植物功能性状与环境的关系是近年来生态学研究的热点, 尤其是植物的叶片性状具有较强的形态结构可塑性, 能够直接反映植物对环境变化的适应策略, 与资源获取和利用有着密切联系(Westoby & Wright, 2003 ; Rozendaal et al ., 2006 ; Bernard- Verdier et al ., 2012 ; 史元春等, 2015 )。植物的株高与其竞争力和繁殖活力有关(Cornelissen et al ., 2003 ), 增加对自身株高的投资能够提高光照的获取效率, 同时也增加了构成和维持支撑结构的投资(Falster & Westoby, 2003 )。生物量分配指植物生长过程中将同化的资源分配到根、茎、叶、花等器官中, 是植物重要的资源配置方式及净碳获取的驱动因子, 也是植物生存与生殖权衡的结果(方炎明, 1996 ; 郝虎东等, 2009 )。在资源匮乏或超过其利用上限的生境, 植物为了规避风险和增强对环境的适应性, 调整其功能性状等可塑性表型及生物量的分配格局来提高生存适合度(史元春等, 2015 )。异质环境中或不同的选择压力下, 现有关于植物功能性状和生物量分配的研究主要侧重于某一方面, 两者之间是否存在共轭变化, 即按照一定的规律或者关联性来匹配, 并且具有协同的动态变化, 目前的直接证据和定量化研究相对较少。因此, 定量化研究不同物种在不同生境下的叶片性状、株高与生物量分配格局的变化及其相互之间的关联性, 有助于理解全球变化背景下环境因子对植物相关功能的影响。
雪被广泛分布在寒冷生命带, 其积累与融化的交替过程直接影响着土壤水分、土壤温度、光照条件以及养分状况, 使得植物在生长节律、繁殖对策和生理生态方面形成一系列独特的适应机制(Jones et al ., 2001 ; 吴彦, 2005 ; 吴彦和Onipchenko, 2007 )。目前, 雪被生态学的研究主要集中在雪被对植物的地理分布、物质循环和土壤微生物动态的影响等方面(陈文年等, 2009 ; 胡霞等, 2012 ; 杨玉莲等, 2012 ; 何伟等, 2013 )。大量研究证据显示1980年以来北半球中高纬度的雪被面积减少了10% (Groisman et al ., 1994 ; Serreze & Hurst, 2000 ; Dye, 2002 ; IPCC, 2001 )。此外, 全球气候变化的趋势预示: 随着较高温度的出现, 高山地区降雪在近几十年将会下降(Serreze & Hurst, 2000 ; Beniston, 2005 ), 导致土壤冻结的增加, 而雪被厚度、融雪时间与生长季长度也将随之发生变化。因此, 雪被提供了季节性的复杂环境梯度(Körner, 2003 ; Pauli et al ., 2015 ), 深刻影响着高山植物的生长发育, 进而影响着高山生态系统的结构与功能(Wipf, 2010 )。然而, 雪被厚度及积雪消融时间长期形成的局域尺度的生境差异及其对植物叶片性状、生物量分配乃至群落结构的作用, 以及叶片性状与具体功能的相关性如何呈现等问题尚不明确, 更是缺乏相关的数据和证据支持。
高山地区的早春植物一般在初春开花, 分布于雪层厚度适中、光照较好的生境, 生长期较长, 且有着雪融水的滋润, 甚至在雪被覆盖下的生长和发育仍然十分活跃(吴彦, 2005 ), 是研究高山植物对不同雪被厚度下异质环境的响应及适应性的较佳材料。为此, 本研究在青藏高原东缘的高山草地选取了3种典型早春植物——紫罗兰报春(Primula purdomii )、甘肃马先蒿(Pedicularis kansuensis )、高原毛茛(Ranunculus tanguticus ), 研究以下两个问题: 1)雪被厚度变化如何影响早春植物的叶片性状、株高和生物量分配格局?2)植物的叶片性状、株高与生物量分配之间具有怎样的相关关系?从而探讨高度适应的植物物种在不同环境梯度如何表现出更好的适合度, 进一步理解全球变化背景下高山植物对不同环境及选择压力的响应和适应机制。
1 材料和方法
1.1 研究区域概况
试验地位于川西高原松潘县卡卡山(32.98° N, 103.67° E), 海拔3950 m, 属于青藏高原东缘岷山山脉的一部分。该区受西南暖湿气流和东南季风控制, 属于典型的山地季风气候, 年平均气温5.7 ℃, 1月平均气温-7.6 ℃, 7月平均气温9.7 ℃, 无绝对无霜期, 年太阳辐射时数1827.5 h, ≥10 ℃年积温428.6 ℃, 年降水量718 mm, 每年11月底或12月初出现持续的积雪覆盖, 积雪厚度在15 cm以上。土壤为棕色草毡土, 土壤pH值为5.42-5.83, 每kg干土含有有机质44.23-62. 00 g、全氮3.51-5.12 g, 平均土层厚度63 cm (胡霞等, 2012 ; Wang et al ., 2014 )。原生乔木以岷江冷杉(Abies fargesii var. faxoniana )为主; 高山灌木有窄叶鲜卑花(Sibiraea angustata )、 鹧鸪杜鹃(Rhododendron zheguense )和鹧鸪柳(Salix zhegushanica ), 以及零星分布的紫果云杉(Picea purpurea )等; 草本植物主要有黑褐穗薹草(Carex atrofusca )、长花马先蒿(Pedicularis longiflora )、长叶火绒草(Leontopodium longifolium )、高原毛茛、甘肃马先蒿、川西小黄菊(Pyrethrum tatsienense )和紫罗兰报春等(Liu et al ., 2010 ; 胡霞等, 2012 ; Wang et al ., 2014 )。
1.2 试验设计
在距山脊最高处50 m左右的斜坡(坡度变化8°-15°)朝南沿坡向下设置3条样带(11 m × 5 m), 相邻样带间距10 m左右。三条样带的垂直高程差为0.51 m, 从雪床的上端到下端, 雪被厚度自然增加, 融雪时间逐渐推迟, 形成了一个自然的融雪梯度(陈文年等, 2011a )。雪被厚度的监测从稳定积雪开始, 积雪消融殆尽结束, 每3天一次, 可根据天气情况适当调整。依据样带的雪被厚度监测数据(表1 )将其划分为厚雪、中雪和浅雪, 最大雪被厚度即为监测期间雪被的最大厚度, 其中浅雪形成时间较晚而消融时间较早, 最大雪被厚度较小, 厚雪则正好相反, 其最大厚度高于浅雪的2倍, 中雪梯度居中。土壤温度采用钮扣式温度计长期监测(置于距土表以下5 cm深处)。选取样带中广泛分布的3种早春优势种高原毛茛、紫罗兰报春和甘肃马先蒿为研究对象。高原毛茛和紫罗兰报春均为多年生草本, 后者多分布于潮湿的生境, 甘肃马先蒿为一年生或两年生草本(中国科学院中国植物志编辑委员会, 1963 , 1980, 1990 )。
1.3 样品采集及测定方法
在每条样带内每个物种随机选取20株左右, 测定其株高(plant height, PH ), 采用全株挖掘法进行取样, 尽可能保证根系完整, 每一植株采集2-3片完全伸展, 且无病虫害、无机械损伤的成熟叶片(李玉霖等, 2005 )。采用平板扫描仪(CanoScan5600F, Canon, Tokyo, Japan)扫描植株鲜叶样品, 然后借助Photoshop CS4和Matlab 7.9计算单叶面积(leaf area, LA )。将所取植株带回实验室, 水浸洗植株地下部分, 将植株分为根、茎、叶、花四类, 分别放入纸袋中, 连同扫描后的叶片于105 ℃杀青30 min后, 在80 ℃条件下烘干至恒质量。用电子天平称量干质量(精度为0.0001 g), 根据Vile等(2005)的方法计算比叶面积(比叶面积=叶面积/叶干质量), 并计算生物量分配, 将花视为繁殖构件。
雪被完全消融后, 采集土壤样品, 每个雪被地段设置5个小区, 每个小区采集5个样品, 然后混匀过筛(2 mm孔径)后测定土壤有机碳含量、全氮含量、全磷含量和土壤含水量, 土壤有机碳测定采用重铬酸钾氧化法(Allen, 1989 ), 全氮测定采用凯氏法(Allen, 1989 ; 鲍士旦, 2000 ), 全磷测定采用钒钼黄比色法(鲍士旦, 2000 )。
1.4 数据分析
首先对不同厚度雪被覆盖下的植物株高、叶片性状和各器官的生物量进行正态性检验, 不符合正态分布的数据通过对数转换, 使之满足正态分布, 利用Levene’s test检验方差齐性。然后, 通过一般线性模型(GLM)进行双因素方差分析, 探究物种和雪被厚度及其交互作用对植物功能性状的影响。运用单因素方差分析, 探究雪被厚度对三种草本植物株高、叶片性状和生物量分配的影响, 并用Tukey法进行多重比较。最后, 对植物性状与生物量指标之间进行Pearson相关分析。统计分析及作图分别在SPSS 19.0和Origin 8.5中完成。
1.5 异速生长分析
根据相关生长模型y = βxα 对不同雪被厚度下的三种草本植物地上/地下生物量及比叶面积-株高进行相关生长分析。分析前对生物量数据进行log10 的对数转换使之符合正态分布。方程可转化为lgy = lgβ + α lgx, x 和y 分别表示地上/地下生物量, β 为标准化常数, α 为异速生长指数。α = 1为等速生长关系, α ≠1为异速生长关系(Niklas & Enquist, 2001 ; Niklas, 2005 ; 丁建林等, 2014 ; 肖遥等, 2014 )。根据Warton和Weber (2002)的方法对同一物种在不同雪被厚度的α 进行异质性检验, 如果α 没有异质性, 则给出一个共同斜率, 若α 不同, 则异速生长关系完全不同, α 相同继续进行截距漂移检验和共轴漂移检验。若截距不同但共轴, 则属于截距漂移; 若截距相同且发生共轴漂移, 则属于共轴漂移; 若截距相同也同轴, 则表明植物的地上/地下生物量在不同雪被厚度下具有完全相同的相关生长关系(Falster et al ., 2006 ; 陶冶和张元明, 2014 )。相关生长模型的参数确定, 采用标准化主轴估计(standardized major axis estimation, SMA)方法(Wright et al ., 2001 ; Warton et al ., 2006 ), 截距漂移检验、共轴漂移检验及α 的方差分析均利用软件SMATR Version 2.0完成。
2 结果
2.1 不同雪被地段环境因子的比较
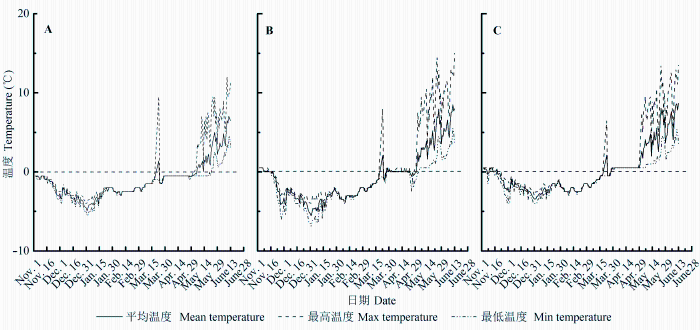
雪被覆盖期, 浅雪和中雪地段土壤日平均温度较厚雪地段低, 且每日波动幅度较大, 中雪地段土壤温度每日最大波动幅度可达3 ℃; 在早春植物的生长季, 厚雪和中雪地段土壤日平均温度略高于浅雪, 波动幅度也较大(图1 )。厚雪地段土壤有机碳含量、全氮含量显著高于浅雪和中雪地段, 全磷含量与浅雪和中雪地段无显著差异, 土壤水分含量均低于中雪地段(p < 0.05)和浅雪地段(p > 0.05); 中雪 和浅雪地段的土壤养分及水分含量均无显著差异 (表2 )。
图1 研究样地土壤温度时间动态。A, 浅雪。B, 中雪。C, 厚雪。
Fig. 1 Temporal dynamics of soil temperatures at study sites. A, Thin snow. B, Medium snow. C, Thick snow.
2.2 不同雪被厚度下三种植物株高和叶片性状的对比
三种植物之间的功能性状差异均极显著(p < 0.01); 雪被厚度对植物的株高、单叶质量、比叶面积的影响极显著(p < 0.01), 而对单叶面积的影响不显著(p > 0.05); 同时, 物种和雪被厚度的交互作用对株高、单叶质量、比叶面积的影响极显著(p < 0.01), 对单叶面积的影响不显著(p > 0.05)(表3 )。即代表着微环境差异的雪被厚度作用于植物的功能性状, 但对单叶面积的影响不大。
紫罗兰报春分布于厚雪和浅雪覆盖地段, 甘肃马先蒿在所有地段中均有分布, 高原毛茛则分布在中雪和浅雪覆盖地段。不同雪被厚度下, 三种植物除单叶面积无显著差异外, 其他功能性状均有显著变化。紫罗兰报春在厚雪地段的单叶质量显著大于浅雪(p < 0.01), 比叶面积的差异亦显著(p < 0.01), 且变化趋势相反, 株高则无显著性差异(p > 0.05); 浅雪地段甘肃马先蒿的单叶质量分别是其在厚雪和中雪的1.45倍(p < 0.05)和1.37倍(p = 0.054), 株高和比叶面积在厚雪和中雪地段均无显著差异, 株高显著低于浅雪地段(p < 0.05), 比叶面积则高于浅雪地段(p < 0.05); 高原毛茛在浅雪地段的单叶质量、株高均显著高于中雪(p < 0.05), 其比叶面积变化趋势则相反(表4 )。紫罗兰报春的比叶面积均显著大于甘肃马先蒿和高原毛茛(p < 0.001), 后两者之间的比叶面积无显著性差异(p = 0.188)。
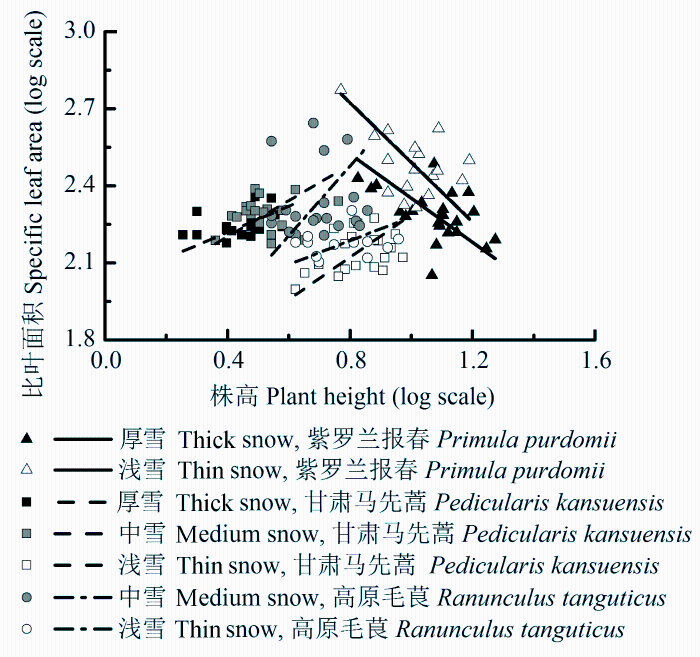
紫罗兰报春和甘肃马先蒿的比叶面积-株高在不同雪被厚度下均有共同的回归斜率, 分别为-0.971、0.667, 株高一定时, 浅雪地段的紫罗兰报春有更高的比叶面积, 而甘肃马先蒿有更低的比叶面积。高原毛茛的比叶面积-株高的回归斜率在不同雪被地段差异显著(p = 0.01)(图2 )。
2.3 不同雪被厚度下三种植物各器官生物量及其分配比例
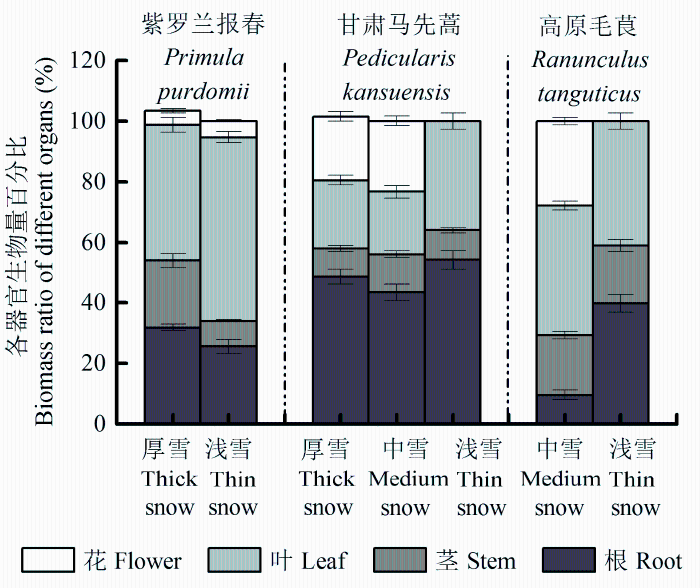
不同雪被厚度下, 紫罗兰报春和甘肃马先蒿的花生物量无显著差异(p > 0.05), 花生物量的分配比例差别亦不大, 前者分别为(4.48 ± 0.69)% (厚雪)、(5.32 ± 0.35)% (浅雪), 后者分别为(21.05 ± 1.55)% (厚雪)、(23.37 ± 1.56)% (中雪)。从厚雪到浅雪地段, 紫罗兰报春的总生物量及其他各构件生物量显著减小(p < 0.05), 茎和根生物量分配比例减小, 叶生物量分配比例增大。甘肃马先蒿总生物量及构件生物量在厚雪和中雪地段间无显著差异(p > 0.05), 且除叶生物量外均显著高于浅雪(p < 0.05); 在中雪地段, 其根、叶生物量分配比例((43.51 ± 2.63)%、(20.55 ± 2.10)%)均最低, 其茎生物量分配比例((12.57 ± 1.07)%)最高。高原毛茛总生物量及构件生物量(花除外)在浅雪地段显著高于中雪(p < 0.05), 与前两种植物生物量的变化相反, 其根生物量分配比例也在浅雪地段最大, 茎、叶生物量分配比例在中雪和浅雪地段差别均不大(表5 ; 图3 )。三种植物根冠比的变化范围在0.11-1.42之间, 其中紫罗兰报春的根冠比随着雪被厚度减小而减小, 另外两种植物则呈现增大趋势, 甘肃马先蒿的根冠比高于其他两种植物。
图2 不同雪被厚度下植物比叶面积和株高的标准主轴回归分析。
Fig. 2 Standardized major axis regression analysis between specific leaf area and plant height on sites with different levels of snow cover.
图3 不同雪被厚度下三种植物各器官生物量百分比(平均值±标准误差)。
Fig. 3 Biomass partitioning of three plant species under different levels of snow cover (mean ± SE ).
2.4 不同雪被厚度下三种植物地上-地下生物量间的相关生长关系
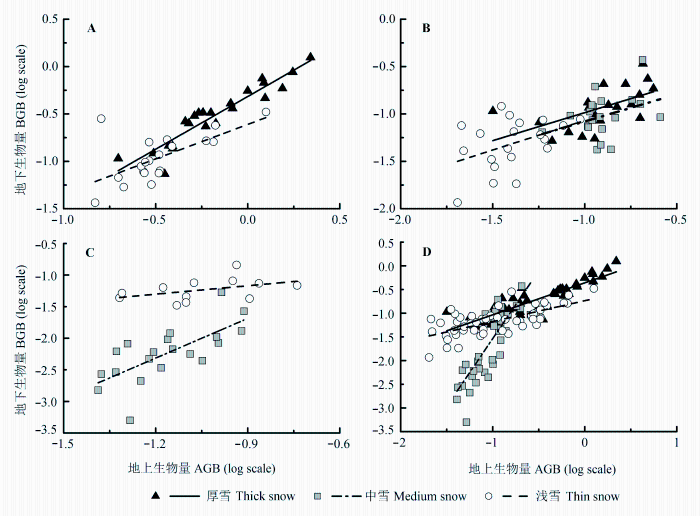
由图4 和表6 可知, 不同雪被厚度下紫罗兰报春的地上/地下生物量的α 差异并不显著, 具有共同斜率且显著大于1 (卡方检验: χ 2 = 4.945, p = 0.026), 地上/地下生物量为异速生长关系, 且地下生物量的增加速率快于地上生物量, 即将更多的生物量分配到地下。进一步检测它们在不同雪被厚度下是否有共同的截距和线性拟合轴。Wald 检验结果表明,紫罗兰报春地上/地下生物量在不同地段具有共同的截距, 发生显著的共轴漂移。
图4 三种植物地上/地下生物量(以10为底取对数)的线性拟合关系。A, 紫罗兰报春。 B, 甘肃马先蒿。 C, 高原毛茛。 D, 全部物种。
Fig. 4 Linear regressions (log10 -log10 ) between aboveground biomass (AGB) and belowground biomass (BGB) of the three plant species. A, Primula purdomii . B, Pedicularis kansuensis . C, Ranunculus tanguticus . D, All species.
将所有样本进行综合检验, 在厚雪和浅雪地段α 差异不显著, 存在共同斜率且显著小于1, 同时发生了截距漂移和共轴漂移。在给定地上生物量时, 浅雪地段的地下生物量大于厚雪, 而α 在中雪地段与其他地段具有异质性, 并显著大于1, 地上生物量的积累快于地下。不同雪被地段, 植物的地上生物量均可较好地解释地下生物量的变异, 解释度分别为75.1% (厚雪)、63.1% (中雪)和46.7% (浅雪)。
2.5 植物株高、叶性状与个体生物量间的相关性
三种植物株高与叶片性状间的相关性基本一致(表7 ), 其株高与比叶面积均呈负相关关系(紫罗兰报春: r = -0.487, p < 0.01; 甘肃马先蒿: r = -0.449, p < 0.01; 高原毛茛: r = -0.185, p > 0.05), 与单叶面积正相关(紫罗兰报春: r = 0.595, p < 0.01; 甘肃马先蒿: r = 0.311, p < 0.05; 高原毛茛: r = 0.196, p > 0.05), 紫罗兰报春的株高与单叶质量极显著负相关, 另外两种植物对应性状之间正相关(甘肃马先蒿: r = 0.489, p < 0.01; 高原毛茛: r = 0.416, p < 0.05)。三种植物的功能性状和个体生物量指标间的关系则具有物种特异性(表5 ), 紫罗兰报春和甘肃马先蒿的地上、地下生物量与各自株高均为极显著负相关, 而高原毛茛的地上、地下生物量与其株高正相关但不显著。紫罗兰报春和高原毛茛的地上、地下生物量和各自比叶面积均负相关(紫罗兰报春: r = -0.497, p < 0.01, r = -0.454, p < 0.01; 高原毛茛: r = -0.166, p > 0.05, r = -0.425, p < 0.05), 与单叶质量和单叶面积均极显著正相关, 而甘肃马先蒿的地上、地下生物量与比叶面积正相关(r = 0.354, p < 0.01; r = 0.212, p > 0.05), 与其单叶质量和单叶面积相关性较弱。紫罗兰报春和甘肃马先蒿的花生物量与各自株高均为正相关(p > 0.05), 与比叶面积负相关(p > 0.05); 高原毛茛的花生物量与前两种植物相反, 与其株高负相关(r = -0.09, p > 0.05), 与其比叶面积正相关(r = 0.031, p > 0.05), 三种植物的花生物量与各自单叶质量和单叶面积的相关性均为正相关, 在紫罗兰报春和高原毛茛中极显著相关, 在高原毛茛中相关性各异(r = 0.27, p > 0.05; r = 0.339, p < 0.05)。甘肃马先蒿和高原毛茛的根冠比与功能性状间的相关性表现较为一致, 即与株高正相关(甘肃马先蒿: r = 0.330, p < 0.01; 高原毛茛: r = 0.236, p > 0.05), 与比叶面积负相关(r = -0.137, p > 0.05; r = -0.418, p < 0.05), 与单叶质量和单叶面积均为正相关(甘肃马先蒿: r = 0.275, p < 0.05, r = 0.245, p > 0.05; 高原毛茛: r = 0.639, p < 0.01, r = 0.293, p > 0.05); 而紫罗兰报春的根冠比与株高负相关(r = -0.178, p > 0.05), 与比叶面积极显著正相关, 与单叶质量负相关, 与单叶面积正相关(p > 0.05)。
3 讨论
3.1 雪被厚度对植物比叶面积和株高的影响
由于地形、风和光照等环境因素的作用, 季节性雪被覆盖地区的雪被厚度存在明显的斑块性差异, 相关的融雪时间及植物生长季长度也随之变化, 导致雪下光照、温度和水分等环境因子再分配, 影响着雪下土壤动物及微生物等生物因子(Wijk, 1986 ; Lipson et al ., 2002 ; Totland & Alatalo, 2002 ; Rixen et al ., 2004 ; 陈文年等, 2011b ; 杨玉莲等, 2012 ), 进而作用于植物的功能性状和生物量分配格局。持续雪被覆盖时期, 厚雪地段土壤日平均温度相对较高, 波动幅度也最小, 土壤微生物依然具有较强的活力, 依然进行着氮矿化过程(Schimel et al ., 2004 ; Hiltbrunner et al ., 2005 ), 为早春植物的生长储备养分, 因此厚雪地段土壤有机碳含量、全氮含量高于中雪和浅雪地段。不同雪被地段土壤体积含水量均在60%以上(表2 ), 可见水分并不构成限制因子, 热量和土壤养分是影响高山植物生长的主要因子(刘琪璟等, 2009 )。综合来看, 厚雪地段土壤状况最优, 中雪地段次之, 浅雪地段相对较差。浅雪地段融雪较早, 植物易受到早期冻融交替或早霜胁迫的影响, 甘肃马先蒿和高原毛茛采取适应性策略, 开花物候有所迟滞。比叶面积-株高之间标准化主轴回归分析(SMA)的结果也表明, 当株高一定时, 浅雪地段紫罗兰报春的比叶面积高于厚雪地段, 厚雪地段甘肃马先蒿的比叶面积则高于浅雪地段, 而与中雪地段无显著差异(图2 )。尽管在不同雪被地段高原毛茛的比叶面积-株高间的回归直线斜率具有显著差异, 但中雪地段的植株比叶面积仍然高于浅雪地段。结果表明, 在土壤状况较好的地段, 甘肃马先蒿和高原毛茛具有相对较大的比叶面积。在厚雪和浅雪地段, 紫罗兰报春的株高无显著差异, 而浅雪地段植株的比叶面积高于厚雪地段。可能由于紫罗兰报春是多年生植物且具有相对粗短的根状茎和肉质长根, 厚雪地段植株的地下部分储存相对较多的营养物质以供生殖生长利用, 而浅雪地段植株主要依赖光合作用生产碳水化合物。
紫罗兰报春的株高在厚雪和浅雪地段无显著差异, 其他两种植物的株高均随着雪被厚度的减小而显著升高。两种植物株高的增加很可能与获取更多光照有关, 另外环境因子对草地群落的物种丰富度有着重要影响(杨元合等, 2004 ; Suding et al ., 2005 ), 随着雪被厚度减小, 生境内物种密度增加(未发表群落调查数据), 竞争压力也会增大, 株高不占优势的甘肃马先蒿和高原毛茛会增加对株高的投资以最大化碳收获。
3.2 生物量分配和异速生长关系
植物通过调节自身的资源配置以适应特定的生境, 形成对一定的环境变化及其复杂性和多样性的适应机制, 在环境变化及胁迫因子作用下具有各自的生长特性和物质分配规律(肖遥等, 2014 )。生物量分配主要取决于植物的遗传特性, 同时也受环境条件的影响(郝虎东等, 2009 ; Poorter et al ., 2012 )。不同的雪被地段, 高山草本植物各器官的生物量如何分配以适应环境的变化, 是研究高山植物生存策略的重要内容。本研究中, 不同雪被厚度覆盖地段下, 三种早春草本植物生物量分配的变化具有物种特异性。在土壤温度及养分状况更优的厚雪地段, 紫罗兰报春总生物量是浅雪地段植株的2倍多。厚雪地段的植株生长速率较高, 可将部分光合产物储存在根状茎和肉质长根中, 为下一阶段的生殖生长积蓄物质和能量, 而浅雪地段的植株生长环境较差, 需要将生物量更多地分配到叶中, 从而使碳收获最大化并达到生殖阈值。厚雪和中雪地段的甘肃马先蒿各部分器官生物量均高于浅雪地段植株, 而根和叶生物量分配比例均在浅雪地段最大, 既能够吸收较多养分, 又可以提高养分利用效率, 有利于其生殖生长。在生境状况相对较好的中雪地段, 高原毛茛各部分器官生物量反而小于浅雪地段植株, 可能是由于植物在浅雪地段具有更长的生长期, 可积累较多的生物量, 但伴随着雪被融化, 也面临着失去雪被保护、遭受霜冻和水分胁迫的风险(陈文年等, 2011b ), 并且浅雪地段土壤养分相对较低, 该地段高原毛茛分配更多的地下生物量, 因此生物量更大的浅雪地段高原毛茛进入生殖阶段晚于中雪地段植株。
异速生长揭示了某种生物或某一类生物的部分与整体或部分与部分间的差异和对比(Niklas & Enquist, 2001 ; Niklas, 2005 ; 李春萍等, 2007 )。植物的异速生长关系是其各器官相对生长速率可塑性的具体体现, 与植物的生物量分配、资源利用及对异质环境的形态适应有着密切关系(Bertalanffy, 1952 ; 陆霞梅等, 2007 )。植物生物量的分配是遗传和自然选择共同作用的结果(Weiner, 2004 ), 既具有物种特异性, 也受环境的影响。在厚雪和浅雪地段, 紫罗兰报春的地上/地下生物量表现出相同的异速生长关系, 且地下生物量的积累速率大于地上生物量的积累速率, 更多生物量分配到地下形成根状茎和肉质根, 以更好地完成生殖生长。甘肃马先蒿和高原毛茛在某些地段地上-地下生物量回归直线并不显著, 可能是样本量不足或者采样过程中样本的损耗造成。本研究结果与陶冶和张元明(2014)对6种类短命植物的地上-地下生物量的相关生长关系的研究结果较为一致, 均不支持Enquist和Niklas (2002)的等速生长假说。总体而言, 三种植物整体样本的地上-地下生物量分配均为异速生长关系, 在厚雪和浅雪地段具有共同的斜率且显著小于1, 地上生物量的积累快于地下, 而在中雪地段则相反, 生长季土壤温度在厚雪和浅雪地段相差不大且均高于中雪。另外, 整体样本中植物地上生物量可以较好地解释地下生物量的变化(表6 )。通过植物的地上部分生物量估算地下部分生物量, 可以探究整个区域内生物量对环境梯度变化的响应。
3.3 植物功能性状和生物量指标间的相关性
当受到胁迫或资源限制时, 植物会适当地调整其功能性状和生物量分配, 从而提高竞争力及其生存适合度(Sultan, 1992 ; 武高林和杜国祯, 2007 )。例如养分不足时, 植物会将生物量更多地分配到地下, 而减少地上部分的生物量分配, 植物的比叶面积和株高等功能性状也会相应减小(武高林等, 2010 ), 因此, 植物功能性状和生物量分配之间存在着某种共轭的变化。这种相互关系对于量化和概括植物面对生境变化的权衡策略及其规律有重要意义(Wright et al ., 2004 ; 陈莹婷和许振柱, 2014 )。本研究中, 紫罗兰报春(主要是株高和比叶面积与生物量指标之间)的相关性较强, 而这种相关性在甘肃马先蒿和高原毛茛相对较弱。由于生活型和个体发育的差异, 不同的物种在面临相同环境变化时, 权衡策略具有物种特异性, 可表征生物量变化的功能性状亦有所不同。因此, 如何选取具有普适性的性状, 表征不同尺度生物量的变化需要以后深入研究, 以丰富中国的植物性状数据库, 并更好地理解全球变化背景下植物功能性状的响应和适应对策。
致谢 感谢中国科学院成都生物研究所王乾老师在植物分类鉴定方面提供的帮助。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Allen SE 1989 ). Chemical Analysis of Ecological Material. 2nd edn. Blackwell, Oxford
, UK.
[本文引用: 2]
[2]
Bao SD Soil and Agricultural Chemistry Analysis . 3rd edn. China Agriculture Press , Beijing . (in Chinese)[鲍士旦 (2000). 土壤农化分析(第3版) . 中国农业出版社 , 北京 .]
[本文引用: 2]
[3]
Beniston M 2005 ). Mountain climates and climatic change: An overview of processes focusing on the European Alps.
Pure and Applied Geophysics , 162 , 1587 -1606 .
[4]
Bernard-Verdier M Navas ML Vellend M 2012 ). Community assembly along a soil depth gradient: Contrasting patterns of plant trait convergence and divergence in a Mediterra- nean rangeland.
Journal of Ecology , 100 , 1422 -1433 .
[本文引用: 1]
[5]
Bertalanffy LV 1952 ). Problems of Life: An Evaluation of Modern Biological and Scientific Thought. Harper
, New York.
[本文引用: 1]
[6]
Chen WN Wu Y Wu N Luo P 2009 ). Changes of five alpine species individual growth along snowmelt gradient.
Journal of Wuhan Botanical Research , 27 , 629 -636 . (in Chinese with English abstract)[陈文年 , 吴彦 , 吴宁 , 罗鹏 (2009 ). 五种高山植物的个体生长在融雪梯度上的变化
. 武汉植物学研究 , 27 , 629 -636 .]
[本文引用: 1]
[7]
Chen WN Wu Y Wu N Luo P 2011 a). Variation in phenology and population distribution pattern of three alpine species along the snowmelt gradient.
Bulletin of Botanical Research , 31 , 206 -212 . (in Chinese with English abstract)[陈文年 , 吴彦 , 吴宁 , 罗鹏 (2011 a). 3种高山植物的物候和种群分布格局在融雪梯度上的变化
. 植物研究 , 31 , 206 -212 .]
[本文引用: 1]
[8]
Chen WN Wu Y Wu N Luo P Wang Q 2011 b). Effects of snowmelt timing on individual growth and reproduction of Pedicularis davidii var. pentodon on the eastern Tibetan Plateau.
Acta Ecologica Sinica , 13 , 3621 -3628 . (in Chinese with English abstract)[陈文年 , 吴彦 , 吴宁 , 罗鹏 , 王乾 (2011 b). 融雪时间对大卫马先蒿生长和繁殖特性的影响
. 生态学报 , 13 , 3621 -3628 .]
[本文引用: 3]
[9]
Chen YT Xu ZZ 2014 ). Review on research of leaf economics spectrum.
Chinese Journal of Plant Ecology , 38 , 1135 -1153 . (in Chinese with English abstract)[陈莹婷 , 许振柱 (2014 ). 植物叶经济谱的研究进展
. 植物生态学报 , 38 , 1135 -1153 .]
[10]
Cornelissen JHC Lavorel S Garnier E Díaz S Buchmann N Gurvich DE Reich PB ter Steege H Morgan HD van der Heijden MGA Pausas JG Poorter H 2003 ). A handbook of protocols for standardised and easy measurement of plant functional traits worldwide.
Australian Journal of Botany , 51 , 335 -380 .
[11]
Delectis Florae Reipublicae Popularis Sinicae , Agendae Academiae Sinicae Edita 1963 ). Flora Reipublicae Popularis Sinicae . Science Press, Beijing . (in Chinese)[中国科学院中国植物志编辑委员会 (1963 ). 中国植物志 . 科学出版社 , 北京 .]
[本文引用: 1]
[12]
Delectis Florae Reipublicae Popularis Sinicae , Agendae Academiae Sinicae Edita 1980 ). Flora Reipublicae Popularis Sinicae . Science Press, Beijing . (in Chinese)[中国科学院中国植物志编辑委员会 (1980 ). 中国植物志 . 科学出版社 , 北京 .]
[本文引用: 1]
[13]
Delectis Florae Reipublicae Popularis Sinicae , Agendae Academiae Sinicae Edita 1990 ). Flora Reipublicae Popularis Sinicae . Science Press, Beijing . (in Chinese)[中国科学院中国植物志编辑委员会 (1990 ). 中国植物志 . 科学出版社 , 北京 .]
[14]
Ding JL Han Y Bao WK Xiang S 2014 ). Biomass allocation strategies of Lilium regale and their altitudinal effects.
Chinese Journal of Applied and Environmental Biology , 20 , 254 -260 . (in Chinese with English abstract)[丁建林 , 韩越 , 包维楷 , 向双 (2014 ). 岷江百合的生物量分配对策及其海拔效应
. 应用与环境生物学报 , 20 , 254 -260 .]
[本文引用: 1]
[15]
Dye DG 2002 ). Variability and trends in the annual snow-cover cycle in northern hemisphere land areas, 1972-2000.
Hydrological Processes , 16 , 3065 -3077 .
[本文引用: 1]
[16]
Enquist BJ Niklas KJ 2002 ). Global allocation rules for patterns of biomass partitioning in seed plants.
Science , 295 , 1517 -1520 .
[17]
Falster DS Warton DI Wright IJ
URL
[本文引用: 1]
[18]
Falster DS Westoby M 2003 ). Plant height and evolutionary games.
Trends in Ecology & Evolution , 18 , 337 -343 .
[19]
Fang YM Plant Reproductive Ecology . Shandong University Press , Jinan . (in Chinese)[方炎明 (1996). 植物生殖生态学 . 山东大学出版社 , 济南 .]
[本文引用: 1]
[20]
Groisman PY Karl TR Knight RW Stenchikov GL 1994 ). Changes of snow cover, temperature, and radiative heat- balance over the northern-hemisphere.
Journal of Climate , 7 , 1633 -1656 .
[21]
Hao HD Tian QS Shi FL Bian XY Li F 2009 ). Allocated dynamics of aboveground biomass and structural biomass in Bromus inermis Leyss.
Chinese Journal of Grassland , 31 (4 ), 85 -90 . (in Chinese with English abstract)[郝虎东 , 田青松 , 石凤翎 , 卞晓燕 , 李芳 (2009 ). 无芒雀麦地上生物量及各构件生物量分配动态
. 中国草地学报 , 31 (4 ), 85 -90 .]
[22]
He W Wu FZ Yang WQ Wu QQ He M Zhao YY 2013 ). Effect of snow patches on leaf litter mass loss of two shrubs in an alpine forest.
Chinese Journal of Plant Ecology , 37 , 306 -316 . (in Chinese with English abstract)[何伟 , 吴福忠 , 杨万勤 , 武启骞 , 何敏 , 赵野逸 (2013 ). 雪被斑块对高山森林两种灌木凋落叶质量损失的影响
. 植物生态学报 , 37 , 306 -316 .]
[23]
Hiltbrunner E Schwikowski M Korner C 2005 ). Inorganic nitrogen storage in alpine snow pack in the Central Alps (Switzerland).
Atmospheric Environment , 39 , 2249 -2259 .
[本文引用: 1]
[24]
Hu X Wu N Wu Y Zuo WQ Guo HX Wang JN 2012 ). Effects of snow cover on the decomposition and nutrient dynamics of Sibiraea angustata leaf litter in Western Sichuan Plateau, Southwest China.
Chinese Journal of Applied Ecology , 23 , 1226 -1232 . (in Chinese with English abstract)[胡霞 , 吴宁 , 吴彦 , 左万庆 , 郭海霞 , 王金牛 (2012 ). 川西高原季节性雪被覆盖对窄叶鲜卑花凋落物分解和养分动态的影响
. 应用生态学报 , 23 , 1226 -1232 .]
[本文引用: 1]
[25]
IPCC (Intergovernmental Panel on Climate Change ) (2001 ). Contribution of working group 1 to the third assessment report of the intergovernmental panel on climate change. In: Houghton JT, Ding Y, Griggs DG, Noguer M, Linden PJ, Xiaosu D eds. Climate Change in 2001: The Scientific Basis. Cambridge University Press, Cambridge, UK.
[本文引用: 1]
[26]
Jones HG Pomeroy JW Walker DA Hoham R 2001). Snow Ecology : An Interdisciplinary Examination of Snow- Covered Ecosystems. Cambridge University Press, Cambridge, UK .
[本文引用: 2]
[27]
Körner C 2003 ). Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems. Springer
, Berlin.
[28]
Li CP Li G Xiao CW 2007 ). The application of allometric relationships in biomass estimation in terrestrial ecosystems.
World Sci-Tech R & D , 29 (2 ), 51 -57 . (in Chinese with English abstract)[李春萍 , 李刚 , 肖春旺 (2007 ). 异速生长关系在陆地生态系统生物量估测中的应用
. 世界科技研究与发展 , 29 (2 ), 51 -57 .]
[29]
Li YL Cui JY Su YZ 2005 ). Specific leaf area and leaf dry matter content of some plants in different dune habitats.
Acta Ecologica Sinica , 25 , 304 -311 . (in Chinese with English abstract)[李玉霖 , 崔建垣 , 苏永中 (2005 ). 不同沙丘生境主要植物比叶面积和叶干物质含量的比较
. 生态学报 , 25 , 304 -311 .]
[本文引用: 1]
[30]
Lipson DA Schadt CW Schmidt SK 2002 ). Changes in soil microbial community structure and function in an alpine dry meadow following spring snow melt.
Microbial Ecology , 43 , 307 -314 .
[31]
Liu L Wu Y Wu N Xu JJ Mao Y Luo P Zhang L 2010 ). Effects of freezing and freeze-thaw cycles on soil microbial biomass and nutrient dynamics under different snow gradients in an alpine meadow (Tibetan Plateau).
Polish Journal of Ecology , 58 , 717 -728 .
[本文引用: 2]
[32]
Liu QJ Xu QQ Zhang GC 2009 ). Impact of alpine snowpacks on primary productivity in Rhododendron aureum community in Changbai Mountain, China.
Acta Ecologica Sinica , 29 , 4035 -4044 . (in Chinese with English abstract)[刘琪璟 , 徐倩倩 , 张国春 (2009 ). 高山带雪斑对牛皮杜鹃群落生产力的影响
. 生态学报 , 29 , 4035 -4044 .]
[33]
Lu XM Zhou CF An SQ Fang C Zhao H Yang Q Yan C 2007 ). Phenotypic plasticity, allometry and invasiveness of plants.
Chinese Journal of Ecology , 26 , 1438 -1444 . (in Chinese with English abstract)[陆霞梅 , 周长芳 , 安树青 , 方超 , 赵晖 , 杨茜 , 颜超 (2007 ). 植物的表型可塑性、异速生长及其入侵能力
. 生态学杂志 , 26 , 1438 -1444 .]
[本文引用: 1]
[34]
Niklas KJ 2005 ). Modelling below- and above-ground biomass for non-woody and woody plants.
Annals of Botany , 95 , 315 -321 .
[本文引用: 1]
[35]
Niklas KJ Enquist BJ 2001 ). Invariant scaling relationships for interspecific plant biomass production rates and body size.
Proceedings of the National Academy of Sciences of the United States of America , 98 , 2922 -2927 .
[36]
Pauli H Gottfried M Lamprecht A Niessner S Rumpf S Winkler M Steinbauer K Grabherr G 2015 ). The GLORIA Field Manual—Standard Multi-Summit Approach, Supplementary Methods and Extra Approaches. 5th edn. GLORIA-Coordination, Austrian Academy of Sciences & University of Natural Resources and Life Sciences
, Vienna.
[本文引用: 3]
[37]
Poorter H Niklas KJ Reich PB Oleksyn J Poot P Mommer L 2012 ). Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control.
New Phytologist , 193 , 30 -50 .
[38]
Rixen C Haeberli W Stoeckli V 2004 ). Ground temperatures under ski pistes with artificial and natural snow.
Arctic Antarctic and Alpine Research , 36 , 419 -427 .
[本文引用: 1]
[39]
Rozendaal DMA Hurtado VH Poorter L 2006 ). Plasticity in leaf traits of 38 tropical tree species in response to light: Relationships with light demand and adult stature.
Functional Ecology , 20 , 207 -216 .
[40]
Schimel JP Bilbrough C Welker JA 2004 ). Increased snow depth affects microbial activity and nitrogen mineralization in two arctic tundra communities.
Soil Biology & Biochemistry , 36 , 217 -227 .
[本文引用: 1]
[41]
Serreze MC Hurst CM 2000 ). Representation of mean arctic precipitation from NCEP-NCAR and ERA reanalyses.
Journal of Climate , 13 , 182 -201 .
[42]
Shi YC Zhao CZ Song QH Du J Chen J Wang JW 2015 ). Slope-related variations in twig and leaf traits of Robinia pseudoacacia in the northern mountains of Lanzhou.
Chinese Journal of Plant Ecology , 39 , 362 -370 . (in Chinese with English abstract)[史元春 , 赵成章 , 宋清华 , 杜晶 , 陈静 , 王继伟 (2015 ). 兰州北山刺槐枝叶性状的坡向差异性
. 植物生态学报 , 39 , 362 -370 .]
[本文引用: 1]
[43]
Suding KN Collins SL Gough L Clark C Cleland EE Gross KL Milchunas DG Pennings S 2005 ). Functional- and abundance-based mechanisms explain diversity loss due to N fertilization.
Proceedings of the National Academy of Sciences of the United States of America , 102 , 4387 -4392 .
[本文引用: 1]
[44]
Sultan SE 1992 ). Phenotypic plasticity and the neo-Darwinian legacy.
Evolutionary Trends in Plants , 6 , 61 -71 .
[本文引用: 1]
[45]
Tao Y Zhang YM 2014 ). Biomass allocation patterns and allometric relationships of six ephemeroid species in Junggar Basin, China.
Acta Prataculturae Sinica , 23 (2 ), 38 -48 . (in Chinese with English abstract)[陶冶 , 张元明 (2014 ). 准噶尔荒漠6种类短命植物生物量分配与异速生长关系
. 草业学报 , 23 (2 ), 38 -48 .]
[46]
Totland O Alatalo JM 2002 ). Effects of temperature and date of snowmelt on growth, reproduction, and flowering phenology in the arctic/alpine herb, Ranunculus glacialis.
Oecologia , 133 , 168 -175 .
[本文引用: 1]
[47]
Vile D Garnier E Shipley B Laurent G Navas ML Roumet C Lavorel S Diaz S Hodgson JG Lloret F Midgley GF Poorter H Rutherford MC Wilson PJ Wright IJ 2005 ). Specific leaf area and dry matter content estimate thick- ness in laminar leaves.
Annals of Botany , 96 , 1129 -1136 .
[48]
Wang JN Shi FS Xu B Wang Q Wu Y Wu N 2014 ). Uptake and recovery of soil nitrogen by bryophytes and vascular plants in an alpine meadow.
Journal of Mountain Science , 11 , 475 -484 .
[本文引用: 1]
[49]
Warton DI Weber NC 2002 ). Common slope tests for bivariate errors-in-variables models.
Biometrical Journal , 44 , 161 -174 .
[本文引用: 2]
[50]
Warton DI Wright IJ Falster DS Westoby M 2006 ). Bivariate line-fitting methods for allometry.
Biological Reviews of the Cambridge Philosophical Society , 81 , 259 -291 .
[本文引用: 2]
[51]
Weiner J 2004 ). Allocation, plasticity and allometry in plants.
Perspectives in Plant Ecology Evolution and Systematics , 6 , 207 -215 .
[本文引用: 1]
[52]
Westoby M Wright IJ 2003 ). The leaf size-twig size spectrum and its relationship to other important spectra of variation among species.
Oecologia , 135 , 621 -628 .
[本文引用: 1]
[53]
Wijk S 1986 ). Performance of Salix herbacea in an alpine snow-bed gradient.
Journal of Ecology , 74 , 675 -684 .
[本文引用: 1]
[54]
Wipf S 2010 ). Phenology, growth, and fecundity of eight subarctic tundra species in response to snowmelt manipulations.
Plant Ecology , 207 , 53 -66 .
[本文引用: 1]
[55]
Wright IJ Reich PB Westoby M 2001 ). Strategy shifts in leaf physiology, structure and nutrient content between species of high- and low-rainfall and high- and low-nutrient habitats.
Functional Ecology , 15 , 423 -434 .
[本文引用: 1]
[56]
Wright IJ Reich PB Westoby M Ackerly DD Baruch Z Bongers F Cavender-Bares J Chapin T Cornelissen JH Diemer M Flexas J Garnier E Groom PK Gulias J Hi- kosaka K Lamont BB Lee T Lee W Lusk C Midgley JJ Navas ML Niinemets U Oleksyn J Osada N Poorter H Poot P Prior L Pyankov VI Roumet C Thomas SC Tjoelker MG Veneklaas EJ Villar R 2004 ). The world- wide leaf economics spectrum.
Nature , 428 , 821 -827 .
[本文引用: 2]
[57]
Wu GL Chen M Du GZ 2010 ). Response of biomass allocation and morphological characteristics to light and nutrient resources for seedlings of three alpine species.
Acta Ecological Sinica , 30 , 60 -66 . (in Chinese with English abstract)[武高林 , 陈敏 , 杜国祯 (2010 ). 三种高寒植物幼苗生物量分配及性状特征对光照和养分的响应
. 生态学报 , 30 , 60 -66 .]
[58]
Wu GL Du GZ 2007 ). Advances in plant morphological growth strategy.
World Sci-Tech R & D , 29 (4 ), 47 -51 . (in Chinese with English abstract)[武高林 , 杜国祯 (2007 ). 植物形态生长对策研究进展
. 世界科技研究与发展 , 29 (4 ), 47 -51 .]
[本文引用: 2]
[59]
Wu Y 2005 ). Effects of seasonal snow cover on plant community.
Journal of Mountain Science , 23 , 40 -46 . (in Chinese with English abstract)[吴彦 (2005 ). 季节性雪被覆盖对植物群落的影响
. 山地学报 , 23 , 40 -46 .]
[本文引用: 1]
[60]
Wu Y Onipchenko VG 2007 ). The impact of snow-cover on alpine vegetation type of different aspects in the west of Sichuan Province.
Acta Ecological Sinica , 27 , 5120 -5129 . (in Chinese with English abstract)[吴彦 , Onipchenko VG 2007 ). 雪被对川西高山植被坡向性分异的影响
. 生态学报 , 27 , 5120 -5129 .]
[本文引用: 1]
[61]
Xiao Y Tao Y Zhang YM 2014 ). Biomass allocation and leaf stoichiometric characteristics in four desert herbaceous plants during different growth periods in the Gurbantünggüt Desert, China.
Chinese Journal of Plant Ecology , 38 , 929 -940 . (in Chinese with English abstract)[肖遥 , 陶冶 , 张元明 (2014 ). 古尔班通古特沙漠4种荒漠草本植物不同生长期的生物量分配与叶片化学计量特征
. 植物生态学报 , 38 , 929 -940 .]
[62]
Yang YH Rao S Hu HF Chen AP Ji CJ Zhu B Zuo WY Li XR Shen HH Wang ZH Tang YH Fang JY 2004 ). Plant species richness of alpine grasslands in relation to environmental factors and biomass on the Tibetan Plateau.
Biodiversity Science , 12 , 200 -205 . (in Chinese with English abstract)[杨元合 , 饶胜 , 胡会峰 , 陈安平 , 吉成均 , 朱彪 , 左闻韵 , 李轩然 , 沈海花 , 王志恒 , 唐艳鸿 , 方精云 (2004 ). 青藏高原高寒草地植物物种丰富度及其与环境因子和生物量的关系
. 生物多样性 , 12 , 200 -205 .]
[本文引用: 1]
[63]
Yang YL Wu FZ He ZH Xu ZF Liu Y Yang WQ Tan B 2012 ). Effects of snow pack removal on soil microbial biomass carbon and nitrogen and the number of soil culturable microorganisms during wintertime in alpine Abies faxoniana forest of western Sichuan, Southwest China.
Chinese Journal of Applied Ecology , 23 , 1809 -1816 . (in Chinese with English abstract)[杨玉莲 , 吴福忠 , 何振华 , 徐振锋 , 刘洋 , 杨万勤 , 谭波 (2012 ). 雪被去除对川西高山冷杉林冬季土壤微生物生物量碳氮和可培养微生物数量的影响
. 应用生态学报 , 23 , 1809 -1816 .]
[本文引用: 1]
Chemical Analysis of Ecological Material. 2nd edn. Blackwell, Oxford
2
1989
... 雪被完全消融后, 采集土壤样品, 每个雪被地段设置5个小区, 每个小区采集5个样品, 然后混匀过筛(2 mm孔径)后测定土壤有机碳含量、全氮含量、全磷含量和土壤含水量, 土壤有机碳测定采用重铬酸钾氧化法(Allen, 1989 ), 全氮测定采用凯氏法(Allen, 1989 ; 鲍士旦, 2000 ), 全磷测定采用钒钼黄比色法(鲍士旦, 2000 ). ...
... ), 全氮测定采用凯氏法(Allen, 1989 ; 鲍士旦, 2000 ), 全磷测定采用钒钼黄比色法(鲍士旦, 2000 ). ...
2
... 雪被完全消融后, 采集土壤样品, 每个雪被地段设置5个小区, 每个小区采集5个样品, 然后混匀过筛(2 mm孔径)后测定土壤有机碳含量、全氮含量、全磷含量和土壤含水量, 土壤有机碳测定采用重铬酸钾氧化法(Allen, 1989 ), 全氮测定采用凯氏法(Allen, 1989 ; 鲍士旦, 2000 ), 全磷测定采用钒钼黄比色法(鲍士旦, 2000 ). ...
... ), 全磷测定采用钒钼黄比色法(鲍士旦, 2000 ). ...
Mountain climates and climatic change: An overview of processes focusing on the European Alps.
2005
Community assembly along a soil depth gradient: Contrasting patterns of plant trait convergence and divergence in a Mediterra- nean rangeland.
1
2012
... 雪被广泛分布在寒冷生命带, 其积累与融化的交替过程直接影响着土壤水分、土壤温度、光照条件以及养分状况, 使得植物在生长节律、繁殖对策和生理生态方面形成一系列独特的适应机制(Jones et al ., 2001 ; 吴彦, 2005 ; 吴彦和Onipchenko, 2007 ).目前, 雪被生态学的研究主要集中在雪被对植物的地理分布、物质循环和土壤微生物动态的影响等方面(陈文年等, 2009 ; 胡霞等, 2012 ; 杨玉莲等, 2012 ; 何伟等, 2013 ).大量研究证据显示1980年以来北半球中高纬度的雪被面积减少了10% (Groisman et al ., 1994 ; Serreze & Hurst, 2000 ; Dye, 2002 ; IPCC, 2001 ).此外, 全球气候变化的趋势预示: 随着较高温度的出现, 高山地区降雪在近几十年将会下降(Serreze & Hurst, 2000 ; Beniston, 2005 ), 导致土壤冻结的增加, 而雪被厚度、融雪时间与生长季长度也将随之发生变化.因此, 雪被提供了季节性的复杂环境梯度(Körner, 2003 ; Pauli et al ., 2015 ), 深刻影响着高山植物的生长发育, 进而影响着高山生态系统的结构与功能(Wipf, 2010 ).然而, 雪被厚度及积雪消融时间长期形成的局域尺度的生境差异及其对植物叶片性状、生物量分配乃至群落结构的作用, 以及叶片性状与具体功能的相关性如何呈现等问题尚不明确, 更是缺乏相关的数据和证据支持. ...
Problems of Life: An Evaluation of Modern Biological and Scientific Thought. Harper
1
1952
... 植物功能性状与环境的关系是近年来生态学研究的热点, 尤其是植物的叶片性状具有较强的形态结构可塑性, 能够直接反映植物对环境变化的适应策略, 与资源获取和利用有着密切联系(Westoby & Wright, 2003 ; Rozendaal et al ., 2006 ; Bernard- Verdier et al ., 2012 ; 史元春等, 2015 ).植物的株高与其竞争力和繁殖活力有关(Cornelissen et al ., 2003 ), 增加对自身株高的投资能够提高光照的获取效率, 同时也增加了构成和维持支撑结构的投资(Falster & Westoby, 2003 ).生物量分配指植物生长过程中将同化的资源分配到根、茎、叶、花等器官中, 是植物重要的资源配置方式及净碳获取的驱动因子, 也是植物生存与生殖权衡的结果(方炎明, 1996 ; 郝虎东等, 2009 ).在资源匮乏或超过其利用上限的生境, 植物为了规避风险和增强对环境的适应性, 调整其功能性状等可塑性表型及生物量的分配格局来提高生存适合度(史元春等, 2015 ).异质环境中或不同的选择压力下, 现有关于植物功能性状和生物量分配的研究主要侧重于某一方面, 两者之间是否存在共轭变化, 即按照一定的规律或者关联性来匹配, 并且具有协同的动态变化, 目前的直接证据和定量化研究相对较少.因此, 定量化研究不同物种在不同生境下的叶片性状、株高与生物量分配格局的变化及其相互之间的关联性, 有助于理解全球变化背景下环境因子对植物相关功能的影响. ...
五种高山植物的个体生长在融雪梯度上的变化
1
2009
... 异速生长揭示了某种生物或某一类生物的部分与整体或部分与部分间的差异和对比(Niklas & Enquist, 2001 ; Niklas, 2005 ; 李春萍等, 2007 ).植物的异速生长关系是其各器官相对生长速率可塑性的具体体现, 与植物的生物量分配、资源利用及对异质环境的形态适应有着密切关系(Bertalanffy, 1952 ; 陆霞梅等, 2007 ).植物生物量的分配是遗传和自然选择共同作用的结果(Weiner, 2004 ), 既具有物种特异性, 也受环境的影响.在厚雪和浅雪地段, 紫罗兰报春的地上/地下生物量表现出相同的异速生长关系, 且地下生物量的积累速率大于地上生物量的积累速率, 更多生物量分配到地下形成根状茎和肉质根, 以更好地完成生殖生长.甘肃马先蒿和高原毛茛在某些地段地上-地下生物量回归直线并不显著, 可能是样本量不足或者采样过程中样本的损耗造成.本研究结果与陶冶和张元明(2014)对6种类短命植物的地上-地下生物量的相关生长关系的研究结果较为一致, 均不支持Enquist和Niklas (2002)的等速生长假说.总体而言, 三种植物整体样本的地上-地下生物量分配均为异速生长关系, 在厚雪和浅雪地段具有共同的斜率且显著小于1, 地上生物量的积累快于地下, 而在中雪地段则相反, 生长季土壤温度在厚雪和浅雪地段相差不大且均高于中雪.另外, 整体样本中植物地上生物量可以较好地解释地下生物量的变化(表6 ).通过植物的地上部分生物量估算地下部分生物量, 可以探究整个区域内生物量对环境梯度变化的响应. ...
a). 3种高山植物的物候和种群分布格局在融雪梯度上的变化
1
2011
... 在距山脊最高处50 m左右的斜坡(坡度变化8°-15°)朝南沿坡向下设置3条样带(11 m × 5 m), 相邻样带间距10 m左右.三条样带的垂直高程差为0.51 m, 从雪床的上端到下端, 雪被厚度自然增加, 融雪时间逐渐推迟, 形成了一个自然的融雪梯度(陈文年等, 2011a ).雪被厚度的监测从稳定积雪开始, 积雪消融殆尽结束, 每3天一次, 可根据天气情况适当调整.依据样带的雪被厚度监测数据(表1 )将其划分为厚雪、中雪和浅雪, 最大雪被厚度即为监测期间雪被的最大厚度, 其中浅雪形成时间较晚而消融时间较早, 最大雪被厚度较小, 厚雪则正好相反, 其最大厚度高于浅雪的2倍, 中雪梯度居中.土壤温度采用钮扣式温度计长期监测(置于距土表以下5 cm深处).选取样带中广泛分布的3种早春优势种高原毛茛、紫罗兰报春和甘肃马先蒿为研究对象.高原毛茛和紫罗兰报春均为多年生草本, 后者多分布于潮湿的生境, 甘肃马先蒿为一年生或两年生草本(中国科学院中国植物志编辑委员会, 1963 , 1980, 1990 ). ...
b). 融雪时间对大卫马先蒿生长和繁殖特性的影响
3
2011
... 雪被广泛分布在寒冷生命带, 其积累与融化的交替过程直接影响着土壤水分、土壤温度、光照条件以及养分状况, 使得植物在生长节律、繁殖对策和生理生态方面形成一系列独特的适应机制(Jones et al ., 2001 ; 吴彦, 2005 ; 吴彦和Onipchenko, 2007 ).目前, 雪被生态学的研究主要集中在雪被对植物的地理分布、物质循环和土壤微生物动态的影响等方面(陈文年等, 2009 ; 胡霞等, 2012 ; 杨玉莲等, 2012 ; 何伟等, 2013 ).大量研究证据显示1980年以来北半球中高纬度的雪被面积减少了10% (Groisman et al ., 1994 ; Serreze & Hurst, 2000 ; Dye, 2002 ; IPCC, 2001 ).此外, 全球气候变化的趋势预示: 随着较高温度的出现, 高山地区降雪在近几十年将会下降(Serreze & Hurst, 2000 ; Beniston, 2005 ), 导致土壤冻结的增加, 而雪被厚度、融雪时间与生长季长度也将随之发生变化.因此, 雪被提供了季节性的复杂环境梯度(Körner, 2003 ; Pauli et al ., 2015 ), 深刻影响着高山植物的生长发育, 进而影响着高山生态系统的结构与功能(Wipf, 2010 ).然而, 雪被厚度及积雪消融时间长期形成的局域尺度的生境差异及其对植物叶片性状、生物量分配乃至群落结构的作用, 以及叶片性状与具体功能的相关性如何呈现等问题尚不明确, 更是缺乏相关的数据和证据支持. ...
... 由于地形、风和光照等环境因素的作用, 季节性雪被覆盖地区的雪被厚度存在明显的斑块性差异, 相关的融雪时间及植物生长季长度也随之变化, 导致雪下光照、温度和水分等环境因子再分配, 影响着雪下土壤动物及微生物等生物因子(Wijk, 1986 ; Lipson et al ., 2002 ; Totland & Alatalo, 2002 ; Rixen et al ., 2004 ; 陈文年等, 2011b ; 杨玉莲等, 2012 ), 进而作用于植物的功能性状和生物量分配格局.持续雪被覆盖时期, 厚雪地段土壤日平均温度相对较高, 波动幅度也最小, 土壤微生物依然具有较强的活力, 依然进行着氮矿化过程(Schimel et al ., 2004 ; Hiltbrunner et al ., 2005 ), 为早春植物的生长储备养分, 因此厚雪地段土壤有机碳含量、全氮含量高于中雪和浅雪地段.不同雪被地段土壤体积含水量均在60%以上(表2 ), 可见水分并不构成限制因子, 热量和土壤养分是影响高山植物生长的主要因子(刘琪璟等, 2009 ).综合来看, 厚雪地段土壤状况最优, 中雪地段次之, 浅雪地段相对较差.浅雪地段融雪较早, 植物易受到早期冻融交替或早霜胁迫的影响, 甘肃马先蒿和高原毛茛采取适应性策略, 开花物候有所迟滞.比叶面积-株高之间标准化主轴回归分析(SMA)的结果也表明, 当株高一定时, 浅雪地段紫罗兰报春的比叶面积高于厚雪地段, 厚雪地段甘肃马先蒿的比叶面积则高于浅雪地段, 而与中雪地段无显著差异(图2 ).尽管在不同雪被地段高原毛茛的比叶面积-株高间的回归直线斜率具有显著差异, 但中雪地段的植株比叶面积仍然高于浅雪地段.结果表明, 在土壤状况较好的地段, 甘肃马先蒿和高原毛茛具有相对较大的比叶面积.在厚雪和浅雪地段, 紫罗兰报春的株高无显著差异, 而浅雪地段植株的比叶面积高于厚雪地段.可能由于紫罗兰报春是多年生植物且具有相对粗短的根状茎和肉质长根, 厚雪地段植株的地下部分储存相对较多的营养物质以供生殖生长利用, 而浅雪地段植株主要依赖光合作用生产碳水化合物. ...
... 植物通过调节自身的资源配置以适应特定的生境, 形成对一定的环境变化及其复杂性和多样性的适应机制, 在环境变化及胁迫因子作用下具有各自的生长特性和物质分配规律(肖遥等, 2014 ).生物量分配主要取决于植物的遗传特性, 同时也受环境条件的影响(郝虎东等, 2009 ; Poorter et al ., 2012 ).不同的雪被地段, 高山草本植物各器官的生物量如何分配以适应环境的变化, 是研究高山植物生存策略的重要内容.本研究中, 不同雪被厚度覆盖地段下, 三种早春草本植物生物量分配的变化具有物种特异性.在土壤温度及养分状况更优的厚雪地段, 紫罗兰报春总生物量是浅雪地段植株的2倍多.厚雪地段的植株生长速率较高, 可将部分光合产物储存在根状茎和肉质长根中, 为下一阶段的生殖生长积蓄物质和能量, 而浅雪地段的植株生长环境较差, 需要将生物量更多地分配到叶中, 从而使碳收获最大化并达到生殖阈值.厚雪和中雪地段的甘肃马先蒿各部分器官生物量均高于浅雪地段植株, 而根和叶生物量分配比例均在浅雪地段最大, 既能够吸收较多养分, 又可以提高养分利用效率, 有利于其生殖生长.在生境状况相对较好的中雪地段, 高原毛茛各部分器官生物量反而小于浅雪地段植株, 可能是由于植物在浅雪地段具有更长的生长期, 可积累较多的生物量, 但伴随着雪被融化, 也面临着失去雪被保护、遭受霜冻和水分胁迫的风险(陈文年等, 2011b ), 并且浅雪地段土壤养分相对较低, 该地段高原毛茛分配更多的地下生物量, 因此生物量更大的浅雪地段高原毛茛进入生殖阶段晚于中雪地段植株. ...
A handbook of protocols for standardised and easy measurement of plant functional traits worldwide.
2003
1
... 在距山脊最高处50 m左右的斜坡(坡度变化8°-15°)朝南沿坡向下设置3条样带(11 m × 5 m), 相邻样带间距10 m左右.三条样带的垂直高程差为0.51 m, 从雪床的上端到下端, 雪被厚度自然增加, 融雪时间逐渐推迟, 形成了一个自然的融雪梯度(陈文年等, 2011a ).雪被厚度的监测从稳定积雪开始, 积雪消融殆尽结束, 每3天一次, 可根据天气情况适当调整.依据样带的雪被厚度监测数据(表1 )将其划分为厚雪、中雪和浅雪, 最大雪被厚度即为监测期间雪被的最大厚度, 其中浅雪形成时间较晚而消融时间较早, 最大雪被厚度较小, 厚雪则正好相反, 其最大厚度高于浅雪的2倍, 中雪梯度居中.土壤温度采用钮扣式温度计长期监测(置于距土表以下5 cm深处).选取样带中广泛分布的3种早春优势种高原毛茛、紫罗兰报春和甘肃马先蒿为研究对象.高原毛茛和紫罗兰报春均为多年生草本, 后者多分布于潮湿的生境, 甘肃马先蒿为一年生或两年生草本(中国科学院中国植物志编辑委员会, 1963 , 1980, 1990 ). ...
1
... 在距山脊最高处50 m左右的斜坡(坡度变化8°-15°)朝南沿坡向下设置3条样带(11 m × 5 m), 相邻样带间距10 m左右.三条样带的垂直高程差为0.51 m, 从雪床的上端到下端, 雪被厚度自然增加, 融雪时间逐渐推迟, 形成了一个自然的融雪梯度(陈文年等, 2011a ).雪被厚度的监测从稳定积雪开始, 积雪消融殆尽结束, 每3天一次, 可根据天气情况适当调整.依据样带的雪被厚度监测数据(表1 )将其划分为厚雪、中雪和浅雪, 最大雪被厚度即为监测期间雪被的最大厚度, 其中浅雪形成时间较晚而消融时间较早, 最大雪被厚度较小, 厚雪则正好相反, 其最大厚度高于浅雪的2倍, 中雪梯度居中.土壤温度采用钮扣式温度计长期监测(置于距土表以下5 cm深处).选取样带中广泛分布的3种早春优势种高原毛茛、紫罗兰报春和甘肃马先蒿为研究对象.高原毛茛和紫罗兰报春均为多年生草本, 后者多分布于潮湿的生境, 甘肃马先蒿为一年生或两年生草本(中国科学院中国植物志编辑委员会, 1963 , 1980, 1990 ). ...
岷江百合的生物量分配对策及其海拔效应
1
2014
... 当受到胁迫或资源限制时, 植物会适当地调整其功能性状和生物量分配, 从而提高竞争力及其生存适合度(Sultan, 1992 ; 武高林和杜国祯, 2007 ).例如养分不足时, 植物会将生物量更多地分配到地下, 而减少地上部分的生物量分配, 植物的比叶面积和株高等功能性状也会相应减小(武高林等, 2010 ), 因此, 植物功能性状和生物量分配之间存在着某种共轭的变化.这种相互关系对于量化和概括植物面对生境变化的权衡策略及其规律有重要意义(Wright et al ., 2004 ; 陈莹婷和许振柱, 2014 ).本研究中, 紫罗兰报春(主要是株高和比叶面积与生物量指标之间)的相关性较强, 而这种相关性在甘肃马先蒿和高原毛茛相对较弱.由于生活型和个体发育的差异, 不同的物种在面临相同环境变化时, 权衡策略具有物种特异性, 可表征生物量变化的功能性状亦有所不同.因此, 如何选取具有普适性的性状, 表征不同尺度生物量的变化需要以后深入研究, 以丰富中国的植物性状数据库, 并更好地理解全球变化背景下植物功能性状的响应和适应对策. ...
Variability and trends in the annual snow-cover cycle in northern hemisphere land areas, 1972-2000.
1
2002
... 植物功能性状与环境的关系是近年来生态学研究的热点, 尤其是植物的叶片性状具有较强的形态结构可塑性, 能够直接反映植物对环境变化的适应策略, 与资源获取和利用有着密切联系(Westoby & Wright, 2003 ; Rozendaal et al ., 2006 ; Bernard- Verdier et al ., 2012 ; 史元春等, 2015 ).植物的株高与其竞争力和繁殖活力有关(Cornelissen et al ., 2003 ), 增加对自身株高的投资能够提高光照的获取效率, 同时也增加了构成和维持支撑结构的投资(Falster & Westoby, 2003 ).生物量分配指植物生长过程中将同化的资源分配到根、茎、叶、花等器官中, 是植物重要的资源配置方式及净碳获取的驱动因子, 也是植物生存与生殖权衡的结果(方炎明, 1996 ; 郝虎东等, 2009 ).在资源匮乏或超过其利用上限的生境, 植物为了规避风险和增强对环境的适应性, 调整其功能性状等可塑性表型及生物量的分配格局来提高生存适合度(史元春等, 2015 ).异质环境中或不同的选择压力下, 现有关于植物功能性状和生物量分配的研究主要侧重于某一方面, 两者之间是否存在共轭变化, 即按照一定的规律或者关联性来匹配, 并且具有协同的动态变化, 目前的直接证据和定量化研究相对较少.因此, 定量化研究不同物种在不同生境下的叶片性状、株高与生物量分配格局的变化及其相互之间的关联性, 有助于理解全球变化背景下环境因子对植物相关功能的影响. ...
Global allocation rules for patterns of biomass partitioning in seed plants.
2002
1
... 根据相关生长模型y = βxα 对不同雪被厚度下的三种草本植物地上/地下生物量及比叶面积-株高进行相关生长分析.分析前对生物量数据进行log10 的对数转换使之符合正态分布.方程可转化为lgy = lgβ + α lgx, x 和y 分别表示地上/地下生物量, β 为标准化常数, α 为异速生长指数.α = 1为等速生长关系, α ≠1为异速生长关系(Niklas & Enquist, 2001 ; Niklas, 2005 ; 丁建林等, 2014 ; 肖遥等, 2014 ).根据Warton和Weber (2002)的方法对同一物种在不同雪被厚度的α 进行异质性检验, 如果α 没有异质性, 则给出一个共同斜率, 若α 不同, 则异速生长关系完全不同, α 相同继续进行截距漂移检验和共轴漂移检验.若截距不同但共轴, 则属于截距漂移; 若截距相同且发生共轴漂移, 则属于共轴漂移; 若截距相同也同轴, 则表明植物的地上/地下生物量在不同雪被厚度下具有完全相同的相关生长关系(Falster et al ., 2006 ; 陶冶和张元明, 2014 ).相关生长模型的参数确定, 采用标准化主轴估计(standardized major axis estimation, SMA)方法(Wright et al ., 2001 ; Warton et al ., 2006 ), 截距漂移检验、共轴漂移检验及α 的方差分析均利用软件SMATR Version 2.0完成. ...
Plant height and evolutionary games.
2003
1
... 植物功能性状与环境的关系是近年来生态学研究的热点, 尤其是植物的叶片性状具有较强的形态结构可塑性, 能够直接反映植物对环境变化的适应策略, 与资源获取和利用有着密切联系(Westoby & Wright, 2003 ; Rozendaal et al ., 2006 ; Bernard- Verdier et al ., 2012 ; 史元春等, 2015 ).植物的株高与其竞争力和繁殖活力有关(Cornelissen et al ., 2003 ), 增加对自身株高的投资能够提高光照的获取效率, 同时也增加了构成和维持支撑结构的投资(Falster & Westoby, 2003 ).生物量分配指植物生长过程中将同化的资源分配到根、茎、叶、花等器官中, 是植物重要的资源配置方式及净碳获取的驱动因子, 也是植物生存与生殖权衡的结果(方炎明, 1996 ; 郝虎东等, 2009 ).在资源匮乏或超过其利用上限的生境, 植物为了规避风险和增强对环境的适应性, 调整其功能性状等可塑性表型及生物量的分配格局来提高生存适合度(史元春等, 2015 ).异质环境中或不同的选择压力下, 现有关于植物功能性状和生物量分配的研究主要侧重于某一方面, 两者之间是否存在共轭变化, 即按照一定的规律或者关联性来匹配, 并且具有协同的动态变化, 目前的直接证据和定量化研究相对较少.因此, 定量化研究不同物种在不同生境下的叶片性状、株高与生物量分配格局的变化及其相互之间的关联性, 有助于理解全球变化背景下环境因子对植物相关功能的影响. ...
Changes of snow cover, temperature, and radiative heat- balance over the northern-hemisphere.
1994
无芒雀麦地上生物量及各构件生物量分配动态
2009
雪被斑块对高山森林两种灌木凋落叶质量损失的影响
2013
Inorganic nitrogen storage in alpine snow pack in the Central Alps (Switzerland).
1
2005
... 根据相关生长模型y = βxα 对不同雪被厚度下的三种草本植物地上/地下生物量及比叶面积-株高进行相关生长分析.分析前对生物量数据进行log10 的对数转换使之符合正态分布.方程可转化为lgy = lgβ + α lgx, x 和y 分别表示地上/地下生物量, β 为标准化常数, α 为异速生长指数.α = 1为等速生长关系, α ≠1为异速生长关系(Niklas & Enquist, 2001 ; Niklas, 2005 ; 丁建林等, 2014 ; 肖遥等, 2014 ).根据Warton和Weber (2002)的方法对同一物种在不同雪被厚度的α 进行异质性检验, 如果α 没有异质性, 则给出一个共同斜率, 若α 不同, 则异速生长关系完全不同, α 相同继续进行截距漂移检验和共轴漂移检验.若截距不同但共轴, 则属于截距漂移; 若截距相同且发生共轴漂移, 则属于共轴漂移; 若截距相同也同轴, 则表明植物的地上/地下生物量在不同雪被厚度下具有完全相同的相关生长关系(Falster et al ., 2006 ; 陶冶和张元明, 2014 ).相关生长模型的参数确定, 采用标准化主轴估计(standardized major axis estimation, SMA)方法(Wright et al ., 2001 ; Warton et al ., 2006 ), 截距漂移检验、共轴漂移检验及α 的方差分析均利用软件SMATR Version 2.0完成. ...
川西高原季节性雪被覆盖对窄叶鲜卑花凋落物分解和养分动态的影响
1
2012
... 雪被广泛分布在寒冷生命带, 其积累与融化的交替过程直接影响着土壤水分、土壤温度、光照条件以及养分状况, 使得植物在生长节律、繁殖对策和生理生态方面形成一系列独特的适应机制(Jones et al ., 2001 ; 吴彦, 2005 ; 吴彦和Onipchenko, 2007 ).目前, 雪被生态学的研究主要集中在雪被对植物的地理分布、物质循环和土壤微生物动态的影响等方面(陈文年等, 2009 ; 胡霞等, 2012 ; 杨玉莲等, 2012 ; 何伟等, 2013 ).大量研究证据显示1980年以来北半球中高纬度的雪被面积减少了10% (Groisman et al ., 1994 ; Serreze & Hurst, 2000 ; Dye, 2002 ; IPCC, 2001 ).此外, 全球气候变化的趋势预示: 随着较高温度的出现, 高山地区降雪在近几十年将会下降(Serreze & Hurst, 2000 ; Beniston, 2005 ), 导致土壤冻结的增加, 而雪被厚度、融雪时间与生长季长度也将随之发生变化.因此, 雪被提供了季节性的复杂环境梯度(Körner, 2003 ; Pauli et al ., 2015 ), 深刻影响着高山植物的生长发育, 进而影响着高山生态系统的结构与功能(Wipf, 2010 ).然而, 雪被厚度及积雪消融时间长期形成的局域尺度的生境差异及其对植物叶片性状、生物量分配乃至群落结构的作用, 以及叶片性状与具体功能的相关性如何呈现等问题尚不明确, 更是缺乏相关的数据和证据支持. ...
1
... 雪被广泛分布在寒冷生命带, 其积累与融化的交替过程直接影响着土壤水分、土壤温度、光照条件以及养分状况, 使得植物在生长节律、繁殖对策和生理生态方面形成一系列独特的适应机制(Jones et al ., 2001 ; 吴彦, 2005 ; 吴彦和Onipchenko, 2007 ).目前, 雪被生态学的研究主要集中在雪被对植物的地理分布、物质循环和土壤微生物动态的影响等方面(陈文年等, 2009 ; 胡霞等, 2012 ; 杨玉莲等, 2012 ; 何伟等, 2013 ).大量研究证据显示1980年以来北半球中高纬度的雪被面积减少了10% (Groisman et al ., 1994 ; Serreze & Hurst, 2000 ; Dye, 2002 ; IPCC, 2001 ).此外, 全球气候变化的趋势预示: 随着较高温度的出现, 高山地区降雪在近几十年将会下降(Serreze & Hurst, 2000 ; Beniston, 2005 ), 导致土壤冻结的增加, 而雪被厚度、融雪时间与生长季长度也将随之发生变化.因此, 雪被提供了季节性的复杂环境梯度(Körner, 2003 ; Pauli et al ., 2015 ), 深刻影响着高山植物的生长发育, 进而影响着高山生态系统的结构与功能(Wipf, 2010 ).然而, 雪被厚度及积雪消融时间长期形成的局域尺度的生境差异及其对植物叶片性状、生物量分配乃至群落结构的作用, 以及叶片性状与具体功能的相关性如何呈现等问题尚不明确, 更是缺乏相关的数据和证据支持. ...
2
... 植物功能性状与环境的关系是近年来生态学研究的热点, 尤其是植物的叶片性状具有较强的形态结构可塑性, 能够直接反映植物对环境变化的适应策略, 与资源获取和利用有着密切联系(Westoby & Wright, 2003 ; Rozendaal et al ., 2006 ; Bernard- Verdier et al ., 2012 ; 史元春等, 2015 ).植物的株高与其竞争力和繁殖活力有关(Cornelissen et al ., 2003 ), 增加对自身株高的投资能够提高光照的获取效率, 同时也增加了构成和维持支撑结构的投资(Falster & Westoby, 2003 ).生物量分配指植物生长过程中将同化的资源分配到根、茎、叶、花等器官中, 是植物重要的资源配置方式及净碳获取的驱动因子, 也是植物生存与生殖权衡的结果(方炎明, 1996 ; 郝虎东等, 2009 ).在资源匮乏或超过其利用上限的生境, 植物为了规避风险和增强对环境的适应性, 调整其功能性状等可塑性表型及生物量的分配格局来提高生存适合度(史元春等, 2015 ).异质环境中或不同的选择压力下, 现有关于植物功能性状和生物量分配的研究主要侧重于某一方面, 两者之间是否存在共轭变化, 即按照一定的规律或者关联性来匹配, 并且具有协同的动态变化, 目前的直接证据和定量化研究相对较少.因此, 定量化研究不同物种在不同生境下的叶片性状、株高与生物量分配格局的变化及其相互之间的关联性, 有助于理解全球变化背景下环境因子对植物相关功能的影响. ...
... 雪被广泛分布在寒冷生命带, 其积累与融化的交替过程直接影响着土壤水分、土壤温度、光照条件以及养分状况, 使得植物在生长节律、繁殖对策和生理生态方面形成一系列独特的适应机制(Jones et al ., 2001 ; 吴彦, 2005 ; 吴彦和Onipchenko, 2007 ).目前, 雪被生态学的研究主要集中在雪被对植物的地理分布、物质循环和土壤微生物动态的影响等方面(陈文年等, 2009 ; 胡霞等, 2012 ; 杨玉莲等, 2012 ; 何伟等, 2013 ).大量研究证据显示1980年以来北半球中高纬度的雪被面积减少了10% (Groisman et al ., 1994 ; Serreze & Hurst, 2000 ; Dye, 2002 ; IPCC, 2001 ).此外, 全球气候变化的趋势预示: 随着较高温度的出现, 高山地区降雪在近几十年将会下降(Serreze & Hurst, 2000 ; Beniston, 2005 ), 导致土壤冻结的增加, 而雪被厚度、融雪时间与生长季长度也将随之发生变化.因此, 雪被提供了季节性的复杂环境梯度(Körner, 2003 ; Pauli et al ., 2015 ), 深刻影响着高山植物的生长发育, 进而影响着高山生态系统的结构与功能(Wipf, 2010 ).然而, 雪被厚度及积雪消融时间长期形成的局域尺度的生境差异及其对植物叶片性状、生物量分配乃至群落结构的作用, 以及叶片性状与具体功能的相关性如何呈现等问题尚不明确, 更是缺乏相关的数据和证据支持. ...
Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems. Springer
2003
异速生长关系在陆地生态系统生物量估测中的应用
2007
不同沙丘生境主要植物比叶面积和叶干物质含量的比较
1
2005
... 雪被广泛分布在寒冷生命带, 其积累与融化的交替过程直接影响着土壤水分、土壤温度、光照条件以及养分状况, 使得植物在生长节律、繁殖对策和生理生态方面形成一系列独特的适应机制(Jones et al ., 2001 ; 吴彦, 2005 ; 吴彦和Onipchenko, 2007 ).目前, 雪被生态学的研究主要集中在雪被对植物的地理分布、物质循环和土壤微生物动态的影响等方面(陈文年等, 2009 ; 胡霞等, 2012 ; 杨玉莲等, 2012 ; 何伟等, 2013 ).大量研究证据显示1980年以来北半球中高纬度的雪被面积减少了10% (Groisman et al ., 1994 ; Serreze & Hurst, 2000 ; Dye, 2002 ; IPCC, 2001 ).此外, 全球气候变化的趋势预示: 随着较高温度的出现, 高山地区降雪在近几十年将会下降(Serreze & Hurst, 2000 ; Beniston, 2005 ), 导致土壤冻结的增加, 而雪被厚度、融雪时间与生长季长度也将随之发生变化.因此, 雪被提供了季节性的复杂环境梯度(Körner, 2003 ; Pauli et al ., 2015 ), 深刻影响着高山植物的生长发育, 进而影响着高山生态系统的结构与功能(Wipf, 2010 ).然而, 雪被厚度及积雪消融时间长期形成的局域尺度的生境差异及其对植物叶片性状、生物量分配乃至群落结构的作用, 以及叶片性状与具体功能的相关性如何呈现等问题尚不明确, 更是缺乏相关的数据和证据支持. ...
Changes in soil microbial community structure and function in an alpine dry meadow following spring snow melt.
2002
Effects of freezing and freeze-thaw cycles on soil microbial biomass and nutrient dynamics under different snow gradients in an alpine meadow (Tibetan Plateau).
2
2010
... 植物功能性状与环境的关系是近年来生态学研究的热点, 尤其是植物的叶片性状具有较强的形态结构可塑性, 能够直接反映植物对环境变化的适应策略, 与资源获取和利用有着密切联系(Westoby & Wright, 2003 ; Rozendaal et al ., 2006 ; Bernard- Verdier et al ., 2012 ; 史元春等, 2015 ).植物的株高与其竞争力和繁殖活力有关(Cornelissen et al ., 2003 ), 增加对自身株高的投资能够提高光照的获取效率, 同时也增加了构成和维持支撑结构的投资(Falster & Westoby, 2003 ).生物量分配指植物生长过程中将同化的资源分配到根、茎、叶、花等器官中, 是植物重要的资源配置方式及净碳获取的驱动因子, 也是植物生存与生殖权衡的结果(方炎明, 1996 ; 郝虎东等, 2009 ).在资源匮乏或超过其利用上限的生境, 植物为了规避风险和增强对环境的适应性, 调整其功能性状等可塑性表型及生物量的分配格局来提高生存适合度(史元春等, 2015 ).异质环境中或不同的选择压力下, 现有关于植物功能性状和生物量分配的研究主要侧重于某一方面, 两者之间是否存在共轭变化, 即按照一定的规律或者关联性来匹配, 并且具有协同的动态变化, 目前的直接证据和定量化研究相对较少.因此, 定量化研究不同物种在不同生境下的叶片性状、株高与生物量分配格局的变化及其相互之间的关联性, 有助于理解全球变化背景下环境因子对植物相关功能的影响. ...
... 植物通过调节自身的资源配置以适应特定的生境, 形成对一定的环境变化及其复杂性和多样性的适应机制, 在环境变化及胁迫因子作用下具有各自的生长特性和物质分配规律(肖遥等, 2014 ).生物量分配主要取决于植物的遗传特性, 同时也受环境条件的影响(郝虎东等, 2009 ; Poorter et al ., 2012 ).不同的雪被地段, 高山草本植物各器官的生物量如何分配以适应环境的变化, 是研究高山植物生存策略的重要内容.本研究中, 不同雪被厚度覆盖地段下, 三种早春草本植物生物量分配的变化具有物种特异性.在土壤温度及养分状况更优的厚雪地段, 紫罗兰报春总生物量是浅雪地段植株的2倍多.厚雪地段的植株生长速率较高, 可将部分光合产物储存在根状茎和肉质长根中, 为下一阶段的生殖生长积蓄物质和能量, 而浅雪地段的植株生长环境较差, 需要将生物量更多地分配到叶中, 从而使碳收获最大化并达到生殖阈值.厚雪和中雪地段的甘肃马先蒿各部分器官生物量均高于浅雪地段植株, 而根和叶生物量分配比例均在浅雪地段最大, 既能够吸收较多养分, 又可以提高养分利用效率, 有利于其生殖生长.在生境状况相对较好的中雪地段, 高原毛茛各部分器官生物量反而小于浅雪地段植株, 可能是由于植物在浅雪地段具有更长的生长期, 可积累较多的生物量, 但伴随着雪被融化, 也面临着失去雪被保护、遭受霜冻和水分胁迫的风险(陈文年等, 2011b ), 并且浅雪地段土壤养分相对较低, 该地段高原毛茛分配更多的地下生物量, 因此生物量更大的浅雪地段高原毛茛进入生殖阶段晚于中雪地段植株. ...
植物的表型可塑性、异速生长及其入侵能力
1
2007
... 雪被广泛分布在寒冷生命带, 其积累与融化的交替过程直接影响着土壤水分、土壤温度、光照条件以及养分状况, 使得植物在生长节律、繁殖对策和生理生态方面形成一系列独特的适应机制(Jones et al ., 2001 ; 吴彦, 2005 ; 吴彦和Onipchenko, 2007 ).目前, 雪被生态学的研究主要集中在雪被对植物的地理分布、物质循环和土壤微生物动态的影响等方面(陈文年等, 2009 ; 胡霞等, 2012 ; 杨玉莲等, 2012 ; 何伟等, 2013 ).大量研究证据显示1980年以来北半球中高纬度的雪被面积减少了10% (Groisman et al ., 1994 ; Serreze & Hurst, 2000 ; Dye, 2002 ; IPCC, 2001 ).此外, 全球气候变化的趋势预示: 随着较高温度的出现, 高山地区降雪在近几十年将会下降(Serreze & Hurst, 2000 ; Beniston, 2005 ), 导致土壤冻结的增加, 而雪被厚度、融雪时间与生长季长度也将随之发生变化.因此, 雪被提供了季节性的复杂环境梯度(Körner, 2003 ; Pauli et al ., 2015 ), 深刻影响着高山植物的生长发育, 进而影响着高山生态系统的结构与功能(Wipf, 2010 ).然而, 雪被厚度及积雪消融时间长期形成的局域尺度的生境差异及其对植物叶片性状、生物量分配乃至群落结构的作用, 以及叶片性状与具体功能的相关性如何呈现等问题尚不明确, 更是缺乏相关的数据和证据支持. ...
Modelling below- and above-ground biomass for non-woody and woody plants.
1
2005
... 由于地形、风和光照等环境因素的作用, 季节性雪被覆盖地区的雪被厚度存在明显的斑块性差异, 相关的融雪时间及植物生长季长度也随之变化, 导致雪下光照、温度和水分等环境因子再分配, 影响着雪下土壤动物及微生物等生物因子(Wijk, 1986 ; Lipson et al ., 2002 ; Totland & Alatalo, 2002 ; Rixen et al ., 2004 ; 陈文年等, 2011b ; 杨玉莲等, 2012 ), 进而作用于植物的功能性状和生物量分配格局.持续雪被覆盖时期, 厚雪地段土壤日平均温度相对较高, 波动幅度也最小, 土壤微生物依然具有较强的活力, 依然进行着氮矿化过程(Schimel et al ., 2004 ; Hiltbrunner et al ., 2005 ), 为早春植物的生长储备养分, 因此厚雪地段土壤有机碳含量、全氮含量高于中雪和浅雪地段.不同雪被地段土壤体积含水量均在60%以上(表2 ), 可见水分并不构成限制因子, 热量和土壤养分是影响高山植物生长的主要因子(刘琪璟等, 2009 ).综合来看, 厚雪地段土壤状况最优, 中雪地段次之, 浅雪地段相对较差.浅雪地段融雪较早, 植物易受到早期冻融交替或早霜胁迫的影响, 甘肃马先蒿和高原毛茛采取适应性策略, 开花物候有所迟滞.比叶面积-株高之间标准化主轴回归分析(SMA)的结果也表明, 当株高一定时, 浅雪地段紫罗兰报春的比叶面积高于厚雪地段, 厚雪地段甘肃马先蒿的比叶面积则高于浅雪地段, 而与中雪地段无显著差异(图2 ).尽管在不同雪被地段高原毛茛的比叶面积-株高间的回归直线斜率具有显著差异, 但中雪地段的植株比叶面积仍然高于浅雪地段.结果表明, 在土壤状况较好的地段, 甘肃马先蒿和高原毛茛具有相对较大的比叶面积.在厚雪和浅雪地段, 紫罗兰报春的株高无显著差异, 而浅雪地段植株的比叶面积高于厚雪地段.可能由于紫罗兰报春是多年生植物且具有相对粗短的根状茎和肉质长根, 厚雪地段植株的地下部分储存相对较多的营养物质以供生殖生长利用, 而浅雪地段植株主要依赖光合作用生产碳水化合物. ...
Invariant scaling relationships for interspecific plant biomass production rates and body size.
2001
The GLORIA Field Manual—Standard Multi-Summit Approach, Supplementary Methods and Extra Approaches. 5th edn. GLORIA-Coordination, Austrian Academy of Sciences & University of Natural Resources and Life Sciences
3
2015
... 雪被广泛分布在寒冷生命带, 其积累与融化的交替过程直接影响着土壤水分、土壤温度、光照条件以及养分状况, 使得植物在生长节律、繁殖对策和生理生态方面形成一系列独特的适应机制(Jones et al ., 2001 ; 吴彦, 2005 ; 吴彦和Onipchenko, 2007 ).目前, 雪被生态学的研究主要集中在雪被对植物的地理分布、物质循环和土壤微生物动态的影响等方面(陈文年等, 2009 ; 胡霞等, 2012 ; 杨玉莲等, 2012 ; 何伟等, 2013 ).大量研究证据显示1980年以来北半球中高纬度的雪被面积减少了10% (Groisman et al ., 1994 ; Serreze & Hurst, 2000 ; Dye, 2002 ; IPCC, 2001 ).此外, 全球气候变化的趋势预示: 随着较高温度的出现, 高山地区降雪在近几十年将会下降(Serreze & Hurst, 2000 ; Beniston, 2005 ), 导致土壤冻结的增加, 而雪被厚度、融雪时间与生长季长度也将随之发生变化.因此, 雪被提供了季节性的复杂环境梯度(Körner, 2003 ; Pauli et al ., 2015 ), 深刻影响着高山植物的生长发育, 进而影响着高山生态系统的结构与功能(Wipf, 2010 ).然而, 雪被厚度及积雪消融时间长期形成的局域尺度的生境差异及其对植物叶片性状、生物量分配乃至群落结构的作用, 以及叶片性状与具体功能的相关性如何呈现等问题尚不明确, 更是缺乏相关的数据和证据支持. ...
... 试验地位于川西高原松潘县卡卡山(32.98° N, 103.67° E), 海拔3950 m, 属于青藏高原东缘岷山山脉的一部分.该区受西南暖湿气流和东南季风控制, 属于典型的山地季风气候, 年平均气温5.7 ℃, 1月平均气温-7.6 ℃, 7月平均气温9.7 ℃, 无绝对无霜期, 年太阳辐射时数1827.5 h, ≥10 ℃年积温428.6 ℃, 年降水量718 mm, 每年11月底或12月初出现持续的积雪覆盖, 积雪厚度在15 cm以上.土壤为棕色草毡土, 土壤pH值为5.42-5.83, 每kg干土含有有机质44.23-62. 00 g、全氮3.51-5.12 g, 平均土层厚度63 cm (胡霞等, 2012 ; Wang et al ., 2014 ).原生乔木以岷江冷杉(Abies fargesii var. faxoniana )为主; 高山灌木有窄叶鲜卑花(Sibiraea angustata )、 鹧鸪杜鹃(Rhododendron zheguense )和鹧鸪柳(Salix zhegushanica ), 以及零星分布的紫果云杉(Picea purpurea )等; 草本植物主要有黑褐穗薹草(Carex atrofusca )、长花马先蒿(Pedicularis longiflora )、长叶火绒草(Leontopodium longifolium )、高原毛茛、甘肃马先蒿、川西小黄菊(Pyrethrum tatsienense )和紫罗兰报春等(Liu et al ., 2010 ; 胡霞等, 2012 ; Wang et al ., 2014 ). ...
... ; 胡霞等, 2012 ; Wang et al ., 2014 ). ...
Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control.
2012
Ground temperatures under ski pistes with artificial and natural snow.
1
2004
... 雪被广泛分布在寒冷生命带, 其积累与融化的交替过程直接影响着土壤水分、土壤温度、光照条件以及养分状况, 使得植物在生长节律、繁殖对策和生理生态方面形成一系列独特的适应机制(Jones et al ., 2001 ; 吴彦, 2005 ; 吴彦和Onipchenko, 2007 ).目前, 雪被生态学的研究主要集中在雪被对植物的地理分布、物质循环和土壤微生物动态的影响等方面(陈文年等, 2009 ; 胡霞等, 2012 ; 杨玉莲等, 2012 ; 何伟等, 2013 ).大量研究证据显示1980年以来北半球中高纬度的雪被面积减少了10% (Groisman et al ., 1994 ; Serreze & Hurst, 2000 ; Dye, 2002 ; IPCC, 2001 ).此外, 全球气候变化的趋势预示: 随着较高温度的出现, 高山地区降雪在近几十年将会下降(Serreze & Hurst, 2000 ; Beniston, 2005 ), 导致土壤冻结的增加, 而雪被厚度、融雪时间与生长季长度也将随之发生变化.因此, 雪被提供了季节性的复杂环境梯度(Körner, 2003 ; Pauli et al ., 2015 ), 深刻影响着高山植物的生长发育, 进而影响着高山生态系统的结构与功能(Wipf, 2010 ).然而, 雪被厚度及积雪消融时间长期形成的局域尺度的生境差异及其对植物叶片性状、生物量分配乃至群落结构的作用, 以及叶片性状与具体功能的相关性如何呈现等问题尚不明确, 更是缺乏相关的数据和证据支持. ...
Plasticity in leaf traits of 38 tropical tree species in response to light: Relationships with light demand and adult stature.
2006
Increased snow depth affects microbial activity and nitrogen mineralization in two arctic tundra communities.
1
2004
... 异速生长揭示了某种生物或某一类生物的部分与整体或部分与部分间的差异和对比(Niklas & Enquist, 2001 ; Niklas, 2005 ; 李春萍等, 2007 ).植物的异速生长关系是其各器官相对生长速率可塑性的具体体现, 与植物的生物量分配、资源利用及对异质环境的形态适应有着密切关系(Bertalanffy, 1952 ; 陆霞梅等, 2007 ).植物生物量的分配是遗传和自然选择共同作用的结果(Weiner, 2004 ), 既具有物种特异性, 也受环境的影响.在厚雪和浅雪地段, 紫罗兰报春的地上/地下生物量表现出相同的异速生长关系, 且地下生物量的积累速率大于地上生物量的积累速率, 更多生物量分配到地下形成根状茎和肉质根, 以更好地完成生殖生长.甘肃马先蒿和高原毛茛在某些地段地上-地下生物量回归直线并不显著, 可能是样本量不足或者采样过程中样本的损耗造成.本研究结果与陶冶和张元明(2014)对6种类短命植物的地上-地下生物量的相关生长关系的研究结果较为一致, 均不支持Enquist和Niklas (2002)的等速生长假说.总体而言, 三种植物整体样本的地上-地下生物量分配均为异速生长关系, 在厚雪和浅雪地段具有共同的斜率且显著小于1, 地上生物量的积累快于地下, 而在中雪地段则相反, 生长季土壤温度在厚雪和浅雪地段相差不大且均高于中雪.另外, 整体样本中植物地上生物量可以较好地解释地下生物量的变化(表6 ).通过植物的地上部分生物量估算地下部分生物量, 可以探究整个区域内生物量对环境梯度变化的响应. ...
Representation of mean arctic precipitation from NCEP-NCAR and ERA reanalyses.
2000
兰州北山刺槐枝叶性状的坡向差异性
1
2015
... 在每条样带内每个物种随机选取20株左右, 测定其株高(plant height, PH ), 采用全株挖掘法进行取样, 尽可能保证根系完整, 每一植株采集2-3片完全伸展, 且无病虫害、无机械损伤的成熟叶片(李玉霖等, 2005 ).采用平板扫描仪(CanoScan5600F, Canon, Tokyo, Japan)扫描植株鲜叶样品, 然后借助Photoshop CS4和Matlab 7.9计算单叶面积(leaf area, LA ).将所取植株带回实验室, 水浸洗植株地下部分, 将植株分为根、茎、叶、花四类, 分别放入纸袋中, 连同扫描后的叶片于105 ℃杀青30 min后, 在80 ℃条件下烘干至恒质量.用电子天平称量干质量(精度为0.0001 g), 根据Vile等(2005)的方法计算比叶面积(比叶面积=叶面积/叶干质量), 并计算生物量分配, 将花视为繁殖构件. ...
Functional- and abundance-based mechanisms explain diversity loss due to N fertilization.
1
2005
... 由于地形、风和光照等环境因素的作用, 季节性雪被覆盖地区的雪被厚度存在明显的斑块性差异, 相关的融雪时间及植物生长季长度也随之变化, 导致雪下光照、温度和水分等环境因子再分配, 影响着雪下土壤动物及微生物等生物因子(Wijk, 1986 ; Lipson et al ., 2002 ; Totland & Alatalo, 2002 ; Rixen et al ., 2004 ; 陈文年等, 2011b ; 杨玉莲等, 2012 ), 进而作用于植物的功能性状和生物量分配格局.持续雪被覆盖时期, 厚雪地段土壤日平均温度相对较高, 波动幅度也最小, 土壤微生物依然具有较强的活力, 依然进行着氮矿化过程(Schimel et al ., 2004 ; Hiltbrunner et al ., 2005 ), 为早春植物的生长储备养分, 因此厚雪地段土壤有机碳含量、全氮含量高于中雪和浅雪地段.不同雪被地段土壤体积含水量均在60%以上(表2 ), 可见水分并不构成限制因子, 热量和土壤养分是影响高山植物生长的主要因子(刘琪璟等, 2009 ).综合来看, 厚雪地段土壤状况最优, 中雪地段次之, 浅雪地段相对较差.浅雪地段融雪较早, 植物易受到早期冻融交替或早霜胁迫的影响, 甘肃马先蒿和高原毛茛采取适应性策略, 开花物候有所迟滞.比叶面积-株高之间标准化主轴回归分析(SMA)的结果也表明, 当株高一定时, 浅雪地段紫罗兰报春的比叶面积高于厚雪地段, 厚雪地段甘肃马先蒿的比叶面积则高于浅雪地段, 而与中雪地段无显著差异(图2 ).尽管在不同雪被地段高原毛茛的比叶面积-株高间的回归直线斜率具有显著差异, 但中雪地段的植株比叶面积仍然高于浅雪地段.结果表明, 在土壤状况较好的地段, 甘肃马先蒿和高原毛茛具有相对较大的比叶面积.在厚雪和浅雪地段, 紫罗兰报春的株高无显著差异, 而浅雪地段植株的比叶面积高于厚雪地段.可能由于紫罗兰报春是多年生植物且具有相对粗短的根状茎和肉质长根, 厚雪地段植株的地下部分储存相对较多的营养物质以供生殖生长利用, 而浅雪地段植株主要依赖光合作用生产碳水化合物. ...
Phenotypic plasticity and the neo-Darwinian legacy.
1
1992
... 试验地位于川西高原松潘县卡卡山(32.98° N, 103.67° E), 海拔3950 m, 属于青藏高原东缘岷山山脉的一部分.该区受西南暖湿气流和东南季风控制, 属于典型的山地季风气候, 年平均气温5.7 ℃, 1月平均气温-7.6 ℃, 7月平均气温9.7 ℃, 无绝对无霜期, 年太阳辐射时数1827.5 h, ≥10 ℃年积温428.6 ℃, 年降水量718 mm, 每年11月底或12月初出现持续的积雪覆盖, 积雪厚度在15 cm以上.土壤为棕色草毡土, 土壤pH值为5.42-5.83, 每kg干土含有有机质44.23-62. 00 g、全氮3.51-5.12 g, 平均土层厚度63 cm (胡霞等, 2012 ; Wang et al ., 2014 ).原生乔木以岷江冷杉(Abies fargesii var. faxoniana )为主; 高山灌木有窄叶鲜卑花(Sibiraea angustata )、 鹧鸪杜鹃(Rhododendron zheguense )和鹧鸪柳(Salix zhegushanica ), 以及零星分布的紫果云杉(Picea purpurea )等; 草本植物主要有黑褐穗薹草(Carex atrofusca )、长花马先蒿(Pedicularis longiflora )、长叶火绒草(Leontopodium longifolium )、高原毛茛、甘肃马先蒿、川西小黄菊(Pyrethrum tatsienense )和紫罗兰报春等(Liu et al ., 2010 ; 胡霞等, 2012 ; Wang et al ., 2014 ). ...
准噶尔荒漠6种类短命植物生物量分配与异速生长关系
2014
Effects of temperature and date of snowmelt on growth, reproduction, and flowering phenology in the arctic/alpine herb, Ranunculus glacialis.
1
2002
... 由于地形、风和光照等环境因素的作用, 季节性雪被覆盖地区的雪被厚度存在明显的斑块性差异, 相关的融雪时间及植物生长季长度也随之变化, 导致雪下光照、温度和水分等环境因子再分配, 影响着雪下土壤动物及微生物等生物因子(Wijk, 1986 ; Lipson et al ., 2002 ; Totland & Alatalo, 2002 ; Rixen et al ., 2004 ; 陈文年等, 2011b ; 杨玉莲等, 2012 ), 进而作用于植物的功能性状和生物量分配格局.持续雪被覆盖时期, 厚雪地段土壤日平均温度相对较高, 波动幅度也最小, 土壤微生物依然具有较强的活力, 依然进行着氮矿化过程(Schimel et al ., 2004 ; Hiltbrunner et al ., 2005 ), 为早春植物的生长储备养分, 因此厚雪地段土壤有机碳含量、全氮含量高于中雪和浅雪地段.不同雪被地段土壤体积含水量均在60%以上(表2 ), 可见水分并不构成限制因子, 热量和土壤养分是影响高山植物生长的主要因子(刘琪璟等, 2009 ).综合来看, 厚雪地段土壤状况最优, 中雪地段次之, 浅雪地段相对较差.浅雪地段融雪较早, 植物易受到早期冻融交替或早霜胁迫的影响, 甘肃马先蒿和高原毛茛采取适应性策略, 开花物候有所迟滞.比叶面积-株高之间标准化主轴回归分析(SMA)的结果也表明, 当株高一定时, 浅雪地段紫罗兰报春的比叶面积高于厚雪地段, 厚雪地段甘肃马先蒿的比叶面积则高于浅雪地段, 而与中雪地段无显著差异(图2 ).尽管在不同雪被地段高原毛茛的比叶面积-株高间的回归直线斜率具有显著差异, 但中雪地段的植株比叶面积仍然高于浅雪地段.结果表明, 在土壤状况较好的地段, 甘肃马先蒿和高原毛茛具有相对较大的比叶面积.在厚雪和浅雪地段, 紫罗兰报春的株高无显著差异, 而浅雪地段植株的比叶面积高于厚雪地段.可能由于紫罗兰报春是多年生植物且具有相对粗短的根状茎和肉质长根, 厚雪地段植株的地下部分储存相对较多的营养物质以供生殖生长利用, 而浅雪地段植株主要依赖光合作用生产碳水化合物. ...
Specific leaf area and dry matter content estimate thick- ness in laminar leaves.
2005
Uptake and recovery of soil nitrogen by bryophytes and vascular plants in an alpine meadow.
1
2014
... 异速生长揭示了某种生物或某一类生物的部分与整体或部分与部分间的差异和对比(Niklas & Enquist, 2001 ; Niklas, 2005 ; 李春萍等, 2007 ).植物的异速生长关系是其各器官相对生长速率可塑性的具体体现, 与植物的生物量分配、资源利用及对异质环境的形态适应有着密切关系(Bertalanffy, 1952 ; 陆霞梅等, 2007 ).植物生物量的分配是遗传和自然选择共同作用的结果(Weiner, 2004 ), 既具有物种特异性, 也受环境的影响.在厚雪和浅雪地段, 紫罗兰报春的地上/地下生物量表现出相同的异速生长关系, 且地下生物量的积累速率大于地上生物量的积累速率, 更多生物量分配到地下形成根状茎和肉质根, 以更好地完成生殖生长.甘肃马先蒿和高原毛茛在某些地段地上-地下生物量回归直线并不显著, 可能是样本量不足或者采样过程中样本的损耗造成.本研究结果与陶冶和张元明(2014)对6种类短命植物的地上-地下生物量的相关生长关系的研究结果较为一致, 均不支持Enquist和Niklas (2002)的等速生长假说.总体而言, 三种植物整体样本的地上-地下生物量分配均为异速生长关系, 在厚雪和浅雪地段具有共同的斜率且显著小于1, 地上生物量的积累快于地下, 而在中雪地段则相反, 生长季土壤温度在厚雪和浅雪地段相差不大且均高于中雪.另外, 整体样本中植物地上生物量可以较好地解释地下生物量的变化(表6 ).通过植物的地上部分生物量估算地下部分生物量, 可以探究整个区域内生物量对环境梯度变化的响应. ...
Common slope tests for bivariate errors-in-variables models.
2
2002
... 根据相关生长模型y = βxα 对不同雪被厚度下的三种草本植物地上/地下生物量及比叶面积-株高进行相关生长分析.分析前对生物量数据进行log10 的对数转换使之符合正态分布.方程可转化为lgy = lgβ + α lgx, x 和y 分别表示地上/地下生物量, β 为标准化常数, α 为异速生长指数.α = 1为等速生长关系, α ≠1为异速生长关系(Niklas & Enquist, 2001 ; Niklas, 2005 ; 丁建林等, 2014 ; 肖遥等, 2014 ).根据Warton和Weber (2002)的方法对同一物种在不同雪被厚度的α 进行异质性检验, 如果α 没有异质性, 则给出一个共同斜率, 若α 不同, 则异速生长关系完全不同, α 相同继续进行截距漂移检验和共轴漂移检验.若截距不同但共轴, 则属于截距漂移; 若截距相同且发生共轴漂移, 则属于共轴漂移; 若截距相同也同轴, 则表明植物的地上/地下生物量在不同雪被厚度下具有完全相同的相关生长关系(Falster et al ., 2006 ; 陶冶和张元明, 2014 ).相关生长模型的参数确定, 采用标准化主轴估计(standardized major axis estimation, SMA)方法(Wright et al ., 2001 ; Warton et al ., 2006 ), 截距漂移检验、共轴漂移检验及α 的方差分析均利用软件SMATR Version 2.0完成. ...
... 异速生长揭示了某种生物或某一类生物的部分与整体或部分与部分间的差异和对比(Niklas & Enquist, 2001 ; Niklas, 2005 ; 李春萍等, 2007 ).植物的异速生长关系是其各器官相对生长速率可塑性的具体体现, 与植物的生物量分配、资源利用及对异质环境的形态适应有着密切关系(Bertalanffy, 1952 ; 陆霞梅等, 2007 ).植物生物量的分配是遗传和自然选择共同作用的结果(Weiner, 2004 ), 既具有物种特异性, 也受环境的影响.在厚雪和浅雪地段, 紫罗兰报春的地上/地下生物量表现出相同的异速生长关系, 且地下生物量的积累速率大于地上生物量的积累速率, 更多生物量分配到地下形成根状茎和肉质根, 以更好地完成生殖生长.甘肃马先蒿和高原毛茛在某些地段地上-地下生物量回归直线并不显著, 可能是样本量不足或者采样过程中样本的损耗造成.本研究结果与陶冶和张元明(2014)对6种类短命植物的地上-地下生物量的相关生长关系的研究结果较为一致, 均不支持Enquist和Niklas (2002)的等速生长假说.总体而言, 三种植物整体样本的地上-地下生物量分配均为异速生长关系, 在厚雪和浅雪地段具有共同的斜率且显著小于1, 地上生物量的积累快于地下, 而在中雪地段则相反, 生长季土壤温度在厚雪和浅雪地段相差不大且均高于中雪.另外, 整体样本中植物地上生物量可以较好地解释地下生物量的变化(表6 ).通过植物的地上部分生物量估算地下部分生物量, 可以探究整个区域内生物量对环境梯度变化的响应. ...
Bivariate line-fitting methods for allometry.
2
2006
... 根据相关生长模型y = βxα 对不同雪被厚度下的三种草本植物地上/地下生物量及比叶面积-株高进行相关生长分析.分析前对生物量数据进行log10 的对数转换使之符合正态分布.方程可转化为lgy = lgβ + α lgx, x 和y 分别表示地上/地下生物量, β 为标准化常数, α 为异速生长指数.α = 1为等速生长关系, α ≠1为异速生长关系(Niklas & Enquist, 2001 ; Niklas, 2005 ; 丁建林等, 2014 ; 肖遥等, 2014 ).根据Warton和Weber (2002)的方法对同一物种在不同雪被厚度的α 进行异质性检验, 如果α 没有异质性, 则给出一个共同斜率, 若α 不同, 则异速生长关系完全不同, α 相同继续进行截距漂移检验和共轴漂移检验.若截距不同但共轴, 则属于截距漂移; 若截距相同且发生共轴漂移, 则属于共轴漂移; 若截距相同也同轴, 则表明植物的地上/地下生物量在不同雪被厚度下具有完全相同的相关生长关系(Falster et al ., 2006 ; 陶冶和张元明, 2014 ).相关生长模型的参数确定, 采用标准化主轴估计(standardized major axis estimation, SMA)方法(Wright et al ., 2001 ; Warton et al ., 2006 ), 截距漂移检验、共轴漂移检验及α 的方差分析均利用软件SMATR Version 2.0完成. ...
... 异速生长揭示了某种生物或某一类生物的部分与整体或部分与部分间的差异和对比(Niklas & Enquist, 2001 ; Niklas, 2005 ; 李春萍等, 2007 ).植物的异速生长关系是其各器官相对生长速率可塑性的具体体现, 与植物的生物量分配、资源利用及对异质环境的形态适应有着密切关系(Bertalanffy, 1952 ; 陆霞梅等, 2007 ).植物生物量的分配是遗传和自然选择共同作用的结果(Weiner, 2004 ), 既具有物种特异性, 也受环境的影响.在厚雪和浅雪地段, 紫罗兰报春的地上/地下生物量表现出相同的异速生长关系, 且地下生物量的积累速率大于地上生物量的积累速率, 更多生物量分配到地下形成根状茎和肉质根, 以更好地完成生殖生长.甘肃马先蒿和高原毛茛在某些地段地上-地下生物量回归直线并不显著, 可能是样本量不足或者采样过程中样本的损耗造成.本研究结果与陶冶和张元明(2014)对6种类短命植物的地上-地下生物量的相关生长关系的研究结果较为一致, 均不支持Enquist和Niklas (2002)的等速生长假说.总体而言, 三种植物整体样本的地上-地下生物量分配均为异速生长关系, 在厚雪和浅雪地段具有共同的斜率且显著小于1, 地上生物量的积累快于地下, 而在中雪地段则相反, 生长季土壤温度在厚雪和浅雪地段相差不大且均高于中雪.另外, 整体样本中植物地上生物量可以较好地解释地下生物量的变化(表6 ).通过植物的地上部分生物量估算地下部分生物量, 可以探究整个区域内生物量对环境梯度变化的响应. ...
Allocation, plasticity and allometry in plants.
1
2004
... 雪被广泛分布在寒冷生命带, 其积累与融化的交替过程直接影响着土壤水分、土壤温度、光照条件以及养分状况, 使得植物在生长节律、繁殖对策和生理生态方面形成一系列独特的适应机制(Jones et al ., 2001 ; 吴彦, 2005 ; 吴彦和Onipchenko, 2007 ).目前, 雪被生态学的研究主要集中在雪被对植物的地理分布、物质循环和土壤微生物动态的影响等方面(陈文年等, 2009 ; 胡霞等, 2012 ; 杨玉莲等, 2012 ; 何伟等, 2013 ).大量研究证据显示1980年以来北半球中高纬度的雪被面积减少了10% (Groisman et al ., 1994 ; Serreze & Hurst, 2000 ; Dye, 2002 ; IPCC, 2001 ).此外, 全球气候变化的趋势预示: 随着较高温度的出现, 高山地区降雪在近几十年将会下降(Serreze & Hurst, 2000 ; Beniston, 2005 ), 导致土壤冻结的增加, 而雪被厚度、融雪时间与生长季长度也将随之发生变化.因此, 雪被提供了季节性的复杂环境梯度(Körner, 2003 ; Pauli et al ., 2015 ), 深刻影响着高山植物的生长发育, 进而影响着高山生态系统的结构与功能(Wipf, 2010 ).然而, 雪被厚度及积雪消融时间长期形成的局域尺度的生境差异及其对植物叶片性状、生物量分配乃至群落结构的作用, 以及叶片性状与具体功能的相关性如何呈现等问题尚不明确, 更是缺乏相关的数据和证据支持. ...
The leaf size-twig size spectrum and its relationship to other important spectra of variation among species.
1
2003
... 植物通过调节自身的资源配置以适应特定的生境, 形成对一定的环境变化及其复杂性和多样性的适应机制, 在环境变化及胁迫因子作用下具有各自的生长特性和物质分配规律(肖遥等, 2014 ).生物量分配主要取决于植物的遗传特性, 同时也受环境条件的影响(郝虎东等, 2009 ; Poorter et al ., 2012 ).不同的雪被地段, 高山草本植物各器官的生物量如何分配以适应环境的变化, 是研究高山植物生存策略的重要内容.本研究中, 不同雪被厚度覆盖地段下, 三种早春草本植物生物量分配的变化具有物种特异性.在土壤温度及养分状况更优的厚雪地段, 紫罗兰报春总生物量是浅雪地段植株的2倍多.厚雪地段的植株生长速率较高, 可将部分光合产物储存在根状茎和肉质长根中, 为下一阶段的生殖生长积蓄物质和能量, 而浅雪地段的植株生长环境较差, 需要将生物量更多地分配到叶中, 从而使碳收获最大化并达到生殖阈值.厚雪和中雪地段的甘肃马先蒿各部分器官生物量均高于浅雪地段植株, 而根和叶生物量分配比例均在浅雪地段最大, 既能够吸收较多养分, 又可以提高养分利用效率, 有利于其生殖生长.在生境状况相对较好的中雪地段, 高原毛茛各部分器官生物量反而小于浅雪地段植株, 可能是由于植物在浅雪地段具有更长的生长期, 可积累较多的生物量, 但伴随着雪被融化, 也面临着失去雪被保护、遭受霜冻和水分胁迫的风险(陈文年等, 2011b ), 并且浅雪地段土壤养分相对较低, 该地段高原毛茛分配更多的地下生物量, 因此生物量更大的浅雪地段高原毛茛进入生殖阶段晚于中雪地段植株. ...
Performance of Salix herbacea in an alpine snow-bed gradient.
1
1986
... 由于地形、风和光照等环境因素的作用, 季节性雪被覆盖地区的雪被厚度存在明显的斑块性差异, 相关的融雪时间及植物生长季长度也随之变化, 导致雪下光照、温度和水分等环境因子再分配, 影响着雪下土壤动物及微生物等生物因子(Wijk, 1986 ; Lipson et al ., 2002 ; Totland & Alatalo, 2002 ; Rixen et al ., 2004 ; 陈文年等, 2011b ; 杨玉莲等, 2012 ), 进而作用于植物的功能性状和生物量分配格局.持续雪被覆盖时期, 厚雪地段土壤日平均温度相对较高, 波动幅度也最小, 土壤微生物依然具有较强的活力, 依然进行着氮矿化过程(Schimel et al ., 2004 ; Hiltbrunner et al ., 2005 ), 为早春植物的生长储备养分, 因此厚雪地段土壤有机碳含量、全氮含量高于中雪和浅雪地段.不同雪被地段土壤体积含水量均在60%以上(表2 ), 可见水分并不构成限制因子, 热量和土壤养分是影响高山植物生长的主要因子(刘琪璟等, 2009 ).综合来看, 厚雪地段土壤状况最优, 中雪地段次之, 浅雪地段相对较差.浅雪地段融雪较早, 植物易受到早期冻融交替或早霜胁迫的影响, 甘肃马先蒿和高原毛茛采取适应性策略, 开花物候有所迟滞.比叶面积-株高之间标准化主轴回归分析(SMA)的结果也表明, 当株高一定时, 浅雪地段紫罗兰报春的比叶面积高于厚雪地段, 厚雪地段甘肃马先蒿的比叶面积则高于浅雪地段, 而与中雪地段无显著差异(图2 ).尽管在不同雪被地段高原毛茛的比叶面积-株高间的回归直线斜率具有显著差异, 但中雪地段的植株比叶面积仍然高于浅雪地段.结果表明, 在土壤状况较好的地段, 甘肃马先蒿和高原毛茛具有相对较大的比叶面积.在厚雪和浅雪地段, 紫罗兰报春的株高无显著差异, 而浅雪地段植株的比叶面积高于厚雪地段.可能由于紫罗兰报春是多年生植物且具有相对粗短的根状茎和肉质长根, 厚雪地段植株的地下部分储存相对较多的营养物质以供生殖生长利用, 而浅雪地段植株主要依赖光合作用生产碳水化合物. ...
Phenology, growth, and fecundity of eight subarctic tundra species in response to snowmelt manipulations.
1
2010
... 植物功能性状与环境的关系是近年来生态学研究的热点, 尤其是植物的叶片性状具有较强的形态结构可塑性, 能够直接反映植物对环境变化的适应策略, 与资源获取和利用有着密切联系(Westoby & Wright, 2003 ; Rozendaal et al ., 2006 ; Bernard- Verdier et al ., 2012 ; 史元春等, 2015 ).植物的株高与其竞争力和繁殖活力有关(Cornelissen et al ., 2003 ), 增加对自身株高的投资能够提高光照的获取效率, 同时也增加了构成和维持支撑结构的投资(Falster & Westoby, 2003 ).生物量分配指植物生长过程中将同化的资源分配到根、茎、叶、花等器官中, 是植物重要的资源配置方式及净碳获取的驱动因子, 也是植物生存与生殖权衡的结果(方炎明, 1996 ; 郝虎东等, 2009 ).在资源匮乏或超过其利用上限的生境, 植物为了规避风险和增强对环境的适应性, 调整其功能性状等可塑性表型及生物量的分配格局来提高生存适合度(史元春等, 2015 ).异质环境中或不同的选择压力下, 现有关于植物功能性状和生物量分配的研究主要侧重于某一方面, 两者之间是否存在共轭变化, 即按照一定的规律或者关联性来匹配, 并且具有协同的动态变化, 目前的直接证据和定量化研究相对较少.因此, 定量化研究不同物种在不同生境下的叶片性状、株高与生物量分配格局的变化及其相互之间的关联性, 有助于理解全球变化背景下环境因子对植物相关功能的影响. ...
Strategy shifts in leaf physiology, structure and nutrient content between species of high- and low-rainfall and high- and low-nutrient habitats.
1
2001
... 由于地形、风和光照等环境因素的作用, 季节性雪被覆盖地区的雪被厚度存在明显的斑块性差异, 相关的融雪时间及植物生长季长度也随之变化, 导致雪下光照、温度和水分等环境因子再分配, 影响着雪下土壤动物及微生物等生物因子(Wijk, 1986 ; Lipson et al ., 2002 ; Totland & Alatalo, 2002 ; Rixen et al ., 2004 ; 陈文年等, 2011b ; 杨玉莲等, 2012 ), 进而作用于植物的功能性状和生物量分配格局.持续雪被覆盖时期, 厚雪地段土壤日平均温度相对较高, 波动幅度也最小, 土壤微生物依然具有较强的活力, 依然进行着氮矿化过程(Schimel et al ., 2004 ; Hiltbrunner et al ., 2005 ), 为早春植物的生长储备养分, 因此厚雪地段土壤有机碳含量、全氮含量高于中雪和浅雪地段.不同雪被地段土壤体积含水量均在60%以上(表2 ), 可见水分并不构成限制因子, 热量和土壤养分是影响高山植物生长的主要因子(刘琪璟等, 2009 ).综合来看, 厚雪地段土壤状况最优, 中雪地段次之, 浅雪地段相对较差.浅雪地段融雪较早, 植物易受到早期冻融交替或早霜胁迫的影响, 甘肃马先蒿和高原毛茛采取适应性策略, 开花物候有所迟滞.比叶面积-株高之间标准化主轴回归分析(SMA)的结果也表明, 当株高一定时, 浅雪地段紫罗兰报春的比叶面积高于厚雪地段, 厚雪地段甘肃马先蒿的比叶面积则高于浅雪地段, 而与中雪地段无显著差异(图2 ).尽管在不同雪被地段高原毛茛的比叶面积-株高间的回归直线斜率具有显著差异, 但中雪地段的植株比叶面积仍然高于浅雪地段.结果表明, 在土壤状况较好的地段, 甘肃马先蒿和高原毛茛具有相对较大的比叶面积.在厚雪和浅雪地段, 紫罗兰报春的株高无显著差异, 而浅雪地段植株的比叶面积高于厚雪地段.可能由于紫罗兰报春是多年生植物且具有相对粗短的根状茎和肉质长根, 厚雪地段植株的地下部分储存相对较多的营养物质以供生殖生长利用, 而浅雪地段植株主要依赖光合作用生产碳水化合物. ...
The world- wide leaf economics spectrum.
2
2004
... 雪被广泛分布在寒冷生命带, 其积累与融化的交替过程直接影响着土壤水分、土壤温度、光照条件以及养分状况, 使得植物在生长节律、繁殖对策和生理生态方面形成一系列独特的适应机制(Jones et al ., 2001 ; 吴彦, 2005 ; 吴彦和Onipchenko, 2007 ).目前, 雪被生态学的研究主要集中在雪被对植物的地理分布、物质循环和土壤微生物动态的影响等方面(陈文年等, 2009 ; 胡霞等, 2012 ; 杨玉莲等, 2012 ; 何伟等, 2013 ).大量研究证据显示1980年以来北半球中高纬度的雪被面积减少了10% (Groisman et al ., 1994 ; Serreze & Hurst, 2000 ; Dye, 2002 ; IPCC, 2001 ).此外, 全球气候变化的趋势预示: 随着较高温度的出现, 高山地区降雪在近几十年将会下降(Serreze & Hurst, 2000 ; Beniston, 2005 ), 导致土壤冻结的增加, 而雪被厚度、融雪时间与生长季长度也将随之发生变化.因此, 雪被提供了季节性的复杂环境梯度(Körner, 2003 ; Pauli et al ., 2015 ), 深刻影响着高山植物的生长发育, 进而影响着高山生态系统的结构与功能(Wipf, 2010 ).然而, 雪被厚度及积雪消融时间长期形成的局域尺度的生境差异及其对植物叶片性状、生物量分配乃至群落结构的作用, 以及叶片性状与具体功能的相关性如何呈现等问题尚不明确, 更是缺乏相关的数据和证据支持. ...
... ).此外, 全球气候变化的趋势预示: 随着较高温度的出现, 高山地区降雪在近几十年将会下降(Serreze & Hurst, 2000 ; Beniston, 2005 ), 导致土壤冻结的增加, 而雪被厚度、融雪时间与生长季长度也将随之发生变化.因此, 雪被提供了季节性的复杂环境梯度(Körner, 2003 ; Pauli et al ., 2015 ), 深刻影响着高山植物的生长发育, 进而影响着高山生态系统的结构与功能(Wipf, 2010 ).然而, 雪被厚度及积雪消融时间长期形成的局域尺度的生境差异及其对植物叶片性状、生物量分配乃至群落结构的作用, 以及叶片性状与具体功能的相关性如何呈现等问题尚不明确, 更是缺乏相关的数据和证据支持. ...
三种高寒植物幼苗生物量分配及性状特征对光照和养分的响应
2010
植物形态生长对策研究进展
2
2007
... 植物功能性状与环境的关系是近年来生态学研究的热点, 尤其是植物的叶片性状具有较强的形态结构可塑性, 能够直接反映植物对环境变化的适应策略, 与资源获取和利用有着密切联系(Westoby & Wright, 2003 ; Rozendaal et al ., 2006 ; Bernard- Verdier et al ., 2012 ; 史元春等, 2015 ).植物的株高与其竞争力和繁殖活力有关(Cornelissen et al ., 2003 ), 增加对自身株高的投资能够提高光照的获取效率, 同时也增加了构成和维持支撑结构的投资(Falster & Westoby, 2003 ).生物量分配指植物生长过程中将同化的资源分配到根、茎、叶、花等器官中, 是植物重要的资源配置方式及净碳获取的驱动因子, 也是植物生存与生殖权衡的结果(方炎明, 1996 ; 郝虎东等, 2009 ).在资源匮乏或超过其利用上限的生境, 植物为了规避风险和增强对环境的适应性, 调整其功能性状等可塑性表型及生物量的分配格局来提高生存适合度(史元春等, 2015 ).异质环境中或不同的选择压力下, 现有关于植物功能性状和生物量分配的研究主要侧重于某一方面, 两者之间是否存在共轭变化, 即按照一定的规律或者关联性来匹配, 并且具有协同的动态变化, 目前的直接证据和定量化研究相对较少.因此, 定量化研究不同物种在不同生境下的叶片性状、株高与生物量分配格局的变化及其相互之间的关联性, 有助于理解全球变化背景下环境因子对植物相关功能的影响. ...
... ).在资源匮乏或超过其利用上限的生境, 植物为了规避风险和增强对环境的适应性, 调整其功能性状等可塑性表型及生物量的分配格局来提高生存适合度(史元春等, 2015 ).异质环境中或不同的选择压力下, 现有关于植物功能性状和生物量分配的研究主要侧重于某一方面, 两者之间是否存在共轭变化, 即按照一定的规律或者关联性来匹配, 并且具有协同的动态变化, 目前的直接证据和定量化研究相对较少.因此, 定量化研究不同物种在不同生境下的叶片性状、株高与生物量分配格局的变化及其相互之间的关联性, 有助于理解全球变化背景下环境因子对植物相关功能的影响. ...
季节性雪被覆盖对植物群落的影响
1
2005
... 紫罗兰报春的株高在厚雪和浅雪地段无显著差异, 其他两种植物的株高均随着雪被厚度的减小而显著升高.两种植物株高的增加很可能与获取更多光照有关, 另外环境因子对草地群落的物种丰富度有着重要影响(杨元合等, 2004 ; Suding et al ., 2005 ), 随着雪被厚度减小, 生境内物种密度增加(未发表群落调查数据), 竞争压力也会增大, 株高不占优势的甘肃马先蒿和高原毛茛会增加对株高的投资以最大化碳收获. ...
雪被对川西高山植被坡向性分异的影响
1
2007
... 当受到胁迫或资源限制时, 植物会适当地调整其功能性状和生物量分配, 从而提高竞争力及其生存适合度(Sultan, 1992 ; 武高林和杜国祯, 2007 ).例如养分不足时, 植物会将生物量更多地分配到地下, 而减少地上部分的生物量分配, 植物的比叶面积和株高等功能性状也会相应减小(武高林等, 2010 ), 因此, 植物功能性状和生物量分配之间存在着某种共轭的变化.这种相互关系对于量化和概括植物面对生境变化的权衡策略及其规律有重要意义(Wright et al ., 2004 ; 陈莹婷和许振柱, 2014 ).本研究中, 紫罗兰报春(主要是株高和比叶面积与生物量指标之间)的相关性较强, 而这种相关性在甘肃马先蒿和高原毛茛相对较弱.由于生活型和个体发育的差异, 不同的物种在面临相同环境变化时, 权衡策略具有物种特异性, 可表征生物量变化的功能性状亦有所不同.因此, 如何选取具有普适性的性状, 表征不同尺度生物量的变化需要以后深入研究, 以丰富中国的植物性状数据库, 并更好地理解全球变化背景下植物功能性状的响应和适应对策. ...
古尔班通古特沙漠4种荒漠草本植物不同生长期的生物量分配与叶片化学计量特征
2014
青藏高原高寒草地植物物种丰富度及其与环境因子和生物量的关系
1
2004
... 根据相关生长模型y = βxα 对不同雪被厚度下的三种草本植物地上/地下生物量及比叶面积-株高进行相关生长分析.分析前对生物量数据进行log10 的对数转换使之符合正态分布.方程可转化为lgy = lgβ + α lgx, x 和y 分别表示地上/地下生物量, β 为标准化常数, α 为异速生长指数.α = 1为等速生长关系, α ≠1为异速生长关系(Niklas & Enquist, 2001 ; Niklas, 2005 ; 丁建林等, 2014 ; 肖遥等, 2014 ).根据Warton和Weber (2002)的方法对同一物种在不同雪被厚度的α 进行异质性检验, 如果α 没有异质性, 则给出一个共同斜率, 若α 不同, 则异速生长关系完全不同, α 相同继续进行截距漂移检验和共轴漂移检验.若截距不同但共轴, 则属于截距漂移; 若截距相同且发生共轴漂移, 则属于共轴漂移; 若截距相同也同轴, 则表明植物的地上/地下生物量在不同雪被厚度下具有完全相同的相关生长关系(Falster et al ., 2006 ; 陶冶和张元明, 2014 ).相关生长模型的参数确定, 采用标准化主轴估计(standardized major axis estimation, SMA)方法(Wright et al ., 2001 ; Warton et al ., 2006 ), 截距漂移检验、共轴漂移检验及α 的方差分析均利用软件SMATR Version 2.0完成. ...
雪被去除对川西高山冷杉林冬季土壤微生物生物量碳氮和可培养微生物数量的影响
1
2012
... 由于地形、风和光照等环境因素的作用, 季节性雪被覆盖地区的雪被厚度存在明显的斑块性差异, 相关的融雪时间及植物生长季长度也随之变化, 导致雪下光照、温度和水分等环境因子再分配, 影响着雪下土壤动物及微生物等生物因子(Wijk, 1986 ; Lipson et al ., 2002 ; Totland & Alatalo, 2002 ; Rixen et al ., 2004 ; 陈文年等, 2011b ; 杨玉莲等, 2012 ), 进而作用于植物的功能性状和生物量分配格局.持续雪被覆盖时期, 厚雪地段土壤日平均温度相对较高, 波动幅度也最小, 土壤微生物依然具有较强的活力, 依然进行着氮矿化过程(Schimel et al ., 2004 ; Hiltbrunner et al ., 2005 ), 为早春植物的生长储备养分, 因此厚雪地段土壤有机碳含量、全氮含量高于中雪和浅雪地段.不同雪被地段土壤体积含水量均在60%以上(表2 ), 可见水分并不构成限制因子, 热量和土壤养分是影响高山植物生长的主要因子(刘琪璟等, 2009 ).综合来看, 厚雪地段土壤状况最优, 中雪地段次之, 浅雪地段相对较差.浅雪地段融雪较早, 植物易受到早期冻融交替或早霜胁迫的影响, 甘肃马先蒿和高原毛茛采取适应性策略, 开花物候有所迟滞.比叶面积-株高之间标准化主轴回归分析(SMA)的结果也表明, 当株高一定时, 浅雪地段紫罗兰报春的比叶面积高于厚雪地段, 厚雪地段甘肃马先蒿的比叶面积则高于浅雪地段, 而与中雪地段无显著差异(图2 ).尽管在不同雪被地段高原毛茛的比叶面积-株高间的回归直线斜率具有显著差异, 但中雪地段的植株比叶面积仍然高于浅雪地段.结果表明, 在土壤状况较好的地段, 甘肃马先蒿和高原毛茛具有相对较大的比叶面积.在厚雪和浅雪地段, 紫罗兰报春的株高无显著差异, 而浅雪地段植株的比叶面积高于厚雪地段.可能由于紫罗兰报春是多年生植物且具有相对粗短的根状茎和肉质长根, 厚雪地段植株的地下部分储存相对较多的营养物质以供生殖生长利用, 而浅雪地段植株主要依赖光合作用生产碳水化合物. ...
 , 徐波
, 徐波

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}