Population structure and spatial pattern of Caragana tibetica communities in Nei Mongol shrub-encroached grassland
ZHANG Pu-Jin1, 2, 3, QING Hua4, ZHANG Lei5, XU Yan-Da6, MU Lan1, 2, YE Ru-Han1, 2, QIU Xiao1, 2, Chang Hong1, 2, SHEN Hai-Hua3, YANG Jie4, *,
1Inner Mongolia Academy of Agricultural and Animal Husbandry Sciences, Hohhot 010031, China2 Inner Mongolia Prataculture Research Center, Chinese Academy of Sciences, Hohhot 010031, China3State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China4School of Ecology and Environment, Inner Mongolia University, Hohhot 010021, China5Inner Mongolia Academy of Forestry Sciences, Hohhot 010010, China6State Environmental Protection Key Laboratory of Regional Ecological Processes and Functions Assessment, Chinese Research Academy of Environmental Sciences, Beijing 100012
Aims The community structure and function of the shrub-encroached grassland, as well as its formation mechanism, is one of hot scientific problems in vegetation science. The spatial pattern analysis is an important means for studying plant population interactions and the relationships between plant population and environment. The shrub-encroached grassland is one major vegetation type in Nei Mongol. Knowledge on the spatial pattern of shrub can improve our understanding on adaptive characteristics of shrub to steppe environment. Methods We chose Caragana tibetica encroached grassland distributed in Ordos City as research objects. The spatial position of each C. tibetica individual in 64 m × 64 m study plot was determined by a method of “three-distance location method”, and then the spatial pattern and intraspecific associations of C. tibetica individuals was analyzed with the point pattern analysis. Important findings The results showed that C. tibetica population was at the rising stage. The mature individuals showed a uniform distribution at the scale of less than 3.7 m, subsequently exhibiting a random distribution with an increased scale. The juvenile individuals showed an aggregation distribution at the scale of 0.3-20.4 m, but a uniform distribution at other scales. The dead C. tibetica individuals were distributed aggregately at the scale from 0.7 to 1.4 m, but randomly at other scales. An indistinctive positive association was found for live C. tibetica individuals to dead individuals at the scale of less than 9.9 m, but no associations was found between them at other scales. The mature individuals negatively interacted with juveniles at the scale of larger than 2.2 m, but unassociated at the other scales.
Keywords:shrub-encroached grassland
;
Caragana tibetica
;
point pattern
;
spatial distribution pattern
ZHANGPu-Jin, QINGHua, ZHANGLei, XUYan-Da, MULan, YERu-Han, QIUXiao, ChangHong, SHENHai-Hua, YANGJie. Population structure and spatial pattern of Caragana tibetica communities in Nei Mongol shrub-encroached grassland. Chinese Journal of Plant Ecology, 2017, 41(2): 165-174 https://doi.org/10.17521/cjpe.2015.0448
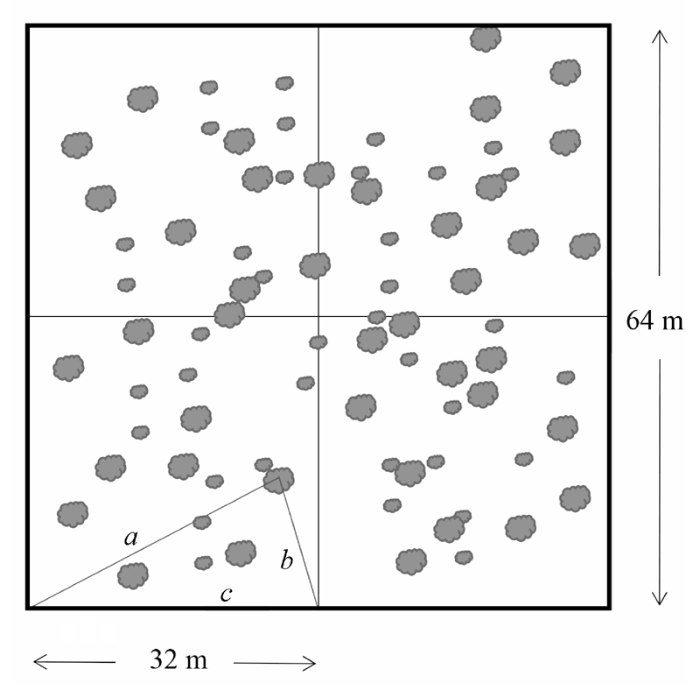
Fig. 1 Schematic view of “three-distance location method”. a and b are the distances of canopy center to each two fixed points, c is the distance of the two fixed points, patches are schematic maps of the shrubs canopy.
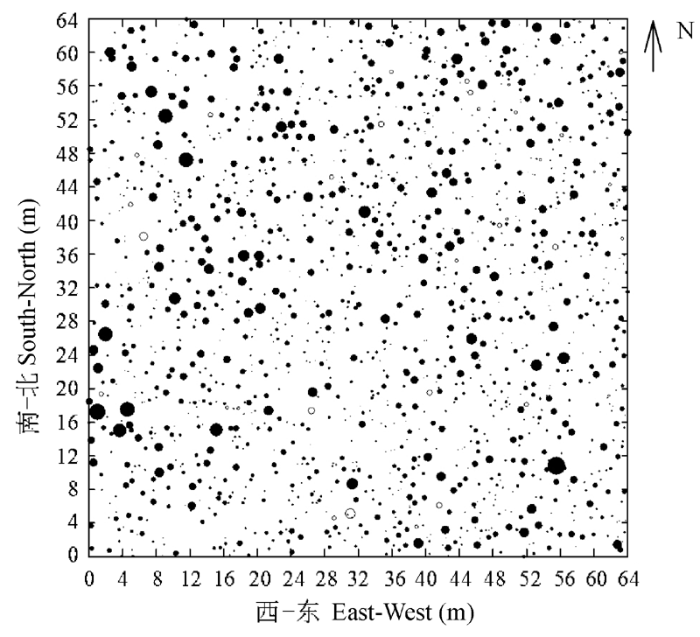
Fig. 2 The distribution of Caragana tibetica in 64 m × 64 m plot. Each circle gives the location of living (filled circle) and dead (empty circle) C. tibetica and circle area is proportional to canopy of C. tibetica.
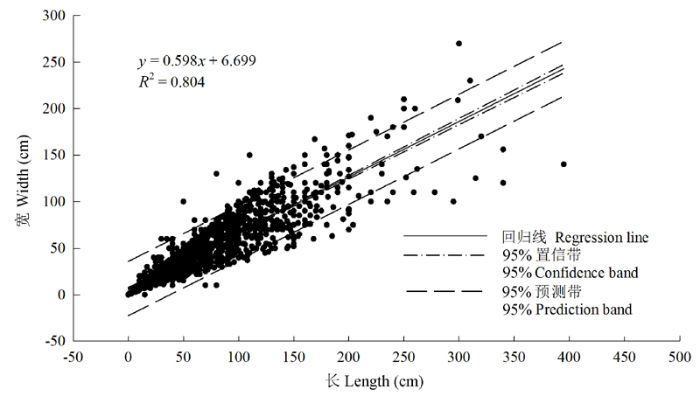
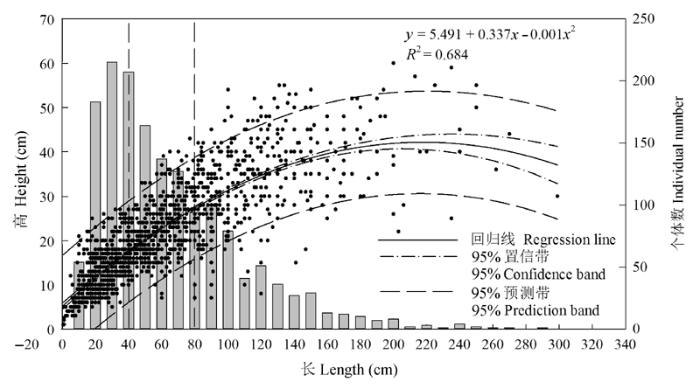
Fig. 4 Relationship (filled circle) between the length and height of Caragana tibetica canopy, and individual number (histogram) in each range of canopy length with a step size of 10 cm, eg. 10 cm indicated a length range of 0-10 cm, the rest can be done in the same manner.
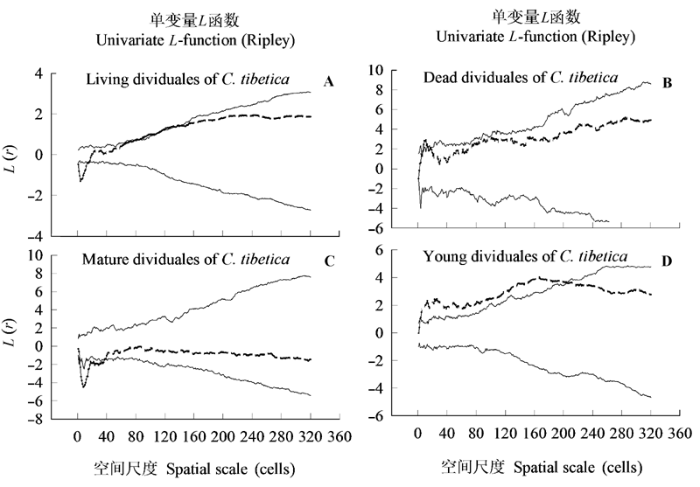
Fig. 5 Ripley’s L-function for patterns of Caragana tibetica individuals under different survival status and developmental stages with confidence envelopes (lines). r referred to radius of sample circle from C. tibetica as center, Confidence envelopes are the highest and lowest L(r) of 999 randomizations of the pattern over the study region.
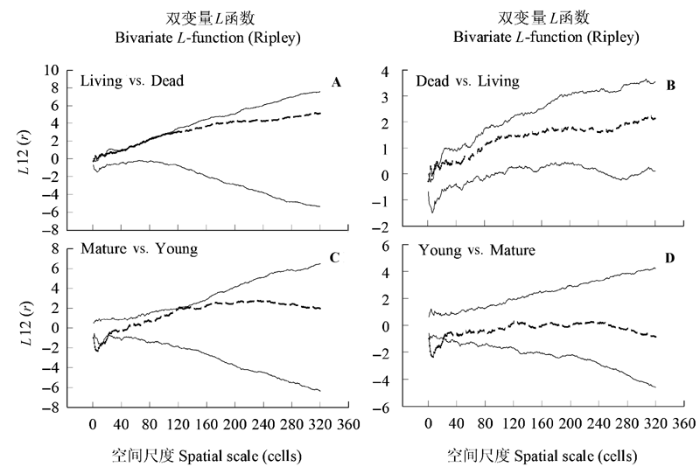
Fig. 6 Ripley’s L-function for spatial association of C. tibetica individuals under different survival status and developmental stages with confidence envelopes (lines). r referred to radius of sample circle from C. tibetica as center, Confidence envelopes are the highest and lowest L(r) of 999 replicates of a random labeling null model for individuals under different survival status and of a population independence null model for individuals under developmental stages.
Ecological vicariance of steppe species and communities on climate gradient in Inner Mongolia and its indication to steppe dynamics under the global change.
Acta Phytoecologica Sinica, 20, 193-206. (in Chinese with English abstract)[李永宏 (1996).
Have southern texas savannas been converted to woodlands in recent history?
1
1989
... 近50年, 越来越多的科学家发现灌木植物在草地中的数量增多和分布范围扩大的现象, 并指出这可能是全球气候和人类活动的改变导致的.随着灌木逐渐增多, 形成了灌木斑块与草本植物群落相复合的植被景观, 它是在干旱半干旱地区出现的一种新的植被类型, 称为灌丛化草原(陈蕾伊等, 2014).灌丛化草原在干旱半干旱区广泛分布, 占干旱半干旱区面积的10%-20% (van Auken, 2000; Eldridge et al., 2011).近年来的研究指出, 草原中灌木的出现改变了光照、温度、水分和养分环境, 也改变了物种组成和群落结构, 这使得灌丛化草原的结构、功能及生态过程有别于草原(Eldridge et al., 2012 ).早在200多年前, 人们就注意到干旱半干旱地区草原的灌丛化现象, 但真正对其形成机制和影响进行研究则始于20世纪80年代后期(Archer, 1989; Grover & Musick, 1990), 并引起全球广泛关注(van Auken, 2000; Jackson et al., 2002; Maestre et al., 2009). ...
Autogenic succession in a subtropical savanna: Conversion of grassland to thorn woodland.
1
1988
... 目前国外主要关注于北美(Buffington & Herbel, 1965; Archer et al., 1988; Knapp et al., 2008)和非洲南部(Eldridge et al., 2011)的灌丛化草原, 集中研究草原灌丛化的成因(Knoop & Walker, 1985; Bond & Midgley, 2001; Coetzee et al., 2007)和灌丛化对生态系统生产力(Phinn et al., 1996; Oba et al., 2000; Knapp et al., 2008)、生物多样性(Eldridge et al., 2011)和土壤碳储量(Jackson et al., 2002)的影响.在国内, 主要集中于内蒙古灌丛化草原的研究(周道玮, 1990; 刘书润, 1991; Chen et al., 2015), 涉及灌丛化的驱动因子(Chen et al., 2015)和灌丛化对土壤水分和养分的影响研究(张璞进等, 2009; 彭海英等, 2014).但关于灌丛化草原植物种群的空间格局研究还没有.植物种群空间格局分析是研究植物种群特征、种群间相互作用及种群与环境关系的重要手段.空间分布格局体现了种群的结构和动态, 可以揭示种群的生态特性和发展趋势; 而空间关联性是指同一物种的不同个体或不同物种个体在空间上的联结性, 体现了群落的形成和演化机制(Greig-Smith, 1983). ...
Contribution to the discussion of dr. Ripley’s paper.
A proposed CO2-controlled mechanism of woody plant invasion in grasslands and savannas.
2
2001
... 目前国外主要关注于北美(Buffington & Herbel, 1965; Archer et al., 1988; Knapp et al., 2008)和非洲南部(Eldridge et al., 2011)的灌丛化草原, 集中研究草原灌丛化的成因(Knoop & Walker, 1985; Bond & Midgley, 2001; Coetzee et al., 2007)和灌丛化对生态系统生产力(Phinn et al., 1996; Oba et al., 2000; Knapp et al., 2008)、生物多样性(Eldridge et al., 2011)和土壤碳储量(Jackson et al., 2002)的影响.在国内, 主要集中于内蒙古灌丛化草原的研究(周道玮, 1990; 刘书润, 1991; Chen et al., 2015), 涉及灌丛化的驱动因子(Chen et al., 2015)和灌丛化对土壤水分和养分的影响研究(张璞进等, 2009; 彭海英等, 2014).但关于灌丛化草原植物种群的空间格局研究还没有.植物种群空间格局分析是研究植物种群特征、种群间相互作用及种群与环境关系的重要手段.空间分布格局体现了种群的结构和动态, 可以揭示种群的生态特性和发展趋势; 而空间关联性是指同一物种的不同个体或不同物种个体在空间上的联结性, 体现了群落的形成和演化机制(Greig-Smith, 1983). ...
Vegetational changes on a semidesert grassland range from 1858 to 1963.
1
1965
... 目前国外主要关注于北美(Buffington & Herbel, 1965; Archer et al., 1988; Knapp et al., 2008)和非洲南部(Eldridge et al., 2011)的灌丛化草原, 集中研究草原灌丛化的成因(Knoop & Walker, 1985; Bond & Midgley, 2001; Coetzee et al., 2007)和灌丛化对生态系统生产力(Phinn et al., 1996; Oba et al., 2000; Knapp et al., 2008)、生物多样性(Eldridge et al., 2011)和土壤碳储量(Jackson et al., 2002)的影响.在国内, 主要集中于内蒙古灌丛化草原的研究(周道玮, 1990; 刘书润, 1991; Chen et al., 2015), 涉及灌丛化的驱动因子(Chen et al., 2015)和灌丛化对土壤水分和养分的影响研究(张璞进等, 2009; 彭海英等, 2014).但关于灌丛化草原植物种群的空间格局研究还没有.植物种群空间格局分析是研究植物种群特征、种群间相互作用及种群与环境关系的重要手段.空间分布格局体现了种群的结构和动态, 可以揭示种群的生态特性和发展趋势; 而空间关联性是指同一物种的不同个体或不同物种个体在空间上的联结性, 体现了群落的形成和演化机制(Greig-Smith, 1983). ...
Climate and native grassland vegetation as drivers of the community structures of shrub-encroached grasslands in Inner Mongolia, China.
2
2015
... 目前国外主要关注于北美(Buffington & Herbel, 1965; Archer et al., 1988; Knapp et al., 2008)和非洲南部(Eldridge et al., 2011)的灌丛化草原, 集中研究草原灌丛化的成因(Knoop & Walker, 1985; Bond & Midgley, 2001; Coetzee et al., 2007)和灌丛化对生态系统生产力(Phinn et al., 1996; Oba et al., 2000; Knapp et al., 2008)、生物多样性(Eldridge et al., 2011)和土壤碳储量(Jackson et al., 2002)的影响.在国内, 主要集中于内蒙古灌丛化草原的研究(周道玮, 1990; 刘书润, 1991; Chen et al., 2015), 涉及灌丛化的驱动因子(Chen et al., 2015)和灌丛化对土壤水分和养分的影响研究(张璞进等, 2009; 彭海英等, 2014).但关于灌丛化草原植物种群的空间格局研究还没有.植物种群空间格局分析是研究植物种群特征、种群间相互作用及种群与环境关系的重要手段.空间分布格局体现了种群的结构和动态, 可以揭示种群的生态特性和发展趋势; 而空间关联性是指同一物种的不同个体或不同物种个体在空间上的联结性, 体现了群落的形成和演化机制(Greig-Smith, 1983). ...
... 近50年, 越来越多的科学家发现灌木植物在草地中的数量增多和分布范围扩大的现象, 并指出这可能是全球气候和人类活动的改变导致的.随着灌木逐渐增多, 形成了灌木斑块与草本植物群落相复合的植被景观, 它是在干旱半干旱地区出现的一种新的植被类型, 称为灌丛化草原(陈蕾伊等, 2014).灌丛化草原在干旱半干旱区广泛分布, 占干旱半干旱区面积的10%-20% (van Auken, 2000; Eldridge et al., 2011).近年来的研究指出, 草原中灌木的出现改变了光照、温度、水分和养分环境, 也改变了物种组成和群落结构, 这使得灌丛化草原的结构、功能及生态过程有别于草原(Eldridge et al., 2012 ).早在200多年前, 人们就注意到干旱半干旱地区草原的灌丛化现象, 但真正对其形成机制和影响进行研究则始于20世纪80年代后期(Archer, 1989; Grover & Musick, 1990), 并引起全球广泛关注(van Auken, 2000; Jackson et al., 2002; Maestre et al., 2009). ...
Overgrazing and bush encroachment by tarchonanthus camphoratus in a semi-arid savanna.
2
2007
... 目前国外主要关注于北美(Buffington & Herbel, 1965; Archer et al., 1988; Knapp et al., 2008)和非洲南部(Eldridge et al., 2011)的灌丛化草原, 集中研究草原灌丛化的成因(Knoop & Walker, 1985; Bond & Midgley, 2001; Coetzee et al., 2007)和灌丛化对生态系统生产力(Phinn et al., 1996; Oba et al., 2000; Knapp et al., 2008)、生物多样性(Eldridge et al., 2011)和土壤碳储量(Jackson et al., 2002)的影响.在国内, 主要集中于内蒙古灌丛化草原的研究(周道玮, 1990; 刘书润, 1991; Chen et al., 2015), 涉及灌丛化的驱动因子(Chen et al., 2015)和灌丛化对土壤水分和养分的影响研究(张璞进等, 2009; 彭海英等, 2014).但关于灌丛化草原植物种群的空间格局研究还没有.植物种群空间格局分析是研究植物种群特征、种群间相互作用及种群与环境关系的重要手段.空间分布格局体现了种群的结构和动态, 可以揭示种群的生态特性和发展趋势; 而空间关联性是指同一物种的不同个体或不同物种个体在空间上的联结性, 体现了群落的形成和演化机制(Greig-Smith, 1983). ...
Impacts of shrub encroachment on ecosystem structure and functioning: Towards a global synthesis.
3
2011
... 近50年, 越来越多的科学家发现灌木植物在草地中的数量增多和分布范围扩大的现象, 并指出这可能是全球气候和人类活动的改变导致的.随着灌木逐渐增多, 形成了灌木斑块与草本植物群落相复合的植被景观, 它是在干旱半干旱地区出现的一种新的植被类型, 称为灌丛化草原(陈蕾伊等, 2014).灌丛化草原在干旱半干旱区广泛分布, 占干旱半干旱区面积的10%-20% (van Auken, 2000; Eldridge et al., 2011).近年来的研究指出, 草原中灌木的出现改变了光照、温度、水分和养分环境, 也改变了物种组成和群落结构, 这使得灌丛化草原的结构、功能及生态过程有别于草原(Eldridge et al., 2012 ).早在200多年前, 人们就注意到干旱半干旱地区草原的灌丛化现象, 但真正对其形成机制和影响进行研究则始于20世纪80年代后期(Archer, 1989; Grover & Musick, 1990), 并引起全球广泛关注(van Auken, 2000; Jackson et al., 2002; Maestre et al., 2009). ...
... 目前国外主要关注于北美(Buffington & Herbel, 1965; Archer et al., 1988; Knapp et al., 2008)和非洲南部(Eldridge et al., 2011)的灌丛化草原, 集中研究草原灌丛化的成因(Knoop & Walker, 1985; Bond & Midgley, 2001; Coetzee et al., 2007)和灌丛化对生态系统生产力(Phinn et al., 1996; Oba et al., 2000; Knapp et al., 2008)、生物多样性(Eldridge et al., 2011)和土壤碳储量(Jackson et al., 2002)的影响.在国内, 主要集中于内蒙古灌丛化草原的研究(周道玮, 1990; 刘书润, 1991; Chen et al., 2015), 涉及灌丛化的驱动因子(Chen et al., 2015)和灌丛化对土壤水分和养分的影响研究(张璞进等, 2009; 彭海英等, 2014).但关于灌丛化草原植物种群的空间格局研究还没有.植物种群空间格局分析是研究植物种群特征、种群间相互作用及种群与环境关系的重要手段.空间分布格局体现了种群的结构和动态, 可以揭示种群的生态特性和发展趋势; 而空间关联性是指同一物种的不同个体或不同物种个体在空间上的联结性, 体现了群落的形成和演化机制(Greig-Smith, 1983). ...
... )、生物多样性(Eldridge et al., 2011)和土壤碳储量(Jackson et al., 2002)的影响.在国内, 主要集中于内蒙古灌丛化草原的研究(周道玮, 1990; 刘书润, 1991; Chen et al., 2015), 涉及灌丛化的驱动因子(Chen et al., 2015)和灌丛化对土壤水分和养分的影响研究(张璞进等, 2009; 彭海英等, 2014).但关于灌丛化草原植物种群的空间格局研究还没有.植物种群空间格局分析是研究植物种群特征、种群间相互作用及种群与环境关系的重要手段.空间分布格局体现了种群的结构和动态, 可以揭示种群的生态特性和发展趋势; 而空间关联性是指同一物种的不同个体或不同物种个体在空间上的联结性, 体现了群落的形成和演化机制(Greig-Smith, 1983). ...
A global database of shrub encroachment effects on ecosystem structure and functioning.
2012
Spatial pattern and neighbour effects on Helianthemum squamatum seedlings in a Mediterranean gypsum community.
1
2005
... 毛刺锦鸡儿幼年个体在0.3-20.4 m之间为聚集分布, 其他范围为随机分布.已有研究表明Helian- themum squamatum幼年个体在小尺度上表现为聚集分布的格局(Escudero et al., 2005), 而毛刺锦鸡儿幼年个体在0-0.3 m的范围内却表现为随机分布的格局.毛刺锦鸡儿种子传播方式主要是靠重力散播, 散播范围小, 主要是散落在母株的附近, 但种子传播是一个随机的过程, 并且受到干旱胁迫, 种子能萌发、发育、成长是一个随机的过程(Wiegand et al., 2000a).这些随机过程的综合也可解释毛刺锦鸡儿幼年个体在更大范围内(>20.4 m)的随机分布格局.在0.3-20.4 m之间的聚集分布格局, 可能是毛刺锦鸡儿适应干旱环境的一种策略.群落中聚集分布的物种可以增强它与群落内其他物种对资源的竞争能力(Raventos et al., 2010).毛刺锦鸡儿幼年个体的聚集分布, 可以提升它们在种群内同其他物种间的竞争能力, 提高幼年个体的存活率, 促进种群的更新和整个种群的存活力.但随着幼年植株的成长, 内部资源难以维持所有幼年个体发育成熟, 特别是在成熟个体的竞争下, 必然会出现自疏现象或密度制约现象(Raventos et al., 2010).这种竞争一方面可能导致由幼年个体发育的成熟个体在小范围内出现均匀分布的空间格局; 另一方面可能导致呈聚集分布的毛刺锦鸡儿幼年个体因竞争而死亡的毛刺锦鸡儿在一定范围内也服从聚集分布. ...
Quantitative plant ecology. In: Anderson JA, Greig-Smith P, Pitelka FA eds. Studies in Ecology
2
1983
... 目前国外主要关注于北美(Buffington & Herbel, 1965; Archer et al., 1988; Knapp et al., 2008)和非洲南部(Eldridge et al., 2011)的灌丛化草原, 集中研究草原灌丛化的成因(Knoop & Walker, 1985; Bond & Midgley, 2001; Coetzee et al., 2007)和灌丛化对生态系统生产力(Phinn et al., 1996; Oba et al., 2000; Knapp et al., 2008)、生物多样性(Eldridge et al., 2011)和土壤碳储量(Jackson et al., 2002)的影响.在国内, 主要集中于内蒙古灌丛化草原的研究(周道玮, 1990; 刘书润, 1991; Chen et al., 2015), 涉及灌丛化的驱动因子(Chen et al., 2015)和灌丛化对土壤水分和养分的影响研究(张璞进等, 2009; 彭海英等, 2014).但关于灌丛化草原植物种群的空间格局研究还没有.植物种群空间格局分析是研究植物种群特征、种群间相互作用及种群与环境关系的重要手段.空间分布格局体现了种群的结构和动态, 可以揭示种群的生态特性和发展趋势; 而空间关联性是指同一物种的不同个体或不同物种个体在空间上的联结性, 体现了群落的形成和演化机制(Greig-Smith, 1983). ...
... 在鄂尔多斯鄂托克旗选择具有代表性和典型性的毛刺锦鸡儿灌丛化短花针茅+无芒隐子草群落为研究样地, 样地面积约50 km2, 地理位置为39.50° N, 107.28° E, 并在样地中设立1个64 m × 64 m的大样方.测定草本群落植物的空间位置有Greig-smith邻接格子样方法(Greig-Smith, 1983)和摄影定位法(王鑫厅等, 2006)法, 它们适合地势平坦的小面积植物群落.对于半干旱干旱地区较为稀疏的灌木群落, 用上述2种方法测定大样方中灌木的空间位置不是最佳的.本研究采用新方法——“三线定位法”: 将64 m × 64 m的大样方分割为4个32 m × 32 m的亚样方(图1).在每一个亚样方中c的长度是固定的, 测量出a和b的长度.通过三角函数计算以确定该灌木的空间坐标.与此同时, 测定灌木冠幅、高度以及存活状态(是否死亡), 经过数据处理和运算, 将得到的4个32 m × 32 m的亚样方数据通过坐标换算合并成为一个64 m × 64 m样方数据. ...
Shrubland encroachment in southern new mexico, USA: An analysis of desertification processes in the American southwest.
1
1990
... 近50年, 越来越多的科学家发现灌木植物在草地中的数量增多和分布范围扩大的现象, 并指出这可能是全球气候和人类活动的改变导致的.随着灌木逐渐增多, 形成了灌木斑块与草本植物群落相复合的植被景观, 它是在干旱半干旱地区出现的一种新的植被类型, 称为灌丛化草原(陈蕾伊等, 2014).灌丛化草原在干旱半干旱区广泛分布, 占干旱半干旱区面积的10%-20% (van Auken, 2000; Eldridge et al., 2011).近年来的研究指出, 草原中灌木的出现改变了光照、温度、水分和养分环境, 也改变了物种组成和群落结构, 这使得灌丛化草原的结构、功能及生态过程有别于草原(Eldridge et al., 2012 ).早在200多年前, 人们就注意到干旱半干旱地区草原的灌丛化现象, 但真正对其形成机制和影响进行研究则始于20世纪80年代后期(Archer, 1989; Grover & Musick, 1990), 并引起全球广泛关注(van Auken, 2000; Jackson et al., 2002; Maestre et al., 2009). ...
Ecosystem carbon loss with woody plant invasion of grasslands.
2
2002
... 近50年, 越来越多的科学家发现灌木植物在草地中的数量增多和分布范围扩大的现象, 并指出这可能是全球气候和人类活动的改变导致的.随着灌木逐渐增多, 形成了灌木斑块与草本植物群落相复合的植被景观, 它是在干旱半干旱地区出现的一种新的植被类型, 称为灌丛化草原(陈蕾伊等, 2014).灌丛化草原在干旱半干旱区广泛分布, 占干旱半干旱区面积的10%-20% (van Auken, 2000; Eldridge et al., 2011).近年来的研究指出, 草原中灌木的出现改变了光照、温度、水分和养分环境, 也改变了物种组成和群落结构, 这使得灌丛化草原的结构、功能及生态过程有别于草原(Eldridge et al., 2012 ).早在200多年前, 人们就注意到干旱半干旱地区草原的灌丛化现象, 但真正对其形成机制和影响进行研究则始于20世纪80年代后期(Archer, 1989; Grover & Musick, 1990), 并引起全球广泛关注(van Auken, 2000; Jackson et al., 2002; Maestre et al., 2009). ...
... 目前国外主要关注于北美(Buffington & Herbel, 1965; Archer et al., 1988; Knapp et al., 2008)和非洲南部(Eldridge et al., 2011)的灌丛化草原, 集中研究草原灌丛化的成因(Knoop & Walker, 1985; Bond & Midgley, 2001; Coetzee et al., 2007)和灌丛化对生态系统生产力(Phinn et al., 1996; Oba et al., 2000; Knapp et al., 2008)、生物多样性(Eldridge et al., 2011)和土壤碳储量(Jackson et al., 2002)的影响.在国内, 主要集中于内蒙古灌丛化草原的研究(周道玮, 1990; 刘书润, 1991; Chen et al., 2015), 涉及灌丛化的驱动因子(Chen et al., 2015)和灌丛化对土壤水分和养分的影响研究(张璞进等, 2009; 彭海英等, 2014).但关于灌丛化草原植物种群的空间格局研究还没有.植物种群空间格局分析是研究植物种群特征、种群间相互作用及种群与环境关系的重要手段.空间分布格局体现了种群的结构和动态, 可以揭示种群的生态特性和发展趋势; 而空间关联性是指同一物种的不同个体或不同物种个体在空间上的联结性, 体现了群落的形成和演化机制(Greig-Smith, 1983). ...
Pattern of self-thinning in jack pine: Testing the random mortality hypothesis.
Shrub encroachment in north american grasslands: Shifts in growth form dominance rapidly alters control of ecosystem carbon inputs.
2
2008
... 目前国外主要关注于北美(Buffington & Herbel, 1965; Archer et al., 1988; Knapp et al., 2008)和非洲南部(Eldridge et al., 2011)的灌丛化草原, 集中研究草原灌丛化的成因(Knoop & Walker, 1985; Bond & Midgley, 2001; Coetzee et al., 2007)和灌丛化对生态系统生产力(Phinn et al., 1996; Oba et al., 2000; Knapp et al., 2008)、生物多样性(Eldridge et al., 2011)和土壤碳储量(Jackson et al., 2002)的影响.在国内, 主要集中于内蒙古灌丛化草原的研究(周道玮, 1990; 刘书润, 1991; Chen et al., 2015), 涉及灌丛化的驱动因子(Chen et al., 2015)和灌丛化对土壤水分和养分的影响研究(张璞进等, 2009; 彭海英等, 2014).但关于灌丛化草原植物种群的空间格局研究还没有.植物种群空间格局分析是研究植物种群特征、种群间相互作用及种群与环境关系的重要手段.空间分布格局体现了种群的结构和动态, 可以揭示种群的生态特性和发展趋势; 而空间关联性是指同一物种的不同个体或不同物种个体在空间上的联结性, 体现了群落的形成和演化机制(Greig-Smith, 1983). ...
... ; Knapp et al., 2008)、生物多样性(Eldridge et al., 2011)和土壤碳储量(Jackson et al., 2002)的影响.在国内, 主要集中于内蒙古灌丛化草原的研究(周道玮, 1990; 刘书润, 1991; Chen et al., 2015), 涉及灌丛化的驱动因子(Chen et al., 2015)和灌丛化对土壤水分和养分的影响研究(张璞进等, 2009; 彭海英等, 2014).但关于灌丛化草原植物种群的空间格局研究还没有.植物种群空间格局分析是研究植物种群特征、种群间相互作用及种群与环境关系的重要手段.空间分布格局体现了种群的结构和动态, 可以揭示种群的生态特性和发展趋势; 而空间关联性是指同一物种的不同个体或不同物种个体在空间上的联结性, 体现了群落的形成和演化机制(Greig-Smith, 1983). ...
Interactions of woody and herb- aceous vegetation in a southern African savanna.
2
1985
... 目前国外主要关注于北美(Buffington & Herbel, 1965; Archer et al., 1988; Knapp et al., 2008)和非洲南部(Eldridge et al., 2011)的灌丛化草原, 集中研究草原灌丛化的成因(Knoop & Walker, 1985; Bond & Midgley, 2001; Coetzee et al., 2007)和灌丛化对生态系统生产力(Phinn et al., 1996; Oba et al., 2000; Knapp et al., 2008)、生物多样性(Eldridge et al., 2011)和土壤碳储量(Jackson et al., 2002)的影响.在国内, 主要集中于内蒙古灌丛化草原的研究(周道玮, 1990; 刘书润, 1991; Chen et al., 2015), 涉及灌丛化的驱动因子(Chen et al., 2015)和灌丛化对土壤水分和养分的影响研究(张璞进等, 2009; 彭海英等, 2014).但关于灌丛化草原植物种群的空间格局研究还没有.植物种群空间格局分析是研究植物种群特征、种群间相互作用及种群与环境关系的重要手段.空间分布格局体现了种群的结构和动态, 可以揭示种群的生态特性和发展趋势; 而空间关联性是指同一物种的不同个体或不同物种个体在空间上的联结性, 体现了群落的形成和演化机制(Greig-Smith, 1983). ...
The problem of pattern and scale in ecology: The Robert H. Macarthur award lecture.
1
1992
... 在调查样方内, 毛刺锦鸡儿种群空间分布格局总的来说体现出多样性的空间分布格局.物种的空间格局与空间尺度之间关系密切, 具有尺度效应(Wiegand et al., 2009; Wang et al., 2010), 这些变化的空间分布格局和种内关系是适应环境的结果, 体现出它们对环境的适应性(Levin, 1992).在干旱的毛刺锦鸡儿灌丛化草原中, 水分是限制性因子, 成熟的毛刺锦鸡儿个体在<3.7 m范围内为均匀分布格局, 是成熟个体间争夺资源(尤其是短缺的水资源)的结果, 一部分个体会因竞争被排斥出去; 在>3.7 m范围为随机分布格局, 是土壤水分资源空间分布格局引起的, 有限的土壤水分资源, 减小了土壤水分的空间自相关距离, 物种间在更大的范围内缺少竞争, 物种趋于随机分布格局(Watkinson, 1980). ...
内蒙古草原植物的生态替代及其对全球变化下草原动态的指示
1
1996
... 灌丛化草原在内蒙古分布较广、面积较大.针茅属(Stipa)植物是灌丛化草原草本层片的优势种, 局部地段以羊草(Leymus chinensis)、冰草(Agropyron cristatum)和无芒隐子草(Cleistogenes songorica)为优势种; 灌木层片主要是锦鸡儿属(Caragana)植物, 一些地方为长柄扁桃(Prunus pedunculata)和猫头刺(Oxytropis aciphylla).灌丛化草原中针茅属和锦鸡儿属植物随着气候逐渐的干旱会出现替代分布现象, 针茅属植物依次出现大针茅(Stipa grandis)、西北针茅(Stipa sareptana var. krylovii)、石生针茅(Stipa tianschanica var. klemenzii)或短花针茅(Stipa breviflora), 这一现象与紧邻的针茅草原相一致(李永宏, 1996); 锦鸡儿属植物依次为小叶锦鸡儿(Car- agana microphylla)、矮锦鸡儿(Caragana pygmaea)或狭叶锦鸡儿(Caragana stenophylla)、毛刺锦鸡儿(Caragana tibetica).内蒙古灌丛化草原以小叶锦鸡儿和毛刺锦鸡儿灌丛化为主.据已有报道, 小叶锦鸡儿灌丛化草原约5000 km2 (周道玮, 1990); 鄂尔多斯市毛刺锦鸡儿灌丛化草原面积约3600 km2 (张璞进, 2011).本研究的前期调查发现, 巴彦淖尔市境内的毛刺锦鸡儿灌丛化草地面积近3000 km2. ...
小叶锦鸡儿灌丛化对锡林河流域草原植被的影响
1
1991
... 目前国外主要关注于北美(Buffington & Herbel, 1965; Archer et al., 1988; Knapp et al., 2008)和非洲南部(Eldridge et al., 2011)的灌丛化草原, 集中研究草原灌丛化的成因(Knoop & Walker, 1985; Bond & Midgley, 2001; Coetzee et al., 2007)和灌丛化对生态系统生产力(Phinn et al., 1996; Oba et al., 2000; Knapp et al., 2008)、生物多样性(Eldridge et al., 2011)和土壤碳储量(Jackson et al., 2002)的影响.在国内, 主要集中于内蒙古灌丛化草原的研究(周道玮, 1990; 刘书润, 1991; Chen et al., 2015), 涉及灌丛化的驱动因子(Chen et al., 2015)和灌丛化对土壤水分和养分的影响研究(张璞进等, 2009; 彭海英等, 2014).但关于灌丛化草原植物种群的空间格局研究还没有.植物种群空间格局分析是研究植物种群特征、种群间相互作用及种群与环境关系的重要手段.空间分布格局体现了种群的结构和动态, 可以揭示种群的生态特性和发展趋势; 而空间关联性是指同一物种的不同个体或不同物种个体在空间上的联结性, 体现了群落的形成和演化机制(Greig-Smith, 1983). ...
The role of nurse plants in the establishment of shrub seedlings in the semi-arid subtropical Andes.
1
2007
... 植株的大小可以很好地指示灌木的发育阶段(Wiegand et al., 2000b; López et al., 2007).已有研究表明, 在干旱的环境中, 灌木的冠幅与高度之间的关系和灌木的发育阶段有显著的相关关系(Ludwig et al., 1975; Reynolds et al., 1999).本文通过分析毛刺锦鸡儿的株高、冠幅以及不同冠幅区间内毛刺锦鸡儿的数量特征来确定毛刺锦鸡儿种群内个体的发育阶段, 进而分析种群结构(Wiegand et al., 2000b). ...
Methods for analysing spatial processes of several types of points
Size-biomass relations of several Chihuahuan desert shrubs.
1
1975
... 植株的大小可以很好地指示灌木的发育阶段(Wiegand et al., 2000b; López et al., 2007).已有研究表明, 在干旱的环境中, 灌木的冠幅与高度之间的关系和灌木的发育阶段有显著的相关关系(Ludwig et al., 1975; Reynolds et al., 1999).本文通过分析毛刺锦鸡儿的株高、冠幅以及不同冠幅区间内毛刺锦鸡儿的数量特征来确定毛刺锦鸡儿种群内个体的发育阶段, 进而分析种群结构(Wiegand et al., 2000b). ...
Shrub encroachment can reverse desertification in semi-arid mediterranean grasslands.
1
2009
... 近50年, 越来越多的科学家发现灌木植物在草地中的数量增多和分布范围扩大的现象, 并指出这可能是全球气候和人类活动的改变导致的.随着灌木逐渐增多, 形成了灌木斑块与草本植物群落相复合的植被景观, 它是在干旱半干旱地区出现的一种新的植被类型, 称为灌丛化草原(陈蕾伊等, 2014).灌丛化草原在干旱半干旱区广泛分布, 占干旱半干旱区面积的10%-20% (van Auken, 2000; Eldridge et al., 2011).近年来的研究指出, 草原中灌木的出现改变了光照、温度、水分和养分环境, 也改变了物种组成和群落结构, 这使得灌丛化草原的结构、功能及生态过程有别于草原(Eldridge et al., 2012 ).早在200多年前, 人们就注意到干旱半干旱地区草原的灌丛化现象, 但真正对其形成机制和影响进行研究则始于20世纪80年代后期(Archer, 1989; Grover & Musick, 1990), 并引起全球广泛关注(van Auken, 2000; Jackson et al., 2002; Maestre et al., 2009). ...
Plant functional types of woody species related to fire disturbance in forest-grassland ecotones.
Bush cover and range condition assessments in relation to landscape and grazing in southern Ethiopia.
1
2000
... 目前国外主要关注于北美(Buffington & Herbel, 1965; Archer et al., 1988; Knapp et al., 2008)和非洲南部(Eldridge et al., 2011)的灌丛化草原, 集中研究草原灌丛化的成因(Knoop & Walker, 1985; Bond & Midgley, 2001; Coetzee et al., 2007)和灌丛化对生态系统生产力(Phinn et al., 1996; Oba et al., 2000; Knapp et al., 2008)、生物多样性(Eldridge et al., 2011)和土壤碳储量(Jackson et al., 2002)的影响.在国内, 主要集中于内蒙古灌丛化草原的研究(周道玮, 1990; 刘书润, 1991; Chen et al., 2015), 涉及灌丛化的驱动因子(Chen et al., 2015)和灌丛化对土壤水分和养分的影响研究(张璞进等, 2009; 彭海英等, 2014).但关于灌丛化草原植物种群的空间格局研究还没有.植物种群空间格局分析是研究植物种群特征、种群间相互作用及种群与环境关系的重要手段.空间分布格局体现了种群的结构和动态, 可以揭示种群的生态特性和发展趋势; 而空间关联性是指同一物种的不同个体或不同物种个体在空间上的联结性, 体现了群落的形成和演化机制(Greig-Smith, 1983). ...
内蒙古典型草原小叶锦鸡儿灌丛化对水分再分配和利用的影响
1
2014
... 目前国外主要关注于北美(Buffington & Herbel, 1965; Archer et al., 1988; Knapp et al., 2008)和非洲南部(Eldridge et al., 2011)的灌丛化草原, 集中研究草原灌丛化的成因(Knoop & Walker, 1985; Bond & Midgley, 2001; Coetzee et al., 2007)和灌丛化对生态系统生产力(Phinn et al., 1996; Oba et al., 2000; Knapp et al., 2008)、生物多样性(Eldridge et al., 2011)和土壤碳储量(Jackson et al., 2002)的影响.在国内, 主要集中于内蒙古灌丛化草原的研究(周道玮, 1990; 刘书润, 1991; Chen et al., 2015), 涉及灌丛化的驱动因子(Chen et al., 2015)和灌丛化对土壤水分和养分的影响研究(张璞进等, 2009; 彭海英等, 2014).但关于灌丛化草原植物种群的空间格局研究还没有.植物种群空间格局分析是研究植物种群特征、种群间相互作用及种群与环境关系的重要手段.空间分布格局体现了种群的结构和动态, 可以揭示种群的生态特性和发展趋势; 而空间关联性是指同一物种的不同个体或不同物种个体在空间上的联结性, 体现了群落的形成和演化机制(Greig-Smith, 1983). ...
Biomass distribution mapping using airborne digital video imagery and spatial statistics in a semi-arid environment.
1
1996
... 目前国外主要关注于北美(Buffington & Herbel, 1965; Archer et al., 1988; Knapp et al., 2008)和非洲南部(Eldridge et al., 2011)的灌丛化草原, 集中研究草原灌丛化的成因(Knoop & Walker, 1985; Bond & Midgley, 2001; Coetzee et al., 2007)和灌丛化对生态系统生产力(Phinn et al., 1996; Oba et al., 2000; Knapp et al., 2008)、生物多样性(Eldridge et al., 2011)和土壤碳储量(Jackson et al., 2002)的影响.在国内, 主要集中于内蒙古灌丛化草原的研究(周道玮, 1990; 刘书润, 1991; Chen et al., 2015), 涉及灌丛化的驱动因子(Chen et al., 2015)和灌丛化对土壤水分和养分的影响研究(张璞进等, 2009; 彭海英等, 2014).但关于灌丛化草原植物种群的空间格局研究还没有.植物种群空间格局分析是研究植物种群特征、种群间相互作用及种群与环境关系的重要手段.空间分布格局体现了种群的结构和动态, 可以揭示种群的生态特性和发展趋势; 而空间关联性是指同一物种的不同个体或不同物种个体在空间上的联结性, 体现了群落的形成和演化机制(Greig-Smith, 1983). ...
Elevated atmospheric CO2 magnifies intraspecific variation in seedling growth of honey mesquite: An assessment of relative growth rates.
Impact of drought on desert shrubs: Effects of seasonality and degree of resource island development.
1
1999
... 植株的大小可以很好地指示灌木的发育阶段(Wiegand et al., 2000b; López et al., 2007).已有研究表明, 在干旱的环境中, 灌木的冠幅与高度之间的关系和灌木的发育阶段有显著的相关关系(Ludwig et al., 1975; Reynolds et al., 1999).本文通过分析毛刺锦鸡儿的株高、冠幅以及不同冠幅区间内毛刺锦鸡儿的数量特征来确定毛刺锦鸡儿种群内个体的发育阶段, 进而分析种群结构(Wiegand et al., 2000b). ...
Species associations in an old-growth temperate forest in north-eastern China.
1
2010
... 在调查样方内, 毛刺锦鸡儿种群空间分布格局总的来说体现出多样性的空间分布格局.物种的空间格局与空间尺度之间关系密切, 具有尺度效应(Wiegand et al., 2009; Wang et al., 2010), 这些变化的空间分布格局和种内关系是适应环境的结果, 体现出它们对环境的适应性(Levin, 1992).在干旱的毛刺锦鸡儿灌丛化草原中, 水分是限制性因子, 成熟的毛刺锦鸡儿个体在<3.7 m范围内为均匀分布格局, 是成熟个体间争夺资源(尤其是短缺的水资源)的结果, 一部分个体会因竞争被排斥出去; 在>3.7 m范围为随机分布格局, 是土壤水分资源空间分布格局引起的, 有限的土壤水分资源, 减小了土壤水分的空间自相关距离, 物种间在更大的范围内缺少竞争, 物种趋于随机分布格局(Watkinson, 1980). ...
植物种群空间分布格局测定的新方法: 摄影定位法
1
2006
... 在鄂尔多斯鄂托克旗选择具有代表性和典型性的毛刺锦鸡儿灌丛化短花针茅+无芒隐子草群落为研究样地, 样地面积约50 km2, 地理位置为39.50° N, 107.28° E, 并在样地中设立1个64 m × 64 m的大样方.测定草本群落植物的空间位置有Greig-smith邻接格子样方法(Greig-Smith, 1983)和摄影定位法(王鑫厅等, 2006)法, 它们适合地势平坦的小面积植物群落.对于半干旱干旱地区较为稀疏的灌木群落, 用上述2种方法测定大样方中灌木的空间位置不是最佳的.本研究采用新方法——“三线定位法”: 将64 m × 64 m的大样方分割为4个32 m × 32 m的亚样方(图1).在每一个亚样方中c的长度是固定的, 测量出a和b的长度.通过三角函数计算以确定该灌木的空间坐标.与此同时, 测定灌木冠幅、高度以及存活状态(是否死亡), 经过数据处理和运算, 将得到的4个32 m × 32 m的亚样方数据通过坐标换算合并成为一个64 m × 64 m样方数据. ...
西鄂尔多斯高原干旱荒漠灌木群落空间分布格局及其竞争关系分析
2010
Density-dependence in single-species populations of plants.
2
1980
... 在调查样方内, 毛刺锦鸡儿种群空间分布格局总的来说体现出多样性的空间分布格局.物种的空间格局与空间尺度之间关系密切, 具有尺度效应(Wiegand et al., 2009; Wang et al., 2010), 这些变化的空间分布格局和种内关系是适应环境的结果, 体现出它们对环境的适应性(Levin, 1992).在干旱的毛刺锦鸡儿灌丛化草原中, 水分是限制性因子, 成熟的毛刺锦鸡儿个体在<3.7 m范围内为均匀分布格局, 是成熟个体间争夺资源(尤其是短缺的水资源)的结果, 一部分个体会因竞争被排斥出去; 在>3.7 m范围为随机分布格局, 是土壤水分资源空间分布格局引起的, 有限的土壤水分资源, 减小了土壤水分的空间自相关距离, 物种间在更大的范围内缺少竞争, 物种趋于随机分布格局(Watkinson, 1980). ...
... 毛刺锦鸡儿存活个体在0-32 m的范围内随着尺度的增加出现了由均匀分布(0-1.4 m)到随机分布(1.4-7.9 m), 到不明显的聚集分布(7.9-14.2 m), 再到随机分布(14.2-32.0 m)的空间格局变化.毛刺锦鸡儿种群在0-1.4 m范围内呈均匀分布格局, 可能是毛刺锦鸡儿种群内个体之间对资源激烈竞争的结果.在两个范围(1.4-7.9 m和14.2-32.0 m)内的随机分布格局可能是限制性环境因子(如土壤水分)空间分布比较均匀和个体间竞争小的作用结果(Watkin- son, 1980).存活个体在一定范围内(7.9-14.2 m)不明显的聚集分布的可能原因是幼年个体在这一范围内呈聚集分布格局.我们的研究结果与王彦阁等(2010)的研究结果相一致, 在干旱环境中灌木在小范围内出现均匀分布格局, 随着取样范围的增大, 分布格局以随机分布为主. ...
Do spatial effects play a role in the spatial distribution of desert-dwelling Acacia raddiana?
1
2000
... 毛刺锦鸡儿幼年个体在0.3-20.4 m之间为聚集分布, 其他范围为随机分布.已有研究表明Helian- themum squamatum幼年个体在小尺度上表现为聚集分布的格局(Escudero et al., 2005), 而毛刺锦鸡儿幼年个体在0-0.3 m的范围内却表现为随机分布的格局.毛刺锦鸡儿种子传播方式主要是靠重力散播, 散播范围小, 主要是散落在母株的附近, 但种子传播是一个随机的过程, 并且受到干旱胁迫, 种子能萌发、发育、成长是一个随机的过程(Wiegand et al., 2000a).这些随机过程的综合也可解释毛刺锦鸡儿幼年个体在更大范围内(>20.4 m)的随机分布格局.在0.3-20.4 m之间的聚集分布格局, 可能是毛刺锦鸡儿适应干旱环境的一种策略.群落中聚集分布的物种可以增强它与群落内其他物种对资源的竞争能力(Raventos et al., 2010).毛刺锦鸡儿幼年个体的聚集分布, 可以提升它们在种群内同其他物种间的竞争能力, 提高幼年个体的存活率, 促进种群的更新和整个种群的存活力.但随着幼年植株的成长, 内部资源难以维持所有幼年个体发育成熟, 特别是在成熟个体的竞争下, 必然会出现自疏现象或密度制约现象(Raventos et al., 2010).这种竞争一方面可能导致由幼年个体发育的成熟个体在小范围内出现均匀分布的空间格局; 另一方面可能导致呈聚集分布的毛刺锦鸡儿幼年个体因竞争而死亡的毛刺锦鸡儿在一定范围内也服从聚集分布. ...
Recruitment in tropical tree species: Revealing complex spatial patterns.
1
2009
... 在调查样方内, 毛刺锦鸡儿种群空间分布格局总的来说体现出多样性的空间分布格局.物种的空间格局与空间尺度之间关系密切, 具有尺度效应(Wiegand et al., 2009; Wang et al., 2010), 这些变化的空间分布格局和种内关系是适应环境的结果, 体现出它们对环境的适应性(Levin, 1992).在干旱的毛刺锦鸡儿灌丛化草原中, 水分是限制性因子, 成熟的毛刺锦鸡儿个体在<3.7 m范围内为均匀分布格局, 是成熟个体间争夺资源(尤其是短缺的水资源)的结果, 一部分个体会因竞争被排斥出去; 在>3.7 m范围为随机分布格局, 是土壤水分资源空间分布格局引起的, 有限的土壤水分资源, 减小了土壤水分的空间自相关距离, 物种间在更大的范围内缺少竞争, 物种趋于随机分布格局(Watkinson, 1980). ...
b). Live fast, die young: Estimating size-age relations and mortality pattern of shrubs species in the semi-arid Karoo, South Africa.
2
2000
... 植株的大小可以很好地指示灌木的发育阶段(Wiegand et al., 2000b; López et al., 2007).已有研究表明, 在干旱的环境中, 灌木的冠幅与高度之间的关系和灌木的发育阶段有显著的相关关系(Ludwig et al., 1975; Reynolds et al., 1999).本文通过分析毛刺锦鸡儿的株高、冠幅以及不同冠幅区间内毛刺锦鸡儿的数量特征来确定毛刺锦鸡儿种群内个体的发育阶段, 进而分析种群结构(Wiegand et al., 2000b). ...
... ).本文通过分析毛刺锦鸡儿的株高、冠幅以及不同冠幅区间内毛刺锦鸡儿的数量特征来确定毛刺锦鸡儿种群内个体的发育阶段, 进而分析种群结构(Wiegand et al., 2000b). ...
Shrubs, granivores and annual plant community stability in an arid ecosystem.
... 灌丛化草原在内蒙古分布较广、面积较大.针茅属(Stipa)植物是灌丛化草原草本层片的优势种, 局部地段以羊草(Leymus chinensis)、冰草(Agropyron cristatum)和无芒隐子草(Cleistogenes songorica)为优势种; 灌木层片主要是锦鸡儿属(Caragana)植物, 一些地方为长柄扁桃(Prunus pedunculata)和猫头刺(Oxytropis aciphylla).灌丛化草原中针茅属和锦鸡儿属植物随着气候逐渐的干旱会出现替代分布现象, 针茅属植物依次出现大针茅(Stipa grandis)、西北针茅(Stipa sareptana var. krylovii)、石生针茅(Stipa tianschanica var. klemenzii)或短花针茅(Stipa breviflora), 这一现象与紧邻的针茅草原相一致(李永宏, 1996); 锦鸡儿属植物依次为小叶锦鸡儿(Car- agana microphylla)、矮锦鸡儿(Caragana pygmaea)或狭叶锦鸡儿(Caragana stenophylla)、毛刺锦鸡儿(Caragana tibetica).内蒙古灌丛化草原以小叶锦鸡儿和毛刺锦鸡儿灌丛化为主.据已有报道, 小叶锦鸡儿灌丛化草原约5000 km2 (周道玮, 1990); 鄂尔多斯市毛刺锦鸡儿灌丛化草原面积约3600 km2 (张璞进, 2011).本研究的前期调查发现, 巴彦淖尔市境内的毛刺锦鸡儿灌丛化草地面积近3000 km2. ...
藏锦鸡儿群落土壤资源空间异质性
1
2009
... 目前国外主要关注于北美(Buffington & Herbel, 1965; Archer et al., 1988; Knapp et al., 2008)和非洲南部(Eldridge et al., 2011)的灌丛化草原, 集中研究草原灌丛化的成因(Knoop & Walker, 1985; Bond & Midgley, 2001; Coetzee et al., 2007)和灌丛化对生态系统生产力(Phinn et al., 1996; Oba et al., 2000; Knapp et al., 2008)、生物多样性(Eldridge et al., 2011)和土壤碳储量(Jackson et al., 2002)的影响.在国内, 主要集中于内蒙古灌丛化草原的研究(周道玮, 1990; 刘书润, 1991; Chen et al., 2015), 涉及灌丛化的驱动因子(Chen et al., 2015)和灌丛化对土壤水分和养分的影响研究(张璞进等, 2009; 彭海英等, 2014).但关于灌丛化草原植物种群的空间格局研究还没有.植物种群空间格局分析是研究植物种群特征、种群间相互作用及种群与环境关系的重要手段.空间分布格局体现了种群的结构和动态, 可以揭示种群的生态特性和发展趋势; 而空间关联性是指同一物种的不同个体或不同物种个体在空间上的联结性, 体现了群落的形成和演化机制(Greig-Smith, 1983). ...
内蒙古小叶锦鸡儿灌丛化草地
3
1990
... 目前国外主要关注于北美(Buffington & Herbel, 1965; Archer et al., 1988; Knapp et al., 2008)和非洲南部(Eldridge et al., 2011)的灌丛化草原, 集中研究草原灌丛化的成因(Knoop & Walker, 1985; Bond & Midgley, 2001; Coetzee et al., 2007)和灌丛化对生态系统生产力(Phinn et al., 1996; Oba et al., 2000; Knapp et al., 2008)、生物多样性(Eldridge et al., 2011)和土壤碳储量(Jackson et al., 2002)的影响.在国内, 主要集中于内蒙古灌丛化草原的研究(周道玮, 1990; 刘书润, 1991; Chen et al., 2015), 涉及灌丛化的驱动因子(Chen et al., 2015)和灌丛化对土壤水分和养分的影响研究(张璞进等, 2009; 彭海英等, 2014).但关于灌丛化草原植物种群的空间格局研究还没有.植物种群空间格局分析是研究植物种群特征、种群间相互作用及种群与环境关系的重要手段.空间分布格局体现了种群的结构和动态, 可以揭示种群的生态特性和发展趋势; 而空间关联性是指同一物种的不同个体或不同物种个体在空间上的联结性, 体现了群落的形成和演化机制(Greig-Smith, 1983). ...
... 灌丛化草原在内蒙古分布较广、面积较大.针茅属(Stipa)植物是灌丛化草原草本层片的优势种, 局部地段以羊草(Leymus chinensis)、冰草(Agropyron cristatum)和无芒隐子草(Cleistogenes songorica)为优势种; 灌木层片主要是锦鸡儿属(Caragana)植物, 一些地方为长柄扁桃(Prunus pedunculata)和猫头刺(Oxytropis aciphylla).灌丛化草原中针茅属和锦鸡儿属植物随着气候逐渐的干旱会出现替代分布现象, 针茅属植物依次出现大针茅(Stipa grandis)、西北针茅(Stipa sareptana var. krylovii)、石生针茅(Stipa tianschanica var. klemenzii)或短花针茅(Stipa breviflora), 这一现象与紧邻的针茅草原相一致(李永宏, 1996); 锦鸡儿属植物依次为小叶锦鸡儿(Car- agana microphylla)、矮锦鸡儿(Caragana pygmaea)或狭叶锦鸡儿(Caragana stenophylla)、毛刺锦鸡儿(Caragana tibetica).内蒙古灌丛化草原以小叶锦鸡儿和毛刺锦鸡儿灌丛化为主.据已有报道, 小叶锦鸡儿灌丛化草原约5000 km2 (周道玮, 1990); 鄂尔多斯市毛刺锦鸡儿灌丛化草原面积约3600 km2 (张璞进, 2011).本研究的前期调查发现, 巴彦淖尔市境内的毛刺锦鸡儿灌丛化草地面积近3000 km2. ...

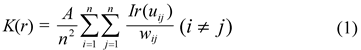

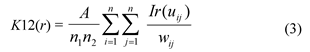
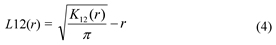


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}