Leaf nitrogen and phosphorus concentration and the empirical regulations in dominant woody plants of shrublands across southern China
LI Jia-Xiang1, 2, XU Wen-Ting1, XIONG Gao-Ming1, WANG Yang1, ZHAO Chang-Ming1, LU Zhi-Jun3, LI Yue-Lin4, XIE Zong-Qiang1, *,
1State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China2College of Forest, Central South University of Forestry & Technology, Changsha 410004, China;3Key Laboratory of Aquatic Botany and Watershed Ecology, Wuhan Botanical Garden, Chinese Academy of Sciences, Wuhan 430074, Chinaand 4South China Botanical Garden, Chinese Academy of Sciences, Guangzhou 510650, China
通讯作者: * 通信作者Author for correspondence (E-mail: xie@ibcas.ac.cn)
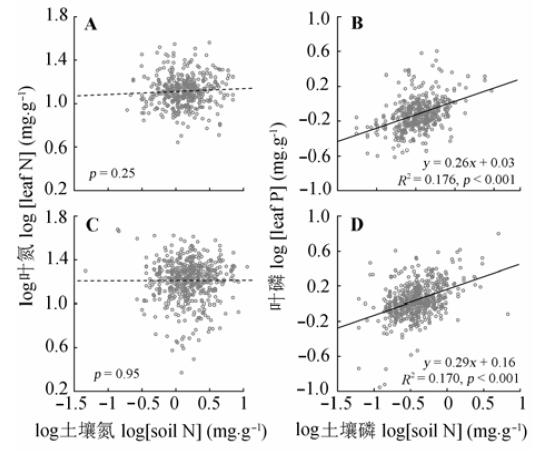
Aims Understanding the changes in N and P concentration in plant organs along the environmental gradients can provide meaningful information to reveal the underline mechanisms for the geochemical cycles and adaptation strategies of plants to the changing environment. In this paper, we aimed to answer: (1) How did the N and P concentration in leaves of evergreen and deciduous woody plants change along the environmental gradients? (2) What were the main factors regulating the N and P concentration in leaves of woody plants in the shrublands across southern China? Methods Using a stratified random sampling method, we sampled 193 dominant woody plants in 462 sites of 12 provinces in southern China. Leaf samples of dominant woody plants, including 91 evergreen and 102 deciduous shrubs, and soil samples at each site were collected. N and P concentration of the leaves and soils were measured after lapping and sieving. Kruskal-Wallis and Nemenyi tests were applied to quantify the difference among the organs and life-forms. For each life-form, the binary linear regression was used to estimate the relationships between leaf log [N] and log [P] concentration and mean annual air temperature (MAT), mean annual precipitation (MAP) and log soil total [N], [P]. The effects of climate, soil and plant life-form on leaf chemical traits were modeled through the general linear models (GLMs) and F-tests. Important findings 1) The geometric means of leaf N and P concentrations of the dominant woody plants were 16.57 mg·g-1 and 1.02 mg·g-1, respectively. The N and P concentration in leaves (17.91 mg·g-1, 1.14 mg·g-1) of deciduous woody plants was higher than those of evergreen woody plants (15.19 mg·g-1, 0.89 mg·g-1). The dependent of leaf P concentration on environmental (climate and soil) appeared more variable than N concentration. 2) Leaf N and P in evergreen woody plants decreased with MAT and but increased with MAP, whereas those in deciduous woody plants showed opposite trends. With increase in MAP, leaf P concentration decreased for both evergreen and deciduous woody plants. 3) Soil N concentration had no significant effect on both evergreen and deciduous woody plants. However, leaf P concentration of the tow increased significantly with soil P concentration. (4) GLMs showed that plant growth form explained 7.6% and 14.4% of variation in leaf N and P, respectively. MAP and soil P concentration contributed 0.8% and 16.4% of the variation in leaf P, respectively. These results suggested that leaf N was mainly influenced by plant growth form, while leaf P concentration was driven by soil, plant life-form, and climate at our study sites.
Keywords:tropical and subtropical region
;
plant functional traits
;
life form
;
ecological stoichiometry
LIJia-Xiang, XUWen-Ting, XIONGGao-Ming, WANGYang, ZHAOChang-Ming, LUZhi-Jun, LIYue-Lin, XIEZong-Qiang. Leaf nitrogen and phosphorus concentration and the empirical regulations in dominant woody plants of shrublands across southern China. Chinese Journal of Plant Ecology, 2017, 41(1): 31-42 https://doi.org/10.17521/cjpe.2016.0251
为了衡量木本植物叶N、P含量与气候和土壤养分之间的关系, 将每个样点上分种叶N、P含量原始值进行对数化转换, 分别与MAT、MAP、土壤N含量和土壤P含量进行一般线性回归(任书杰等, 2007; Hong et al., 2014)。同时, 采用广义线性模型(GLM)来衡量生活型、MAT、MAP、土壤N含量和P含量分别对木本植物叶N含量和P含量变异的解释量(Chen et al., 2013; Zhao et al., 2014)。所有的统计分析和作图分别在R 3.2.1和Excel 2013中进行。
Table 1 Leaf N and P concentration of dominant woody plants in shrublands across southern China
生活型 Life-form
N含量 N content (mg·g-1)
P含量 P content (mg·g-1)
n
GM
Range
CV
n
GM
Range
CV
常绿木本植物 Evergreen woody plants
91
15.19a
7.68-33.00
0.31
91
0.89a
0.37-3.81
0.52
落叶木本植物 Deciduous woody plants
102
17.91b
5.06-43.78
0.35
102
1.14b
0.18-4.04
0.47
所有木本植物 All woody plants
193
16.57
5.06-43.78
0.35
193
1.02
0.18-4.04
0.50
GM, n, Range and CV were geometric mean values, statistical samples, variation range and coefficient of variation, respectively. “a” and “b” indicated significant difference (p < 0.05) between evergreen and deciduous woody plants.GM、n、Range和CV分别为几何平均数、样本数、范围和变异系数; a和b表示常绿和落叶木本器官元素含量差异显著(p < 0.05)。
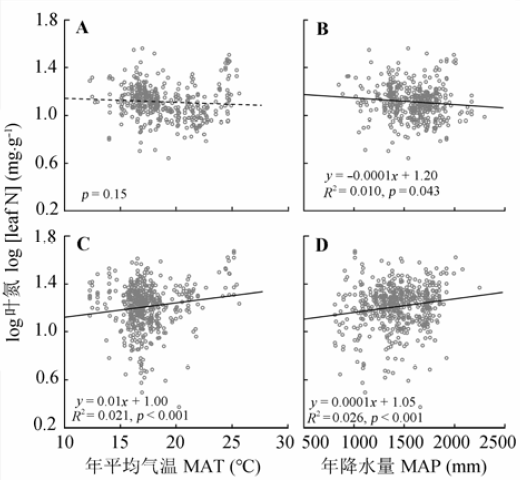
Fig. 1 Effects of mean annual temperature (MAT) and mean annual precipitation (MAP) on leaf N concentration of evergreen (A, B) and deciduous woody plants (C, D).
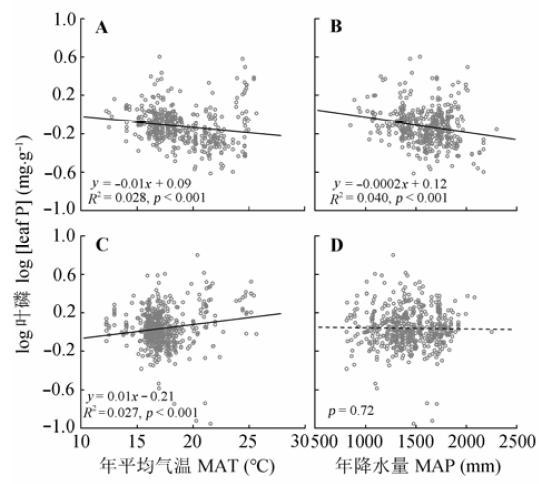
Fig. 2 Effects of mean annual temperature (MAT) and mean annual precipitation (MAP) on leaf P concentrations of evergreen (A, B) and deciduous woody plants (C, D).
Table 2 Summary of general linear models for leaf N and P content of woody plants
因子 Factor
叶片氮含量 Leaf N content
叶片磷含量 Leaf P content
DF
MS
F
SS%
DF
MS
F
SS%
生活型 Life form
1
2.08
78.51
7.6
1
5.79
199.75
14.4
年平均气温 MAT (℃)
1
0.07
2.68
1
0.00
0.05
年降水量 MAP (mm)
1
0.09
3.24
1
0.31
10.52
0.8
土壤氮含量 Soil N content (mg·g-1)
1
0.03
1.22
1
0.37
12.91
0.9
土壤磷含量 Soil P content (mg·g-1)
1
0.63
23.58
2.3
1
6.57
226.60
16.4
DF, MS, F and SS% represent freedom degree, mean sum of squares, F value and interpretation quantity of each factor. F values in bold indicate p < 0.05. MAT, mean annual temperature; MAP, mean annual precipitation.DF、MS、F和SS%分别表示自由度、平均残差平方和、F值和各因子的解释量; 加粗的F值表示p < 0.05。
The Editorial Committee of Vegetation Map of China, Chinese Academy ofSciences (2007). Vegetation Map of the People’s Republic of China 1:1000000. Geological Publishing House, Beijing. (in Chinese)[中国科学院中国植被图编辑委员会 (2007). 中华人民共和国植被图1:1000000. 地质出版社, 北京.]
Technical specification of field investigation and laboratory analysis for carbon sequestration in shrubland ecosystems
. In: Compilation Group of Observation and Investigation for Carbon Sequestration in Terrestrial Ecosystems ed. Observation and Investigation for Carbon Sequestration in Terrestrial Ecosystems. Science Press, Beijing. 145-191. (in Chinese)[谢宗强, 唐志尧 (2015).
The mineral nutrition of wild plants revisited: A re-evaluation of processes and patterns
1
2000
... 氮(N)和磷(P)是植物生命活动所需的大量元素, 其含量大小系植物的主要功能性状, 在生态系统功能和动态中起关键作用(Koerselman & Meuleman, 1996; Reich, 2005).为了最大化地满足生长, 植物通过平衡限制性资源的分配, 在气候、土壤养分等改变时调整分配策略以适应环境限制(Reich & Oleksyn, 2004), 因此, 气候、土壤成为影响植物营养器官N、P含量的主要环境因子.研究发现叶片N、P含量与气候和土壤的关系因各区域限制性因子的差异而不同(Reich & Oleksyn, 2004; Han et al., 2005; Kerkhoff et al., 2005; He et al., 2008; 丁凡等, 2011; 王晶苑等, 2011; Sardans & Peñuelas, 2013; Hong et al., 2014; Yang et al., 2014); 同时, 相同功能型植物往往趋于分享一套关键的植物功能性状来实现对环境的趋同适应和对生态系统的相似作用(Díaz & Cabido, 2001).因此, 探索不同功能型植物器官中N、P含量沿环境梯度的分异规律, 有助于揭示陆地生态系统物质循环和植物养分适应策略的过程和机制(Aerts & Chapin, 2000; 曾德慧和陈广生, 2005; 刘超等, 2012). ...
Stoichiometry and nutrition of plant growth in natural communities
2
2008
... 植物生活型是叶N、P含量的一个主要决定因子, 以往大量研究已注意到不同生活型植物器官N、P化学计量存在显著差异(Reich & Oleksyn, 2004; Han et al., 2008; Chen et al., 2013; Sardans & Peñuelas, 2013; Pan et al., 2015), 发现生命周期长且生长慢的植物器官N、P含量较生命周期短且生长迅速的植物高, 并认为植物器官N、P含量是植物代谢能力以及养分利用效率的综合反映(Han et al., 2005; Ågren, 2008).本研究中, 常绿和落叶木本植物叶N、P含量存在显著差异, 落叶木本植物叶的N、P含量(17.91 mg·g-1, 1.14 mg·g-1)显著高于常绿木本植物叶(15.19 mg·g-1, 0.89 mg·g-1)(表1), 生活型可分别解释叶片N、P含量7.6%和14.4%的变异(表2).该结果主要是因为常绿木本植物叶寿命较长, 与落叶木本植物相比, 通常具有更长的养分驻留时间、保守的养分利用策略和更高的养分利用效率; 为了适应低养环境, 常绿木本植物需要投资更多能量用于叶片保护和结构支撑, 从而导致其比落叶木本植物更低的光合速率和养分含量(Edwards et al., 2014), 但常绿性意味着具有较长的光合时间, 保证其具有较高的养分利用效率, 从而减少对土壤养分的依赖.然而, 落叶木本植物通过叶片凋落减少其在不利季节的蒸散和呼吸; 虽然落叶习性意味着较短的叶生命周期, 却有较高的凋落物分解和养分重吸收率, 有着较常绿植物高的光合速率和养分含量.结果证明不同生活型植物的物种特性对养分利用策略的影响较大, 进而决定植物器官的营养元素含量(He et al., 2008; Wu et al., 2012), 支持生长速率假说(Ågren, 2008). ...
... ), 支持生长速率假说(Ågren, 2008). ...
Plant stoichiometry at different scales: Element concentration patterns reflect environment more than genotype
1
2012
... 植物的营养元素含量是植物在一定生境下吸收营养元素的能力, 也是植物与环境之间相互作用的结果.在全球或国家尺度上, 已有研究表明植物叶N、P含量随MAT增高和MAP增加逐渐减少(Güsewell, 2004; Hedin, 2004; McGroddy et al., 2004; He et al., 2006; Ågren & Weih, 2012; Chen et al., 2013).然而, 中国南方灌丛中常绿和落叶木本植物叶N、P含量与气候因子的关系不同(图1, 图2), 其中常绿木本植物叶N、P含量与Han等(2005)与Reich和Oleksyn (2004)的研究结果存在一样的温度趋势, 结果支持温度-植物生理假说: 叶片N、P含量的增加可补偿低温下代谢速率的下降, 因此随温度的增加, 叶片N、P含量呈降低趋势(Reich & Oleksyn, 2004).同时, 较高的叶片N、P含量也是植物适应干旱条件的反映(Yang et al., 2014), 也可能是较多的降水可增强土壤N、P的淋溶作用, 土壤养分的可获得性降低, 从而导致常绿木本植物叶N、P含量呈降低的趋势.然而, 落叶木本植物叶N、P含量随MAT和MAP的升高而呈增加趋势, 可能与较高的温度和较多的降水会加快养分周转, 增加土壤养分的可获得性有关; 再者, 在温暖湿润的环境中, 落叶木本植物生长较快, 较高的叶N、P含量有助于满足植物快速生长的养分需求, 因此落叶木本植物叶N、P含量随年平均气温和年降水量的增加呈增加趋势. ...
Leaf nitrogen and phosphorus concentrations of woody plants differ in responses to climate, soil and plant growth form
5
2013
... 灌丛作为一种重要的自然植被类型, 不同于森林的主要特点是以灌木生活型植物为优势成分, 具有类型多样、分布广泛、生产力高、萌生能力强等特点, 其分布面积约占中国国土面积的20%, 约占中国亚热带国土面积的25% (中国科学院中国植被图编辑委员会, 2007), 是陆地生态系统养分循环的重要场所(胡会峰等, 2006; Piao et al., 2009).截至目前, 关于植物养分循环的研究多聚焦于森林或草地(张丽霞, 2003; He et al., 2006, 2008; 任书杰等, 2007; 甘露等, 2008; 丁凡等, 2011; 林志斌等, 2011; Chen et al., 2013; Cornelissen et al., 2014; Hong et al., 2014; Nottingham et al., 2015; Pan et al., 2015; Petter et al., 2016), 发现不同功能型植物叶N、P含量差异显著, 且落叶木本植物叶的N、P含量显著高于常绿木本植物.尽管植物在灌丛中的生长环境和地位与森林或草地截然不同, 但关于灌丛中植物器官N、P格局却较少受到重视, 且相关研究多在北方(韩文轩等, 2009; Yang et al., 2014; 李单凤等, 2015); 再者, 众多研究在区域和全球尺度阐释了叶N、P的地理变异机制(Elser et al., 2007), 但大部分研究并没有检验不同功能型植物叶N、P对环境因子的响应规律(Chen et al., 2013).因此, 在较大范围内研究灌丛生态系统不同生活型优势植物N、P含量及其与环境因子的关系, 有助于深入了解植物N、P养分循环和植物养分适应策略的过程和机制. ...
... 为了衡量木本植物叶N、P含量与气候和土壤养分之间的关系, 将每个样点上分种叶N、P含量原始值进行对数化转换, 分别与MAT、MAP、土壤N含量和土壤P含量进行一般线性回归(任书杰等, 2007; Hong et al., 2014).同时, 采用广义线性模型(GLM)来衡量生活型、MAT、MAP、土壤N含量和P含量分别对木本植物叶N含量和P含量变异的解释量(Chen et al., 2013; Zhao et al., 2014).所有的统计分析和作图分别在R 3.2.1和Excel 2013中进行. ...
... 植物生活型是叶N、P含量的一个主要决定因子, 以往大量研究已注意到不同生活型植物器官N、P化学计量存在显著差异(Reich & Oleksyn, 2004; Han et al., 2008; Chen et al., 2013; Sardans & Peñuelas, 2013; Pan et al., 2015), 发现生命周期长且生长慢的植物器官N、P含量较生命周期短且生长迅速的植物高, 并认为植物器官N、P含量是植物代谢能力以及养分利用效率的综合反映(Han et al., 2005; Ågren, 2008).本研究中, 常绿和落叶木本植物叶N、P含量存在显著差异, 落叶木本植物叶的N、P含量(17.91 mg·g-1, 1.14 mg·g-1)显著高于常绿木本植物叶(15.19 mg·g-1, 0.89 mg·g-1)(表1), 生活型可分别解释叶片N、P含量7.6%和14.4%的变异(表2).该结果主要是因为常绿木本植物叶寿命较长, 与落叶木本植物相比, 通常具有更长的养分驻留时间、保守的养分利用策略和更高的养分利用效率; 为了适应低养环境, 常绿木本植物需要投资更多能量用于叶片保护和结构支撑, 从而导致其比落叶木本植物更低的光合速率和养分含量(Edwards et al., 2014), 但常绿性意味着具有较长的光合时间, 保证其具有较高的养分利用效率, 从而减少对土壤养分的依赖.然而, 落叶木本植物通过叶片凋落减少其在不利季节的蒸散和呼吸; 虽然落叶习性意味着较短的叶生命周期, 却有较高的凋落物分解和养分重吸收率, 有着较常绿植物高的光合速率和养分含量.结果证明不同生活型植物的物种特性对养分利用策略的影响较大, 进而决定植物器官的营养元素含量(He et al., 2008; Wu et al., 2012), 支持生长速率假说(Ågren, 2008). ...
... 植物生活型是叶N、P含量的一个主要决定因子, 以往大量研究已注意到不同生活型植物器官N、P化学计量存在显著差异(Reich & Oleksyn, 2004; Han et al., 2008; Chen et al., 2013; Sardans & Peñuelas, 2013; Pan et al., 2015), 发现生命周期长且生长慢的植物器官N、P含量较生命周期短且生长迅速的植物高, 并认为植物器官N、P含量是植物代谢能力以及养分利用效率的综合反映(Han et al., 2005; Ågren, 2008).本研究中, 常绿和落叶木本植物叶N、P含量存在显著差异, 落叶木本植物叶的N、P含量(17.91 mg·g-1, 1.14 mg·g-1)显著高于常绿木本植物叶(15.19 mg·g-1, 0.89 mg·g-1)(表1), 生活型可分别解释叶片N、P含量7.6%和14.4%的变异(表2).该结果主要是因为常绿木本植物叶寿命较长, 与落叶木本植物相比, 通常具有更长的养分驻留时间、保守的养分利用策略和更高的养分利用效率; 为了适应低养环境, 常绿木本植物需要投资更多能量用于叶片保护和结构支撑, 从而导致其比落叶木本植物更低的光合速率和养分含量(Edwards et al., 2014), 但常绿性意味着具有较长的光合时间, 保证其具有较高的养分利用效率, 从而减少对土壤养分的依赖.然而, 落叶木本植物通过叶片凋落减少其在不利季节的蒸散和呼吸; 虽然落叶习性意味着较短的叶生命周期, 却有较高的凋落物分解和养分重吸收率, 有着较常绿植物高的光合速率和养分含量.结果证明不同生活型植物的物种特性对养分利用策略的影响较大, 进而决定植物器官的营养元素含量(He et al., 2008; Wu et al., 2012), 支持生长速率假说(Ågren, 2008). ...
Stoichiometry of root and leaf nitrogen and phosphorus in a dry alpine steppe on the northern Tibetan Plateau
3
2014
... 氮(N)和磷(P)是植物生命活动所需的大量元素, 其含量大小系植物的主要功能性状, 在生态系统功能和动态中起关键作用(Koerselman & Meuleman, 1996; Reich, 2005).为了最大化地满足生长, 植物通过平衡限制性资源的分配, 在气候、土壤养分等改变时调整分配策略以适应环境限制(Reich & Oleksyn, 2004), 因此, 气候、土壤成为影响植物营养器官N、P含量的主要环境因子.研究发现叶片N、P含量与气候和土壤的关系因各区域限制性因子的差异而不同(Reich & Oleksyn, 2004; Han et al., 2005; Kerkhoff et al., 2005; He et al., 2008; 丁凡等, 2011; 王晶苑等, 2011; Sardans & Peñuelas, 2013; Hong et al., 2014; Yang et al., 2014); 同时, 相同功能型植物往往趋于分享一套关键的植物功能性状来实现对环境的趋同适应和对生态系统的相似作用(Díaz & Cabido, 2001).因此, 探索不同功能型植物器官中N、P含量沿环境梯度的分异规律, 有助于揭示陆地生态系统物质循环和植物养分适应策略的过程和机制(Aerts & Chapin, 2000; 曾德慧和陈广生, 2005; 刘超等, 2012). ...
... 灌丛作为一种重要的自然植被类型, 不同于森林的主要特点是以灌木生活型植物为优势成分, 具有类型多样、分布广泛、生产力高、萌生能力强等特点, 其分布面积约占中国国土面积的20%, 约占中国亚热带国土面积的25% (中国科学院中国植被图编辑委员会, 2007), 是陆地生态系统养分循环的重要场所(胡会峰等, 2006; Piao et al., 2009).截至目前, 关于植物养分循环的研究多聚焦于森林或草地(张丽霞, 2003; He et al., 2006, 2008; 任书杰等, 2007; 甘露等, 2008; 丁凡等, 2011; 林志斌等, 2011; Chen et al., 2013; Cornelissen et al., 2014; Hong et al., 2014; Nottingham et al., 2015; Pan et al., 2015; Petter et al., 2016), 发现不同功能型植物叶N、P含量差异显著, 且落叶木本植物叶的N、P含量显著高于常绿木本植物.尽管植物在灌丛中的生长环境和地位与森林或草地截然不同, 但关于灌丛中植物器官N、P格局却较少受到重视, 且相关研究多在北方(韩文轩等, 2009; Yang et al., 2014; 李单凤等, 2015); 再者, 众多研究在区域和全球尺度阐释了叶N、P的地理变异机制(Elser et al., 2007), 但大部分研究并没有检验不同功能型植物叶N、P对环境因子的响应规律(Chen et al., 2013).因此, 在较大范围内研究灌丛生态系统不同生活型优势植物N、P含量及其与环境因子的关系, 有助于深入了解植物N、P养分循环和植物养分适应策略的过程和机制. ...
... 为了衡量木本植物叶N、P含量与气候和土壤养分之间的关系, 将每个样点上分种叶N、P含量原始值进行对数化转换, 分别与MAT、MAP、土壤N含量和土壤P含量进行一般线性回归(任书杰等, 2007; Hong et al., 2014).同时, 采用广义线性模型(GLM)来衡量生活型、MAT、MAP、土壤N含量和P含量分别对木本植物叶N含量和P含量变异的解释量(Chen et al., 2013; Zhao et al., 2014).所有的统计分析和作图分别在R 3.2.1和Excel 2013中进行. ...
中国主要灌丛植被碳储量
1
2006
... 灌丛作为一种重要的自然植被类型, 不同于森林的主要特点是以灌木生活型植物为优势成分, 具有类型多样、分布广泛、生产力高、萌生能力强等特点, 其分布面积约占中国国土面积的20%, 约占中国亚热带国土面积的25% (中国科学院中国植被图编辑委员会, 2007), 是陆地生态系统养分循环的重要场所(胡会峰等, 2006; Piao et al., 2009).截至目前, 关于植物养分循环的研究多聚焦于森林或草地(张丽霞, 2003; He et al., 2006, 2008; 任书杰等, 2007; 甘露等, 2008; 丁凡等, 2011; 林志斌等, 2011; Chen et al., 2013; Cornelissen et al., 2014; Hong et al., 2014; Nottingham et al., 2015; Pan et al., 2015; Petter et al., 2016), 发现不同功能型植物叶N、P含量差异显著, 且落叶木本植物叶的N、P含量显著高于常绿木本植物.尽管植物在灌丛中的生长环境和地位与森林或草地截然不同, 但关于灌丛中植物器官N、P格局却较少受到重视, 且相关研究多在北方(韩文轩等, 2009; Yang et al., 2014; 李单凤等, 2015); 再者, 众多研究在区域和全球尺度阐释了叶N、P的地理变异机制(Elser et al., 2007), 但大部分研究并没有检验不同功能型植物叶N、P对环境因子的响应规律(Chen et al., 2013).因此, 在较大范围内研究灌丛生态系统不同生活型优势植物N、P含量及其与环境因子的关系, 有助于深入了解植物N、P养分循环和植物养分适应策略的过程和机制. ...
Plant allometry, stoichiometry and the temperature-dependence of primary productivity
1
2005
... 氮(N)和磷(P)是植物生命活动所需的大量元素, 其含量大小系植物的主要功能性状, 在生态系统功能和动态中起关键作用(Koerselman & Meuleman, 1996; Reich, 2005).为了最大化地满足生长, 植物通过平衡限制性资源的分配, 在气候、土壤养分等改变时调整分配策略以适应环境限制(Reich & Oleksyn, 2004), 因此, 气候、土壤成为影响植物营养器官N、P含量的主要环境因子.研究发现叶片N、P含量与气候和土壤的关系因各区域限制性因子的差异而不同(Reich & Oleksyn, 2004; Han et al., 2005; Kerkhoff et al., 2005; He et al., 2008; 丁凡等, 2011; 王晶苑等, 2011; Sardans & Peñuelas, 2013; Hong et al., 2014; Yang et al., 2014); 同时, 相同功能型植物往往趋于分享一套关键的植物功能性状来实现对环境的趋同适应和对生态系统的相似作用(Díaz & Cabido, 2001).因此, 探索不同功能型植物器官中N、P含量沿环境梯度的分异规律, 有助于揭示陆地生态系统物质循环和植物养分适应策略的过程和机制(Aerts & Chapin, 2000; 曾德慧和陈广生, 2005; 刘超等, 2012). ...
The vegetation N:P ratio: A new tool to detect the nature of nutrient limitation
1
1996
... 氮(N)和磷(P)是植物生命活动所需的大量元素, 其含量大小系植物的主要功能性状, 在生态系统功能和动态中起关键作用(Koerselman & Meuleman, 1996; Reich, 2005).为了最大化地满足生长, 植物通过平衡限制性资源的分配, 在气候、土壤养分等改变时调整分配策略以适应环境限制(Reich & Oleksyn, 2004), 因此, 气候、土壤成为影响植物营养器官N、P含量的主要环境因子.研究发现叶片N、P含量与气候和土壤的关系因各区域限制性因子的差异而不同(Reich & Oleksyn, 2004; Han et al., 2005; Kerkhoff et al., 2005; He et al., 2008; 丁凡等, 2011; 王晶苑等, 2011; Sardans & Peñuelas, 2013; Hong et al., 2014; Yang et al., 2014); 同时, 相同功能型植物往往趋于分享一套关键的植物功能性状来实现对环境的趋同适应和对生态系统的相似作用(Díaz & Cabido, 2001).因此, 探索不同功能型植物器官中N、P含量沿环境梯度的分异规律, 有助于揭示陆地生态系统物质循环和植物养分适应策略的过程和机制(Aerts & Chapin, 2000; 曾德慧和陈广生, 2005; 刘超等, 2012). ...
黄土高原优势灌丛营养器官化学计量特征的环境分异和机制
3
2015
... 灌丛作为一种重要的自然植被类型, 不同于森林的主要特点是以灌木生活型植物为优势成分, 具有类型多样、分布广泛、生产力高、萌生能力强等特点, 其分布面积约占中国国土面积的20%, 约占中国亚热带国土面积的25% (中国科学院中国植被图编辑委员会, 2007), 是陆地生态系统养分循环的重要场所(胡会峰等, 2006; Piao et al., 2009).截至目前, 关于植物养分循环的研究多聚焦于森林或草地(张丽霞, 2003; He et al., 2006, 2008; 任书杰等, 2007; 甘露等, 2008; 丁凡等, 2011; 林志斌等, 2011; Chen et al., 2013; Cornelissen et al., 2014; Hong et al., 2014; Nottingham et al., 2015; Pan et al., 2015; Petter et al., 2016), 发现不同功能型植物叶N、P含量差异显著, 且落叶木本植物叶的N、P含量显著高于常绿木本植物.尽管植物在灌丛中的生长环境和地位与森林或草地截然不同, 但关于灌丛中植物器官N、P格局却较少受到重视, 且相关研究多在北方(韩文轩等, 2009; Yang et al., 2014; 李单凤等, 2015); 再者, 众多研究在区域和全球尺度阐释了叶N、P的地理变异机制(Elser et al., 2007), 但大部分研究并没有检验不同功能型植物叶N、P对环境因子的响应规律(Chen et al., 2013).因此, 在较大范围内研究灌丛生态系统不同生活型优势植物N、P含量及其与环境因子的关系, 有助于深入了解植物N、P养分循环和植物养分适应策略的过程和机制. ...
Tree growth changes with climate and forest type are associated with relative allocation of nutrients, especially phosphorus, to leaves and wood
2
2013
... 氮(N)和磷(P)是植物生命活动所需的大量元素, 其含量大小系植物的主要功能性状, 在生态系统功能和动态中起关键作用(Koerselman & Meuleman, 1996; Reich, 2005).为了最大化地满足生长, 植物通过平衡限制性资源的分配, 在气候、土壤养分等改变时调整分配策略以适应环境限制(Reich & Oleksyn, 2004), 因此, 气候、土壤成为影响植物营养器官N、P含量的主要环境因子.研究发现叶片N、P含量与气候和土壤的关系因各区域限制性因子的差异而不同(Reich & Oleksyn, 2004; Han et al., 2005; Kerkhoff et al., 2005; He et al., 2008; 丁凡等, 2011; 王晶苑等, 2011; Sardans & Peñuelas, 2013; Hong et al., 2014; Yang et al., 2014); 同时, 相同功能型植物往往趋于分享一套关键的植物功能性状来实现对环境的趋同适应和对生态系统的相似作用(Díaz & Cabido, 2001).因此, 探索不同功能型植物器官中N、P含量沿环境梯度的分异规律, 有助于揭示陆地生态系统物质循环和植物养分适应策略的过程和机制(Aerts & Chapin, 2000; 曾德慧和陈广生, 2005; 刘超等, 2012). ...
... 植物生活型是叶N、P含量的一个主要决定因子, 以往大量研究已注意到不同生活型植物器官N、P化学计量存在显著差异(Reich & Oleksyn, 2004; Han et al., 2008; Chen et al., 2013; Sardans & Peñuelas, 2013; Pan et al., 2015), 发现生命周期长且生长慢的植物器官N、P含量较生命周期短且生长迅速的植物高, 并认为植物器官N、P含量是植物代谢能力以及养分利用效率的综合反映(Han et al., 2005; Ågren, 2008).本研究中, 常绿和落叶木本植物叶N、P含量存在显著差异, 落叶木本植物叶的N、P含量(17.91 mg·g-1, 1.14 mg·g-1)显著高于常绿木本植物叶(15.19 mg·g-1, 0.89 mg·g-1)(表1), 生活型可分别解释叶片N、P含量7.6%和14.4%的变异(表2).该结果主要是因为常绿木本植物叶寿命较长, 与落叶木本植物相比, 通常具有更长的养分驻留时间、保守的养分利用策略和更高的养分利用效率; 为了适应低养环境, 常绿木本植物需要投资更多能量用于叶片保护和结构支撑, 从而导致其比落叶木本植物更低的光合速率和养分含量(Edwards et al., 2014), 但常绿性意味着具有较长的光合时间, 保证其具有较高的养分利用效率, 从而减少对土壤养分的依赖.然而, 落叶木本植物通过叶片凋落减少其在不利季节的蒸散和呼吸; 虽然落叶习性意味着较短的叶生命周期, 却有较高的凋落物分解和养分重吸收率, 有着较常绿植物高的光合速率和养分含量.结果证明不同生活型植物的物种特性对养分利用策略的影响较大, 进而决定植物器官的营养元素含量(He et al., 2008; Wu et al., 2012), 支持生长速率假说(Ågren, 2008). ...
Global biogeography of plant chemistry: Filling in the blanks
1
2005
... 氮(N)和磷(P)是植物生命活动所需的大量元素, 其含量大小系植物的主要功能性状, 在生态系统功能和动态中起关键作用(Koerselman & Meuleman, 1996; Reich, 2005).为了最大化地满足生长, 植物通过平衡限制性资源的分配, 在气候、土壤养分等改变时调整分配策略以适应环境限制(Reich & Oleksyn, 2004), 因此, 气候、土壤成为影响植物营养器官N、P含量的主要环境因子.研究发现叶片N、P含量与气候和土壤的关系因各区域限制性因子的差异而不同(Reich & Oleksyn, 2004; Han et al., 2005; Kerkhoff et al., 2005; He et al., 2008; 丁凡等, 2011; 王晶苑等, 2011; Sardans & Peñuelas, 2013; Hong et al., 2014; Yang et al., 2014); 同时, 相同功能型植物往往趋于分享一套关键的植物功能性状来实现对环境的趋同适应和对生态系统的相似作用(Díaz & Cabido, 2001).因此, 探索不同功能型植物器官中N、P含量沿环境梯度的分异规律, 有助于揭示陆地生态系统物质循环和植物养分适应策略的过程和机制(Aerts & Chapin, 2000; 曾德慧和陈广生, 2005; 刘超等, 2012). ...
中国东部南北样带654种植物叶氮和磷的化学计量学特征研究
3
2007
... 灌丛作为一种重要的自然植被类型, 不同于森林的主要特点是以灌木生活型植物为优势成分, 具有类型多样、分布广泛、生产力高、萌生能力强等特点, 其分布面积约占中国国土面积的20%, 约占中国亚热带国土面积的25% (中国科学院中国植被图编辑委员会, 2007), 是陆地生态系统养分循环的重要场所(胡会峰等, 2006; Piao et al., 2009).截至目前, 关于植物养分循环的研究多聚焦于森林或草地(张丽霞, 2003; He et al., 2006, 2008; 任书杰等, 2007; 甘露等, 2008; 丁凡等, 2011; 林志斌等, 2011; Chen et al., 2013; Cornelissen et al., 2014; Hong et al., 2014; Nottingham et al., 2015; Pan et al., 2015; Petter et al., 2016), 发现不同功能型植物叶N、P含量差异显著, 且落叶木本植物叶的N、P含量显著高于常绿木本植物.尽管植物在灌丛中的生长环境和地位与森林或草地截然不同, 但关于灌丛中植物器官N、P格局却较少受到重视, 且相关研究多在北方(韩文轩等, 2009; Yang et al., 2014; 李单凤等, 2015); 再者, 众多研究在区域和全球尺度阐释了叶N、P的地理变异机制(Elser et al., 2007), 但大部分研究并没有检验不同功能型植物叶N、P对环境因子的响应规律(Chen et al., 2013).因此, 在较大范围内研究灌丛生态系统不同生活型优势植物N、P含量及其与环境因子的关系, 有助于深入了解植物N、P养分循环和植物养分适应策略的过程和机制. ...
... 为了衡量木本植物叶N、P含量与气候和土壤养分之间的关系, 将每个样点上分种叶N、P含量原始值进行对数化转换, 分别与MAT、MAP、土壤N含量和土壤P含量进行一般线性回归(任书杰等, 2007; Hong et al., 2014).同时, 采用广义线性模型(GLM)来衡量生活型、MAT、MAP、土壤N含量和P含量分别对木本植物叶N含量和P含量变异的解释量(Chen et al., 2013; Zhao et al., 2014).所有的统计分析和作图分别在R 3.2.1和Excel 2013中进行. ...
... 植物的营养元素含量是植物在一定生境下吸收营养元素的能力, 也是植物与环境之间相互作用的结果.在全球或国家尺度上, 已有研究表明植物叶N、P含量随MAT增高和MAP增加逐渐减少(Güsewell, 2004; Hedin, 2004; McGroddy et al., 2004; He et al., 2006; Ågren & Weih, 2012; Chen et al., 2013).然而, 中国南方灌丛中常绿和落叶木本植物叶N、P含量与气候因子的关系不同(图1, 图2), 其中常绿木本植物叶N、P含量与Han等(2005)与Reich和Oleksyn (2004)的研究结果存在一样的温度趋势, 结果支持温度-植物生理假说: 叶片N、P含量的增加可补偿低温下代谢速率的下降, 因此随温度的增加, 叶片N、P含量呈降低趋势(Reich & Oleksyn, 2004).同时, 较高的叶片N、P含量也是植物适应干旱条件的反映(Yang et al., 2014), 也可能是较多的降水可增强土壤N、P的淋溶作用, 土壤养分的可获得性降低, 从而导致常绿木本植物叶N、P含量呈降低的趋势.然而, 落叶木本植物叶N、P含量随MAT和MAP的升高而呈增加趋势, 可能与较高的温度和较多的降水会加快养分周转, 增加土壤养分的可获得性有关; 再者, 在温暖湿润的环境中, 落叶木本植物生长较快, 较高的叶N、P含量有助于满足植物快速生长的养分需求, 因此落叶木本植物叶N、P含量随年平均气温和年降水量的增加呈增加趋势. ...
生态化学计量学——复杂生命系统奥秘的探索
1
2005
... 氮(N)和磷(P)是植物生命活动所需的大量元素, 其含量大小系植物的主要功能性状, 在生态系统功能和动态中起关键作用(Koerselman & Meuleman, 1996; Reich, 2005).为了最大化地满足生长, 植物通过平衡限制性资源的分配, 在气候、土壤养分等改变时调整分配策略以适应环境限制(Reich & Oleksyn, 2004), 因此, 气候、土壤成为影响植物营养器官N、P含量的主要环境因子.研究发现叶片N、P含量与气候和土壤的关系因各区域限制性因子的差异而不同(Reich & Oleksyn, 2004; Han et al., 2005; Kerkhoff et al., 2005; He et al., 2008; 丁凡等, 2011; 王晶苑等, 2011; Sardans & Peñuelas, 2013; Hong et al., 2014; Yang et al., 2014); 同时, 相同功能型植物往往趋于分享一套关键的植物功能性状来实现对环境的趋同适应和对生态系统的相似作用(Díaz & Cabido, 2001).因此, 探索不同功能型植物器官中N、P含量沿环境梯度的分异规律, 有助于揭示陆地生态系统物质循环和植物养分适应策略的过程和机制(Aerts & Chapin, 2000; 曾德慧和陈广生, 2005; 刘超等, 2012). ...
植物N:P计量化学: 中国高等植物的分异规律与野外实验初步验证
1
2003
... 灌丛作为一种重要的自然植被类型, 不同于森林的主要特点是以灌木生活型植物为优势成分, 具有类型多样、分布广泛、生产力高、萌生能力强等特点, 其分布面积约占中国国土面积的20%, 约占中国亚热带国土面积的25% (中国科学院中国植被图编辑委员会, 2007), 是陆地生态系统养分循环的重要场所(胡会峰等, 2006; Piao et al., 2009).截至目前, 关于植物养分循环的研究多聚焦于森林或草地(张丽霞, 2003; He et al., 2006, 2008; 任书杰等, 2007; 甘露等, 2008; 丁凡等, 2011; 林志斌等, 2011; Chen et al., 2013; Cornelissen et al., 2014; Hong et al., 2014; Nottingham et al., 2015; Pan et al., 2015; Petter et al., 2016), 发现不同功能型植物叶N、P含量差异显著, 且落叶木本植物叶的N、P含量显著高于常绿木本植物.尽管植物在灌丛中的生长环境和地位与森林或草地截然不同, 但关于灌丛中植物器官N、P格局却较少受到重视, 且相关研究多在北方(韩文轩等, 2009; Yang et al., 2014; 李单凤等, 2015); 再者, 众多研究在区域和全球尺度阐释了叶N、P的地理变异机制(Elser et al., 2007), 但大部分研究并没有检验不同功能型植物叶N、P对环境因子的响应规律(Chen et al., 2013).因此, 在较大范围内研究灌丛生态系统不同生活型优势植物N、P含量及其与环境因子的关系, 有助于深入了解植物N、P养分循环和植物养分适应策略的过程和机制. ...
The altitudinal patterns of leaf C:N:P stoichi- ometry are regulated by plant growth form, climate and soil on Changbai Mountain, China
1
2014
... 为了衡量木本植物叶N、P含量与气候和土壤养分之间的关系, 将每个样点上分种叶N、P含量原始值进行对数化转换, 分别与MAT、MAP、土壤N含量和土壤P含量进行一般线性回归(任书杰等, 2007; Hong et al., 2014).同时, 采用广义线性模型(GLM)来衡量生活型、MAT、MAP、土壤N含量和P含量分别对木本植物叶N含量和P含量变异的解释量(Chen et al., 2013; Zhao et al., 2014).所有的统计分析和作图分别在R 3.2.1和Excel 2013中进行. ...



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}