全球变化在全世界范围内以前所未有的速度影响着生物多样性的变化, 进而使自然生态系统功能产生不可预知的改变(Grime, 1997 ; Cardinale et al ., 2012 ; MacDougall et al ., 2013 ; Valencia et al ., 2015 ), 尤其是随着物种消失速率的加快, 越来越多的学者开始关注物种多样性的改变和由之而引起的群落结构变化对生态系统功能的影响(Grime, 1997 ; Tilman et al ., 1997 ; Díaz & Cabido, 2001 ; Díaz et al ., 2007 ; Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 )。20多年来, 许多研究工作围绕这一问题展开, 但这些工作大多仅关注物种多样性和群落结构与单一或少数生态系统功能之间的关系(Hector et al ., 1999 ; Waide et al ., 1999 ; Simova et al ., 2013 ; Valencia et al ., 2015 ), 对生态系统多种功能同时评价和研究的工作还不多见, 而生态系统最为重要的价值是同时提供多种功能和服务的能力(Sanderson et al ., 2004 ; Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ; Soliveres et al ., 2014 ), 因此物种多样性与生态系统多功能性(multifunctionality)之间关系的研究显得十分 重要。
“multifunctionality”一词首先由Sanderson等(2004)提出, 他们认为经营者在管理牧场和草场生态系统时往往只关注初级生产力的提高(因为生产输出与经济效益直接相关), 而忽略了生态系统的其他价值(如环境和美学效益), 指出从可持续发展的角度出发, 生态系统同时执行多种功能的能力“multifunctionality”更应该被重视, 并从环境效益和以人类为中心的经济效益两个角度出发提供了体现multifunctionality的15个功能特征。Hector和Bagchi (2007)引用“multifunctionality”这一概念在《Nature》上发表文章, 认为生态系统的多种服务或过程就是“ecosystem ‘multifunctionality’ ”, 并第一次量化研究了物种多样性同时对8个联合的生态系统过程所起的作用。Gamfeldt等(2008)、Zavaleta等(2010)又引用Hector和Bagchi (2007)的这篇文章, 分别在《Ecology》和《PNAS》上发文探讨和量化生物多样性对维持高水平“multifunctionality”的重要性, 认为“multifunctionality”即为生态系统同时维持多个生态功能或服务的能力。自此, 随着引用的增多, “multifunctionality”一词开始被人们广泛认可并正式使用。Maestre等(2012b)在《Science》上发表文章, 应用14个土壤变量(分别体现C、N、P循环三个生态系统过程)来综合评价全球干旱地区的生态系统多功能性, 其选用的指标和评价方法几乎成为近年来生态系统多功能性研究的标准方法(Maestre et al ., 2012a ; Byrnes et al ., 2014 ; Soliveres et al ., 2014 ; Wagg et al ., 2014 ; Valencia et al ., 2015 )。随后, Maestre等(2012a)在《Journal of Ecology》上, Soli- veres等(2014)在《Global Ecology and Biogeography》上、Wagg等(2014)在《PANS》上发表的多功能性研究论文均采用了Maestre等的文章的土壤变量和评价方法, 最新由Valencia等(2015)发表在《New Phytologist》上的文章同样引用了这些土壤变量及其评价方法, 这些文章的发表, 标志着生态系统多功能性(multifunctionality)概念正在逐步完善, 其评价方法也趋于统一。
虽然Maestre等的多功能性(multifunctionality)评价指标和量化方法被许多学者所采用, 但也存在一些争议(Byrnes et al ., 2014 ; Lefcheck et al ., 2015 ), 本文笔者认为这一方法中对每个样方多功能性指数(M )的计算只是14个评价指标的Z-scores简单平均, 并没有考虑各个指标间的权重, 显然默认每个指标有同等的地位和作用并不符合自然生态系统的真实情况。另外, Maestre等(2012b)选择的14个土壤指标涉及C、N、P的循环与固存, 尽管C、N、P这三大物质循环在生态系统中占据着重要的地位, 但土壤同时也有涵养水分、平衡和缓冲外界干扰、为微生物提供稳定的环境、为各种物理生化反应提供介质和可持续地供给植物营养等多重功能(王仁忠, 1998 ; 王玉辉等, 2002 ; Sanderson et al ., 2004 ; Pei et al ., 2008 ), 仅用C、N、P的循环与固存这三个功能来体现生态系统的“多”功能性, 不免略显勉强。因此本文用改进的量化方法, 尝试选取农业实践中较为常见、测定更为简易的土壤指标, 并结合具体的立地条件, 对内蒙古三种不同利用方式下的草原群落进行多功能性的评价, 探讨物种多样性与多功能性指数之间的关系, 试图达到以下目的: (1)结合立地条件, 选取更为合适的土壤多功能性评价指标; (2)与Maestre等人的评价方法相对比, 建立更为科学的多功能性指数评价方法; (3)评价不同土地利用方式下草原生态系统的多功能性, 探讨物种多样性对多功能性的影响。
1 材料和方法
1.1 实验样地
实验在内蒙古辉河国家自然保护区(118.80°- 119.75° E, 48.15°-49.00° N)内进行。此保护区主要位于内蒙古自治区呼伦贝尔市鄂温克族自治旗, 处于大兴安岭西北坡山地向呼伦贝尔高原的过渡地段, 海拔800-1000 m。地带性土壤类型为黑钙土和栗钙土。保护区总面积达3468.48 km2 , 草甸草原面积1610.48 km2 。区域气候属中温带大陆性季风气候, 冬季漫长寒冷, 夏季温润短促, 年平均气温为-2.4-2.2 ℃; 无霜期为100-120天; 2008到2014年的平均年降水量为375.03 mm, 70%的降水集中在6-8月份。该保护区属于全国17个生物多样性保护关键区域, 于2007年加入“中国生物圈保护区网络” (资料来源于辉河国家级自然保护区管理局)。
1.2 样地设置和植被调查
2008年在地形均一、地势平坦的自由放牧草场设置4个样地(样地间距为200-600 m), 每个样地 (2970 m2 )对应分为两部分, 分别进行围封和刈割管理。围封样地无放牧无干扰; 刈割样地每年秋季8月20日左右刈割一次, 无其他干扰; 围封和刈割样地均用围栏封育, 围栏外依然保持自由放牧, 载畜量为3.9-4.5只羊·hm-2 , 结合生物量、草地盖度和高度等情况, 我们界定放牧草场为重度放牧。2014年分别在每个围封和刈割样地中心用机械排列法各设置15个样方, 样方行距和列距均为5 m, 总计围封和刈割样方各为60个。由于放牧样地物种组成和群落结构较为简单, 且匀质化程度较高, 故在自由放牧草场设置两个样地, 每个样地设置10个样方, 共计20个样方。
在每个1 m × 1 m的样方内进行群落调查, 记录每个物种的多度、盖度和高度, 多度为样方内的密度, 盖度由同一调查人员统一估测, 每个样方中中等高度的个体高度记为该物种的高度值。
1.3 多功能性评价指标的选取与测定
尽管按照最初的概念, 多功能性是指生态系统同时维持各种生态系统功能和服务的能力, 其评价指标应涉及经济、环境、景观美学等各方面的指标(Sanderson et al ., 2004 ), 但随着多功能性概念的发展, 狭义的概念被人们提出, 同时维持多个生态功能或服务的能力即可称为“多功能性”, 这便导致不同的研究中各研究者选用的指标和数量并不相同, 涉及的生态系统功能和数量也各不相同(Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ; Zavaleta et al ., 2010 ; Maestre et al ., 2012a , 2012b ; Wagg et al ., 2014 ; Lefcheck et al ., 2015 )。近年来多功能性的研究中, 评价指标的选取多参考Maestre等(2012a, 2012b)的研究, 由于土壤-植被是相互依赖、互为反馈的复合体, 并且土壤是生态系统的养分库和种子库, 土壤指标很大程度上能反映生态系统的多个功能(如C、N、P的固存, 水土保持, 承载野生动植物等)(Sanderson et al ., 2004 ; Maestre et al ., 2012a ), 因此本文也采用Maestre等(2012a, 2012b)的这一方法, 用土壤变量进行生态系统多功能性的评价。
在每个样方分别采集环刀样和原状土样, 同时按梅花形取样法用土钻取0-20 cm表层土混合样。在实验室测取与C、N、P三大物质循环和固存有关的指标: 土壤有机质(OM )、全碳(TC )、全氮(TN )、碱解氮(AN )、全磷(TP )和速效磷(AvP ) (Hector & Bagchi, 2007 ; 王明君等, 2007 ; 闫玉春等, 2009 ); 与土壤涵养水分有关的指标(Sanderson et al ., 2004 ): 土壤自然含水量(SMC )、土壤容重(BD )、毛管持水量(CMC ); 与土壤呼吸、养分转化和循环有关的指标: pH值、阳离子交换量(CEC ), C:N、总孔隙度(BP )、毛管孔隙度(CP )、非毛管孔隙度(NCP )、通气孔隙度(AP )等16个生产实践中简单易测但又对土壤功能体现较为全面的指标, 这些指标涉及土壤的水、肥、气、热等各方面和多个生态系统功能(王玉辉等, 2002 ; 张蕴薇等, 2002 ; Pei et al ., 2008 ; 王向涛等, 2010 ; 蒋德明等, 2013 )。尽管本文选取的指标并不全面, 但均是土壤调查中常采用的指标, 它们既能很好地反映土壤肥力状况、稳定性较高, 又易于调查和测定(王仁忠, 1998 ; 王玉辉等, 2002 ; 曹成有等, 2011 ; 陈慧颖等, 2014 ; 王忆慧等, 2015 )。
1.4 多功能性的量化
1.4.1 功能指标的筛选
土壤可以供给植物生长所需的营养, 影响植物的生长和分布; 不同地区、不同经营方式下的草原群落中, 对植物生长和分布起作用的土壤因子各不相同, 因此在最终确定多功能性指标的过程中需建立起群落植物分布和各土壤因子的联系, 这一联系通过以下多元分析完成。首先对植被调查中物种-样方矩阵进行除趋势对应分析(detrended correspondence analysis, DCA), 分析结果表明几个排序轴中最大的梯度长度(lengths of gradient)为3.149, 本文选用约束型排序中的线性模型冗余分析(redundance analysis, RDA)对物种-样方矩阵和土壤因子-样方矩阵进行综合分析, 分析结果可以很好地显示植物分布受土壤因子的影响状况, 同时也显示出16个功能指标中有几个对植物样方的分布没有显著影响(见2.2节), 据此本文对剩余的指标进行最终的多功能性量化。这一过程主要由CANOCO 4.5完成, 每个指标均进行蒙特卡罗(Monte Carlo)检验。
1.4.2 样方和样地尺度多功能性的评价
RDA的分析结果帮助我们完成多功能性量化指标的筛选过程, 对筛选出的有效功能指标进行数据标准化, 去除量纲和单位的不同, 继而进行Bartlett球形度检验(Bartlett’s Test of Sphericit), 经过检验后的数据经因子分析得到相关系数矩阵、特征值及贡献率和因子载荷矩阵, 由下列公式计算各样方和各样地的多功能性指数M:
M = ∑ai zi
zi = ∑wij xij
式中, ai 为各因子的方差贡献率, zi 为因子得分; wij 为第i 个变量在第j 个因子处的因子得分系数, xij 为第i 个变量在第j 个因子处的标准化值。以上各个指标的计算均由SASTISTICA 8.0完成。
同时按照Maestre等(2012a, 2012b)的方法, 计算出多功能性指数, 与本文的方法进行比较。每个样方中所有功能指标标准化后的Z-Scores的平均值即为该样方的多功能性指数M' (Maestre et al ., 2012a , 2012b ; Soliveres et al ., 2014 ; Valencia et al ., 2015 )。
1.5 生物多样性指数
分别计算样方和样地尺度的物种丰富度指数、Shannon-Wiener指数、Simpson指数和Evenness指数等多样性指数。
(1)物种丰富度指数: Margalef指数(Ma ), Ma = (S - 1) / lnN
(2) Simpson指数: D = 1 - ∑Pi 2
(3) Shannon-Wiener指数: H ′ = - ∑Pi lnPi
(4) 均匀度指数: Pielou指数(Jsw ), Jsw = H' /lnS
式中, S 为物种数, N 为群落中所有物种的总密度, Pi 为第i 个物种的相对多度。
运用单因素方差分析和Duncan差异显著性检验来分析各土壤指标和物种多样性指数在不同管理方式下的差异, 显著性水平设定为p = 0.05。用简单回归分析物种多样性与多功能性之间的关系, 并拟合最佳回归模型。
2 结果和分析
2.1 土壤功能指标和群落多样性指数
经过7年的围栏处理, 土壤pH值和TN 在3个群落中无显著差异, 除AvP 为放牧和刈割高于围封样地外, 刈割和围封土壤的各项功能指标均显著优于放牧样地(图1 ), 表现为有较高的有机质、TC 、TP 和AN 含量, 较好的土壤通气性(NCP、CP、AP 和BP 高), 较强的土壤持水(SMC 和CMC 高而BD 较低)和保肥能力(高的CEC 和适宜的C:N)。刈割和围封之间绝大多数功能指标没有显著差异, 唯有土壤自然含水量为围封样地高于刈割, 但刈割样地土壤中的TC 和C:N的含量却显著高于围封样地。与之对应的植物群落结构和多样性指数呈现同样的变化规律(表1 ), 放牧样地的个体密度、物种丰富度和生物多样性指数均显著低于刈割和围封样地, 后两者之间并没有显著差异, 但刈割样地的各项指数在数值上均高于围封。植物和土壤数据表明, 过度放牧严重破坏了土壤结构, 降低了生物多样性, 中度干扰的刈割管理虽然在生物量、种群高度与群落盖度等方面低于围封(另文发表), 但有助于维持生物多样性。
2.2 土壤功能指标对植物分布的影响
单一指标间的比较不能给予不同管理方式下草原群落多功能性的总体评价, 也不能判定具体样地中不同土壤指标对植物样方的影响作用。RDA的分析结果表明各土壤因子对植物分布有显著影响(p =0.0020, Monte Carlo test), 两个环境轴能解释66.3%的信息量, 其中第一轴主要由阳离子交换量、土壤自然持水量、毛管最大持水量、容重和速效磷等因子决定, 解释信息量高达51.4%, 表明在本实验区域内植物主要沿土壤水分和保肥性梯度分布。用于多功能性评价的16个土壤功能指标中, TC 、BP 和AP 3个指标的箭头连线最短, 且与植物样方的分布无显著的相关性(p > 0.05), 表明这3个指标对本实验区域的植物分布影响作用不大, 因此本文未采用这三个指标, 而是用其余指标进行多功能性的评价。
图1 刈割、围封、放牧三种利用方式下草原群落的土壤功能指标(平均值±标准误差)。相同小写字母表示数据间差异不显著。C, 刈割群落; E, 围封群落; G, 放牧群落。AN, 碱解氮; AP, 通气孔隙度; AvP, 速效磷; BD, 土壤容重; BP, 总孔隙度; CEC, 阳离子交换量; CMC, 毛管持水量; CP, 毛管孔隙度; NCP, 非毛管孔隙度; OM, 土壤有机质; SMC, 土壤含水量; TC, 全碳; TN, 全氮; TP, 全磷。
Fig. 1 Soil functional indexes in three communities with treatments of clipping, enclosure and grazing (mean ± SE ). Same lowercase letters indicate non-significant difference between different communities. C, clipping community; E, enclosure community; G, grazing community. AN, available N; AP, aeration porosity; AvP, available P; BD, soil bulk density; BP, bulk porosity; CEC, cation exchange capacity; CMC, capillary moisture capacity; CP, capillary porosity; NCP, non-capillary porosity; OM, organic matter; SMC, soil moisture content; TC, Total C; TN, Total N; TP, total P.
图2 植物样方与土壤因子的冗余分析。AN, 碱解氮; AP, 通气孔隙度; AvP, 速效磷; BD, 土壤容重; BP, 总孔隙度; CEC, 阳离子交换量; CMC, 毛管持水量; CP, 毛管孔隙度; NCP, 非毛管孔隙度; OM, 土壤有机质; SMC, 土壤含水量; TC, 全碳; TN, 全氮; TP, 全磷。
Fig. 2 The effect of soil factors on plant distribution patterns based on Redundance Analysis. AN, available N; AP, aeration porosity; AvP, available P; BD, soil bulk density; BP, bulk porosity; CEC, cation exchange capacity; CMC, capillary moisture capacity; CP, capillary porosity; NCP, non-capillary porosity; OM, organic matter; SMC, soil moisture content; TC, Total C; TN, Total N; TP, total P.
2.3 多功能性的量化
对筛选出的13个功能指标, 采用因子分析的方法完成多变量的降维和公因子的提取(Bartlett球形检验显著), 并计算各指标和公因子的相关系数矩阵、公因子特征值和贡献率及因子载荷矩阵(图3 )。依据特征值>1原则, 得到4个公因子, 4个公因子的特征值累计为10.69, 方差贡献率累计为82.22%。其中公因子1主要受TP、CEC、OM、CMC、SMC 和BD 支配, 公因子2主要受AN、TN 和CP 支配, 公因子3主要受C:N和TN 支配, 而公因子4单独受AvP 支配。
多功能性的综合量化结果为刈割样地(0.2178)>围封样地(0.0704)>放牧样地(-0.8031), 其中放牧样地因为最小的CP 和较高的TN 含量, 使其在公因子2上有较高的得分, 其余3个公因子上的得分均为最低; 围封样地有最低的AvP 含量, 致使其在公因子4上有最高的得分, 其在公因子1上的得分也较高, 但是由于AN 含量和C:N低于刈割样地、CP 高于刈割样地, 导致其在公因子2和3上的得分低于刈割样地; 而刈割样地在各公因子上的指标均较好, 故有最优的多功能性评价得分(表2 )。这一结果与不同群落间单一土壤指标的比较有很好的一致性, 可以相互印证, 但量化后的结果更为直观。
图3 因子特征值、方差贡献率和因子载荷。黑色柱子表示因子载荷>0.6或<-0.6。AN, 碱解氮; AP, 通气孔隙度; AvP, 速效磷; BD, 土壤容重; BP, 总孔隙度; CEC, 阳离子交换量; CMC, 毛管持水量; CP, 毛管孔隙度; NCP, 非毛管孔隙度; OM, 土壤有机质; SMC, 土壤含水量; TC, 全碳; TN, 全氮; TP, 全磷。
Fig. 3 The eigenvalues, percent of variance explained and factor loadings. Black bars indicate the factor loadings >0.6 or <-0.6. AN, available N; AP, aeration porosity; AvP, available P; BD, soil bulk density; BP, bulk porosity; CEC, cation exchange capacity; CMC, capillary moisture capacity; CP, capillary porosity; NCP, non-capillary porosity; OM, organic matter; SMC, soil moisture content; TC, total C; TN, total N; TP, total P.
2.4 多功能性与物种多样性之间的关系
三种不同利用方式下草原群落的多功能性与Shannon-Wiener指数和Simpson指数在样方尺度上有弱的正相关关系(R 2 = 0.0980, p = 0.0005; R 2 = 0.0805, p = 0.0009, 图表中未显示), 在样地尺度上均没有显著的相关关系(p > 0.05); 与之相比, 群落的多功能性指数(M )与均匀度指数和Margalef物种丰富度指数不仅在样方尺度上有较好的相关性 (R 2 = 0.1871, p < 0.0001; R 2 = 0.1601, p < 0.0001), 在样地尺度上的相关性更强(R 2 = 0.5921, p = 0.0093; R 2 = 0.7499, p = 0.0007)。群落的多功能性与均匀度指数在样方和样地尺度上均呈显著的正相关关系, 而与物种丰富度指数在样方和样地尺度上均呈单峰曲线关系(图4 ), 这一结果表明, 物种数目适中且分布均匀的群落可能拥有较好的多功能性。
图4 样方和样地尺度群落多功能性与均匀度和物种丰富度之间的关系。A、B, 样地尺度。C、D, 样方尺度。图中圆形表示围封样地, 三角形表示刈割样地, 菱形表示放牧样地。
Fig. 4 Relationships between multifunctionality and community evenness and Margalef index at quadrat and sites scale. A, B, site scale. C, D, quadrat scale. Diamonds represent grazing sites, circles represent enclosure sites and triangles represent clipping sites.
图5 样方和样地尺度上两种量化方法得到的多功能性指数。A, 样方尺度。B, 样地尺度。1), 本文方法得出的多功能性指数; 2), Maestre等(2012a, 2012b)对多变量的Z-scores简单平均得出的多功能性指数。菱形代表放牧样地, 圆形代表围封样地, 三角形代表刈割样地。
Fig. 5 Correlations between multifunctionality indexes calculated by two methods at quadrat and site scale. A, quadrat scale. B, site scale. 1), the multifunctionality index calculated by the author’method; 2), the multifunctionality index calculated by the method of Maestre et al. (2012a, 2012b). Diamonds represent grazing sites, circles represent enclosure sites and triangles represent clipping sites.
3 讨论
3.1 不同土地利用方式下的土壤功能指标和群落结构
放牧、刈割和围封是内蒙古草原最主要的三种利用方式, 分析三种方式下草原群落的物种多样性和生态系统功能变化对草原的可持续经营与管理有重要的指导意义。重度放牧的草地, 所有种群均受放牧压力的影响, 导致种群个体小型化, 生态位收缩(王炜等, 2000 ), 光合面积减少(Carrera et al ., 2008 ), 群落的盖度、高度和生产力下降(陈芙蓉等, 2013 ; 王忆慧等, 2015 ), 地表植被受到破坏, 地表裸露面积增大, 水分蒸散量增大(王玉辉等, 2002 )。牲畜的践踏作用又增大了土壤紧实度, 造成非毛管孔隙度的减少、增加了土壤容重, 也降低了毛管孔隙度和通气孔隙度, 使土壤的透气性、渗透力和蓄水能力降低(王玉辉等, 2002 ; 张蕴薇等, 2002 ; Carrera et al ., 2008 )。生产力下降也导致了凋落物的归还减少, 致使土壤中的有机质和氮含量减少(王玉辉等, 2002 ; 王明君等, 2007 ; Carrera et al ., 2008 ; Pei et al ., 2008 ), 伴随着有机质的减少, 土壤团聚体和地表结皮破坏(王向涛等, 2010 ), 使土壤黏粒减少, 砂粒增多, 进而导致植物-土壤界面无法保持平衡, 造成草地沙化(王明君等, 2007 ); 而N作为北方草原主要的限制因子, 土壤中N的减少反过来又会使群落光合作用等生态系统功能受到影响。另外, 本研究地中牛羊在晚间归宿, 伴随着牲畜排泄物在圈舍的累积和畜产品的产出, 各种营养元素被带离生态系统, 导致营养元素从土壤中流失(牛得草等, 2013 )。牲畜的选择性采食也使劣质牧草(N等营养含量较低、木质素等结构性碳较高)增多, 凋落物品质变差(较高的C:N和木质素:N)、分解速率降低, 抑制微生物活性, 使进入土壤中的有机质和营养元素进一步减少(王忆慧等, 2015 )。土壤结构的破坏、水分和营养元素的缺失导致重度放牧草地不再拥有适宜的群落微生境, 不仅不利于现有植物的生长发育(王炜等, 2000 ; 王玉辉等, 2002 ), 而且也不适宜机会物种的定居和拓殖, 所以导致重度放牧样地生物多样性的下降和土壤功能指标的恶化。因此与刈割和围封相比, 本研究中长期重度放牧的草地表现出以上各种退化现象; 群落的生物多样性显著降低(表1 ), 各土壤功能指标较差(图1 ), 表现出最低的多功能性评价指数(表2 )。
在退化草原的恢复过程中, 围封禁牧因投资少、见效快等优点被广泛应用(徐粒等, 2014 )。虽然短时间内, 围封可以显著提高草地物种多样性和生产力, 但围封时间过长则不利于群落维持较高的多样性和生产力水平(闫玉春等, 2009 ; 徐粒等, 2014 )。本研究中刈割和围封群落的各多样性指数之间虽然不存在统计上的显著差异, 但数值上刈割总是优于围封(表1 ), 说明经过7年或之后更长的时间, 刈割处理可能更有利于维持群落的生物多样性水平, 2015年的群落调查结果即为刈割的各多样性指标显著高于围封(另文发表)。这可能是因为随着围封时间的延长, 凋落物和根系周转向土壤输入更多的有机质和营养元素(闫玉春等, 2009 ; 杨浩等, 2009 ), 土壤结构得到改善, 土壤的持水能力得到增强, 生产力和养分积累得到增加(Bai et al ., 2007 ), 养分和水分的竞争转向光资源的竞争, 次优种的竞争性排除导致物种多样性的下降(Rajaniemi, 2002 ; Partel et al ., 2007 ; Hautier et al ., 2009 )。另外, 草地凋落物、枯枝层的增多也抑制了植物的再生和耐阴幼苗的形成, 同样不利于草原的繁殖更新和群落稳定(石红霄等, 2014 )。而反观刈割, 其适度的干扰不仅有利于植物的补偿性生长, 也有利于N、P等元素的矿化。
本文的研究结果还表明, 刈割和围封群落的多数土壤功能指标无显著差异, 都具有较好的容重、孔隙度指标和较高的有机质含量; 围封除有较高的土壤自然含水量外, TC 、C:N和AvP 含量均低于刈割群落(图1 )。这可能与凋落物的分解速度有关, 割草能够提高植物叶片的N含量, 使凋落物中C:N和木质素:N降低, 从而刺激微生物活性, 加速凋落物的分解和结构性碳的回归(Madritch & Cardinale, 2007 ; Walter et al ., 2013 ); 同时, 割草减少凋落物积累, 使地表增温, 有利于好氧细菌生长, 促使植被中的营养元素回归(陈慧颖等, 2014 )。与之相反, 围封群落中没有生物量的移出, 随着可溶性碳和养分的快速分解, 累积的凋落物和立枯物中有大量的结构性碳, 难分解物质的比重增加, 导致分解速率降低(Hossain & Sugiyama, 2008 ; 王忆慧等, 2015 ), 归还土壤中的营养物质减少。因此, 尽管刈割和围封的土壤指标大多没有显著差异, 但凋落物组成和分解速率的差异, 可能导致刈割群落有更高的养分循环效率, 使刈割群落的土壤有较高的TC 、C:N和其他营养元素含量, 所以刈割较之围封有更高的多功能性评价指数。
3.2 多功能性评价
由于地区不同, 生态系统的限制因子和影响植物分布的土壤因子也各不相同, 因此对多功能性的评价应适时适地进行, 常用的某些土壤指标也许在具体的实验地区对植物的影响并不明显。土壤-植被是相互作用、互为反馈的复合体(张静妮等, 2010 ; Delgado-Baquerizo et al ., 2013 ), 土壤系统的多功能性应该通过对植物的影响而体现。本研究中植物样方与环境因子的RDA分析表明, 土壤变量对植物分布有很好的解释效果, 尤其是CEC 、SMC 和BD 等综合作用的水肥轴(第一轴, 图2 ), 能够解释51.4%的信息量; 但其中TC、BP 和AP 等三个因子对植物分布没有显著影响, 因此针对本研究样地,本文舍弃这三个因子而对剩余的13个土壤因子进行多功能性指数的评价。
Maestre等人的多功能性指数(M' )只是评价指标的Z-scores的简单平均(Maestre et al ., 2012a ; 2012b ; Valencia et al ., 2015 ), 如果评价指标间有较高的相关性, 这一方法的评价结果会包含较多的冗余信息(Valencia et al ., 2015 ), 但因为相关性较高的指标数量相对于总体较多的评价指标来说是可以被接受的, 而且这种方法简单而直接, 所以被越来越多的学者接受和采用(Byrnes et al ., 2014 ; Soliveres et al ., 2014 ; Wagg et al ., 2014 ; Lefcheck et al ., 2015 ; Valencia et al ., 2015 )。本文采用因子分析的方法对筛选后的土壤指标进行多功能性指数的量化, 尽管这一方法与Maestre等人的方法有很好的相关性(图5 ), 尤其是在样地尺度上(R 2 = 0.9231, p < 0.0001); 但因子分析通过降维的方法可适当降低多变量间因为相关而造成的冗余信息, 而且本文作者也认为, 当选用的评价指标更多、样本量更大时, 这一方法可能更为适用。本文得出的公因子情形与RDA分析的结果相一致(图2 , 图3 ), 可以相互印证; 它在降低变量间冗余信息的同时, 也消除了Z-Scores简单平均这一方法默认每个功能指标在生态系统中有同等地位的不合理性。
3.3 生物多样性对多功能性的影响
由4个多样性指数与多功能性的相关性比较来看, 均匀度和物种丰富度指数对多功能性的影响要强于Shannon-Wiener指数和Simpson指数。而均匀度和物种丰富度指数对多功能性的影响均为样方尺度小于样地尺度, 表明均匀度和物种丰富度指数与多功能性之间的关系均有尺度依赖性。当然, 这一结果也不排除是由样本量的不同而造成, 这一问题会在未来研究中加以重视。本研究中物种的丰富度指数与生态系统的多功能性呈单峰曲线关系, 这一结果在许多研究单一功能与物种多样性关系的实验中并不鲜见(Waide et al ., 1999 ; Isbell et al ., 2009 ), 如生产力处于中等时物种丰富度最大(Kassen et al ., 2000 ; Partel et al ., 2007 ), 尽管生产力和物种丰富度之间的关系是不确定的(Adler et al ., 2011 ; Maestre et al ., 2012b ); 但最新的研究结果表明仅关注单一功能和物种间的关系低估了生物多样性对生态系统功能的重要性, 物种丰富的群落更能维持较高水平的生态系统多功能性(Zavaleta et al ., 2010 ; Lefcheck et al ., 2015 )。在已有的物种丰富度与多功能性的研究中, 由于一个物种不仅在生态系统的一个功能里起作用, 在其他的功能里可能同样起作用, 这些同时支持不同功能的物种重叠比例在0.2-0.5之间(Hector & Bagchi, 2007 ), 因此随着研究中涉及的生态系统功能数增多, 物种丰富度与之呈正的饱和关系(Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 )。这可以部分支持本文的单峰曲线模型, 但更深层的原因可能需要更大的样本从更多的尺度进行分析。Maestre等(2012b)对全球范围内224个干旱地区生态系统的物种丰富度和多功能性之间的关系进行了研究, 发现非生物因子中年平均气温和土壤的砂粒含量对多功能性预测能力最强(均与多功能性呈负相关), 但按照赤池信息准则(Akaka’s Information Criterion, AICc)从255个模型中选出的8个最优模型(解释信息量>55%)均包含物种丰富度这一变量, 表明物种丰富度本身虽然只能解释一小部分多功能性的变异, 但却是不可或缺的。由于物种均匀分布可以使资源的互补性利用更为充分, 而资源的互补性利用可使多功能性增加(Maestre et al ., 2012b ), 这可以解释本文中均匀度与多功能性指数的正相关关系。在Maestre等(2012a)的地衣微型生物土壤结皮模式实验中发现, 均匀度与物种丰富度的交互作用对多功能性有显著影响, 但单独与物种组成、空间分布格局和物种丰富度相比, 其对多功能性的相对重要性最小, Maestre等(2012a)认为这是因为二者的交互作用很大程度上受单个物种影响, 优势种数量与均匀度有倒数关系, 当一个优势种支配了某一特定的功能时, 均匀度就与这个功能呈负相关关系。本研究相对这种微宇宙实验尺度更大, 多功能性的影响因素更多, 也更为复杂; 而且三种土地利用方式下的草地群落中, 一些优势种还不足以对某些生态系统功能起决定作用; 尤其在放牧和刈割群落中, 按照胁迫梯度假说和生物多样性-生态系统功能理论(Hector et al ., 1999 ), 群落物种的正相互作用和资源的互补性利用可能仍起主导作用, 因此, 本研究中多功能性与物种均匀度呈正相关关系。
本文尝试引用生态系统多功能性这一概念对具体的草原群落进行评价, 并对现行主流的多功能性评价方法进行了改进, 使其更为科学合理。当然, 本研究与目前的绝大多数研究一样, 没有考虑不同营养级的物种多样性, 只是关注植物物种多样性, 尤其是缺乏对养分循环有重要驱动作用的微生物群落的考虑(Hector & Bagchi, 2007 ; Maestre et al ., 2012b )。另外, 指标的选取方面, 尽管土壤指标在很大程度上可以体现生态系统的多种功能, 例如保持水土、涵养水分和承载动植物, 并且土壤是生态系统的主要养分库(Sanderson et al ., 2004 ; Gamfeldt et al ., 2008 ; Valencia et al ., 2015 ), 但生态系统并不仅仅由土壤系统组成, 而是一定空间内共同栖居着的所有生物与其环境之间由于不断进行物质循环、能量流动和信息传递而形成的统一整体, 各个环境因子都是不可或缺的。生态系统的服务和功能是多方位的, 因此多功能性(multifunctionlity)的概念在未来仍需进一步明确和完善, 其量化方法也需进一步发展。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Adler PB Seabloom EW Borer ET Hillebrand H Hautier Y Hector A Harpole WS O'Halloran LR Grace JB Anderson TM Bakker JD Biederman LA Brown CS Buckley YM Calabrese LB Chu CJ Cleland EE Collins SL Cottingham KL Crawley MJ Damschen EI Davies KF DeCrappeo NM Fay PA Firn J Frater P Gasarch EI Gruner DS Hagenah N Hille Ris Lambers J Humphries H Jin VL Kay AD Kirkman KP Klein JA Knops JMH La Pierre KJ Lambrinos JG Li W MacDougall AS McCulley RL Melbourne BA Mitchell CE Moore JL Morgan JW Mortensen B Orrock JL Prober SM Pyke DA Risch AC Schuetz M Smith MD Stevens CJ Sullivan LL Wang G Wragg PD Wright JP Yang LH 2011 ). Productivity is a poor predictor of plant species richness.
Science , 333 , 1750 -1753 .
[本文引用: 1]
[2]
Bai Y Wu J Pan Q Huang J Wang Q Li F Buyantuyev A Han X 2007 ). Positive linear relationship between productivity and diversity: Evidence from the Eurasian Steppe.
Journal of Applied Ecology , 44 , 1023 -1034 .
[本文引用: 1]
[3]
Byrnes JEK Gamfeldt L Isbell F Lefcheck JS Griffin JN Hector A Cardinale BJ Hooper DU Dee LE Emmett Duffy J 2014 ). Investigating the relationship between biodiversity and ecosystem multifunctionality, challenges and solutions.
Methods in Ecology and Evolution , 5 , 111 -124 .
[本文引用: 3]
[4]
Cao CY Shao JF Jiang DM Cui ZB 2011 ). Effects of fence enclosure on soil nutrients and biological activities in highly degraded grasslands.
Journal of Northeastern University (Natural Science) , 32 , 427 -430 . (in Chinese with English abstract)[曹成有 , 邵建飞 , 蒋德明 , 崔振波 (2011 ). 围栏封育对重度退化草地土壤养分和生物活性的影响
. 东北大学学报(自然科学版) , 32 , 427 -430 .]
[5]
Cardinale BJ Duffy JE Gonzalez A Hooper DU Perrings C Venail P Narwani A Mace GM Tilman D Wardle DA Kinzig AP Daily GC Loreau M Grace JB Larigauderie A Srivastava DS Naeem S 2012 ). Biodiversity loss and its impact on humanity.
Nature , 486 , 59 -67 .
[本文引用: 1]
[6]
Carrera AL Bertiller MB Larreguy C 2008 ). Leaf litterfall, fine-root production, and decomposition in shrublands with different canopy structure induced by grazing in the Patagonian Monte, Argentina.
Plant and Soil , 311 , 39 -50 .
[本文引用: 1]
[7]
Chen FR Cheng JM Liu W Zhu RB Yang XM Zhao XY Su JS 2013 ). Effects of different disturbances on diversity and biomass of communities in the typical steppe of loess region.
Acta Ecologica Sinica , 33 , 2856 -2866 . (in Chinese with English abstract)[陈芙蓉 , 程积民 , 刘伟 , 朱仁斌 , 杨晓梅 , 赵新宇 , 苏纪帅 (2013 ).不同干扰对黄土区典型草原物种多样性和生物量的影响
.生态学报 , 33 , 2856 -2866 .]
[本文引用: 3]
[8]
Chen HY Zhang JH Huang YM Gong JR 2014 ). Traits related to carbon sequestration of common plant species in a Stipa grandis steppe in Nei Mongol under different land-uses.
Chinese Journal of Plant Ecology , 38 , 821 -832 . (in Chinese with English abstract)[陈慧颖 , 张景慧 , 黄永梅 , 龚吉蕊 (2014 ). 内蒙古大针茅草原常见植物在不同土地利用方式下的固碳相关属性
. 植物生态学报 , 38 , 821 -832 .]
[9]
Delgado-Baquerizo M Maestre FT Gallardo A Bowker MA Wallenstein MD Quero JL Ochoa V Gozalo B García-Gómez M Soliveres S García-Palacios P Berdugo M Valencia E Escolar C Arredondo T Barraza-Zepeda C Bran D Carreira JA Chaieb M Conceição AA Derak M Eldridge DJ Escudero A Espinosa CI Gaitán J Gatica MG Gómez-González S Guzman E Gutiérrez JR Florentino A Hepper E Hernández RM Huber-Sannwald E Jankju M Liu J Mau RL Miriti M Monerris J Naseri K Noumi Z Polo V Prina A Pucheta E Ramírez E Ramírez-Collantes DA Romão R Tighe M Torres D Torres-Díaz C Ungar ED Val J Wamiti W Wang D Zaady E 2013 ). Decoupling of soil nutrient cycles as a function of aridity in global drylands.
Nature , 502 , 672 -676 .
[本文引用: 1]
[10]
Díaz S Cabido M 2001 ). Vive la difference: Plant functional diversity matters to ecosystem processes.
Trends in Ecology & Evolution , 16 , 646 -655 .
[11]
Díaz S Lavorel S de Bello F Quetier F Grigulis K Robson TM 2007 ). Incorporating plant functional diversity effects in ecosystem service assessments.
Proceedings of the National Academy of Sciences of the United States of America , 104 , 20684 -20689 .
[本文引用: 2]
[12]
Gamfeldt L Hillebrand H Jonsson PR 2008 ). Multiple functions increase the importance of biodiversity for overall ecosystem functioning.
Ecology , 89 , 1223 -1231 .
[本文引用: 1]
[13]
Grime JP 1997 ). Biodiversity and ecosystem function: The debate deepens.
Science , 277 , 1260 -1261 .
[本文引用: 1]
[14]
Hautier Y Niklaus PA Hector A 2009 ). Competition for light causes plant biodiversity loss after eutrophication.
Science , 324 , 636 -638 .
[本文引用: 1]
[15]
Hector A Bagchi R 2007 ). Biodiversity and ecosystem multifunctionality.
Nature , 448 , 188 -190 .
[本文引用: 5]
[16]
Hector A Schmid B Beierkuhnlein C Caldeira MC Diemer M Dimitrakopoulos PG Finn JA Freitas H Giller PS Good J Harris R Hogberg P Huss-Danell K Joshi J Jumpponen A Korner C Leadley PW Loreau M Minns A Mulder C O'Donovan G Otway SJ Pereira JS Prinz A Read DJ Scherer-Lorenzen M Schulze ED Siamantziouras A Spehn EM Terry AC Troumbis AY Woodward FI Yachi S Lawton JH 1999 ). Plant diversity and productivity experiments in European grasslands.
Science , 286 , 1123 -1127 .
[本文引用: 2]
[17]
Hossain MZ Sugiyama S 2008 ). Effects of chemical composi- tion on the rate and temporal pattern of decomposition in grassland species leaf litter.
Grassland Science , 54 , 40 -44 .
[本文引用: 1]
[18]
Isbell FI Polley HW Wilsey BJ 2009 ). Biodiversity, productivity and the temporal stability of productivity, patterns and processes.
Ecology Letters , 12 , 443 -451 .
[本文引用: 7]
[19]
Jiang DM Miao RH Toshio OD Zhou QL 2013 ). Effects of fence enclosure on vegetation restoration and soil properties in Horqin Sandy Land.
Ecology and Environmental Sciences , 22 , 40 -46 . (in Chinese with English abstract)[蒋德明 , 苗仁辉 , 押田敏雄 , 周全来 (2013 ). 封育对科尔沁沙地植被恢复和土壤特性的影响
. 生态环境学报 , 22 , 40 -46 .]
[本文引用: 2]
[20]
Kassen R Buckling A Bell G Rainey PB 2000 ). Diversity peaks at intermediate productivity in a laboratory microcosm.
Nature , 406 , 508 -512 .
[本文引用: 1]
[21]
Lefcheck JS Byrnes JEK Isbell F Gamfeldt L Griffin JN Eisenhauer N Hensel MJS Hector A Cardinale BJ Duffy JE 2015 ). Biodiversity enhances ecosystem multifunc- tionality across trophic levels and habitats.
Nature Communications , 6 , 6936 .
[本文引用: 1]
[22]
MacDougall AS McCann KS Gellner G Turkington R 2013 ). Diversity loss with persistent human disturbance increases vulnerability to ecosystem collapse.
Nature , 494 , 86 -89 .
[23]
Madritch MD Cardinale BJ 2007 ). Impacts of tree species diversity on litter decomposition in northern temperate forests of Wisconsin, USA: A multi-site experiment along a latitudinal gradient.
Plant and Soil , 292 , 147 -159 .
[本文引用: 1]
[24]
Maestre FT Castillo-Monroy AP Bowker MA Ochoa-Hueso R 2012 a). Species richness effects on ecosystem multifunctionality depend on evenness, composition and spatial pattern.
Journal of Ecology , 100 , 317 -330 .
[本文引用: 5]
[25]
Maestre FT Quero JL Gotelli NJ Escudero A Ochoa V Delgado-Baquerizo M Garcia-Gomez M Bowker MA Soliveres S Escolar C Garcia-Palacios P Berdugo M Valencia E Gozalo B Gallardo A Aguilera L Arredondo T Blones J Boeken B Bran D Conceicao AA Cabrera O Chaieb M Derak M Eldridge DJ Espinosa CI Floren- tino A Gaitan J Gatica MG Ghiloufi W Gomez- Gonzalez S Gutierrez JR Hernandez RM Huang X Huber-Sannwald E Jankju M Miriti M Monerris J Mau RL Morici E Naseri K Ospina A Polo V Prina A Puch- eta E Ramirez-Collantes DA Romao R Tighe M Torres-Diaz C Val J Veiga JP Wang D Zaady E 2012 b). Plant species richness and ecosystem multifunctionality in global drylands.
Science , 335 , 214 -218 .
[本文引用: 11]
[26]
Niu DC Jiang SG Qin Y Zhang BL Cao GT Fu H 2013 ). Effects of grazing and fencing on soil microorganisms and enzymes activities.
Partacultural Science , 30 , 528 -534 . (in Chinese with English abstract)[牛得草 , 江世高 , 秦燕 , 张宝林 , 曹格图 , 傅华 (2013 ). 围封与放牧对土壤微生物和酶活性的影响
. 草业科学 , 30 , 528 -534 .]
[本文引用: 1]
[27]
Partel M Laanisto L Zobel M 2007 ). Contrasting plant productivity-diversity relationships across latitude, the role of evolutionary history.
Ecology , 88 , 1091 -1097 .
[本文引用: 1]
[28]
Pei S Fu H Wan C 2008 ). Changes in soil properties and vegetation following exclosure and grazing in degraded Alxa desert steppe of Inner Mongolia, China. Agriculture,
Ecosystems and Environment , 124 , 33 -39 .
[29]
Rajaniemi TK 2002 ). Why does fertilization reduce plant species diversity? Testing three competition-based hypotheses.
Journal of Ecology , 90 , 316 -324 .
[30]
Sanderson MA Skinner RH Barker DJ Edwards GR Tracy BF Wedin DA 2004 ). Plant species diversity and management of temperate forage and grazing land ecosystems.
Crop Science , 44 , 1132 -1144 .
[31]
Shi HX Fan YJ Hou XY Yang YP Wu XH Yang TT Li P 2014 ). Analysis of plant community characteristics of Kobresia pygmaea meadow in the three headwaters under fencing and grazing.
Chinese Journal of Grassland , 36 (3 ), 67 -72 . (in Chinese with English abstract)[石红霄 , 范月君 , 侯向阳 , 杨玉平 , 吴新宏 , 杨婷婷 , 李鹏 (2014 ). 三江源区围栏与放牧高山嵩草草甸植物群落特征分析
. 中国草地学报 , 36 (3 ), 67 -72 .]
[本文引用: 1]
[32]
Simova I Li YM Storch D 2013 ). Relationship between species richness and productivity in plants: The role of sampling effect, heterogeneity and species pool.
Journal of Ecology , 101 , 161 -170 .
[本文引用: 2]
[33]
Soliveres S Maestre FT Eldridge DJ Delgado-Baquerizo M Quero JL Bowker MA Gallardo A 2014 ). Plant diversity and ecosystem multifunctionality peak at intermediate levels of woody cover in global drylands.
Global Ecology and Biogeography , 23 , 1408 -1416 .
[本文引用: 3]
[34]
Tilman D Knops J Wedin D Reich P Ritchie M Siemann E 1997 ). The influence of functional diversity and compos- ition on ecosystem processes.
Science , 277 , 1300 -1302 .
[本文引用: 1]
[35]
Valencia E Maestre FT Le Bagousse-Pinguet Y Quero JL Tamme R Borger L Garcia-Gomez M Gross N 2015 ). Functional diversity enhances the resistance of ecosystem multifunctionality to aridity in Mediterranean drylands.
New Phytologist , 206 , 660 -71 .
[本文引用: 6]
[36]
Wagg C Bender SF Widmer F van der Heijden MGA 2014 ). Soil biodiversity and soil community composition determine ecosystem multifunctionality.
Proceedings of the National Academy of Sciences of the United States of America , 111 , 5266 -5270 .
[37]
Waide RB Willig MR Steiner CF Mittelbach G Gough L Dodson SI Juday GP Parmenter R 1999 ). The relationship between productivity and species richness.
Annual Review of Ecology and Systematics , 30 , 257 -300 .
[本文引用: 1]
[38]
Walter J Hein R Beierkuhnlein C Hammerl V Jentsch A Schädler M Schuerings J Kreyling J 2013 ). Combined effects of multifactor climate change and land-use on decomposition in temperate grassland.
Soil Biology & Biochemistry , 60 , 10 -18 .
[本文引用: 1]
[39]
Wang MJ Han GD Zhao ML Chen HJ Wang Z Hao XL Bo T 2007 ). The effects of different grazing intensity on soil organic carbon content in meadow steppe.
Partacultural Science , 24 (10 ), 6 -10 . (in Chinese with English abstract)[王明君 , 韩国栋 , 赵萌莉 , 陈海军 , 王珍 , 郝晓莉 , 薄涛 (2007 ). 草甸草原不同放牧强度对土壤有机碳含量的影响
. 草业科学 , 24 (10 ), 6 -10 .]
[本文引用: 4]
[40]
Wang RZ 1998 ). A study on the effects of grazing and mowing disturbances in Leymus chinensis grassland in Songnen Plain.
Acta Ecologica Sinica , 18 , 100 -103 . (in Chinese with English abstract)[王仁忠 (1998 ). 放牧和刈割干扰对松嫩草原羊草草地影响的研究
. 生态学报 , 18 , 100 -103 .]
[本文引用: 1]
[41]
Wang W Liang CZ Liu ZL Hao DY 2000 ). Analysis of the plant individual behaviour during the degradation and restoring succession in steppe community.
Acta Phytoecologica Sinica , 24 , 268 -274 . (in Chinese with English abstract)[王炜 , 梁存柱 , 刘钟龄 , 郝敦元 (2000 ). 草原群落退化与恢复演替中的植物个体行为分析
. 植物生态学报 , 24 , 268 -274 .]
[本文引用: 8]
[42]
Wang XT Zhang SH Chen DD Tan YR Sun DS Du GZ 2010 ). The effects of natural grazing intensity on plant community and soil nutrients in alpine meadow.
Acta Agrestia Sinica , 18 , 510 -516 . (in Chinese with English abstract)[王向涛 , 张世虎 , 陈懂懂 , 谈嫣蓉 , 孙大帅 , 杜国祯 (2010 ). 不同放牧强度下高寒草甸植被特征和土壤养分变化研究
. 草地学报 , 18 , 510 -516 .]
[本文引用: 3]
[43]
Wang YH Gong JR Liu M Huang YM Yan X Zhang ZY Xu S Luo QP 2015 ). Effects of grassland-use on soil respiration and litter decomposition.
Chinese Journal of Plant Ecology , 39 , 239 -248 . (in Chinese with English abstract)[王忆慧 , 龚吉蕊 , 刘敏 , 黄永梅 , 晏欣 , 张梓瑜 , 徐沙 , 罗亲普 (2015 ). 草地利用方式对土壤呼吸和凋落物分解的影响
. 植物生态学报 , 39 , 239 -248 .]
[本文引用: 2]
[44]
Wang YH He XY Zhou GS 2002 ). Study on the Responses of Leymus chinensis steppe to grazing in songnen plain.
Acta Agrestia Sinica , 10 (1 ), 45 -49 . (in Chinese with English abstract)[王玉辉 , 何兴元 , 周广胜 (2002 ). 放牧强度对羊草草原的影响
. 草地学报 , 10 (1 ), 45 -49 .]
[本文引用: 1]
[45]
Xu L Gao Q Wang YL 2014 ). Species richness within a six-year slope exclosure in a temperate grassland and its relationship with aboveground biomass.
Ecology and Environmental Sciences , 23 , 398 -405 . (in Chinese with English abstract)[徐粒 , 高琼 , 王亚林 (2014 ). 围封6年对温带典型草原坡地物种多样性及其与地上生物量的关系的影响
. 生态环境学报 , 23 , 398 -405 .]
[46]
Yan YC Tang HP Xin XP Wang X 2009 ). Advances in research on the effects of exclosure on grasslands.
Acta Ecologica Sinica , 29 , 5039 -5046 . (in Chinese with English abstract)[闫玉春 , 唐海萍 , 辛晓平 , 王旭 (2009 ). 围封对草地的影响研究进展
. 生态学报 , 29 , 5039 -5046 .]
[本文引用: 3]
[47]
Yang H Bai YF Li YH Han XG 2009 ). Response of plant species composition and community structure to long-term grazing in typical steppe of Inner Mongolia.
Chinese Journal of Plant Ecology , 33 , 499 -507 . (in Chinese with English abstract)[杨浩 , 白永飞 , 李永宏 , 韩兴国 (2009 ). 内蒙古典型草原物种组成和群落结构对长期放牧的响应
. 植物生态学报 , 33 , 499 -507 .]
[48]
Zavaleta ES Pasari JR Hulvey KB Tilman GD 2010 ). Sustaining multiple ecosystem functions in grassland communities requires higher biodiversity.
Proceedings of the National Academy of Sciences of the United States of America , 107 , 1443 -1446 .
[本文引用: 2]
[49]
Zhang JN Lai X Li G Zhao JN Zhang YS Yang DL 2010 ). Response of plant diversity and soil nutrient condition to grazing disturbance in Stipa baicalensis Roshev. grassland.
Acta Agrestia Sinica , 18 , 177 -182 . (in Chinese with English abstract)[张静妮 , 赖欣 , 李刚 , 赵建宁 , 张永生 , 杨殿林 (2010 ). 贝加尔针茅草原植物多样性及土壤养分对放牧干扰的响应
. 草地学报 , 18 , 177 -182 .]
[50]
Zhang YW Han JG Li ZQ 2002 ). A study of the effects of different grazing intensities on soil physical properties.
Acta Agrestia Sinica , 10 , 74 -78 . (in Chinese with English abstract)[张蕴薇 , 韩建国 , 李志强 (2002 ). 放牧强度对土壤物理性质的影响
. 草地学报 , 10 , 74 -78 .]
[本文引用: 2]
Productivity is a poor predictor of plant species richness.
1
2011
... 由4个多样性指数与多功能性的相关性比较来看, 均匀度和物种丰富度指数对多功能性的影响要强于Shannon-Wiener指数和Simpson指数.而均匀度和物种丰富度指数对多功能性的影响均为样方尺度小于样地尺度, 表明均匀度和物种丰富度指数与多功能性之间的关系均有尺度依赖性.当然, 这一结果也不排除是由样本量的不同而造成, 这一问题会在未来研究中加以重视.本研究中物种的丰富度指数与生态系统的多功能性呈单峰曲线关系, 这一结果在许多研究单一功能与物种多样性关系的实验中并不鲜见(Waide et al ., 1999 ; Isbell et al ., 2009 ), 如生产力处于中等时物种丰富度最大(Kassen et al ., 2000 ; Partel et al ., 2007 ), 尽管生产力和物种丰富度之间的关系是不确定的(Adler et al ., 2011 ; Maestre et al ., 2012b ); 但最新的研究结果表明仅关注单一功能和物种间的关系低估了生物多样性对生态系统功能的重要性, 物种丰富的群落更能维持较高水平的生态系统多功能性(Zavaleta et al ., 2010 ; Lefcheck et al ., 2015 ).在已有的物种丰富度与多功能性的研究中, 由于一个物种不仅在生态系统的一个功能里起作用, 在其他的功能里可能同样起作用, 这些同时支持不同功能的物种重叠比例在0.2-0.5之间(Hector & Bagchi, 2007 ), 因此随着研究中涉及的生态系统功能数增多, 物种丰富度与之呈正的饱和关系(Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ).这可以部分支持本文的单峰曲线模型, 但更深层的原因可能需要更大的样本从更多的尺度进行分析.Maestre等(2012b)对全球范围内224个干旱地区生态系统的物种丰富度和多功能性之间的关系进行了研究, 发现非生物因子中年平均气温和土壤的砂粒含量对多功能性预测能力最强(均与多功能性呈负相关), 但按照赤池信息准则(Akaka’s Information Criterion, AICc)从255个模型中选出的8个最优模型(解释信息量>55%)均包含物种丰富度这一变量, 表明物种丰富度本身虽然只能解释一小部分多功能性的变异, 但却是不可或缺的.由于物种均匀分布可以使资源的互补性利用更为充分, 而资源的互补性利用可使多功能性增加(Maestre et al ., 2012b ), 这可以解释本文中均匀度与多功能性指数的正相关关系.在Maestre等(2012a)的地衣微型生物土壤结皮模式实验中发现, 均匀度与物种丰富度的交互作用对多功能性有显著影响, 但单独与物种组成、空间分布格局和物种丰富度相比, 其对多功能性的相对重要性最小, Maestre等(2012a)认为这是因为二者的交互作用很大程度上受单个物种影响, 优势种数量与均匀度有倒数关系, 当一个优势种支配了某一特定的功能时, 均匀度就与这个功能呈负相关关系.本研究相对这种微宇宙实验尺度更大, 多功能性的影响因素更多, 也更为复杂; 而且三种土地利用方式下的草地群落中, 一些优势种还不足以对某些生态系统功能起决定作用; 尤其在放牧和刈割群落中, 按照胁迫梯度假说和生物多样性-生态系统功能理论(Hector et al ., 1999 ), 群落物种的正相互作用和资源的互补性利用可能仍起主导作用, 因此, 本研究中多功能性与物种均匀度呈正相关关系. ...
Positive linear relationship between productivity and diversity: Evidence from the Eurasian Steppe.
1
2007
... 在退化草原的恢复过程中, 围封禁牧因投资少、见效快等优点被广泛应用(徐粒等, 2014 ).虽然短时间内, 围封可以显著提高草地物种多样性和生产力, 但围封时间过长则不利于群落维持较高的多样性和生产力水平(闫玉春等, 2009 ; 徐粒等, 2014 ).本研究中刈割和围封群落的各多样性指数之间虽然不存在统计上的显著差异, 但数值上刈割总是优于围封(表1 ), 说明经过7年或之后更长的时间, 刈割处理可能更有利于维持群落的生物多样性水平, 2015年的群落调查结果即为刈割的各多样性指标显著高于围封(另文发表).这可能是因为随着围封时间的延长, 凋落物和根系周转向土壤输入更多的有机质和营养元素(闫玉春等, 2009 ; 杨浩等, 2009 ), 土壤结构得到改善, 土壤的持水能力得到增强, 生产力和养分积累得到增加(Bai et al ., 2007 ), 养分和水分的竞争转向光资源的竞争, 次优种的竞争性排除导致物种多样性的下降(Rajaniemi, 2002 ; Partel et al ., 2007 ; Hautier et al ., 2009 ).另外, 草地凋落物、枯枝层的增多也抑制了植物的再生和耐阴幼苗的形成, 同样不利于草原的繁殖更新和群落稳定(石红霄等, 2014 ).而反观刈割, 其适度的干扰不仅有利于植物的补偿性生长, 也有利于N、P等元素的矿化. ...
Investigating the relationship between biodiversity and ecosystem multifunctionality, challenges and solutions.
3
2014
... “multifunctionality”一词首先由Sanderson等(2004)提出, 他们认为经营者在管理牧场和草场生态系统时往往只关注初级生产力的提高(因为生产输出与经济效益直接相关), 而忽略了生态系统的其他价值(如环境和美学效益), 指出从可持续发展的角度出发, 生态系统同时执行多种功能的能力“multifunctionality”更应该被重视, 并从环境效益和以人类为中心的经济效益两个角度出发提供了体现multifunctionality的15个功能特征.Hector和Bagchi (2007)引用“multifunctionality”这一概念在《Nature》上发表文章, 认为生态系统的多种服务或过程就是“ecosystem ‘multifunctionality’ ”, 并第一次量化研究了物种多样性同时对8个联合的生态系统过程所起的作用.Gamfeldt等(2008)、Zavaleta等(2010)又引用Hector和Bagchi (2007)的这篇文章, 分别在《Ecology》和《PNAS》上发文探讨和量化生物多样性对维持高水平“multifunctionality”的重要性, 认为“multifunctionality”即为生态系统同时维持多个生态功能或服务的能力.自此, 随着引用的增多, “multifunctionality”一词开始被人们广泛认可并正式使用.Maestre等(2012b)在《Science》上发表文章, 应用14个土壤变量(分别体现C、N、P循环三个生态系统过程)来综合评价全球干旱地区的生态系统多功能性, 其选用的指标和评价方法几乎成为近年来生态系统多功能性研究的标准方法(Maestre et al ., 2012a ; Byrnes et al ., 2014 ; Soliveres et al ., 2014 ; Wagg et al ., 2014 ; Valencia et al ., 2015 ).随后, Maestre等(2012a)在《Journal of Ecology》上, Soli- veres等(2014)在《Global Ecology and Biogeography》上、Wagg等(2014)在《PANS》上发表的多功能性研究论文均采用了Maestre等的文章的土壤变量和评价方法, 最新由Valencia等(2015)发表在《New Phytologist》上的文章同样引用了这些土壤变量及其评价方法, 这些文章的发表, 标志着生态系统多功能性(multifunctionality)概念正在逐步完善, 其评价方法也趋于统一. ...
... 虽然Maestre等的多功能性(multifunctionality)评价指标和量化方法被许多学者所采用, 但也存在一些争议(Byrnes et al ., 2014 ; Lefcheck et al ., 2015 ), 本文笔者认为这一方法中对每个样方多功能性指数(M )的计算只是14个评价指标的Z-scores简单平均, 并没有考虑各个指标间的权重, 显然默认每个指标有同等的地位和作用并不符合自然生态系统的真实情况.另外, Maestre等(2012b)选择的14个土壤指标涉及C、N、P的循环与固存, 尽管C、N、P这三大物质循环在生态系统中占据着重要的地位, 但土壤同时也有涵养水分、平衡和缓冲外界干扰、为微生物提供稳定的环境、为各种物理生化反应提供介质和可持续地供给植物营养等多重功能(王仁忠, 1998 ; 王玉辉等, 2002 ; Sanderson et al ., 2004 ; Pei et al ., 2008 ), 仅用C、N、P的循环与固存这三个功能来体现生态系统的“多”功能性, 不免略显勉强.因此本文用改进的量化方法, 尝试选取农业实践中较为常见、测定更为简易的土壤指标, 并结合具体的立地条件, 对内蒙古三种不同利用方式下的草原群落进行多功能性的评价, 探讨物种多样性与多功能性指数之间的关系, 试图达到以下目的: (1)结合立地条件, 选取更为合适的土壤多功能性评价指标; (2)与Maestre等人的评价方法相对比, 建立更为科学的多功能性指数评价方法; (3)评价不同土地利用方式下草原生态系统的多功能性, 探讨物种多样性对多功能性的影响. ...
... Maestre等人的多功能性指数(M' )只是评价指标的Z-scores的简单平均(Maestre et al ., 2012a ; 2012b ; Valencia et al ., 2015 ), 如果评价指标间有较高的相关性, 这一方法的评价结果会包含较多的冗余信息(Valencia et al ., 2015 ), 但因为相关性较高的指标数量相对于总体较多的评价指标来说是可以被接受的, 而且这种方法简单而直接, 所以被越来越多的学者接受和采用(Byrnes et al ., 2014 ; Soliveres et al ., 2014 ; Wagg et al ., 2014 ; Lefcheck et al ., 2015 ; Valencia et al ., 2015 ).本文采用因子分析的方法对筛选后的土壤指标进行多功能性指数的量化, 尽管这一方法与Maestre等人的方法有很好的相关性(图5 ), 尤其是在样地尺度上(R 2 = 0.9231, p < 0.0001); 但因子分析通过降维的方法可适当降低多变量间因为相关而造成的冗余信息, 而且本文作者也认为, 当选用的评价指标更多、样本量更大时, 这一方法可能更为适用.本文得出的公因子情形与RDA分析的结果相一致(图2 , 图3 ), 可以相互印证; 它在降低变量间冗余信息的同时, 也消除了Z-Scores简单平均这一方法默认每个功能指标在生态系统中有同等地位的不合理性. ...
围栏封育对重度退化草地土壤养分和生物活性的影响
2011
Biodiversity loss and its impact on humanity.
1
2012
... 在每个样方分别采集环刀样和原状土样, 同时按梅花形取样法用土钻取0-20 cm表层土混合样.在实验室测取与C、N、P三大物质循环和固存有关的指标: 土壤有机质(OM )、全碳(TC )、全氮(TN )、碱解氮(AN )、全磷(TP )和速效磷(AvP ) (Hector & Bagchi, 2007 ; 王明君等, 2007 ; 闫玉春等, 2009 ); 与土壤涵养水分有关的指标(Sanderson et al ., 2004 ): 土壤自然含水量(SMC )、土壤容重(BD )、毛管持水量(CMC ); 与土壤呼吸、养分转化和循环有关的指标: pH值、阳离子交换量(CEC ), C:N、总孔隙度(BP )、毛管孔隙度(CP )、非毛管孔隙度(NCP )、通气孔隙度(AP )等16个生产实践中简单易测但又对土壤功能体现较为全面的指标, 这些指标涉及土壤的水、肥、气、热等各方面和多个生态系统功能(王玉辉等, 2002 ; 张蕴薇等, 2002 ; Pei et al ., 2008 ; 王向涛等, 2010 ; 蒋德明等, 2013 ).尽管本文选取的指标并不全面, 但均是土壤调查中常采用的指标, 它们既能很好地反映土壤肥力状况、稳定性较高, 又易于调查和测定(王仁忠, 1998 ; 王玉辉等, 2002 ; 曹成有等, 2011 ; 陈慧颖等, 2014 ; 王忆慧等, 2015 ). ...
Leaf litterfall, fine-root production, and decomposition in shrublands with different canopy structure induced by grazing in the Patagonian Monte, Argentina.
1
2008
... 全球变化在全世界范围内以前所未有的速度影响着生物多样性的变化, 进而使自然生态系统功能产生不可预知的改变(Grime, 1997 ; Cardinale et al ., 2012 ; MacDougall et al ., 2013 ; Valencia et al ., 2015 ), 尤其是随着物种消失速率的加快, 越来越多的学者开始关注物种多样性的改变和由之而引起的群落结构变化对生态系统功能的影响(Grime, 1997 ; Tilman et al ., 1997 ; Díaz & Cabido, 2001 ; Díaz et al ., 2007 ; Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ).20多年来, 许多研究工作围绕这一问题展开, 但这些工作大多仅关注物种多样性和群落结构与单一或少数生态系统功能之间的关系(Hector et al ., 1999 ; Waide et al ., 1999 ; Simova et al ., 2013 ; Valencia et al ., 2015 ), 对生态系统多种功能同时评价和研究的工作还不多见, 而生态系统最为重要的价值是同时提供多种功能和服务的能力(Sanderson et al ., 2004 ; Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ; Soliveres et al ., 2014 ), 因此物种多样性与生态系统多功能性(multifunctionality)之间关系的研究显得十分 重要. ...
不同干扰对黄土区典型草原物种多样性和生物量的影响
3
2013
... 放牧、刈割和围封是内蒙古草原最主要的三种利用方式, 分析三种方式下草原群落的物种多样性和生态系统功能变化对草原的可持续经营与管理有重要的指导意义.重度放牧的草地, 所有种群均受放牧压力的影响, 导致种群个体小型化, 生态位收缩(王炜等, 2000 ), 光合面积减少(Carrera et al ., 2008 ), 群落的盖度、高度和生产力下降(陈芙蓉等, 2013 ; 王忆慧等, 2015 ), 地表植被受到破坏, 地表裸露面积增大, 水分蒸散量增大(王玉辉等, 2002 ).牲畜的践踏作用又增大了土壤紧实度, 造成非毛管孔隙度的减少、增加了土壤容重, 也降低了毛管孔隙度和通气孔隙度, 使土壤的透气性、渗透力和蓄水能力降低(王玉辉等, 2002 ; 张蕴薇等, 2002 ; Carrera et al ., 2008 ).生产力下降也导致了凋落物的归还减少, 致使土壤中的有机质和氮含量减少(王玉辉等, 2002 ; 王明君等, 2007 ; Carrera et al ., 2008 ; Pei et al ., 2008 ), 伴随着有机质的减少, 土壤团聚体和地表结皮破坏(王向涛等, 2010 ), 使土壤黏粒减少, 砂粒增多, 进而导致植物-土壤界面无法保持平衡, 造成草地沙化(王明君等, 2007 ); 而N作为北方草原主要的限制因子, 土壤中N的减少反过来又会使群落光合作用等生态系统功能受到影响.另外, 本研究地中牛羊在晚间归宿, 伴随着牲畜排泄物在圈舍的累积和畜产品的产出, 各种营养元素被带离生态系统, 导致营养元素从土壤中流失(牛得草等, 2013 ).牲畜的选择性采食也使劣质牧草(N等营养含量较低、木质素等结构性碳较高)增多, 凋落物品质变差(较高的C:N和木质素:N)、分解速率降低, 抑制微生物活性, 使进入土壤中的有机质和营养元素进一步减少(王忆慧等, 2015 ).土壤结构的破坏、水分和营养元素的缺失导致重度放牧草地不再拥有适宜的群落微生境, 不仅不利于现有植物的生长发育(王炜等, 2000 ; 王玉辉等, 2002 ), 而且也不适宜机会物种的定居和拓殖, 所以导致重度放牧样地生物多样性的下降和土壤功能指标的恶化.因此与刈割和围封相比, 本研究中长期重度放牧的草地表现出以上各种退化现象; 群落的生物多样性显著降低(表1 ), 各土壤功能指标较差(图1 ), 表现出最低的多功能性评价指数(表2 ). ...
... ; Carrera et al ., 2008 ).生产力下降也导致了凋落物的归还减少, 致使土壤中的有机质和氮含量减少(王玉辉等, 2002 ; 王明君等, 2007 ; Carrera et al ., 2008 ; Pei et al ., 2008 ), 伴随着有机质的减少, 土壤团聚体和地表结皮破坏(王向涛等, 2010 ), 使土壤黏粒减少, 砂粒增多, 进而导致植物-土壤界面无法保持平衡, 造成草地沙化(王明君等, 2007 ); 而N作为北方草原主要的限制因子, 土壤中N的减少反过来又会使群落光合作用等生态系统功能受到影响.另外, 本研究地中牛羊在晚间归宿, 伴随着牲畜排泄物在圈舍的累积和畜产品的产出, 各种营养元素被带离生态系统, 导致营养元素从土壤中流失(牛得草等, 2013 ).牲畜的选择性采食也使劣质牧草(N等营养含量较低、木质素等结构性碳较高)增多, 凋落物品质变差(较高的C:N和木质素:N)、分解速率降低, 抑制微生物活性, 使进入土壤中的有机质和营养元素进一步减少(王忆慧等, 2015 ).土壤结构的破坏、水分和营养元素的缺失导致重度放牧草地不再拥有适宜的群落微生境, 不仅不利于现有植物的生长发育(王炜等, 2000 ; 王玉辉等, 2002 ), 而且也不适宜机会物种的定居和拓殖, 所以导致重度放牧样地生物多样性的下降和土壤功能指标的恶化.因此与刈割和围封相比, 本研究中长期重度放牧的草地表现出以上各种退化现象; 群落的生物多样性显著降低(表1 ), 各土壤功能指标较差(图1 ), 表现出最低的多功能性评价指数(表2 ). ...
... ; Carrera et al ., 2008 ; Pei et al ., 2008 ), 伴随着有机质的减少, 土壤团聚体和地表结皮破坏(王向涛等, 2010 ), 使土壤黏粒减少, 砂粒增多, 进而导致植物-土壤界面无法保持平衡, 造成草地沙化(王明君等, 2007 ); 而N作为北方草原主要的限制因子, 土壤中N的减少反过来又会使群落光合作用等生态系统功能受到影响.另外, 本研究地中牛羊在晚间归宿, 伴随着牲畜排泄物在圈舍的累积和畜产品的产出, 各种营养元素被带离生态系统, 导致营养元素从土壤中流失(牛得草等, 2013 ).牲畜的选择性采食也使劣质牧草(N等营养含量较低、木质素等结构性碳较高)增多, 凋落物品质变差(较高的C:N和木质素:N)、分解速率降低, 抑制微生物活性, 使进入土壤中的有机质和营养元素进一步减少(王忆慧等, 2015 ).土壤结构的破坏、水分和营养元素的缺失导致重度放牧草地不再拥有适宜的群落微生境, 不仅不利于现有植物的生长发育(王炜等, 2000 ; 王玉辉等, 2002 ), 而且也不适宜机会物种的定居和拓殖, 所以导致重度放牧样地生物多样性的下降和土壤功能指标的恶化.因此与刈割和围封相比, 本研究中长期重度放牧的草地表现出以上各种退化现象; 群落的生物多样性显著降低(表1 ), 各土壤功能指标较差(图1 ), 表现出最低的多功能性评价指数(表2 ). ...
内蒙古大针茅草原常见植物在不同土地利用方式下的固碳相关属性
2014
Decoupling of soil nutrient cycles as a function of aridity in global drylands.
1
2013
... 放牧、刈割和围封是内蒙古草原最主要的三种利用方式, 分析三种方式下草原群落的物种多样性和生态系统功能变化对草原的可持续经营与管理有重要的指导意义.重度放牧的草地, 所有种群均受放牧压力的影响, 导致种群个体小型化, 生态位收缩(王炜等, 2000 ), 光合面积减少(Carrera et al ., 2008 ), 群落的盖度、高度和生产力下降(陈芙蓉等, 2013 ; 王忆慧等, 2015 ), 地表植被受到破坏, 地表裸露面积增大, 水分蒸散量增大(王玉辉等, 2002 ).牲畜的践踏作用又增大了土壤紧实度, 造成非毛管孔隙度的减少、增加了土壤容重, 也降低了毛管孔隙度和通气孔隙度, 使土壤的透气性、渗透力和蓄水能力降低(王玉辉等, 2002 ; 张蕴薇等, 2002 ; Carrera et al ., 2008 ).生产力下降也导致了凋落物的归还减少, 致使土壤中的有机质和氮含量减少(王玉辉等, 2002 ; 王明君等, 2007 ; Carrera et al ., 2008 ; Pei et al ., 2008 ), 伴随着有机质的减少, 土壤团聚体和地表结皮破坏(王向涛等, 2010 ), 使土壤黏粒减少, 砂粒增多, 进而导致植物-土壤界面无法保持平衡, 造成草地沙化(王明君等, 2007 ); 而N作为北方草原主要的限制因子, 土壤中N的减少反过来又会使群落光合作用等生态系统功能受到影响.另外, 本研究地中牛羊在晚间归宿, 伴随着牲畜排泄物在圈舍的累积和畜产品的产出, 各种营养元素被带离生态系统, 导致营养元素从土壤中流失(牛得草等, 2013 ).牲畜的选择性采食也使劣质牧草(N等营养含量较低、木质素等结构性碳较高)增多, 凋落物品质变差(较高的C:N和木质素:N)、分解速率降低, 抑制微生物活性, 使进入土壤中的有机质和营养元素进一步减少(王忆慧等, 2015 ).土壤结构的破坏、水分和营养元素的缺失导致重度放牧草地不再拥有适宜的群落微生境, 不仅不利于现有植物的生长发育(王炜等, 2000 ; 王玉辉等, 2002 ), 而且也不适宜机会物种的定居和拓殖, 所以导致重度放牧样地生物多样性的下降和土壤功能指标的恶化.因此与刈割和围封相比, 本研究中长期重度放牧的草地表现出以上各种退化现象; 群落的生物多样性显著降低(表1 ), 各土壤功能指标较差(图1 ), 表现出最低的多功能性评价指数(表2 ). ...
Vive la difference: Plant functional diversity matters to ecosystem processes.
2001
Incorporating plant functional diversity effects in ecosystem service assessments.
2
2007
... 在每个样方分别采集环刀样和原状土样, 同时按梅花形取样法用土钻取0-20 cm表层土混合样.在实验室测取与C、N、P三大物质循环和固存有关的指标: 土壤有机质(OM )、全碳(TC )、全氮(TN )、碱解氮(AN )、全磷(TP )和速效磷(AvP ) (Hector & Bagchi, 2007 ; 王明君等, 2007 ; 闫玉春等, 2009 ); 与土壤涵养水分有关的指标(Sanderson et al ., 2004 ): 土壤自然含水量(SMC )、土壤容重(BD )、毛管持水量(CMC ); 与土壤呼吸、养分转化和循环有关的指标: pH值、阳离子交换量(CEC ), C:N、总孔隙度(BP )、毛管孔隙度(CP )、非毛管孔隙度(NCP )、通气孔隙度(AP )等16个生产实践中简单易测但又对土壤功能体现较为全面的指标, 这些指标涉及土壤的水、肥、气、热等各方面和多个生态系统功能(王玉辉等, 2002 ; 张蕴薇等, 2002 ; Pei et al ., 2008 ; 王向涛等, 2010 ; 蒋德明等, 2013 ).尽管本文选取的指标并不全面, 但均是土壤调查中常采用的指标, 它们既能很好地反映土壤肥力状况、稳定性较高, 又易于调查和测定(王仁忠, 1998 ; 王玉辉等, 2002 ; 曹成有等, 2011 ; 陈慧颖等, 2014 ; 王忆慧等, 2015 ). ...
... 本文的研究结果还表明, 刈割和围封群落的多数土壤功能指标无显著差异, 都具有较好的容重、孔隙度指标和较高的有机质含量; 围封除有较高的土壤自然含水量外, TC 、C:N和AvP 含量均低于刈割群落(图1 ).这可能与凋落物的分解速度有关, 割草能够提高植物叶片的N含量, 使凋落物中C:N和木质素:N降低, 从而刺激微生物活性, 加速凋落物的分解和结构性碳的回归(Madritch & Cardinale, 2007 ; Walter et al ., 2013 ); 同时, 割草减少凋落物积累, 使地表增温, 有利于好氧细菌生长, 促使植被中的营养元素回归(陈慧颖等, 2014 ).与之相反, 围封群落中没有生物量的移出, 随着可溶性碳和养分的快速分解, 累积的凋落物和立枯物中有大量的结构性碳, 难分解物质的比重增加, 导致分解速率降低(Hossain & Sugiyama, 2008 ; 王忆慧等, 2015 ), 归还土壤中的营养物质减少.因此, 尽管刈割和围封的土壤指标大多没有显著差异, 但凋落物组成和分解速率的差异, 可能导致刈割群落有更高的养分循环效率, 使刈割群落的土壤有较高的TC 、C:N和其他营养元素含量, 所以刈割较之围封有更高的多功能性评价指数. ...
Multiple functions increase the importance of biodiversity for overall ecosystem functioning.
1
2008
... 由于地区不同, 生态系统的限制因子和影响植物分布的土壤因子也各不相同, 因此对多功能性的评价应适时适地进行, 常用的某些土壤指标也许在具体的实验地区对植物的影响并不明显.土壤-植被是相互作用、互为反馈的复合体(张静妮等, 2010 ; Delgado-Baquerizo et al ., 2013 ), 土壤系统的多功能性应该通过对植物的影响而体现.本研究中植物样方与环境因子的RDA分析表明, 土壤变量对植物分布有很好的解释效果, 尤其是CEC 、SMC 和BD 等综合作用的水肥轴(第一轴, 图2 ), 能够解释51.4%的信息量; 但其中TC、BP 和AP 等三个因子对植物分布没有显著影响, 因此针对本研究样地,本文舍弃这三个因子而对剩余的13个土壤因子进行多功能性指数的评价. ...
Biodiversity and ecosystem function: The debate deepens.
1
1997
... 全球变化在全世界范围内以前所未有的速度影响着生物多样性的变化, 进而使自然生态系统功能产生不可预知的改变(Grime, 1997 ; Cardinale et al ., 2012 ; MacDougall et al ., 2013 ; Valencia et al ., 2015 ), 尤其是随着物种消失速率的加快, 越来越多的学者开始关注物种多样性的改变和由之而引起的群落结构变化对生态系统功能的影响(Grime, 1997 ; Tilman et al ., 1997 ; Díaz & Cabido, 2001 ; Díaz et al ., 2007 ; Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ).20多年来, 许多研究工作围绕这一问题展开, 但这些工作大多仅关注物种多样性和群落结构与单一或少数生态系统功能之间的关系(Hector et al ., 1999 ; Waide et al ., 1999 ; Simova et al ., 2013 ; Valencia et al ., 2015 ), 对生态系统多种功能同时评价和研究的工作还不多见, 而生态系统最为重要的价值是同时提供多种功能和服务的能力(Sanderson et al ., 2004 ; Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ; Soliveres et al ., 2014 ), 因此物种多样性与生态系统多功能性(multifunctionality)之间关系的研究显得十分 重要. ...
Competition for light causes plant biodiversity loss after eutrophication.
1
2009
... 全球变化在全世界范围内以前所未有的速度影响着生物多样性的变化, 进而使自然生态系统功能产生不可预知的改变(Grime, 1997 ; Cardinale et al ., 2012 ; MacDougall et al ., 2013 ; Valencia et al ., 2015 ), 尤其是随着物种消失速率的加快, 越来越多的学者开始关注物种多样性的改变和由之而引起的群落结构变化对生态系统功能的影响(Grime, 1997 ; Tilman et al ., 1997 ; Díaz & Cabido, 2001 ; Díaz et al ., 2007 ; Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ).20多年来, 许多研究工作围绕这一问题展开, 但这些工作大多仅关注物种多样性和群落结构与单一或少数生态系统功能之间的关系(Hector et al ., 1999 ; Waide et al ., 1999 ; Simova et al ., 2013 ; Valencia et al ., 2015 ), 对生态系统多种功能同时评价和研究的工作还不多见, 而生态系统最为重要的价值是同时提供多种功能和服务的能力(Sanderson et al ., 2004 ; Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ; Soliveres et al ., 2014 ), 因此物种多样性与生态系统多功能性(multifunctionality)之间关系的研究显得十分 重要. ...
Biodiversity and ecosystem multifunctionality.
5
2007
... 全球变化在全世界范围内以前所未有的速度影响着生物多样性的变化, 进而使自然生态系统功能产生不可预知的改变(Grime, 1997 ; Cardinale et al ., 2012 ; MacDougall et al ., 2013 ; Valencia et al ., 2015 ), 尤其是随着物种消失速率的加快, 越来越多的学者开始关注物种多样性的改变和由之而引起的群落结构变化对生态系统功能的影响(Grime, 1997 ; Tilman et al ., 1997 ; Díaz & Cabido, 2001 ; Díaz et al ., 2007 ; Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ).20多年来, 许多研究工作围绕这一问题展开, 但这些工作大多仅关注物种多样性和群落结构与单一或少数生态系统功能之间的关系(Hector et al ., 1999 ; Waide et al ., 1999 ; Simova et al ., 2013 ; Valencia et al ., 2015 ), 对生态系统多种功能同时评价和研究的工作还不多见, 而生态系统最为重要的价值是同时提供多种功能和服务的能力(Sanderson et al ., 2004 ; Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ; Soliveres et al ., 2014 ), 因此物种多样性与生态系统多功能性(multifunctionality)之间关系的研究显得十分 重要. ...
... ; Gamfeldt et al ., 2008 ; Soliveres et al ., 2014 ), 因此物种多样性与生态系统多功能性(multifunctionality)之间关系的研究显得十分 重要. ...
... 尽管按照最初的概念, 多功能性是指生态系统同时维持各种生态系统功能和服务的能力, 其评价指标应涉及经济、环境、景观美学等各方面的指标(Sanderson et al ., 2004 ), 但随着多功能性概念的发展, 狭义的概念被人们提出, 同时维持多个生态功能或服务的能力即可称为“多功能性”, 这便导致不同的研究中各研究者选用的指标和数量并不相同, 涉及的生态系统功能和数量也各不相同(Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ; Zavaleta et al ., 2010 ; Maestre et al ., 2012a , 2012b ; Wagg et al ., 2014 ; Lefcheck et al ., 2015 ).近年来多功能性的研究中, 评价指标的选取多参考Maestre等(2012a, 2012b)的研究, 由于土壤-植被是相互依赖、互为反馈的复合体, 并且土壤是生态系统的养分库和种子库, 土壤指标很大程度上能反映生态系统的多个功能(如C、N、P的固存, 水土保持, 承载野生动植物等)(Sanderson et al ., 2004 ; Maestre et al ., 2012a ), 因此本文也采用Maestre等(2012a, 2012b)的这一方法, 用土壤变量进行生态系统多功能性的评价. ...
... 由4个多样性指数与多功能性的相关性比较来看, 均匀度和物种丰富度指数对多功能性的影响要强于Shannon-Wiener指数和Simpson指数.而均匀度和物种丰富度指数对多功能性的影响均为样方尺度小于样地尺度, 表明均匀度和物种丰富度指数与多功能性之间的关系均有尺度依赖性.当然, 这一结果也不排除是由样本量的不同而造成, 这一问题会在未来研究中加以重视.本研究中物种的丰富度指数与生态系统的多功能性呈单峰曲线关系, 这一结果在许多研究单一功能与物种多样性关系的实验中并不鲜见(Waide et al ., 1999 ; Isbell et al ., 2009 ), 如生产力处于中等时物种丰富度最大(Kassen et al ., 2000 ; Partel et al ., 2007 ), 尽管生产力和物种丰富度之间的关系是不确定的(Adler et al ., 2011 ; Maestre et al ., 2012b ); 但最新的研究结果表明仅关注单一功能和物种间的关系低估了生物多样性对生态系统功能的重要性, 物种丰富的群落更能维持较高水平的生态系统多功能性(Zavaleta et al ., 2010 ; Lefcheck et al ., 2015 ).在已有的物种丰富度与多功能性的研究中, 由于一个物种不仅在生态系统的一个功能里起作用, 在其他的功能里可能同样起作用, 这些同时支持不同功能的物种重叠比例在0.2-0.5之间(Hector & Bagchi, 2007 ), 因此随着研究中涉及的生态系统功能数增多, 物种丰富度与之呈正的饱和关系(Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ).这可以部分支持本文的单峰曲线模型, 但更深层的原因可能需要更大的样本从更多的尺度进行分析.Maestre等(2012b)对全球范围内224个干旱地区生态系统的物种丰富度和多功能性之间的关系进行了研究, 发现非生物因子中年平均气温和土壤的砂粒含量对多功能性预测能力最强(均与多功能性呈负相关), 但按照赤池信息准则(Akaka’s Information Criterion, AICc)从255个模型中选出的8个最优模型(解释信息量>55%)均包含物种丰富度这一变量, 表明物种丰富度本身虽然只能解释一小部分多功能性的变异, 但却是不可或缺的.由于物种均匀分布可以使资源的互补性利用更为充分, 而资源的互补性利用可使多功能性增加(Maestre et al ., 2012b ), 这可以解释本文中均匀度与多功能性指数的正相关关系.在Maestre等(2012a)的地衣微型生物土壤结皮模式实验中发现, 均匀度与物种丰富度的交互作用对多功能性有显著影响, 但单独与物种组成、空间分布格局和物种丰富度相比, 其对多功能性的相对重要性最小, Maestre等(2012a)认为这是因为二者的交互作用很大程度上受单个物种影响, 优势种数量与均匀度有倒数关系, 当一个优势种支配了某一特定的功能时, 均匀度就与这个功能呈负相关关系.本研究相对这种微宇宙实验尺度更大, 多功能性的影响因素更多, 也更为复杂; 而且三种土地利用方式下的草地群落中, 一些优势种还不足以对某些生态系统功能起决定作用; 尤其在放牧和刈割群落中, 按照胁迫梯度假说和生物多样性-生态系统功能理论(Hector et al ., 1999 ), 群落物种的正相互作用和资源的互补性利用可能仍起主导作用, 因此, 本研究中多功能性与物种均匀度呈正相关关系. ...
... 本文尝试引用生态系统多功能性这一概念对具体的草原群落进行评价, 并对现行主流的多功能性评价方法进行了改进, 使其更为科学合理.当然, 本研究与目前的绝大多数研究一样, 没有考虑不同营养级的物种多样性, 只是关注植物物种多样性, 尤其是缺乏对养分循环有重要驱动作用的微生物群落的考虑(Hector & Bagchi, 2007 ; Maestre et al ., 2012b ).另外, 指标的选取方面, 尽管土壤指标在很大程度上可以体现生态系统的多种功能, 例如保持水土、涵养水分和承载动植物, 并且土壤是生态系统的主要养分库(Sanderson et al ., 2004 ; Gamfeldt et al ., 2008 ; Valencia et al ., 2015 ), 但生态系统并不仅仅由土壤系统组成, 而是一定空间内共同栖居着的所有生物与其环境之间由于不断进行物质循环、能量流动和信息传递而形成的统一整体, 各个环境因子都是不可或缺的.生态系统的服务和功能是多方位的, 因此多功能性(multifunctionlity)的概念在未来仍需进一步明确和完善, 其量化方法也需进一步发展. ...
Plant diversity and productivity experiments in European grasslands.
2
1999
... 全球变化在全世界范围内以前所未有的速度影响着生物多样性的变化, 进而使自然生态系统功能产生不可预知的改变(Grime, 1997 ; Cardinale et al ., 2012 ; MacDougall et al ., 2013 ; Valencia et al ., 2015 ), 尤其是随着物种消失速率的加快, 越来越多的学者开始关注物种多样性的改变和由之而引起的群落结构变化对生态系统功能的影响(Grime, 1997 ; Tilman et al ., 1997 ; Díaz & Cabido, 2001 ; Díaz et al ., 2007 ; Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ).20多年来, 许多研究工作围绕这一问题展开, 但这些工作大多仅关注物种多样性和群落结构与单一或少数生态系统功能之间的关系(Hector et al ., 1999 ; Waide et al ., 1999 ; Simova et al ., 2013 ; Valencia et al ., 2015 ), 对生态系统多种功能同时评价和研究的工作还不多见, 而生态系统最为重要的价值是同时提供多种功能和服务的能力(Sanderson et al ., 2004 ; Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ; Soliveres et al ., 2014 ), 因此物种多样性与生态系统多功能性(multifunctionality)之间关系的研究显得十分 重要. ...
... ), 尤其是随着物种消失速率的加快, 越来越多的学者开始关注物种多样性的改变和由之而引起的群落结构变化对生态系统功能的影响(Grime, 1997 ; Tilman et al ., 1997 ; Díaz & Cabido, 2001 ; Díaz et al ., 2007 ; Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ).20多年来, 许多研究工作围绕这一问题展开, 但这些工作大多仅关注物种多样性和群落结构与单一或少数生态系统功能之间的关系(Hector et al ., 1999 ; Waide et al ., 1999 ; Simova et al ., 2013 ; Valencia et al ., 2015 ), 对生态系统多种功能同时评价和研究的工作还不多见, 而生态系统最为重要的价值是同时提供多种功能和服务的能力(Sanderson et al ., 2004 ; Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ; Soliveres et al ., 2014 ), 因此物种多样性与生态系统多功能性(multifunctionality)之间关系的研究显得十分 重要. ...
Effects of chemical composi- tion on the rate and temporal pattern of decomposition in grassland species leaf litter.
1
2008
... 在退化草原的恢复过程中, 围封禁牧因投资少、见效快等优点被广泛应用(徐粒等, 2014 ).虽然短时间内, 围封可以显著提高草地物种多样性和生产力, 但围封时间过长则不利于群落维持较高的多样性和生产力水平(闫玉春等, 2009 ; 徐粒等, 2014 ).本研究中刈割和围封群落的各多样性指数之间虽然不存在统计上的显著差异, 但数值上刈割总是优于围封(表1 ), 说明经过7年或之后更长的时间, 刈割处理可能更有利于维持群落的生物多样性水平, 2015年的群落调查结果即为刈割的各多样性指标显著高于围封(另文发表).这可能是因为随着围封时间的延长, 凋落物和根系周转向土壤输入更多的有机质和营养元素(闫玉春等, 2009 ; 杨浩等, 2009 ), 土壤结构得到改善, 土壤的持水能力得到增强, 生产力和养分积累得到增加(Bai et al ., 2007 ), 养分和水分的竞争转向光资源的竞争, 次优种的竞争性排除导致物种多样性的下降(Rajaniemi, 2002 ; Partel et al ., 2007 ; Hautier et al ., 2009 ).另外, 草地凋落物、枯枝层的增多也抑制了植物的再生和耐阴幼苗的形成, 同样不利于草原的繁殖更新和群落稳定(石红霄等, 2014 ).而反观刈割, 其适度的干扰不仅有利于植物的补偿性生长, 也有利于N、P等元素的矿化. ...
Biodiversity, productivity and the temporal stability of productivity, patterns and processes.
7
2009
... 全球变化在全世界范围内以前所未有的速度影响着生物多样性的变化, 进而使自然生态系统功能产生不可预知的改变(Grime, 1997 ; Cardinale et al ., 2012 ; MacDougall et al ., 2013 ; Valencia et al ., 2015 ), 尤其是随着物种消失速率的加快, 越来越多的学者开始关注物种多样性的改变和由之而引起的群落结构变化对生态系统功能的影响(Grime, 1997 ; Tilman et al ., 1997 ; Díaz & Cabido, 2001 ; Díaz et al ., 2007 ; Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ).20多年来, 许多研究工作围绕这一问题展开, 但这些工作大多仅关注物种多样性和群落结构与单一或少数生态系统功能之间的关系(Hector et al ., 1999 ; Waide et al ., 1999 ; Simova et al ., 2013 ; Valencia et al ., 2015 ), 对生态系统多种功能同时评价和研究的工作还不多见, 而生态系统最为重要的价值是同时提供多种功能和服务的能力(Sanderson et al ., 2004 ; Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ; Soliveres et al ., 2014 ), 因此物种多样性与生态系统多功能性(multifunctionality)之间关系的研究显得十分 重要. ...
... ; Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ; Soliveres et al ., 2014 ), 因此物种多样性与生态系统多功能性(multifunctionality)之间关系的研究显得十分 重要. ...
... 尽管按照最初的概念, 多功能性是指生态系统同时维持各种生态系统功能和服务的能力, 其评价指标应涉及经济、环境、景观美学等各方面的指标(Sanderson et al ., 2004 ), 但随着多功能性概念的发展, 狭义的概念被人们提出, 同时维持多个生态功能或服务的能力即可称为“多功能性”, 这便导致不同的研究中各研究者选用的指标和数量并不相同, 涉及的生态系统功能和数量也各不相同(Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ; Zavaleta et al ., 2010 ; Maestre et al ., 2012a , 2012b ; Wagg et al ., 2014 ; Lefcheck et al ., 2015 ).近年来多功能性的研究中, 评价指标的选取多参考Maestre等(2012a, 2012b)的研究, 由于土壤-植被是相互依赖、互为反馈的复合体, 并且土壤是生态系统的养分库和种子库, 土壤指标很大程度上能反映生态系统的多个功能(如C、N、P的固存, 水土保持, 承载野生动植物等)(Sanderson et al ., 2004 ; Maestre et al ., 2012a ), 因此本文也采用Maestre等(2012a, 2012b)的这一方法, 用土壤变量进行生态系统多功能性的评价. ...
... 在每个样方分别采集环刀样和原状土样, 同时按梅花形取样法用土钻取0-20 cm表层土混合样.在实验室测取与C、N、P三大物质循环和固存有关的指标: 土壤有机质(OM )、全碳(TC )、全氮(TN )、碱解氮(AN )、全磷(TP )和速效磷(AvP ) (Hector & Bagchi, 2007 ; 王明君等, 2007 ; 闫玉春等, 2009 ); 与土壤涵养水分有关的指标(Sanderson et al ., 2004 ): 土壤自然含水量(SMC )、土壤容重(BD )、毛管持水量(CMC ); 与土壤呼吸、养分转化和循环有关的指标: pH值、阳离子交换量(CEC ), C:N、总孔隙度(BP )、毛管孔隙度(CP )、非毛管孔隙度(NCP )、通气孔隙度(AP )等16个生产实践中简单易测但又对土壤功能体现较为全面的指标, 这些指标涉及土壤的水、肥、气、热等各方面和多个生态系统功能(王玉辉等, 2002 ; 张蕴薇等, 2002 ; Pei et al ., 2008 ; 王向涛等, 2010 ; 蒋德明等, 2013 ).尽管本文选取的指标并不全面, 但均是土壤调查中常采用的指标, 它们既能很好地反映土壤肥力状况、稳定性较高, 又易于调查和测定(王仁忠, 1998 ; 王玉辉等, 2002 ; 曹成有等, 2011 ; 陈慧颖等, 2014 ; 王忆慧等, 2015 ). ...
... 由4个多样性指数与多功能性的相关性比较来看, 均匀度和物种丰富度指数对多功能性的影响要强于Shannon-Wiener指数和Simpson指数.而均匀度和物种丰富度指数对多功能性的影响均为样方尺度小于样地尺度, 表明均匀度和物种丰富度指数与多功能性之间的关系均有尺度依赖性.当然, 这一结果也不排除是由样本量的不同而造成, 这一问题会在未来研究中加以重视.本研究中物种的丰富度指数与生态系统的多功能性呈单峰曲线关系, 这一结果在许多研究单一功能与物种多样性关系的实验中并不鲜见(Waide et al ., 1999 ; Isbell et al ., 2009 ), 如生产力处于中等时物种丰富度最大(Kassen et al ., 2000 ; Partel et al ., 2007 ), 尽管生产力和物种丰富度之间的关系是不确定的(Adler et al ., 2011 ; Maestre et al ., 2012b ); 但最新的研究结果表明仅关注单一功能和物种间的关系低估了生物多样性对生态系统功能的重要性, 物种丰富的群落更能维持较高水平的生态系统多功能性(Zavaleta et al ., 2010 ; Lefcheck et al ., 2015 ).在已有的物种丰富度与多功能性的研究中, 由于一个物种不仅在生态系统的一个功能里起作用, 在其他的功能里可能同样起作用, 这些同时支持不同功能的物种重叠比例在0.2-0.5之间(Hector & Bagchi, 2007 ), 因此随着研究中涉及的生态系统功能数增多, 物种丰富度与之呈正的饱和关系(Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ).这可以部分支持本文的单峰曲线模型, 但更深层的原因可能需要更大的样本从更多的尺度进行分析.Maestre等(2012b)对全球范围内224个干旱地区生态系统的物种丰富度和多功能性之间的关系进行了研究, 发现非生物因子中年平均气温和土壤的砂粒含量对多功能性预测能力最强(均与多功能性呈负相关), 但按照赤池信息准则(Akaka’s Information Criterion, AICc)从255个模型中选出的8个最优模型(解释信息量>55%)均包含物种丰富度这一变量, 表明物种丰富度本身虽然只能解释一小部分多功能性的变异, 但却是不可或缺的.由于物种均匀分布可以使资源的互补性利用更为充分, 而资源的互补性利用可使多功能性增加(Maestre et al ., 2012b ), 这可以解释本文中均匀度与多功能性指数的正相关关系.在Maestre等(2012a)的地衣微型生物土壤结皮模式实验中发现, 均匀度与物种丰富度的交互作用对多功能性有显著影响, 但单独与物种组成、空间分布格局和物种丰富度相比, 其对多功能性的相对重要性最小, Maestre等(2012a)认为这是因为二者的交互作用很大程度上受单个物种影响, 优势种数量与均匀度有倒数关系, 当一个优势种支配了某一特定的功能时, 均匀度就与这个功能呈负相关关系.本研究相对这种微宇宙实验尺度更大, 多功能性的影响因素更多, 也更为复杂; 而且三种土地利用方式下的草地群落中, 一些优势种还不足以对某些生态系统功能起决定作用; 尤其在放牧和刈割群落中, 按照胁迫梯度假说和生物多样性-生态系统功能理论(Hector et al ., 1999 ), 群落物种的正相互作用和资源的互补性利用可能仍起主导作用, 因此, 本研究中多功能性与物种均匀度呈正相关关系. ...
... ), 因此随着研究中涉及的生态系统功能数增多, 物种丰富度与之呈正的饱和关系(Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ).这可以部分支持本文的单峰曲线模型, 但更深层的原因可能需要更大的样本从更多的尺度进行分析.Maestre等(2012b)对全球范围内224个干旱地区生态系统的物种丰富度和多功能性之间的关系进行了研究, 发现非生物因子中年平均气温和土壤的砂粒含量对多功能性预测能力最强(均与多功能性呈负相关), 但按照赤池信息准则(Akaka’s Information Criterion, AICc)从255个模型中选出的8个最优模型(解释信息量>55%)均包含物种丰富度这一变量, 表明物种丰富度本身虽然只能解释一小部分多功能性的变异, 但却是不可或缺的.由于物种均匀分布可以使资源的互补性利用更为充分, 而资源的互补性利用可使多功能性增加(Maestre et al ., 2012b ), 这可以解释本文中均匀度与多功能性指数的正相关关系.在Maestre等(2012a)的地衣微型生物土壤结皮模式实验中发现, 均匀度与物种丰富度的交互作用对多功能性有显著影响, 但单独与物种组成、空间分布格局和物种丰富度相比, 其对多功能性的相对重要性最小, Maestre等(2012a)认为这是因为二者的交互作用很大程度上受单个物种影响, 优势种数量与均匀度有倒数关系, 当一个优势种支配了某一特定的功能时, 均匀度就与这个功能呈负相关关系.本研究相对这种微宇宙实验尺度更大, 多功能性的影响因素更多, 也更为复杂; 而且三种土地利用方式下的草地群落中, 一些优势种还不足以对某些生态系统功能起决定作用; 尤其在放牧和刈割群落中, 按照胁迫梯度假说和生物多样性-生态系统功能理论(Hector et al ., 1999 ), 群落物种的正相互作用和资源的互补性利用可能仍起主导作用, 因此, 本研究中多功能性与物种均匀度呈正相关关系. ...
... 本文尝试引用生态系统多功能性这一概念对具体的草原群落进行评价, 并对现行主流的多功能性评价方法进行了改进, 使其更为科学合理.当然, 本研究与目前的绝大多数研究一样, 没有考虑不同营养级的物种多样性, 只是关注植物物种多样性, 尤其是缺乏对养分循环有重要驱动作用的微生物群落的考虑(Hector & Bagchi, 2007 ; Maestre et al ., 2012b ).另外, 指标的选取方面, 尽管土壤指标在很大程度上可以体现生态系统的多种功能, 例如保持水土、涵养水分和承载动植物, 并且土壤是生态系统的主要养分库(Sanderson et al ., 2004 ; Gamfeldt et al ., 2008 ; Valencia et al ., 2015 ), 但生态系统并不仅仅由土壤系统组成, 而是一定空间内共同栖居着的所有生物与其环境之间由于不断进行物质循环、能量流动和信息传递而形成的统一整体, 各个环境因子都是不可或缺的.生态系统的服务和功能是多方位的, 因此多功能性(multifunctionlity)的概念在未来仍需进一步明确和完善, 其量化方法也需进一步发展. ...
封育对科尔沁沙地植被恢复和土壤特性的影响
2
2013
... 全球变化在全世界范围内以前所未有的速度影响着生物多样性的变化, 进而使自然生态系统功能产生不可预知的改变(Grime, 1997 ; Cardinale et al ., 2012 ; MacDougall et al ., 2013 ; Valencia et al ., 2015 ), 尤其是随着物种消失速率的加快, 越来越多的学者开始关注物种多样性的改变和由之而引起的群落结构变化对生态系统功能的影响(Grime, 1997 ; Tilman et al ., 1997 ; Díaz & Cabido, 2001 ; Díaz et al ., 2007 ; Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ).20多年来, 许多研究工作围绕这一问题展开, 但这些工作大多仅关注物种多样性和群落结构与单一或少数生态系统功能之间的关系(Hector et al ., 1999 ; Waide et al ., 1999 ; Simova et al ., 2013 ; Valencia et al ., 2015 ), 对生态系统多种功能同时评价和研究的工作还不多见, 而生态系统最为重要的价值是同时提供多种功能和服务的能力(Sanderson et al ., 2004 ; Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ; Soliveres et al ., 2014 ), 因此物种多样性与生态系统多功能性(multifunctionality)之间关系的研究显得十分 重要. ...
... 由4个多样性指数与多功能性的相关性比较来看, 均匀度和物种丰富度指数对多功能性的影响要强于Shannon-Wiener指数和Simpson指数.而均匀度和物种丰富度指数对多功能性的影响均为样方尺度小于样地尺度, 表明均匀度和物种丰富度指数与多功能性之间的关系均有尺度依赖性.当然, 这一结果也不排除是由样本量的不同而造成, 这一问题会在未来研究中加以重视.本研究中物种的丰富度指数与生态系统的多功能性呈单峰曲线关系, 这一结果在许多研究单一功能与物种多样性关系的实验中并不鲜见(Waide et al ., 1999 ; Isbell et al ., 2009 ), 如生产力处于中等时物种丰富度最大(Kassen et al ., 2000 ; Partel et al ., 2007 ), 尽管生产力和物种丰富度之间的关系是不确定的(Adler et al ., 2011 ; Maestre et al ., 2012b ); 但最新的研究结果表明仅关注单一功能和物种间的关系低估了生物多样性对生态系统功能的重要性, 物种丰富的群落更能维持较高水平的生态系统多功能性(Zavaleta et al ., 2010 ; Lefcheck et al ., 2015 ).在已有的物种丰富度与多功能性的研究中, 由于一个物种不仅在生态系统的一个功能里起作用, 在其他的功能里可能同样起作用, 这些同时支持不同功能的物种重叠比例在0.2-0.5之间(Hector & Bagchi, 2007 ), 因此随着研究中涉及的生态系统功能数增多, 物种丰富度与之呈正的饱和关系(Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ).这可以部分支持本文的单峰曲线模型, 但更深层的原因可能需要更大的样本从更多的尺度进行分析.Maestre等(2012b)对全球范围内224个干旱地区生态系统的物种丰富度和多功能性之间的关系进行了研究, 发现非生物因子中年平均气温和土壤的砂粒含量对多功能性预测能力最强(均与多功能性呈负相关), 但按照赤池信息准则(Akaka’s Information Criterion, AICc)从255个模型中选出的8个最优模型(解释信息量>55%)均包含物种丰富度这一变量, 表明物种丰富度本身虽然只能解释一小部分多功能性的变异, 但却是不可或缺的.由于物种均匀分布可以使资源的互补性利用更为充分, 而资源的互补性利用可使多功能性增加(Maestre et al ., 2012b ), 这可以解释本文中均匀度与多功能性指数的正相关关系.在Maestre等(2012a)的地衣微型生物土壤结皮模式实验中发现, 均匀度与物种丰富度的交互作用对多功能性有显著影响, 但单独与物种组成、空间分布格局和物种丰富度相比, 其对多功能性的相对重要性最小, Maestre等(2012a)认为这是因为二者的交互作用很大程度上受单个物种影响, 优势种数量与均匀度有倒数关系, 当一个优势种支配了某一特定的功能时, 均匀度就与这个功能呈负相关关系.本研究相对这种微宇宙实验尺度更大, 多功能性的影响因素更多, 也更为复杂; 而且三种土地利用方式下的草地群落中, 一些优势种还不足以对某些生态系统功能起决定作用; 尤其在放牧和刈割群落中, 按照胁迫梯度假说和生物多样性-生态系统功能理论(Hector et al ., 1999 ), 群落物种的正相互作用和资源的互补性利用可能仍起主导作用, 因此, 本研究中多功能性与物种均匀度呈正相关关系. ...
Diversity peaks at intermediate productivity in a laboratory microcosm.
1
2000
... 本文的研究结果还表明, 刈割和围封群落的多数土壤功能指标无显著差异, 都具有较好的容重、孔隙度指标和较高的有机质含量; 围封除有较高的土壤自然含水量外, TC 、C:N和AvP 含量均低于刈割群落(图1 ).这可能与凋落物的分解速度有关, 割草能够提高植物叶片的N含量, 使凋落物中C:N和木质素:N降低, 从而刺激微生物活性, 加速凋落物的分解和结构性碳的回归(Madritch & Cardinale, 2007 ; Walter et al ., 2013 ); 同时, 割草减少凋落物积累, 使地表增温, 有利于好氧细菌生长, 促使植被中的营养元素回归(陈慧颖等, 2014 ).与之相反, 围封群落中没有生物量的移出, 随着可溶性碳和养分的快速分解, 累积的凋落物和立枯物中有大量的结构性碳, 难分解物质的比重增加, 导致分解速率降低(Hossain & Sugiyama, 2008 ; 王忆慧等, 2015 ), 归还土壤中的营养物质减少.因此, 尽管刈割和围封的土壤指标大多没有显著差异, 但凋落物组成和分解速率的差异, 可能导致刈割群落有更高的养分循环效率, 使刈割群落的土壤有较高的TC 、C:N和其他营养元素含量, 所以刈割较之围封有更高的多功能性评价指数. ...
Biodiversity enhances ecosystem multifunc- tionality across trophic levels and habitats.
1
2015
... 由4个多样性指数与多功能性的相关性比较来看, 均匀度和物种丰富度指数对多功能性的影响要强于Shannon-Wiener指数和Simpson指数.而均匀度和物种丰富度指数对多功能性的影响均为样方尺度小于样地尺度, 表明均匀度和物种丰富度指数与多功能性之间的关系均有尺度依赖性.当然, 这一结果也不排除是由样本量的不同而造成, 这一问题会在未来研究中加以重视.本研究中物种的丰富度指数与生态系统的多功能性呈单峰曲线关系, 这一结果在许多研究单一功能与物种多样性关系的实验中并不鲜见(Waide et al ., 1999 ; Isbell et al ., 2009 ), 如生产力处于中等时物种丰富度最大(Kassen et al ., 2000 ; Partel et al ., 2007 ), 尽管生产力和物种丰富度之间的关系是不确定的(Adler et al ., 2011 ; Maestre et al ., 2012b ); 但最新的研究结果表明仅关注单一功能和物种间的关系低估了生物多样性对生态系统功能的重要性, 物种丰富的群落更能维持较高水平的生态系统多功能性(Zavaleta et al ., 2010 ; Lefcheck et al ., 2015 ).在已有的物种丰富度与多功能性的研究中, 由于一个物种不仅在生态系统的一个功能里起作用, 在其他的功能里可能同样起作用, 这些同时支持不同功能的物种重叠比例在0.2-0.5之间(Hector & Bagchi, 2007 ), 因此随着研究中涉及的生态系统功能数增多, 物种丰富度与之呈正的饱和关系(Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ).这可以部分支持本文的单峰曲线模型, 但更深层的原因可能需要更大的样本从更多的尺度进行分析.Maestre等(2012b)对全球范围内224个干旱地区生态系统的物种丰富度和多功能性之间的关系进行了研究, 发现非生物因子中年平均气温和土壤的砂粒含量对多功能性预测能力最强(均与多功能性呈负相关), 但按照赤池信息准则(Akaka’s Information Criterion, AICc)从255个模型中选出的8个最优模型(解释信息量>55%)均包含物种丰富度这一变量, 表明物种丰富度本身虽然只能解释一小部分多功能性的变异, 但却是不可或缺的.由于物种均匀分布可以使资源的互补性利用更为充分, 而资源的互补性利用可使多功能性增加(Maestre et al ., 2012b ), 这可以解释本文中均匀度与多功能性指数的正相关关系.在Maestre等(2012a)的地衣微型生物土壤结皮模式实验中发现, 均匀度与物种丰富度的交互作用对多功能性有显著影响, 但单独与物种组成、空间分布格局和物种丰富度相比, 其对多功能性的相对重要性最小, Maestre等(2012a)认为这是因为二者的交互作用很大程度上受单个物种影响, 优势种数量与均匀度有倒数关系, 当一个优势种支配了某一特定的功能时, 均匀度就与这个功能呈负相关关系.本研究相对这种微宇宙实验尺度更大, 多功能性的影响因素更多, 也更为复杂; 而且三种土地利用方式下的草地群落中, 一些优势种还不足以对某些生态系统功能起决定作用; 尤其在放牧和刈割群落中, 按照胁迫梯度假说和生物多样性-生态系统功能理论(Hector et al ., 1999 ), 群落物种的正相互作用和资源的互补性利用可能仍起主导作用, 因此, 本研究中多功能性与物种均匀度呈正相关关系. ...
Diversity loss with persistent human disturbance increases vulnerability to ecosystem collapse.
2013
Impacts of tree species diversity on litter decomposition in northern temperate forests of Wisconsin, USA: A multi-site experiment along a latitudinal gradient.
1
2007
... 在每个样方分别采集环刀样和原状土样, 同时按梅花形取样法用土钻取0-20 cm表层土混合样.在实验室测取与C、N、P三大物质循环和固存有关的指标: 土壤有机质(OM )、全碳(TC )、全氮(TN )、碱解氮(AN )、全磷(TP )和速效磷(AvP ) (Hector & Bagchi, 2007 ; 王明君等, 2007 ; 闫玉春等, 2009 ); 与土壤涵养水分有关的指标(Sanderson et al ., 2004 ): 土壤自然含水量(SMC )、土壤容重(BD )、毛管持水量(CMC ); 与土壤呼吸、养分转化和循环有关的指标: pH值、阳离子交换量(CEC ), C:N、总孔隙度(BP )、毛管孔隙度(CP )、非毛管孔隙度(NCP )、通气孔隙度(AP )等16个生产实践中简单易测但又对土壤功能体现较为全面的指标, 这些指标涉及土壤的水、肥、气、热等各方面和多个生态系统功能(王玉辉等, 2002 ; 张蕴薇等, 2002 ; Pei et al ., 2008 ; 王向涛等, 2010 ; 蒋德明等, 2013 ).尽管本文选取的指标并不全面, 但均是土壤调查中常采用的指标, 它们既能很好地反映土壤肥力状况、稳定性较高, 又易于调查和测定(王仁忠, 1998 ; 王玉辉等, 2002 ; 曹成有等, 2011 ; 陈慧颖等, 2014 ; 王忆慧等, 2015 ). ...
a). Species richness effects on ecosystem multifunctionality depend on evenness, composition and spatial pattern.
5
2012
... “multifunctionality”一词首先由Sanderson等(2004)提出, 他们认为经营者在管理牧场和草场生态系统时往往只关注初级生产力的提高(因为生产输出与经济效益直接相关), 而忽略了生态系统的其他价值(如环境和美学效益), 指出从可持续发展的角度出发, 生态系统同时执行多种功能的能力“multifunctionality”更应该被重视, 并从环境效益和以人类为中心的经济效益两个角度出发提供了体现multifunctionality的15个功能特征.Hector和Bagchi (2007)引用“multifunctionality”这一概念在《Nature》上发表文章, 认为生态系统的多种服务或过程就是“ecosystem ‘multifunctionality’ ”, 并第一次量化研究了物种多样性同时对8个联合的生态系统过程所起的作用.Gamfeldt等(2008)、Zavaleta等(2010)又引用Hector和Bagchi (2007)的这篇文章, 分别在《Ecology》和《PNAS》上发文探讨和量化生物多样性对维持高水平“multifunctionality”的重要性, 认为“multifunctionality”即为生态系统同时维持多个生态功能或服务的能力.自此, 随着引用的增多, “multifunctionality”一词开始被人们广泛认可并正式使用.Maestre等(2012b)在《Science》上发表文章, 应用14个土壤变量(分别体现C、N、P循环三个生态系统过程)来综合评价全球干旱地区的生态系统多功能性, 其选用的指标和评价方法几乎成为近年来生态系统多功能性研究的标准方法(Maestre et al ., 2012a ; Byrnes et al ., 2014 ; Soliveres et al ., 2014 ; Wagg et al ., 2014 ; Valencia et al ., 2015 ).随后, Maestre等(2012a)在《Journal of Ecology》上, Soli- veres等(2014)在《Global Ecology and Biogeography》上、Wagg等(2014)在《PANS》上发表的多功能性研究论文均采用了Maestre等的文章的土壤变量和评价方法, 最新由Valencia等(2015)发表在《New Phytologist》上的文章同样引用了这些土壤变量及其评价方法, 这些文章的发表, 标志着生态系统多功能性(multifunctionality)概念正在逐步完善, 其评价方法也趋于统一. ...
... 尽管按照最初的概念, 多功能性是指生态系统同时维持各种生态系统功能和服务的能力, 其评价指标应涉及经济、环境、景观美学等各方面的指标(Sanderson et al ., 2004 ), 但随着多功能性概念的发展, 狭义的概念被人们提出, 同时维持多个生态功能或服务的能力即可称为“多功能性”, 这便导致不同的研究中各研究者选用的指标和数量并不相同, 涉及的生态系统功能和数量也各不相同(Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ; Zavaleta et al ., 2010 ; Maestre et al ., 2012a , 2012b ; Wagg et al ., 2014 ; Lefcheck et al ., 2015 ).近年来多功能性的研究中, 评价指标的选取多参考Maestre等(2012a, 2012b)的研究, 由于土壤-植被是相互依赖、互为反馈的复合体, 并且土壤是生态系统的养分库和种子库, 土壤指标很大程度上能反映生态系统的多个功能(如C、N、P的固存, 水土保持, 承载野生动植物等)(Sanderson et al ., 2004 ; Maestre et al ., 2012a ), 因此本文也采用Maestre等(2012a, 2012b)的这一方法, 用土壤变量进行生态系统多功能性的评价. ...
... ; Maestre et al ., 2012a ), 因此本文也采用Maestre等(2012a, 2012b)的这一方法, 用土壤变量进行生态系统多功能性的评价. ...
... 同时按照Maestre等(2012a, 2012b)的方法, 计算出多功能性指数, 与本文的方法进行比较.每个样方中所有功能指标标准化后的Z-Scores的平均值即为该样方的多功能性指数M' (Maestre et al ., 2012a , 2012b ; Soliveres et al ., 2014 ; Valencia et al ., 2015 ). ...
... 由4个多样性指数与多功能性的相关性比较来看, 均匀度和物种丰富度指数对多功能性的影响要强于Shannon-Wiener指数和Simpson指数.而均匀度和物种丰富度指数对多功能性的影响均为样方尺度小于样地尺度, 表明均匀度和物种丰富度指数与多功能性之间的关系均有尺度依赖性.当然, 这一结果也不排除是由样本量的不同而造成, 这一问题会在未来研究中加以重视.本研究中物种的丰富度指数与生态系统的多功能性呈单峰曲线关系, 这一结果在许多研究单一功能与物种多样性关系的实验中并不鲜见(Waide et al ., 1999 ; Isbell et al ., 2009 ), 如生产力处于中等时物种丰富度最大(Kassen et al ., 2000 ; Partel et al ., 2007 ), 尽管生产力和物种丰富度之间的关系是不确定的(Adler et al ., 2011 ; Maestre et al ., 2012b ); 但最新的研究结果表明仅关注单一功能和物种间的关系低估了生物多样性对生态系统功能的重要性, 物种丰富的群落更能维持较高水平的生态系统多功能性(Zavaleta et al ., 2010 ; Lefcheck et al ., 2015 ).在已有的物种丰富度与多功能性的研究中, 由于一个物种不仅在生态系统的一个功能里起作用, 在其他的功能里可能同样起作用, 这些同时支持不同功能的物种重叠比例在0.2-0.5之间(Hector & Bagchi, 2007 ), 因此随着研究中涉及的生态系统功能数增多, 物种丰富度与之呈正的饱和关系(Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ).这可以部分支持本文的单峰曲线模型, 但更深层的原因可能需要更大的样本从更多的尺度进行分析.Maestre等(2012b)对全球范围内224个干旱地区生态系统的物种丰富度和多功能性之间的关系进行了研究, 发现非生物因子中年平均气温和土壤的砂粒含量对多功能性预测能力最强(均与多功能性呈负相关), 但按照赤池信息准则(Akaka’s Information Criterion, AICc)从255个模型中选出的8个最优模型(解释信息量>55%)均包含物种丰富度这一变量, 表明物种丰富度本身虽然只能解释一小部分多功能性的变异, 但却是不可或缺的.由于物种均匀分布可以使资源的互补性利用更为充分, 而资源的互补性利用可使多功能性增加(Maestre et al ., 2012b ), 这可以解释本文中均匀度与多功能性指数的正相关关系.在Maestre等(2012a)的地衣微型生物土壤结皮模式实验中发现, 均匀度与物种丰富度的交互作用对多功能性有显著影响, 但单独与物种组成、空间分布格局和物种丰富度相比, 其对多功能性的相对重要性最小, Maestre等(2012a)认为这是因为二者的交互作用很大程度上受单个物种影响, 优势种数量与均匀度有倒数关系, 当一个优势种支配了某一特定的功能时, 均匀度就与这个功能呈负相关关系.本研究相对这种微宇宙实验尺度更大, 多功能性的影响因素更多, 也更为复杂; 而且三种土地利用方式下的草地群落中, 一些优势种还不足以对某些生态系统功能起决定作用; 尤其在放牧和刈割群落中, 按照胁迫梯度假说和生物多样性-生态系统功能理论(Hector et al ., 1999 ), 群落物种的正相互作用和资源的互补性利用可能仍起主导作用, 因此, 本研究中多功能性与物种均匀度呈正相关关系. ...
b). Plant species richness and ecosystem multifunctionality in global drylands.
11
2012
... 虽然Maestre等的多功能性(multifunctionality)评价指标和量化方法被许多学者所采用, 但也存在一些争议(Byrnes et al ., 2014 ; Lefcheck et al ., 2015 ), 本文笔者认为这一方法中对每个样方多功能性指数(M )的计算只是14个评价指标的Z-scores简单平均, 并没有考虑各个指标间的权重, 显然默认每个指标有同等的地位和作用并不符合自然生态系统的真实情况.另外, Maestre等(2012b)选择的14个土壤指标涉及C、N、P的循环与固存, 尽管C、N、P这三大物质循环在生态系统中占据着重要的地位, 但土壤同时也有涵养水分、平衡和缓冲外界干扰、为微生物提供稳定的环境、为各种物理生化反应提供介质和可持续地供给植物营养等多重功能(王仁忠, 1998 ; 王玉辉等, 2002 ; Sanderson et al ., 2004 ; Pei et al ., 2008 ), 仅用C、N、P的循环与固存这三个功能来体现生态系统的“多”功能性, 不免略显勉强.因此本文用改进的量化方法, 尝试选取农业实践中较为常见、测定更为简易的土壤指标, 并结合具体的立地条件, 对内蒙古三种不同利用方式下的草原群落进行多功能性的评价, 探讨物种多样性与多功能性指数之间的关系, 试图达到以下目的: (1)结合立地条件, 选取更为合适的土壤多功能性评价指标; (2)与Maestre等人的评价方法相对比, 建立更为科学的多功能性指数评价方法; (3)评价不同土地利用方式下草原生态系统的多功能性, 探讨物种多样性对多功能性的影响. ...
... 尽管按照最初的概念, 多功能性是指生态系统同时维持各种生态系统功能和服务的能力, 其评价指标应涉及经济、环境、景观美学等各方面的指标(Sanderson et al ., 2004 ), 但随着多功能性概念的发展, 狭义的概念被人们提出, 同时维持多个生态功能或服务的能力即可称为“多功能性”, 这便导致不同的研究中各研究者选用的指标和数量并不相同, 涉及的生态系统功能和数量也各不相同(Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ; Zavaleta et al ., 2010 ; Maestre et al ., 2012a , 2012b ; Wagg et al ., 2014 ; Lefcheck et al ., 2015 ).近年来多功能性的研究中, 评价指标的选取多参考Maestre等(2012a, 2012b)的研究, 由于土壤-植被是相互依赖、互为反馈的复合体, 并且土壤是生态系统的养分库和种子库, 土壤指标很大程度上能反映生态系统的多个功能(如C、N、P的固存, 水土保持, 承载野生动植物等)(Sanderson et al ., 2004 ; Maestre et al ., 2012a ), 因此本文也采用Maestre等(2012a, 2012b)的这一方法, 用土壤变量进行生态系统多功能性的评价. ...
... ; Lefcheck et al ., 2015 ).近年来多功能性的研究中, 评价指标的选取多参考Maestre等(2012a, 2012b)的研究, 由于土壤-植被是相互依赖、互为反馈的复合体, 并且土壤是生态系统的养分库和种子库, 土壤指标很大程度上能反映生态系统的多个功能(如C、N、P的固存, 水土保持, 承载野生动植物等)(Sanderson et al ., 2004 ; Maestre et al ., 2012a ), 因此本文也采用Maestre等(2012a, 2012b)的这一方法, 用土壤变量进行生态系统多功能性的评价. ...
... 同时按照Maestre等(2012a, 2012b)的方法, 计算出多功能性指数, 与本文的方法进行比较.每个样方中所有功能指标标准化后的Z-Scores的平均值即为该样方的多功能性指数M' (Maestre et al ., 2012a , 2012b ; Soliveres et al ., 2014 ; Valencia et al ., 2015 ). ...
... Maestre等人的多功能性指数(M' )只是评价指标的Z-scores的简单平均(Maestre et al ., 2012a ; 2012b ; Valencia et al ., 2015 ), 如果评价指标间有较高的相关性, 这一方法的评价结果会包含较多的冗余信息(Valencia et al ., 2015 ), 但因为相关性较高的指标数量相对于总体较多的评价指标来说是可以被接受的, 而且这种方法简单而直接, 所以被越来越多的学者接受和采用(Byrnes et al ., 2014 ; Soliveres et al ., 2014 ; Wagg et al ., 2014 ; Lefcheck et al ., 2015 ; Valencia et al ., 2015 ).本文采用因子分析的方法对筛选后的土壤指标进行多功能性指数的量化, 尽管这一方法与Maestre等人的方法有很好的相关性(图5 ), 尤其是在样地尺度上(R 2 = 0.9231, p < 0.0001); 但因子分析通过降维的方法可适当降低多变量间因为相关而造成的冗余信息, 而且本文作者也认为, 当选用的评价指标更多、样本量更大时, 这一方法可能更为适用.本文得出的公因子情形与RDA分析的结果相一致(图2 , 图3 ), 可以相互印证; 它在降低变量间冗余信息的同时, 也消除了Z-Scores简单平均这一方法默认每个功能指标在生态系统中有同等地位的不合理性. ...
... ; 2012b ; Valencia et al ., 2015 ), 如果评价指标间有较高的相关性, 这一方法的评价结果会包含较多的冗余信息(Valencia et al ., 2015 ), 但因为相关性较高的指标数量相对于总体较多的评价指标来说是可以被接受的, 而且这种方法简单而直接, 所以被越来越多的学者接受和采用(Byrnes et al ., 2014 ; Soliveres et al ., 2014 ; Wagg et al ., 2014 ; Lefcheck et al ., 2015 ; Valencia et al ., 2015 ).本文采用因子分析的方法对筛选后的土壤指标进行多功能性指数的量化, 尽管这一方法与Maestre等人的方法有很好的相关性(图5 ), 尤其是在样地尺度上(R 2 = 0.9231, p < 0.0001); 但因子分析通过降维的方法可适当降低多变量间因为相关而造成的冗余信息, 而且本文作者也认为, 当选用的评价指标更多、样本量更大时, 这一方法可能更为适用.本文得出的公因子情形与RDA分析的结果相一致(图2 , 图3 ), 可以相互印证; 它在降低变量间冗余信息的同时, 也消除了Z-Scores简单平均这一方法默认每个功能指标在生态系统中有同等地位的不合理性. ...
... ; Lefcheck et al ., 2015 ; Valencia et al ., 2015 ).本文采用因子分析的方法对筛选后的土壤指标进行多功能性指数的量化, 尽管这一方法与Maestre等人的方法有很好的相关性(图5 ), 尤其是在样地尺度上(R 2 = 0.9231, p < 0.0001); 但因子分析通过降维的方法可适当降低多变量间因为相关而造成的冗余信息, 而且本文作者也认为, 当选用的评价指标更多、样本量更大时, 这一方法可能更为适用.本文得出的公因子情形与RDA分析的结果相一致(图2 , 图3 ), 可以相互印证; 它在降低变量间冗余信息的同时, 也消除了Z-Scores简单平均这一方法默认每个功能指标在生态系统中有同等地位的不合理性. ...
... 由4个多样性指数与多功能性的相关性比较来看, 均匀度和物种丰富度指数对多功能性的影响要强于Shannon-Wiener指数和Simpson指数.而均匀度和物种丰富度指数对多功能性的影响均为样方尺度小于样地尺度, 表明均匀度和物种丰富度指数与多功能性之间的关系均有尺度依赖性.当然, 这一结果也不排除是由样本量的不同而造成, 这一问题会在未来研究中加以重视.本研究中物种的丰富度指数与生态系统的多功能性呈单峰曲线关系, 这一结果在许多研究单一功能与物种多样性关系的实验中并不鲜见(Waide et al ., 1999 ; Isbell et al ., 2009 ), 如生产力处于中等时物种丰富度最大(Kassen et al ., 2000 ; Partel et al ., 2007 ), 尽管生产力和物种丰富度之间的关系是不确定的(Adler et al ., 2011 ; Maestre et al ., 2012b ); 但最新的研究结果表明仅关注单一功能和物种间的关系低估了生物多样性对生态系统功能的重要性, 物种丰富的群落更能维持较高水平的生态系统多功能性(Zavaleta et al ., 2010 ; Lefcheck et al ., 2015 ).在已有的物种丰富度与多功能性的研究中, 由于一个物种不仅在生态系统的一个功能里起作用, 在其他的功能里可能同样起作用, 这些同时支持不同功能的物种重叠比例在0.2-0.5之间(Hector & Bagchi, 2007 ), 因此随着研究中涉及的生态系统功能数增多, 物种丰富度与之呈正的饱和关系(Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ).这可以部分支持本文的单峰曲线模型, 但更深层的原因可能需要更大的样本从更多的尺度进行分析.Maestre等(2012b)对全球范围内224个干旱地区生态系统的物种丰富度和多功能性之间的关系进行了研究, 发现非生物因子中年平均气温和土壤的砂粒含量对多功能性预测能力最强(均与多功能性呈负相关), 但按照赤池信息准则(Akaka’s Information Criterion, AICc)从255个模型中选出的8个最优模型(解释信息量>55%)均包含物种丰富度这一变量, 表明物种丰富度本身虽然只能解释一小部分多功能性的变异, 但却是不可或缺的.由于物种均匀分布可以使资源的互补性利用更为充分, 而资源的互补性利用可使多功能性增加(Maestre et al ., 2012b ), 这可以解释本文中均匀度与多功能性指数的正相关关系.在Maestre等(2012a)的地衣微型生物土壤结皮模式实验中发现, 均匀度与物种丰富度的交互作用对多功能性有显著影响, 但单独与物种组成、空间分布格局和物种丰富度相比, 其对多功能性的相对重要性最小, Maestre等(2012a)认为这是因为二者的交互作用很大程度上受单个物种影响, 优势种数量与均匀度有倒数关系, 当一个优势种支配了某一特定的功能时, 均匀度就与这个功能呈负相关关系.本研究相对这种微宇宙实验尺度更大, 多功能性的影响因素更多, 也更为复杂; 而且三种土地利用方式下的草地群落中, 一些优势种还不足以对某些生态系统功能起决定作用; 尤其在放牧和刈割群落中, 按照胁迫梯度假说和生物多样性-生态系统功能理论(Hector et al ., 1999 ), 群落物种的正相互作用和资源的互补性利用可能仍起主导作用, 因此, 本研究中多功能性与物种均匀度呈正相关关系. ...
... ; Lefcheck et al ., 2015 ).在已有的物种丰富度与多功能性的研究中, 由于一个物种不仅在生态系统的一个功能里起作用, 在其他的功能里可能同样起作用, 这些同时支持不同功能的物种重叠比例在0.2-0.5之间(Hector & Bagchi, 2007 ), 因此随着研究中涉及的生态系统功能数增多, 物种丰富度与之呈正的饱和关系(Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ).这可以部分支持本文的单峰曲线模型, 但更深层的原因可能需要更大的样本从更多的尺度进行分析.Maestre等(2012b)对全球范围内224个干旱地区生态系统的物种丰富度和多功能性之间的关系进行了研究, 发现非生物因子中年平均气温和土壤的砂粒含量对多功能性预测能力最强(均与多功能性呈负相关), 但按照赤池信息准则(Akaka’s Information Criterion, AICc)从255个模型中选出的8个最优模型(解释信息量>55%)均包含物种丰富度这一变量, 表明物种丰富度本身虽然只能解释一小部分多功能性的变异, 但却是不可或缺的.由于物种均匀分布可以使资源的互补性利用更为充分, 而资源的互补性利用可使多功能性增加(Maestre et al ., 2012b ), 这可以解释本文中均匀度与多功能性指数的正相关关系.在Maestre等(2012a)的地衣微型生物土壤结皮模式实验中发现, 均匀度与物种丰富度的交互作用对多功能性有显著影响, 但单独与物种组成、空间分布格局和物种丰富度相比, 其对多功能性的相对重要性最小, Maestre等(2012a)认为这是因为二者的交互作用很大程度上受单个物种影响, 优势种数量与均匀度有倒数关系, 当一个优势种支配了某一特定的功能时, 均匀度就与这个功能呈负相关关系.本研究相对这种微宇宙实验尺度更大, 多功能性的影响因素更多, 也更为复杂; 而且三种土地利用方式下的草地群落中, 一些优势种还不足以对某些生态系统功能起决定作用; 尤其在放牧和刈割群落中, 按照胁迫梯度假说和生物多样性-生态系统功能理论(Hector et al ., 1999 ), 群落物种的正相互作用和资源的互补性利用可能仍起主导作用, 因此, 本研究中多功能性与物种均匀度呈正相关关系. ...
... ).这可以部分支持本文的单峰曲线模型, 但更深层的原因可能需要更大的样本从更多的尺度进行分析.Maestre等(2012b)对全球范围内224个干旱地区生态系统的物种丰富度和多功能性之间的关系进行了研究, 发现非生物因子中年平均气温和土壤的砂粒含量对多功能性预测能力最强(均与多功能性呈负相关), 但按照赤池信息准则(Akaka’s Information Criterion, AICc)从255个模型中选出的8个最优模型(解释信息量>55%)均包含物种丰富度这一变量, 表明物种丰富度本身虽然只能解释一小部分多功能性的变异, 但却是不可或缺的.由于物种均匀分布可以使资源的互补性利用更为充分, 而资源的互补性利用可使多功能性增加(Maestre et al ., 2012b ), 这可以解释本文中均匀度与多功能性指数的正相关关系.在Maestre等(2012a)的地衣微型生物土壤结皮模式实验中发现, 均匀度与物种丰富度的交互作用对多功能性有显著影响, 但单独与物种组成、空间分布格局和物种丰富度相比, 其对多功能性的相对重要性最小, Maestre等(2012a)认为这是因为二者的交互作用很大程度上受单个物种影响, 优势种数量与均匀度有倒数关系, 当一个优势种支配了某一特定的功能时, 均匀度就与这个功能呈负相关关系.本研究相对这种微宇宙实验尺度更大, 多功能性的影响因素更多, 也更为复杂; 而且三种土地利用方式下的草地群落中, 一些优势种还不足以对某些生态系统功能起决定作用; 尤其在放牧和刈割群落中, 按照胁迫梯度假说和生物多样性-生态系统功能理论(Hector et al ., 1999 ), 群落物种的正相互作用和资源的互补性利用可能仍起主导作用, 因此, 本研究中多功能性与物种均匀度呈正相关关系. ...
... 本文尝试引用生态系统多功能性这一概念对具体的草原群落进行评价, 并对现行主流的多功能性评价方法进行了改进, 使其更为科学合理.当然, 本研究与目前的绝大多数研究一样, 没有考虑不同营养级的物种多样性, 只是关注植物物种多样性, 尤其是缺乏对养分循环有重要驱动作用的微生物群落的考虑(Hector & Bagchi, 2007 ; Maestre et al ., 2012b ).另外, 指标的选取方面, 尽管土壤指标在很大程度上可以体现生态系统的多种功能, 例如保持水土、涵养水分和承载动植物, 并且土壤是生态系统的主要养分库(Sanderson et al ., 2004 ; Gamfeldt et al ., 2008 ; Valencia et al ., 2015 ), 但生态系统并不仅仅由土壤系统组成, 而是一定空间内共同栖居着的所有生物与其环境之间由于不断进行物质循环、能量流动和信息传递而形成的统一整体, 各个环境因子都是不可或缺的.生态系统的服务和功能是多方位的, 因此多功能性(multifunctionlity)的概念在未来仍需进一步明确和完善, 其量化方法也需进一步发展. ...
围封与放牧对土壤微生物和酶活性的影响
1
2013
... 全球变化在全世界范围内以前所未有的速度影响着生物多样性的变化, 进而使自然生态系统功能产生不可预知的改变(Grime, 1997 ; Cardinale et al ., 2012 ; MacDougall et al ., 2013 ; Valencia et al ., 2015 ), 尤其是随着物种消失速率的加快, 越来越多的学者开始关注物种多样性的改变和由之而引起的群落结构变化对生态系统功能的影响(Grime, 1997 ; Tilman et al ., 1997 ; Díaz & Cabido, 2001 ; Díaz et al ., 2007 ; Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ).20多年来, 许多研究工作围绕这一问题展开, 但这些工作大多仅关注物种多样性和群落结构与单一或少数生态系统功能之间的关系(Hector et al ., 1999 ; Waide et al ., 1999 ; Simova et al ., 2013 ; Valencia et al ., 2015 ), 对生态系统多种功能同时评价和研究的工作还不多见, 而生态系统最为重要的价值是同时提供多种功能和服务的能力(Sanderson et al ., 2004 ; Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ; Soliveres et al ., 2014 ), 因此物种多样性与生态系统多功能性(multifunctionality)之间关系的研究显得十分 重要. ...
Contrasting plant productivity-diversity relationships across latitude, the role of evolutionary history.
1
2007
... 本文的研究结果还表明, 刈割和围封群落的多数土壤功能指标无显著差异, 都具有较好的容重、孔隙度指标和较高的有机质含量; 围封除有较高的土壤自然含水量外, TC 、C:N和AvP 含量均低于刈割群落(图1 ).这可能与凋落物的分解速度有关, 割草能够提高植物叶片的N含量, 使凋落物中C:N和木质素:N降低, 从而刺激微生物活性, 加速凋落物的分解和结构性碳的回归(Madritch & Cardinale, 2007 ; Walter et al ., 2013 ); 同时, 割草减少凋落物积累, 使地表增温, 有利于好氧细菌生长, 促使植被中的营养元素回归(陈慧颖等, 2014 ).与之相反, 围封群落中没有生物量的移出, 随着可溶性碳和养分的快速分解, 累积的凋落物和立枯物中有大量的结构性碳, 难分解物质的比重增加, 导致分解速率降低(Hossain & Sugiyama, 2008 ; 王忆慧等, 2015 ), 归还土壤中的营养物质减少.因此, 尽管刈割和围封的土壤指标大多没有显著差异, 但凋落物组成和分解速率的差异, 可能导致刈割群落有更高的养分循环效率, 使刈割群落的土壤有较高的TC 、C:N和其他营养元素含量, 所以刈割较之围封有更高的多功能性评价指数. ...
Changes in soil properties and vegetation following exclosure and grazing in degraded Alxa desert steppe of Inner Mongolia, China. Agriculture,
2008
Why does fertilization reduce plant species diversity? Testing three competition-based hypotheses.
2002
Plant species diversity and management of temperate forage and grazing land ecosystems.
2004
三江源区围栏与放牧高山嵩草草甸植物群落特征分析
1
2014
... 放牧、刈割和围封是内蒙古草原最主要的三种利用方式, 分析三种方式下草原群落的物种多样性和生态系统功能变化对草原的可持续经营与管理有重要的指导意义.重度放牧的草地, 所有种群均受放牧压力的影响, 导致种群个体小型化, 生态位收缩(王炜等, 2000 ), 光合面积减少(Carrera et al ., 2008 ), 群落的盖度、高度和生产力下降(陈芙蓉等, 2013 ; 王忆慧等, 2015 ), 地表植被受到破坏, 地表裸露面积增大, 水分蒸散量增大(王玉辉等, 2002 ).牲畜的践踏作用又增大了土壤紧实度, 造成非毛管孔隙度的减少、增加了土壤容重, 也降低了毛管孔隙度和通气孔隙度, 使土壤的透气性、渗透力和蓄水能力降低(王玉辉等, 2002 ; 张蕴薇等, 2002 ; Carrera et al ., 2008 ).生产力下降也导致了凋落物的归还减少, 致使土壤中的有机质和氮含量减少(王玉辉等, 2002 ; 王明君等, 2007 ; Carrera et al ., 2008 ; Pei et al ., 2008 ), 伴随着有机质的减少, 土壤团聚体和地表结皮破坏(王向涛等, 2010 ), 使土壤黏粒减少, 砂粒增多, 进而导致植物-土壤界面无法保持平衡, 造成草地沙化(王明君等, 2007 ); 而N作为北方草原主要的限制因子, 土壤中N的减少反过来又会使群落光合作用等生态系统功能受到影响.另外, 本研究地中牛羊在晚间归宿, 伴随着牲畜排泄物在圈舍的累积和畜产品的产出, 各种营养元素被带离生态系统, 导致营养元素从土壤中流失(牛得草等, 2013 ).牲畜的选择性采食也使劣质牧草(N等营养含量较低、木质素等结构性碳较高)增多, 凋落物品质变差(较高的C:N和木质素:N)、分解速率降低, 抑制微生物活性, 使进入土壤中的有机质和营养元素进一步减少(王忆慧等, 2015 ).土壤结构的破坏、水分和营养元素的缺失导致重度放牧草地不再拥有适宜的群落微生境, 不仅不利于现有植物的生长发育(王炜等, 2000 ; 王玉辉等, 2002 ), 而且也不适宜机会物种的定居和拓殖, 所以导致重度放牧样地生物多样性的下降和土壤功能指标的恶化.因此与刈割和围封相比, 本研究中长期重度放牧的草地表现出以上各种退化现象; 群落的生物多样性显著降低(表1 ), 各土壤功能指标较差(图1 ), 表现出最低的多功能性评价指数(表2 ). ...
Relationship between species richness and productivity in plants: The role of sampling effect, heterogeneity and species pool.
2
2013
... 在退化草原的恢复过程中, 围封禁牧因投资少、见效快等优点被广泛应用(徐粒等, 2014 ).虽然短时间内, 围封可以显著提高草地物种多样性和生产力, 但围封时间过长则不利于群落维持较高的多样性和生产力水平(闫玉春等, 2009 ; 徐粒等, 2014 ).本研究中刈割和围封群落的各多样性指数之间虽然不存在统计上的显著差异, 但数值上刈割总是优于围封(表1 ), 说明经过7年或之后更长的时间, 刈割处理可能更有利于维持群落的生物多样性水平, 2015年的群落调查结果即为刈割的各多样性指标显著高于围封(另文发表).这可能是因为随着围封时间的延长, 凋落物和根系周转向土壤输入更多的有机质和营养元素(闫玉春等, 2009 ; 杨浩等, 2009 ), 土壤结构得到改善, 土壤的持水能力得到增强, 生产力和养分积累得到增加(Bai et al ., 2007 ), 养分和水分的竞争转向光资源的竞争, 次优种的竞争性排除导致物种多样性的下降(Rajaniemi, 2002 ; Partel et al ., 2007 ; Hautier et al ., 2009 ).另外, 草地凋落物、枯枝层的增多也抑制了植物的再生和耐阴幼苗的形成, 同样不利于草原的繁殖更新和群落稳定(石红霄等, 2014 ).而反观刈割, 其适度的干扰不仅有利于植物的补偿性生长, 也有利于N、P等元素的矿化. ...
... 由4个多样性指数与多功能性的相关性比较来看, 均匀度和物种丰富度指数对多功能性的影响要强于Shannon-Wiener指数和Simpson指数.而均匀度和物种丰富度指数对多功能性的影响均为样方尺度小于样地尺度, 表明均匀度和物种丰富度指数与多功能性之间的关系均有尺度依赖性.当然, 这一结果也不排除是由样本量的不同而造成, 这一问题会在未来研究中加以重视.本研究中物种的丰富度指数与生态系统的多功能性呈单峰曲线关系, 这一结果在许多研究单一功能与物种多样性关系的实验中并不鲜见(Waide et al ., 1999 ; Isbell et al ., 2009 ), 如生产力处于中等时物种丰富度最大(Kassen et al ., 2000 ; Partel et al ., 2007 ), 尽管生产力和物种丰富度之间的关系是不确定的(Adler et al ., 2011 ; Maestre et al ., 2012b ); 但最新的研究结果表明仅关注单一功能和物种间的关系低估了生物多样性对生态系统功能的重要性, 物种丰富的群落更能维持较高水平的生态系统多功能性(Zavaleta et al ., 2010 ; Lefcheck et al ., 2015 ).在已有的物种丰富度与多功能性的研究中, 由于一个物种不仅在生态系统的一个功能里起作用, 在其他的功能里可能同样起作用, 这些同时支持不同功能的物种重叠比例在0.2-0.5之间(Hector & Bagchi, 2007 ), 因此随着研究中涉及的生态系统功能数增多, 物种丰富度与之呈正的饱和关系(Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ).这可以部分支持本文的单峰曲线模型, 但更深层的原因可能需要更大的样本从更多的尺度进行分析.Maestre等(2012b)对全球范围内224个干旱地区生态系统的物种丰富度和多功能性之间的关系进行了研究, 发现非生物因子中年平均气温和土壤的砂粒含量对多功能性预测能力最强(均与多功能性呈负相关), 但按照赤池信息准则(Akaka’s Information Criterion, AICc)从255个模型中选出的8个最优模型(解释信息量>55%)均包含物种丰富度这一变量, 表明物种丰富度本身虽然只能解释一小部分多功能性的变异, 但却是不可或缺的.由于物种均匀分布可以使资源的互补性利用更为充分, 而资源的互补性利用可使多功能性增加(Maestre et al ., 2012b ), 这可以解释本文中均匀度与多功能性指数的正相关关系.在Maestre等(2012a)的地衣微型生物土壤结皮模式实验中发现, 均匀度与物种丰富度的交互作用对多功能性有显著影响, 但单独与物种组成、空间分布格局和物种丰富度相比, 其对多功能性的相对重要性最小, Maestre等(2012a)认为这是因为二者的交互作用很大程度上受单个物种影响, 优势种数量与均匀度有倒数关系, 当一个优势种支配了某一特定的功能时, 均匀度就与这个功能呈负相关关系.本研究相对这种微宇宙实验尺度更大, 多功能性的影响因素更多, 也更为复杂; 而且三种土地利用方式下的草地群落中, 一些优势种还不足以对某些生态系统功能起决定作用; 尤其在放牧和刈割群落中, 按照胁迫梯度假说和生物多样性-生态系统功能理论(Hector et al ., 1999 ), 群落物种的正相互作用和资源的互补性利用可能仍起主导作用, 因此, 本研究中多功能性与物种均匀度呈正相关关系. ...
Plant diversity and ecosystem multifunctionality peak at intermediate levels of woody cover in global drylands.
3
2014
... 虽然Maestre等的多功能性(multifunctionality)评价指标和量化方法被许多学者所采用, 但也存在一些争议(Byrnes et al ., 2014 ; Lefcheck et al ., 2015 ), 本文笔者认为这一方法中对每个样方多功能性指数(M )的计算只是14个评价指标的Z-scores简单平均, 并没有考虑各个指标间的权重, 显然默认每个指标有同等的地位和作用并不符合自然生态系统的真实情况.另外, Maestre等(2012b)选择的14个土壤指标涉及C、N、P的循环与固存, 尽管C、N、P这三大物质循环在生态系统中占据着重要的地位, 但土壤同时也有涵养水分、平衡和缓冲外界干扰、为微生物提供稳定的环境、为各种物理生化反应提供介质和可持续地供给植物营养等多重功能(王仁忠, 1998 ; 王玉辉等, 2002 ; Sanderson et al ., 2004 ; Pei et al ., 2008 ), 仅用C、N、P的循环与固存这三个功能来体现生态系统的“多”功能性, 不免略显勉强.因此本文用改进的量化方法, 尝试选取农业实践中较为常见、测定更为简易的土壤指标, 并结合具体的立地条件, 对内蒙古三种不同利用方式下的草原群落进行多功能性的评价, 探讨物种多样性与多功能性指数之间的关系, 试图达到以下目的: (1)结合立地条件, 选取更为合适的土壤多功能性评价指标; (2)与Maestre等人的评价方法相对比, 建立更为科学的多功能性指数评价方法; (3)评价不同土地利用方式下草原生态系统的多功能性, 探讨物种多样性对多功能性的影响. ...
... 在每个样方分别采集环刀样和原状土样, 同时按梅花形取样法用土钻取0-20 cm表层土混合样.在实验室测取与C、N、P三大物质循环和固存有关的指标: 土壤有机质(OM )、全碳(TC )、全氮(TN )、碱解氮(AN )、全磷(TP )和速效磷(AvP ) (Hector & Bagchi, 2007 ; 王明君等, 2007 ; 闫玉春等, 2009 ); 与土壤涵养水分有关的指标(Sanderson et al ., 2004 ): 土壤自然含水量(SMC )、土壤容重(BD )、毛管持水量(CMC ); 与土壤呼吸、养分转化和循环有关的指标: pH值、阳离子交换量(CEC ), C:N、总孔隙度(BP )、毛管孔隙度(CP )、非毛管孔隙度(NCP )、通气孔隙度(AP )等16个生产实践中简单易测但又对土壤功能体现较为全面的指标, 这些指标涉及土壤的水、肥、气、热等各方面和多个生态系统功能(王玉辉等, 2002 ; 张蕴薇等, 2002 ; Pei et al ., 2008 ; 王向涛等, 2010 ; 蒋德明等, 2013 ).尽管本文选取的指标并不全面, 但均是土壤调查中常采用的指标, 它们既能很好地反映土壤肥力状况、稳定性较高, 又易于调查和测定(王仁忠, 1998 ; 王玉辉等, 2002 ; 曹成有等, 2011 ; 陈慧颖等, 2014 ; 王忆慧等, 2015 ). ...
... 放牧、刈割和围封是内蒙古草原最主要的三种利用方式, 分析三种方式下草原群落的物种多样性和生态系统功能变化对草原的可持续经营与管理有重要的指导意义.重度放牧的草地, 所有种群均受放牧压力的影响, 导致种群个体小型化, 生态位收缩(王炜等, 2000 ), 光合面积减少(Carrera et al ., 2008 ), 群落的盖度、高度和生产力下降(陈芙蓉等, 2013 ; 王忆慧等, 2015 ), 地表植被受到破坏, 地表裸露面积增大, 水分蒸散量增大(王玉辉等, 2002 ).牲畜的践踏作用又增大了土壤紧实度, 造成非毛管孔隙度的减少、增加了土壤容重, 也降低了毛管孔隙度和通气孔隙度, 使土壤的透气性、渗透力和蓄水能力降低(王玉辉等, 2002 ; 张蕴薇等, 2002 ; Carrera et al ., 2008 ).生产力下降也导致了凋落物的归还减少, 致使土壤中的有机质和氮含量减少(王玉辉等, 2002 ; 王明君等, 2007 ; Carrera et al ., 2008 ; Pei et al ., 2008 ), 伴随着有机质的减少, 土壤团聚体和地表结皮破坏(王向涛等, 2010 ), 使土壤黏粒减少, 砂粒增多, 进而导致植物-土壤界面无法保持平衡, 造成草地沙化(王明君等, 2007 ); 而N作为北方草原主要的限制因子, 土壤中N的减少反过来又会使群落光合作用等生态系统功能受到影响.另外, 本研究地中牛羊在晚间归宿, 伴随着牲畜排泄物在圈舍的累积和畜产品的产出, 各种营养元素被带离生态系统, 导致营养元素从土壤中流失(牛得草等, 2013 ).牲畜的选择性采食也使劣质牧草(N等营养含量较低、木质素等结构性碳较高)增多, 凋落物品质变差(较高的C:N和木质素:N)、分解速率降低, 抑制微生物活性, 使进入土壤中的有机质和营养元素进一步减少(王忆慧等, 2015 ).土壤结构的破坏、水分和营养元素的缺失导致重度放牧草地不再拥有适宜的群落微生境, 不仅不利于现有植物的生长发育(王炜等, 2000 ; 王玉辉等, 2002 ), 而且也不适宜机会物种的定居和拓殖, 所以导致重度放牧样地生物多样性的下降和土壤功能指标的恶化.因此与刈割和围封相比, 本研究中长期重度放牧的草地表现出以上各种退化现象; 群落的生物多样性显著降低(表1 ), 各土壤功能指标较差(图1 ), 表现出最低的多功能性评价指数(表2 ). ...
The influence of functional diversity and compos- ition on ecosystem processes.
1
1997
... 在退化草原的恢复过程中, 围封禁牧因投资少、见效快等优点被广泛应用(徐粒等, 2014 ).虽然短时间内, 围封可以显著提高草地物种多样性和生产力, 但围封时间过长则不利于群落维持较高的多样性和生产力水平(闫玉春等, 2009 ; 徐粒等, 2014 ).本研究中刈割和围封群落的各多样性指数之间虽然不存在统计上的显著差异, 但数值上刈割总是优于围封(表1 ), 说明经过7年或之后更长的时间, 刈割处理可能更有利于维持群落的生物多样性水平, 2015年的群落调查结果即为刈割的各多样性指标显著高于围封(另文发表).这可能是因为随着围封时间的延长, 凋落物和根系周转向土壤输入更多的有机质和营养元素(闫玉春等, 2009 ; 杨浩等, 2009 ), 土壤结构得到改善, 土壤的持水能力得到增强, 生产力和养分积累得到增加(Bai et al ., 2007 ), 养分和水分的竞争转向光资源的竞争, 次优种的竞争性排除导致物种多样性的下降(Rajaniemi, 2002 ; Partel et al ., 2007 ; Hautier et al ., 2009 ).另外, 草地凋落物、枯枝层的增多也抑制了植物的再生和耐阴幼苗的形成, 同样不利于草原的繁殖更新和群落稳定(石红霄等, 2014 ).而反观刈割, 其适度的干扰不仅有利于植物的补偿性生长, 也有利于N、P等元素的矿化. ...
Functional diversity enhances the resistance of ecosystem multifunctionality to aridity in Mediterranean drylands.
6
2015
... 全球变化在全世界范围内以前所未有的速度影响着生物多样性的变化, 进而使自然生态系统功能产生不可预知的改变(Grime, 1997 ; Cardinale et al ., 2012 ; MacDougall et al ., 2013 ; Valencia et al ., 2015 ), 尤其是随着物种消失速率的加快, 越来越多的学者开始关注物种多样性的改变和由之而引起的群落结构变化对生态系统功能的影响(Grime, 1997 ; Tilman et al ., 1997 ; Díaz & Cabido, 2001 ; Díaz et al ., 2007 ; Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ).20多年来, 许多研究工作围绕这一问题展开, 但这些工作大多仅关注物种多样性和群落结构与单一或少数生态系统功能之间的关系(Hector et al ., 1999 ; Waide et al ., 1999 ; Simova et al ., 2013 ; Valencia et al ., 2015 ), 对生态系统多种功能同时评价和研究的工作还不多见, 而生态系统最为重要的价值是同时提供多种功能和服务的能力(Sanderson et al ., 2004 ; Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ; Soliveres et al ., 2014 ), 因此物种多样性与生态系统多功能性(multifunctionality)之间关系的研究显得十分 重要. ...
... 虽然Maestre等的多功能性(multifunctionality)评价指标和量化方法被许多学者所采用, 但也存在一些争议(Byrnes et al ., 2014 ; Lefcheck et al ., 2015 ), 本文笔者认为这一方法中对每个样方多功能性指数(M )的计算只是14个评价指标的Z-scores简单平均, 并没有考虑各个指标间的权重, 显然默认每个指标有同等的地位和作用并不符合自然生态系统的真实情况.另外, Maestre等(2012b)选择的14个土壤指标涉及C、N、P的循环与固存, 尽管C、N、P这三大物质循环在生态系统中占据着重要的地位, 但土壤同时也有涵养水分、平衡和缓冲外界干扰、为微生物提供稳定的环境、为各种物理生化反应提供介质和可持续地供给植物营养等多重功能(王仁忠, 1998 ; 王玉辉等, 2002 ; Sanderson et al ., 2004 ; Pei et al ., 2008 ), 仅用C、N、P的循环与固存这三个功能来体现生态系统的“多”功能性, 不免略显勉强.因此本文用改进的量化方法, 尝试选取农业实践中较为常见、测定更为简易的土壤指标, 并结合具体的立地条件, 对内蒙古三种不同利用方式下的草原群落进行多功能性的评价, 探讨物种多样性与多功能性指数之间的关系, 试图达到以下目的: (1)结合立地条件, 选取更为合适的土壤多功能性评价指标; (2)与Maestre等人的评价方法相对比, 建立更为科学的多功能性指数评价方法; (3)评价不同土地利用方式下草原生态系统的多功能性, 探讨物种多样性对多功能性的影响. ...
... 尽管按照最初的概念, 多功能性是指生态系统同时维持各种生态系统功能和服务的能力, 其评价指标应涉及经济、环境、景观美学等各方面的指标(Sanderson et al ., 2004 ), 但随着多功能性概念的发展, 狭义的概念被人们提出, 同时维持多个生态功能或服务的能力即可称为“多功能性”, 这便导致不同的研究中各研究者选用的指标和数量并不相同, 涉及的生态系统功能和数量也各不相同(Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ; Zavaleta et al ., 2010 ; Maestre et al ., 2012a , 2012b ; Wagg et al ., 2014 ; Lefcheck et al ., 2015 ).近年来多功能性的研究中, 评价指标的选取多参考Maestre等(2012a, 2012b)的研究, 由于土壤-植被是相互依赖、互为反馈的复合体, 并且土壤是生态系统的养分库和种子库, 土壤指标很大程度上能反映生态系统的多个功能(如C、N、P的固存, 水土保持, 承载野生动植物等)(Sanderson et al ., 2004 ; Maestre et al ., 2012a ), 因此本文也采用Maestre等(2012a, 2012b)的这一方法, 用土壤变量进行生态系统多功能性的评价. ...
... ).近年来多功能性的研究中, 评价指标的选取多参考Maestre等(2012a, 2012b)的研究, 由于土壤-植被是相互依赖、互为反馈的复合体, 并且土壤是生态系统的养分库和种子库, 土壤指标很大程度上能反映生态系统的多个功能(如C、N、P的固存, 水土保持, 承载野生动植物等)(Sanderson et al ., 2004 ; Maestre et al ., 2012a ), 因此本文也采用Maestre等(2012a, 2012b)的这一方法, 用土壤变量进行生态系统多功能性的评价. ...
... 在每个样方分别采集环刀样和原状土样, 同时按梅花形取样法用土钻取0-20 cm表层土混合样.在实验室测取与C、N、P三大物质循环和固存有关的指标: 土壤有机质(OM )、全碳(TC )、全氮(TN )、碱解氮(AN )、全磷(TP )和速效磷(AvP ) (Hector & Bagchi, 2007 ; 王明君等, 2007 ; 闫玉春等, 2009 ); 与土壤涵养水分有关的指标(Sanderson et al ., 2004 ): 土壤自然含水量(SMC )、土壤容重(BD )、毛管持水量(CMC ); 与土壤呼吸、养分转化和循环有关的指标: pH值、阳离子交换量(CEC ), C:N、总孔隙度(BP )、毛管孔隙度(CP )、非毛管孔隙度(NCP )、通气孔隙度(AP )等16个生产实践中简单易测但又对土壤功能体现较为全面的指标, 这些指标涉及土壤的水、肥、气、热等各方面和多个生态系统功能(王玉辉等, 2002 ; 张蕴薇等, 2002 ; Pei et al ., 2008 ; 王向涛等, 2010 ; 蒋德明等, 2013 ).尽管本文选取的指标并不全面, 但均是土壤调查中常采用的指标, 它们既能很好地反映土壤肥力状况、稳定性较高, 又易于调查和测定(王仁忠, 1998 ; 王玉辉等, 2002 ; 曹成有等, 2011 ; 陈慧颖等, 2014 ; 王忆慧等, 2015 ). ...
... 本文尝试引用生态系统多功能性这一概念对具体的草原群落进行评价, 并对现行主流的多功能性评价方法进行了改进, 使其更为科学合理.当然, 本研究与目前的绝大多数研究一样, 没有考虑不同营养级的物种多样性, 只是关注植物物种多样性, 尤其是缺乏对养分循环有重要驱动作用的微生物群落的考虑(Hector & Bagchi, 2007 ; Maestre et al ., 2012b ).另外, 指标的选取方面, 尽管土壤指标在很大程度上可以体现生态系统的多种功能, 例如保持水土、涵养水分和承载动植物, 并且土壤是生态系统的主要养分库(Sanderson et al ., 2004 ; Gamfeldt et al ., 2008 ; Valencia et al ., 2015 ), 但生态系统并不仅仅由土壤系统组成, 而是一定空间内共同栖居着的所有生物与其环境之间由于不断进行物质循环、能量流动和信息传递而形成的统一整体, 各个环境因子都是不可或缺的.生态系统的服务和功能是多方位的, 因此多功能性(multifunctionlity)的概念在未来仍需进一步明确和完善, 其量化方法也需进一步发展. ...
Soil biodiversity and soil community composition determine ecosystem multifunctionality.
2014
The relationship between productivity and species richness.
1
1999
... 在退化草原的恢复过程中, 围封禁牧因投资少、见效快等优点被广泛应用(徐粒等, 2014 ).虽然短时间内, 围封可以显著提高草地物种多样性和生产力, 但围封时间过长则不利于群落维持较高的多样性和生产力水平(闫玉春等, 2009 ; 徐粒等, 2014 ).本研究中刈割和围封群落的各多样性指数之间虽然不存在统计上的显著差异, 但数值上刈割总是优于围封(表1 ), 说明经过7年或之后更长的时间, 刈割处理可能更有利于维持群落的生物多样性水平, 2015年的群落调查结果即为刈割的各多样性指标显著高于围封(另文发表).这可能是因为随着围封时间的延长, 凋落物和根系周转向土壤输入更多的有机质和营养元素(闫玉春等, 2009 ; 杨浩等, 2009 ), 土壤结构得到改善, 土壤的持水能力得到增强, 生产力和养分积累得到增加(Bai et al ., 2007 ), 养分和水分的竞争转向光资源的竞争, 次优种的竞争性排除导致物种多样性的下降(Rajaniemi, 2002 ; Partel et al ., 2007 ; Hautier et al ., 2009 ).另外, 草地凋落物、枯枝层的增多也抑制了植物的再生和耐阴幼苗的形成, 同样不利于草原的繁殖更新和群落稳定(石红霄等, 2014 ).而反观刈割, 其适度的干扰不仅有利于植物的补偿性生长, 也有利于N、P等元素的矿化. ...
Combined effects of multifactor climate change and land-use on decomposition in temperate grassland.
1
2013
... 全球变化在全世界范围内以前所未有的速度影响着生物多样性的变化, 进而使自然生态系统功能产生不可预知的改变(Grime, 1997 ; Cardinale et al ., 2012 ; MacDougall et al ., 2013 ; Valencia et al ., 2015 ), 尤其是随着物种消失速率的加快, 越来越多的学者开始关注物种多样性的改变和由之而引起的群落结构变化对生态系统功能的影响(Grime, 1997 ; Tilman et al ., 1997 ; Díaz & Cabido, 2001 ; Díaz et al ., 2007 ; Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ).20多年来, 许多研究工作围绕这一问题展开, 但这些工作大多仅关注物种多样性和群落结构与单一或少数生态系统功能之间的关系(Hector et al ., 1999 ; Waide et al ., 1999 ; Simova et al ., 2013 ; Valencia et al ., 2015 ), 对生态系统多种功能同时评价和研究的工作还不多见, 而生态系统最为重要的价值是同时提供多种功能和服务的能力(Sanderson et al ., 2004 ; Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ; Soliveres et al ., 2014 ), 因此物种多样性与生态系统多功能性(multifunctionality)之间关系的研究显得十分 重要. ...
草甸草原不同放牧强度对土壤有机碳含量的影响
4
2007
... 全球变化在全世界范围内以前所未有的速度影响着生物多样性的变化, 进而使自然生态系统功能产生不可预知的改变(Grime, 1997 ; Cardinale et al ., 2012 ; MacDougall et al ., 2013 ; Valencia et al ., 2015 ), 尤其是随着物种消失速率的加快, 越来越多的学者开始关注物种多样性的改变和由之而引起的群落结构变化对生态系统功能的影响(Grime, 1997 ; Tilman et al ., 1997 ; Díaz & Cabido, 2001 ; Díaz et al ., 2007 ; Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ).20多年来, 许多研究工作围绕这一问题展开, 但这些工作大多仅关注物种多样性和群落结构与单一或少数生态系统功能之间的关系(Hector et al ., 1999 ; Waide et al ., 1999 ; Simova et al ., 2013 ; Valencia et al ., 2015 ), 对生态系统多种功能同时评价和研究的工作还不多见, 而生态系统最为重要的价值是同时提供多种功能和服务的能力(Sanderson et al ., 2004 ; Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ; Soliveres et al ., 2014 ), 因此物种多样性与生态系统多功能性(multifunctionality)之间关系的研究显得十分 重要. ...
... “multifunctionality”一词首先由Sanderson等(2004)提出, 他们认为经营者在管理牧场和草场生态系统时往往只关注初级生产力的提高(因为生产输出与经济效益直接相关), 而忽略了生态系统的其他价值(如环境和美学效益), 指出从可持续发展的角度出发, 生态系统同时执行多种功能的能力“multifunctionality”更应该被重视, 并从环境效益和以人类为中心的经济效益两个角度出发提供了体现multifunctionality的15个功能特征.Hector和Bagchi (2007)引用“multifunctionality”这一概念在《Nature》上发表文章, 认为生态系统的多种服务或过程就是“ecosystem ‘multifunctionality’ ”, 并第一次量化研究了物种多样性同时对8个联合的生态系统过程所起的作用.Gamfeldt等(2008)、Zavaleta等(2010)又引用Hector和Bagchi (2007)的这篇文章, 分别在《Ecology》和《PNAS》上发文探讨和量化生物多样性对维持高水平“multifunctionality”的重要性, 认为“multifunctionality”即为生态系统同时维持多个生态功能或服务的能力.自此, 随着引用的增多, “multifunctionality”一词开始被人们广泛认可并正式使用.Maestre等(2012b)在《Science》上发表文章, 应用14个土壤变量(分别体现C、N、P循环三个生态系统过程)来综合评价全球干旱地区的生态系统多功能性, 其选用的指标和评价方法几乎成为近年来生态系统多功能性研究的标准方法(Maestre et al ., 2012a ; Byrnes et al ., 2014 ; Soliveres et al ., 2014 ; Wagg et al ., 2014 ; Valencia et al ., 2015 ).随后, Maestre等(2012a)在《Journal of Ecology》上, Soli- veres等(2014)在《Global Ecology and Biogeography》上、Wagg等(2014)在《PANS》上发表的多功能性研究论文均采用了Maestre等的文章的土壤变量和评价方法, 最新由Valencia等(2015)发表在《New Phytologist》上的文章同样引用了这些土壤变量及其评价方法, 这些文章的发表, 标志着生态系统多功能性(multifunctionality)概念正在逐步完善, 其评价方法也趋于统一. ...
... 同时按照Maestre等(2012a, 2012b)的方法, 计算出多功能性指数, 与本文的方法进行比较.每个样方中所有功能指标标准化后的Z-Scores的平均值即为该样方的多功能性指数M' (Maestre et al ., 2012a , 2012b ; Soliveres et al ., 2014 ; Valencia et al ., 2015 ). ...
... Maestre等人的多功能性指数(M' )只是评价指标的Z-scores的简单平均(Maestre et al ., 2012a ; 2012b ; Valencia et al ., 2015 ), 如果评价指标间有较高的相关性, 这一方法的评价结果会包含较多的冗余信息(Valencia et al ., 2015 ), 但因为相关性较高的指标数量相对于总体较多的评价指标来说是可以被接受的, 而且这种方法简单而直接, 所以被越来越多的学者接受和采用(Byrnes et al ., 2014 ; Soliveres et al ., 2014 ; Wagg et al ., 2014 ; Lefcheck et al ., 2015 ; Valencia et al ., 2015 ).本文采用因子分析的方法对筛选后的土壤指标进行多功能性指数的量化, 尽管这一方法与Maestre等人的方法有很好的相关性(图5 ), 尤其是在样地尺度上(R 2 = 0.9231, p < 0.0001); 但因子分析通过降维的方法可适当降低多变量间因为相关而造成的冗余信息, 而且本文作者也认为, 当选用的评价指标更多、样本量更大时, 这一方法可能更为适用.本文得出的公因子情形与RDA分析的结果相一致(图2 , 图3 ), 可以相互印证; 它在降低变量间冗余信息的同时, 也消除了Z-Scores简单平均这一方法默认每个功能指标在生态系统中有同等地位的不合理性. ...
放牧和刈割干扰对松嫩草原羊草草地影响的研究
1
1998
... 全球变化在全世界范围内以前所未有的速度影响着生物多样性的变化, 进而使自然生态系统功能产生不可预知的改变(Grime, 1997 ; Cardinale et al ., 2012 ; MacDougall et al ., 2013 ; Valencia et al ., 2015 ), 尤其是随着物种消失速率的加快, 越来越多的学者开始关注物种多样性的改变和由之而引起的群落结构变化对生态系统功能的影响(Grime, 1997 ; Tilman et al ., 1997 ; Díaz & Cabido, 2001 ; Díaz et al ., 2007 ; Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ).20多年来, 许多研究工作围绕这一问题展开, 但这些工作大多仅关注物种多样性和群落结构与单一或少数生态系统功能之间的关系(Hector et al ., 1999 ; Waide et al ., 1999 ; Simova et al ., 2013 ; Valencia et al ., 2015 ), 对生态系统多种功能同时评价和研究的工作还不多见, 而生态系统最为重要的价值是同时提供多种功能和服务的能力(Sanderson et al ., 2004 ; Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ; Soliveres et al ., 2014 ), 因此物种多样性与生态系统多功能性(multifunctionality)之间关系的研究显得十分 重要. ...
草原群落退化与恢复演替中的植物个体行为分析
8
2000
... 全球变化在全世界范围内以前所未有的速度影响着生物多样性的变化, 进而使自然生态系统功能产生不可预知的改变(Grime, 1997 ; Cardinale et al ., 2012 ; MacDougall et al ., 2013 ; Valencia et al ., 2015 ), 尤其是随着物种消失速率的加快, 越来越多的学者开始关注物种多样性的改变和由之而引起的群落结构变化对生态系统功能的影响(Grime, 1997 ; Tilman et al ., 1997 ; Díaz & Cabido, 2001 ; Díaz et al ., 2007 ; Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ).20多年来, 许多研究工作围绕这一问题展开, 但这些工作大多仅关注物种多样性和群落结构与单一或少数生态系统功能之间的关系(Hector et al ., 1999 ; Waide et al ., 1999 ; Simova et al ., 2013 ; Valencia et al ., 2015 ), 对生态系统多种功能同时评价和研究的工作还不多见, 而生态系统最为重要的价值是同时提供多种功能和服务的能力(Sanderson et al ., 2004 ; Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ; Soliveres et al ., 2014 ), 因此物种多样性与生态系统多功能性(multifunctionality)之间关系的研究显得十分 重要. ...
... ; Valencia et al ., 2015 ), 对生态系统多种功能同时评价和研究的工作还不多见, 而生态系统最为重要的价值是同时提供多种功能和服务的能力(Sanderson et al ., 2004 ; Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ; Soliveres et al ., 2014 ), 因此物种多样性与生态系统多功能性(multifunctionality)之间关系的研究显得十分 重要. ...
... “multifunctionality”一词首先由Sanderson等(2004)提出, 他们认为经营者在管理牧场和草场生态系统时往往只关注初级生产力的提高(因为生产输出与经济效益直接相关), 而忽略了生态系统的其他价值(如环境和美学效益), 指出从可持续发展的角度出发, 生态系统同时执行多种功能的能力“multifunctionality”更应该被重视, 并从环境效益和以人类为中心的经济效益两个角度出发提供了体现multifunctionality的15个功能特征.Hector和Bagchi (2007)引用“multifunctionality”这一概念在《Nature》上发表文章, 认为生态系统的多种服务或过程就是“ecosystem ‘multifunctionality’ ”, 并第一次量化研究了物种多样性同时对8个联合的生态系统过程所起的作用.Gamfeldt等(2008)、Zavaleta等(2010)又引用Hector和Bagchi (2007)的这篇文章, 分别在《Ecology》和《PNAS》上发文探讨和量化生物多样性对维持高水平“multifunctionality”的重要性, 认为“multifunctionality”即为生态系统同时维持多个生态功能或服务的能力.自此, 随着引用的增多, “multifunctionality”一词开始被人们广泛认可并正式使用.Maestre等(2012b)在《Science》上发表文章, 应用14个土壤变量(分别体现C、N、P循环三个生态系统过程)来综合评价全球干旱地区的生态系统多功能性, 其选用的指标和评价方法几乎成为近年来生态系统多功能性研究的标准方法(Maestre et al ., 2012a ; Byrnes et al ., 2014 ; Soliveres et al ., 2014 ; Wagg et al ., 2014 ; Valencia et al ., 2015 ).随后, Maestre等(2012a)在《Journal of Ecology》上, Soli- veres等(2014)在《Global Ecology and Biogeography》上、Wagg等(2014)在《PANS》上发表的多功能性研究论文均采用了Maestre等的文章的土壤变量和评价方法, 最新由Valencia等(2015)发表在《New Phytologist》上的文章同样引用了这些土壤变量及其评价方法, 这些文章的发表, 标志着生态系统多功能性(multifunctionality)概念正在逐步完善, 其评价方法也趋于统一. ...
... 同时按照Maestre等(2012a, 2012b)的方法, 计算出多功能性指数, 与本文的方法进行比较.每个样方中所有功能指标标准化后的Z-Scores的平均值即为该样方的多功能性指数M' (Maestre et al ., 2012a , 2012b ; Soliveres et al ., 2014 ; Valencia et al ., 2015 ). ...
... Maestre等人的多功能性指数(M' )只是评价指标的Z-scores的简单平均(Maestre et al ., 2012a ; 2012b ; Valencia et al ., 2015 ), 如果评价指标间有较高的相关性, 这一方法的评价结果会包含较多的冗余信息(Valencia et al ., 2015 ), 但因为相关性较高的指标数量相对于总体较多的评价指标来说是可以被接受的, 而且这种方法简单而直接, 所以被越来越多的学者接受和采用(Byrnes et al ., 2014 ; Soliveres et al ., 2014 ; Wagg et al ., 2014 ; Lefcheck et al ., 2015 ; Valencia et al ., 2015 ).本文采用因子分析的方法对筛选后的土壤指标进行多功能性指数的量化, 尽管这一方法与Maestre等人的方法有很好的相关性(图5 ), 尤其是在样地尺度上(R 2 = 0.9231, p < 0.0001); 但因子分析通过降维的方法可适当降低多变量间因为相关而造成的冗余信息, 而且本文作者也认为, 当选用的评价指标更多、样本量更大时, 这一方法可能更为适用.本文得出的公因子情形与RDA分析的结果相一致(图2 , 图3 ), 可以相互印证; 它在降低变量间冗余信息的同时, 也消除了Z-Scores简单平均这一方法默认每个功能指标在生态系统中有同等地位的不合理性. ...
... ), 如果评价指标间有较高的相关性, 这一方法的评价结果会包含较多的冗余信息(Valencia et al ., 2015 ), 但因为相关性较高的指标数量相对于总体较多的评价指标来说是可以被接受的, 而且这种方法简单而直接, 所以被越来越多的学者接受和采用(Byrnes et al ., 2014 ; Soliveres et al ., 2014 ; Wagg et al ., 2014 ; Lefcheck et al ., 2015 ; Valencia et al ., 2015 ).本文采用因子分析的方法对筛选后的土壤指标进行多功能性指数的量化, 尽管这一方法与Maestre等人的方法有很好的相关性(图5 ), 尤其是在样地尺度上(R 2 = 0.9231, p < 0.0001); 但因子分析通过降维的方法可适当降低多变量间因为相关而造成的冗余信息, 而且本文作者也认为, 当选用的评价指标更多、样本量更大时, 这一方法可能更为适用.本文得出的公因子情形与RDA分析的结果相一致(图2 , 图3 ), 可以相互印证; 它在降低变量间冗余信息的同时, 也消除了Z-Scores简单平均这一方法默认每个功能指标在生态系统中有同等地位的不合理性. ...
... ; Valencia et al ., 2015 ).本文采用因子分析的方法对筛选后的土壤指标进行多功能性指数的量化, 尽管这一方法与Maestre等人的方法有很好的相关性(图5 ), 尤其是在样地尺度上(R 2 = 0.9231, p < 0.0001); 但因子分析通过降维的方法可适当降低多变量间因为相关而造成的冗余信息, 而且本文作者也认为, 当选用的评价指标更多、样本量更大时, 这一方法可能更为适用.本文得出的公因子情形与RDA分析的结果相一致(图2 , 图3 ), 可以相互印证; 它在降低变量间冗余信息的同时, 也消除了Z-Scores简单平均这一方法默认每个功能指标在生态系统中有同等地位的不合理性. ...
... 本文尝试引用生态系统多功能性这一概念对具体的草原群落进行评价, 并对现行主流的多功能性评价方法进行了改进, 使其更为科学合理.当然, 本研究与目前的绝大多数研究一样, 没有考虑不同营养级的物种多样性, 只是关注植物物种多样性, 尤其是缺乏对养分循环有重要驱动作用的微生物群落的考虑(Hector & Bagchi, 2007 ; Maestre et al ., 2012b ).另外, 指标的选取方面, 尽管土壤指标在很大程度上可以体现生态系统的多种功能, 例如保持水土、涵养水分和承载动植物, 并且土壤是生态系统的主要养分库(Sanderson et al ., 2004 ; Gamfeldt et al ., 2008 ; Valencia et al ., 2015 ), 但生态系统并不仅仅由土壤系统组成, 而是一定空间内共同栖居着的所有生物与其环境之间由于不断进行物质循环、能量流动和信息传递而形成的统一整体, 各个环境因子都是不可或缺的.生态系统的服务和功能是多方位的, 因此多功能性(multifunctionlity)的概念在未来仍需进一步明确和完善, 其量化方法也需进一步发展. ...
不同放牧强度下高寒草甸植被特征和土壤养分变化研究
3
2010
... “multifunctionality”一词首先由Sanderson等(2004)提出, 他们认为经营者在管理牧场和草场生态系统时往往只关注初级生产力的提高(因为生产输出与经济效益直接相关), 而忽略了生态系统的其他价值(如环境和美学效益), 指出从可持续发展的角度出发, 生态系统同时执行多种功能的能力“multifunctionality”更应该被重视, 并从环境效益和以人类为中心的经济效益两个角度出发提供了体现multifunctionality的15个功能特征.Hector和Bagchi (2007)引用“multifunctionality”这一概念在《Nature》上发表文章, 认为生态系统的多种服务或过程就是“ecosystem ‘multifunctionality’ ”, 并第一次量化研究了物种多样性同时对8个联合的生态系统过程所起的作用.Gamfeldt等(2008)、Zavaleta等(2010)又引用Hector和Bagchi (2007)的这篇文章, 分别在《Ecology》和《PNAS》上发文探讨和量化生物多样性对维持高水平“multifunctionality”的重要性, 认为“multifunctionality”即为生态系统同时维持多个生态功能或服务的能力.自此, 随着引用的增多, “multifunctionality”一词开始被人们广泛认可并正式使用.Maestre等(2012b)在《Science》上发表文章, 应用14个土壤变量(分别体现C、N、P循环三个生态系统过程)来综合评价全球干旱地区的生态系统多功能性, 其选用的指标和评价方法几乎成为近年来生态系统多功能性研究的标准方法(Maestre et al ., 2012a ; Byrnes et al ., 2014 ; Soliveres et al ., 2014 ; Wagg et al ., 2014 ; Valencia et al ., 2015 ).随后, Maestre等(2012a)在《Journal of Ecology》上, Soli- veres等(2014)在《Global Ecology and Biogeography》上、Wagg等(2014)在《PANS》上发表的多功能性研究论文均采用了Maestre等的文章的土壤变量和评价方法, 最新由Valencia等(2015)发表在《New Phytologist》上的文章同样引用了这些土壤变量及其评价方法, 这些文章的发表, 标志着生态系统多功能性(multifunctionality)概念正在逐步完善, 其评价方法也趋于统一. ...
... 尽管按照最初的概念, 多功能性是指生态系统同时维持各种生态系统功能和服务的能力, 其评价指标应涉及经济、环境、景观美学等各方面的指标(Sanderson et al ., 2004 ), 但随着多功能性概念的发展, 狭义的概念被人们提出, 同时维持多个生态功能或服务的能力即可称为“多功能性”, 这便导致不同的研究中各研究者选用的指标和数量并不相同, 涉及的生态系统功能和数量也各不相同(Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ; Zavaleta et al ., 2010 ; Maestre et al ., 2012a , 2012b ; Wagg et al ., 2014 ; Lefcheck et al ., 2015 ).近年来多功能性的研究中, 评价指标的选取多参考Maestre等(2012a, 2012b)的研究, 由于土壤-植被是相互依赖、互为反馈的复合体, 并且土壤是生态系统的养分库和种子库, 土壤指标很大程度上能反映生态系统的多个功能(如C、N、P的固存, 水土保持, 承载野生动植物等)(Sanderson et al ., 2004 ; Maestre et al ., 2012a ), 因此本文也采用Maestre等(2012a, 2012b)的这一方法, 用土壤变量进行生态系统多功能性的评价. ...
... Maestre等人的多功能性指数(M' )只是评价指标的Z-scores的简单平均(Maestre et al ., 2012a ; 2012b ; Valencia et al ., 2015 ), 如果评价指标间有较高的相关性, 这一方法的评价结果会包含较多的冗余信息(Valencia et al ., 2015 ), 但因为相关性较高的指标数量相对于总体较多的评价指标来说是可以被接受的, 而且这种方法简单而直接, 所以被越来越多的学者接受和采用(Byrnes et al ., 2014 ; Soliveres et al ., 2014 ; Wagg et al ., 2014 ; Lefcheck et al ., 2015 ; Valencia et al ., 2015 ).本文采用因子分析的方法对筛选后的土壤指标进行多功能性指数的量化, 尽管这一方法与Maestre等人的方法有很好的相关性(图5 ), 尤其是在样地尺度上(R 2 = 0.9231, p < 0.0001); 但因子分析通过降维的方法可适当降低多变量间因为相关而造成的冗余信息, 而且本文作者也认为, 当选用的评价指标更多、样本量更大时, 这一方法可能更为适用.本文得出的公因子情形与RDA分析的结果相一致(图2 , 图3 ), 可以相互印证; 它在降低变量间冗余信息的同时, 也消除了Z-Scores简单平均这一方法默认每个功能指标在生态系统中有同等地位的不合理性. ...
草地利用方式对土壤呼吸和凋落物分解的影响
2
2015
... 全球变化在全世界范围内以前所未有的速度影响着生物多样性的变化, 进而使自然生态系统功能产生不可预知的改变(Grime, 1997 ; Cardinale et al ., 2012 ; MacDougall et al ., 2013 ; Valencia et al ., 2015 ), 尤其是随着物种消失速率的加快, 越来越多的学者开始关注物种多样性的改变和由之而引起的群落结构变化对生态系统功能的影响(Grime, 1997 ; Tilman et al ., 1997 ; Díaz & Cabido, 2001 ; Díaz et al ., 2007 ; Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ).20多年来, 许多研究工作围绕这一问题展开, 但这些工作大多仅关注物种多样性和群落结构与单一或少数生态系统功能之间的关系(Hector et al ., 1999 ; Waide et al ., 1999 ; Simova et al ., 2013 ; Valencia et al ., 2015 ), 对生态系统多种功能同时评价和研究的工作还不多见, 而生态系统最为重要的价值是同时提供多种功能和服务的能力(Sanderson et al ., 2004 ; Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ; Soliveres et al ., 2014 ), 因此物种多样性与生态系统多功能性(multifunctionality)之间关系的研究显得十分 重要. ...
... 由4个多样性指数与多功能性的相关性比较来看, 均匀度和物种丰富度指数对多功能性的影响要强于Shannon-Wiener指数和Simpson指数.而均匀度和物种丰富度指数对多功能性的影响均为样方尺度小于样地尺度, 表明均匀度和物种丰富度指数与多功能性之间的关系均有尺度依赖性.当然, 这一结果也不排除是由样本量的不同而造成, 这一问题会在未来研究中加以重视.本研究中物种的丰富度指数与生态系统的多功能性呈单峰曲线关系, 这一结果在许多研究单一功能与物种多样性关系的实验中并不鲜见(Waide et al ., 1999 ; Isbell et al ., 2009 ), 如生产力处于中等时物种丰富度最大(Kassen et al ., 2000 ; Partel et al ., 2007 ), 尽管生产力和物种丰富度之间的关系是不确定的(Adler et al ., 2011 ; Maestre et al ., 2012b ); 但最新的研究结果表明仅关注单一功能和物种间的关系低估了生物多样性对生态系统功能的重要性, 物种丰富的群落更能维持较高水平的生态系统多功能性(Zavaleta et al ., 2010 ; Lefcheck et al ., 2015 ).在已有的物种丰富度与多功能性的研究中, 由于一个物种不仅在生态系统的一个功能里起作用, 在其他的功能里可能同样起作用, 这些同时支持不同功能的物种重叠比例在0.2-0.5之间(Hector & Bagchi, 2007 ), 因此随着研究中涉及的生态系统功能数增多, 物种丰富度与之呈正的饱和关系(Hector & Bagchi, 2007 ; Gamfeldt et al ., 2008 ).这可以部分支持本文的单峰曲线模型, 但更深层的原因可能需要更大的样本从更多的尺度进行分析.Maestre等(2012b)对全球范围内224个干旱地区生态系统的物种丰富度和多功能性之间的关系进行了研究, 发现非生物因子中年平均气温和土壤的砂粒含量对多功能性预测能力最强(均与多功能性呈负相关), 但按照赤池信息准则(Akaka’s Information Criterion, AICc)从255个模型中选出的8个最优模型(解释信息量>55%)均包含物种丰富度这一变量, 表明物种丰富度本身虽然只能解释一小部分多功能性的变异, 但却是不可或缺的.由于物种均匀分布可以使资源的互补性利用更为充分, 而资源的互补性利用可使多功能性增加(Maestre et al ., 2012b ), 这可以解释本文中均匀度与多功能性指数的正相关关系.在Maestre等(2012a)的地衣微型生物土壤结皮模式实验中发现, 均匀度与物种丰富度的交互作用对多功能性有显著影响, 但单独与物种组成、空间分布格局和物种丰富度相比, 其对多功能性的相对重要性最小, Maestre等(2012a)认为这是因为二者的交互作用很大程度上受单个物种影响, 优势种数量与均匀度有倒数关系, 当一个优势种支配了某一特定的功能时, 均匀度就与这个功能呈负相关关系.本研究相对这种微宇宙实验尺度更大, 多功能性的影响因素更多, 也更为复杂; 而且三种土地利用方式下的草地群落中, 一些优势种还不足以对某些生态系统功能起决定作用; 尤其在放牧和刈割群落中, 按照胁迫梯度假说和生物多样性-生态系统功能理论(Hector et al ., 1999 ), 群落物种的正相互作用和资源的互补性利用可能仍起主导作用, 因此, 本研究中多功能性与物种均匀度呈正相关关系. ...
放牧强度对羊草草原的影响
1
2002
... 本文的研究结果还表明, 刈割和围封群落的多数土壤功能指标无显著差异, 都具有较好的容重、孔隙度指标和较高的有机质含量; 围封除有较高的土壤自然含水量外, TC 、C:N和AvP 含量均低于刈割群落(图1 ).这可能与凋落物的分解速度有关, 割草能够提高植物叶片的N含量, 使凋落物中C:N和木质素:N降低, 从而刺激微生物活性, 加速凋落物的分解和结构性碳的回归(Madritch & Cardinale, 2007 ; Walter et al ., 2013 ); 同时, 割草减少凋落物积累, 使地表增温, 有利于好氧细菌生长, 促使植被中的营养元素回归(陈慧颖等, 2014 ).与之相反, 围封群落中没有生物量的移出, 随着可溶性碳和养分的快速分解, 累积的凋落物和立枯物中有大量的结构性碳, 难分解物质的比重增加, 导致分解速率降低(Hossain & Sugiyama, 2008 ; 王忆慧等, 2015 ), 归还土壤中的营养物质减少.因此, 尽管刈割和围封的土壤指标大多没有显著差异, 但凋落物组成和分解速率的差异, 可能导致刈割群落有更高的养分循环效率, 使刈割群落的土壤有较高的TC 、C:N和其他营养元素含量, 所以刈割较之围封有更高的多功能性评价指数. ...
围封6年对温带典型草原坡地物种多样性及其与地上生物量的关系的影响
2014
围封对草地的影响研究进展
3
2009
... 在每个样方分别采集环刀样和原状土样, 同时按梅花形取样法用土钻取0-20 cm表层土混合样.在实验室测取与C、N、P三大物质循环和固存有关的指标: 土壤有机质(OM )、全碳(TC )、全氮(TN )、碱解氮(AN )、全磷(TP )和速效磷(AvP ) (Hector & Bagchi, 2007 ; 王明君等, 2007 ; 闫玉春等, 2009 ); 与土壤涵养水分有关的指标(Sanderson et al ., 2004 ): 土壤自然含水量(SMC )、土壤容重(BD )、毛管持水量(CMC ); 与土壤呼吸、养分转化和循环有关的指标: pH值、阳离子交换量(CEC ), C:N、总孔隙度(BP )、毛管孔隙度(CP )、非毛管孔隙度(NCP )、通气孔隙度(AP )等16个生产实践中简单易测但又对土壤功能体现较为全面的指标, 这些指标涉及土壤的水、肥、气、热等各方面和多个生态系统功能(王玉辉等, 2002 ; 张蕴薇等, 2002 ; Pei et al ., 2008 ; 王向涛等, 2010 ; 蒋德明等, 2013 ).尽管本文选取的指标并不全面, 但均是土壤调查中常采用的指标, 它们既能很好地反映土壤肥力状况、稳定性较高, 又易于调查和测定(王仁忠, 1998 ; 王玉辉等, 2002 ; 曹成有等, 2011 ; 陈慧颖等, 2014 ; 王忆慧等, 2015 ). ...
... 放牧、刈割和围封是内蒙古草原最主要的三种利用方式, 分析三种方式下草原群落的物种多样性和生态系统功能变化对草原的可持续经营与管理有重要的指导意义.重度放牧的草地, 所有种群均受放牧压力的影响, 导致种群个体小型化, 生态位收缩(王炜等, 2000 ), 光合面积减少(Carrera et al ., 2008 ), 群落的盖度、高度和生产力下降(陈芙蓉等, 2013 ; 王忆慧等, 2015 ), 地表植被受到破坏, 地表裸露面积增大, 水分蒸散量增大(王玉辉等, 2002 ).牲畜的践踏作用又增大了土壤紧实度, 造成非毛管孔隙度的减少、增加了土壤容重, 也降低了毛管孔隙度和通气孔隙度, 使土壤的透气性、渗透力和蓄水能力降低(王玉辉等, 2002 ; 张蕴薇等, 2002 ; Carrera et al ., 2008 ).生产力下降也导致了凋落物的归还减少, 致使土壤中的有机质和氮含量减少(王玉辉等, 2002 ; 王明君等, 2007 ; Carrera et al ., 2008 ; Pei et al ., 2008 ), 伴随着有机质的减少, 土壤团聚体和地表结皮破坏(王向涛等, 2010 ), 使土壤黏粒减少, 砂粒增多, 进而导致植物-土壤界面无法保持平衡, 造成草地沙化(王明君等, 2007 ); 而N作为北方草原主要的限制因子, 土壤中N的减少反过来又会使群落光合作用等生态系统功能受到影响.另外, 本研究地中牛羊在晚间归宿, 伴随着牲畜排泄物在圈舍的累积和畜产品的产出, 各种营养元素被带离生态系统, 导致营养元素从土壤中流失(牛得草等, 2013 ).牲畜的选择性采食也使劣质牧草(N等营养含量较低、木质素等结构性碳较高)增多, 凋落物品质变差(较高的C:N和木质素:N)、分解速率降低, 抑制微生物活性, 使进入土壤中的有机质和营养元素进一步减少(王忆慧等, 2015 ).土壤结构的破坏、水分和营养元素的缺失导致重度放牧草地不再拥有适宜的群落微生境, 不仅不利于现有植物的生长发育(王炜等, 2000 ; 王玉辉等, 2002 ), 而且也不适宜机会物种的定居和拓殖, 所以导致重度放牧样地生物多样性的下降和土壤功能指标的恶化.因此与刈割和围封相比, 本研究中长期重度放牧的草地表现出以上各种退化现象; 群落的生物多样性显著降低(表1 ), 各土壤功能指标较差(图1 ), 表现出最低的多功能性评价指数(表2 ). ...
... ), 使土壤黏粒减少, 砂粒增多, 进而导致植物-土壤界面无法保持平衡, 造成草地沙化(王明君等, 2007 ); 而N作为北方草原主要的限制因子, 土壤中N的减少反过来又会使群落光合作用等生态系统功能受到影响.另外, 本研究地中牛羊在晚间归宿, 伴随着牲畜排泄物在圈舍的累积和畜产品的产出, 各种营养元素被带离生态系统, 导致营养元素从土壤中流失(牛得草等, 2013 ).牲畜的选择性采食也使劣质牧草(N等营养含量较低、木质素等结构性碳较高)增多, 凋落物品质变差(较高的C:N和木质素:N)、分解速率降低, 抑制微生物活性, 使进入土壤中的有机质和营养元素进一步减少(王忆慧等, 2015 ).土壤结构的破坏、水分和营养元素的缺失导致重度放牧草地不再拥有适宜的群落微生境, 不仅不利于现有植物的生长发育(王炜等, 2000 ; 王玉辉等, 2002 ), 而且也不适宜机会物种的定居和拓殖, 所以导致重度放牧样地生物多样性的下降和土壤功能指标的恶化.因此与刈割和围封相比, 本研究中长期重度放牧的草地表现出以上各种退化现象; 群落的生物多样性显著降低(表1 ), 各土壤功能指标较差(图1 ), 表现出最低的多功能性评价指数(表2 ). ...
内蒙古典型草原物种组成和群落结构对长期放牧的响应
2009
Sustaining multiple ecosystem functions in grassland communities requires higher biodiversity.
2
2010
... 虽然Maestre等的多功能性(multifunctionality)评价指标和量化方法被许多学者所采用, 但也存在一些争议(Byrnes et al ., 2014 ; Lefcheck et al ., 2015 ), 本文笔者认为这一方法中对每个样方多功能性指数(M )的计算只是14个评价指标的Z-scores简单平均, 并没有考虑各个指标间的权重, 显然默认每个指标有同等的地位和作用并不符合自然生态系统的真实情况.另外, Maestre等(2012b)选择的14个土壤指标涉及C、N、P的循环与固存, 尽管C、N、P这三大物质循环在生态系统中占据着重要的地位, 但土壤同时也有涵养水分、平衡和缓冲外界干扰、为微生物提供稳定的环境、为各种物理生化反应提供介质和可持续地供给植物营养等多重功能(王仁忠, 1998 ; 王玉辉等, 2002 ; Sanderson et al ., 2004 ; Pei et al ., 2008 ), 仅用C、N、P的循环与固存这三个功能来体现生态系统的“多”功能性, 不免略显勉强.因此本文用改进的量化方法, 尝试选取农业实践中较为常见、测定更为简易的土壤指标, 并结合具体的立地条件, 对内蒙古三种不同利用方式下的草原群落进行多功能性的评价, 探讨物种多样性与多功能性指数之间的关系, 试图达到以下目的: (1)结合立地条件, 选取更为合适的土壤多功能性评价指标; (2)与Maestre等人的评价方法相对比, 建立更为科学的多功能性指数评价方法; (3)评价不同土地利用方式下草原生态系统的多功能性, 探讨物种多样性对多功能性的影响. ...
... 在每个样方分别采集环刀样和原状土样, 同时按梅花形取样法用土钻取0-20 cm表层土混合样.在实验室测取与C、N、P三大物质循环和固存有关的指标: 土壤有机质(OM )、全碳(TC )、全氮(TN )、碱解氮(AN )、全磷(TP )和速效磷(AvP ) (Hector & Bagchi, 2007 ; 王明君等, 2007 ; 闫玉春等, 2009 ); 与土壤涵养水分有关的指标(Sanderson et al ., 2004 ): 土壤自然含水量(SMC )、土壤容重(BD )、毛管持水量(CMC ); 与土壤呼吸、养分转化和循环有关的指标: pH值、阳离子交换量(CEC ), C:N、总孔隙度(BP )、毛管孔隙度(CP )、非毛管孔隙度(NCP )、通气孔隙度(AP )等16个生产实践中简单易测但又对土壤功能体现较为全面的指标, 这些指标涉及土壤的水、肥、气、热等各方面和多个生态系统功能(王玉辉等, 2002 ; 张蕴薇等, 2002 ; Pei et al ., 2008 ; 王向涛等, 2010 ; 蒋德明等, 2013 ).尽管本文选取的指标并不全面, 但均是土壤调查中常采用的指标, 它们既能很好地反映土壤肥力状况、稳定性较高, 又易于调查和测定(王仁忠, 1998 ; 王玉辉等, 2002 ; 曹成有等, 2011 ; 陈慧颖等, 2014 ; 王忆慧等, 2015 ). ...
贝加尔针茅草原植物多样性及土壤养分对放牧干扰的响应
2010
放牧强度对土壤物理性质的影响
2
2002
... 放牧、刈割和围封是内蒙古草原最主要的三种利用方式, 分析三种方式下草原群落的物种多样性和生态系统功能变化对草原的可持续经营与管理有重要的指导意义.重度放牧的草地, 所有种群均受放牧压力的影响, 导致种群个体小型化, 生态位收缩(王炜等, 2000 ), 光合面积减少(Carrera et al ., 2008 ), 群落的盖度、高度和生产力下降(陈芙蓉等, 2013 ; 王忆慧等, 2015 ), 地表植被受到破坏, 地表裸露面积增大, 水分蒸散量增大(王玉辉等, 2002 ).牲畜的践踏作用又增大了土壤紧实度, 造成非毛管孔隙度的减少、增加了土壤容重, 也降低了毛管孔隙度和通气孔隙度, 使土壤的透气性、渗透力和蓄水能力降低(王玉辉等, 2002 ; 张蕴薇等, 2002 ; Carrera et al ., 2008 ).生产力下降也导致了凋落物的归还减少, 致使土壤中的有机质和氮含量减少(王玉辉等, 2002 ; 王明君等, 2007 ; Carrera et al ., 2008 ; Pei et al ., 2008 ), 伴随着有机质的减少, 土壤团聚体和地表结皮破坏(王向涛等, 2010 ), 使土壤黏粒减少, 砂粒增多, 进而导致植物-土壤界面无法保持平衡, 造成草地沙化(王明君等, 2007 ); 而N作为北方草原主要的限制因子, 土壤中N的减少反过来又会使群落光合作用等生态系统功能受到影响.另外, 本研究地中牛羊在晚间归宿, 伴随着牲畜排泄物在圈舍的累积和畜产品的产出, 各种营养元素被带离生态系统, 导致营养元素从土壤中流失(牛得草等, 2013 ).牲畜的选择性采食也使劣质牧草(N等营养含量较低、木质素等结构性碳较高)增多, 凋落物品质变差(较高的C:N和木质素:N)、分解速率降低, 抑制微生物活性, 使进入土壤中的有机质和营养元素进一步减少(王忆慧等, 2015 ).土壤结构的破坏、水分和营养元素的缺失导致重度放牧草地不再拥有适宜的群落微生境, 不仅不利于现有植物的生长发育(王炜等, 2000 ; 王玉辉等, 2002 ), 而且也不适宜机会物种的定居和拓殖, 所以导致重度放牧样地生物多样性的下降和土壤功能指标的恶化.因此与刈割和围封相比, 本研究中长期重度放牧的草地表现出以上各种退化现象; 群落的生物多样性显著降低(表1 ), 各土壤功能指标较差(图1 ), 表现出最低的多功能性评价指数(表2 ). ...
... ).土壤结构的破坏、水分和营养元素的缺失导致重度放牧草地不再拥有适宜的群落微生境, 不仅不利于现有植物的生长发育(王炜等, 2000 ; 王玉辉等, 2002 ), 而且也不适宜机会物种的定居和拓殖, 所以导致重度放牧样地生物多样性的下降和土壤功能指标的恶化.因此与刈割和围封相比, 本研究中长期重度放牧的草地表现出以上各种退化现象; 群落的生物多样性显著降低(表1 ), 各土壤功能指标较差(图1 ), 表现出最低的多功能性评价指数(表2 ). ...



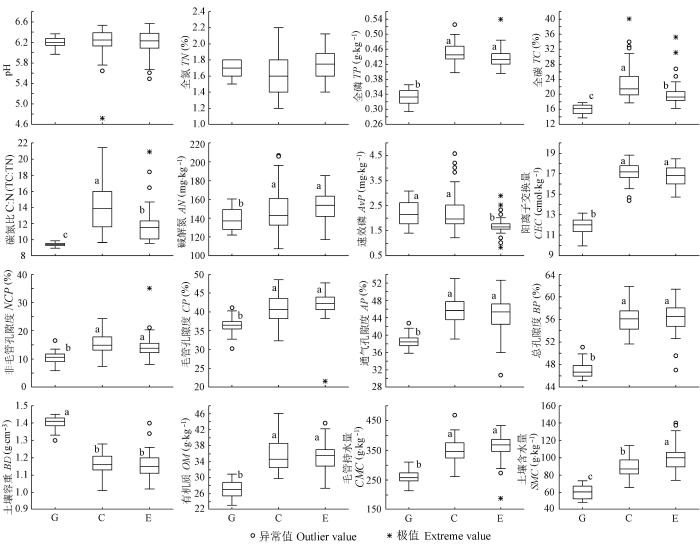
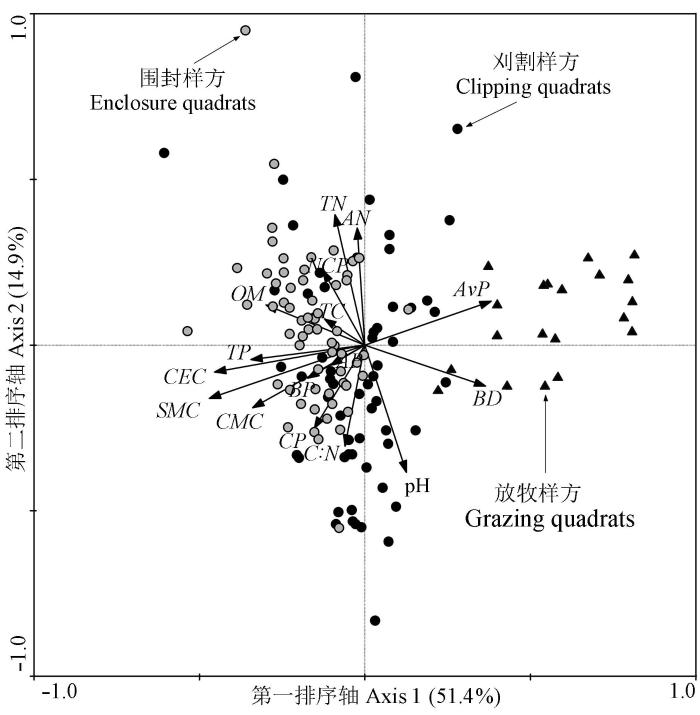
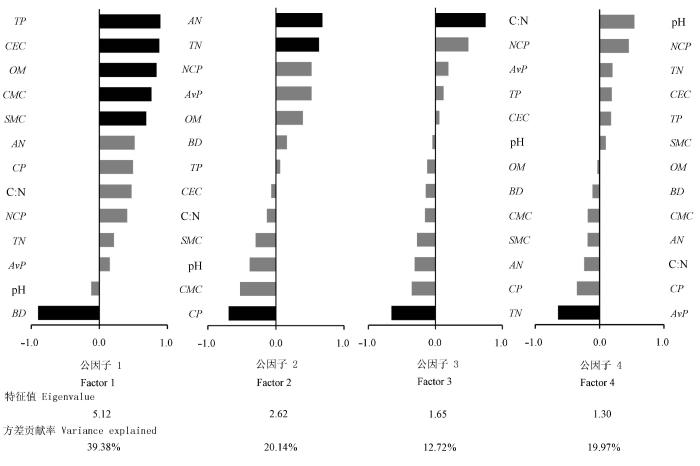
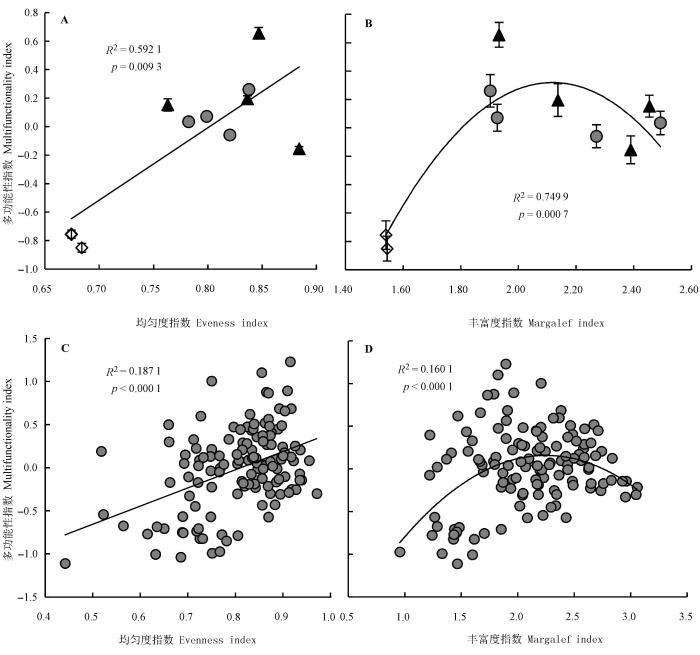
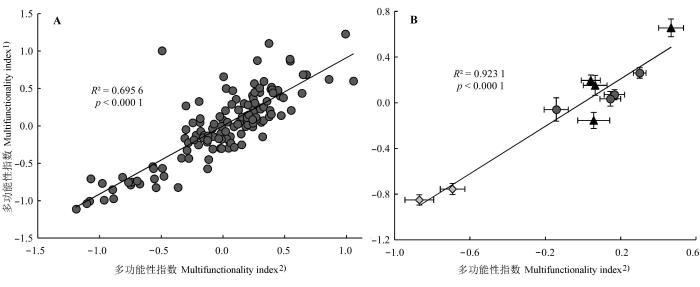
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}