作为生态系统最基本的特征指标, 生物量直接反映了生态系统生产力水平的高低(Keeling & Phillips, 2007 ), 同时作为衡量植被固碳能力的重要参数, 生物量的准确测定在碳循环与气候变化相关研究中具有重要意义(Dai et al ., 2013 )。碳密度是指单位土地面积的碳存储量, 是反映生态系统碳存储能力的重要指标之一, 作为估算生态系统碳储量和研究生态系统与大气碳交换的重要参数, 其准确估算是判定碳汇、制定缓解全球变化的合理政策措施的重要依据(吕超群和孙书存, 2004 )。
灌丛通常是指高度在3 m以内、茎干具有多个分枝且主干不明显的木本植物。在以往研究中, 灌丛常被当作介于森林和草地之间的一个过渡类型, 并没有受到足够的重视。在五年一次的中国森林资源清查工作中, 灌丛(灌木林)也只有一个简单的面积信息① (① 森林资源管理司(2010). 全国森林资源统计——第七次全国森林资源清查.)。目前有关生物量和碳密度方面的研究, 灌丛远远滞后于森林和草地(胡会峰等, 2006 )。随着全球气候变暖和人类活动的加强, 灌丛入侵草地、森林等问题越来越突出(Sturm et al ., 2001 ; Montane et al ., 2007 ), 草地向灌丛生态系统的演替加快, 进而增大灌丛分布面积, 这被认为是我国陆地生态系统碳储量增加的主要原因(王玲, 2009 ; 朴世龙等, 2010 )。然而灌丛植被的碳汇能力是目前我国陆地生态系统碳汇研究中最不确定的一个因素(Piao et al ., 2009 ), 因此加强灌丛群落生物量和碳密度相关研究, 对于准确估算我国陆地生态系统碳汇功能和固碳潜力具有重要意义。
在西藏, 灌丛分布相当普遍, 几乎除羌塘高原西北部以外都有分布, 其面积达13 × 106 hm2 , 已经超过森林, 成为西藏陆地生态系统中分布面积仅次于草地的一个重要类型(中国科学院中国植被图编辑委员会, 2001 ), 因此灌丛是西藏植被碳汇估算中不可或缺的组成部分。然而, 目前有关西藏灌丛生物量方面的研究鲜有报道(王启基等, 1991 ; 李英年等, 2006 ; 梁倍等, 2013 ), 涉及根系生物量的研究则更少(高巧等, 2014 )。西藏雅鲁藏布江(以下简称雅江)中游地区是西藏经济社会发展的核心区域, 而砂生槐(Sophora moorcroftiana )灌丛正是该区分布最广、最具代表性的灌丛类型, 其受人类活动的影响较大, 因此探索该区砂生槐灌丛群落的生物量现状及其分配规律对于指导当地生态建设具有现实意义。砂生槐灌丛主要分布在雅江流域中段的宽谷、两侧低山及拉萨河、年楚河等主要支流的宽谷内。作为典型的河谷灌丛, 砂生槐具有良好的防风固沙、涵养水源等重要生态作用(彭跃明, 1997 )。以往研究主要集中在砂生槐的种群和群落结构、繁殖特性、固沙特性和物种多样性等方面(赵文智, 1998 ; 郭其强等, 2009 ; 李海东等, 2011 ; 王文娟等, 2011 ; 杨兆平等, 2011 ; 赵垦田等, 2013 ), 尚未见到有关群落生物量和碳密度方面的报道。本研究采用相对生长法对西藏雅鲁藏布江流域中游的砂生槐灌丛进行地上和地下不同器官的生物量调查, 利用样方法测定了草本植物和凋落物的生物量, 并结合实测的全碳含量对灌丛植被碳密度进行了详尽计算, 以期为准确评价西藏灌丛生态系统碳收支和碳平衡提供基础数据, 为指导当地生态建设提供科学依据。
1 研究区域和方法
1.1 研究区概况
调查工作主要在西藏自治区的贡嘎县、扎囊县、南木林县、谢通门县、曲水县、日喀则市和拉萨市境内开展, 地处雅鲁藏布江流域中游的宽谷、两侧低山及拉萨河的宽谷, 海拔3500-3900 m, 气候温暖干燥, 年平均气温5-9 ℃, 年降水量300-450 mm。地表多风化碎石, 土壤以沙土为主。由于河谷中常有风成沙地或沙丘, 使喜干暖、抗风沙并耐贫瘠的砂生槐得以大量分布, 成为该区干暖河谷中最具代表性的灌丛植物。常形成成片的单优群落, 在固定、半固定沙丘及附近地区常与半灌木状的藏沙蒿(Artemisia wellbyi )形成共优种群落, 而在一些坡地上则与野丁香(Leptodermis potanini )形成共优种群落。其他常见的伴生物种有固沙草(Orinus thoroldii )、毛瓣棘豆(Oxytropis sericopetala )、白草(Pennisetum flaccidum )和小角柱花(又叫小蓝雪花Ceratostigma minus )等。
1.2 研究方法
1.2.1 样点选择与样地设置
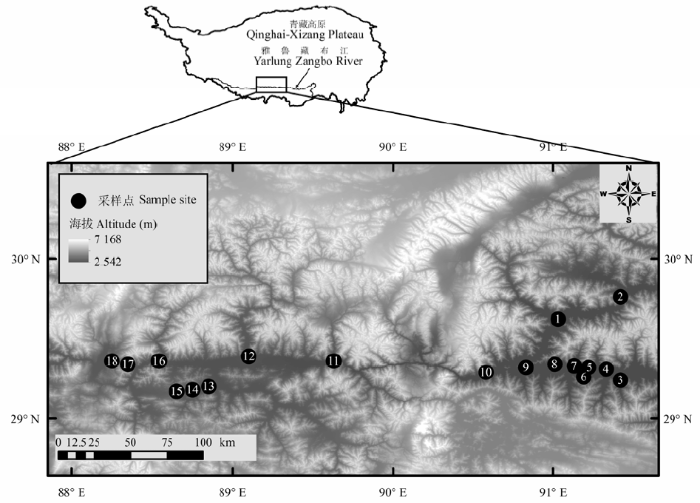
基于《1:1000000中国植被图集》(中国科学院中国植被图编辑委员会, 2001 ), 我们于2011-2013年对该区砂生槐灌丛进行了实地调查。考虑到交通的可达性, 我们主要在公路沿线开展工作, 选取典型的砂生槐分布区进行调查, 样点的选择标准包括: 1)群落总分布面积>5 hm2 , 2)距公路200 m以上, 3)原则上两样点间距离>30 km。最后选择了18个砂生槐灌丛调查样点(图1 )。
图1 砂生槐调查样点分布图。
Fig. 1 Distribution of sampled sites for Sophora moorcroftiana shrublands.
1.2.2 灌木层生物量估算
每个样点设置一个20 m × 20 m的代表性样地, 在样地内沿其中一条对角线设置3个5 m × 5 m的样方, 测量样方内每一株灌木的高度(H )和冠幅(包括长度和垂直于长轴的宽度), 将每丛灌木的垂直投影面积视为一个标准椭圆, 利用冠幅长、宽计算灌木投影面积(C )。每个样点选取3-5株不同大小的标准株, 共获取52株标准株, 将每株标准株分枝、叶、根称取鲜质量, 取样后带回实验室, 在70 ℃下烘干至恒质量(48 h), 称取样品干质量数据。根据样品干料率以推算各器官干质量。分别采用线性、指数、对数及幂函数模型建立各器官生物量与调查因子(H 、C )及其组合(生物量体积, V = C × H )的回归方程, 依据决定系数R 2 的大小评价方程的优劣, 选出拟合度好且具有生物学意义的数学模型来最终描述各器官生物量和各调查因子或其组合的关系, 进而利用该关系模型估算调查样地内灌木层各器官生物量。
1.2.3 草本层、凋落物生物量估算
利用样方收获法估算草本层生物量, 即在每个样方内选择1个50 cm × 50 cm的小样方, 收获该样方中的所有草本, 分别称取地上部分和地下部分鲜质量, 取样带回室内, 烘干至恒质量, 通过样品干料率计算草本层地上与地下生物量。
凋落物生物量估算类似草本层生物量估算, 在每个灌木样方内设置1个1 m × 1 m的样方作为凋落物生物量调查小样方, 收集样方内的所有凋落物, 称取鲜质量并取样带回室内, 测定干料率后推算单位土地面积的凋落物量。
1.2.4 生物量碳密度估算及东、西部碳密度比较
利用各器官生物量和对应的实测全碳含量估算灌丛碳密度。所有样品均在植被与环境变化国家重点实验室进行全碳含量测定, 测定方法为重铬酸钾- 硫酸氧化法。
由于砂生槐灌丛具有在宽谷地带成片分布的特点, 而拉萨与日喀则之间的高山峡谷正好将本研究中的雅江中游地带分割成东、西两个砂生槐灌丛较集中的宽谷区域, 加之该区域具有西高东低的地势特征, 大致可以90° E为界将18个采样点分划为东、西两部分, 从而进一步比较不同区域砂生槐灌丛碳密度的差异。
2 结果和分析
2.1 砂生槐灌丛灌木层生物量
砂生槐灌木不同器官干质量均与调查因子显著相关(表1 ), 就拟合方程而言, 对数模型和指数模型的拟合效果较差(R 2 = 0.35-0.65), 幂函数模型和线性模型拟合效果较好(R 2 = 0.50-0.94), 而幂函数模型(R 2 = 0.53-0.94)又优于线性模型(R 2 = 0.50-0.90), 因此能更好地描述各器官干质量与调查因子的关系。比较不同器官的幂函数模型, 发现植株冠幅(R 2 = 0.86-0.94)或生物量体积(R 2 = 0.82-0.92)对器官生物量的预测能力远高于植株高度(R 2 = 0.53-0.75), 其中, 冠幅能帮助我们更好地预测灌木的地上生物量, 而生物量体积有助于我们更好地预测地下部分, 因此, 我们选用关系最好的3个模型——Wb = 0.0011C 1.279 , Wl = 0.003C 1.039 , Wr = 0.024V 0.789 , 计算样地中砂生槐灌木的生物量。
18个样点的估算结果表明, 西藏雅江流域典型地段砂生槐灌丛灌木层平均总生物量为4.08 Mg·hm-2 , 变化范围在1.63-7.71 Mg·hm-2 , 其中, 枝条的平均生物量为1.57 Mg·hm-2 (0.62-3.16 Mg·hm-2 ), 占灌木层生物量的38.48%; 叶的平均生物量为0.43 Mg·hm-2 (0.17-0.76 Mg·hm-2 ), 占10.54%; 根的平均生物量为2.08 Mg·hm-2 (0.84-4.12 Mg·hm-2 ), 占50.98% (表2 )。砂生槐灌木层地下与地上生物量之比为1.05。
2.2 草本层生物量及凋落物量
砂生槐灌丛群落的草本层平均生物量为1.53 Mg·hm-2 , 变化范围在0.61-4.92 Mg·hm-2 。其中, 地上部分平均生物量为0.67 Mg·hm-2 (0.53-2.93 Mg·hm-2 ), 占草本层总生物量的43.79%; 地下部分为0.86 Mg·hm-2 , 占56.21%, 且变异较大, 最低仅为0.08 Mg·hm-2 , 最高可达2.81 Mg·hm-2 (表3 )。总体来看, 地下部分生物量略高于地上部分, 但差异并不显著(p > 0.05)。平均凋落物量为0.1 Mg·hm-2 , 变化范围在0.08-0.27 Mg·hm-2 。
2.3 砂生槐灌丛群落总生物量及碳密度
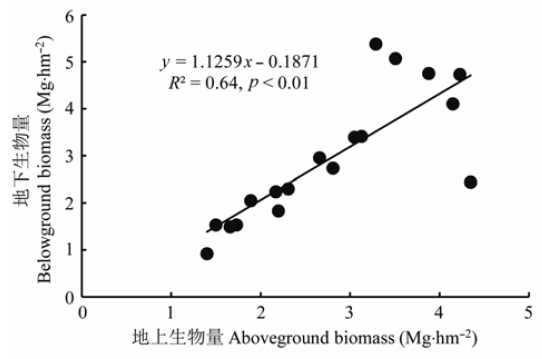
西藏雅鲁藏布江流域中段砂生槐灌丛群落平均总生物量为5.71 Mg·hm-2 , 变化范围在2.32-8.96 Mg·hm-2 , 其中灌木层平均为4.08 Mg·hm-2 , 占群落总生物量的71.45%; 草本层为1.53 Mg·hm-2 , 占26.80%; 凋落物量仅为0.1 Mg·hm-2 , 占1.75%。进一步分析群落地上生物量与地下生物量的关系, 发现二者显著正相关(图2 ), 表明利用地上生物量的测定数据估算研究区范围内的砂生槐灌丛植被的地下生物量是可行的。
图2 砂生槐灌丛地上部分与地下部分生物量关系。
Fig. 2 The relationship between aboveground and belowground biomass of Sophora moorcroftiana communities.
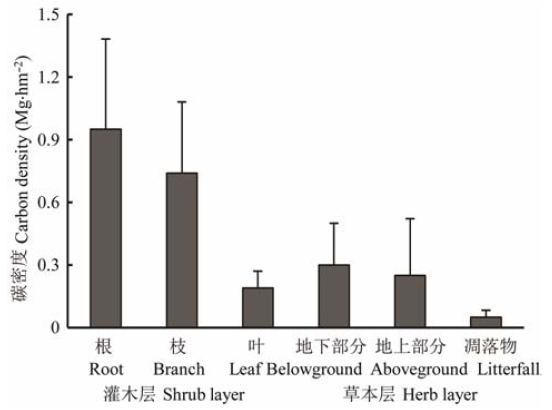
砂生槐灌丛各部分实测的平均全碳含量分别为: 枝47.02%, 叶44.92%, 根45.93%, 草本地上部分36.37%, 草本地下部分33.79%, 凋落物43.49%。利用这些实测的不同器官全碳含量数据, 结合各样点生物量计算出砂生槐灌丛平均碳密度为2.48 Mg·hm-2 , 变化范围在1.03-3.90 Mg·hm-2 , 其中, 灌木层、草本层和凋落物碳密度分别为1.88 Mg·hm-2 、0.55 Mg·hm-2 和0.05 Mg·hm-2 (图3 )。灌木层各器官碳密度所占比例分别是: 枝39.89%, 叶10.11%, 根50.00%, 碳密度主要集中在根和枝, 二者占89.89%。各部分碳密度大小顺序为灌木层根>灌木层枝>灌木层叶>草本层地下部分>草本层地上部分>凋落物。
图3 砂生槐灌丛碳密度分布(平均值±标准误差)。
Fig. 3 The distribution of carbon density (mean ± SE, Mg·hm-2 ) within Sophora moorcroftiana shrubs.
进一步比较东、西部样点碳密度的差异, 发现西部地区砂生槐灌木层碳密度显著高于东部(p < 0.05, 表4 ), 草本层和凋落物则表现为东部>西部, 但差异不显著(p > 0.05, 表4 )。
3 讨论
3.1 砂生槐灌丛对高原干旱河谷环境的适应
青藏高原由于海拔高、辐射强、温度低, 除了东南部较湿润, 高原大部分地区都具干旱半干旱气候特点, 因此高原灌丛通常较低矮、个体之间冠层不连续, 为典型的离散型灌丛。本研究中砂生槐灌丛属于干旱河谷灌丛, 其群落地上部分生物量(不含凋落物)介于1.4-6.9 Mg·hm-2 , 平均为3.53 Mg·hm-2 , 与青藏高原东缘的岷江干旱河谷灌丛地上生物量相当(1.4-4.8 Mg·hm-2 ; 刘国华等, 2003 ; 王勇军等, 2010 ), 略高于青海海北高寒金露梅灌丛(1.5-2.6 Mg·hm-2 ; 王启基等, 1991 ; 李英年等, 2006 )。目前有关高原灌丛根系生物量的报道很少, 本研究中砂生槐灌丛根系生物量在0.8-4.1 Mg·hm-2 之间, 平均为2.1 Mg·hm-2 , 高于青海海北高寒金露梅灌丛(1.5 Mg·hm-2 ; 李英年等, 2006 )和四川甘孜州高寒柳灌丛和杜鹃灌丛(1.3-1.8 Mg·hm-2 ; 高巧等, 2014 ), 这与砂生槐的深根系特点以及对干旱河谷环境的适应有关。砂生槐为典型深根系物种, 主根平均深入地下3 m, 侧根在距地表0 .7-1.5 m出现, 根幅大, 平均为1.9 × 2.3 m2 (彭跃明, 1997 ; 孙旭, 2010 )。从砂生槐器官生物量的分配来看, 根生物量占比超过一半, 使得平均根冠比达1.05, 从植被总体生物量的分配来看, 地下与地上生物量的关系斜率甚至达1.13 (图2 ), 即干旱地区植物把更多生物量分配到用于吸收水分和养分以及固定、支撑植物体的根系, 而不是地上部分的光合器官, 这些特点使砂生槐灌丛具有很强的防风固沙作用, 也体现了这一植被类型对河谷干旱环境的适应。
本研究中砂生槐灌丛植被平均碳密度为2.48 Mg·hm-2 , 与付达夫(2014)针对本研究物种灌丛碳密度估算结果(约2.5 Mg·hm-2 )一致, 但远低于胡 会峰等(2006)估算的中国主要灌丛植被平均碳密度(10.88 Mg·hm-2 )及付达夫(2014)估算的西藏灌木林碳密度(8.53 Mg·hm-2 ), 也低于四川甘孜州高寒矮灌丛(3.20 Mg·hm-2 ), 这与砂生槐灌丛所处生境以及群落盖度较低有关——在干旱的河谷地带, 为减少对水分和(或)养分资源的竞争, 砂生槐种群具有较强的自疏作用, 个体间距较大, 导致植被冠层不连续, 灌木层盖度在12%-53%之间, 平均仅为30.5%, 明显低于四川甘孜州高寒矮灌丛(22%-85%, 高巧等, 2014 )。砂生槐灌丛群落碳密度在研究区东部与西部存在显著差异, 这是由于东、西部灌丛盖度存在差异所致, 即西部地区植被盖度明显高于东部(表4 ), 究其原因, 主要与东、西部的自然环境(尤其是海拔)以及人为干扰强度的差异有关。研究区东部平均海拔较西部低200-300 m, 因此平均气温高1-2 ℃, 而年降水量略低或相近, 导致东部地区蒸发量相对较高, 因此焚风效应(过山气流在背风坡下沉而变得干热)形成的干旱河谷气候在研究区东部比在西部更明显, 这与杨志刚等(2013)利用Penman-Monteith模型计算的雅江流域潜在蒸散量结果一致, 即潜在蒸散量在位于雅江中游东部的泽当明显高于西部的日喀则(杨志刚等, 2013 )。显然, 东部地区较高的蒸散量加剧了河谷的干旱化程度, 进一步限制了该区砂生槐灌丛植被盖度的扩张。因此, 深入探究不同器官生物量分配与气候、土壤因子的关系, 将有助于揭示砂生槐灌丛的地理分异机制。此外, 研究区东部地区人口密度更大, 人类活动和放牧干扰强度较大, 牲畜践踏和啃食对砂生槐灌丛的生长和繁殖均具有不可忽视的作用, 这可能也是导致研究区东部植被盖度较低的原因之一。在未来气候变化背景下, 如果蒸散持续降低(吴绍洪等, 2005 ; Yin et al ., 2013 ), 将有助于砂生槐灌丛生物量和碳密度的增加, 这无疑对于雅江流域水土保持和防风固沙等生态工程具有重要指导意义。
3.2 碳密度不同估算方法的误差比较
目前有关碳密度的研究大部分采用平均碳密度方法(胡会峰等, 2006 ; 付达夫, 2014 ; 李亚男等, 2014 ), 即碳密度等于生物量乘以换算系数0.5或0.45。然而不同物种、不同器官之间的碳含量是存在差异的, 且碳含量通常达不到50% (宋永昌, 2001 ), 用平均碳密度法势必会高估生态系统碳密度。本研究通过实测不同器官全碳含量, 结合植被生物量计算了灌丛植被碳密度, 与利用换算系数的估算方法相比, 也发现以0.5为换算系数的估算结果比实际结果偏高, 总体高约15%, 其中灌木层偏高6%-11%, 草本层偏高36%-44% (表5 )。以0.45作为换算系数的总体估算结果与实际结果较接近(偏高3.6%), 表现为灌木层略微偏低(0-4%), 草本层偏高(20%-30%)(表5 )。综上所述, 在区域尺度上估算西藏雅江流域砂生槐灌丛群落碳密度时, 可以考虑用0.45作为碳含量的换算系数。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Dai L Jia J Yu D Lewis BJ Zhou L Zhou W Zhao W Jiang L 2013 ). Effects of climate change on biomass carbon sequestration in old-growth forest ecosystems on Changbai Mountain in Northeast China
.Forest Ecology and Management , 300 , 106 -116 .
[本文引用: 1]
[2]
Fu DF 2014 ). Shrub wood carbon reserve estimation in Tibet Autonomous Region
.Central South Forest Inventory and Planning , 33 , 4 -7 . (in Chinese with English abstract)[付达夫 (2014 ). 西藏自治区灌木林碳储量估算
. 中南林业调查规划 , 33 , 4 -7 .]
[本文引用: 1]
[3]
Gao Q Yang XC Yin CY Liu Q 2014 ). Estimation of biomass allocation and carbon density in alpine dwarf shrubs in Ganzi Zangzu Autonomous Prefecture of Sichuan Province, China
.Journal of Plant Ecology , 38 , 355 -365 . (in Chinese with English abstract)[高巧 , 阳小成 , 尹春英 , 刘庆 (2014 ). 四川省甘孜藏族自治州高寒矮灌丛生物量分配及其碳密度的估算
. 植物生态学报 , 38 , 355 -365 .]
[本文引用: 3]
[4]
Guo QQ Luo DQ Fang JP Ren DZ Wang ZH Fu JF 2009 ). Research status, utilization and protection measures of Sophora moorcroftiana in Tibet
. Journal of Northwest Forestry University , 24 , 98 -101 . (in Chinese with English abstract)[郭其强 , 罗大庆 , 方江平 , 仁德智 , 王贞红 , 傅军锋 (2009 ). 西藏砂生槐的研究现状及其利用与保护对策
. 西北林学院学报 , 24 , 98 -101 .]
[本文引用: 1]
[5]
Hu HF Wang ZH Liu GH Fu BJ 2006 ). Vegetation carbon storage of major shrublands in China
.Journal of Plant Ecology (Chinese Version) , 30 , 539 -544 . (in Chinese with English abstract)[胡会峰 , 王志恒 , 刘国华 , 傅伯杰 (2006 ). 中国主要灌丛植被碳储量
. 植物生态学报 , 30 , 539 -544 .]
[本文引用: 2]
[6]
Keeling HC Phillips OL 2007 ). The global relationship between forest productivity and biomass
.Global Ecology and Biogeography , 16 , 618 -631 .
[本文引用: 1]
[7]
Li HD Shen WS She GH Sun M Yuan L Lin NF 2011 ). Population structure and point pattern of Sophora moor- croftiana in Tibet
. Journal of Desert Research , 31 , 1443 -1448 . (in Chinese with English abstract)[李海东 , 沈渭寿 , 佘光辉 , 孙明 , 袁磊 , 林乃峰 (2011 ). 西藏砂生槐种群结构与点格局分析
. 中国沙漠 , 31 , 1443 -1448 .]
[本文引用: 1]
[8]
Li YN Yu XF Xu ZQ Liu LL Yao WX Wang L 2014 ). Carbon density characteristics of two typical shrub communities in the northern mountain region of Hebei
.Scientia Silvae Sinicae , 50 (6 ), 28 -33 . (in Chinese with English abstract)[李亚男 , 虞晓凡 , 许中旗 , 刘乐乐 , 姚卫星 , 王丽 (2014 ). 冀北山地2种典型灌丛的碳密度特征
. 林业科学 , 50 (6 ), 28 -33 .]
[本文引用: 1]
[9]
Li YN Zhao L Wang QX Du MY Gu S Xu SX Zang FW Zhao XQ 2006 ). Estimation of biomass and annual turnover quantities of Potentilla froticosa shrub
. Acta Agrestia Sinica , 14 , 72 -76 . (in Chinese with English abstract)[李英年 , 赵亮 , 王勤学 , 杜明远 , 古松 , 徐世晓 , 张发伟 , 赵新全 (2006 ). 高寒金露梅灌丛生物量及年周转量
. 草地学报 , 14 , 72 -76 .]
[本文引用: 3]
[10]
Liang B Di L Zhao CY Peng SZ Peng HH Wang C 2013 ). Spatial distribution of aboveground biomass of shrubs in Tianlaochi catchment of the Qilian Mountains
.Chinese. Journal of Applied Ecology , 25 , 367 -373 . (in Chinese with English abstract)[梁倍 , 邸利 , 赵传燕 , 彭守璋 , 彭焕华 , 王超 (2013 ). 祁连山天老池流域灌丛地上生物量空间分布
. 应用生态学报 , 25 , 367 -373 .]
[本文引用: 1]
[11]
Liu GH Zhang JY Zhang YX Zhou JY Guan WB Ma KM Fu BJ 2003 ). Distribution regulation of aboveground biomass of three main shrub types in the dry valley of Minjiang River
.Journal of Mountain Science , 21 , 24 -32 . (in Chinese with English abstract)[刘国华 , 张洁瑜 , 张育新 , 周建云 , 关文彬 , 马克明 , 傅伯杰 (2003 ). 岷江干旱河谷三种主要灌丛地上生物量的分布规律
. 山地学报 , 21 , 24 -32 .]
[本文引用: 1]
[12]
Lü CQ Sun SC 2004 ). A review on the distribution patterns of carbon density in density in terrestrial ecosystems
.Acta Phytoecologica Sinica , 28 , 692 -703 . (in Chinese with English abstract)[吕超群 , 孙书存 (2004 ). 陆地生态系统碳密度格局研究概述
. 植物生态学报 , 28 , 692 -703 .]
[本文引用: 1]
[13]
Montane F Rovira P Casals P 2007 ). Shrub encroachment into mesic mountain grasslands in the Iberian peninsula: Effects of plant quality and temperature on soil C and N stocks
.Global Biogeochemical Cycles , 21 , 1 -10 .
[本文引用: 1]
[14]
Peng YM 1997 ). Study on sand fixation effect of Sophora moorcroftiana in Yaluzangbu river middle valley, Tibet
. Forestry Science and Technology , 22 , 6 -8 . (in Chinese with English abstract)[彭跃明 (1997 ). 西藏雅鲁藏布江中部流域砂生槐固沙作用研究
. 林业科技 , 22 , 6 -8 .]
[本文引用: 2]
[15]
Piao SL Fang JY Ciais P Peylin P Huang Y Sitch S Wang T 2009 ). The carbon balance of terrestrial ecosystems in China
.Nature, 458 , 1009 -1014 .
[本文引用: 1]
[16]
Piao SL Fang JY Huang Y 2010 ). The carbon balance of terrestrial ecosystems in China
.China Basic Science , 12 , 20 -22 . (in Chinese with English abstract)[朴世龙 , 方精云 , 黄耀 (2010 ). 中国陆地生态系统碳收支
. 中国基础科学 , 12 , 20 -22 .]
[本文引用: 1]
[17]
Song YC 2001 ). Vegetation Ecology . East China Normal University Press , Shanghai . 353 -422 . (in Chinese)[宋永昌 (2001 ). 植被生态学 . 华东师范大学出版社 , 上海 . 353 -422 .]
[本文引用: 1]
[18]
Sturm M Racine C Tape K 2001 ). Increasing shrub abundance in the Arctic
.Nature , 411 , 546 -547 .
[本文引用: 1]
[19]
Sun X 2010 ). Study on Community Structure and Biomass of Xizang Sophora moorcroftiana
. Master degree disserta- tion, Agricultural and Animal Husbandry College of Tibet University, Linzhi, Xizang . (in Chinese with English abstract)[孙旭 (2010 ). 西藏砂生槐群落结构与生物量研究
. 硕士学位论文, 西藏大学农牧学院, 西藏林芝 .]
[本文引用: 1]
[20]
The Editorial Committee of Vegetation Map of China, Chinese Academy of Sciences (2001 ). 1:1000000 Vegetation Atlas of China . Science Press , Beijing . (in Chinese)[中国科学院中国植被图编辑委员会 (2001 ). 1:1000000中国植被图集 . 科学出版社 , 北京 .]
[本文引用: 2]
[21]
Wang L 2009 ). Study of Biomass and Its Models of Main Shrub Community Type in Northwest Sichuan
. Master degree dissertation, Sichuan Agricultural University, Ya’an, Sichuan . (in Chinese with English abstract)[王玲 (2009 ). 川西北地区主要灌丛类型生物量及其模型研究
. 硕士学位论文, 四川农业大学, 四川雅安 .]
[本文引用: 1]
[22]
Wang QJ Zhou XM Zhang YQ Zhao XQ 1991 ). Structure characteristics and biomass of Potentilla froticosa shrub in Qinghai Plateau
. Acta Botanica Boreali-Occidentalia Sinica , 11 , 333 -340 . (in Chinese with English abstract)[王启基 , 周兴民 , 张堰青 , 赵新全 (1991 ). 青藏高原金露梅灌丛的结构特征及其生物量
. 西北植物学报 , 11 , 333 -340 .]
[本文引用: 2]
[23]
Wang WJ He DH Tang XQ Gong WF 2011 ). Effects of different temperature and sand burial depths on seed germination seedling growth ofSophora moorcroftiana
. Journal of Desert Research , 31 , 1437 -1442 . (in Chinese with English abstract)[王文娟 , 贺达汉 , 唐小琴 , 巩文峰 (2011 ). 不同温度和沙埋深度对砂生槐种子萌发及幼苗生长的影响
. 中国沙漠 , 31 , 1437 -1442 .]
[本文引用: 1]
[24]
Wang YJ Huang CD Zhang J Yang WQ Wang XS 2010 ). Species diversity, biomass and their relationship of shrubberies in an arid valley of the Minjiang River
.Arid Zone Research , 27 , 567 -572 . (in Chinese with English abstract)[王勇军 , 黄从德 , 张健 , 杨万勤 , 王宪帅 (2010 ). 岷江干旱河谷灌丛物种多样性、生物量及其关系
. 干旱区研究 , 27 , 567 -572 .]
[本文引用: 1]
[25]
Wu SH Yin YH Zheng D Yang QY 2005 ). Climate changes in the Tibetan Plateau during the last three decades
.Acta Geographica Sinica , 60 , 3 -11 . (in Chinese with English abstract)[吴绍洪 , 尹云鹤 , 郑度 , 杨勤业 (2005 ). 青藏高原近30年气候变化趋势,
地理学报 , 60 , 3 -11 .]
[本文引用: 1]
[26]
Yang ZG Tang XP Lu HY Ma PF Hong JC 2013 ). The hanges of potential evapotranspiration over Yarlung Zangbo River Basin during 1961-2010
.Acta Geographica Sinica , 68 , 1263 -1268 . (in Chinese with English abstract)[杨志刚 , 唐小萍 , 路红亚 , 马鹏飞 , 洪建昌 (2013 ). 近50年雅鲁藏布江流域潜在蒸散量的变化特征
. 地理学报 , 68 , 1263 -1268 .]
[本文引用: 1]
[27]
Yang ZP Shen WS Sun M Sun J Li HD 2011 ). Structural characteristics ofSophora moorcroftiana community on wind-sandy land in middle reaches of Yaluzangbu River
. Chinese Journal of Applied Ecology , 21 , 1121 -1126 . (in Chinese with English abstract)[杨兆平 , 沈渭寿 , 孙明 , 孙俊 , 李海东 (2011 ). 雅鲁藏布江中游河谷风沙化土地砂生槐群落结构特征
. 应用生态学报 , 21 , 1121 -1126 .]
[本文引用: 1]
[28]
Yin YH Wu SH Zhao DS 2013 ). Past and future spatiotemporal changes in evapotranspiration and effective moisture on the Tibetan Plateau
.Journal of Geophysical Research: Atmosphere, 118 , 10850 -10860 .
[本文引用: 1]
[29]
Zhao KT Yang XL Ma HP Zhang XJ 2013 ). Analyses on community characteristics and soil microorganism dynamics during ecological restoration ofSophora moorcroftiana in the semi-arid valley of Lhasa
. Scientia Silvae Sinicae , 49 (2 ), 25 -20 . (in Chinese with English abstract)[赵垦田 , 杨小林 , 马和平 , 张新军 (2013 ). 拉萨半干旱河谷砂生槐灌丛生态恢复过程的群落特征与土壤微生物动态分析
. 林业科学 , 49 (2 ), 25 -20 .]
[本文引用: 1]
[30]
Zhao WZ 1998 ). A preliminary study on the arenaceous adaptability ofSophora moorcroftiana
. Acta Phytoecologica Sinica , 22 , 379 -384 . (in Chinese with English abstract)[赵文智 (1998 ). 砂生槐沙生适应性初步研究
. 植物生态学报 , 22 , 379 -384 .]
[本文引用: 1]
Effects of climate change on biomass carbon sequestration in old-growth forest ecosystems on Changbai Mountain in Northeast China
1
2013
... 作为生态系统最基本的特征指标, 生物量直接反映了生态系统生产力水平的高低(Keeling & Phillips, 2007 ), 同时作为衡量植被固碳能力的重要参数, 生物量的准确测定在碳循环与气候变化相关研究中具有重要意义(Dai et al ., 2013 ).碳密度是指单位土地面积的碳存储量, 是反映生态系统碳存储能力的重要指标之一, 作为估算生态系统碳储量和研究生态系统与大气碳交换的重要参数, 其准确估算是判定碳汇、制定缓解全球变化的合理政策措施的重要依据(吕超群和孙书存, 2004 ). ...
西藏自治区灌木林碳储量估算
1
2014
... 目前有关碳密度的研究大部分采用平均碳密度方法(胡会峰等, 2006 ; 付达夫, 2014 ; 李亚男等, 2014 ), 即碳密度等于生物量乘以换算系数0.5或0.45.然而不同物种、不同器官之间的碳含量是存在差异的, 且碳含量通常达不到50% (宋永昌, 2001 ), 用平均碳密度法势必会高估生态系统碳密度.本研究通过实测不同器官全碳含量, 结合植被生物量计算了灌丛植被碳密度, 与利用换算系数的估算方法相比, 也发现以0.5为换算系数的估算结果比实际结果偏高, 总体高约15%, 其中灌木层偏高6%-11%, 草本层偏高36%-44% (表5 ).以0.45作为换算系数的总体估算结果与实际结果较接近(偏高3.6%), 表现为灌木层略微偏低(0-4%), 草本层偏高(20%-30%)(表5 ).综上所述, 在区域尺度上估算西藏雅江流域砂生槐灌丛群落碳密度时, 可以考虑用0.45作为碳含量的换算系数. ...
四川省甘孜藏族自治州高寒矮灌丛生物量分配及其碳密度的估算
3
2014
... 在西藏, 灌丛分布相当普遍, 几乎除羌塘高原西北部以外都有分布, 其面积达13 × 106 hm2 , 已经超过森林, 成为西藏陆地生态系统中分布面积仅次于草地的一个重要类型(中国科学院中国植被图编辑委员会, 2001 ), 因此灌丛是西藏植被碳汇估算中不可或缺的组成部分.然而, 目前有关西藏灌丛生物量方面的研究鲜有报道(王启基等, 1991 ; 李英年等, 2006 ; 梁倍等, 2013 ), 涉及根系生物量的研究则更少(高巧等, 2014 ).西藏雅鲁藏布江(以下简称雅江)中游地区是西藏经济社会发展的核心区域, 而砂生槐(Sophora moorcroftiana )灌丛正是该区分布最广、最具代表性的灌丛类型, 其受人类活动的影响较大, 因此探索该区砂生槐灌丛群落的生物量现状及其分配规律对于指导当地生态建设具有现实意义.砂生槐灌丛主要分布在雅江流域中段的宽谷、两侧低山及拉萨河、年楚河等主要支流的宽谷内.作为典型的河谷灌丛, 砂生槐具有良好的防风固沙、涵养水源等重要生态作用(彭跃明, 1997 ).以往研究主要集中在砂生槐的种群和群落结构、繁殖特性、固沙特性和物种多样性等方面(赵文智, 1998 ; 郭其强等, 2009 ; 李海东等, 2011 ; 王文娟等, 2011 ; 杨兆平等, 2011 ; 赵垦田等, 2013 ), 尚未见到有关群落生物量和碳密度方面的报道.本研究采用相对生长法对西藏雅鲁藏布江流域中游的砂生槐灌丛进行地上和地下不同器官的生物量调查, 利用样方法测定了草本植物和凋落物的生物量, 并结合实测的全碳含量对灌丛植被碳密度进行了详尽计算, 以期为准确评价西藏灌丛生态系统碳收支和碳平衡提供基础数据, 为指导当地生态建设提供科学依据. ...
... 青藏高原由于海拔高、辐射强、温度低, 除了东南部较湿润, 高原大部分地区都具干旱半干旱气候特点, 因此高原灌丛通常较低矮、个体之间冠层不连续, 为典型的离散型灌丛.本研究中砂生槐灌丛属于干旱河谷灌丛, 其群落地上部分生物量(不含凋落物)介于1.4-6.9 Mg·hm-2 , 平均为3.53 Mg·hm-2 , 与青藏高原东缘的岷江干旱河谷灌丛地上生物量相当(1.4-4.8 Mg·hm-2 ; 刘国华等, 2003 ; 王勇军等, 2010 ), 略高于青海海北高寒金露梅灌丛(1.5-2.6 Mg·hm-2 ; 王启基等, 1991 ; 李英年等, 2006 ).目前有关高原灌丛根系生物量的报道很少, 本研究中砂生槐灌丛根系生物量在0.8-4.1 Mg·hm-2 之间, 平均为2.1 Mg·hm-2 , 高于青海海北高寒金露梅灌丛(1.5 Mg·hm-2 ; 李英年等, 2006 )和四川甘孜州高寒柳灌丛和杜鹃灌丛(1.3-1.8 Mg·hm-2 ; 高巧等, 2014 ), 这与砂生槐的深根系特点以及对干旱河谷环境的适应有关.砂生槐为典型深根系物种, 主根平均深入地下3 m, 侧根在距地表0 .7-1.5 m出现, 根幅大, 平均为1.9 × 2.3 m2 (彭跃明, 1997 ; 孙旭, 2010 ).从砂生槐器官生物量的分配来看, 根生物量占比超过一半, 使得平均根冠比达1.05, 从植被总体生物量的分配来看, 地下与地上生物量的关系斜率甚至达1.13 (图2 ), 即干旱地区植物把更多生物量分配到用于吸收水分和养分以及固定、支撑植物体的根系, 而不是地上部分的光合器官, 这些特点使砂生槐灌丛具有很强的防风固沙作用, 也体现了这一植被类型对河谷干旱环境的适应. ...
... 本研究中砂生槐灌丛植被平均碳密度为2.48 Mg·hm-2 , 与付达夫(2014)针对本研究物种灌丛碳密度估算结果(约2.5 Mg·hm-2 )一致, 但远低于胡 会峰等(2006)估算的中国主要灌丛植被平均碳密度(10.88 Mg·hm-2 )及付达夫(2014)估算的西藏灌木林碳密度(8.53 Mg·hm-2 ), 也低于四川甘孜州高寒矮灌丛(3.20 Mg·hm-2 ), 这与砂生槐灌丛所处生境以及群落盖度较低有关——在干旱的河谷地带, 为减少对水分和(或)养分资源的竞争, 砂生槐种群具有较强的自疏作用, 个体间距较大, 导致植被冠层不连续, 灌木层盖度在12%-53%之间, 平均仅为30.5%, 明显低于四川甘孜州高寒矮灌丛(22%-85%, 高巧等, 2014 ).砂生槐灌丛群落碳密度在研究区东部与西部存在显著差异, 这是由于东、西部灌丛盖度存在差异所致, 即西部地区植被盖度明显高于东部(表4 ), 究其原因, 主要与东、西部的自然环境(尤其是海拔)以及人为干扰强度的差异有关.研究区东部平均海拔较西部低200-300 m, 因此平均气温高1-2 ℃, 而年降水量略低或相近, 导致东部地区蒸发量相对较高, 因此焚风效应(过山气流在背风坡下沉而变得干热)形成的干旱河谷气候在研究区东部比在西部更明显, 这与杨志刚等(2013)利用Penman-Monteith模型计算的雅江流域潜在蒸散量结果一致, 即潜在蒸散量在位于雅江中游东部的泽当明显高于西部的日喀则(杨志刚等, 2013 ).显然, 东部地区较高的蒸散量加剧了河谷的干旱化程度, 进一步限制了该区砂生槐灌丛植被盖度的扩张.因此, 深入探究不同器官生物量分配与气候、土壤因子的关系, 将有助于揭示砂生槐灌丛的地理分异机制.此外, 研究区东部地区人口密度更大, 人类活动和放牧干扰强度较大, 牲畜践踏和啃食对砂生槐灌丛的生长和繁殖均具有不可忽视的作用, 这可能也是导致研究区东部植被盖度较低的原因之一.在未来气候变化背景下, 如果蒸散持续降低(吴绍洪等, 2005 ; Yin et al ., 2013 ), 将有助于砂生槐灌丛生物量和碳密度的增加, 这无疑对于雅江流域水土保持和防风固沙等生态工程具有重要指导意义. ...
西藏砂生槐的研究现状及其利用与保护对策
1
2009
... 在西藏, 灌丛分布相当普遍, 几乎除羌塘高原西北部以外都有分布, 其面积达13 × 106 hm2 , 已经超过森林, 成为西藏陆地生态系统中分布面积仅次于草地的一个重要类型(中国科学院中国植被图编辑委员会, 2001 ), 因此灌丛是西藏植被碳汇估算中不可或缺的组成部分.然而, 目前有关西藏灌丛生物量方面的研究鲜有报道(王启基等, 1991 ; 李英年等, 2006 ; 梁倍等, 2013 ), 涉及根系生物量的研究则更少(高巧等, 2014 ).西藏雅鲁藏布江(以下简称雅江)中游地区是西藏经济社会发展的核心区域, 而砂生槐(Sophora moorcroftiana )灌丛正是该区分布最广、最具代表性的灌丛类型, 其受人类活动的影响较大, 因此探索该区砂生槐灌丛群落的生物量现状及其分配规律对于指导当地生态建设具有现实意义.砂生槐灌丛主要分布在雅江流域中段的宽谷、两侧低山及拉萨河、年楚河等主要支流的宽谷内.作为典型的河谷灌丛, 砂生槐具有良好的防风固沙、涵养水源等重要生态作用(彭跃明, 1997 ).以往研究主要集中在砂生槐的种群和群落结构、繁殖特性、固沙特性和物种多样性等方面(赵文智, 1998 ; 郭其强等, 2009 ; 李海东等, 2011 ; 王文娟等, 2011 ; 杨兆平等, 2011 ; 赵垦田等, 2013 ), 尚未见到有关群落生物量和碳密度方面的报道.本研究采用相对生长法对西藏雅鲁藏布江流域中游的砂生槐灌丛进行地上和地下不同器官的生物量调查, 利用样方法测定了草本植物和凋落物的生物量, 并结合实测的全碳含量对灌丛植被碳密度进行了详尽计算, 以期为准确评价西藏灌丛生态系统碳收支和碳平衡提供基础数据, 为指导当地生态建设提供科学依据. ...
中国主要灌丛植被碳储量
2
2006
... 灌丛通常是指高度在3 m以内、茎干具有多个分枝且主干不明显的木本植物.在以往研究中, 灌丛常被当作介于森林和草地之间的一个过渡类型, 并没有受到足够的重视.在五年一次的中国森林资源清查工作中, 灌丛(灌木林)也只有一个简单的面积信息① (① 森林资源管理司(2010). 全国森林资源统计——第七次全国森林资源清查.).目前有关生物量和碳密度方面的研究, 灌丛远远滞后于森林和草地(胡会峰等, 2006 ).随着全球气候变暖和人类活动的加强, 灌丛入侵草地、森林等问题越来越突出(Sturm et al ., 2001 ; Montane et al ., 2007 ), 草地向灌丛生态系统的演替加快, 进而增大灌丛分布面积, 这被认为是我国陆地生态系统碳储量增加的主要原因(王玲, 2009 ; 朴世龙等, 2010 ).然而灌丛植被的碳汇能力是目前我国陆地生态系统碳汇研究中最不确定的一个因素(Piao et al ., 2009 ), 因此加强灌丛群落生物量和碳密度相关研究, 对于准确估算我国陆地生态系统碳汇功能和固碳潜力具有重要意义. ...
... 目前有关碳密度的研究大部分采用平均碳密度方法(胡会峰等, 2006 ; 付达夫, 2014 ; 李亚男等, 2014 ), 即碳密度等于生物量乘以换算系数0.5或0.45.然而不同物种、不同器官之间的碳含量是存在差异的, 且碳含量通常达不到50% (宋永昌, 2001 ), 用平均碳密度法势必会高估生态系统碳密度.本研究通过实测不同器官全碳含量, 结合植被生物量计算了灌丛植被碳密度, 与利用换算系数的估算方法相比, 也发现以0.5为换算系数的估算结果比实际结果偏高, 总体高约15%, 其中灌木层偏高6%-11%, 草本层偏高36%-44% (表5 ).以0.45作为换算系数的总体估算结果与实际结果较接近(偏高3.6%), 表现为灌木层略微偏低(0-4%), 草本层偏高(20%-30%)(表5 ).综上所述, 在区域尺度上估算西藏雅江流域砂生槐灌丛群落碳密度时, 可以考虑用0.45作为碳含量的换算系数. ...
The global relationship between forest productivity and biomass
1
2007
... 作为生态系统最基本的特征指标, 生物量直接反映了生态系统生产力水平的高低(Keeling & Phillips, 2007 ), 同时作为衡量植被固碳能力的重要参数, 生物量的准确测定在碳循环与气候变化相关研究中具有重要意义(Dai et al ., 2013 ).碳密度是指单位土地面积的碳存储量, 是反映生态系统碳存储能力的重要指标之一, 作为估算生态系统碳储量和研究生态系统与大气碳交换的重要参数, 其准确估算是判定碳汇、制定缓解全球变化的合理政策措施的重要依据(吕超群和孙书存, 2004 ). ...
西藏砂生槐种群结构与点格局分析
1
2011
... 在西藏, 灌丛分布相当普遍, 几乎除羌塘高原西北部以外都有分布, 其面积达13 × 106 hm2 , 已经超过森林, 成为西藏陆地生态系统中分布面积仅次于草地的一个重要类型(中国科学院中国植被图编辑委员会, 2001 ), 因此灌丛是西藏植被碳汇估算中不可或缺的组成部分.然而, 目前有关西藏灌丛生物量方面的研究鲜有报道(王启基等, 1991 ; 李英年等, 2006 ; 梁倍等, 2013 ), 涉及根系生物量的研究则更少(高巧等, 2014 ).西藏雅鲁藏布江(以下简称雅江)中游地区是西藏经济社会发展的核心区域, 而砂生槐(Sophora moorcroftiana )灌丛正是该区分布最广、最具代表性的灌丛类型, 其受人类活动的影响较大, 因此探索该区砂生槐灌丛群落的生物量现状及其分配规律对于指导当地生态建设具有现实意义.砂生槐灌丛主要分布在雅江流域中段的宽谷、两侧低山及拉萨河、年楚河等主要支流的宽谷内.作为典型的河谷灌丛, 砂生槐具有良好的防风固沙、涵养水源等重要生态作用(彭跃明, 1997 ).以往研究主要集中在砂生槐的种群和群落结构、繁殖特性、固沙特性和物种多样性等方面(赵文智, 1998 ; 郭其强等, 2009 ; 李海东等, 2011 ; 王文娟等, 2011 ; 杨兆平等, 2011 ; 赵垦田等, 2013 ), 尚未见到有关群落生物量和碳密度方面的报道.本研究采用相对生长法对西藏雅鲁藏布江流域中游的砂生槐灌丛进行地上和地下不同器官的生物量调查, 利用样方法测定了草本植物和凋落物的生物量, 并结合实测的全碳含量对灌丛植被碳密度进行了详尽计算, 以期为准确评价西藏灌丛生态系统碳收支和碳平衡提供基础数据, 为指导当地生态建设提供科学依据. ...
冀北山地2种典型灌丛的碳密度特征
1
2014
... 目前有关碳密度的研究大部分采用平均碳密度方法(胡会峰等, 2006 ; 付达夫, 2014 ; 李亚男等, 2014 ), 即碳密度等于生物量乘以换算系数0.5或0.45.然而不同物种、不同器官之间的碳含量是存在差异的, 且碳含量通常达不到50% (宋永昌, 2001 ), 用平均碳密度法势必会高估生态系统碳密度.本研究通过实测不同器官全碳含量, 结合植被生物量计算了灌丛植被碳密度, 与利用换算系数的估算方法相比, 也发现以0.5为换算系数的估算结果比实际结果偏高, 总体高约15%, 其中灌木层偏高6%-11%, 草本层偏高36%-44% (表5 ).以0.45作为换算系数的总体估算结果与实际结果较接近(偏高3.6%), 表现为灌木层略微偏低(0-4%), 草本层偏高(20%-30%)(表5 ).综上所述, 在区域尺度上估算西藏雅江流域砂生槐灌丛群落碳密度时, 可以考虑用0.45作为碳含量的换算系数. ...
高寒金露梅灌丛生物量及年周转量
3
2006
... 在西藏, 灌丛分布相当普遍, 几乎除羌塘高原西北部以外都有分布, 其面积达13 × 106 hm2 , 已经超过森林, 成为西藏陆地生态系统中分布面积仅次于草地的一个重要类型(中国科学院中国植被图编辑委员会, 2001 ), 因此灌丛是西藏植被碳汇估算中不可或缺的组成部分.然而, 目前有关西藏灌丛生物量方面的研究鲜有报道(王启基等, 1991 ; 李英年等, 2006 ; 梁倍等, 2013 ), 涉及根系生物量的研究则更少(高巧等, 2014 ).西藏雅鲁藏布江(以下简称雅江)中游地区是西藏经济社会发展的核心区域, 而砂生槐(Sophora moorcroftiana )灌丛正是该区分布最广、最具代表性的灌丛类型, 其受人类活动的影响较大, 因此探索该区砂生槐灌丛群落的生物量现状及其分配规律对于指导当地生态建设具有现实意义.砂生槐灌丛主要分布在雅江流域中段的宽谷、两侧低山及拉萨河、年楚河等主要支流的宽谷内.作为典型的河谷灌丛, 砂生槐具有良好的防风固沙、涵养水源等重要生态作用(彭跃明, 1997 ).以往研究主要集中在砂生槐的种群和群落结构、繁殖特性、固沙特性和物种多样性等方面(赵文智, 1998 ; 郭其强等, 2009 ; 李海东等, 2011 ; 王文娟等, 2011 ; 杨兆平等, 2011 ; 赵垦田等, 2013 ), 尚未见到有关群落生物量和碳密度方面的报道.本研究采用相对生长法对西藏雅鲁藏布江流域中游的砂生槐灌丛进行地上和地下不同器官的生物量调查, 利用样方法测定了草本植物和凋落物的生物量, 并结合实测的全碳含量对灌丛植被碳密度进行了详尽计算, 以期为准确评价西藏灌丛生态系统碳收支和碳平衡提供基础数据, 为指导当地生态建设提供科学依据. ...
... 青藏高原由于海拔高、辐射强、温度低, 除了东南部较湿润, 高原大部分地区都具干旱半干旱气候特点, 因此高原灌丛通常较低矮、个体之间冠层不连续, 为典型的离散型灌丛.本研究中砂生槐灌丛属于干旱河谷灌丛, 其群落地上部分生物量(不含凋落物)介于1.4-6.9 Mg·hm-2 , 平均为3.53 Mg·hm-2 , 与青藏高原东缘的岷江干旱河谷灌丛地上生物量相当(1.4-4.8 Mg·hm-2 ; 刘国华等, 2003 ; 王勇军等, 2010 ), 略高于青海海北高寒金露梅灌丛(1.5-2.6 Mg·hm-2 ; 王启基等, 1991 ; 李英年等, 2006 ).目前有关高原灌丛根系生物量的报道很少, 本研究中砂生槐灌丛根系生物量在0.8-4.1 Mg·hm-2 之间, 平均为2.1 Mg·hm-2 , 高于青海海北高寒金露梅灌丛(1.5 Mg·hm-2 ; 李英年等, 2006 )和四川甘孜州高寒柳灌丛和杜鹃灌丛(1.3-1.8 Mg·hm-2 ; 高巧等, 2014 ), 这与砂生槐的深根系特点以及对干旱河谷环境的适应有关.砂生槐为典型深根系物种, 主根平均深入地下3 m, 侧根在距地表0 .7-1.5 m出现, 根幅大, 平均为1.9 × 2.3 m2 (彭跃明, 1997 ; 孙旭, 2010 ).从砂生槐器官生物量的分配来看, 根生物量占比超过一半, 使得平均根冠比达1.05, 从植被总体生物量的分配来看, 地下与地上生物量的关系斜率甚至达1.13 (图2 ), 即干旱地区植物把更多生物量分配到用于吸收水分和养分以及固定、支撑植物体的根系, 而不是地上部分的光合器官, 这些特点使砂生槐灌丛具有很强的防风固沙作用, 也体现了这一植被类型对河谷干旱环境的适应. ...
... ; 李英年等, 2006 )和四川甘孜州高寒柳灌丛和杜鹃灌丛(1.3-1.8 Mg·hm-2 ; 高巧等, 2014 ), 这与砂生槐的深根系特点以及对干旱河谷环境的适应有关.砂生槐为典型深根系物种, 主根平均深入地下3 m, 侧根在距地表0 .7-1.5 m出现, 根幅大, 平均为1.9 × 2.3 m2 (彭跃明, 1997 ; 孙旭, 2010 ).从砂生槐器官生物量的分配来看, 根生物量占比超过一半, 使得平均根冠比达1.05, 从植被总体生物量的分配来看, 地下与地上生物量的关系斜率甚至达1.13 (图2 ), 即干旱地区植物把更多生物量分配到用于吸收水分和养分以及固定、支撑植物体的根系, 而不是地上部分的光合器官, 这些特点使砂生槐灌丛具有很强的防风固沙作用, 也体现了这一植被类型对河谷干旱环境的适应. ...
祁连山天老池流域灌丛地上生物量空间分布
1
2013
... 在西藏, 灌丛分布相当普遍, 几乎除羌塘高原西北部以外都有分布, 其面积达13 × 106 hm2 , 已经超过森林, 成为西藏陆地生态系统中分布面积仅次于草地的一个重要类型(中国科学院中国植被图编辑委员会, 2001 ), 因此灌丛是西藏植被碳汇估算中不可或缺的组成部分.然而, 目前有关西藏灌丛生物量方面的研究鲜有报道(王启基等, 1991 ; 李英年等, 2006 ; 梁倍等, 2013 ), 涉及根系生物量的研究则更少(高巧等, 2014 ).西藏雅鲁藏布江(以下简称雅江)中游地区是西藏经济社会发展的核心区域, 而砂生槐(Sophora moorcroftiana )灌丛正是该区分布最广、最具代表性的灌丛类型, 其受人类活动的影响较大, 因此探索该区砂生槐灌丛群落的生物量现状及其分配规律对于指导当地生态建设具有现实意义.砂生槐灌丛主要分布在雅江流域中段的宽谷、两侧低山及拉萨河、年楚河等主要支流的宽谷内.作为典型的河谷灌丛, 砂生槐具有良好的防风固沙、涵养水源等重要生态作用(彭跃明, 1997 ).以往研究主要集中在砂生槐的种群和群落结构、繁殖特性、固沙特性和物种多样性等方面(赵文智, 1998 ; 郭其强等, 2009 ; 李海东等, 2011 ; 王文娟等, 2011 ; 杨兆平等, 2011 ; 赵垦田等, 2013 ), 尚未见到有关群落生物量和碳密度方面的报道.本研究采用相对生长法对西藏雅鲁藏布江流域中游的砂生槐灌丛进行地上和地下不同器官的生物量调查, 利用样方法测定了草本植物和凋落物的生物量, 并结合实测的全碳含量对灌丛植被碳密度进行了详尽计算, 以期为准确评价西藏灌丛生态系统碳收支和碳平衡提供基础数据, 为指导当地生态建设提供科学依据. ...
岷江干旱河谷三种主要灌丛地上生物量的分布规律
1
2003
... 青藏高原由于海拔高、辐射强、温度低, 除了东南部较湿润, 高原大部分地区都具干旱半干旱气候特点, 因此高原灌丛通常较低矮、个体之间冠层不连续, 为典型的离散型灌丛.本研究中砂生槐灌丛属于干旱河谷灌丛, 其群落地上部分生物量(不含凋落物)介于1.4-6.9 Mg·hm-2 , 平均为3.53 Mg·hm-2 , 与青藏高原东缘的岷江干旱河谷灌丛地上生物量相当(1.4-4.8 Mg·hm-2 ; 刘国华等, 2003 ; 王勇军等, 2010 ), 略高于青海海北高寒金露梅灌丛(1.5-2.6 Mg·hm-2 ; 王启基等, 1991 ; 李英年等, 2006 ).目前有关高原灌丛根系生物量的报道很少, 本研究中砂生槐灌丛根系生物量在0.8-4.1 Mg·hm-2 之间, 平均为2.1 Mg·hm-2 , 高于青海海北高寒金露梅灌丛(1.5 Mg·hm-2 ; 李英年等, 2006 )和四川甘孜州高寒柳灌丛和杜鹃灌丛(1.3-1.8 Mg·hm-2 ; 高巧等, 2014 ), 这与砂生槐的深根系特点以及对干旱河谷环境的适应有关.砂生槐为典型深根系物种, 主根平均深入地下3 m, 侧根在距地表0 .7-1.5 m出现, 根幅大, 平均为1.9 × 2.3 m2 (彭跃明, 1997 ; 孙旭, 2010 ).从砂生槐器官生物量的分配来看, 根生物量占比超过一半, 使得平均根冠比达1.05, 从植被总体生物量的分配来看, 地下与地上生物量的关系斜率甚至达1.13 (图2 ), 即干旱地区植物把更多生物量分配到用于吸收水分和养分以及固定、支撑植物体的根系, 而不是地上部分的光合器官, 这些特点使砂生槐灌丛具有很强的防风固沙作用, 也体现了这一植被类型对河谷干旱环境的适应. ...
陆地生态系统碳密度格局研究概述
1
2004
... 作为生态系统最基本的特征指标, 生物量直接反映了生态系统生产力水平的高低(Keeling & Phillips, 2007 ), 同时作为衡量植被固碳能力的重要参数, 生物量的准确测定在碳循环与气候变化相关研究中具有重要意义(Dai et al ., 2013 ).碳密度是指单位土地面积的碳存储量, 是反映生态系统碳存储能力的重要指标之一, 作为估算生态系统碳储量和研究生态系统与大气碳交换的重要参数, 其准确估算是判定碳汇、制定缓解全球变化的合理政策措施的重要依据(吕超群和孙书存, 2004 ). ...
Shrub encroachment into mesic mountain grasslands in the Iberian peninsula: Effects of plant quality and temperature on soil C and N stocks
1
2007
... 灌丛通常是指高度在3 m以内、茎干具有多个分枝且主干不明显的木本植物.在以往研究中, 灌丛常被当作介于森林和草地之间的一个过渡类型, 并没有受到足够的重视.在五年一次的中国森林资源清查工作中, 灌丛(灌木林)也只有一个简单的面积信息① (① 森林资源管理司(2010). 全国森林资源统计——第七次全国森林资源清查.).目前有关生物量和碳密度方面的研究, 灌丛远远滞后于森林和草地(胡会峰等, 2006 ).随着全球气候变暖和人类活动的加强, 灌丛入侵草地、森林等问题越来越突出(Sturm et al ., 2001 ; Montane et al ., 2007 ), 草地向灌丛生态系统的演替加快, 进而增大灌丛分布面积, 这被认为是我国陆地生态系统碳储量增加的主要原因(王玲, 2009 ; 朴世龙等, 2010 ).然而灌丛植被的碳汇能力是目前我国陆地生态系统碳汇研究中最不确定的一个因素(Piao et al ., 2009 ), 因此加强灌丛群落生物量和碳密度相关研究, 对于准确估算我国陆地生态系统碳汇功能和固碳潜力具有重要意义. ...
西藏雅鲁藏布江中部流域砂生槐固沙作用研究
2
1997
... 在西藏, 灌丛分布相当普遍, 几乎除羌塘高原西北部以外都有分布, 其面积达13 × 106 hm2 , 已经超过森林, 成为西藏陆地生态系统中分布面积仅次于草地的一个重要类型(中国科学院中国植被图编辑委员会, 2001 ), 因此灌丛是西藏植被碳汇估算中不可或缺的组成部分.然而, 目前有关西藏灌丛生物量方面的研究鲜有报道(王启基等, 1991 ; 李英年等, 2006 ; 梁倍等, 2013 ), 涉及根系生物量的研究则更少(高巧等, 2014 ).西藏雅鲁藏布江(以下简称雅江)中游地区是西藏经济社会发展的核心区域, 而砂生槐(Sophora moorcroftiana )灌丛正是该区分布最广、最具代表性的灌丛类型, 其受人类活动的影响较大, 因此探索该区砂生槐灌丛群落的生物量现状及其分配规律对于指导当地生态建设具有现实意义.砂生槐灌丛主要分布在雅江流域中段的宽谷、两侧低山及拉萨河、年楚河等主要支流的宽谷内.作为典型的河谷灌丛, 砂生槐具有良好的防风固沙、涵养水源等重要生态作用(彭跃明, 1997 ).以往研究主要集中在砂生槐的种群和群落结构、繁殖特性、固沙特性和物种多样性等方面(赵文智, 1998 ; 郭其强等, 2009 ; 李海东等, 2011 ; 王文娟等, 2011 ; 杨兆平等, 2011 ; 赵垦田等, 2013 ), 尚未见到有关群落生物量和碳密度方面的报道.本研究采用相对生长法对西藏雅鲁藏布江流域中游的砂生槐灌丛进行地上和地下不同器官的生物量调查, 利用样方法测定了草本植物和凋落物的生物量, 并结合实测的全碳含量对灌丛植被碳密度进行了详尽计算, 以期为准确评价西藏灌丛生态系统碳收支和碳平衡提供基础数据, 为指导当地生态建设提供科学依据. ...
... 青藏高原由于海拔高、辐射强、温度低, 除了东南部较湿润, 高原大部分地区都具干旱半干旱气候特点, 因此高原灌丛通常较低矮、个体之间冠层不连续, 为典型的离散型灌丛.本研究中砂生槐灌丛属于干旱河谷灌丛, 其群落地上部分生物量(不含凋落物)介于1.4-6.9 Mg·hm-2 , 平均为3.53 Mg·hm-2 , 与青藏高原东缘的岷江干旱河谷灌丛地上生物量相当(1.4-4.8 Mg·hm-2 ; 刘国华等, 2003 ; 王勇军等, 2010 ), 略高于青海海北高寒金露梅灌丛(1.5-2.6 Mg·hm-2 ; 王启基等, 1991 ; 李英年等, 2006 ).目前有关高原灌丛根系生物量的报道很少, 本研究中砂生槐灌丛根系生物量在0.8-4.1 Mg·hm-2 之间, 平均为2.1 Mg·hm-2 , 高于青海海北高寒金露梅灌丛(1.5 Mg·hm-2 ; 李英年等, 2006 )和四川甘孜州高寒柳灌丛和杜鹃灌丛(1.3-1.8 Mg·hm-2 ; 高巧等, 2014 ), 这与砂生槐的深根系特点以及对干旱河谷环境的适应有关.砂生槐为典型深根系物种, 主根平均深入地下3 m, 侧根在距地表0 .7-1.5 m出现, 根幅大, 平均为1.9 × 2.3 m2 (彭跃明, 1997 ; 孙旭, 2010 ).从砂生槐器官生物量的分配来看, 根生物量占比超过一半, 使得平均根冠比达1.05, 从植被总体生物量的分配来看, 地下与地上生物量的关系斜率甚至达1.13 (图2 ), 即干旱地区植物把更多生物量分配到用于吸收水分和养分以及固定、支撑植物体的根系, 而不是地上部分的光合器官, 这些特点使砂生槐灌丛具有很强的防风固沙作用, 也体现了这一植被类型对河谷干旱环境的适应. ...
The carbon balance of terrestrial ecosystems in China
1
2009
... 灌丛通常是指高度在3 m以内、茎干具有多个分枝且主干不明显的木本植物.在以往研究中, 灌丛常被当作介于森林和草地之间的一个过渡类型, 并没有受到足够的重视.在五年一次的中国森林资源清查工作中, 灌丛(灌木林)也只有一个简单的面积信息① (① 森林资源管理司(2010). 全国森林资源统计——第七次全国森林资源清查.).目前有关生物量和碳密度方面的研究, 灌丛远远滞后于森林和草地(胡会峰等, 2006 ).随着全球气候变暖和人类活动的加强, 灌丛入侵草地、森林等问题越来越突出(Sturm et al ., 2001 ; Montane et al ., 2007 ), 草地向灌丛生态系统的演替加快, 进而增大灌丛分布面积, 这被认为是我国陆地生态系统碳储量增加的主要原因(王玲, 2009 ; 朴世龙等, 2010 ).然而灌丛植被的碳汇能力是目前我国陆地生态系统碳汇研究中最不确定的一个因素(Piao et al ., 2009 ), 因此加强灌丛群落生物量和碳密度相关研究, 对于准确估算我国陆地生态系统碳汇功能和固碳潜力具有重要意义. ...
中国陆地生态系统碳收支
1
2010
... 灌丛通常是指高度在3 m以内、茎干具有多个分枝且主干不明显的木本植物.在以往研究中, 灌丛常被当作介于森林和草地之间的一个过渡类型, 并没有受到足够的重视.在五年一次的中国森林资源清查工作中, 灌丛(灌木林)也只有一个简单的面积信息① (① 森林资源管理司(2010). 全国森林资源统计——第七次全国森林资源清查.).目前有关生物量和碳密度方面的研究, 灌丛远远滞后于森林和草地(胡会峰等, 2006 ).随着全球气候变暖和人类活动的加强, 灌丛入侵草地、森林等问题越来越突出(Sturm et al ., 2001 ; Montane et al ., 2007 ), 草地向灌丛生态系统的演替加快, 进而增大灌丛分布面积, 这被认为是我国陆地生态系统碳储量增加的主要原因(王玲, 2009 ; 朴世龙等, 2010 ).然而灌丛植被的碳汇能力是目前我国陆地生态系统碳汇研究中最不确定的一个因素(Piao et al ., 2009 ), 因此加强灌丛群落生物量和碳密度相关研究, 对于准确估算我国陆地生态系统碳汇功能和固碳潜力具有重要意义. ...
1
2001
... 目前有关碳密度的研究大部分采用平均碳密度方法(胡会峰等, 2006 ; 付达夫, 2014 ; 李亚男等, 2014 ), 即碳密度等于生物量乘以换算系数0.5或0.45.然而不同物种、不同器官之间的碳含量是存在差异的, 且碳含量通常达不到50% (宋永昌, 2001 ), 用平均碳密度法势必会高估生态系统碳密度.本研究通过实测不同器官全碳含量, 结合植被生物量计算了灌丛植被碳密度, 与利用换算系数的估算方法相比, 也发现以0.5为换算系数的估算结果比实际结果偏高, 总体高约15%, 其中灌木层偏高6%-11%, 草本层偏高36%-44% (表5 ).以0.45作为换算系数的总体估算结果与实际结果较接近(偏高3.6%), 表现为灌木层略微偏低(0-4%), 草本层偏高(20%-30%)(表5 ).综上所述, 在区域尺度上估算西藏雅江流域砂生槐灌丛群落碳密度时, 可以考虑用0.45作为碳含量的换算系数. ...
Increasing shrub abundance in the Arctic
1
2001
... 灌丛通常是指高度在3 m以内、茎干具有多个分枝且主干不明显的木本植物.在以往研究中, 灌丛常被当作介于森林和草地之间的一个过渡类型, 并没有受到足够的重视.在五年一次的中国森林资源清查工作中, 灌丛(灌木林)也只有一个简单的面积信息① (① 森林资源管理司(2010). 全国森林资源统计——第七次全国森林资源清查.).目前有关生物量和碳密度方面的研究, 灌丛远远滞后于森林和草地(胡会峰等, 2006 ).随着全球气候变暖和人类活动的加强, 灌丛入侵草地、森林等问题越来越突出(Sturm et al ., 2001 ; Montane et al ., 2007 ), 草地向灌丛生态系统的演替加快, 进而增大灌丛分布面积, 这被认为是我国陆地生态系统碳储量增加的主要原因(王玲, 2009 ; 朴世龙等, 2010 ).然而灌丛植被的碳汇能力是目前我国陆地生态系统碳汇研究中最不确定的一个因素(Piao et al ., 2009 ), 因此加强灌丛群落生物量和碳密度相关研究, 对于准确估算我国陆地生态系统碳汇功能和固碳潜力具有重要意义. ...
西藏砂生槐群落结构与生物量研究
1
2010
... 青藏高原由于海拔高、辐射强、温度低, 除了东南部较湿润, 高原大部分地区都具干旱半干旱气候特点, 因此高原灌丛通常较低矮、个体之间冠层不连续, 为典型的离散型灌丛.本研究中砂生槐灌丛属于干旱河谷灌丛, 其群落地上部分生物量(不含凋落物)介于1.4-6.9 Mg·hm-2 , 平均为3.53 Mg·hm-2 , 与青藏高原东缘的岷江干旱河谷灌丛地上生物量相当(1.4-4.8 Mg·hm-2 ; 刘国华等, 2003 ; 王勇军等, 2010 ), 略高于青海海北高寒金露梅灌丛(1.5-2.6 Mg·hm-2 ; 王启基等, 1991 ; 李英年等, 2006 ).目前有关高原灌丛根系生物量的报道很少, 本研究中砂生槐灌丛根系生物量在0.8-4.1 Mg·hm-2 之间, 平均为2.1 Mg·hm-2 , 高于青海海北高寒金露梅灌丛(1.5 Mg·hm-2 ; 李英年等, 2006 )和四川甘孜州高寒柳灌丛和杜鹃灌丛(1.3-1.8 Mg·hm-2 ; 高巧等, 2014 ), 这与砂生槐的深根系特点以及对干旱河谷环境的适应有关.砂生槐为典型深根系物种, 主根平均深入地下3 m, 侧根在距地表0 .7-1.5 m出现, 根幅大, 平均为1.9 × 2.3 m2 (彭跃明, 1997 ; 孙旭, 2010 ).从砂生槐器官生物量的分配来看, 根生物量占比超过一半, 使得平均根冠比达1.05, 从植被总体生物量的分配来看, 地下与地上生物量的关系斜率甚至达1.13 (图2 ), 即干旱地区植物把更多生物量分配到用于吸收水分和养分以及固定、支撑植物体的根系, 而不是地上部分的光合器官, 这些特点使砂生槐灌丛具有很强的防风固沙作用, 也体现了这一植被类型对河谷干旱环境的适应. ...
2
2001
... 在西藏, 灌丛分布相当普遍, 几乎除羌塘高原西北部以外都有分布, 其面积达13 × 106 hm2 , 已经超过森林, 成为西藏陆地生态系统中分布面积仅次于草地的一个重要类型(中国科学院中国植被图编辑委员会, 2001 ), 因此灌丛是西藏植被碳汇估算中不可或缺的组成部分.然而, 目前有关西藏灌丛生物量方面的研究鲜有报道(王启基等, 1991 ; 李英年等, 2006 ; 梁倍等, 2013 ), 涉及根系生物量的研究则更少(高巧等, 2014 ).西藏雅鲁藏布江(以下简称雅江)中游地区是西藏经济社会发展的核心区域, 而砂生槐(Sophora moorcroftiana )灌丛正是该区分布最广、最具代表性的灌丛类型, 其受人类活动的影响较大, 因此探索该区砂生槐灌丛群落的生物量现状及其分配规律对于指导当地生态建设具有现实意义.砂生槐灌丛主要分布在雅江流域中段的宽谷、两侧低山及拉萨河、年楚河等主要支流的宽谷内.作为典型的河谷灌丛, 砂生槐具有良好的防风固沙、涵养水源等重要生态作用(彭跃明, 1997 ).以往研究主要集中在砂生槐的种群和群落结构、繁殖特性、固沙特性和物种多样性等方面(赵文智, 1998 ; 郭其强等, 2009 ; 李海东等, 2011 ; 王文娟等, 2011 ; 杨兆平等, 2011 ; 赵垦田等, 2013 ), 尚未见到有关群落生物量和碳密度方面的报道.本研究采用相对生长法对西藏雅鲁藏布江流域中游的砂生槐灌丛进行地上和地下不同器官的生物量调查, 利用样方法测定了草本植物和凋落物的生物量, 并结合实测的全碳含量对灌丛植被碳密度进行了详尽计算, 以期为准确评价西藏灌丛生态系统碳收支和碳平衡提供基础数据, 为指导当地生态建设提供科学依据. ...
... 基于《1:1000000中国植被图集》(中国科学院中国植被图编辑委员会, 2001 ), 我们于2011-2013年对该区砂生槐灌丛进行了实地调查.考虑到交通的可达性, 我们主要在公路沿线开展工作, 选取典型的砂生槐分布区进行调查, 样点的选择标准包括: 1)群落总分布面积>5 hm2 , 2)距公路200 m以上, 3)原则上两样点间距离>30 km.最后选择了18个砂生槐灌丛调查样点(图1 ). ...
川西北地区主要灌丛类型生物量及其模型研究
1
2009
... 灌丛通常是指高度在3 m以内、茎干具有多个分枝且主干不明显的木本植物.在以往研究中, 灌丛常被当作介于森林和草地之间的一个过渡类型, 并没有受到足够的重视.在五年一次的中国森林资源清查工作中, 灌丛(灌木林)也只有一个简单的面积信息① (① 森林资源管理司(2010). 全国森林资源统计——第七次全国森林资源清查.).目前有关生物量和碳密度方面的研究, 灌丛远远滞后于森林和草地(胡会峰等, 2006 ).随着全球气候变暖和人类活动的加强, 灌丛入侵草地、森林等问题越来越突出(Sturm et al ., 2001 ; Montane et al ., 2007 ), 草地向灌丛生态系统的演替加快, 进而增大灌丛分布面积, 这被认为是我国陆地生态系统碳储量增加的主要原因(王玲, 2009 ; 朴世龙等, 2010 ).然而灌丛植被的碳汇能力是目前我国陆地生态系统碳汇研究中最不确定的一个因素(Piao et al ., 2009 ), 因此加强灌丛群落生物量和碳密度相关研究, 对于准确估算我国陆地生态系统碳汇功能和固碳潜力具有重要意义. ...
青藏高原金露梅灌丛的结构特征及其生物量
2
1991
... 在西藏, 灌丛分布相当普遍, 几乎除羌塘高原西北部以外都有分布, 其面积达13 × 106 hm2 , 已经超过森林, 成为西藏陆地生态系统中分布面积仅次于草地的一个重要类型(中国科学院中国植被图编辑委员会, 2001 ), 因此灌丛是西藏植被碳汇估算中不可或缺的组成部分.然而, 目前有关西藏灌丛生物量方面的研究鲜有报道(王启基等, 1991 ; 李英年等, 2006 ; 梁倍等, 2013 ), 涉及根系生物量的研究则更少(高巧等, 2014 ).西藏雅鲁藏布江(以下简称雅江)中游地区是西藏经济社会发展的核心区域, 而砂生槐(Sophora moorcroftiana )灌丛正是该区分布最广、最具代表性的灌丛类型, 其受人类活动的影响较大, 因此探索该区砂生槐灌丛群落的生物量现状及其分配规律对于指导当地生态建设具有现实意义.砂生槐灌丛主要分布在雅江流域中段的宽谷、两侧低山及拉萨河、年楚河等主要支流的宽谷内.作为典型的河谷灌丛, 砂生槐具有良好的防风固沙、涵养水源等重要生态作用(彭跃明, 1997 ).以往研究主要集中在砂生槐的种群和群落结构、繁殖特性、固沙特性和物种多样性等方面(赵文智, 1998 ; 郭其强等, 2009 ; 李海东等, 2011 ; 王文娟等, 2011 ; 杨兆平等, 2011 ; 赵垦田等, 2013 ), 尚未见到有关群落生物量和碳密度方面的报道.本研究采用相对生长法对西藏雅鲁藏布江流域中游的砂生槐灌丛进行地上和地下不同器官的生物量调查, 利用样方法测定了草本植物和凋落物的生物量, 并结合实测的全碳含量对灌丛植被碳密度进行了详尽计算, 以期为准确评价西藏灌丛生态系统碳收支和碳平衡提供基础数据, 为指导当地生态建设提供科学依据. ...
... 青藏高原由于海拔高、辐射强、温度低, 除了东南部较湿润, 高原大部分地区都具干旱半干旱气候特点, 因此高原灌丛通常较低矮、个体之间冠层不连续, 为典型的离散型灌丛.本研究中砂生槐灌丛属于干旱河谷灌丛, 其群落地上部分生物量(不含凋落物)介于1.4-6.9 Mg·hm-2 , 平均为3.53 Mg·hm-2 , 与青藏高原东缘的岷江干旱河谷灌丛地上生物量相当(1.4-4.8 Mg·hm-2 ; 刘国华等, 2003 ; 王勇军等, 2010 ), 略高于青海海北高寒金露梅灌丛(1.5-2.6 Mg·hm-2 ; 王启基等, 1991 ; 李英年等, 2006 ).目前有关高原灌丛根系生物量的报道很少, 本研究中砂生槐灌丛根系生物量在0.8-4.1 Mg·hm-2 之间, 平均为2.1 Mg·hm-2 , 高于青海海北高寒金露梅灌丛(1.5 Mg·hm-2 ; 李英年等, 2006 )和四川甘孜州高寒柳灌丛和杜鹃灌丛(1.3-1.8 Mg·hm-2 ; 高巧等, 2014 ), 这与砂生槐的深根系特点以及对干旱河谷环境的适应有关.砂生槐为典型深根系物种, 主根平均深入地下3 m, 侧根在距地表0 .7-1.5 m出现, 根幅大, 平均为1.9 × 2.3 m2 (彭跃明, 1997 ; 孙旭, 2010 ).从砂生槐器官生物量的分配来看, 根生物量占比超过一半, 使得平均根冠比达1.05, 从植被总体生物量的分配来看, 地下与地上生物量的关系斜率甚至达1.13 (图2 ), 即干旱地区植物把更多生物量分配到用于吸收水分和养分以及固定、支撑植物体的根系, 而不是地上部分的光合器官, 这些特点使砂生槐灌丛具有很强的防风固沙作用, 也体现了这一植被类型对河谷干旱环境的适应. ...
不同温度和沙埋深度对砂生槐种子萌发及幼苗生长的影响
1
2011
... 在西藏, 灌丛分布相当普遍, 几乎除羌塘高原西北部以外都有分布, 其面积达13 × 106 hm2 , 已经超过森林, 成为西藏陆地生态系统中分布面积仅次于草地的一个重要类型(中国科学院中国植被图编辑委员会, 2001 ), 因此灌丛是西藏植被碳汇估算中不可或缺的组成部分.然而, 目前有关西藏灌丛生物量方面的研究鲜有报道(王启基等, 1991 ; 李英年等, 2006 ; 梁倍等, 2013 ), 涉及根系生物量的研究则更少(高巧等, 2014 ).西藏雅鲁藏布江(以下简称雅江)中游地区是西藏经济社会发展的核心区域, 而砂生槐(Sophora moorcroftiana )灌丛正是该区分布最广、最具代表性的灌丛类型, 其受人类活动的影响较大, 因此探索该区砂生槐灌丛群落的生物量现状及其分配规律对于指导当地生态建设具有现实意义.砂生槐灌丛主要分布在雅江流域中段的宽谷、两侧低山及拉萨河、年楚河等主要支流的宽谷内.作为典型的河谷灌丛, 砂生槐具有良好的防风固沙、涵养水源等重要生态作用(彭跃明, 1997 ).以往研究主要集中在砂生槐的种群和群落结构、繁殖特性、固沙特性和物种多样性等方面(赵文智, 1998 ; 郭其强等, 2009 ; 李海东等, 2011 ; 王文娟等, 2011 ; 杨兆平等, 2011 ; 赵垦田等, 2013 ), 尚未见到有关群落生物量和碳密度方面的报道.本研究采用相对生长法对西藏雅鲁藏布江流域中游的砂生槐灌丛进行地上和地下不同器官的生物量调查, 利用样方法测定了草本植物和凋落物的生物量, 并结合实测的全碳含量对灌丛植被碳密度进行了详尽计算, 以期为准确评价西藏灌丛生态系统碳收支和碳平衡提供基础数据, 为指导当地生态建设提供科学依据. ...
岷江干旱河谷灌丛物种多样性、生物量及其关系
1
2010
... 青藏高原由于海拔高、辐射强、温度低, 除了东南部较湿润, 高原大部分地区都具干旱半干旱气候特点, 因此高原灌丛通常较低矮、个体之间冠层不连续, 为典型的离散型灌丛.本研究中砂生槐灌丛属于干旱河谷灌丛, 其群落地上部分生物量(不含凋落物)介于1.4-6.9 Mg·hm-2 , 平均为3.53 Mg·hm-2 , 与青藏高原东缘的岷江干旱河谷灌丛地上生物量相当(1.4-4.8 Mg·hm-2 ; 刘国华等, 2003 ; 王勇军等, 2010 ), 略高于青海海北高寒金露梅灌丛(1.5-2.6 Mg·hm-2 ; 王启基等, 1991 ; 李英年等, 2006 ).目前有关高原灌丛根系生物量的报道很少, 本研究中砂生槐灌丛根系生物量在0.8-4.1 Mg·hm-2 之间, 平均为2.1 Mg·hm-2 , 高于青海海北高寒金露梅灌丛(1.5 Mg·hm-2 ; 李英年等, 2006 )和四川甘孜州高寒柳灌丛和杜鹃灌丛(1.3-1.8 Mg·hm-2 ; 高巧等, 2014 ), 这与砂生槐的深根系特点以及对干旱河谷环境的适应有关.砂生槐为典型深根系物种, 主根平均深入地下3 m, 侧根在距地表0 .7-1.5 m出现, 根幅大, 平均为1.9 × 2.3 m2 (彭跃明, 1997 ; 孙旭, 2010 ).从砂生槐器官生物量的分配来看, 根生物量占比超过一半, 使得平均根冠比达1.05, 从植被总体生物量的分配来看, 地下与地上生物量的关系斜率甚至达1.13 (图2 ), 即干旱地区植物把更多生物量分配到用于吸收水分和养分以及固定、支撑植物体的根系, 而不是地上部分的光合器官, 这些特点使砂生槐灌丛具有很强的防风固沙作用, 也体现了这一植被类型对河谷干旱环境的适应. ...
青藏高原近30年气候变化趋势,
1
2005
... 本研究中砂生槐灌丛植被平均碳密度为2.48 Mg·hm-2 , 与付达夫(2014)针对本研究物种灌丛碳密度估算结果(约2.5 Mg·hm-2 )一致, 但远低于胡 会峰等(2006)估算的中国主要灌丛植被平均碳密度(10.88 Mg·hm-2 )及付达夫(2014)估算的西藏灌木林碳密度(8.53 Mg·hm-2 ), 也低于四川甘孜州高寒矮灌丛(3.20 Mg·hm-2 ), 这与砂生槐灌丛所处生境以及群落盖度较低有关——在干旱的河谷地带, 为减少对水分和(或)养分资源的竞争, 砂生槐种群具有较强的自疏作用, 个体间距较大, 导致植被冠层不连续, 灌木层盖度在12%-53%之间, 平均仅为30.5%, 明显低于四川甘孜州高寒矮灌丛(22%-85%, 高巧等, 2014 ).砂生槐灌丛群落碳密度在研究区东部与西部存在显著差异, 这是由于东、西部灌丛盖度存在差异所致, 即西部地区植被盖度明显高于东部(表4 ), 究其原因, 主要与东、西部的自然环境(尤其是海拔)以及人为干扰强度的差异有关.研究区东部平均海拔较西部低200-300 m, 因此平均气温高1-2 ℃, 而年降水量略低或相近, 导致东部地区蒸发量相对较高, 因此焚风效应(过山气流在背风坡下沉而变得干热)形成的干旱河谷气候在研究区东部比在西部更明显, 这与杨志刚等(2013)利用Penman-Monteith模型计算的雅江流域潜在蒸散量结果一致, 即潜在蒸散量在位于雅江中游东部的泽当明显高于西部的日喀则(杨志刚等, 2013 ).显然, 东部地区较高的蒸散量加剧了河谷的干旱化程度, 进一步限制了该区砂生槐灌丛植被盖度的扩张.因此, 深入探究不同器官生物量分配与气候、土壤因子的关系, 将有助于揭示砂生槐灌丛的地理分异机制.此外, 研究区东部地区人口密度更大, 人类活动和放牧干扰强度较大, 牲畜践踏和啃食对砂生槐灌丛的生长和繁殖均具有不可忽视的作用, 这可能也是导致研究区东部植被盖度较低的原因之一.在未来气候变化背景下, 如果蒸散持续降低(吴绍洪等, 2005 ; Yin et al ., 2013 ), 将有助于砂生槐灌丛生物量和碳密度的增加, 这无疑对于雅江流域水土保持和防风固沙等生态工程具有重要指导意义. ...
近50年雅鲁藏布江流域潜在蒸散量的变化特征
1
2013
... 本研究中砂生槐灌丛植被平均碳密度为2.48 Mg·hm-2 , 与付达夫(2014)针对本研究物种灌丛碳密度估算结果(约2.5 Mg·hm-2 )一致, 但远低于胡 会峰等(2006)估算的中国主要灌丛植被平均碳密度(10.88 Mg·hm-2 )及付达夫(2014)估算的西藏灌木林碳密度(8.53 Mg·hm-2 ), 也低于四川甘孜州高寒矮灌丛(3.20 Mg·hm-2 ), 这与砂生槐灌丛所处生境以及群落盖度较低有关——在干旱的河谷地带, 为减少对水分和(或)养分资源的竞争, 砂生槐种群具有较强的自疏作用, 个体间距较大, 导致植被冠层不连续, 灌木层盖度在12%-53%之间, 平均仅为30.5%, 明显低于四川甘孜州高寒矮灌丛(22%-85%, 高巧等, 2014 ).砂生槐灌丛群落碳密度在研究区东部与西部存在显著差异, 这是由于东、西部灌丛盖度存在差异所致, 即西部地区植被盖度明显高于东部(表4 ), 究其原因, 主要与东、西部的自然环境(尤其是海拔)以及人为干扰强度的差异有关.研究区东部平均海拔较西部低200-300 m, 因此平均气温高1-2 ℃, 而年降水量略低或相近, 导致东部地区蒸发量相对较高, 因此焚风效应(过山气流在背风坡下沉而变得干热)形成的干旱河谷气候在研究区东部比在西部更明显, 这与杨志刚等(2013)利用Penman-Monteith模型计算的雅江流域潜在蒸散量结果一致, 即潜在蒸散量在位于雅江中游东部的泽当明显高于西部的日喀则(杨志刚等, 2013 ).显然, 东部地区较高的蒸散量加剧了河谷的干旱化程度, 进一步限制了该区砂生槐灌丛植被盖度的扩张.因此, 深入探究不同器官生物量分配与气候、土壤因子的关系, 将有助于揭示砂生槐灌丛的地理分异机制.此外, 研究区东部地区人口密度更大, 人类活动和放牧干扰强度较大, 牲畜践踏和啃食对砂生槐灌丛的生长和繁殖均具有不可忽视的作用, 这可能也是导致研究区东部植被盖度较低的原因之一.在未来气候变化背景下, 如果蒸散持续降低(吴绍洪等, 2005 ; Yin et al ., 2013 ), 将有助于砂生槐灌丛生物量和碳密度的增加, 这无疑对于雅江流域水土保持和防风固沙等生态工程具有重要指导意义. ...
雅鲁藏布江中游河谷风沙化土地砂生槐群落结构特征
1
2011
... 在西藏, 灌丛分布相当普遍, 几乎除羌塘高原西北部以外都有分布, 其面积达13 × 106 hm2 , 已经超过森林, 成为西藏陆地生态系统中分布面积仅次于草地的一个重要类型(中国科学院中国植被图编辑委员会, 2001 ), 因此灌丛是西藏植被碳汇估算中不可或缺的组成部分.然而, 目前有关西藏灌丛生物量方面的研究鲜有报道(王启基等, 1991 ; 李英年等, 2006 ; 梁倍等, 2013 ), 涉及根系生物量的研究则更少(高巧等, 2014 ).西藏雅鲁藏布江(以下简称雅江)中游地区是西藏经济社会发展的核心区域, 而砂生槐(Sophora moorcroftiana )灌丛正是该区分布最广、最具代表性的灌丛类型, 其受人类活动的影响较大, 因此探索该区砂生槐灌丛群落的生物量现状及其分配规律对于指导当地生态建设具有现实意义.砂生槐灌丛主要分布在雅江流域中段的宽谷、两侧低山及拉萨河、年楚河等主要支流的宽谷内.作为典型的河谷灌丛, 砂生槐具有良好的防风固沙、涵养水源等重要生态作用(彭跃明, 1997 ).以往研究主要集中在砂生槐的种群和群落结构、繁殖特性、固沙特性和物种多样性等方面(赵文智, 1998 ; 郭其强等, 2009 ; 李海东等, 2011 ; 王文娟等, 2011 ; 杨兆平等, 2011 ; 赵垦田等, 2013 ), 尚未见到有关群落生物量和碳密度方面的报道.本研究采用相对生长法对西藏雅鲁藏布江流域中游的砂生槐灌丛进行地上和地下不同器官的生物量调查, 利用样方法测定了草本植物和凋落物的生物量, 并结合实测的全碳含量对灌丛植被碳密度进行了详尽计算, 以期为准确评价西藏灌丛生态系统碳收支和碳平衡提供基础数据, 为指导当地生态建设提供科学依据. ...
Past and future spatiotemporal changes in evapotranspiration and effective moisture on the Tibetan Plateau
1
2013
... 本研究中砂生槐灌丛植被平均碳密度为2.48 Mg·hm-2 , 与付达夫(2014)针对本研究物种灌丛碳密度估算结果(约2.5 Mg·hm-2 )一致, 但远低于胡 会峰等(2006)估算的中国主要灌丛植被平均碳密度(10.88 Mg·hm-2 )及付达夫(2014)估算的西藏灌木林碳密度(8.53 Mg·hm-2 ), 也低于四川甘孜州高寒矮灌丛(3.20 Mg·hm-2 ), 这与砂生槐灌丛所处生境以及群落盖度较低有关——在干旱的河谷地带, 为减少对水分和(或)养分资源的竞争, 砂生槐种群具有较强的自疏作用, 个体间距较大, 导致植被冠层不连续, 灌木层盖度在12%-53%之间, 平均仅为30.5%, 明显低于四川甘孜州高寒矮灌丛(22%-85%, 高巧等, 2014 ).砂生槐灌丛群落碳密度在研究区东部与西部存在显著差异, 这是由于东、西部灌丛盖度存在差异所致, 即西部地区植被盖度明显高于东部(表4 ), 究其原因, 主要与东、西部的自然环境(尤其是海拔)以及人为干扰强度的差异有关.研究区东部平均海拔较西部低200-300 m, 因此平均气温高1-2 ℃, 而年降水量略低或相近, 导致东部地区蒸发量相对较高, 因此焚风效应(过山气流在背风坡下沉而变得干热)形成的干旱河谷气候在研究区东部比在西部更明显, 这与杨志刚等(2013)利用Penman-Monteith模型计算的雅江流域潜在蒸散量结果一致, 即潜在蒸散量在位于雅江中游东部的泽当明显高于西部的日喀则(杨志刚等, 2013 ).显然, 东部地区较高的蒸散量加剧了河谷的干旱化程度, 进一步限制了该区砂生槐灌丛植被盖度的扩张.因此, 深入探究不同器官生物量分配与气候、土壤因子的关系, 将有助于揭示砂生槐灌丛的地理分异机制.此外, 研究区东部地区人口密度更大, 人类活动和放牧干扰强度较大, 牲畜践踏和啃食对砂生槐灌丛的生长和繁殖均具有不可忽视的作用, 这可能也是导致研究区东部植被盖度较低的原因之一.在未来气候变化背景下, 如果蒸散持续降低(吴绍洪等, 2005 ; Yin et al ., 2013 ), 将有助于砂生槐灌丛生物量和碳密度的增加, 这无疑对于雅江流域水土保持和防风固沙等生态工程具有重要指导意义. ...
拉萨半干旱河谷砂生槐灌丛生态恢复过程的群落特征与土壤微生物动态分析
1
2013
... 在西藏, 灌丛分布相当普遍, 几乎除羌塘高原西北部以外都有分布, 其面积达13 × 106 hm2 , 已经超过森林, 成为西藏陆地生态系统中分布面积仅次于草地的一个重要类型(中国科学院中国植被图编辑委员会, 2001 ), 因此灌丛是西藏植被碳汇估算中不可或缺的组成部分.然而, 目前有关西藏灌丛生物量方面的研究鲜有报道(王启基等, 1991 ; 李英年等, 2006 ; 梁倍等, 2013 ), 涉及根系生物量的研究则更少(高巧等, 2014 ).西藏雅鲁藏布江(以下简称雅江)中游地区是西藏经济社会发展的核心区域, 而砂生槐(Sophora moorcroftiana )灌丛正是该区分布最广、最具代表性的灌丛类型, 其受人类活动的影响较大, 因此探索该区砂生槐灌丛群落的生物量现状及其分配规律对于指导当地生态建设具有现实意义.砂生槐灌丛主要分布在雅江流域中段的宽谷、两侧低山及拉萨河、年楚河等主要支流的宽谷内.作为典型的河谷灌丛, 砂生槐具有良好的防风固沙、涵养水源等重要生态作用(彭跃明, 1997 ).以往研究主要集中在砂生槐的种群和群落结构、繁殖特性、固沙特性和物种多样性等方面(赵文智, 1998 ; 郭其强等, 2009 ; 李海东等, 2011 ; 王文娟等, 2011 ; 杨兆平等, 2011 ; 赵垦田等, 2013 ), 尚未见到有关群落生物量和碳密度方面的报道.本研究采用相对生长法对西藏雅鲁藏布江流域中游的砂生槐灌丛进行地上和地下不同器官的生物量调查, 利用样方法测定了草本植物和凋落物的生物量, 并结合实测的全碳含量对灌丛植被碳密度进行了详尽计算, 以期为准确评价西藏灌丛生态系统碳收支和碳平衡提供基础数据, 为指导当地生态建设提供科学依据. ...
砂生槐沙生适应性初步研究
1
1998
... 在西藏, 灌丛分布相当普遍, 几乎除羌塘高原西北部以外都有分布, 其面积达13 × 106 hm2 , 已经超过森林, 成为西藏陆地生态系统中分布面积仅次于草地的一个重要类型(中国科学院中国植被图编辑委员会, 2001 ), 因此灌丛是西藏植被碳汇估算中不可或缺的组成部分.然而, 目前有关西藏灌丛生物量方面的研究鲜有报道(王启基等, 1991 ; 李英年等, 2006 ; 梁倍等, 2013 ), 涉及根系生物量的研究则更少(高巧等, 2014 ).西藏雅鲁藏布江(以下简称雅江)中游地区是西藏经济社会发展的核心区域, 而砂生槐(Sophora moorcroftiana )灌丛正是该区分布最广、最具代表性的灌丛类型, 其受人类活动的影响较大, 因此探索该区砂生槐灌丛群落的生物量现状及其分配规律对于指导当地生态建设具有现实意义.砂生槐灌丛主要分布在雅江流域中段的宽谷、两侧低山及拉萨河、年楚河等主要支流的宽谷内.作为典型的河谷灌丛, 砂生槐具有良好的防风固沙、涵养水源等重要生态作用(彭跃明, 1997 ).以往研究主要集中在砂生槐的种群和群落结构、繁殖特性、固沙特性和物种多样性等方面(赵文智, 1998 ; 郭其强等, 2009 ; 李海东等, 2011 ; 王文娟等, 2011 ; 杨兆平等, 2011 ; 赵垦田等, 2013 ), 尚未见到有关群落生物量和碳密度方面的报道.本研究采用相对生长法对西藏雅鲁藏布江流域中游的砂生槐灌丛进行地上和地下不同器官的生物量调查, 利用样方法测定了草本植物和凋落物的生物量, 并结合实测的全碳含量对灌丛植被碳密度进行了详尽计算, 以期为准确评价西藏灌丛生态系统碳收支和碳平衡提供基础数据, 为指导当地生态建设提供科学依据. ...
 , 沈维
, 沈维

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}