Aims Trees with different wood properties display variations in xylem anatomy and leaf vein structure, which may influence tree water transport efficiency and water-use strategy, and consequently constrain tree survival, growth and distribution. However, the effects of wood properties on leaf hydraulic conductance and vulnerability and their potential trade-offs at leaf level are not well understood. Our aims were to examine variations in leaf hydraulic traits of trees with different wood properties and explore potential trade-offs between leaf hydraulic efficiency and safety. Methods Nine tree species with different wood properties were selected for measuring the leaf hydraulic traits, including three diffuse-porous species (Populus davidiana, Tilia amurensis, Betula platyphylla), three ring-porous species (Quercus mongolica, Fraxinus mandshurica, Juglans mandshurica), and three non-porous species (Picea koraiensis, Pinus sylvestris var. mongolica, Pinus koraiensis). Four dominant and healthy trees per species were randomly selected. The hydraulic traits measured included leaf hydraulic conductance on leaf area (Karea) and dry mass (Kmass) basis, leaf hydraulic vulnerability (P50), and leaf water potential at turgor loss point (TLP), while the leaf structural traits were leaf dry mass content (LDMC), leaf density (LD) and leaf mass per unit area (LMA). Important findings The Karea, Kmass, and P50 differed significantly among the tree species with different woody properties (p < 0.05). Both Karea and Kmass were the lowest for the non-porous trees, and did not differ significantly between the diffuse-porous and ring-porous trees. The ring-porous trees had the highest P50 values, while the diffuse-porous and non-porous trees showed no significant differences in P50. Both Karea and Kmass were negatively correlated with P50 (p < 0.05) for all the trees, and the relationships for the diffuse-porous, ring-porous, and non-porous trees were fitted into linear, power, exponential functions, respectively. This indicates that significant trade-offs exist between leaf hydraulic efficiency and safety. The Kmass was correlated (p < 0.01) with TLP in a negative linear function for the diffuse- and ring-porous trees and in a negative exponential function for the non-porous trees. The P50 increased with increasing TLP. These results suggest that apoplastic and symplastic drought resistance are strictly coordinated in order to protect living cells from approaching their critical water status under water stresses. The Kmass was negatively correlated (p < 0.01) with LDMC, LD, or LMA, while the P50 was positively correlated with LDMC and LD; this suggests that variations in Kmass and P50 are driven by similar changes in structural traits regardless of wood traits. We conclude that the tree tolerance to hydraulic dysfunction increases with increasing carbon investment in the leaf hydraulic system.
JINYing, WANGChuan-Kuan. Leaf hydraulic traits and their trade-offs for nine Chinese temperate tree species with different wood properties. Chinese Journal of Plant Ecology, 2016, 40(7): 702-710 https://doi.org/10.17521/cjpe.2016.0064
研究地位于黑龙江帽儿山森林生态站(45.40° N, 127.67° E)。平均海拔400 m, 平均坡度10°-15°。地带性土壤为暗棕色森林土。气候属大陆性温带季风气候, 四季分明, 夏季湿热, 冬季干冷, 年降水量629 mm, 约50%的降水量集中在6-8月, 年蒸发量864 mm, 年平均气温3.1 ℃。1月份平均气温-18.5 ℃, 7月份平均气温22.0 ℃。无霜期120-140天。现有植被是原地带性植被——阔叶红松林屡遭人为干扰后演替成的天然次生林和人工林, 代表了东北东部山区典型的森林类型(Wang et al., 2013)。
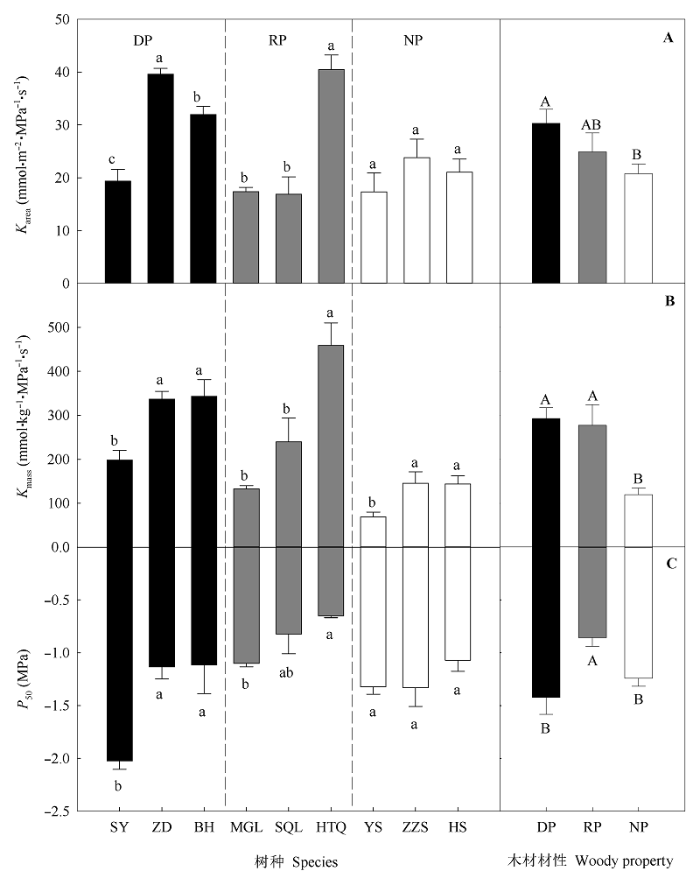
Fig. 1 Comparisons of leaf hydraulic traits among the tree species with different wood properties (mean ± SE). Different uppercase and lowercase letters above columns indicate significant differences among different wood properties and among different tree species with the same wood property, respectively (p < 0.05). Karea and Kmass, leaf hydraulic conductance per leaf area and dry mass, respectively; P50, leaf hydraulic vulnerability. See Table 1 for the codes of tree species and wood properties.
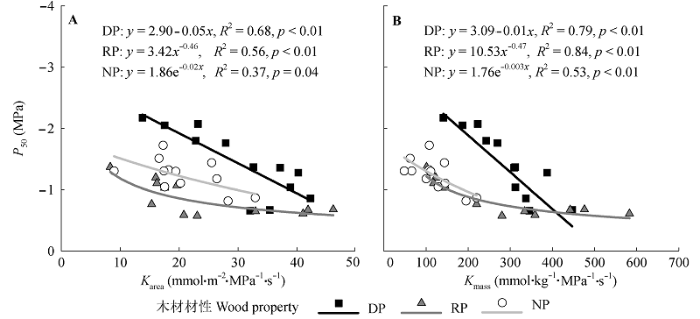
Fig. 2 Relationships between leaf hydraulic efficiency and hydraulic vulnerability (P50) of the trees with wood properties. Karea and Kmass, leaf hydraulic conductance per leaf area and dry mass, respectively. The codes of wood properties are listed in Table 1.
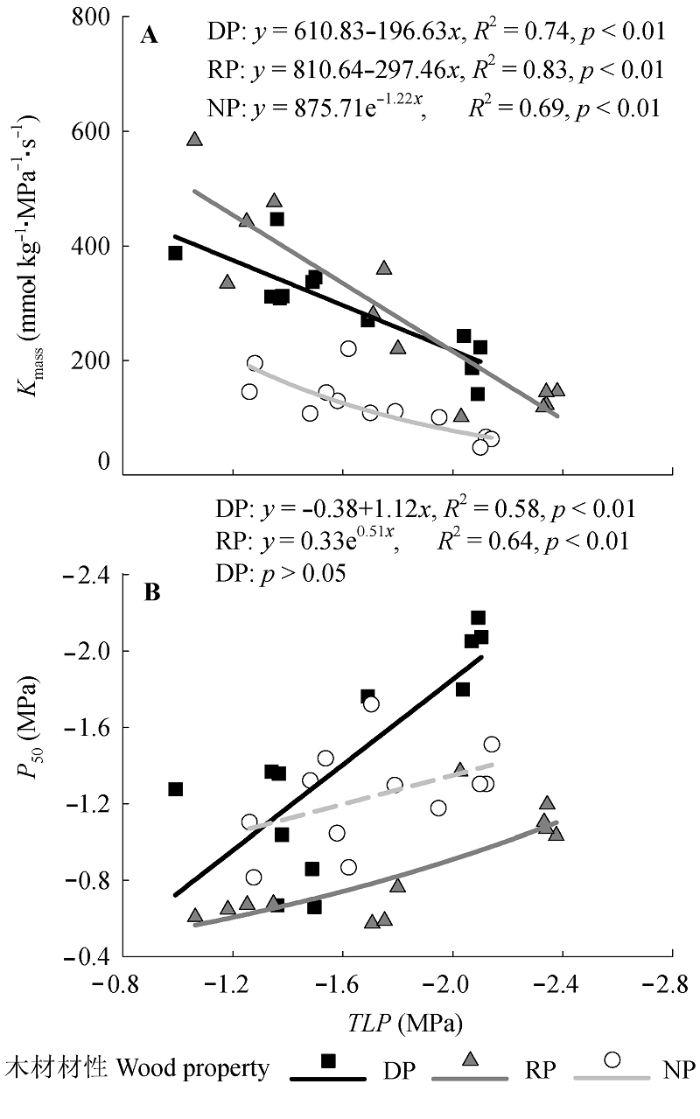
Fig. 3 Relationships between leaf water potential at turgor loss point (TLP) and (A) leaf-mass-based hydraulic conductance (Kmass) or (B) leaf hydraulic vulnerability (P50) of the trees with wood properties. The dash line denotes non-significant (p > 0.05). The codes of wood properties are listed in Table 1.
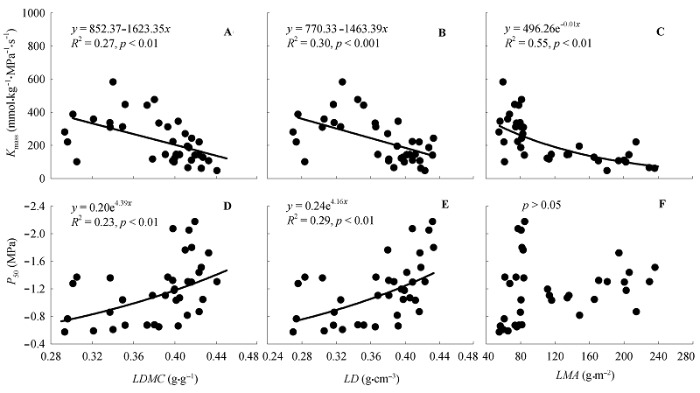
Fig. 4 The relationships between leaf hydraulics and structural traits of all tree species. Kmass, leaf hydraulic conductance per dry mass; LD, leaf density; LDMC, leaf dry mass content; LMA, leaf mass per unit area; P50, leaf hydraulic vulnerability.
3 讨论
3.1 不同材性树种叶导水率和水力脆弱性差异
不同材性树木的叶导水率差异显著, 以无孔材树种的Kleaf最低、而散孔材和环孔材树种的Kleaf差异不显著(图1)。左力翔等(2012)在我国西北干旱地区的测定结果也显示散孔材和环孔材树种的叶导水率无显著差异。这可能是由于散孔材和环孔材树种的Kleaf变异性较大导致的(图1)。从解剖结构看: 尽管环孔材导管直径可能大于散孔材, 可以承载的水分运输较强, 但由于导管密度与导管直径负相关, 因此其较低的导管密度抵消了其较大直径导致的高导水率(Giordano et al., 1978; Nardini et al., 2012a)。无孔材树种的Kleaf最低, 是由于其叶的水分运输依赖于由单一主脉组成的输水结构, 且输水单元为直径较窄的管胞, 因此, 其导水能力较差; 而散孔材和环孔材树种的叶水分运输依赖于高度分支的叶脉网络把水分输送到蒸发表面, 且输水单元为直径较大的导管(Brodribb et al., 2005; Sperry et al., 2006, 2008)。此外, 本研究中的无孔材树种均为常绿针叶树种(表1), 其叶导水率和P50均较低(图1), 这说明它们具有较强的栓塞阻力, 为此其叶水力系统的构建需要有较大的碳投资(Simonin et al., 2012), 这也可能是避免冬季冻融栓塞的一种对策(Feild & Brodribb, 2001)。
叶导水率也能在一定程度上反映树木对不同生境适应能力。一般来说, 生长在水分条件较好的立地上的树木的叶导水率和栓塞阻力较高, 而生长在水分条件较差的立地上的树木的叶导水率和栓塞阻力较低(Bucci et al., 2003; Nardini et al., 2003, 2012a)。本研究中的环孔材胡桃楸, 生长在湿润的沟谷地带(表1), 其叶导水率和P50在9个树种中最高(图1); 相反, 环孔材蒙古栎生长在干旱的山脊地带、散孔材山杨常生长在较干旱的山坡上部(表1; Zhang et al., 2014), 其叶导水率和P50较低。由此可见, 在小尺度范围内, 叶导水率和P50可影响树木的分布。叶导水率高的树种, 适宜生长在湿润的立地上, 对光和养分的竞争能力较强, 具有较高的光合和生长速率。相反, 叶导水率低的树种, 常有较高的栓塞阻力, 可在干旱条件下生存, 但同时也降低了其在资源丰富地区的竞争能力(Nardini et al., 2012a)。
Dynamic changes in hydraulic conductivity in petioles of two savanna tree species: Factors and mechanisms contributing to the refilling of embolized vessels
Disentangling the relative importance of climate, size and competition on tree growth in Iberian forests: Implications for forest management under global change
Xylem density, biomechanics and anatomical traits correlate with water stress in 17 evergreen shrub species of the Mediterranean-type climate region of South Africa
Moving water well: Comparing hydraulic efficiency in twigs and trunks of coniferous, ring-porous, and diffuse-porous saplings from temperate and tropical forests
Strong hydraulic segmentation and leaf senescence due to dehydration may trigger die-back in Nothofagus dombeyi under severe droughts: A comparison with the co-occurring Austrocedrus chilensis
Are symplast tolerance to intense drought conditions and xylem vulnerability to cavitation coordinated? An integrated analysis of photosynthetic, hydraulic and leaf level processes in two Mediterranean drought-resistant species
Effects of seasonal drought on water status, leaf spectral traits and fluorescence parameters in Tarenna depauperata Hutchins, a Chinese savanna evergreen species
.Plant Science Journal, 34, 117-126. (in Chinese with English abstract)[张树斌, 张教林, 曹坤芳 (2016).
Leaf hydraulic capacity in ferns, conifers and angiosperms: Impacts on photosynthetic maxima
4
2005
... 叶片既是植物进行光合作用的主要场所, 也是植物进行气体交换、水分蒸腾和运输的门户, 其形态结构特征直接影响植物的生理活动和生态功能(潘昕等, 2015; Zhu et al., 2015).叶片在进行光合作用的同时不可避免地丧失水分, 高水分传导效率能够保证更有效的水分运输, 补充植物蒸腾散失的水分, 以保持叶片较高的气孔导度, 从而维持高光合速率(Zhang et al., 2015; 张树斌等, 2016).因此, 叶片水力在限制树木生长、竞争和分布中起着关键的作用(Brodribb et al., 2005; Hao et al., 2008; Villagra et al., 2013).叶导水率(基于叶面积(Karea)或基于叶质量(Kmass))是指单位时间、单位水势压力梯度下流经叶的水流量(Sack & Holbrook, 2006), 表征叶的水分运输阻力.虽然叶水分运输路径在整个植株中占的比例较小, 但其水分运输阻力占整株的比例高达60%.叶水力脆弱性(P50)是指叶导水率降低50%时所对应的叶水势, 反映叶的栓塞阻力或水力安全.一般而言, 在枝水平上抗栓塞能力的增加常伴随水力效率的降低或碳消耗的增加(Bucci et al., 2006; Jacobsen et al., 2007), 但在叶水平上是否存在雷同的权衡关系尚不清楚(Nardini et al., 2012a; Villagra et al., 2013; Nardini & Luglio, 2014).以往研究报道叶水平的P50与Karea缺乏相关性, 因此, 人们常认为在叶水平上不存在水力效率与水力安全的权衡(Blackman et al., 2010; Scoffoni et al., 2011, 2012; Bucci et al. 2012; Nardini et al., 2012a).然而, 近期一些研究发现, Kmass与P50之间存在显著的相关关系(Nardini et al., 2012a; Nardini & Luglio, 2014).这引发人们思考: 考虑到叶肉组织中的水分运输和碳投资, 叶水力效率与水力安全之间是否存在一种更为普适的权衡关系?对这个问题的回答, 可深入理解叶生物物理结构构建与生理代谢的关系. ...
... 不同材性树种叶片的木质部结构和叶脉分布模式不同, 从而会导致水分运输效率和水分利用策略的差异(Brodribb et al., 2005; McCulloh et al., 2010; Scholz et al., 2014).无孔材树种依靠结构简单的单一主脉和直径较小的管胞输导水分; 散孔材和环孔材树种则依靠结构复杂的网状叶脉系统和直径较大的导管进行水分运输, 其中散孔材和环孔材树种叶脉木质部导管直径、导管密度和叶肉细胞结构可能也存在差异(Brodribb et al., 2005; 左力翔等, 2012).这些内部结构特性的差异使3种不同材性的树种在应对水分胁迫时采取躲避或忍受对策(李吉跃和翟洪波, 2000; Gomez-Aparicio et al., 2011; Carnicer et al., 2013; Coll et al., 2013).近年来关于叶水力学特性的研究已经成为植物生理生态的研究热点(潘莹萍和陈亚鹏, 2014; 张志亮等, 2014; 龚容和高琼, 2015; 金鹰和王传宽, 2015), 但国内对叶水力特性的报道不多.左力翔等(2012)报道我国西北干旱地区的散孔材和环孔材树种的叶导水率和水力脆弱性均无显著差异.我国东北森林地处水热同步的温带地区, 生长季树木受干旱胁迫的情况不多, 不同材性树种叶水力特性是否存在差异、叶水力效率与水力安全是否存在权衡尚不清楚.为此, 本研究在东北温带森林中选择了9种常见树种, 分别代表无孔材、散孔材和环孔材树种, 比较不同材性的树种叶水力性状的差异, 并探索叶水力效率与水力安全的权衡关系, 以便深入理解不同树种在群落中的共存机制. ...
... ).无孔材树种依靠结构简单的单一主脉和直径较小的管胞输导水分; 散孔材和环孔材树种则依靠结构复杂的网状叶脉系统和直径较大的导管进行水分运输, 其中散孔材和环孔材树种叶脉木质部导管直径、导管密度和叶肉细胞结构可能也存在差异(Brodribb et al., 2005; 左力翔等, 2012).这些内部结构特性的差异使3种不同材性的树种在应对水分胁迫时采取躲避或忍受对策(李吉跃和翟洪波, 2000; Gomez-Aparicio et al., 2011; Carnicer et al., 2013; Coll et al., 2013).近年来关于叶水力学特性的研究已经成为植物生理生态的研究热点(潘莹萍和陈亚鹏, 2014; 张志亮等, 2014; 龚容和高琼, 2015; 金鹰和王传宽, 2015), 但国内对叶水力特性的报道不多.左力翔等(2012)报道我国西北干旱地区的散孔材和环孔材树种的叶导水率和水力脆弱性均无显著差异.我国东北森林地处水热同步的温带地区, 生长季树木受干旱胁迫的情况不多, 不同材性树种叶水力特性是否存在差异、叶水力效率与水力安全是否存在权衡尚不清楚.为此, 本研究在东北温带森林中选择了9种常见树种, 分别代表无孔材、散孔材和环孔材树种, 比较不同材性的树种叶水力性状的差异, 并探索叶水力效率与水力安全的权衡关系, 以便深入理解不同树种在群落中的共存机制. ...
... 不同材性树木的叶导水率差异显著, 以无孔材树种的Kleaf最低、而散孔材和环孔材树种的Kleaf差异不显著(图1).左力翔等(2012)在我国西北干旱地区的测定结果也显示散孔材和环孔材树种的叶导水率无显著差异.这可能是由于散孔材和环孔材树种的Kleaf变异性较大导致的(图1).从解剖结构看: 尽管环孔材导管直径可能大于散孔材, 可以承载的水分运输较强, 但由于导管密度与导管直径负相关, 因此其较低的导管密度抵消了其较大直径导致的高导水率(Giordano et al., 1978; Nardini et al., 2012a).无孔材树种的Kleaf最低, 是由于其叶的水分运输依赖于由单一主脉组成的输水结构, 且输水单元为直径较窄的管胞, 因此, 其导水能力较差; 而散孔材和环孔材树种的叶水分运输依赖于高度分支的叶脉网络把水分输送到蒸发表面, 且输水单元为直径较大的导管(Brodribb et al., 2005; Sperry et al., 2006, 2008).此外, 本研究中的无孔材树种均为常绿针叶树种(表1), 其叶导水率和P50均较低(图1), 这说明它们具有较强的栓塞阻力, 为此其叶水力系统的构建需要有较大的碳投资(Simonin et al., 2012), 这也可能是避免冬季冻融栓塞的一种对策(Feild & Brodribb, 2001). ...
Hydraulic differences along the water transport system of South American Nothofagus species: Do leaves protect the stem functionality?
1
2012
... 叶片既是植物进行光合作用的主要场所, 也是植物进行气体交换、水分蒸腾和运输的门户, 其形态结构特征直接影响植物的生理活动和生态功能(潘昕等, 2015; Zhu et al., 2015).叶片在进行光合作用的同时不可避免地丧失水分, 高水分传导效率能够保证更有效的水分运输, 补充植物蒸腾散失的水分, 以保持叶片较高的气孔导度, 从而维持高光合速率(Zhang et al., 2015; 张树斌等, 2016).因此, 叶片水力在限制树木生长、竞争和分布中起着关键的作用(Brodribb et al., 2005; Hao et al., 2008; Villagra et al., 2013).叶导水率(基于叶面积(Karea)或基于叶质量(Kmass))是指单位时间、单位水势压力梯度下流经叶的水流量(Sack & Holbrook, 2006), 表征叶的水分运输阻力.虽然叶水分运输路径在整个植株中占的比例较小, 但其水分运输阻力占整株的比例高达60%.叶水力脆弱性(P50)是指叶导水率降低50%时所对应的叶水势, 反映叶的栓塞阻力或水力安全.一般而言, 在枝水平上抗栓塞能力的增加常伴随水力效率的降低或碳消耗的增加(Bucci et al., 2006; Jacobsen et al., 2007), 但在叶水平上是否存在雷同的权衡关系尚不清楚(Nardini et al., 2012a; Villagra et al., 2013; Nardini & Luglio, 2014).以往研究报道叶水平的P50与Karea缺乏相关性, 因此, 人们常认为在叶水平上不存在水力效率与水力安全的权衡(Blackman et al., 2010; Scoffoni et al., 2011, 2012; Bucci et al. 2012; Nardini et al., 2012a).然而, 近期一些研究发现, Kmass与P50之间存在显著的相关关系(Nardini et al., 2012a; Nardini & Luglio, 2014).这引发人们思考: 考虑到叶肉组织中的水分运输和碳投资, 叶水力效率与水力安全之间是否存在一种更为普适的权衡关系?对这个问题的回答, 可深入理解叶生物物理结构构建与生理代谢的关系. ...
Nutrient availability constrains the hydraulic architecture and water relations of savannah trees
1
2006
... 叶片既是植物进行光合作用的主要场所, 也是植物进行气体交换、水分蒸腾和运输的门户, 其形态结构特征直接影响植物的生理活动和生态功能(潘昕等, 2015; Zhu et al., 2015).叶片在进行光合作用的同时不可避免地丧失水分, 高水分传导效率能够保证更有效的水分运输, 补充植物蒸腾散失的水分, 以保持叶片较高的气孔导度, 从而维持高光合速率(Zhang et al., 2015; 张树斌等, 2016).因此, 叶片水力在限制树木生长、竞争和分布中起着关键的作用(Brodribb et al., 2005; Hao et al., 2008; Villagra et al., 2013).叶导水率(基于叶面积(Karea)或基于叶质量(Kmass))是指单位时间、单位水势压力梯度下流经叶的水流量(Sack & Holbrook, 2006), 表征叶的水分运输阻力.虽然叶水分运输路径在整个植株中占的比例较小, 但其水分运输阻力占整株的比例高达60%.叶水力脆弱性(P50)是指叶导水率降低50%时所对应的叶水势, 反映叶的栓塞阻力或水力安全.一般而言, 在枝水平上抗栓塞能力的增加常伴随水力效率的降低或碳消耗的增加(Bucci et al., 2006; Jacobsen et al., 2007), 但在叶水平上是否存在雷同的权衡关系尚不清楚(Nardini et al., 2012a; Villagra et al., 2013; Nardini & Luglio, 2014).以往研究报道叶水平的P50与Karea缺乏相关性, 因此, 人们常认为在叶水平上不存在水力效率与水力安全的权衡(Blackman et al., 2010; Scoffoni et al., 2011, 2012; Bucci et al. 2012; Nardini et al., 2012a).然而, 近期一些研究发现, Kmass与P50之间存在显著的相关关系(Nardini et al., 2012a; Nardini & Luglio, 2014).这引发人们思考: 考虑到叶肉组织中的水分运输和碳投资, 叶水力效率与水力安全之间是否存在一种更为普适的权衡关系?对这个问题的回答, 可深入理解叶生物物理结构构建与生理代谢的关系. ...
Dynamic changes in hydraulic conductivity in petioles of two savanna tree species: Factors and mechanisms contributing to the refilling of embolized vessels
1
2003
... 叶导水率也能在一定程度上反映树木对不同生境适应能力.一般来说, 生长在水分条件较好的立地上的树木的叶导水率和栓塞阻力较高, 而生长在水分条件较差的立地上的树木的叶导水率和栓塞阻力较低(Bucci et al., 2003; Nardini et al., 2003, 2012a).本研究中的环孔材胡桃楸, 生长在湿润的沟谷地带(表1), 其叶导水率和P50在9个树种中最高(图1); 相反, 环孔材蒙古栎生长在干旱的山脊地带、散孔材山杨常生长在较干旱的山坡上部(表1; Zhang et al., 2014), 其叶导水率和P50较低.由此可见, 在小尺度范围内, 叶导水率和P50可影响树木的分布.叶导水率高的树种, 适宜生长在湿润的立地上, 对光和养分的竞争能力较强, 具有较高的光合和生长速率.相反, 叶导水率低的树种, 常有较高的栓塞阻力, 可在干旱条件下生存, 但同时也降低了其在资源丰富地区的竞争能力(Nardini et al., 2012a). ...
Contrasting trait syndromes in angiosperms and conifers are associated with different responses of tree growth to temperature on a large scale
1
2013
... 不同材性树种叶片的木质部结构和叶脉分布模式不同, 从而会导致水分运输效率和水分利用策略的差异(Brodribb et al., 2005; McCulloh et al., 2010; Scholz et al., 2014).无孔材树种依靠结构简单的单一主脉和直径较小的管胞输导水分; 散孔材和环孔材树种则依靠结构复杂的网状叶脉系统和直径较大的导管进行水分运输, 其中散孔材和环孔材树种叶脉木质部导管直径、导管密度和叶肉细胞结构可能也存在差异(Brodribb et al., 2005; 左力翔等, 2012).这些内部结构特性的差异使3种不同材性的树种在应对水分胁迫时采取躲避或忍受对策(李吉跃和翟洪波, 2000; Gomez-Aparicio et al., 2011; Carnicer et al., 2013; Coll et al., 2013).近年来关于叶水力学特性的研究已经成为植物生理生态的研究热点(潘莹萍和陈亚鹏, 2014; 张志亮等, 2014; 龚容和高琼, 2015; 金鹰和王传宽, 2015), 但国内对叶水力特性的报道不多.左力翔等(2012)报道我国西北干旱地区的散孔材和环孔材树种的叶导水率和水力脆弱性均无显著差异.我国东北森林地处水热同步的温带地区, 生长季树木受干旱胁迫的情况不多, 不同材性树种叶水力特性是否存在差异、叶水力效率与水力安全是否存在权衡尚不清楚.为此, 本研究在东北温带森林中选择了9种常见树种, 分别代表无孔材、散孔材和环孔材树种, 比较不同材性的树种叶水力性状的差异, 并探索叶水力效率与水力安全的权衡关系, 以便深入理解不同树种在群落中的共存机制. ...
Multivariate effect gradients driving forest demographic responses in the Iberian Peninsula
1
2013
... 不同材性树种叶片的木质部结构和叶脉分布模式不同, 从而会导致水分运输效率和水分利用策略的差异(Brodribb et al., 2005; McCulloh et al., 2010; Scholz et al., 2014).无孔材树种依靠结构简单的单一主脉和直径较小的管胞输导水分; 散孔材和环孔材树种则依靠结构复杂的网状叶脉系统和直径较大的导管进行水分运输, 其中散孔材和环孔材树种叶脉木质部导管直径、导管密度和叶肉细胞结构可能也存在差异(Brodribb et al., 2005; 左力翔等, 2012).这些内部结构特性的差异使3种不同材性的树种在应对水分胁迫时采取躲避或忍受对策(李吉跃和翟洪波, 2000; Gomez-Aparicio et al., 2011; Carnicer et al., 2013; Coll et al., 2013).近年来关于叶水力学特性的研究已经成为植物生理生态的研究热点(潘莹萍和陈亚鹏, 2014; 张志亮等, 2014; 龚容和高琼, 2015; 金鹰和王传宽, 2015), 但国内对叶水力特性的报道不多.左力翔等(2012)报道我国西北干旱地区的散孔材和环孔材树种的叶导水率和水力脆弱性均无显著差异.我国东北森林地处水热同步的温带地区, 生长季树木受干旱胁迫的情况不多, 不同材性树种叶水力特性是否存在差异、叶水力效率与水力安全是否存在权衡尚不清楚.为此, 本研究在东北温带森林中选择了9种常见树种, 分别代表无孔材、散孔材和环孔材树种, 比较不同材性的树种叶水力性状的差异, 并探索叶水力效率与水力安全的权衡关系, 以便深入理解不同树种在群落中的共存机制. ...
Stem water transport and freeze-thaw xylem embolism in conifers and angiosperms in a Tasmanian treeline heath
1
2001
... 不同材性树木的叶导水率差异显著, 以无孔材树种的Kleaf最低、而散孔材和环孔材树种的Kleaf差异不显著(图1).左力翔等(2012)在我国西北干旱地区的测定结果也显示散孔材和环孔材树种的叶导水率无显著差异.这可能是由于散孔材和环孔材树种的Kleaf变异性较大导致的(图1).从解剖结构看: 尽管环孔材导管直径可能大于散孔材, 可以承载的水分运输较强, 但由于导管密度与导管直径负相关, 因此其较低的导管密度抵消了其较大直径导致的高导水率(Giordano et al., 1978; Nardini et al., 2012a).无孔材树种的Kleaf最低, 是由于其叶的水分运输依赖于由单一主脉组成的输水结构, 且输水单元为直径较窄的管胞, 因此, 其导水能力较差; 而散孔材和环孔材树种的叶水分运输依赖于高度分支的叶脉网络把水分输送到蒸发表面, 且输水单元为直径较大的导管(Brodribb et al., 2005; Sperry et al., 2006, 2008).此外, 本研究中的无孔材树种均为常绿针叶树种(表1), 其叶导水率和P50均较低(图1), 这说明它们具有较强的栓塞阻力, 为此其叶水力系统的构建需要有较大的碳投资(Simonin et al., 2012), 这也可能是避免冬季冻融栓塞的一种对策(Feild & Brodribb, 2001). ...
Flow in xylem vessels and Poiseuille’s law
1
1978
... 不同材性树木的叶导水率差异显著, 以无孔材树种的Kleaf最低、而散孔材和环孔材树种的Kleaf差异不显著(图1).左力翔等(2012)在我国西北干旱地区的测定结果也显示散孔材和环孔材树种的叶导水率无显著差异.这可能是由于散孔材和环孔材树种的Kleaf变异性较大导致的(图1).从解剖结构看: 尽管环孔材导管直径可能大于散孔材, 可以承载的水分运输较强, 但由于导管密度与导管直径负相关, 因此其较低的导管密度抵消了其较大直径导致的高导水率(Giordano et al., 1978; Nardini et al., 2012a).无孔材树种的Kleaf最低, 是由于其叶的水分运输依赖于由单一主脉组成的输水结构, 且输水单元为直径较窄的管胞, 因此, 其导水能力较差; 而散孔材和环孔材树种的叶水分运输依赖于高度分支的叶脉网络把水分输送到蒸发表面, 且输水单元为直径较大的导管(Brodribb et al., 2005; Sperry et al., 2006, 2008).此外, 本研究中的无孔材树种均为常绿针叶树种(表1), 其叶导水率和P50均较低(图1), 这说明它们具有较强的栓塞阻力, 为此其叶水力系统的构建需要有较大的碳投资(Simonin et al., 2012), 这也可能是避免冬季冻融栓塞的一种对策(Feild & Brodribb, 2001). ...
Disentangling the relative importance of climate, size and competition on tree growth in Iberian forests: Implications for forest management under global change
1
2011
... 不同材性树种叶片的木质部结构和叶脉分布模式不同, 从而会导致水分运输效率和水分利用策略的差异(Brodribb et al., 2005; McCulloh et al., 2010; Scholz et al., 2014).无孔材树种依靠结构简单的单一主脉和直径较小的管胞输导水分; 散孔材和环孔材树种则依靠结构复杂的网状叶脉系统和直径较大的导管进行水分运输, 其中散孔材和环孔材树种叶脉木质部导管直径、导管密度和叶肉细胞结构可能也存在差异(Brodribb et al., 2005; 左力翔等, 2012).这些内部结构特性的差异使3种不同材性的树种在应对水分胁迫时采取躲避或忍受对策(李吉跃和翟洪波, 2000; Gomez-Aparicio et al., 2011; Carnicer et al., 2013; Coll et al., 2013).近年来关于叶水力学特性的研究已经成为植物生理生态的研究热点(潘莹萍和陈亚鹏, 2014; 张志亮等, 2014; 龚容和高琼, 2015; 金鹰和王传宽, 2015), 但国内对叶水力特性的报道不多.左力翔等(2012)报道我国西北干旱地区的散孔材和环孔材树种的叶导水率和水力脆弱性均无显著差异.我国东北森林地处水热同步的温带地区, 生长季树木受干旱胁迫的情况不多, 不同材性树种叶水力特性是否存在差异、叶水力效率与水力安全是否存在权衡尚不清楚.为此, 本研究在东北温带森林中选择了9种常见树种, 分别代表无孔材、散孔材和环孔材树种, 比较不同材性的树种叶水力性状的差异, 并探索叶水力效率与水力安全的权衡关系, 以便深入理解不同树种在群落中的共存机制. ...
叶片结构的水力学特性对植物生理功能影响的研究进展
1
2015
... 不同材性树种叶片的木质部结构和叶脉分布模式不同, 从而会导致水分运输效率和水分利用策略的差异(Brodribb et al., 2005; McCulloh et al., 2010; Scholz et al., 2014).无孔材树种依靠结构简单的单一主脉和直径较小的管胞输导水分; 散孔材和环孔材树种则依靠结构复杂的网状叶脉系统和直径较大的导管进行水分运输, 其中散孔材和环孔材树种叶脉木质部导管直径、导管密度和叶肉细胞结构可能也存在差异(Brodribb et al., 2005; 左力翔等, 2012).这些内部结构特性的差异使3种不同材性的树种在应对水分胁迫时采取躲避或忍受对策(李吉跃和翟洪波, 2000; Gomez-Aparicio et al., 2011; Carnicer et al., 2013; Coll et al., 2013).近年来关于叶水力学特性的研究已经成为植物生理生态的研究热点(潘莹萍和陈亚鹏, 2014; 张志亮等, 2014; 龚容和高琼, 2015; 金鹰和王传宽, 2015), 但国内对叶水力特性的报道不多.左力翔等(2012)报道我国西北干旱地区的散孔材和环孔材树种的叶导水率和水力脆弱性均无显著差异.我国东北森林地处水热同步的温带地区, 生长季树木受干旱胁迫的情况不多, 不同材性树种叶水力特性是否存在差异、叶水力效率与水力安全是否存在权衡尚不清楚.为此, 本研究在东北温带森林中选择了9种常见树种, 分别代表无孔材、散孔材和环孔材树种, 比较不同材性的树种叶水力性状的差异, 并探索叶水力效率与水力安全的权衡关系, 以便深入理解不同树种在群落中的共存机制. ...
Stem and leaf hydraulics of congeneric tree species from adjacent tropical savanna and forest ecosystems
1
2008
... 叶片既是植物进行光合作用的主要场所, 也是植物进行气体交换、水分蒸腾和运输的门户, 其形态结构特征直接影响植物的生理活动和生态功能(潘昕等, 2015; Zhu et al., 2015).叶片在进行光合作用的同时不可避免地丧失水分, 高水分传导效率能够保证更有效的水分运输, 补充植物蒸腾散失的水分, 以保持叶片较高的气孔导度, 从而维持高光合速率(Zhang et al., 2015; 张树斌等, 2016).因此, 叶片水力在限制树木生长、竞争和分布中起着关键的作用(Brodribb et al., 2005; Hao et al., 2008; Villagra et al., 2013).叶导水率(基于叶面积(Karea)或基于叶质量(Kmass))是指单位时间、单位水势压力梯度下流经叶的水流量(Sack & Holbrook, 2006), 表征叶的水分运输阻力.虽然叶水分运输路径在整个植株中占的比例较小, 但其水分运输阻力占整株的比例高达60%.叶水力脆弱性(P50)是指叶导水率降低50%时所对应的叶水势, 反映叶的栓塞阻力或水力安全.一般而言, 在枝水平上抗栓塞能力的增加常伴随水力效率的降低或碳消耗的增加(Bucci et al., 2006; Jacobsen et al., 2007), 但在叶水平上是否存在雷同的权衡关系尚不清楚(Nardini et al., 2012a; Villagra et al., 2013; Nardini & Luglio, 2014).以往研究报道叶水平的P50与Karea缺乏相关性, 因此, 人们常认为在叶水平上不存在水力效率与水力安全的权衡(Blackman et al., 2010; Scoffoni et al., 2011, 2012; Bucci et al. 2012; Nardini et al., 2012a).然而, 近期一些研究发现, Kmass与P50之间存在显著的相关关系(Nardini et al., 2012a; Nardini & Luglio, 2014).这引发人们思考: 考虑到叶肉组织中的水分运输和碳投资, 叶水力效率与水力安全之间是否存在一种更为普适的权衡关系?对这个问题的回答, 可深入理解叶生物物理结构构建与生理代谢的关系. ...
Hydraulic failure and tree dieback are associated with high wood density in a temperate forest under extreme drought
1
2011
... 本研究结果显示, Kmass与LDMC、LD、LMA显著负相关; P50与LDMC、LD显著正相关(图4).这说明高的栓塞阻力与厚木质部导管壁、高主脉密度或导管密度有关, 而这些均与高LD和LMA有关(Blackman et al., 2010; Scoffoni et al., 2011; Nardini et al., 2012a).在枝水平上的研究也发现相似的结果, 即: 高木质部栓塞阻力与高木材密度相关(Meinzer et al., 2009; Hoffmann et al., 2011; Nardini et al., 2013); 而高木材密度意味着厚导管壁或大量的机械组织.因此, 为了增加栓塞阻力, 树木将大部分碳用于构建木材密度, 从而降低了用于生长的碳投资(Nardini & Luglio, 2014).这些结果表明, Kmass和P50的高低均与叶形态-解剖特性相关.叶导水率低的树种需要构建一个安全的木质部以降低栓塞风险, 而构建安全的木质部需要对叶水力系统构建投资较多的碳(即高LDMC和LD), 这就提高了叶的水分供给成本(Wikberg & Ögren, 2004; Simonin et al., 2012; Villagra et al., 2013; Nardini & Luglio, 2014). ...
Xylem density, biomechanics and anatomical traits correlate with water stress in 17 evergreen shrub species of the Mediterranean-type climate region of South Africa
1
2007
... 叶片既是植物进行光合作用的主要场所, 也是植物进行气体交换、水分蒸腾和运输的门户, 其形态结构特征直接影响植物的生理活动和生态功能(潘昕等, 2015; Zhu et al., 2015).叶片在进行光合作用的同时不可避免地丧失水分, 高水分传导效率能够保证更有效的水分运输, 补充植物蒸腾散失的水分, 以保持叶片较高的气孔导度, 从而维持高光合速率(Zhang et al., 2015; 张树斌等, 2016).因此, 叶片水力在限制树木生长、竞争和分布中起着关键的作用(Brodribb et al., 2005; Hao et al., 2008; Villagra et al., 2013).叶导水率(基于叶面积(Karea)或基于叶质量(Kmass))是指单位时间、单位水势压力梯度下流经叶的水流量(Sack & Holbrook, 2006), 表征叶的水分运输阻力.虽然叶水分运输路径在整个植株中占的比例较小, 但其水分运输阻力占整株的比例高达60%.叶水力脆弱性(P50)是指叶导水率降低50%时所对应的叶水势, 反映叶的栓塞阻力或水力安全.一般而言, 在枝水平上抗栓塞能力的增加常伴随水力效率的降低或碳消耗的增加(Bucci et al., 2006; Jacobsen et al., 2007), 但在叶水平上是否存在雷同的权衡关系尚不清楚(Nardini et al., 2012a; Villagra et al., 2013; Nardini & Luglio, 2014).以往研究报道叶水平的P50与Karea缺乏相关性, 因此, 人们常认为在叶水平上不存在水力效率与水力安全的权衡(Blackman et al., 2010; Scoffoni et al., 2011, 2012; Bucci et al. 2012; Nardini et al., 2012a).然而, 近期一些研究发现, Kmass与P50之间存在显著的相关关系(Nardini et al., 2012a; Nardini & Luglio, 2014).这引发人们思考: 考虑到叶肉组织中的水分运输和碳投资, 叶水力效率与水力安全之间是否存在一种更为普适的权衡关系?对这个问题的回答, 可深入理解叶生物物理结构构建与生理代谢的关系. ...
植物叶片水力与经济性状权衡关系的研究进展
1
2015
... 不同材性树种叶片的木质部结构和叶脉分布模式不同, 从而会导致水分运输效率和水分利用策略的差异(Brodribb et al., 2005; McCulloh et al., 2010; Scholz et al., 2014).无孔材树种依靠结构简单的单一主脉和直径较小的管胞输导水分; 散孔材和环孔材树种则依靠结构复杂的网状叶脉系统和直径较大的导管进行水分运输, 其中散孔材和环孔材树种叶脉木质部导管直径、导管密度和叶肉细胞结构可能也存在差异(Brodribb et al., 2005; 左力翔等, 2012).这些内部结构特性的差异使3种不同材性的树种在应对水分胁迫时采取躲避或忍受对策(李吉跃和翟洪波, 2000; Gomez-Aparicio et al., 2011; Carnicer et al., 2013; Coll et al., 2013).近年来关于叶水力学特性的研究已经成为植物生理生态的研究热点(潘莹萍和陈亚鹏, 2014; 张志亮等, 2014; 龚容和高琼, 2015; 金鹰和王传宽, 2015), 但国内对叶水力特性的报道不多.左力翔等(2012)报道我国西北干旱地区的散孔材和环孔材树种的叶导水率和水力脆弱性均无显著差异.我国东北森林地处水热同步的温带地区, 生长季树木受干旱胁迫的情况不多, 不同材性树种叶水力特性是否存在差异、叶水力效率与水力安全是否存在权衡尚不清楚.为此, 本研究在东北温带森林中选择了9种常见树种, 分别代表无孔材、散孔材和环孔材树种, 比较不同材性的树种叶水力性状的差异, 并探索叶水力效率与水力安全的权衡关系, 以便深入理解不同树种在群落中的共存机制. ...
Evidence for xylem embolism as a primary factor in dehydration-induced declines in leaf hydraulic conductance
1
2012
... 本研究发现, Kmass和P50均与质外体的抗旱阻力(TLP)有关(图3), 与前人的研究(Blackman et al., 2010; Scoffoni et al., 2012; Villagra et al., 2013)一致.TLP低的树种, 其叶导水率低, 但干旱诱导的质外体抗栓塞阻力高.P50和TLP的正相关关系表明共质体和质外体抗旱阻力的彼此协调; 在树木面临水分胁迫时, 膨压损失引起的木质部外水力特性变化与木质部内栓塞抵抗力的共同作用导致气孔关闭、蒸腾降低, 从而保护活细胞、防止其接近水分临界值 (Kim & Steudle, 2007; Blackman et al., 2010; Vila- grosa et al. 2010; Johnson et al., 2012). ...
Light and turgor affect the water permeability (aquaporins) of parenchyma cells in the midrib of
1
2007
... 本研究发现, Kmass和P50均与质外体的抗旱阻力(TLP)有关(图3), 与前人的研究(Blackman et al., 2010; Scoffoni et al., 2012; Villagra et al., 2013)一致.TLP低的树种, 其叶导水率低, 但干旱诱导的质外体抗栓塞阻力高.P50和TLP的正相关关系表明共质体和质外体抗旱阻力的彼此协调; 在树木面临水分胁迫时, 膨压损失引起的木质部外水力特性变化与木质部内栓塞抵抗力的共同作用导致气孔关闭、蒸腾降低, 从而保护活细胞、防止其接近水分临界值 (Kim & Steudle, 2007; Blackman et al., 2010; Vila- grosa et al. 2010; Johnson et al., 2012). ...
木本植物水力结构与抗旱性
1
2000
... 不同材性树种叶片的木质部结构和叶脉分布模式不同, 从而会导致水分运输效率和水分利用策略的差异(Brodribb et al., 2005; McCulloh et al., 2010; Scholz et al., 2014).无孔材树种依靠结构简单的单一主脉和直径较小的管胞输导水分; 散孔材和环孔材树种则依靠结构复杂的网状叶脉系统和直径较大的导管进行水分运输, 其中散孔材和环孔材树种叶脉木质部导管直径、导管密度和叶肉细胞结构可能也存在差异(Brodribb et al., 2005; 左力翔等, 2012).这些内部结构特性的差异使3种不同材性的树种在应对水分胁迫时采取躲避或忍受对策(李吉跃和翟洪波, 2000; Gomez-Aparicio et al., 2011; Carnicer et al., 2013; Coll et al., 2013).近年来关于叶水力学特性的研究已经成为植物生理生态的研究热点(潘莹萍和陈亚鹏, 2014; 张志亮等, 2014; 龚容和高琼, 2015; 金鹰和王传宽, 2015), 但国内对叶水力特性的报道不多.左力翔等(2012)报道我国西北干旱地区的散孔材和环孔材树种的叶导水率和水力脆弱性均无显著差异.我国东北森林地处水热同步的温带地区, 生长季树木受干旱胁迫的情况不多, 不同材性树种叶水力特性是否存在差异、叶水力效率与水力安全是否存在权衡尚不清楚.为此, 本研究在东北温带森林中选择了9种常见树种, 分别代表无孔材、散孔材和环孔材树种, 比较不同材性的树种叶水力性状的差异, 并探索叶水力效率与水力安全的权衡关系, 以便深入理解不同树种在群落中的共存机制. ...
Moving water well: Comparing hydraulic efficiency in twigs and trunks of coniferous, ring-porous, and diffuse-porous saplings from temperate and tropical forests
1
2010
... 不同材性树种叶片的木质部结构和叶脉分布模式不同, 从而会导致水分运输效率和水分利用策略的差异(Brodribb et al., 2005; McCulloh et al., 2010; Scholz et al., 2014).无孔材树种依靠结构简单的单一主脉和直径较小的管胞输导水分; 散孔材和环孔材树种则依靠结构复杂的网状叶脉系统和直径较大的导管进行水分运输, 其中散孔材和环孔材树种叶脉木质部导管直径、导管密度和叶肉细胞结构可能也存在差异(Brodribb et al., 2005; 左力翔等, 2012).这些内部结构特性的差异使3种不同材性的树种在应对水分胁迫时采取躲避或忍受对策(李吉跃和翟洪波, 2000; Gomez-Aparicio et al., 2011; Carnicer et al., 2013; Coll et al., 2013).近年来关于叶水力学特性的研究已经成为植物生理生态的研究热点(潘莹萍和陈亚鹏, 2014; 张志亮等, 2014; 龚容和高琼, 2015; 金鹰和王传宽, 2015), 但国内对叶水力特性的报道不多.左力翔等(2012)报道我国西北干旱地区的散孔材和环孔材树种的叶导水率和水力脆弱性均无显著差异.我国东北森林地处水热同步的温带地区, 生长季树木受干旱胁迫的情况不多, 不同材性树种叶水力特性是否存在差异、叶水力效率与水力安全是否存在权衡尚不清楚.为此, 本研究在东北温带森林中选择了9种常见树种, 分别代表无孔材、散孔材和环孔材树种, 比较不同材性的树种叶水力性状的差异, 并探索叶水力效率与水力安全的权衡关系, 以便深入理解不同树种在群落中的共存机制. ...
Xylem hydraulic safety margins in woody plants: Coordination of stomatal control of xylem tension with hydraulic capacitance
1
2009
... 本研究结果显示, Kmass与LDMC、LD、LMA显著负相关; P50与LDMC、LD显著正相关(图4).这说明高的栓塞阻力与厚木质部导管壁、高主脉密度或导管密度有关, 而这些均与高LD和LMA有关(Blackman et al., 2010; Scoffoni et al., 2011; Nardini et al., 2012a).在枝水平上的研究也发现相似的结果, 即: 高木质部栓塞阻力与高木材密度相关(Meinzer et al., 2009; Hoffmann et al., 2011; Nardini et al., 2013); 而高木材密度意味着厚导管壁或大量的机械组织.因此, 为了增加栓塞阻力, 树木将大部分碳用于构建木材密度, 从而降低了用于生长的碳投资(Nardini & Luglio, 2014).这些结果表明, Kmass和P50的高低均与叶形态-解剖特性相关.叶导水率低的树种需要构建一个安全的木质部以降低栓塞风险, 而构建安全的木质部需要对叶水力系统构建投资较多的碳(即高LDMC和LD), 这就提高了叶的水分供给成本(Wikberg & Ögren, 2004; Simonin et al., 2012; Villagra et al., 2013; Nardini & Luglio, 2014). ...
Shoot desiccation and hydraulic failure in temperate woody angiosperms during an extreme summer drought
1
2013
... 本研究结果显示, Kmass与LDMC、LD、LMA显著负相关; P50与LDMC、LD显著正相关(图4).这说明高的栓塞阻力与厚木质部导管壁、高主脉密度或导管密度有关, 而这些均与高LD和LMA有关(Blackman et al., 2010; Scoffoni et al., 2011; Nardini et al., 2012a).在枝水平上的研究也发现相似的结果, 即: 高木质部栓塞阻力与高木材密度相关(Meinzer et al., 2009; Hoffmann et al., 2011; Nardini et al., 2013); 而高木材密度意味着厚导管壁或大量的机械组织.因此, 为了增加栓塞阻力, 树木将大部分碳用于构建木材密度, 从而降低了用于生长的碳投资(Nardini & Luglio, 2014).这些结果表明, Kmass和P50的高低均与叶形态-解剖特性相关.叶导水率低的树种需要构建一个安全的木质部以降低栓塞风险, 而构建安全的木质部需要对叶水力系统构建投资较多的碳(即高LDMC和LD), 这就提高了叶的水分供给成本(Wikberg & Ögren, 2004; Simonin et al., 2012; Villagra et al., 2013; Nardini & Luglio, 2014). ...
Leaf hydraulic capacity and drought vulnerability: Possible trade-offs and correlations with climate across three major biomes
5
2014
... 叶片既是植物进行光合作用的主要场所, 也是植物进行气体交换、水分蒸腾和运输的门户, 其形态结构特征直接影响植物的生理活动和生态功能(潘昕等, 2015; Zhu et al., 2015).叶片在进行光合作用的同时不可避免地丧失水分, 高水分传导效率能够保证更有效的水分运输, 补充植物蒸腾散失的水分, 以保持叶片较高的气孔导度, 从而维持高光合速率(Zhang et al., 2015; 张树斌等, 2016).因此, 叶片水力在限制树木生长、竞争和分布中起着关键的作用(Brodribb et al., 2005; Hao et al., 2008; Villagra et al., 2013).叶导水率(基于叶面积(Karea)或基于叶质量(Kmass))是指单位时间、单位水势压力梯度下流经叶的水流量(Sack & Holbrook, 2006), 表征叶的水分运输阻力.虽然叶水分运输路径在整个植株中占的比例较小, 但其水分运输阻力占整株的比例高达60%.叶水力脆弱性(P50)是指叶导水率降低50%时所对应的叶水势, 反映叶的栓塞阻力或水力安全.一般而言, 在枝水平上抗栓塞能力的增加常伴随水力效率的降低或碳消耗的增加(Bucci et al., 2006; Jacobsen et al., 2007), 但在叶水平上是否存在雷同的权衡关系尚不清楚(Nardini et al., 2012a; Villagra et al., 2013; Nardini & Luglio, 2014).以往研究报道叶水平的P50与Karea缺乏相关性, 因此, 人们常认为在叶水平上不存在水力效率与水力安全的权衡(Blackman et al., 2010; Scoffoni et al., 2011, 2012; Bucci et al. 2012; Nardini et al., 2012a).然而, 近期一些研究发现, Kmass与P50之间存在显著的相关关系(Nardini et al., 2012a; Nardini & Luglio, 2014).这引发人们思考: 考虑到叶肉组织中的水分运输和碳投资, 叶水力效率与水力安全之间是否存在一种更为普适的权衡关系?对这个问题的回答, 可深入理解叶生物物理结构构建与生理代谢的关系. ...
... 以往研究通常用叶面积来标准化叶导水率(Karea), 但近期用叶质量来标准化叶导水率(Kmass)的研究越来越多(Nardini et al., 2012b; Scoffoni et al., 2012; Simonin et al., 2012; Nardini & Luglio, 2014).近期研究认为, Karea侧重于叶片-大气界面上的表面生理过程, 忽略了叶肉组织中的水分运输, 只能衡量叶水力系统给叶片蒸发表面提供水分的效率.对于叶片较薄或叶脉末端距离叶蒸发表面较近、水分在叶肉组织中运输距离较短的树种而言, 用Karea可很好地衡量叶水分运输效率(Nardini et al., 2012a).但如果叶片中的水流从叶脉组织出来后并不是直接到达表皮, 而是要在叶肉组织中流经较长的距离(Zwien- iecki et al., 2007)时, 用Kmass才能充分地反映出叶水力系统给整个叶组织提供水分的效率.此外, Kmass还可以估计构建叶水力系统的碳投资(Nardini et al., 2012b; Simonin et al., 2012).本研究同时比较了不同材性Karea和Kmass的差异, 结果相似, 但无孔材Kmass显著低于散孔材和环孔材; 这说明用叶质量标准化的Kmass估测针叶树种整个叶组织的导水率更恰当. ...
... 本研究结果显示, Kmass与LDMC、LD、LMA显著负相关; P50与LDMC、LD显著正相关(图4).这说明高的栓塞阻力与厚木质部导管壁、高主脉密度或导管密度有关, 而这些均与高LD和LMA有关(Blackman et al., 2010; Scoffoni et al., 2011; Nardini et al., 2012a).在枝水平上的研究也发现相似的结果, 即: 高木质部栓塞阻力与高木材密度相关(Meinzer et al., 2009; Hoffmann et al., 2011; Nardini et al., 2013); 而高木材密度意味着厚导管壁或大量的机械组织.因此, 为了增加栓塞阻力, 树木将大部分碳用于构建木材密度, 从而降低了用于生长的碳投资(Nardini & Luglio, 2014).这些结果表明, Kmass和P50的高低均与叶形态-解剖特性相关.叶导水率低的树种需要构建一个安全的木质部以降低栓塞风险, 而构建安全的木质部需要对叶水力系统构建投资较多的碳(即高LDMC和LD), 这就提高了叶的水分供给成本(Wikberg & Ögren, 2004; Simonin et al., 2012; Villagra et al., 2013; Nardini & Luglio, 2014). ...
Alternative methods for scaling leaf hydraulic conductance offer new insights into the structure-function relationships of sun and shade leaves
2
2012
... 以往研究通常用叶面积来标准化叶导水率(Karea), 但近期用叶质量来标准化叶导水率(Kmass)的研究越来越多(Nardini et al., 2012b; Scoffoni et al., 2012; Simonin et al., 2012; Nardini & Luglio, 2014).近期研究认为, Karea侧重于叶片-大气界面上的表面生理过程, 忽略了叶肉组织中的水分运输, 只能衡量叶水力系统给叶片蒸发表面提供水分的效率.对于叶片较薄或叶脉末端距离叶蒸发表面较近、水分在叶肉组织中运输距离较短的树种而言, 用Karea可很好地衡量叶水分运输效率(Nardini et al., 2012a).但如果叶片中的水流从叶脉组织出来后并不是直接到达表皮, 而是要在叶肉组织中流经较长的距离(Zwien- iecki et al., 2007)时, 用Kmass才能充分地反映出叶水力系统给整个叶组织提供水分的效率.此外, Kmass还可以估计构建叶水力系统的碳投资(Nardini et al., 2012b; Simonin et al., 2012).本研究同时比较了不同材性Karea和Kmass的差异, 结果相似, 但无孔材Kmass显著低于散孔材和环孔材; 这说明用叶质量标准化的Kmass估测针叶树种整个叶组织的导水率更恰当. ...
... 还可以估计构建叶水力系统的碳投资(Nardini et al., 2012b; Simonin et al., 2012).本研究同时比较了不同材性Karea和Kmass的差异, 结果相似, 但无孔材Kmass显著低于散孔材和环孔材; 这说明用叶质量标准化的Kmass估测针叶树种整个叶组织的导水率更恰当. ...
Changes in leaf hydraulic conductance correlate with leaf vein embolism in Cercis siliquastrum L
1
2003
... 叶导水率也能在一定程度上反映树木对不同生境适应能力.一般来说, 生长在水分条件较好的立地上的树木的叶导水率和栓塞阻力较高, 而生长在水分条件较差的立地上的树木的叶导水率和栓塞阻力较低(Bucci et al., 2003; Nardini et al., 2003, 2012a).本研究中的环孔材胡桃楸, 生长在湿润的沟谷地带(表1), 其叶导水率和P50在9个树种中最高(图1); 相反, 环孔材蒙古栎生长在干旱的山脊地带、散孔材山杨常生长在较干旱的山坡上部(表1; Zhang et al., 2014), 其叶导水率和P50较低.由此可见, 在小尺度范围内, 叶导水率和P50可影响树木的分布.叶导水率高的树种, 适宜生长在湿润的立地上, 对光和养分的竞争能力较强, 具有较高的光合和生长速率.相反, 叶导水率低的树种, 常有较高的栓塞阻力, 可在干旱条件下生存, 但同时也降低了其在资源丰富地区的竞争能力(Nardini et al., 2012a). ...
基于叶片解剖结构对青藏高原25种灌木的抗旱性评价
1
2015
... 叶片既是植物进行光合作用的主要场所, 也是植物进行气体交换、水分蒸腾和运输的门户, 其形态结构特征直接影响植物的生理活动和生态功能(潘昕等, 2015; Zhu et al., 2015).叶片在进行光合作用的同时不可避免地丧失水分, 高水分传导效率能够保证更有效的水分运输, 补充植物蒸腾散失的水分, 以保持叶片较高的气孔导度, 从而维持高光合速率(Zhang et al., 2015; 张树斌等, 2016).因此, 叶片水力在限制树木生长、竞争和分布中起着关键的作用(Brodribb et al., 2005; Hao et al., 2008; Villagra et al., 2013).叶导水率(基于叶面积(Karea)或基于叶质量(Kmass))是指单位时间、单位水势压力梯度下流经叶的水流量(Sack & Holbrook, 2006), 表征叶的水分运输阻力.虽然叶水分运输路径在整个植株中占的比例较小, 但其水分运输阻力占整株的比例高达60%.叶水力脆弱性(P50)是指叶导水率降低50%时所对应的叶水势, 反映叶的栓塞阻力或水力安全.一般而言, 在枝水平上抗栓塞能力的增加常伴随水力效率的降低或碳消耗的增加(Bucci et al., 2006; Jacobsen et al., 2007), 但在叶水平上是否存在雷同的权衡关系尚不清楚(Nardini et al., 2012a; Villagra et al., 2013; Nardini & Luglio, 2014).以往研究报道叶水平的P50与Karea缺乏相关性, 因此, 人们常认为在叶水平上不存在水力效率与水力安全的权衡(Blackman et al., 2010; Scoffoni et al., 2011, 2012; Bucci et al. 2012; Nardini et al., 2012a).然而, 近期一些研究发现, Kmass与P50之间存在显著的相关关系(Nardini et al., 2012a; Nardini & Luglio, 2014).这引发人们思考: 考虑到叶肉组织中的水分运输和碳投资, 叶水力效率与水力安全之间是否存在一种更为普适的权衡关系?对这个问题的回答, 可深入理解叶生物物理结构构建与生理代谢的关系. ...
叶片水力性状研究进展
1
2014
... 不同材性树种叶片的木质部结构和叶脉分布模式不同, 从而会导致水分运输效率和水分利用策略的差异(Brodribb et al., 2005; McCulloh et al., 2010; Scholz et al., 2014).无孔材树种依靠结构简单的单一主脉和直径较小的管胞输导水分; 散孔材和环孔材树种则依靠结构复杂的网状叶脉系统和直径较大的导管进行水分运输, 其中散孔材和环孔材树种叶脉木质部导管直径、导管密度和叶肉细胞结构可能也存在差异(Brodribb et al., 2005; 左力翔等, 2012).这些内部结构特性的差异使3种不同材性的树种在应对水分胁迫时采取躲避或忍受对策(李吉跃和翟洪波, 2000; Gomez-Aparicio et al., 2011; Carnicer et al., 2013; Coll et al., 2013).近年来关于叶水力学特性的研究已经成为植物生理生态的研究热点(潘莹萍和陈亚鹏, 2014; 张志亮等, 2014; 龚容和高琼, 2015; 金鹰和王传宽, 2015), 但国内对叶水力特性的报道不多.左力翔等(2012)报道我国西北干旱地区的散孔材和环孔材树种的叶导水率和水力脆弱性均无显著差异.我国东北森林地处水热同步的温带地区, 生长季树木受干旱胁迫的情况不多, 不同材性树种叶水力特性是否存在差异、叶水力效率与水力安全是否存在权衡尚不清楚.为此, 本研究在东北温带森林中选择了9种常见树种, 分别代表无孔材、散孔材和环孔材树种, 比较不同材性的树种叶水力性状的差异, 并探索叶水力效率与水力安全的权衡关系, 以便深入理解不同树种在群落中的共存机制. ...
Leaf hydraulics
1
2006
... 叶片既是植物进行光合作用的主要场所, 也是植物进行气体交换、水分蒸腾和运输的门户, 其形态结构特征直接影响植物的生理活动和生态功能(潘昕等, 2015; Zhu et al., 2015).叶片在进行光合作用的同时不可避免地丧失水分, 高水分传导效率能够保证更有效的水分运输, 补充植物蒸腾散失的水分, 以保持叶片较高的气孔导度, 从而维持高光合速率(Zhang et al., 2015; 张树斌等, 2016).因此, 叶片水力在限制树木生长、竞争和分布中起着关键的作用(Brodribb et al., 2005; Hao et al., 2008; Villagra et al., 2013).叶导水率(基于叶面积(Karea)或基于叶质量(Kmass))是指单位时间、单位水势压力梯度下流经叶的水流量(Sack & Holbrook, 2006), 表征叶的水分运输阻力.虽然叶水分运输路径在整个植株中占的比例较小, 但其水分运输阻力占整株的比例高达60%.叶水力脆弱性(P50)是指叶导水率降低50%时所对应的叶水势, 反映叶的栓塞阻力或水力安全.一般而言, 在枝水平上抗栓塞能力的增加常伴随水力效率的降低或碳消耗的增加(Bucci et al., 2006; Jacobsen et al., 2007), 但在叶水平上是否存在雷同的权衡关系尚不清楚(Nardini et al., 2012a; Villagra et al., 2013; Nardini & Luglio, 2014).以往研究报道叶水平的P50与Karea缺乏相关性, 因此, 人们常认为在叶水平上不存在水力效率与水力安全的权衡(Blackman et al., 2010; Scoffoni et al., 2011, 2012; Bucci et al. 2012; Nardini et al., 2012a).然而, 近期一些研究发现, Kmass与P50之间存在显著的相关关系(Nardini et al., 2012a; Nardini & Luglio, 2014).这引发人们思考: 考虑到叶肉组织中的水分运输和碳投资, 叶水力效率与水力安全之间是否存在一种更为普适的权衡关系?对这个问题的回答, 可深入理解叶生物物理结构构建与生理代谢的关系. ...
Strong hydraulic segmentation and leaf senescence due to dehydration may trigger die-back in Nothofagus dombeyi under severe droughts: A comparison with the co-occurring Austrocedrus chilensis
1
2014
... 不同材性树种叶片的木质部结构和叶脉分布模式不同, 从而会导致水分运输效率和水分利用策略的差异(Brodribb et al., 2005; McCulloh et al., 2010; Scholz et al., 2014).无孔材树种依靠结构简单的单一主脉和直径较小的管胞输导水分; 散孔材和环孔材树种则依靠结构复杂的网状叶脉系统和直径较大的导管进行水分运输, 其中散孔材和环孔材树种叶脉木质部导管直径、导管密度和叶肉细胞结构可能也存在差异(Brodribb et al., 2005; 左力翔等, 2012).这些内部结构特性的差异使3种不同材性的树种在应对水分胁迫时采取躲避或忍受对策(李吉跃和翟洪波, 2000; Gomez-Aparicio et al., 2011; Carnicer et al., 2013; Coll et al., 2013).近年来关于叶水力学特性的研究已经成为植物生理生态的研究热点(潘莹萍和陈亚鹏, 2014; 张志亮等, 2014; 龚容和高琼, 2015; 金鹰和王传宽, 2015), 但国内对叶水力特性的报道不多.左力翔等(2012)报道我国西北干旱地区的散孔材和环孔材树种的叶导水率和水力脆弱性均无显著差异.我国东北森林地处水热同步的温带地区, 生长季树木受干旱胁迫的情况不多, 不同材性树种叶水力特性是否存在差异、叶水力效率与水力安全是否存在权衡尚不清楚.为此, 本研究在东北温带森林中选择了9种常见树种, 分别代表无孔材、散孔材和环孔材树种, 比较不同材性的树种叶水力性状的差异, 并探索叶水力效率与水力安全的权衡关系, 以便深入理解不同树种在群落中的共存机制. ...
Dynamics of leaf hydraulic conductance with water status: Quantification and analysis of species differences under steady state
4
2012
... 叶片既是植物进行光合作用的主要场所, 也是植物进行气体交换、水分蒸腾和运输的门户, 其形态结构特征直接影响植物的生理活动和生态功能(潘昕等, 2015; Zhu et al., 2015).叶片在进行光合作用的同时不可避免地丧失水分, 高水分传导效率能够保证更有效的水分运输, 补充植物蒸腾散失的水分, 以保持叶片较高的气孔导度, 从而维持高光合速率(Zhang et al., 2015; 张树斌等, 2016).因此, 叶片水力在限制树木生长、竞争和分布中起着关键的作用(Brodribb et al., 2005; Hao et al., 2008; Villagra et al., 2013).叶导水率(基于叶面积(Karea)或基于叶质量(Kmass))是指单位时间、单位水势压力梯度下流经叶的水流量(Sack & Holbrook, 2006), 表征叶的水分运输阻力.虽然叶水分运输路径在整个植株中占的比例较小, 但其水分运输阻力占整株的比例高达60%.叶水力脆弱性(P50)是指叶导水率降低50%时所对应的叶水势, 反映叶的栓塞阻力或水力安全.一般而言, 在枝水平上抗栓塞能力的增加常伴随水力效率的降低或碳消耗的增加(Bucci et al., 2006; Jacobsen et al., 2007), 但在叶水平上是否存在雷同的权衡关系尚不清楚(Nardini et al., 2012a; Villagra et al., 2013; Nardini & Luglio, 2014).以往研究报道叶水平的P50与Karea缺乏相关性, 因此, 人们常认为在叶水平上不存在水力效率与水力安全的权衡(Blackman et al., 2010; Scoffoni et al., 2011, 2012; Bucci et al. 2012; Nardini et al., 2012a).然而, 近期一些研究发现, Kmass与P50之间存在显著的相关关系(Nardini et al., 2012a; Nardini & Luglio, 2014).这引发人们思考: 考虑到叶肉组织中的水分运输和碳投资, 叶水力效率与水力安全之间是否存在一种更为普适的权衡关系?对这个问题的回答, 可深入理解叶生物物理结构构建与生理代谢的关系. ...
... 以往研究通常用叶面积来标准化叶导水率(Karea), 但近期用叶质量来标准化叶导水率(Kmass)的研究越来越多(Nardini et al., 2012b; Scoffoni et al., 2012; Simonin et al., 2012; Nardini & Luglio, 2014).近期研究认为, Karea侧重于叶片-大气界面上的表面生理过程, 忽略了叶肉组织中的水分运输, 只能衡量叶水力系统给叶片蒸发表面提供水分的效率.对于叶片较薄或叶脉末端距离叶蒸发表面较近、水分在叶肉组织中运输距离较短的树种而言, 用Karea可很好地衡量叶水分运输效率(Nardini et al., 2012a).但如果叶片中的水流从叶脉组织出来后并不是直接到达表皮, 而是要在叶肉组织中流经较长的距离(Zwien- iecki et al., 2007)时, 用Kmass才能充分地反映出叶水力系统给整个叶组织提供水分的效率.此外, Kmass还可以估计构建叶水力系统的碳投资(Nardini et al., 2012b; Simonin et al., 2012).本研究同时比较了不同材性Karea和Kmass的差异, 结果相似, 但无孔材Kmass显著低于散孔材和环孔材; 这说明用叶质量标准化的Kmass估测针叶树种整个叶组织的导水率更恰当. ...
... 我们发现, 即便有材性效应, 但所有树种的P50均随Karea或Kmass的增大而显著下降(图2), 表明这些树种的叶水力效率与水力安全存在不同程度的权衡关系; 这与前人基于叶质量标准化叶导水率研究叶片水力效率与安全的权衡关系(Blackman et al., 2010; Scoffoni et al., 2012)一致, 说明树木常以降低导水率和提高碳投资为代价来增加栓塞引起的水力失调阻力. ...
... 本研究发现, Kmass和P50均与质外体的抗旱阻力(TLP)有关(图3), 与前人的研究(Blackman et al., 2010; Scoffoni et al., 2012; Villagra et al., 2013)一致.TLP低的树种, 其叶导水率低, 但干旱诱导的质外体抗栓塞阻力高.P50和TLP的正相关关系表明共质体和质外体抗旱阻力的彼此协调; 在树木面临水分胁迫时, 膨压损失引起的木质部外水力特性变化与木质部内栓塞抵抗力的共同作用导致气孔关闭、蒸腾降低, 从而保护活细胞、防止其接近水分临界值 (Kim & Steudle, 2007; Blackman et al., 2010; Vila- grosa et al. 2010; Johnson et al., 2012). ...
Decline of leaf hydraulic conductance with dehydration: Relationship to leaf size and venation architecture
2
2011
... 叶片既是植物进行光合作用的主要场所, 也是植物进行气体交换、水分蒸腾和运输的门户, 其形态结构特征直接影响植物的生理活动和生态功能(潘昕等, 2015; Zhu et al., 2015).叶片在进行光合作用的同时不可避免地丧失水分, 高水分传导效率能够保证更有效的水分运输, 补充植物蒸腾散失的水分, 以保持叶片较高的气孔导度, 从而维持高光合速率(Zhang et al., 2015; 张树斌等, 2016).因此, 叶片水力在限制树木生长、竞争和分布中起着关键的作用(Brodribb et al., 2005; Hao et al., 2008; Villagra et al., 2013).叶导水率(基于叶面积(Karea)或基于叶质量(Kmass))是指单位时间、单位水势压力梯度下流经叶的水流量(Sack & Holbrook, 2006), 表征叶的水分运输阻力.虽然叶水分运输路径在整个植株中占的比例较小, 但其水分运输阻力占整株的比例高达60%.叶水力脆弱性(P50)是指叶导水率降低50%时所对应的叶水势, 反映叶的栓塞阻力或水力安全.一般而言, 在枝水平上抗栓塞能力的增加常伴随水力效率的降低或碳消耗的增加(Bucci et al., 2006; Jacobsen et al., 2007), 但在叶水平上是否存在雷同的权衡关系尚不清楚(Nardini et al., 2012a; Villagra et al., 2013; Nardini & Luglio, 2014).以往研究报道叶水平的P50与Karea缺乏相关性, 因此, 人们常认为在叶水平上不存在水力效率与水力安全的权衡(Blackman et al., 2010; Scoffoni et al., 2011, 2012; Bucci et al. 2012; Nardini et al., 2012a).然而, 近期一些研究发现, Kmass与P50之间存在显著的相关关系(Nardini et al., 2012a; Nardini & Luglio, 2014).这引发人们思考: 考虑到叶肉组织中的水分运输和碳投资, 叶水力效率与水力安全之间是否存在一种更为普适的权衡关系?对这个问题的回答, 可深入理解叶生物物理结构构建与生理代谢的关系. ...
... 本研究结果显示, Kmass与LDMC、LD、LMA显著负相关; P50与LDMC、LD显著正相关(图4).这说明高的栓塞阻力与厚木质部导管壁、高主脉密度或导管密度有关, 而这些均与高LD和LMA有关(Blackman et al., 2010; Scoffoni et al., 2011; Nardini et al., 2012a).在枝水平上的研究也发现相似的结果, 即: 高木质部栓塞阻力与高木材密度相关(Meinzer et al., 2009; Hoffmann et al., 2011; Nardini et al., 2013); 而高木材密度意味着厚导管壁或大量的机械组织.因此, 为了增加栓塞阻力, 树木将大部分碳用于构建木材密度, 从而降低了用于生长的碳投资(Nardini & Luglio, 2014).这些结果表明, Kmass和P50的高低均与叶形态-解剖特性相关.叶导水率低的树种需要构建一个安全的木质部以降低栓塞风险, 而构建安全的木质部需要对叶水力系统构建投资较多的碳(即高LDMC和LD), 这就提高了叶的水分供给成本(Wikberg & Ögren, 2004; Simonin et al., 2012; Villagra et al., 2013; Nardini & Luglio, 2014). ...
Hydraulic conductance of leaves correlates with leaf lifespan: Implications for lifetime carbon gain
4
2012
... 不同材性树木的叶导水率差异显著, 以无孔材树种的Kleaf最低、而散孔材和环孔材树种的Kleaf差异不显著(图1).左力翔等(2012)在我国西北干旱地区的测定结果也显示散孔材和环孔材树种的叶导水率无显著差异.这可能是由于散孔材和环孔材树种的Kleaf变异性较大导致的(图1).从解剖结构看: 尽管环孔材导管直径可能大于散孔材, 可以承载的水分运输较强, 但由于导管密度与导管直径负相关, 因此其较低的导管密度抵消了其较大直径导致的高导水率(Giordano et al., 1978; Nardini et al., 2012a).无孔材树种的Kleaf最低, 是由于其叶的水分运输依赖于由单一主脉组成的输水结构, 且输水单元为直径较窄的管胞, 因此, 其导水能力较差; 而散孔材和环孔材树种的叶水分运输依赖于高度分支的叶脉网络把水分输送到蒸发表面, 且输水单元为直径较大的导管(Brodribb et al., 2005; Sperry et al., 2006, 2008).此外, 本研究中的无孔材树种均为常绿针叶树种(表1), 其叶导水率和P50均较低(图1), 这说明它们具有较强的栓塞阻力, 为此其叶水力系统的构建需要有较大的碳投资(Simonin et al., 2012), 这也可能是避免冬季冻融栓塞的一种对策(Feild & Brodribb, 2001). ...
... 以往研究通常用叶面积来标准化叶导水率(Karea), 但近期用叶质量来标准化叶导水率(Kmass)的研究越来越多(Nardini et al., 2012b; Scoffoni et al., 2012; Simonin et al., 2012; Nardini & Luglio, 2014).近期研究认为, Karea侧重于叶片-大气界面上的表面生理过程, 忽略了叶肉组织中的水分运输, 只能衡量叶水力系统给叶片蒸发表面提供水分的效率.对于叶片较薄或叶脉末端距离叶蒸发表面较近、水分在叶肉组织中运输距离较短的树种而言, 用Karea可很好地衡量叶水分运输效率(Nardini et al., 2012a).但如果叶片中的水流从叶脉组织出来后并不是直接到达表皮, 而是要在叶肉组织中流经较长的距离(Zwien- iecki et al., 2007)时, 用Kmass才能充分地反映出叶水力系统给整个叶组织提供水分的效率.此外, Kmass还可以估计构建叶水力系统的碳投资(Nardini et al., 2012b; Simonin et al., 2012).本研究同时比较了不同材性Karea和Kmass的差异, 结果相似, 但无孔材Kmass显著低于散孔材和环孔材; 这说明用叶质量标准化的Kmass估测针叶树种整个叶组织的导水率更恰当. ...
... ; Simonin et al., 2012).本研究同时比较了不同材性Karea和Kmass的差异, 结果相似, 但无孔材Kmass显著低于散孔材和环孔材; 这说明用叶质量标准化的Kmass估测针叶树种整个叶组织的导水率更恰当. ...
... 本研究结果显示, Kmass与LDMC、LD、LMA显著负相关; P50与LDMC、LD显著正相关(图4).这说明高的栓塞阻力与厚木质部导管壁、高主脉密度或导管密度有关, 而这些均与高LD和LMA有关(Blackman et al., 2010; Scoffoni et al., 2011; Nardini et al., 2012a).在枝水平上的研究也发现相似的结果, 即: 高木质部栓塞阻力与高木材密度相关(Meinzer et al., 2009; Hoffmann et al., 2011; Nardini et al., 2013); 而高木材密度意味着厚导管壁或大量的机械组织.因此, 为了增加栓塞阻力, 树木将大部分碳用于构建木材密度, 从而降低了用于生长的碳投资(Nardini & Luglio, 2014).这些结果表明, Kmass和P50的高低均与叶形态-解剖特性相关.叶导水率低的树种需要构建一个安全的木质部以降低栓塞风险, 而构建安全的木质部需要对叶水力系统构建投资较多的碳(即高LDMC和LD), 这就提高了叶的水分供给成本(Wikberg & Ögren, 2004; Simonin et al., 2012; Villagra et al., 2013; Nardini & Luglio, 2014). ...
Size and function in conifer tracheids and angiosperm vessels
1
2006
... 不同材性树木的叶导水率差异显著, 以无孔材树种的Kleaf最低、而散孔材和环孔材树种的Kleaf差异不显著(图1).左力翔等(2012)在我国西北干旱地区的测定结果也显示散孔材和环孔材树种的叶导水率无显著差异.这可能是由于散孔材和环孔材树种的Kleaf变异性较大导致的(图1).从解剖结构看: 尽管环孔材导管直径可能大于散孔材, 可以承载的水分运输较强, 但由于导管密度与导管直径负相关, 因此其较低的导管密度抵消了其较大直径导致的高导水率(Giordano et al., 1978; Nardini et al., 2012a).无孔材树种的Kleaf最低, 是由于其叶的水分运输依赖于由单一主脉组成的输水结构, 且输水单元为直径较窄的管胞, 因此, 其导水能力较差; 而散孔材和环孔材树种的叶水分运输依赖于高度分支的叶脉网络把水分输送到蒸发表面, 且输水单元为直径较大的导管(Brodribb et al., 2005; Sperry et al., 2006, 2008).此外, 本研究中的无孔材树种均为常绿针叶树种(表1), 其叶导水率和P50均较低(图1), 这说明它们具有较强的栓塞阻力, 为此其叶水力系统的构建需要有较大的碳投资(Simonin et al., 2012), 这也可能是避免冬季冻融栓塞的一种对策(Feild & Brodribb, 2001). ...
Safety and efficiency conflicts in hydraulic architecture: Scaling from tissues to trees
1
2008
... 不同材性树木的叶导水率差异显著, 以无孔材树种的Kleaf最低、而散孔材和环孔材树种的Kleaf差异不显著(图1).左力翔等(2012)在我国西北干旱地区的测定结果也显示散孔材和环孔材树种的叶导水率无显著差异.这可能是由于散孔材和环孔材树种的Kleaf变异性较大导致的(图1).从解剖结构看: 尽管环孔材导管直径可能大于散孔材, 可以承载的水分运输较强, 但由于导管密度与导管直径负相关, 因此其较低的导管密度抵消了其较大直径导致的高导水率(Giordano et al., 1978; Nardini et al., 2012a).无孔材树种的Kleaf最低, 是由于其叶的水分运输依赖于由单一主脉组成的输水结构, 且输水单元为直径较窄的管胞, 因此, 其导水能力较差; 而散孔材和环孔材树种的叶水分运输依赖于高度分支的叶脉网络把水分输送到蒸发表面, 且输水单元为直径较大的导管(Brodribb et al., 2005; Sperry et al., 2006, 2008).此外, 本研究中的无孔材树种均为常绿针叶树种(表1), 其叶导水率和P50均较低(图1), 这说明它们具有较强的栓塞阻力, 为此其叶水力系统的构建需要有较大的碳投资(Simonin et al., 2012), 这也可能是避免冬季冻融栓塞的一种对策(Feild & Brodribb, 2001). ...
The measurement of the turgor pressure and the water relations of plants by the pressure- bomb technique
1972
Are symplast tolerance to intense drought conditions and xylem vulnerability to cavitation coordinated? An integrated analysis of photosynthetic, hydraulic and leaf level processes in two Mediterranean drought-resistant species
1
2010
... 本研究发现, Kmass和P50均与质外体的抗旱阻力(TLP)有关(图3), 与前人的研究(Blackman et al., 2010; Scoffoni et al., 2012; Villagra et al., 2013)一致.TLP低的树种, 其叶导水率低, 但干旱诱导的质外体抗栓塞阻力高.P50和TLP的正相关关系表明共质体和质外体抗旱阻力的彼此协调; 在树木面临水分胁迫时, 膨压损失引起的木质部外水力特性变化与木质部内栓塞抵抗力的共同作用导致气孔关闭、蒸腾降低, 从而保护活细胞、防止其接近水分临界值 (Kim & Steudle, 2007; Blackman et al., 2010; Vila- grosa et al. 2010; Johnson et al., 2012). ...
Functional relationships between leaf hydraulics and leaf economic traits in response to nutrient addition in subtropical tree species
4
2013
... 叶片既是植物进行光合作用的主要场所, 也是植物进行气体交换、水分蒸腾和运输的门户, 其形态结构特征直接影响植物的生理活动和生态功能(潘昕等, 2015; Zhu et al., 2015).叶片在进行光合作用的同时不可避免地丧失水分, 高水分传导效率能够保证更有效的水分运输, 补充植物蒸腾散失的水分, 以保持叶片较高的气孔导度, 从而维持高光合速率(Zhang et al., 2015; 张树斌等, 2016).因此, 叶片水力在限制树木生长、竞争和分布中起着关键的作用(Brodribb et al., 2005; Hao et al., 2008; Villagra et al., 2013).叶导水率(基于叶面积(Karea)或基于叶质量(Kmass))是指单位时间、单位水势压力梯度下流经叶的水流量(Sack & Holbrook, 2006), 表征叶的水分运输阻力.虽然叶水分运输路径在整个植株中占的比例较小, 但其水分运输阻力占整株的比例高达60%.叶水力脆弱性(P50)是指叶导水率降低50%时所对应的叶水势, 反映叶的栓塞阻力或水力安全.一般而言, 在枝水平上抗栓塞能力的增加常伴随水力效率的降低或碳消耗的增加(Bucci et al., 2006; Jacobsen et al., 2007), 但在叶水平上是否存在雷同的权衡关系尚不清楚(Nardini et al., 2012a; Villagra et al., 2013; Nardini & Luglio, 2014).以往研究报道叶水平的P50与Karea缺乏相关性, 因此, 人们常认为在叶水平上不存在水力效率与水力安全的权衡(Blackman et al., 2010; Scoffoni et al., 2011, 2012; Bucci et al. 2012; Nardini et al., 2012a).然而, 近期一些研究发现, Kmass与P50之间存在显著的相关关系(Nardini et al., 2012a; Nardini & Luglio, 2014).这引发人们思考: 考虑到叶肉组织中的水分运输和碳投资, 叶水力效率与水力安全之间是否存在一种更为普适的权衡关系?对这个问题的回答, 可深入理解叶生物物理结构构建与生理代谢的关系. ...
... ; Villagra et al., 2013; Nardini & Luglio, 2014).以往研究报道叶水平的P50与Karea缺乏相关性, 因此, 人们常认为在叶水平上不存在水力效率与水力安全的权衡(Blackman et al., 2010; Scoffoni et al., 2011, 2012; Bucci et al. 2012; Nardini et al., 2012a).然而, 近期一些研究发现, Kmass与P50之间存在显著的相关关系(Nardini et al., 2012a; Nardini & Luglio, 2014).这引发人们思考: 考虑到叶肉组织中的水分运输和碳投资, 叶水力效率与水力安全之间是否存在一种更为普适的权衡关系?对这个问题的回答, 可深入理解叶生物物理结构构建与生理代谢的关系. ...
... 本研究发现, Kmass和P50均与质外体的抗旱阻力(TLP)有关(图3), 与前人的研究(Blackman et al., 2010; Scoffoni et al., 2012; Villagra et al., 2013)一致.TLP低的树种, 其叶导水率低, 但干旱诱导的质外体抗栓塞阻力高.P50和TLP的正相关关系表明共质体和质外体抗旱阻力的彼此协调; 在树木面临水分胁迫时, 膨压损失引起的木质部外水力特性变化与木质部内栓塞抵抗力的共同作用导致气孔关闭、蒸腾降低, 从而保护活细胞、防止其接近水分临界值 (Kim & Steudle, 2007; Blackman et al., 2010; Vila- grosa et al. 2010; Johnson et al., 2012). ...
... 本研究结果显示, Kmass与LDMC、LD、LMA显著负相关; P50与LDMC、LD显著正相关(图4).这说明高的栓塞阻力与厚木质部导管壁、高主脉密度或导管密度有关, 而这些均与高LD和LMA有关(Blackman et al., 2010; Scoffoni et al., 2011; Nardini et al., 2012a).在枝水平上的研究也发现相似的结果, 即: 高木质部栓塞阻力与高木材密度相关(Meinzer et al., 2009; Hoffmann et al., 2011; Nardini et al., 2013); 而高木材密度意味着厚导管壁或大量的机械组织.因此, 为了增加栓塞阻力, 树木将大部分碳用于构建木材密度, 从而降低了用于生长的碳投资(Nardini & Luglio, 2014).这些结果表明, Kmass和P50的高低均与叶形态-解剖特性相关.叶导水率低的树种需要构建一个安全的木质部以降低栓塞风险, 而构建安全的木质部需要对叶水力系统构建投资较多的碳(即高LDMC和LD), 这就提高了叶的水分供给成本(Wikberg & Ögren, 2004; Simonin et al., 2012; Villagra et al., 2013; Nardini & Luglio, 2014). ...
Seasonality of soil CO2 efflux in a temperate forest: Biophysical effects of snowpack and spring freeze-thaw cycles
1
2013
... 研究地位于黑龙江帽儿山森林生态站(45.40° N, 127.67° E).平均海拔400 m, 平均坡度10°-15°.地带性土壤为暗棕色森林土.气候属大陆性温带季风气候, 四季分明, 夏季湿热, 冬季干冷, 年降水量629 mm, 约50%的降水量集中在6-8月, 年蒸发量864 mm, 年平均气温3.1 ℃.1月份平均气温-18.5 ℃, 7月份平均气温22.0 ℃.无霜期120-140天.现有植被是原地带性植被——阔叶红松林屡遭人为干扰后演替成的天然次生林和人工林, 代表了东北东部山区典型的森林类型(Wang et al., 2013). ...
Interrelationships between water use and growth traits in biomass-producing willows
1
2004
... 本研究结果显示, Kmass与LDMC、LD、LMA显著负相关; P50与LDMC、LD显著正相关(图4).这说明高的栓塞阻力与厚木质部导管壁、高主脉密度或导管密度有关, 而这些均与高LD和LMA有关(Blackman et al., 2010; Scoffoni et al., 2011; Nardini et al., 2012a).在枝水平上的研究也发现相似的结果, 即: 高木质部栓塞阻力与高木材密度相关(Meinzer et al., 2009; Hoffmann et al., 2011; Nardini et al., 2013); 而高木材密度意味着厚导管壁或大量的机械组织.因此, 为了增加栓塞阻力, 树木将大部分碳用于构建木材密度, 从而降低了用于生长的碳投资(Nardini & Luglio, 2014).这些结果表明, Kmass和P50的高低均与叶形态-解剖特性相关.叶导水率低的树种需要构建一个安全的木质部以降低栓塞风险, 而构建安全的木质部需要对叶水力系统构建投资较多的碳(即高LDMC和LD), 这就提高了叶的水分供给成本(Wikberg & Ögren, 2004; Simonin et al., 2012; Villagra et al., 2013; Nardini & Luglio, 2014). ...
Spatial variations in non-structural carbohydrates in stems of twelve temperate tree species
1
2014
... 叶导水率也能在一定程度上反映树木对不同生境适应能力.一般来说, 生长在水分条件较好的立地上的树木的叶导水率和栓塞阻力较高, 而生长在水分条件较差的立地上的树木的叶导水率和栓塞阻力较低(Bucci et al., 2003; Nardini et al., 2003, 2012a).本研究中的环孔材胡桃楸, 生长在湿润的沟谷地带(表1), 其叶导水率和P50在9个树种中最高(图1); 相反, 环孔材蒙古栎生长在干旱的山脊地带、散孔材山杨常生长在较干旱的山坡上部(表1; Zhang et al., 2014), 其叶导水率和P50较低.由此可见, 在小尺度范围内, 叶导水率和P50可影响树木的分布.叶导水率高的树种, 适宜生长在湿润的立地上, 对光和养分的竞争能力较强, 具有较高的光合和生长速率.相反, 叶导水率低的树种, 常有较高的栓塞阻力, 可在干旱条件下生存, 但同时也降低了其在资源丰富地区的竞争能力(Nardini et al., 2012a). ...

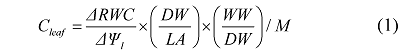


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}