔对环境因素变化的反应更加敏感(Bush et al., 2008) , 但是在Litvak等(2012)的研究中并未发现木材结构影响气孔导度斜率的现象.而随着测量高度的增高, 气孔导度对饱和水汽压亏缺响应的敏感性也会降低.叶肉导度和Vcmax也通过影响光合作用影响植物的水分利用效率(Wang et al., 2013a).Flexas等(2013)分析了44种不同物种水分利用效率的规律, 发现水分利用效率和叶肉导度与气孔导度的比值成正比.这个关系表明水分利用效率只有在固碳能力的提高高于水分丢失增加(气孔导度升高)的情况下, 才可以升高.可以预测, 同样的关系也应该在气孔导度斜率及Vcmax和气孔导度的比值间存在.临界氮利用效率也被证明和临界水分利用效率有互补的关系(Palmroth et al., 2013). ...
Comparing model predictions and experimental data for the response of stomatal conductance and guard cell turgor to manipulations of cuticular conductance, leaf-to-air vapour pressure difference and temperature: Feedback mechanisms are able to account for all observations.
Towards an improved and more flexible representation of water stress in coupled photosynthesis-stomatal conductance models.
2011
Effects of stand age and tree species on canopy transpiration and average stomatal conductance of boreal forests.
2
2005
... 以往研究显示叶片水势(Oren et al., 1999)、叶片、枝条、根部和整株植物水力传导(Salleo et al., 2000; Addington et al., 2004; Bunce, 2006; Domec et al., 2009)、叶片相对含水量(Ewers et al., 2005)等会影响气孔导度对饱和水汽压亏缺的响应.而由于树龄和树高都会影响水力传导和气孔导度, 进而都会影响气孔导度斜率(Schäfer et al., 2000; Franks, 2004; Ewers et al., 2005; Buckley & Roberts, 2006; Ambrose et al., 2010; Woodruff et al., 2010).木材密度和气孔导度斜率之间也存在着显著的负相关关系(Héroult et al., 2013; Lin et al., 2015).植物结构属性, 如根叶比、根深等以及水力传导结构属性也会影响气孔导度斜率(Litvak et al., 2012; McCulloh & Woodruff, 2012).如与散孔材木相比, 环孔材木的气孔对环境因素变化的反应更加敏感(Bush et al., 2008) , 但是在Litvak等(2012)的研究中并未发现木材结构影响气孔导度斜率的现象.而随着测量高度的增高, 气孔导度对饱和水汽压亏缺响应的敏感性也会降低.叶肉导度和Vcmax也通过影响光合作用影响植物的水分利用效率(Wang et al., 2013a).Flexas等(2013)分析了44种不同物种水分利用效率的规律, 发现水分利用效率和叶肉导度与气孔导度的比值成正比.这个关系表明水分利用效率只有在固碳能力的提高高于水分丢失增加(气孔导度升高)的情况下, 才可以升高.可以预测, 同样的关系也应该在气孔导度斜率及Vcmax和气孔导度的比值间存在.临界氮利用效率也被证明和临界水分利用效率有互补的关系(Palmroth et al., 2013). ...
... ; Ewers et al., 2005; Buckley & Roberts, 2006; Ambrose et al., 2010; Woodruff et al., 2010).木材密度和气孔导度斜率之间也存在着显著的负相关关系(Héroult et al., 2013; Lin et al., 2015).植物结构属性, 如根叶比、根深等以及水力传导结构属性也会影响气孔导度斜率(Litvak et al., 2012; McCulloh & Woodruff, 2012).如与散孔材木相比, 环孔材木的气孔对环境因素变化的反应更加敏感(Bush et al., 2008) , 但是在Litvak等(2012)的研究中并未发现木材结构影响气孔导度斜率的现象.而随着测量高度的增高, 气孔导度对饱和水汽压亏缺响应的敏感性也会降低.叶肉导度和Vcmax也通过影响光合作用影响植物的水分利用效率(Wang et al., 2013a).Flexas等(2013)分析了44种不同物种水分利用效率的规律, 发现水分利用效率和叶肉导度与气孔导度的比值成正比.这个关系表明水分利用效率只有在固碳能力的提高高于水分丢失增加(气孔导度升高)的情况下, 才可以升高.可以预测, 同样的关系也应该在气孔导度斜率及Vcmax和气孔导度的比值间存在.临界氮利用效率也被证明和临界水分利用效率有互补的关系(Palmroth et al., 2013). ...
Intercomparison of sugar maple (Acer saccharum Marsh.) stand transpiration responses to environmental conditions from the Western Great Lakes Region of the United States.
1
2008
... 气孔导度斜率的大小与气孔导度和气孔密度有关(Oren et al., 1999; Pataki & Oren, 2003; Ewers et al., 2008).当气孔导度较小时, 一点小小的改变就会给叶温带来极大的影响, 这时的气孔表现极其敏感.最高气孔导度斜率往往出现在0.4以下的低气孔导度条件下, 0.4以上的气孔导度, 响应机制会变得比较平缓(Hetherington & Woodward, 2003). ...
A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species.
1
1980
... mol-1 H2O), 实验证明这一数值在一定时间内不会随温湿度变化而改变(Hall & Schulze, 1980; Field et al., 1982), 但是会因土壤含水量减小而下降(Farquhar et al., 1980).最优气孔理论被广为接受, 而且可以为气孔导度模型所借鉴, 以研究不同环境和生物因素对气孔行为的影响.然而由于对λ的有限理解及由其所推出的模型无法准确描述气孔对CO2的响应规律(Thomas et al., 1999), 这一理论还没有得到广泛 应用. ...
A portable system for measuring carbon dioxide and water vapour exchange of leaves.
1
1982
... mol-1 H2O), 实验证明这一数值在一定时间内不会随温湿度变化而改变(Hall & Schulze, 1980; Field et al., 1982), 但是会因土壤含水量减小而下降(Farquhar et al., 1980).最优气孔理论被广为接受, 而且可以为气孔导度模型所借鉴, 以研究不同环境和生物因素对气孔行为的影响.然而由于对λ的有限理解及由其所推出的模型无法准确描述气孔对CO2的响应规律(Thomas et al., 1999), 这一理论还没有得到广泛 应用. ...
Diffusional conductances to CO2 as a target for increasing photosynthesis and photosynthetic water-use efficiency.
2013
Stomatal control and hydraulic conductance, with special reference to tall trees.
1
2004
... 以往研究显示叶片水势(Oren et al., 1999)、叶片、枝条、根部和整株植物水力传导(Salleo et al., 2000; Addington et al., 2004; Bunce, 2006; Domec et al., 2009)、叶片相对含水量(Ewers et al., 2005)等会影响气孔导度对饱和水汽压亏缺的响应.而由于树龄和树高都会影响水力传导和气孔导度, 进而都会影响气孔导度斜率(Schäfer et al., 2000; Franks, 2004; Ewers et al., 2005; Buckley & Roberts, 2006; Ambrose et al., 2010; Woodruff et al., 2010).木材密度和气孔导度斜率之间也存在着显著的负相关关系(Héroult et al., 2013; Lin et al., 2015).植物结构属性, 如根叶比、根深等以及水力传导结构属性也会影响气孔导度斜率(Litvak et al., 2012; McCulloh & Woodruff, 2012).如与散孔材木相比, 环孔材木的气孔对环境因素变化的反应更加敏感(Bush et al., 2008) , 但是在Litvak等(2012)的研究中并未发现木材结构影响气孔导度斜率的现象.而随着测量高度的增高, 气孔导度对饱和水汽压亏缺响应的敏感性也会降低.叶肉导度和Vcmax也通过影响光合作用影响植物的水分利用效率(Wang et al., 2013a).Flexas等(2013)分析了44种不同物种水分利用效率的规律, 发现水分利用效率和叶肉导度与气孔导度的比值成正比.这个关系表明水分利用效率只有在固碳能力的提高高于水分丢失增加(气孔导度升高)的情况下, 才可以升高.可以预测, 同样的关系也应该在气孔导度斜率及Vcmax和气孔导度的比值间存在.临界氮利用效率也被证明和临界水分利用效率有互补的关系(Palmroth et al., 2013). ...
干旱胁迫对杨树光合生理指标的影响
1
2006
... 在很多生态系统模型中, 土壤干旱胁迫对气孔导度影响的模拟通常是通过改变水分胁迫与光合作用/气孔导度之间的作用关系来实现的(Sala & Tenhunen, 1996; Verhoef & Allen, 2000).但也有模型是通过改变水分胁迫与光合作用或者光合作用参数Vcmax和最大电子传输速率(Jmax)的关系, 模拟水分胁迫下光合作用的改变进而模拟气孔导度的改变(Sellers et al., 1996; Ronda et al., 2001; Best et al., 2011).虽然在轻度干旱情况下, 上述几种做法可以模拟出水分胁迫对叶片尺度碳水通量的影响, 但在严重干旱情况下无法同时准确模拟光合作用和蒸腾作用(Verhoef & Allen, 2000; Reichstein et al., 2002; Keenan et al., 2009).Egea等(2011)提议, 在干旱条件下可以通过先限制叶肉导度, 进而限制气孔导度和光合作用参数的方案来提高光合作用和气孔导度模拟的准确性.Zhou等(2013)的研究结果也表明模拟干旱状况下的气孔导度, 必须同时考虑干旱对气孔和非气孔因素(如Vcmax)的综合影响.由于对干旱状况下气孔导度预测的不准确性, 现有的模型一般都倾向于高估光合作用和蒸腾作用对干旱的响应(Dietze, 2014).与以前从气孔(物理过程)或者非气孔(生化过程)的角度出发研究干旱对光合作用和气孔导度的影响不同, 气孔导度机理模型中的气孔导度斜率在干旱环境下的变化规律包含了干旱对气孔(生物物理因素)和非气孔因素, 包括最大光合效率和临界水分利用效率的综合影响, 为研究全球变化下陆地生态系统碳水耦合的适应性提供了一个简明且综合的框架.Manzoni等(2011)计算了50种植物的临界水分利用效率, 发现在忽略表皮水分丢失的情况下, 植物临界水分利用效率在轻微干旱的情况下增高, 而在严重干旱的情况下下降; 如果不忽略表皮水分丢失, 植物临界水分利用效率会随着水分胁迫的增加而提高; 而不同的植物功能型和不同的气候条件下, 反应也会不同.总体说来, 土壤水分较少时, 植物的水分利用效率会增高, 此时气孔导度斜率值也会降低(Lin et al., 2015).在环境水分条件与气孔导度斜率的变化规律之间还有很多值得讨论的地方, 我国学者对于水分利用的研究主要集中在单叶水分利用效率(付士磊等, 2006; 杨建伟等, 2006; 全先奎和王传宽, 2015)、气孔导度时空变异的调查(司建华等, 2008)、气孔导度对水分环境的响应(鱼腾飞等, 2012; 袁国富等, 2012)、单叶或者幼苗蒸腾速率(宋炳煜等, 2004; 岳广阳等, 2009)、树干液流密度(赵平等, 2006)、土壤水分定位监测(尹春梅等, 2008), 以及涡度协相关法测量生态系统水分利用效率(Zhou et al., 2013)等方面.土壤干旱如何影响不同功能型和物种的气孔导度斜率和临界水分利用效率, 还有待于进一步的研究. ...
Modelling stomatal conductance of field-grown sunflower under varying soil water content and leaf environment: Comparison of three models of stomatal response to leaf environment and coupling with an abscisic acid-based model of stomatal response to soil drying.
2002
Trends in wood density and structure are linked to prevention of xylem implosion by negative pressure.
1
2001
... 不同植物功能型的气孔导度斜率值大多差异较大, 农作物的气孔导度斜率值最大, 裸子植物树种的斜率值最小(Zhou et al., 2013).Medley等(2011)比较了8种不同物种的气孔导度斜率, 发现被子植物气孔导度斜率高于针叶树种.Lin等(2015)发现: 对于被子植物来说, 随着植物干材密度的增加, 气孔导度斜率值会下降, 而裸子植物却没有这样的规律.这可能与被子植物提供支撑作用和水分运输作用的是两套系统, 而裸子植物中管胞同时提供了这两项功能相关(Hacke et al., 2001).Way等(2014)发现: C3植物与C3-C4中间体植物相比, 临界水分利用效率并没有显著差异, 而具有C4循环的近C4中间体植物的临界水分利用效率比C3-C4中间体植物显著增强. ...
Drought effects on transpiration and leaf water status of cowpea in controlled environments.
1
1980
... mol-1 H2O), 实验证明这一数值在一定时间内不会随温湿度变化而改变(Hall & Schulze, 1980; Field et al., 1982), 但是会因土壤含水量减小而下降(Farquhar et al., 1980).最优气孔理论被广为接受, 而且可以为气孔导度模型所借鉴, 以研究不同环境和生物因素对气孔行为的影响.然而由于对λ的有限理解及由其所推出的模型无法准确描述气孔对CO2的响应规律(Thomas et al., 1999), 这一理论还没有得到广泛 应用. ...
Modelling photosynthesis of cotton grown in elevated CO2.
1
1992
... 虽然光合作用、气孔导度的调查数据在文献中大量存在, 但是由于实验设计的不同以及缺少饱和水汽压亏缺等环境参数的信息, 很少有实验数据能够满足对气孔导度斜率进行模拟计算的要求(Zhou et al., 2013).以往大多数关于气孔导度斜率的研究都是在温室里的短期试验, 在自然条件下的研究还很少(Medlyn et al., 2011).对于增高的CO2浓度下气孔导度对环境因子的适应性变化的研究结果也不一致.在高CO2浓度的未来环境下, 植物会降低气孔导度和提高光合速率(Long et al., 2004; Bernacchi et al., 2005).有学者在培养箱中升高CO2浓度对大豆(Glycine max)和棉花(Gossypium hirsutum)进行培养研究得出, 气孔导度斜率会因CO2浓度的增加而降低(Harley et al., 1992; Bunce, 2004).但是Leakey等(2006)在FACE (free-air CO2 enrichment)实验样地对大豆进行了生长和生理参数测量, 他们的实验结果却表明气孔导度斜率不会因外界CO2浓度的升高而改变, 并且指出前人所得的气孔导度斜率降低的结论可能是增高的CO2浓度对气孔导度带来的瞬时改变的结果.Medlyn等(2001)也测定了生长在高CO2浓度下6种不同树种的生理参数, 发现CO2浓度升高并没有改变气孔导度斜率.这一结果也在Katul等(2009)的研究中得到证实, Katul等(2009)也发现CO2浓度升高并没有改变Ball-Berry模型中的光合作用与气孔导度的线性关系的斜率. ...
Optimal stomatal conductance in relation to photosynthesis in climatically contrasting Eucalyptus species under drought.
2
2013
... Medlyn等(2011)的机理模型与Ball-Berry和Leuning的半经验模型相比, 其对气孔导度的模拟更加准确, 且模型中的g1与临界水分利用效率具有一定的比例关系(方程(4)), 这一参数因此具有了生物学含义, 可以用来描述植物的水分利用策略(Medlyn et al., 2011; Héroult et al., 2013). ...
... 以往研究显示叶片水势(Oren et al., 1999)、叶片、枝条、根部和整株植物水力传导(Salleo et al., 2000; Addington et al., 2004; Bunce, 2006; Domec et al., 2009)、叶片相对含水量(Ewers et al., 2005)等会影响气孔导度对饱和水汽压亏缺的响应.而由于树龄和树高都会影响水力传导和气孔导度, 进而都会影响气孔导度斜率(Schäfer et al., 2000; Franks, 2004; Ewers et al., 2005; Buckley & Roberts, 2006; Ambrose et al., 2010; Woodruff et al., 2010).木材密度和气孔导度斜率之间也存在着显著的负相关关系(Héroult et al., 2013; Lin et al., 2015).植物结构属性, 如根叶比、根深等以及水力传导结构属性也会影响气孔导度斜率(Litvak et al., 2012; McCulloh & Woodruff, 2012).如与散孔材木相比, 环孔材木的气孔对环境因素变化的反应更加敏感(Bush et al., 2008) , 但是在Litvak等(2012)的研究中并未发现木材结构影响气孔导度斜率的现象.而随着测量高度的增高, 气孔导度对饱和水汽压亏缺响应的敏感性也会降低.叶肉导度和Vcmax也通过影响光合作用影响植物的水分利用效率(Wang et al., 2013a).Flexas等(2013)分析了44种不同物种水分利用效率的规律, 发现水分利用效率和叶肉导度与气孔导度的比值成正比.这个关系表明水分利用效率只有在固碳能力的提高高于水分丢失增加(气孔导度升高)的情况下, 才可以升高.可以预测, 同样的关系也应该在气孔导度斜率及Vcmax和气孔导度的比值间存在.临界氮利用效率也被证明和临界水分利用效率有互补的关系(Palmroth et al., 2013). ...
The role of stomata in sensing and driving environmental change.
... 气孔导度斜率的大小与气孔导度和气孔密度有关(Oren et al., 1999; Pataki & Oren, 2003; Ewers et al., 2008).当气孔导度较小时, 一点小小的改变就会给叶温带来极大的影响, 这时的气孔表现极其敏感.最高气孔导度斜率往往出现在0.4以下的低气孔导度条件下, 0.4以上的气孔导度, 响应机制会变得比较平缓(Hetherington & Woodward, 2003). ...
1
... 以碳积累为代表的光合作用和以水分消耗为代表的蒸腾作用是所有植物功能模型的基础, 准确地模拟光合作用和蒸腾作用也是生态系统模型、植被模型、地球系统模型能够有效模拟碳、水和能量循环的必要条件(Wang et al., 2013b, 2015, 2016).由于以Farquhar等(1980)的光合作用模型为代表的机理模型的广泛应用, 通过对物种生物属性和环境参数(如光合最大羧化速率(Vcmax)、CO2浓度、温度和光照)的测量, 可以有效地模拟光合作用(Wullschleger, 1993; Kattge et al., 2009).相对于光合作用的机理模型, 蒸腾作用通常都是由经验或者半经验模型模拟出的气孔导度值来计算.过去40年里, 气孔导度的模拟工作大量开展.概括来讲, 对气孔导度的模拟有以下几种方式: (1)经验模型.大多通过气孔导度与多个环境因素如空气湿度(Lange et al., 1971)、水汽压亏缺(VPD)(Monteith, 1995)、叶片温度(Jarvis, 1976; Jones, 1992)和臭氧(Uddling et al., 2009)等的统计关系建立模型; (2)半经验模型.基于植物生理特性的假设, 但是仍然和经验模型结合, 如基于气孔导度和光合作用(Ball et al., 1987; Leuning, 1990, 1995; Misson et al., 2004b)、脱落酸(ABA) (Tardieu & Davies, 1993; Guteschick & Simonneau, 2002)、水分传导(Sperry et al., 1998; Oren et al., 1999; Tuzet et al., 2003)和膨压调节(Dewar, 2002; Buckley et al., 2003)等的关系建立的模型, 其参数不具有生物学属性; (3)机理模型.基于植物生理特性的假设, 其参数具有生物学特性, 并具有可以应用在各种环境条件下的潜力(Dewar, 2002; Medlyn et al., 2011; Prentice et al., 2014).很多文献已经综述过气孔导度的经验与半经验模型, 本文将侧重阐述气孔导度机理模型的研究. ...
The interpretation of the variations in leaf water potential and stomatal conductance found in canopies in the field.
1976
Influence of leaf area index prescriptions on simulations of heat, moisture, and carbon fluxes.
1
2014
... 以碳积累为代表的光合作用和以水分消耗为代表的蒸腾作用是所有植物功能模型的基础, 准确地模拟光合作用和蒸腾作用也是生态系统模型、植被模型、地球系统模型能够有效模拟碳、水和能量循环的必要条件(Wang et al., 2013b, 2015, 2016).由于以Farquhar等(1980)的光合作用模型为代表的机理模型的广泛应用, 通过对物种生物属性和环境参数(如光合最大羧化速率(Vcmax)、CO2浓度、温度和光照)的测量, 可以有效地模拟光合作用(Wullschleger, 1993; Kattge et al., 2009).相对于光合作用的机理模型, 蒸腾作用通常都是由经验或者半经验模型模拟出的气孔导度值来计算.过去40年里, 气孔导度的模拟工作大量开展.概括来讲, 对气孔导度的模拟有以下几种方式: (1)经验模型.大多通过气孔导度与多个环境因素如空气湿度(Lange et al., 1971)、水汽压亏缺(VPD)(Monteith, 1995)、叶片温度(Jarvis, 1976; Jones, 1992)和臭氧(Uddling et al., 2009)等的统计关系建立模型; (2)半经验模型.基于植物生理特性的假设, 但是仍然和经验模型结合, 如基于气孔导度和光合作用(Ball et al., 1987; Leuning, 1990, 1995; Misson et al., 2004b)、脱落酸(ABA) (Tardieu & Davies, 1993; Guteschick & Simonneau, 2002)、水分传导(Sperry et al., 1998; Oren et al., 1999; Tuzet et al., 2003)和膨压调节(Dewar, 2002; Buckley et al., 2003)等的关系建立的模型, 其参数不具有生物学属性; (3)机理模型.基于植物生理特性的假设, 其参数具有生物学特性, 并具有可以应用在各种环境条件下的潜力(Dewar, 2002; Medlyn et al., 2011; Prentice et al., 2014).很多文献已经综述过气孔导度的经验与半经验模型, 本文将侧重阐述气孔导度机理模型的研究. ...
Quantifying photosynthetic capacity and its relationship to leaf nitrogen content for global-scale terrestrial biosphere models.
2009
Leaf stomatal responses to vapour pressure deficit under current and CO2-enriched atmosphere explained by the economics of gas exchange.
1
2009
... 在很多生态系统模型中, 土壤干旱胁迫对气孔导度影响的模拟通常是通过改变水分胁迫与光合作用/气孔导度之间的作用关系来实现的(Sala & Tenhunen, 1996; Verhoef & Allen, 2000).但也有模型是通过改变水分胁迫与光合作用或者光合作用参数Vcmax和最大电子传输速率(Jmax)的关系, 模拟水分胁迫下光合作用的改变进而模拟气孔导度的改变(Sellers et al., 1996; Ronda et al., 2001; Best et al., 2011).虽然在轻度干旱情况下, 上述几种做法可以模拟出水分胁迫对叶片尺度碳水通量的影响, 但在严重干旱情况下无法同时准确模拟光合作用和蒸腾作用(Verhoef & Allen, 2000; Reichstein et al., 2002; Keenan et al., 2009).Egea等(2011)提议, 在干旱条件下可以通过先限制叶肉导度, 进而限制气孔导度和光合作用参数的方案来提高光合作用和气孔导度模拟的准确性.Zhou等(2013)的研究结果也表明模拟干旱状况下的气孔导度, 必须同时考虑干旱对气孔和非气孔因素(如Vcmax)的综合影响.由于对干旱状况下气孔导度预测的不准确性, 现有的模型一般都倾向于高估光合作用和蒸腾作用对干旱的响应(Dietze, 2014).与以前从气孔(物理过程)或者非气孔(生化过程)的角度出发研究干旱对光合作用和气孔导度的影响不同, 气孔导度机理模型中的气孔导度斜率在干旱环境下的变化规律包含了干旱对气孔(生物物理因素)和非气孔因素, 包括最大光合效率和临界水分利用效率的综合影响, 为研究全球变化下陆地生态系统碳水耦合的适应性提供了一个简明且综合的框架.Manzoni等(2011)计算了50种植物的临界水分利用效率, 发现在忽略表皮水分丢失的情况下, 植物临界水分利用效率在轻微干旱的情况下增高, 而在严重干旱的情况下下降; 如果不忽略表皮水分丢失, 植物临界水分利用效率会随着水分胁迫的增加而提高; 而不同的植物功能型和不同的气候条件下, 反应也会不同.总体说来, 土壤水分较少时, 植物的水分利用效率会增高, 此时气孔导度斜率值也会降低(Lin et al., 2015).在环境水分条件与气孔导度斜率的变化规律之间还有很多值得讨论的地方, 我国学者对于水分利用的研究主要集中在单叶水分利用效率(付士磊等, 2006; 杨建伟等, 2006; 全先奎和王传宽, 2015)、气孔导度时空变异的调查(司建华等, 2008)、气孔导度对水分环境的响应(鱼腾飞等, 2012; 袁国富等, 2012)、单叶或者幼苗蒸腾速率(宋炳煜等, 2004; 岳广阳等, 2009)、树干液流密度(赵平等, 2006)、土壤水分定位监测(尹春梅等, 2008), 以及涡度协相关法测量生态系统水分利用效率(Zhou et al., 2013)等方面.土壤干旱如何影响不同功能型和物种的气孔导度斜率和临界水分利用效率, 还有待于进一步的研究. ...
Improved understanding of drought controls on seasonal variation in mediterranean forest canopy CO2 and water fluxes through combined in situ measurements and ecosystem modelling.
1
2009
... 此后多人对Ball-Berry模型进行了优化、改进和应用, 如加入叶片水势的作用等(Sellers et al., 1992; Kim & Lieth, 2003; Tuzet et al., 2003; Yu et al., 2004), 但其本质上依然是半经验模型.在对Ball-Berry模型的众多改进中, Yu等(2004)的模型较为突出.此模型结合了Jarvis模型和Ball-Berry模型的优点, 采用了更便于计算的模型参数, 基于气孔与光合作用的联系及光合作用与光照之间的联系, 推导出了双曲线式的气孔导度模型, 使得直接从气象参数计算气孔导度成为可能.模型简洁、实用, 对于实测数据符合较好. ...
A coupled model of photosynthesis, stomatal conductance and transpiration for a rose leaf (Rosa hybrida L.).
2
2003
... 目前盛行的生态系统模型多是通过减小m值来模拟干旱对气孔导度和光合作用的影响(Wang et al., 2013b, 2015); 或者通过减小光合作用模型中的Vcmax来间接地减小气孔导度和光合作用来实现干旱条件下的生长模拟(Krinner et al., 2005).没有叶片尺度上气孔导度模拟的准确性, 陆地生态系统模型将无法准确地预测冠层、生态系统乃至区域尺度上碳、水循环的变化(Damour et al., 2010).成功模拟气孔行为的机理模型将为理解碳水耦合的适应过程以及陆地植被模型对植被结构、功能和演替的准确把握提供一个有效的框架. ...
... 大多数地球系统模型或者植被模型都采用经验或者半经验模型来模拟气孔导度(Sellers et al., 1992, 1997; Bonan, 1995; Wang et al., 2013b, 2015).虽然对于经验或半经验模型来说, 可以利用实验数据矫正参数, 但是在现实中, 并非如此.如上文中所提到的, 多数模型都只是区分了C3和C4植物的气孔导度模型参数m值(Krinner et al., 2005; Oleson et al., 2013). De Kauw等(2015)首次将气孔导度机理模型(Medlyn et al., 2011)耦合到陆地表面模型CABLE (Community Atmosphere Biosphere Land Exchange)中, 研究结果显示: 相比于气孔导度经验模型, 耦合气孔导度机理模型导致在常绿针叶林、苔原和C4草原区域, 年蒸腾量减少了30%.Kala等(2014)进一步耦合气孔导度机理模型、CABLE和ACCESS (Australian Community Climate and Earth Systems Simulator)模型, 研究结果显示在北半球的夏季, 新的模型预测的温带森林潜热通量每天减少0.5-1.0 mm, 直接导致每天最高和最低温度升高了1 ℃, 而极端高温升高了1.5 ℃.地表的能量平衡也将进一步影响净初级生产力和陆地碳平衡.由此可见, 深入研究气孔导度斜率的变化规律并提高气孔导度预测的准确性, 可大大减小生态系统模型对碳水通量预测的不确定性, 这对区域尺度生态系统碳水平衡的研究至关重要.然而, 由于对气孔导度斜率变化规律的理解还十分有限, 气孔导度机理模型在植被和地球系统模型中的应用还需要加强. ...
A dynamic global vegetation model for studies of the coupled atmosphere-biosphere system.
1
2005
... 以碳积累为代表的光合作用和以水分消耗为代表的蒸腾作用是所有植物功能模型的基础, 准确地模拟光合作用和蒸腾作用也是生态系统模型、植被模型、地球系统模型能够有效模拟碳、水和能量循环的必要条件(Wang et al., 2013b, 2015, 2016).由于以Farquhar等(1980)的光合作用模型为代表的机理模型的广泛应用, 通过对物种生物属性和环境参数(如光合最大羧化速率(Vcmax)、CO2浓度、温度和光照)的测量, 可以有效地模拟光合作用(Wullschleger, 1993; Kattge et al., 2009).相对于光合作用的机理模型, 蒸腾作用通常都是由经验或者半经验模型模拟出的气孔导度值来计算.过去40年里, 气孔导度的模拟工作大量开展.概括来讲, 对气孔导度的模拟有以下几种方式: (1)经验模型.大多通过气孔导度与多个环境因素如空气湿度(Lange et al., 1971)、水汽压亏缺(VPD)(Monteith, 1995)、叶片温度(Jarvis, 1976; Jones, 1992)和臭氧(Uddling et al., 2009)等的统计关系建立模型; (2)半经验模型.基于植物生理特性的假设, 但是仍然和经验模型结合, 如基于气孔导度和光合作用(Ball et al., 1987; Leuning, 1990, 1995; Misson et al., 2004b)、脱落酸(ABA) (Tardieu & Davies, 1993; Guteschick & Simonneau, 2002)、水分传导(Sperry et al., 1998; Oren et al., 1999; Tuzet et al., 2003)和膨压调节(Dewar, 2002; Buckley et al., 2003)等的关系建立的模型, 其参数不具有生物学属性; (3)机理模型.基于植物生理特性的假设, 其参数具有生物学特性, 并具有可以应用在各种环境条件下的潜力(Dewar, 2002; Medlyn et al., 2011; Prentice et al., 2014).很多文献已经综述过气孔导度的经验与半经验模型, 本文将侧重阐述气孔导度机理模型的研究. ...
Responses of stomata to changes in humidity.
1971
Long-term growth of soybean at elevated [CO2] does not cause acclimation of stomatal conductance under fully open-air conditions.
1
2006
... 以碳积累为代表的光合作用和以水分消耗为代表的蒸腾作用是所有植物功能模型的基础, 准确地模拟光合作用和蒸腾作用也是生态系统模型、植被模型、地球系统模型能够有效模拟碳、水和能量循环的必要条件(Wang et al., 2013b, 2015, 2016).由于以Farquhar等(1980)的光合作用模型为代表的机理模型的广泛应用, 通过对物种生物属性和环境参数(如光合最大羧化速率(Vcmax)、CO2浓度、温度和光照)的测量, 可以有效地模拟光合作用(Wullschleger, 1993; Kattge et al., 2009).相对于光合作用的机理模型, 蒸腾作用通常都是由经验或者半经验模型模拟出的气孔导度值来计算.过去40年里, 气孔导度的模拟工作大量开展.概括来讲, 对气孔导度的模拟有以下几种方式: (1)经验模型.大多通过气孔导度与多个环境因素如空气湿度(Lange et al., 1971)、水汽压亏缺(VPD)(Monteith, 1995)、叶片温度(Jarvis, 1976; Jones, 1992)和臭氧(Uddling et al., 2009)等的统计关系建立模型; (2)半经验模型.基于植物生理特性的假设, 但是仍然和经验模型结合, 如基于气孔导度和光合作用(Ball et al., 1987; Leuning, 1990, 1995; Misson et al., 2004b)、脱落酸(ABA) (Tardieu & Davies, 1993; Guteschick & Simonneau, 2002)、水分传导(Sperry et al., 1998; Oren et al., 1999; Tuzet et al., 2003)和膨压调节(Dewar, 2002; Buckley et al., 2003)等的关系建立的模型, 其参数不具有生物学属性; (3)机理模型.基于植物生理特性的假设, 其参数具有生物学特性, 并具有可以应用在各种环境条件下的潜力(Dewar, 2002; Medlyn et al., 2011; Prentice et al., 2014).很多文献已经综述过气孔导度的经验与半经验模型, 本文将侧重阐述气孔导度机理模型的研究. ...
Modelling stomatal behaviour and and photosynthesis of
1
1990
... 以碳积累为代表的光合作用和以水分消耗为代表的蒸腾作用是所有植物功能模型的基础, 准确地模拟光合作用和蒸腾作用也是生态系统模型、植被模型、地球系统模型能够有效模拟碳、水和能量循环的必要条件(Wang et al., 2013b, 2015, 2016).由于以Farquhar等(1980)的光合作用模型为代表的机理模型的广泛应用, 通过对物种生物属性和环境参数(如光合最大羧化速率(Vcmax)、CO2浓度、温度和光照)的测量, 可以有效地模拟光合作用(Wullschleger, 1993; Kattge et al., 2009).相对于光合作用的机理模型, 蒸腾作用通常都是由经验或者半经验模型模拟出的气孔导度值来计算.过去40年里, 气孔导度的模拟工作大量开展.概括来讲, 对气孔导度的模拟有以下几种方式: (1)经验模型.大多通过气孔导度与多个环境因素如空气湿度(Lange et al., 1971)、水汽压亏缺(VPD)(Monteith, 1995)、叶片温度(Jarvis, 1976; Jones, 1992)和臭氧(Uddling et al., 2009)等的统计关系建立模型; (2)半经验模型.基于植物生理特性的假设, 但是仍然和经验模型结合, 如基于气孔导度和光合作用(Ball et al., 1987; Leuning, 1990, 1995; Misson et al., 2004b)、脱落酸(ABA) (Tardieu & Davies, 1993; Guteschick & Simonneau, 2002)、水分传导(Sperry et al., 1998; Oren et al., 1999; Tuzet et al., 2003)和膨压调节(Dewar, 2002; Buckley et al., 2003)等的关系建立的模型, 其参数不具有生物学属性; (3)机理模型.基于植物生理特性的假设, 其参数具有生物学特性, 并具有可以应用在各种环境条件下的潜力(Dewar, 2002; Medlyn et al., 2011; Prentice et al., 2014).很多文献已经综述过气孔导度的经验与半经验模型, 本文将侧重阐述气孔导度机理模型的研究. ...
A critical appraisal of a combined stomatal- photosynthesis model for C3 plants.
... 在很多生态系统模型中, 土壤干旱胁迫对气孔导度影响的模拟通常是通过改变水分胁迫与光合作用/气孔导度之间的作用关系来实现的(Sala & Tenhunen, 1996; Verhoef & Allen, 2000).但也有模型是通过改变水分胁迫与光合作用或者光合作用参数Vcmax和最大电子传输速率(Jmax)的关系, 模拟水分胁迫下光合作用的改变进而模拟气孔导度的改变(Sellers et al., 1996; Ronda et al., 2001; Best et al., 2011).虽然在轻度干旱情况下, 上述几种做法可以模拟出水分胁迫对叶片尺度碳水通量的影响, 但在严重干旱情况下无法同时准确模拟光合作用和蒸腾作用(Verhoef & Allen, 2000; Reichstein et al., 2002; Keenan et al., 2009).Egea等(2011)提议, 在干旱条件下可以通过先限制叶肉导度, 进而限制气孔导度和光合作用参数的方案来提高光合作用和气孔导度模拟的准确性.Zhou等(2013)的研究结果也表明模拟干旱状况下的气孔导度, 必须同时考虑干旱对气孔和非气孔因素(如Vcmax)的综合影响.由于对干旱状况下气孔导度预测的不准确性, 现有的模型一般都倾向于高估光合作用和蒸腾作用对干旱的响应(Dietze, 2014).与以前从气孔(物理过程)或者非气孔(生化过程)的角度出发研究干旱对光合作用和气孔导度的影响不同, 气孔导度机理模型中的气孔导度斜率在干旱环境下的变化规律包含了干旱对气孔(生物物理因素)和非气孔因素, 包括最大光合效率和临界水分利用效率的综合影响, 为研究全球变化下陆地生态系统碳水耦合的适应性提供了一个简明且综合的框架.Manzoni等(2011)计算了50种植物的临界水分利用效率, 发现在忽略表皮水分丢失的情况下, 植物临界水分利用效率在轻微干旱的情况下增高, 而在严重干旱的情况下下降; 如果不忽略表皮水分丢失, 植物临界水分利用效率会随着水分胁迫的增加而提高; 而不同的植物功能型和不同的气候条件下, 反应也会不同.总体说来, 土壤水分较少时, 植物的水分利用效率会增高, 此时气孔导度斜率值也会降低(Lin et al., 2015).在环境水分条件与气孔导度斜率的变化规律之间还有很多值得讨论的地方, 我国学者对于水分利用的研究主要集中在单叶水分利用效率(付士磊等, 2006; 杨建伟等, 2006; 全先奎和王传宽, 2015)、气孔导度时空变异的调查(司建华等, 2008)、气孔导度对水分环境的响应(鱼腾飞等, 2012; 袁国富等, 2012)、单叶或者幼苗蒸腾速率(宋炳煜等, 2004; 岳广阳等, 2009)、树干液流密度(赵平等, 2006)、土壤水分定位监测(尹春梅等, 2008), 以及涡度协相关法测量生态系统水分利用效率(Zhou et al., 2013)等方面.土壤干旱如何影响不同功能型和物种的气孔导度斜率和临界水分利用效率, 还有待于进一步的研究. ...
... 以往研究显示叶片水势(Oren et al., 1999)、叶片、枝条、根部和整株植物水力传导(Salleo et al., 2000; Addington et al., 2004; Bunce, 2006; Domec et al., 2009)、叶片相对含水量(Ewers et al., 2005)等会影响气孔导度对饱和水汽压亏缺的响应.而由于树龄和树高都会影响水力传导和气孔导度, 进而都会影响气孔导度斜率(Schäfer et al., 2000; Franks, 2004; Ewers et al., 2005; Buckley & Roberts, 2006; Ambrose et al., 2010; Woodruff et al., 2010).木材密度和气孔导度斜率之间也存在着显著的负相关关系(Héroult et al., 2013; Lin et al., 2015).植物结构属性, 如根叶比、根深等以及水力传导结构属性也会影响气孔导度斜率(Litvak et al., 2012; McCulloh & Woodruff, 2012).如与散孔材木相比, 环孔材木的气孔对环境因素变化的反应更加敏感(Bush et al., 2008) , 但是在Litvak等(2012)的研究中并未发现木材结构影响气孔导度斜率的现象.而随着测量高度的增高, 气孔导度对饱和水汽压亏缺响应的敏感性也会降低.叶肉导度和Vcmax也通过影响光合作用影响植物的水分利用效率(Wang et al., 2013a).Flexas等(2013)分析了44种不同物种水分利用效率的规律, 发现水分利用效率和叶肉导度与气孔导度的比值成正比.这个关系表明水分利用效率只有在固碳能力的提高高于水分丢失增加(气孔导度升高)的情况下, 才可以升高.可以预测, 同样的关系也应该在气孔导度斜率及Vcmax和气孔导度的比值间存在.临界氮利用效率也被证明和临界水分利用效率有互补的关系(Palmroth et al., 2013). ...
Optimal stomatal behaviour around the world.
1
2015
... 以往研究显示叶片水势(Oren et al., 1999)、叶片、枝条、根部和整株植物水力传导(Salleo et al., 2000; Addington et al., 2004; Bunce, 2006; Domec et al., 2009)、叶片相对含水量(Ewers et al., 2005)等会影响气孔导度对饱和水汽压亏缺的响应.而由于树龄和树高都会影响水力传导和气孔导度, 进而都会影响气孔导度斜率(Schäfer et al., 2000; Franks, 2004; Ewers et al., 2005; Buckley & Roberts, 2006; Ambrose et al., 2010; Woodruff et al., 2010).木材密度和气孔导度斜率之间也存在着显著的负相关关系(Héroult et al., 2013; Lin et al., 2015).植物结构属性, 如根叶比、根深等以及水力传导结构属性也会影响气孔导度斜率(Litvak et al., 2012; McCulloh & Woodruff, 2012).如与散孔材木相比, 环孔材木的气孔对环境因素变化的反应更加敏感(Bush et al., 2008) , 但是在Litvak等(2012)的研究中并未发现木材结构影响气孔导度斜率的现象.而随着测量高度的增高, 气孔导度对饱和水汽压亏缺响应的敏感性也会降低.叶肉导度和Vcmax也通过影响光合作用影响植物的水分利用效率(Wang et al., 2013a).Flexas等(2013)分析了44种不同物种水分利用效率的规律, 发现水分利用效率和叶肉导度与气孔导度的比值成正比.这个关系表明水分利用效率只有在固碳能力的提高高于水分丢失增加(气孔导度升高)的情况下, 才可以升高.可以预测, 同样的关系也应该在气孔导度斜率及Vcmax和气孔导度的比值间存在.临界氮利用效率也被证明和临界水分利用效率有互补的关系(Palmroth et al., 2013). ...
Transpiration sensitivity of urban trees in a semi-arid climate is constrained by xylem vulnerability to cavitation.
1
2012
... 虽然光合作用、气孔导度的调查数据在文献中大量存在, 但是由于实验设计的不同以及缺少饱和水汽压亏缺等环境参数的信息, 很少有实验数据能够满足对气孔导度斜率进行模拟计算的要求(Zhou et al., 2013).以往大多数关于气孔导度斜率的研究都是在温室里的短期试验, 在自然条件下的研究还很少(Medlyn et al., 2011).对于增高的CO2浓度下气孔导度对环境因子的适应性变化的研究结果也不一致.在高CO2浓度的未来环境下, 植物会降低气孔导度和提高光合速率(Long et al., 2004; Bernacchi et al., 2005).有学者在培养箱中升高CO2浓度对大豆(Glycine max)和棉花(Gossypium hirsutum)进行培养研究得出, 气孔导度斜率会因CO2浓度的增加而降低(Harley et al., 1992; Bunce, 2004).但是Leakey等(2006)在FACE (free-air CO2 enrichment)实验样地对大豆进行了生长和生理参数测量, 他们的实验结果却表明气孔导度斜率不会因外界CO2浓度的升高而改变, 并且指出前人所得的气孔导度斜率降低的结论可能是增高的CO2浓度对气孔导度带来的瞬时改变的结果.Medlyn等(2001)也测定了生长在高CO2浓度下6种不同树种的生理参数, 发现CO2浓度升高并没有改变气孔导度斜率.这一结果也在Katul等(2009)的研究中得到证实, Katul等(2009)也发现CO2浓度升高并没有改变Ball-Berry模型中的光合作用与气孔导度的线性关系的斜率. ...
Rising atmospheric carbon dioxide: Plants FACE the Future.
1
2004
... 经验模型通常都忽略了叶片生理特性和气孔导度之间的关系, 也经常缺少反馈机制, 并且需要对模型参数进行大量的调试, 这些都限制了气孔导度经验模型的应用.Jarvis模型(方程1)是气孔导度经验模型, 最早由Jarvis (1976)提出, 后来经过许多学者的优化改进(Stewart, 1988; White et al., 1999; Macfarlane et al., 2004; Misson et al., 2004a; Noe & Giersch, 2004). ...
The apparent feed-forward response to vapour pressure deficit of stomata in droughted, field-grown Eucalyptus globulus Labill.
1
2004
... 实验观测也表明气孔导度斜率与黎明叶片水势之间存在指数关系, 通过实验数据拟合可以得出二者之间的经验方程(Mäkelä et al., 1996; Tuzet et al., 2003; Zhou et al., 2013). ...
Optimal control of gas exchange during drought: Theoretical analysis.
1996
Optimizing stomatal conductance for maximum carbon gain under water stress: A meta-analysis across plant functional types and climates.
1
2011
... 以往研究显示叶片水势(Oren et al., 1999)、叶片、枝条、根部和整株植物水力传导(Salleo et al., 2000; Addington et al., 2004; Bunce, 2006; Domec et al., 2009)、叶片相对含水量(Ewers et al., 2005)等会影响气孔导度对饱和水汽压亏缺的响应.而由于树龄和树高都会影响水力传导和气孔导度, 进而都会影响气孔导度斜率(Schäfer et al., 2000; Franks, 2004; Ewers et al., 2005; Buckley & Roberts, 2006; Ambrose et al., 2010; Woodruff et al., 2010).木材密度和气孔导度斜率之间也存在着显著的负相关关系(Héroult et al., 2013; Lin et al., 2015).植物结构属性, 如根叶比、根深等以及水力传导结构属性也会影响气孔导度斜率(Litvak et al., 2012; McCulloh & Woodruff, 2012).如与散孔材木相比, 环孔材木的气孔对环境因素变化的反应更加敏感(Bush et al., 2008) , 但是在Litvak等(2012)的研究中并未发现木材结构影响气孔导度斜率的现象.而随着测量高度的增高, 气孔导度对饱和水汽压亏缺响应的敏感性也会降低.叶肉导度和Vcmax也通过影响光合作用影响植物的水分利用效率(Wang et al., 2013a).Flexas等(2013)分析了44种不同物种水分利用效率的规律, 发现水分利用效率和叶肉导度与气孔导度的比值成正比.这个关系表明水分利用效率只有在固碳能力的提高高于水分丢失增加(气孔导度升高)的情况下, 才可以升高.可以预测, 同样的关系也应该在气孔导度斜率及Vcmax和气孔导度的比值间存在.临界氮利用效率也被证明和临界水分利用效率有互补的关系(Palmroth et al., 2013). ...
Linking stomatal sensitivity and whole-tree hydraulic architecture.
2012
Stomatal conductance of forest species after long-term exposure to elevated CO2 concentration: A synthesis.
8
2001
... 以碳积累为代表的光合作用和以水分消耗为代表的蒸腾作用是所有植物功能模型的基础, 准确地模拟光合作用和蒸腾作用也是生态系统模型、植被模型、地球系统模型能够有效模拟碳、水和能量循环的必要条件(Wang et al., 2013b, 2015, 2016).由于以Farquhar等(1980)的光合作用模型为代表的机理模型的广泛应用, 通过对物种生物属性和环境参数(如光合最大羧化速率(Vcmax)、CO2浓度、温度和光照)的测量, 可以有效地模拟光合作用(Wullschleger, 1993; Kattge et al., 2009).相对于光合作用的机理模型, 蒸腾作用通常都是由经验或者半经验模型模拟出的气孔导度值来计算.过去40年里, 气孔导度的模拟工作大量开展.概括来讲, 对气孔导度的模拟有以下几种方式: (1)经验模型.大多通过气孔导度与多个环境因素如空气湿度(Lange et al., 1971)、水汽压亏缺(VPD)(Monteith, 1995)、叶片温度(Jarvis, 1976; Jones, 1992)和臭氧(Uddling et al., 2009)等的统计关系建立模型; (2)半经验模型.基于植物生理特性的假设, 但是仍然和经验模型结合, 如基于气孔导度和光合作用(Ball et al., 1987; Leuning, 1990, 1995; Misson et al., 2004b)、脱落酸(ABA) (Tardieu & Davies, 1993; Guteschick & Simonneau, 2002)、水分传导(Sperry et al., 1998; Oren et al., 1999; Tuzet et al., 2003)和膨压调节(Dewar, 2002; Buckley et al., 2003)等的关系建立的模型, 其参数不具有生物学属性; (3)机理模型.基于植物生理特性的假设, 其参数具有生物学特性, 并具有可以应用在各种环境条件下的潜力(Dewar, 2002; Medlyn et al., 2011; Prentice et al., 2014).很多文献已经综述过气孔导度的经验与半经验模型, 本文将侧重阐述气孔导度机理模型的研究. ...
... 依据最优气孔导度理论并借鉴Ball-Berry的半经验模型发展的气孔导度机理模型(推导过程见Medlyn et al., 2011)(方程3), 可以将模型中的斜率参数与广受肯定的最优气孔理论中的临界水分利用效率(λ = ∂A/∂E, mol CO2·mol-1 H2O)(Damour et al., 2010; Medlyn et al., 2011)联系在一起(方程4), 使得模型参数具有了重要的生物学含义. ...
... ; Medlyn et al., 2011)联系在一起(方程4), 使得模型参数具有了重要的生物学含义. ...
... Medlyn等(2011)的机理模型与Ball-Berry和Leuning的半经验模型相比, 其对气孔导度的模拟更加准确, 且模型中的g1与临界水分利用效率具有一定的比例关系(方程(4)), 这一参数因此具有了生物学含义, 可以用来描述植物的水分利用策略(Medlyn et al., 2011; Héroult et al., 2013). ...
... 对于外界环境的变化, 气孔导度斜率也会做出不同的响应.不同植物物种的水分利用策略不同, 在同样的湿度环境下气孔导度斜率会有差异, 同样的物种在不同水分环境下气孔导度斜率也会不同(Zhou et al., 2013).Lin等(2015)对超过1万条光合测量数据综合分析后发现, 在植物生长适宜的外界温湿度下, 随着温湿度升高, 气孔导度斜率都会增加.对于气孔导度斜率随温度升高而增加的原因, 有两种解释: 一是气孔导度斜率与CO2补偿点浓度相关(Medlyn et al., 2011), 而这一浓度会随温度升高而增加(Bernacchi et al., 2001); 二是水分子黏性会随温度升高而降低, 这就降低了运输水分的消耗, 从而增高了气孔导度斜率(Prentice et al., 2014).Héroult等(2013)报道了产地不同的4种桉属植物在相同环境条件下, 气孔导度斜率随着自然产地不同而不同, 自然生长在湿润地区的植物气孔导度斜率比在半湿润地区的植物高.在湿润环境生长的长叶松(Pinus palustris)气孔关闭对增加的饱和水汽压亏缺的反应比在干旱环境下更加敏感(Addingtong et al., 2006). ...
... 虽然光合作用、气孔导度的调查数据在文献中大量存在, 但是由于实验设计的不同以及缺少饱和水汽压亏缺等环境参数的信息, 很少有实验数据能够满足对气孔导度斜率进行模拟计算的要求(Zhou et al., 2013).以往大多数关于气孔导度斜率的研究都是在温室里的短期试验, 在自然条件下的研究还很少(Medlyn et al., 2011).对于增高的CO2浓度下气孔导度对环境因子的适应性变化的研究结果也不一致.在高CO2浓度的未来环境下, 植物会降低气孔导度和提高光合速率(Long et al., 2004; Bernacchi et al., 2005).有学者在培养箱中升高CO2浓度对大豆(Glycine max)和棉花(Gossypium hirsutum)进行培养研究得出, 气孔导度斜率会因CO2浓度的增加而降低(Harley et al., 1992; Bunce, 2004).但是Leakey等(2006)在FACE (free-air CO2 enrichment)实验样地对大豆进行了生长和生理参数测量, 他们的实验结果却表明气孔导度斜率不会因外界CO2浓度的升高而改变, 并且指出前人所得的气孔导度斜率降低的结论可能是增高的CO2浓度对气孔导度带来的瞬时改变的结果.Medlyn等(2001)也测定了生长在高CO2浓度下6种不同树种的生理参数, 发现CO2浓度升高并没有改变气孔导度斜率.这一结果也在Katul等(2009)的研究中得到证实, Katul等(2009)也发现CO2浓度升高并没有改变Ball-Berry模型中的光合作用与气孔导度的线性关系的斜率. ...
... 大多数地球系统模型或者植被模型都采用经验或者半经验模型来模拟气孔导度(Sellers et al., 1992, 1997; Bonan, 1995; Wang et al., 2013b, 2015).虽然对于经验或半经验模型来说, 可以利用实验数据矫正参数, 但是在现实中, 并非如此.如上文中所提到的, 多数模型都只是区分了C3和C4植物的气孔导度模型参数m值(Krinner et al., 2005; Oleson et al., 2013). De Kauw等(2015)首次将气孔导度机理模型(Medlyn et al., 2011)耦合到陆地表面模型CABLE (Community Atmosphere Biosphere Land Exchange)中, 研究结果显示: 相比于气孔导度经验模型, 耦合气孔导度机理模型导致在常绿针叶林、苔原和C4草原区域, 年蒸腾量减少了30%.Kala等(2014)进一步耦合气孔导度机理模型、CABLE和ACCESS (Australian Community Climate and Earth Systems Simulator)模型, 研究结果显示在北半球的夏季, 新的模型预测的温带森林潜热通量每天减少0.5-1.0 mm, 直接导致每天最高和最低温度升高了1 ℃, 而极端高温升高了1.5 ℃.地表的能量平衡也将进一步影响净初级生产力和陆地碳平衡.由此可见, 深入研究气孔导度斜率的变化规律并提高气孔导度预测的准确性, 可大大减小生态系统模型对碳水通量预测的不确定性, 这对区域尺度生态系统碳水平衡的研究至关重要.然而, 由于对气孔导度斜率变化规律的理解还十分有限, 气孔导度机理模型在植被和地球系统模型中的应用还需要加强. ...
Reconciling the optimal and empirical approaches to modelling stomatal conductance.
2011
a). A comparison of three approaches to modeling leaf gas exchange in annually drought-stressed ponderosa pine forests.
2004
b). Dendroecological analysis of climatic effects on Quercus petraea and Pinus halepensis radial growth using the process-based MAIDEN model.
1
2004
... 以碳积累为代表的光合作用和以水分消耗为代表的蒸腾作用是所有植物功能模型的基础, 准确地模拟光合作用和蒸腾作用也是生态系统模型、植被模型、地球系统模型能够有效模拟碳、水和能量循环的必要条件(Wang et al., 2013b, 2015, 2016).由于以Farquhar等(1980)的光合作用模型为代表的机理模型的广泛应用, 通过对物种生物属性和环境参数(如光合最大羧化速率(Vcmax)、CO2浓度、温度和光照)的测量, 可以有效地模拟光合作用(Wullschleger, 1993; Kattge et al., 2009).相对于光合作用的机理模型, 蒸腾作用通常都是由经验或者半经验模型模拟出的气孔导度值来计算.过去40年里, 气孔导度的模拟工作大量开展.概括来讲, 对气孔导度的模拟有以下几种方式: (1)经验模型.大多通过气孔导度与多个环境因素如空气湿度(Lange et al., 1971)、水汽压亏缺(VPD)(Monteith, 1995)、叶片温度(Jarvis, 1976; Jones, 1992)和臭氧(Uddling et al., 2009)等的统计关系建立模型; (2)半经验模型.基于植物生理特性的假设, 但是仍然和经验模型结合, 如基于气孔导度和光合作用(Ball et al., 1987; Leuning, 1990, 1995; Misson et al., 2004b)、脱落酸(ABA) (Tardieu & Davies, 1993; Guteschick & Simonneau, 2002)、水分传导(Sperry et al., 1998; Oren et al., 1999; Tuzet et al., 2003)和膨压调节(Dewar, 2002; Buckley et al., 2003)等的关系建立的模型, 其参数不具有生物学属性; (3)机理模型.基于植物生理特性的假设, 其参数具有生物学特性, 并具有可以应用在各种环境条件下的潜力(Dewar, 2002; Medlyn et al., 2011; Prentice et al., 2014).很多文献已经综述过气孔导度的经验与半经验模型, 本文将侧重阐述气孔导度机理模型的研究. ...
A reinterpretation of stomatal responses to humidity.
... 经验模型通常都忽略了叶片生理特性和气孔导度之间的关系, 也经常缺少反馈机制, 并且需要对模型参数进行大量的调试, 这些都限制了气孔导度经验模型的应用.Jarvis模型(方程1)是气孔导度经验模型, 最早由Jarvis (1976)提出, 后来经过许多学者的优化改进(Stewart, 1988; White et al., 1999; Macfarlane et al., 2004; Misson et al., 2004a; Noe & Giersch, 2004). ...
A simple dynamic model of photosynthesis in oak leaves: Coupling leaf conductance and photosynthetic carbon fixation by a variable intracellular CO2 pool.
1
2004
... 大多数地球系统模型或者植被模型都采用经验或者半经验模型来模拟气孔导度(Sellers et al., 1992, 1997; Bonan, 1995; Wang et al., 2013b, 2015).虽然对于经验或半经验模型来说, 可以利用实验数据矫正参数, 但是在现实中, 并非如此.如上文中所提到的, 多数模型都只是区分了C3和C4植物的气孔导度模型参数m值(Krinner et al., 2005; Oleson et al., 2013). De Kauw等(2015)首次将气孔导度机理模型(Medlyn et al., 2011)耦合到陆地表面模型CABLE (Community Atmosphere Biosphere Land Exchange)中, 研究结果显示: 相比于气孔导度经验模型, 耦合气孔导度机理模型导致在常绿针叶林、苔原和C4草原区域, 年蒸腾量减少了30%.Kala等(2014)进一步耦合气孔导度机理模型、CABLE和ACCESS (Australian Community Climate and Earth Systems Simulator)模型, 研究结果显示在北半球的夏季, 新的模型预测的温带森林潜热通量每天减少0.5-1.0 mm, 直接导致每天最高和最低温度升高了1 ℃, 而极端高温升高了1.5 ℃.地表的能量平衡也将进一步影响净初级生产力和陆地碳平衡.由此可见, 深入研究气孔导度斜率的变化规律并提高气孔导度预测的准确性, 可大大减小生态系统模型对碳水通量预测的不确定性, 这对区域尺度生态系统碳水平衡的研究至关重要.然而, 由于对气孔导度斜率变化规律的理解还十分有限, 气孔导度机理模型在植被和地球系统模型中的应用还需要加强. ...
Interactions between urbanization, heat stress, and climate change.
3
2013
... 以碳积累为代表的光合作用和以水分消耗为代表的蒸腾作用是所有植物功能模型的基础, 准确地模拟光合作用和蒸腾作用也是生态系统模型、植被模型、地球系统模型能够有效模拟碳、水和能量循环的必要条件(Wang et al., 2013b, 2015, 2016).由于以Farquhar等(1980)的光合作用模型为代表的机理模型的广泛应用, 通过对物种生物属性和环境参数(如光合最大羧化速率(Vcmax)、CO2浓度、温度和光照)的测量, 可以有效地模拟光合作用(Wullschleger, 1993; Kattge et al., 2009).相对于光合作用的机理模型, 蒸腾作用通常都是由经验或者半经验模型模拟出的气孔导度值来计算.过去40年里, 气孔导度的模拟工作大量开展.概括来讲, 对气孔导度的模拟有以下几种方式: (1)经验模型.大多通过气孔导度与多个环境因素如空气湿度(Lange et al., 1971)、水汽压亏缺(VPD)(Monteith, 1995)、叶片温度(Jarvis, 1976; Jones, 1992)和臭氧(Uddling et al., 2009)等的统计关系建立模型; (2)半经验模型.基于植物生理特性的假设, 但是仍然和经验模型结合, 如基于气孔导度和光合作用(Ball et al., 1987; Leuning, 1990, 1995; Misson et al., 2004b)、脱落酸(ABA) (Tardieu & Davies, 1993; Guteschick & Simonneau, 2002)、水分传导(Sperry et al., 1998; Oren et al., 1999; Tuzet et al., 2003)和膨压调节(Dewar, 2002; Buckley et al., 2003)等的关系建立的模型, 其参数不具有生物学属性; (3)机理模型.基于植物生理特性的假设, 其参数具有生物学特性, 并具有可以应用在各种环境条件下的潜力(Dewar, 2002; Medlyn et al., 2011; Prentice et al., 2014).很多文献已经综述过气孔导度的经验与半经验模型, 本文将侧重阐述气孔导度机理模型的研究. ...
... 以往研究显示叶片水势(Oren et al., 1999)、叶片、枝条、根部和整株植物水力传导(Salleo et al., 2000; Addington et al., 2004; Bunce, 2006; Domec et al., 2009)、叶片相对含水量(Ewers et al., 2005)等会影响气孔导度对饱和水汽压亏缺的响应.而由于树龄和树高都会影响水力传导和气孔导度, 进而都会影响气孔导度斜率(Schäfer et al., 2000; Franks, 2004; Ewers et al., 2005; Buckley & Roberts, 2006; Ambrose et al., 2010; Woodruff et al., 2010).木材密度和气孔导度斜率之间也存在着显著的负相关关系(Héroult et al., 2013; Lin et al., 2015).植物结构属性, 如根叶比、根深等以及水力传导结构属性也会影响气孔导度斜率(Litvak et al., 2012; McCulloh & Woodruff, 2012).如与散孔材木相比, 环孔材木的气孔对环境因素变化的反应更加敏感(Bush et al., 2008) , 但是在Litvak等(2012)的研究中并未发现木材结构影响气孔导度斜率的现象.而随着测量高度的增高, 气孔导度对饱和水汽压亏缺响应的敏感性也会降低.叶肉导度和Vcmax也通过影响光合作用影响植物的水分利用效率(Wang et al., 2013a).Flexas等(2013)分析了44种不同物种水分利用效率的规律, 发现水分利用效率和叶肉导度与气孔导度的比值成正比.这个关系表明水分利用效率只有在固碳能力的提高高于水分丢失增加(气孔导度升高)的情况下, 才可以升高.可以预测, 同样的关系也应该在气孔导度斜率及Vcmax和气孔导度的比值间存在.临界氮利用效率也被证明和临界水分利用效率有互补的关系(Palmroth et al., 2013). ...
... 气孔导度斜率的大小与气孔导度和气孔密度有关(Oren et al., 1999; Pataki & Oren, 2003; Ewers et al., 2008).当气孔导度较小时, 一点小小的改变就会给叶温带来极大的影响, 这时的气孔表现极其敏感.最高气孔导度斜率往往出现在0.4以下的低气孔导度条件下, 0.4以上的气孔导度, 响应机制会变得比较平缓(Hetherington & Woodward, 2003). ...
Survey and synthesis of intra-and interspecific variation in stomatal sensitivity to vapour pressure deficit.
1
1999
... 以往研究显示叶片水势(Oren et al., 1999)、叶片、枝条、根部和整株植物水力传导(Salleo et al., 2000; Addington et al., 2004; Bunce, 2006; Domec et al., 2009)、叶片相对含水量(Ewers et al., 2005)等会影响气孔导度对饱和水汽压亏缺的响应.而由于树龄和树高都会影响水力传导和气孔导度, 进而都会影响气孔导度斜率(Schäfer et al., 2000; Franks, 2004; Ewers et al., 2005; Buckley & Roberts, 2006; Ambrose et al., 2010; Woodruff et al., 2010).木材密度和气孔导度斜率之间也存在着显著的负相关关系(Héroult et al., 2013; Lin et al., 2015).植物结构属性, 如根叶比、根深等以及水力传导结构属性也会影响气孔导度斜率(Litvak et al., 2012; McCulloh & Woodruff, 2012).如与散孔材木相比, 环孔材木的气孔对环境因素变化的反应更加敏感(Bush et al., 2008) , 但是在Litvak等(2012)的研究中并未发现木材结构影响气孔导度斜率的现象.而随着测量高度的增高, 气孔导度对饱和水汽压亏缺响应的敏感性也会降低.叶肉导度和Vcmax也通过影响光合作用影响植物的水分利用效率(Wang et al., 2013a).Flexas等(2013)分析了44种不同物种水分利用效率的规律, 发现水分利用效率和叶肉导度与气孔导度的比值成正比.这个关系表明水分利用效率只有在固碳能力的提高高于水分丢失增加(气孔导度升高)的情况下, 才可以升高.可以预测, 同样的关系也应该在气孔导度斜率及Vcmax和气孔导度的比值间存在.临界氮利用效率也被证明和临界水分利用效率有互补的关系(Palmroth et al., 2013). ...
On the complementary relationship between marginal nitrogen and water-use efficiencies among Pinus taeda leaves grown under ambient and CO2-enriched environments.
1
2013
... 气孔导度斜率的大小与气孔导度和气孔密度有关(Oren et al., 1999; Pataki & Oren, 2003; Ewers et al., 2008).当气孔导度较小时, 一点小小的改变就会给叶温带来极大的影响, 这时的气孔表现极其敏感.最高气孔导度斜率往往出现在0.4以下的低气孔导度条件下, 0.4以上的气孔导度, 响应机制会变得比较平缓(Hetherington & Woodward, 2003). ...
Species differences in stomatal control of water loss at the canopy scale in a mature bottomland deciduous forest.
2
2003
... 以碳积累为代表的光合作用和以水分消耗为代表的蒸腾作用是所有植物功能模型的基础, 准确地模拟光合作用和蒸腾作用也是生态系统模型、植被模型、地球系统模型能够有效模拟碳、水和能量循环的必要条件(Wang et al., 2013b, 2015, 2016).由于以Farquhar等(1980)的光合作用模型为代表的机理模型的广泛应用, 通过对物种生物属性和环境参数(如光合最大羧化速率(Vcmax)、CO2浓度、温度和光照)的测量, 可以有效地模拟光合作用(Wullschleger, 1993; Kattge et al., 2009).相对于光合作用的机理模型, 蒸腾作用通常都是由经验或者半经验模型模拟出的气孔导度值来计算.过去40年里, 气孔导度的模拟工作大量开展.概括来讲, 对气孔导度的模拟有以下几种方式: (1)经验模型.大多通过气孔导度与多个环境因素如空气湿度(Lange et al., 1971)、水汽压亏缺(VPD)(Monteith, 1995)、叶片温度(Jarvis, 1976; Jones, 1992)和臭氧(Uddling et al., 2009)等的统计关系建立模型; (2)半经验模型.基于植物生理特性的假设, 但是仍然和经验模型结合, 如基于气孔导度和光合作用(Ball et al., 1987; Leuning, 1990, 1995; Misson et al., 2004b)、脱落酸(ABA) (Tardieu & Davies, 1993; Guteschick & Simonneau, 2002)、水分传导(Sperry et al., 1998; Oren et al., 1999; Tuzet et al., 2003)和膨压调节(Dewar, 2002; Buckley et al., 2003)等的关系建立的模型, 其参数不具有生物学属性; (3)机理模型.基于植物生理特性的假设, 其参数具有生物学特性, 并具有可以应用在各种环境条件下的潜力(Dewar, 2002; Medlyn et al., 2011; Prentice et al., 2014).很多文献已经综述过气孔导度的经验与半经验模型, 本文将侧重阐述气孔导度机理模型的研究. ...
... 对于外界环境的变化, 气孔导度斜率也会做出不同的响应.不同植物物种的水分利用策略不同, 在同样的湿度环境下气孔导度斜率会有差异, 同样的物种在不同水分环境下气孔导度斜率也会不同(Zhou et al., 2013).Lin等(2015)对超过1万条光合测量数据综合分析后发现, 在植物生长适宜的外界温湿度下, 随着温湿度升高, 气孔导度斜率都会增加.对于气孔导度斜率随温度升高而增加的原因, 有两种解释: 一是气孔导度斜率与CO2补偿点浓度相关(Medlyn et al., 2011), 而这一浓度会随温度升高而增加(Bernacchi et al., 2001); 二是水分子黏性会随温度升高而降低, 这就降低了运输水分的消耗, 从而增高了气孔导度斜率(Prentice et al., 2014).Héroult等(2013)报道了产地不同的4种桉属植物在相同环境条件下, 气孔导度斜率随着自然产地不同而不同, 自然生长在湿润地区的植物气孔导度斜率比在半湿润地区的植物高.在湿润环境生长的长叶松(Pinus palustris)气孔关闭对增加的饱和水汽压亏缺的反应比在干旱环境下更加敏感(Addingtong et al., 2006). ...
Balancing the costs of carbon gain and water transport: Testing a new theoretical framework for plant functional ecology.
1
2014
... 在很多生态系统模型中, 土壤干旱胁迫对气孔导度影响的模拟通常是通过改变水分胁迫与光合作用/气孔导度之间的作用关系来实现的(Sala & Tenhunen, 1996; Verhoef & Allen, 2000).但也有模型是通过改变水分胁迫与光合作用或者光合作用参数Vcmax和最大电子传输速率(Jmax)的关系, 模拟水分胁迫下光合作用的改变进而模拟气孔导度的改变(Sellers et al., 1996; Ronda et al., 2001; Best et al., 2011).虽然在轻度干旱情况下, 上述几种做法可以模拟出水分胁迫对叶片尺度碳水通量的影响, 但在严重干旱情况下无法同时准确模拟光合作用和蒸腾作用(Verhoef & Allen, 2000; Reichstein et al., 2002; Keenan et al., 2009).Egea等(2011)提议, 在干旱条件下可以通过先限制叶肉导度, 进而限制气孔导度和光合作用参数的方案来提高光合作用和气孔导度模拟的准确性.Zhou等(2013)的研究结果也表明模拟干旱状况下的气孔导度, 必须同时考虑干旱对气孔和非气孔因素(如Vcmax)的综合影响.由于对干旱状况下气孔导度预测的不准确性, 现有的模型一般都倾向于高估光合作用和蒸腾作用对干旱的响应(Dietze, 2014).与以前从气孔(物理过程)或者非气孔(生化过程)的角度出发研究干旱对光合作用和气孔导度的影响不同, 气孔导度机理模型中的气孔导度斜率在干旱环境下的变化规律包含了干旱对气孔(生物物理因素)和非气孔因素, 包括最大光合效率和临界水分利用效率的综合影响, 为研究全球变化下陆地生态系统碳水耦合的适应性提供了一个简明且综合的框架.Manzoni等(2011)计算了50种植物的临界水分利用效率, 发现在忽略表皮水分丢失的情况下, 植物临界水分利用效率在轻微干旱的情况下增高, 而在严重干旱的情况下下降; 如果不忽略表皮水分丢失, 植物临界水分利用效率会随着水分胁迫的增加而提高; 而不同的植物功能型和不同的气候条件下, 反应也会不同.总体说来, 土壤水分较少时, 植物的水分利用效率会增高, 此时气孔导度斜率值也会降低(Lin et al., 2015).在环境水分条件与气孔导度斜率的变化规律之间还有很多值得讨论的地方, 我国学者对于水分利用的研究主要集中在单叶水分利用效率(付士磊等, 2006; 杨建伟等, 2006; 全先奎和王传宽, 2015)、气孔导度时空变异的调查(司建华等, 2008)、气孔导度对水分环境的响应(鱼腾飞等, 2012; 袁国富等, 2012)、单叶或者幼苗蒸腾速率(宋炳煜等, 2004; 岳广阳等, 2009)、树干液流密度(赵平等, 2006)、土壤水分定位监测(尹春梅等, 2008), 以及涡度协相关法测量生态系统水分利用效率(Zhou et al., 2013)等方面.土壤干旱如何影响不同功能型和物种的气孔导度斜率和临界水分利用效率, 还有待于进一步的研究. ...
帽儿山17个种源落叶松针叶的水分利用效率比较
1
2015
... 在很多生态系统模型中, 土壤干旱胁迫对气孔导度影响的模拟通常是通过改变水分胁迫与光合作用/气孔导度之间的作用关系来实现的(Sala & Tenhunen, 1996; Verhoef & Allen, 2000).但也有模型是通过改变水分胁迫与光合作用或者光合作用参数Vcmax和最大电子传输速率(Jmax)的关系, 模拟水分胁迫下光合作用的改变进而模拟气孔导度的改变(Sellers et al., 1996; Ronda et al., 2001; Best et al., 2011).虽然在轻度干旱情况下, 上述几种做法可以模拟出水分胁迫对叶片尺度碳水通量的影响, 但在严重干旱情况下无法同时准确模拟光合作用和蒸腾作用(Verhoef & Allen, 2000; Reichstein et al., 2002; Keenan et al., 2009).Egea等(2011)提议, 在干旱条件下可以通过先限制叶肉导度, 进而限制气孔导度和光合作用参数的方案来提高光合作用和气孔导度模拟的准确性.Zhou等(2013)的研究结果也表明模拟干旱状况下的气孔导度, 必须同时考虑干旱对气孔和非气孔因素(如Vcmax)的综合影响.由于对干旱状况下气孔导度预测的不准确性, 现有的模型一般都倾向于高估光合作用和蒸腾作用对干旱的响应(Dietze, 2014).与以前从气孔(物理过程)或者非气孔(生化过程)的角度出发研究干旱对光合作用和气孔导度的影响不同, 气孔导度机理模型中的气孔导度斜率在干旱环境下的变化规律包含了干旱对气孔(生物物理因素)和非气孔因素, 包括最大光合效率和临界水分利用效率的综合影响, 为研究全球变化下陆地生态系统碳水耦合的适应性提供了一个简明且综合的框架.Manzoni等(2011)计算了50种植物的临界水分利用效率, 发现在忽略表皮水分丢失的情况下, 植物临界水分利用效率在轻微干旱的情况下增高, 而在严重干旱的情况下下降; 如果不忽略表皮水分丢失, 植物临界水分利用效率会随着水分胁迫的增加而提高; 而不同的植物功能型和不同的气候条件下, 反应也会不同.总体说来, 土壤水分较少时, 植物的水分利用效率会增高, 此时气孔导度斜率值也会降低(Lin et al., 2015).在环境水分条件与气孔导度斜率的变化规律之间还有很多值得讨论的地方, 我国学者对于水分利用的研究主要集中在单叶水分利用效率(付士磊等, 2006; 杨建伟等, 2006; 全先奎和王传宽, 2015)、气孔导度时空变异的调查(司建华等, 2008)、气孔导度对水分环境的响应(鱼腾飞等, 2012; 袁国富等, 2012)、单叶或者幼苗蒸腾速率(宋炳煜等, 2004; 岳广阳等, 2009)、树干液流密度(赵平等, 2006)、土壤水分定位监测(尹春梅等, 2008), 以及涡度协相关法测量生态系统水分利用效率(Zhou et al., 2013)等方面.土壤干旱如何影响不同功能型和物种的气孔导度斜率和临界水分利用效率, 还有待于进一步的研究. ...
Severe drought effects on ecosystem CO2 and H2O fluxes at three Mediterranean evergreen sites: Revision of current hypotheses?
1
2002
... 在很多生态系统模型中, 土壤干旱胁迫对气孔导度影响的模拟通常是通过改变水分胁迫与光合作用/气孔导度之间的作用关系来实现的(Sala & Tenhunen, 1996; Verhoef & Allen, 2000).但也有模型是通过改变水分胁迫与光合作用或者光合作用参数Vcmax和最大电子传输速率(Jmax)的关系, 模拟水分胁迫下光合作用的改变进而模拟气孔导度的改变(Sellers et al., 1996; Ronda et al., 2001; Best et al., 2011).虽然在轻度干旱情况下, 上述几种做法可以模拟出水分胁迫对叶片尺度碳水通量的影响, 但在严重干旱情况下无法同时准确模拟光合作用和蒸腾作用(Verhoef & Allen, 2000; Reichstein et al., 2002; Keenan et al., 2009).Egea等(2011)提议, 在干旱条件下可以通过先限制叶肉导度, 进而限制气孔导度和光合作用参数的方案来提高光合作用和气孔导度模拟的准确性.Zhou等(2013)的研究结果也表明模拟干旱状况下的气孔导度, 必须同时考虑干旱对气孔和非气孔因素(如Vcmax)的综合影响.由于对干旱状况下气孔导度预测的不准确性, 现有的模型一般都倾向于高估光合作用和蒸腾作用对干旱的响应(Dietze, 2014).与以前从气孔(物理过程)或者非气孔(生化过程)的角度出发研究干旱对光合作用和气孔导度的影响不同, 气孔导度机理模型中的气孔导度斜率在干旱环境下的变化规律包含了干旱对气孔(生物物理因素)和非气孔因素, 包括最大光合效率和临界水分利用效率的综合影响, 为研究全球变化下陆地生态系统碳水耦合的适应性提供了一个简明且综合的框架.Manzoni等(2011)计算了50种植物的临界水分利用效率, 发现在忽略表皮水分丢失的情况下, 植物临界水分利用效率在轻微干旱的情况下增高, 而在严重干旱的情况下下降; 如果不忽略表皮水分丢失, 植物临界水分利用效率会随着水分胁迫的增加而提高; 而不同的植物功能型和不同的气候条件下, 反应也会不同.总体说来, 土壤水分较少时, 植物的水分利用效率会增高, 此时气孔导度斜率值也会降低(Lin et al., 2015).在环境水分条件与气孔导度斜率的变化规律之间还有很多值得讨论的地方, 我国学者对于水分利用的研究主要集中在单叶水分利用效率(付士磊等, 2006; 杨建伟等, 2006; 全先奎和王传宽, 2015)、气孔导度时空变异的调查(司建华等, 2008)、气孔导度对水分环境的响应(鱼腾飞等, 2012; 袁国富等, 2012)、单叶或者幼苗蒸腾速率(宋炳煜等, 2004; 岳广阳等, 2009)、树干液流密度(赵平等, 2006)、土壤水分定位监测(尹春梅等, 2008), 以及涡度协相关法测量生态系统水分利用效率(Zhou et al., 2013)等方面.土壤干旱如何影响不同功能型和物种的气孔导度斜率和临界水分利用效率, 还有待于进一步的研究. ...
Representation of the canopy conductance in modeling the surface energy budget for low vegetation.
1
2001
... 在很多生态系统模型中, 土壤干旱胁迫对气孔导度影响的模拟通常是通过改变水分胁迫与光合作用/气孔导度之间的作用关系来实现的(Sala & Tenhunen, 1996; Verhoef & Allen, 2000).但也有模型是通过改变水分胁迫与光合作用或者光合作用参数Vcmax和最大电子传输速率(Jmax)的关系, 模拟水分胁迫下光合作用的改变进而模拟气孔导度的改变(Sellers et al., 1996; Ronda et al., 2001; Best et al., 2011).虽然在轻度干旱情况下, 上述几种做法可以模拟出水分胁迫对叶片尺度碳水通量的影响, 但在严重干旱情况下无法同时准确模拟光合作用和蒸腾作用(Verhoef & Allen, 2000; Reichstein et al., 2002; Keenan et al., 2009).Egea等(2011)提议, 在干旱条件下可以通过先限制叶肉导度, 进而限制气孔导度和光合作用参数的方案来提高光合作用和气孔导度模拟的准确性.Zhou等(2013)的研究结果也表明模拟干旱状况下的气孔导度, 必须同时考虑干旱对气孔和非气孔因素(如Vcmax)的综合影响.由于对干旱状况下气孔导度预测的不准确性, 现有的模型一般都倾向于高估光合作用和蒸腾作用对干旱的响应(Dietze, 2014).与以前从气孔(物理过程)或者非气孔(生化过程)的角度出发研究干旱对光合作用和气孔导度的影响不同, 气孔导度机理模型中的气孔导度斜率在干旱环境下的变化规律包含了干旱对气孔(生物物理因素)和非气孔因素, 包括最大光合效率和临界水分利用效率的综合影响, 为研究全球变化下陆地生态系统碳水耦合的适应性提供了一个简明且综合的框架.Manzoni等(2011)计算了50种植物的临界水分利用效率, 发现在忽略表皮水分丢失的情况下, 植物临界水分利用效率在轻微干旱的情况下增高, 而在严重干旱的情况下下降; 如果不忽略表皮水分丢失, 植物临界水分利用效率会随着水分胁迫的增加而提高; 而不同的植物功能型和不同的气候条件下, 反应也会不同.总体说来, 土壤水分较少时, 植物的水分利用效率会增高, 此时气孔导度斜率值也会降低(Lin et al., 2015).在环境水分条件与气孔导度斜率的变化规律之间还有很多值得讨论的地方, 我国学者对于水分利用的研究主要集中在单叶水分利用效率(付士磊等, 2006; 杨建伟等, 2006; 全先奎和王传宽, 2015)、气孔导度时空变异的调查(司建华等, 2008)、气孔导度对水分环境的响应(鱼腾飞等, 2012; 袁国富等, 2012)、单叶或者幼苗蒸腾速率(宋炳煜等, 2004; 岳广阳等, 2009)、树干液流密度(赵平等, 2006)、土壤水分定位监测(尹春梅等, 2008), 以及涡度协相关法测量生态系统水分利用效率(Zhou et al., 2013)等方面.土壤干旱如何影响不同功能型和物种的气孔导度斜率和临界水分利用效率, 还有待于进一步的研究. ...
Simulations of canopy net photosynthesis and transpiration in Quercus ilex L. under the influence of seasonal drought.
1
1996
... 以往研究显示叶片水势(Oren et al., 1999)、叶片、枝条、根部和整株植物水力传导(Salleo et al., 2000; Addington et al., 2004; Bunce, 2006; Domec et al., 2009)、叶片相对含水量(Ewers et al., 2005)等会影响气孔导度对饱和水汽压亏缺的响应.而由于树龄和树高都会影响水力传导和气孔导度, 进而都会影响气孔导度斜率(Schäfer et al., 2000; Franks, 2004; Ewers et al., 2005; Buckley & Roberts, 2006; Ambrose et al., 2010; Woodruff et al., 2010).木材密度和气孔导度斜率之间也存在着显著的负相关关系(Héroult et al., 2013; Lin et al., 2015).植物结构属性, 如根叶比、根深等以及水力传导结构属性也会影响气孔导度斜率(Litvak et al., 2012; McCulloh & Woodruff, 2012).如与散孔材木相比, 环孔材木的气孔对环境因素变化的反应更加敏感(Bush et al., 2008) , 但是在Litvak等(2012)的研究中并未发现木材结构影响气孔导度斜率的现象.而随着测量高度的增高, 气孔导度对饱和水汽压亏缺响应的敏感性也会降低.叶肉导度和Vcmax也通过影响光合作用影响植物的水分利用效率(Wang et al., 2013a).Flexas等(2013)分析了44种不同物种水分利用效率的规律, 发现水分利用效率和叶肉导度与气孔导度的比值成正比.这个关系表明水分利用效率只有在固碳能力的提高高于水分丢失增加(气孔导度升高)的情况下, 才可以升高.可以预测, 同样的关系也应该在气孔导度斜率及Vcmax和气孔导度的比值间存在.临界氮利用效率也被证明和临界水分利用效率有互补的关系(Palmroth et al., 2013). ...
Xylem cavitation and hydraulic control of stomatal conductance in laurel (Laurus nobilis L.).
2000
Stomatal movement: Its regulation and regulatory role: A review.
1
1927
... 以往研究显示叶片水势(Oren et al., 1999)、叶片、枝条、根部和整株植物水力传导(Salleo et al., 2000; Addington et al., 2004; Bunce, 2006; Domec et al., 2009)、叶片相对含水量(Ewers et al., 2005)等会影响气孔导度对饱和水汽压亏缺的响应.而由于树龄和树高都会影响水力传导和气孔导度, 进而都会影响气孔导度斜率(Schäfer et al., 2000; Franks, 2004; Ewers et al., 2005; Buckley & Roberts, 2006; Ambrose et al., 2010; Woodruff et al., 2010).木材密度和气孔导度斜率之间也存在着显著的负相关关系(Héroult et al., 2013; Lin et al., 2015).植物结构属性, 如根叶比、根深等以及水力传导结构属性也会影响气孔导度斜率(Litvak et al., 2012; McCulloh & Woodruff, 2012).如与散孔材木相比, 环孔材木的气孔对环境因素变化的反应更加敏感(Bush et al., 2008) , 但是在Litvak等(2012)的研究中并未发现木材结构影响气孔导度斜率的现象.而随着测量高度的增高, 气孔导度对饱和水汽压亏缺响应的敏感性也会降低.叶肉导度和Vcmax也通过影响光合作用影响植物的水分利用效率(Wang et al., 2013a).Flexas等(2013)分析了44种不同物种水分利用效率的规律, 发现水分利用效率和叶肉导度与气孔导度的比值成正比.这个关系表明水分利用效率只有在固碳能力的提高高于水分丢失增加(气孔导度升高)的情况下, 才可以升高.可以预测, 同样的关系也应该在气孔导度斜率及Vcmax和气孔导度的比值间存在.临界氮利用效率也被证明和临界水分利用效率有互补的关系(Palmroth et al., 2013). ...
The effect of tree height on crown level stomatal conductance.
2
2000
... 此后多人对Ball-Berry模型进行了优化、改进和应用, 如加入叶片水势的作用等(Sellers et al., 1992; Kim & Lieth, 2003; Tuzet et al., 2003; Yu et al., 2004), 但其本质上依然是半经验模型.在对Ball-Berry模型的众多改进中, Yu等(2004)的模型较为突出.此模型结合了Jarvis模型和Ball-Berry模型的优点, 采用了更便于计算的模型参数, 基于气孔与光合作用的联系及光合作用与光照之间的联系, 推导出了双曲线式的气孔导度模型, 使得直接从气象参数计算气孔导度成为可能.模型简洁、实用, 对于实测数据符合较好. ...
... 大多数地球系统模型或者植被模型都采用经验或者半经验模型来模拟气孔导度(Sellers et al., 1992, 1997; Bonan, 1995; Wang et al., 2013b, 2015).虽然对于经验或半经验模型来说, 可以利用实验数据矫正参数, 但是在现实中, 并非如此.如上文中所提到的, 多数模型都只是区分了C3和C4植物的气孔导度模型参数m值(Krinner et al., 2005; Oleson et al., 2013). De Kauw等(2015)首次将气孔导度机理模型(Medlyn et al., 2011)耦合到陆地表面模型CABLE (Community Atmosphere Biosphere Land Exchange)中, 研究结果显示: 相比于气孔导度经验模型, 耦合气孔导度机理模型导致在常绿针叶林、苔原和C4草原区域, 年蒸腾量减少了30%.Kala等(2014)进一步耦合气孔导度机理模型、CABLE和ACCESS (Australian Community Climate and Earth Systems Simulator)模型, 研究结果显示在北半球的夏季, 新的模型预测的温带森林潜热通量每天减少0.5-1.0 mm, 直接导致每天最高和最低温度升高了1 ℃, 而极端高温升高了1.5 ℃.地表的能量平衡也将进一步影响净初级生产力和陆地碳平衡.由此可见, 深入研究气孔导度斜率的变化规律并提高气孔导度预测的准确性, 可大大减小生态系统模型对碳水通量预测的不确定性, 这对区域尺度生态系统碳水平衡的研究至关重要.然而, 由于对气孔导度斜率变化规律的理解还十分有限, 气孔导度机理模型在植被和地球系统模型中的应用还需要加强. ...
Canopy reflectance, photosynthesis, and transpiration. III. A reanalysis using improved leaf models and a new canopy integration scheme.
2
1992
... Ball-Berry模型的特点是可以把复杂的环境因子影响综合到光合作用、湿度和CO2浓度3个要素中.它简化了气孔导度与环境因子的关系, 但是它把光合作用速率作为模型的输入参数处理, 因而无法应用这个模型直接从环境因子计算出气孔导度.Collatz等(1991)耦合了该气孔模型与光合作用生化模型, 建立联立方程组, 以便同时解出光合速率和气孔导度.Ball-Berry气孔导度模型同Farquhar等(1980)光合作用模型结合(FvCB模型), 可以有效地模拟叶片尺度光合作用和气孔导度.FvCB模型被广泛应用到大量陆地生态系统模型中, 并结合气候模式来预测全球变化对植被和大气之间碳、水循环的影响(Sellers et al., 1997; Damour et al., 2010).因此这两个模型的准确性, 将直接影响大量生态系统模型、区域模型乃至地球系统模型预测的可靠性. ...
... 大多数地球系统模型或者植被模型都采用经验或者半经验模型来模拟气孔导度(Sellers et al., 1992, 1997; Bonan, 1995; Wang et al., 2013b, 2015).虽然对于经验或半经验模型来说, 可以利用实验数据矫正参数, 但是在现实中, 并非如此.如上文中所提到的, 多数模型都只是区分了C3和C4植物的气孔导度模型参数m值(Krinner et al., 2005; Oleson et al., 2013). De Kauw等(2015)首次将气孔导度机理模型(Medlyn et al., 2011)耦合到陆地表面模型CABLE (Community Atmosphere Biosphere Land Exchange)中, 研究结果显示: 相比于气孔导度经验模型, 耦合气孔导度机理模型导致在常绿针叶林、苔原和C4草原区域, 年蒸腾量减少了30%.Kala等(2014)进一步耦合气孔导度机理模型、CABLE和ACCESS (Australian Community Climate and Earth Systems Simulator)模型, 研究结果显示在北半球的夏季, 新的模型预测的温带森林潜热通量每天减少0.5-1.0 mm, 直接导致每天最高和最低温度升高了1 ℃, 而极端高温升高了1.5 ℃.地表的能量平衡也将进一步影响净初级生产力和陆地碳平衡.由此可见, 深入研究气孔导度斜率的变化规律并提高气孔导度预测的准确性, 可大大减小生态系统模型对碳水通量预测的不确定性, 这对区域尺度生态系统碳水平衡的研究至关重要.然而, 由于对气孔导度斜率变化规律的理解还十分有限, 气孔导度机理模型在植被和地球系统模型中的应用还需要加强. ...
Modeling the exchanges of energy, water, and carbon between continents and the atmosphere.
1
1997
... 在很多生态系统模型中, 土壤干旱胁迫对气孔导度影响的模拟通常是通过改变水分胁迫与光合作用/气孔导度之间的作用关系来实现的(Sala & Tenhunen, 1996; Verhoef & Allen, 2000).但也有模型是通过改变水分胁迫与光合作用或者光合作用参数Vcmax和最大电子传输速率(Jmax)的关系, 模拟水分胁迫下光合作用的改变进而模拟气孔导度的改变(Sellers et al., 1996; Ronda et al., 2001; Best et al., 2011).虽然在轻度干旱情况下, 上述几种做法可以模拟出水分胁迫对叶片尺度碳水通量的影响, 但在严重干旱情况下无法同时准确模拟光合作用和蒸腾作用(Verhoef & Allen, 2000; Reichstein et al., 2002; Keenan et al., 2009).Egea等(2011)提议, 在干旱条件下可以通过先限制叶肉导度, 进而限制气孔导度和光合作用参数的方案来提高光合作用和气孔导度模拟的准确性.Zhou等(2013)的研究结果也表明模拟干旱状况下的气孔导度, 必须同时考虑干旱对气孔和非气孔因素(如Vcmax)的综合影响.由于对干旱状况下气孔导度预测的不准确性, 现有的模型一般都倾向于高估光合作用和蒸腾作用对干旱的响应(Dietze, 2014).与以前从气孔(物理过程)或者非气孔(生化过程)的角度出发研究干旱对光合作用和气孔导度的影响不同, 气孔导度机理模型中的气孔导度斜率在干旱环境下的变化规律包含了干旱对气孔(生物物理因素)和非气孔因素, 包括最大光合效率和临界水分利用效率的综合影响, 为研究全球变化下陆地生态系统碳水耦合的适应性提供了一个简明且综合的框架.Manzoni等(2011)计算了50种植物的临界水分利用效率, 发现在忽略表皮水分丢失的情况下, 植物临界水分利用效率在轻微干旱的情况下增高, 而在严重干旱的情况下下降; 如果不忽略表皮水分丢失, 植物临界水分利用效率会随着水分胁迫的增加而提高; 而不同的植物功能型和不同的气候条件下, 反应也会不同.总体说来, 土壤水分较少时, 植物的水分利用效率会增高, 此时气孔导度斜率值也会降低(Lin et al., 2015).在环境水分条件与气孔导度斜率的变化规律之间还有很多值得讨论的地方, 我国学者对于水分利用的研究主要集中在单叶水分利用效率(付士磊等, 2006; 杨建伟等, 2006; 全先奎和王传宽, 2015)、气孔导度时空变异的调查(司建华等, 2008)、气孔导度对水分环境的响应(鱼腾飞等, 2012; 袁国富等, 2012)、单叶或者幼苗蒸腾速率(宋炳煜等, 2004; 岳广阳等, 2009)、树干液流密度(赵平等, 2006)、土壤水分定位监测(尹春梅等, 2008), 以及涡度协相关法测量生态系统水分利用效率(Zhou et al., 2013)等方面.土壤干旱如何影响不同功能型和物种的气孔导度斜率和临界水分利用效率, 还有待于进一步的研究. ...
A revised land surface parameterization (SiB2) for atmospheric GCMs. Part I: Model formulation.
1
1996
... 在很多生态系统模型中, 土壤干旱胁迫对气孔导度影响的模拟通常是通过改变水分胁迫与光合作用/气孔导度之间的作用关系来实现的(Sala & Tenhunen, 1996; Verhoef & Allen, 2000).但也有模型是通过改变水分胁迫与光合作用或者光合作用参数Vcmax和最大电子传输速率(Jmax)的关系, 模拟水分胁迫下光合作用的改变进而模拟气孔导度的改变(Sellers et al., 1996; Ronda et al., 2001; Best et al., 2011).虽然在轻度干旱情况下, 上述几种做法可以模拟出水分胁迫对叶片尺度碳水通量的影响, 但在严重干旱情况下无法同时准确模拟光合作用和蒸腾作用(Verhoef & Allen, 2000; Reichstein et al., 2002; Keenan et al., 2009).Egea等(2011)提议, 在干旱条件下可以通过先限制叶肉导度, 进而限制气孔导度和光合作用参数的方案来提高光合作用和气孔导度模拟的准确性.Zhou等(2013)的研究结果也表明模拟干旱状况下的气孔导度, 必须同时考虑干旱对气孔和非气孔因素(如Vcmax)的综合影响.由于对干旱状况下气孔导度预测的不准确性, 现有的模型一般都倾向于高估光合作用和蒸腾作用对干旱的响应(Dietze, 2014).与以前从气孔(物理过程)或者非气孔(生化过程)的角度出发研究干旱对光合作用和气孔导度的影响不同, 气孔导度机理模型中的气孔导度斜率在干旱环境下的变化规律包含了干旱对气孔(生物物理因素)和非气孔因素, 包括最大光合效率和临界水分利用效率的综合影响, 为研究全球变化下陆地生态系统碳水耦合的适应性提供了一个简明且综合的框架.Manzoni等(2011)计算了50种植物的临界水分利用效率, 发现在忽略表皮水分丢失的情况下, 植物临界水分利用效率在轻微干旱的情况下增高, 而在严重干旱的情况下下降; 如果不忽略表皮水分丢失, 植物临界水分利用效率会随着水分胁迫的增加而提高; 而不同的植物功能型和不同的气候条件下, 反应也会不同.总体说来, 土壤水分较少时, 植物的水分利用效率会增高, 此时气孔导度斜率值也会降低(Lin et al., 2015).在环境水分条件与气孔导度斜率的变化规律之间还有很多值得讨论的地方, 我国学者对于水分利用的研究主要集中在单叶水分利用效率(付士磊等, 2006; 杨建伟等, 2006; 全先奎和王传宽, 2015)、气孔导度时空变异的调查(司建华等, 2008)、气孔导度对水分环境的响应(鱼腾飞等, 2012; 袁国富等, 2012)、单叶或者幼苗蒸腾速率(宋炳煜等, 2004; 岳广阳等, 2009)、树干液流密度(赵平等, 2006)、土壤水分定位监测(尹春梅等, 2008), 以及涡度协相关法测量生态系统水分利用效率(Zhou et al., 2013)等方面.土壤干旱如何影响不同功能型和物种的气孔导度斜率和临界水分利用效率, 还有待于进一步的研究. ...
A SVAT scheme describing energy and CO2 fluxes for multi-component vegetation: Calibration and test for a Sahelian savannah.
2000
Analyzing the impact of climate and management factors on the productivity and soil carbon sequestration of poplar plantations.
3
2016
... 以碳积累为代表的光合作用和以水分消耗为代表的蒸腾作用是所有植物功能模型的基础, 准确地模拟光合作用和蒸腾作用也是生态系统模型、植被模型、地球系统模型能够有效模拟碳、水和能量循环的必要条件(Wang et al., 2013b, 2015, 2016).由于以Farquhar等(1980)的光合作用模型为代表的机理模型的广泛应用, 通过对物种生物属性和环境参数(如光合最大羧化速率(Vcmax)、CO2浓度、温度和光照)的测量, 可以有效地模拟光合作用(Wullschleger, 1993; Kattge et al., 2009).相对于光合作用的机理模型, 蒸腾作用通常都是由经验或者半经验模型模拟出的气孔导度值来计算.过去40年里, 气孔导度的模拟工作大量开展.概括来讲, 对气孔导度的模拟有以下几种方式: (1)经验模型.大多通过气孔导度与多个环境因素如空气湿度(Lange et al., 1971)、水汽压亏缺(VPD)(Monteith, 1995)、叶片温度(Jarvis, 1976; Jones, 1992)和臭氧(Uddling et al., 2009)等的统计关系建立模型; (2)半经验模型.基于植物生理特性的假设, 但是仍然和经验模型结合, 如基于气孔导度和光合作用(Ball et al., 1987; Leuning, 1990, 1995; Misson et al., 2004b)、脱落酸(ABA) (Tardieu & Davies, 1993; Guteschick & Simonneau, 2002)、水分传导(Sperry et al., 1998; Oren et al., 1999; Tuzet et al., 2003)和膨压调节(Dewar, 2002; Buckley et al., 2003)等的关系建立的模型, 其参数不具有生物学属性; (3)机理模型.基于植物生理特性的假设, 其参数具有生物学特性, 并具有可以应用在各种环境条件下的潜力(Dewar, 2002; Medlyn et al., 2011; Prentice et al., 2014).很多文献已经综述过气孔导度的经验与半经验模型, 本文将侧重阐述气孔导度机理模型的研究. ...
... 目前盛行的生态系统模型多是通过减小m值来模拟干旱对气孔导度和光合作用的影响(Wang et al., 2013b, 2015); 或者通过减小光合作用模型中的Vcmax来间接地减小气孔导度和光合作用来实现干旱条件下的生长模拟(Krinner et al., 2005).没有叶片尺度上气孔导度模拟的准确性, 陆地生态系统模型将无法准确地预测冠层、生态系统乃至区域尺度上碳、水循环的变化(Damour et al., 2010).成功模拟气孔行为的机理模型将为理解碳水耦合的适应过程以及陆地植被模型对植被结构、功能和演替的准确把握提供一个有效的框架. ...
... 大多数地球系统模型或者植被模型都采用经验或者半经验模型来模拟气孔导度(Sellers et al., 1992, 1997; Bonan, 1995; Wang et al., 2013b, 2015).虽然对于经验或半经验模型来说, 可以利用实验数据矫正参数, 但是在现实中, 并非如此.如上文中所提到的, 多数模型都只是区分了C3和C4植物的气孔导度模型参数m值(Krinner et al., 2005; Oleson et al., 2013). De Kauw等(2015)首次将气孔导度机理模型(Medlyn et al., 2011)耦合到陆地表面模型CABLE (Community Atmosphere Biosphere Land Exchange)中, 研究结果显示: 相比于气孔导度经验模型, 耦合气孔导度机理模型导致在常绿针叶林、苔原和C4草原区域, 年蒸腾量减少了30%.Kala等(2014)进一步耦合气孔导度机理模型、CABLE和ACCESS (Australian Community Climate and Earth Systems Simulator)模型, 研究结果显示在北半球的夏季, 新的模型预测的温带森林潜热通量每天减少0.5-1.0 mm, 直接导致每天最高和最低温度升高了1 ℃, 而极端高温升高了1.5 ℃.地表的能量平衡也将进一步影响净初级生产力和陆地碳平衡.由此可见, 深入研究气孔导度斜率的变化规律并提高气孔导度预测的准确性, 可大大减小生态系统模型对碳水通量预测的不确定性, 这对区域尺度生态系统碳水平衡的研究至关重要.然而, 由于对气孔导度斜率变化规律的理解还十分有限, 气孔导度机理模型在植被和地球系统模型中的应用还需要加强. ...
A physiological and biophysical model of coppice willow (Salix spp.) production yields for the contiguous USA in current and future climate scenarios.
2015
a). Predicting yields of short-rotation hybrid poplar (Populus spp.) for the contiguous US through model-data synthesis.
2013
b). Ecophysiological screening of tree species for biomass production: Trade-off between production and water use.
2013
Increasing water use efficiency along the C3 to C4 evolutionary pathway: A stomatal optimization perspective.
1
2014
... 经验模型通常都忽略了叶片生理特性和气孔导度之间的关系, 也经常缺少反馈机制, 并且需要对模型参数进行大量的调试, 这些都限制了气孔导度经验模型的应用.Jarvis模型(方程1)是气孔导度经验模型, 最早由Jarvis (1976)提出, 后来经过许多学者的优化改进(Stewart, 1988; White et al., 1999; Macfarlane et al., 2004; Misson et al., 2004a; Noe & Giersch, 2004). ...
Quantifying the effect of cumulative water stress on stomatal conductance of Eucalyptus globulus and Eucalyptus nitens: A phenomenological approach.
1
1999
... 以往研究显示叶片水势(Oren et al., 1999)、叶片、枝条、根部和整株植物水力传导(Salleo et al., 2000; Addington et al., 2004; Bunce, 2006; Domec et al., 2009)、叶片相对含水量(Ewers et al., 2005)等会影响气孔导度对饱和水汽压亏缺的响应.而由于树龄和树高都会影响水力传导和气孔导度, 进而都会影响气孔导度斜率(Schäfer et al., 2000; Franks, 2004; Ewers et al., 2005; Buckley & Roberts, 2006; Ambrose et al., 2010; Woodruff et al., 2010).木材密度和气孔导度斜率之间也存在着显著的负相关关系(Héroult et al., 2013; Lin et al., 2015).植物结构属性, 如根叶比、根深等以及水力传导结构属性也会影响气孔导度斜率(Litvak et al., 2012; McCulloh & Woodruff, 2012).如与散孔材木相比, 环孔材木的气孔对环境因素变化的反应更加敏感(Bush et al., 2008) , 但是在Litvak等(2012)的研究中并未发现木材结构影响气孔导度斜率的现象.而随着测量高度的增高, 气孔导度对饱和水汽压亏缺响应的敏感性也会降低.叶肉导度和Vcmax也通过影响光合作用影响植物的水分利用效率(Wang et al., 2013a).Flexas等(2013)分析了44种不同物种水分利用效率的规律, 发现水分利用效率和叶肉导度与气孔导度的比值成正比.这个关系表明水分利用效率只有在固碳能力的提高高于水分丢失增加(气孔导度升高)的情况下, 才可以升高.可以预测, 同样的关系也应该在气孔导度斜率及Vcmax和气孔导度的比值间存在.临界氮利用效率也被证明和临界水分利用效率有互补的关系(Palmroth et al., 2013). ...
Height- related trends in stomatal sensitivity to leaf-to-air vapour pressure deficit in a tall conifer.
1
2010
... 以碳积累为代表的光合作用和以水分消耗为代表的蒸腾作用是所有植物功能模型的基础, 准确地模拟光合作用和蒸腾作用也是生态系统模型、植被模型、地球系统模型能够有效模拟碳、水和能量循环的必要条件(Wang et al., 2013b, 2015, 2016).由于以Farquhar等(1980)的光合作用模型为代表的机理模型的广泛应用, 通过对物种生物属性和环境参数(如光合最大羧化速率(Vcmax)、CO2浓度、温度和光照)的测量, 可以有效地模拟光合作用(Wullschleger, 1993; Kattge et al., 2009).相对于光合作用的机理模型, 蒸腾作用通常都是由经验或者半经验模型模拟出的气孔导度值来计算.过去40年里, 气孔导度的模拟工作大量开展.概括来讲, 对气孔导度的模拟有以下几种方式: (1)经验模型.大多通过气孔导度与多个环境因素如空气湿度(Lange et al., 1971)、水汽压亏缺(VPD)(Monteith, 1995)、叶片温度(Jarvis, 1976; Jones, 1992)和臭氧(Uddling et al., 2009)等的统计关系建立模型; (2)半经验模型.基于植物生理特性的假设, 但是仍然和经验模型结合, 如基于气孔导度和光合作用(Ball et al., 1987; Leuning, 1990, 1995; Misson et al., 2004b)、脱落酸(ABA) (Tardieu & Davies, 1993; Guteschick & Simonneau, 2002)、水分传导(Sperry et al., 1998; Oren et al., 1999; Tuzet et al., 2003)和膨压调节(Dewar, 2002; Buckley et al., 2003)等的关系建立的模型, 其参数不具有生物学属性; (3)机理模型.基于植物生理特性的假设, 其参数具有生物学特性, 并具有可以应用在各种环境条件下的潜力(Dewar, 2002; Medlyn et al., 2011; Prentice et al., 2014).很多文献已经综述过气孔导度的经验与半经验模型, 本文将侧重阐述气孔导度机理模型的研究. ...
Biochemical limitations to carbon assimilation in C3 plants—A retrospective analysis of the A/Ci curves from 109 species.
1
1993
... 在很多生态系统模型中, 土壤干旱胁迫对气孔导度影响的模拟通常是通过改变水分胁迫与光合作用/气孔导度之间的作用关系来实现的(Sala & Tenhunen, 1996; Verhoef & Allen, 2000).但也有模型是通过改变水分胁迫与光合作用或者光合作用参数Vcmax和最大电子传输速率(Jmax)的关系, 模拟水分胁迫下光合作用的改变进而模拟气孔导度的改变(Sellers et al., 1996; Ronda et al., 2001; Best et al., 2011).虽然在轻度干旱情况下, 上述几种做法可以模拟出水分胁迫对叶片尺度碳水通量的影响, 但在严重干旱情况下无法同时准确模拟光合作用和蒸腾作用(Verhoef & Allen, 2000; Reichstein et al., 2002; Keenan et al., 2009).Egea等(2011)提议, 在干旱条件下可以通过先限制叶肉导度, 进而限制气孔导度和光合作用参数的方案来提高光合作用和气孔导度模拟的准确性.Zhou等(2013)的研究结果也表明模拟干旱状况下的气孔导度, 必须同时考虑干旱对气孔和非气孔因素(如Vcmax)的综合影响.由于对干旱状况下气孔导度预测的不准确性, 现有的模型一般都倾向于高估光合作用和蒸腾作用对干旱的响应(Dietze, 2014).与以前从气孔(物理过程)或者非气孔(生化过程)的角度出发研究干旱对光合作用和气孔导度的影响不同, 气孔导度机理模型中的气孔导度斜率在干旱环境下的变化规律包含了干旱对气孔(生物物理因素)和非气孔因素, 包括最大光合效率和临界水分利用效率的综合影响, 为研究全球变化下陆地生态系统碳水耦合的适应性提供了一个简明且综合的框架.Manzoni等(2011)计算了50种植物的临界水分利用效率, 发现在忽略表皮水分丢失的情况下, 植物临界水分利用效率在轻微干旱的情况下增高, 而在严重干旱的情况下下降; 如果不忽略表皮水分丢失, 植物临界水分利用效率会随着水分胁迫的增加而提高; 而不同的植物功能型和不同的气候条件下, 反应也会不同.总体说来, 土壤水分较少时, 植物的水分利用效率会增高, 此时气孔导度斜率值也会降低(Lin et al., 2015).在环境水分条件与气孔导度斜率的变化规律之间还有很多值得讨论的地方, 我国学者对于水分利用的研究主要集中在单叶水分利用效率(付士磊等, 2006; 杨建伟等, 2006; 全先奎和王传宽, 2015)、气孔导度时空变异的调查(司建华等, 2008)、气孔导度对水分环境的响应(鱼腾飞等, 2012; 袁国富等, 2012)、单叶或者幼苗蒸腾速率(宋炳煜等, 2004; 岳广阳等, 2009)、树干液流密度(赵平等, 2006)、土壤水分定位监测(尹春梅等, 2008), 以及涡度协相关法测量生态系统水分利用效率(Zhou et al., 2013)等方面.土壤干旱如何影响不同功能型和物种的气孔导度斜率和临界水分利用效率, 还有待于进一步的研究. ...
黄土高原常用造林树种水分利用特征
1
2006
... 在很多生态系统模型中, 土壤干旱胁迫对气孔导度影响的模拟通常是通过改变水分胁迫与光合作用/气孔导度之间的作用关系来实现的(Sala & Tenhunen, 1996; Verhoef & Allen, 2000).但也有模型是通过改变水分胁迫与光合作用或者光合作用参数Vcmax和最大电子传输速率(Jmax)的关系, 模拟水分胁迫下光合作用的改变进而模拟气孔导度的改变(Sellers et al., 1996; Ronda et al., 2001; Best et al., 2011).虽然在轻度干旱情况下, 上述几种做法可以模拟出水分胁迫对叶片尺度碳水通量的影响, 但在严重干旱情况下无法同时准确模拟光合作用和蒸腾作用(Verhoef & Allen, 2000; Reichstein et al., 2002; Keenan et al., 2009).Egea等(2011)提议, 在干旱条件下可以通过先限制叶肉导度, 进而限制气孔导度和光合作用参数的方案来提高光合作用和气孔导度模拟的准确性.Zhou等(2013)的研究结果也表明模拟干旱状况下的气孔导度, 必须同时考虑干旱对气孔和非气孔因素(如Vcmax)的综合影响.由于对干旱状况下气孔导度预测的不准确性, 现有的模型一般都倾向于高估光合作用和蒸腾作用对干旱的响应(Dietze, 2014).与以前从气孔(物理过程)或者非气孔(生化过程)的角度出发研究干旱对光合作用和气孔导度的影响不同, 气孔导度机理模型中的气孔导度斜率在干旱环境下的变化规律包含了干旱对气孔(生物物理因素)和非气孔因素, 包括最大光合效率和临界水分利用效率的综合影响, 为研究全球变化下陆地生态系统碳水耦合的适应性提供了一个简明且综合的框架.Manzoni等(2011)计算了50种植物的临界水分利用效率, 发现在忽略表皮水分丢失的情况下, 植物临界水分利用效率在轻微干旱的情况下增高, 而在严重干旱的情况下下降; 如果不忽略表皮水分丢失, 植物临界水分利用效率会随着水分胁迫的增加而提高; 而不同的植物功能型和不同的气候条件下, 反应也会不同.总体说来, 土壤水分较少时, 植物的水分利用效率会增高, 此时气孔导度斜率值也会降低(Lin et al., 2015).在环境水分条件与气孔导度斜率的变化规律之间还有很多值得讨论的地方, 我国学者对于水分利用的研究主要集中在单叶水分利用效率(付士磊等, 2006; 杨建伟等, 2006; 全先奎和王传宽, 2015)、气孔导度时空变异的调查(司建华等, 2008)、气孔导度对水分环境的响应(鱼腾飞等, 2012; 袁国富等, 2012)、单叶或者幼苗蒸腾速率(宋炳煜等, 2004; 岳广阳等, 2009)、树干液流密度(赵平等, 2006)、土壤水分定位监测(尹春梅等, 2008), 以及涡度协相关法测量生态系统水分利用效率(Zhou et al., 2013)等方面.土壤干旱如何影响不同功能型和物种的气孔导度斜率和临界水分利用效率, 还有待于进一步的研究. ...
 , 胡亚林
, 胡亚林
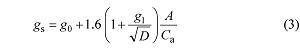



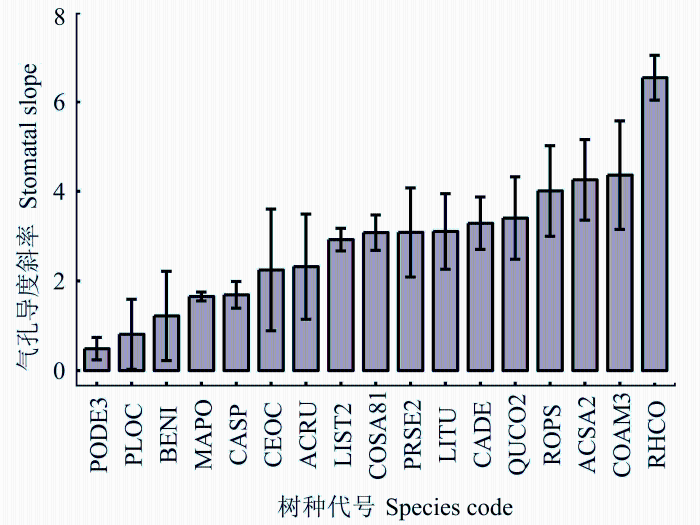
{kind=link}
{kind=link}