维持体内K+ /Na+ 平衡是植物适应盐胁迫环境的一个关键手段, 利用先进技术手段和转基因技术, 人们对其机制开展了大量有益的探索研究, 发现植物一方面通过限制Na+ 的进入和增加Na+ 的外排或区隔化, 以减少体内或细胞质中的Na+ 浓度, 另一方面限制体内K+ 流失, 从而实现体内K+ /Na+ 平衡(Chao et al ., 2013 ; Feki et al ., 2014 )。早期的研究结果显示细胞质中的Na+ 外排主要由质膜上的Na+ /H+ 逆向转运蛋白(SOS1)负责, 并由H+ -ATPase酶负责提供驱动力, 同时引起细胞膜电位重新极化, 减少Na+ 的流入(Shi et al ., 2000 , 2003 ; Maathuis, 2006 )。另外, K+ 的吸收会受到Na+ 的竞争性抑制, 并且诱导通过外向K+ 通道而外流的K+ 增加(Demidchik & Maathuis, 2007 ; Britto & Kronzucker, 2008 ; Shabala & Cuin, 2008 )。同时, 有研究显示在盐胁迫下大麦(Hordeum vulgare )和小麦(Triticum aestivum )耐盐型品种根系的K+ 外流量均小于盐敏感型品种(Chen et al ., 2005 ; Cuin et al ., 2008 )。因此, 了解植物根系区域Na+ 和K+ 离子流情况, 对于理解植物耐盐能力非常重要。
沙枣(Elaeagnus angustifolia )是胡颓子科胡颓子属的落叶小乔木, 主要分布于我国西北五省, 在北京、天津和山东等地也开展了引种栽培, 具有耐旱、耐盐碱和抗风沙的特性, 能提高土壤肥力, 改善土壤环境, 已经被列为半干旱地区和盐土地防护林带的重要树种(郭丽君和王玉涛, 2008 ; 管文轲和徐娜, 2012 ; 杨升等, 2014 )。大量的研究结果显示, 沙枣幼苗在盐胁迫下, 根、茎和叶片中Na+ 含量增加, 而K+ 含量减少(张宝泽等, 1992 ; 刘宝玉, 2007 ; 刘正祥等, 2014 )。前期研究也发现, 沙枣幼苗随着NaCl浓度的增加, 根、茎和叶中的Na+ 含量显著增加, 而K+ 含量显著减少, 并且阿拉尔种源沙枣幼苗与银川种源幼苗间存在显著性差异, 阿拉尔种源沙枣幼苗能积累更多的Na+ 在根中, 而叶片中Na+ 含量较少, K+ 含量在NaCl胁迫下的降幅也低于银川种源(杨升等, 2014 , 2016 )。但是这些结果都是植株在试验中积累下来的一个总体含量, 是静态的, 而研究探讨盐胁迫下Na+ 和K+ 在沙枣体内具有怎样的动态运移机制, 并且比较不同种源间的差异, 有利于进一步理解沙枣的耐盐机制。因此, 本研究选取沙枣银川种源(YC, 盐敏感型)和阿拉尔种源(ALE, 耐盐型)水培苗为试验材料(杨升等, 2015 , 2016 ), 采用非损伤微测技术(non-invasive micro-test technology, NMT)研究: (1)在150 mmol·L-1 NaCl胁迫24 h后, 沙枣幼苗根系Na+ 和K+ 的稳态离子流; (2) 150 mmol·L-1 NaCl瞬时处理, K+ 和H+ 离子流的动态特征; (3)盐胁迫下, 分别用阿米洛利(Na+ /H+ 逆向转运体抑制剂, Amiloride) 和氯化四乙胺(K+ 通道抑制剂, TEA)处理后, Na+ 和K+ 的离子流特性, 以期从离子动态转运方面探讨沙枣耐盐机制和沙枣种源间K+ /Na+ 平衡的调控差异, 为进一步发掘优良耐盐沙枣种质资源提供理论参考依据。
1 材料和方法
1.1 试验材料
2012年10月下旬, 分别从新疆阿拉尔和宁夏银川采集50株沙枣母树上的成熟果实。处理后, 12月上旬将饱满无虫眼沙枣种子沙藏。次年3月上旬于温室内, 采用四分法, 将沙藏后的种子置于塑料盆中, 并用塑料膜遮盖。第二天(萌芽率达70%), 将沙枣种子播种于营养钵(10 cm × 10 cm, 口径×高)中, 基质为蛭石。期间, 浇洒改良的Hoagland营养液, 注意病虫害防治。当幼苗生长到8-10片叶龄、株高约10 cm时, 取长势基本一致的幼苗, 洗净根部蛭石, 置于塑料花盆(40 cm × 35 cm × 15 cm, 长×宽×高)中开展水培, 80株/种源, 花盆中全天通气。在长出新根之前用自来水培养, 2天换一次水, 之后用1/4 Hoagland营养液培养(蒸馏水配制), 3天换一次营养液。
1.2 试验设计
1.2.1 稳态离子流测定
水培20天后, 每个种源各选取24株大小、长势基本一致的苗木, 其中, 每个种源中16株用含 150 mmol·L-1 NaCl的1/4 Hoagland营养液培养24 h, 8株作为对照。然后进行Na+ 和K+ 的稳态离子流动态变化测定, 测量10 min。
1.2.2 离子动力学测定
水培19天后, 每个种源挑选大小、长势基本一致的沙枣幼苗16株。测量5-6 min后, 加入NaCl溶液(此时测试液中NaCl浓度为150 mmol·L-1 ), 然后继续测定沙枣根尖K+ 和H+ 的动力学离子流30-35 min。
1.2.3 离子流药理学测定
水培20天后, 每个种源选取大小、长势基本一致的沙枣幼苗32株, 其中, 每个种源中24株用含 150 mmol·L-1 NaCl的1/4 Hoagland营养液培养24 h, 8株作为对照。分别用100 µmol·L-1 Amiloride (Kong et al ., 2012 )和20 mmol·L-1 TEA (Wegner et al ., 2011 )处理NaCl胁迫24 h后的根系30 min, 再测定根尖的Na+ 和K+ 的离子流。
1.3 试验方法
1.3.1 离子流测定的前期准备
在美国扬格(旭月北京)测试中心利用NMT (NMT100 Series, Younger USA LLC, Amherst, USA)测定盐胁迫下根尖Na+ 、K+ 和H+ 的离子流特性(孙健, 2011 ; Kong et al ., 2012 )。所用测试液: 0.1 mmol·L-1 KCl, 0.1 mmol·L-1 MgCl2 , 0.1 mmol·L-1 CaCl2 , 0.5 mmol·L-1 NaCl, 0.2 mmol·L-1 Na2 SO4 , 0.3 mmol·L-1 2-N-吗啉代乙烷磺酸二水(MES), pH 6.0。前期试验确定了沙枣幼苗根系的最佳扫描位置距根尖顶端为600 μm (对照)和450 μm (150 mmol·L-1 NaCl)(刘正祥, 2013 )。
1.3.2 离子流测定的实验方案
稳态离子流测定(孙健, 2011 ; 刘正祥, 2013 ): 选取大小、部位基本一致的嫩根, 剪取3-4 cm的完整根尖, 先用去离子水冲洗干净, 再在测试液中平衡30 min。然后, 固定在培养皿中, 定点测量根尖Na+ 和K+ 的离子流, 待离子流基本稳定后, 测量 10 min, 每处理水平测量6条根尖(即6次重复)。
离子动力学测定(孙健, 2011 ; 刘正祥, 2013 ): 剪取3-4 cm的完整根尖, 先用去离子水冲洗干净, 再在测试液中平衡30 min。然后, 固定在培养皿中, 测量K+ 和H+ 的离子流, 待稳定测量5-6 min后, 加入一定量的NaCl溶液(pH 6.0), 使测试液中NaCl浓度为150 mmol·L-1 , 再定点测量K+ 和H+ 的离子流30-35 min, 每个种源测量6条根尖。由于扩散效应, 加入NaCl后的前3 min数据给予剔除。
离子流药理学(Kong et al ., 2012 ): 选取大小、部位基本一致的嫩根, 剪取3-4 cm的完整根尖。先冲洗干净, 然后, 对照和盐胁迫离子流测定用基本测试液平衡30 min, 而用于药理学离子流测定的根尖用含Amiloride或TEA的基本测试液平衡30 min。平衡后, 固定在培养皿中, 测定根尖相应位置(对照为600 μm, NaCl处理为450 μm) Na+ 和K+ 的离子流, 持续测定10 min。
1.4 数据处理与分析
测得的离子流数据先导入Excel表格中, 剔除异常数据, 然后利用Mageflux软件计算离子流速。用Microsoft Excel 2003软件整理数据和制图, 运用SPSS 16.0软件进行单因素方差分析和最小显著性差异法多重比较(p < 0.05)。
2 结果和分析
2.1 NaCl胁迫对沙枣幼苗根系稳态离子流的影响
2.1.1 稳态Na+ 流
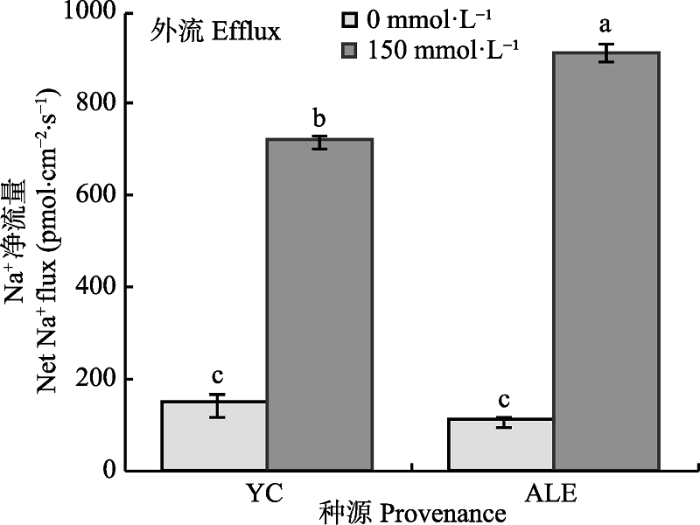
从图1 可以看出, 在对照条件下, 两个种源沙枣幼苗根系的Na+ 流均为外流, 其净流量均低于 200 pmol·cm-2 ·s-1 , 两个种源间无显著性差异, 而在150 mmol·L-1 NaCl胁迫处理24 h后, 与对照相比, 沙枣根系的Na+ 外流的净流量显著增加, 并且两个种源间差异明显。NaCl胁迫下, 阿拉尔种源根系Na+ 外流的净流量显著高于银川种源, 分别为912和720 pmol·cm-2 ·s-1 。由此说明, 盐胁迫下, 沙枣幼苗根系Na+ 的外排能力显著提高, 并且与银川种源相比, 阿拉尔种源沙枣幼苗表现出更强的排Na+ 能力。
图1 NaCl胁迫对两个种源沙枣幼苗根系稳态Na+ 流的影响(平均值±标准误差, n = 4)。ALE, 阿拉尔; YC, 银川。不同小写字母表示不同处理和种源间差异显著(p < 0.05)。
Fig. 1 Effects of NaCl stress on steady-state Na+ flux at apical regions of two provenances of Elaeagnus angustifolia (mean ± SE, n = 4). ALE, Alaer; YC, Yinchuan. Different lowercase letters indicate significant differences between different treatments and provenances (p < 0.05).
2.1.2 稳态K+ 流
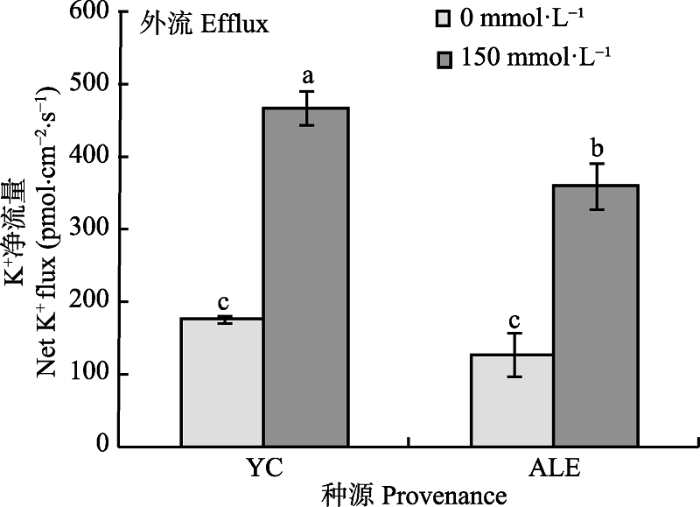
K+ 是植物代谢过程中的重要物质, 限制体内Na+ 积累, 减少体内K+ 流失, 可以提高植物耐盐性(Gouiaa et al ., 2012 )。如图2 所示, 在对照条件下, 银川和阿拉尔两个种源沙枣根系在对照条件下K+ 外流的净流量仅分别为180和159 pmol·cm-2 ·s-1 , 而在150 mmol·L-1 NaCl胁迫下, 两个种源沙枣根系K+ 外流的净流量显著增大, 分别为469和360 pmol·cm-2 ·s-1 ,并且银川种源沙枣幼苗K+ 外流净流量明显高于阿拉尔种源, 这表明NaCl胁迫能够激发沙枣根系中K+ 外流, 并且银川种源沙枣幼苗根系保留K+ 能力弱于阿拉尔种源。
图2 NaCl胁迫对两个种源沙枣幼苗根系稳态K+ 流的影响(平均值±标准误差, n = 4)。ALE, 阿拉尔; YC, 银川。不同小写字母表示不同处理和种源间差异显著(p < 0.05)。
Fig. 2 Effects of NaCl stress on steady-state K+ flux at apical regions of two provenances of Elaeagnus angustifolia (mean ± SE, n = 4). ALE, Alaer; YC, Yinchuan. Different lowercase letters indicate significant differences between different treatments and provenances (p < 0.05).
2.2 NaCl胁迫对不同种源沙枣幼苗根系离子流动力学的影响
2.2.1 瞬时K+ 流的动态特性
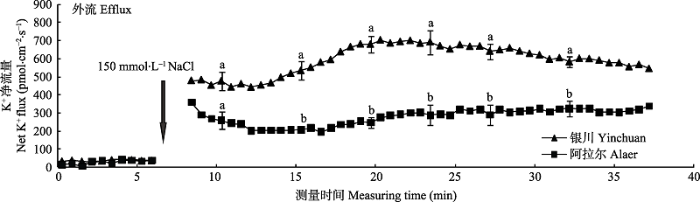
由图3 可知, NaCl胁迫前, 两个种源沙枣幼苗根系K+ 流表现出平稳的轻微外流。在瞬时加入NaCl (150 mmol·L-1 )后, 沙枣幼苗根系K+ 的外流立刻显著增加, 但是外排强度和变化趋势种源间存在差异。加入NaCl后, 银川种源沙枣幼苗根系的K+ 外排始终高于阿拉尔种源, 并且在10-17 min间, 银川种源K+ 的外流逐渐增加, 其最大值为701 pmol·cm-2 ·s-1 , 随后呈缓慢下降趋势, 而NaCl诱导阿拉尔种源K+ 外排在前5 min内呈现出下降趋势, 最终维持在200- 320 pmol·cm-2 ·s-1 。这说明在受到NaCl刺激后, 沙枣幼苗根系的K+ 外流立刻显著增加, 引起K+ 损失, 并且阿拉尔种源沙枣幼苗根系的保留K+ 能力强于银川种源。
图3 NaCl胁迫诱导两个种源沙枣幼苗根系瞬时K+ 流的动态差异(平均值±标准误差, n = 4)。不同小写字母表示同一时间点种源间差异显著(p < 0.05)。
Fig. 3 Differences of NaCl stress on transient K+ kinetics at apical regions of two provenances of Elaeagnus angustifolia (mean ± SE, n = 4). Different lowercase letters indicate significant differences between provenances at the same time (p < 0.05).
2.2.2 瞬时H+ 流动态特性
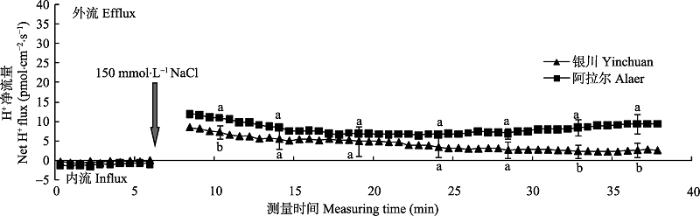
由图4 可知, 两个种源沙枣幼苗根系的H+ 净流量在NaCl胁迫前呈现出轻微的内流, 但NaCl胁迫诱导后, H+ 流马上由内流转为外流, 并且阿拉尔种源的H+ 外流大于银川种源。随着时间的延长, 沙枣幼苗根系的H+ 净流量逐渐下降, 大约胁迫15 min后, H+ 净流量趋于平稳, 但阿拉尔种源根系维持在6.38-9.41 pmol·cm-2 ·s-1 之间, 高于银川种源(2.21- 5.50 pmol·cm-2 ·s-1 ), 并且随着时间延长, 差异越来越明显。由此说明, NaCl胁迫能够诱导沙枣根系的H+ 外流, 并且耐盐型沙枣种源的外流量大于盐敏感型种源, 进而表明耐盐型沙枣种源根系具有更强的质子泵活性, 由此抑制膜电位去极化, 减少K+ 流失。
图4 NaCl胁迫诱导两个种源沙枣幼苗根系瞬时H+ 流的动态差异(平均值±标准误差, n = 4)。不同小写字母表示同一时间点种源间差异显著(p < 0.05)。
Fig. 4 Differences of NaCl stress on transient H+ kinetics at apical regions of two provenances of Elaeagnus angustifolia (mean ± SE, n = 4). Different lowercase letters indicate significant differences between provenances at the same time (p < 0.05).
2.3 NaCl胁迫下两个种源沙枣幼苗根系离子流的药理学特性
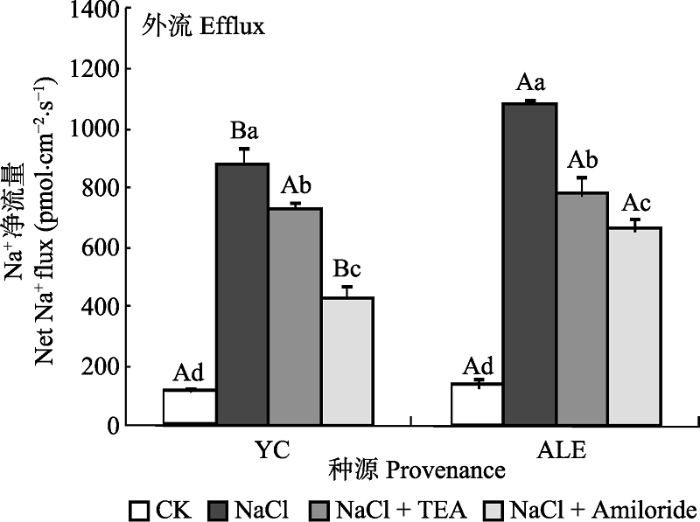
2.3.1 两个种源沙枣幼苗根系在不同处理下的Na+ 流
如图5 所示, 银川种源和阿拉尔种源沙枣幼苗根系在各处理下, Na+ 净流量均为外流, 在对照条件下, 外流量最小, 而NaCl胁迫下最大。在TEA和Amiloride处理后, Na+ 的净流量均显著减少。同时, Amiloride作为Na+ /H+ 逆向转运蛋白抑制剂, 对Na+ 外流的限制作用比K+ 通道蛋白抑制剂TEA显著。在NaCl胁迫下, 阿拉尔种源的Na+ 净流量处于1β000-1β200 pmol·cm-2 ·s-1 之间, 而银川种源则维持在800-1β000 pmol·cm-2 ·s-1 。在NaCl + Amiloride处理后, 银川种源的Na+ 净外流量仍然显著低于阿拉尔种源, 与仅NaCl处理相比, 分别下降了 449 pmol·cm-2 ·s-1 和429 pmol·cm-2 ·s-1 。在NaCl + TEA处理后, 沙枣幼苗根系的Na+ 净流量在阿拉尔和银川种源间差异不明显, 分别为775和727 pmol·cm-2 ·s-1 。进一步说明在NaCl胁迫下, 阿拉尔种源沙枣幼苗质膜Na+ 外排能力强于银川种源。
图5 两种抑制剂对NaCl胁迫后两个种源沙枣根系Na+ 流的影响(平均值±标准误差, n = 4)。ALE, 阿拉尔; YC, 银川。不同小写字母表示同一种源不同处理间差异显著(p < 0.05); 不同大写字母表示相同处理不同种源间差异显著(p < 0.05)。
Fig. 5 Effects of two kinds of inhibitors on net Na+ flux at apical regions of two provenances of Elaeagnus angustifolia treated by NaCl stress (mean ± SE, n = 4). ALE, Alaer; YC, Yinchuan. Different lowercase letters indicate significant differences among different treatments in the same provenance (p < 0.05), While different capital letters indicate significant difference between provenances at the same treatment (p < 0.05).
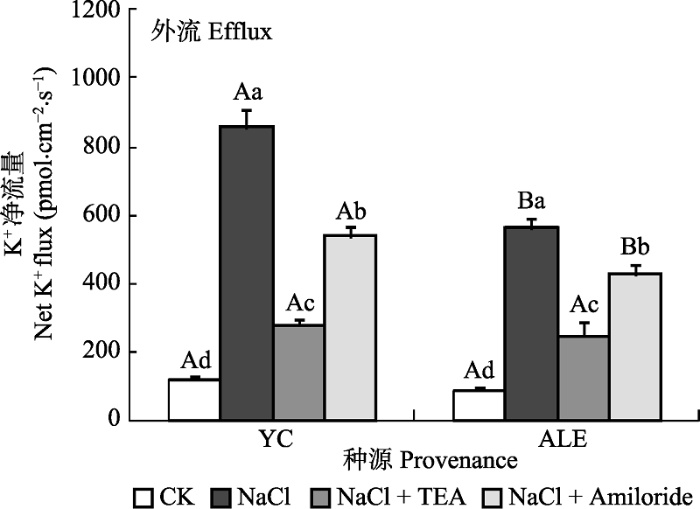
2.3.2 两个种源沙枣幼苗根系在不同处理下的K+ 流
如图6 所示, 在各处理下, 银川和阿拉尔种源沙枣幼苗根系的K+ 净流量均呈现出外流, K+ 净流量在对照条件下分别仅为117和87 pmol·cm-2 ·s-1 , 当NaCl胁迫24 h后, 分别增加到859和560 pmol·cm-2 ·s-1 。当加入抑制剂处理后, K+ 的外流均显著减少, 而Amiloride的抑制效果弱于TEA。在NaCl和NaCl+ Amiloride处理条件下, 银川种源沙枣幼苗根尖的K+ 净流量均明显高于阿拉尔种源, 而在CK和NaCl+ TEA处理下, 两个种源间无明显差异。表明NaCl胁迫环境可以引起沙枣体内K+ 的流失, 银川种源沙枣幼苗根系保留K+ 的能力弱于阿拉尔种源, 并且K+ 通道蛋白和Na+ /H+ 逆向转运体活性能影响质膜上的K+ 运移。
图6 两种抑制剂对NaCl胁迫后两个种源沙枣根系K+ 流的影响(平均值±标准误差, n = 4)。ALE, 阿拉尔; YC, 银川。不同小写字母表示同一种源不同处理间差异显著(p < 0.05); 不同大写字母表示相同处理不同种源间差异显著(p < 0.05)。
Fig. 6 Effects of two kinds of inhibitors on net K+ flux at apical regions of two provenances of Elaeagnus angustifolia treated by NaCl stress (mean ± SE, n = 4). ALE, Alaer; YC, Yinchuan. Different lowercase letters indicate significant differences among different treatments in the same provenance (p < 0.05), While different capital letters indicate significant difference between provenances at the same treatment (p < 0.05).
3 讨论
植物维持细胞溶质中较高的K+ /Na+ 是评判植物耐盐性的一个重要因素, 植物主要通过限制Na+ 吸收, 增加Na+ 外排和区隔化, 以及控制K+ 外流量来实现较高的K+ /Na+ (Maathuis & Amtmann, 1999 ; Chen et al ., 2007a ; Cuin et al ., 2012 )。有研究发现, 耐盐性较强的品种在盐胁迫下具有更强的外排Na+ 能力或保留K+ 能力(Chen et al ., 2007b , 2008 ; Cuin et al ., 2008 ; Sun et al ., 2009 )。但是也有研究显示K+ 外流不能预测不同基因型水稻(Oryza sativa )的耐盐性(Coskun et al ., 2013 )。在本试验中, NaCl胁迫后, 沙枣幼苗根系的稳态Na+ 和K+ 外流量均显著增加, 并且耐盐型沙枣种源的Na+ 净流量明显大于盐敏感型种源, 而K+ 外流的净流量小于盐敏感型种源。这表明NaCl胁迫引起沙枣幼苗Na+ 积累而K+ 流失, 但耐盐型沙枣种源幼苗根系具有更好的Na+ 外排能力和保留K+ 能力。
在高等植物中, 质膜Na+ /H+ 逆向转运蛋白除了负责调节根部Na+ 外排和体内Na+ 长距离运输外, 还可以调控细胞内的K+ 平衡、pH值和Ca2+ 信号转导 (马清等, 2011 ; 孙健, 2011 )。质膜Na+ /H+ 逆向转运体在盐胁迫下被诱导合成或者激活, 进而将Na+ 从细胞质外排到细胞外, 但是排Na+ 需要质膜两侧产生的H+ 电化学势梯度来完成, 而质膜H+ -ATPase利用水解ATP产生的能量将H+ 泵出细胞质能形成H+ 电化学势梯度(Rausch et al ., 1996 ; 陈观平等, 2006 ; 马清等, 2011 )。同时, Olias等(2009)对番茄(Solanum lycopersicum )研究发现质膜Na+ /H+ 逆向转运体(SlSOS1)不仅在NaCl胁迫下维持离子平衡起重要作用, 而且对于Na+ 在各器官间的区隔化也发挥关键作用。Cuin等(2011)研究认为Na+ /H+ 逆向转运体可以作为小麦耐盐性筛选的一个量化指标。大量转基因试验证实质膜Na+ /H+ 逆向转运体的转基因植物也都表现出更强的耐盐能力(Yang et al ., 2009 ; Yue et al ., 2012 ; Feki et al ., 2014 )。在本试验中, NaCl胁迫下, 沙枣幼苗根系Na+ 外排显著增加, 耐盐型沙枣种源的Na+ 外流速率大于盐敏感型种源, 并且瞬时盐胁迫下, H+ 由内流变为外流, 耐盐型种源沙枣幼苗根系的H+ 外流量明显大于盐敏感型种源, 说明耐盐型种源沙枣幼苗在盐胁迫下具有更强的Na+ /H+ 逆向转运体和质膜H+ -ATPase活性。本研究的药理学试验显示, Na+ /H+ 逆向转运体抑制剂Amiloride显著减少Na+ 的外排, 并且阿拉尔种源在NaCl和NaCl+ Amiloride处理下Na+ 净外排量均显著高于银川种源, 这进一步表明质膜Na+ /H+ 逆向转运蛋白在沙枣根尖外排Na+ 过程中起着非常重要的作用, 但是两个种源在抑制量间无显著差异(449 pmol·cm-2 ·s-1 和 429 pmol·cm-2 ·s-1 ), 并且在NaCl + TEA处理后两个种源根尖的Na+ 净流量也无明显差异, 这可能与抑制剂浓度, 处理时间和试验材料有关, 也可能沙枣的排Na+ 机制有其未知的特殊性,有待进一步研究探讨。
另外, 在NaCl胁迫下, 细胞去极化程度会加强, 由此促使通过外向K+ 通道和非选择性阳离子通道外流的K+ 增加(Chen et al ., 2007a ; Shabala & Cuin, 2008 ; Sun et al ., 2009 )。在本试验中, 在NaCl胁迫 24 h和瞬时NaCl胁迫后, K+ 外流量显著增加, 但受到K+ 通道抑制剂TEA的限制, 这与对大麦(Chen et al ., 2007a )的研究结论相同。另外, 在TEA处理后, 两个种源间的K+ 外流差异不明显, 由此推测两个种源的K+ 外流在其他通道途径间无差异, 经K+ 通道而发生的K+ 外流是引起种源间K+ 外流和保持K+ /Na+ 平衡能力差异的一个主要原因。
综上所述, 在NaCl胁迫环境下, 沙枣幼苗为了降低体内Na+ 的积累, 通过质膜Na+ /H+ 逆向转运体将Na+ 排到细胞外, 同时, NaCl胁迫会使细胞去极化程度加强, 增加K+ 外流。阿拉尔种源的沙枣幼苗根系表现出更强的Na+ /H+ 逆向转运体活性, 更多的Na+ 外排, 从而减少Na+ 向叶片组织的运输, 并减少NaCl胁迫下根系细胞中的K+ 外排, 进而更好地保持细胞质中K+ /Na+ 平衡, 由此表现出更强的耐盐能力。
致谢 感谢温州市森林生态科技创新团队项目(C20150002)资助。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Britto DT Kronzucker HJ 2008 ). Cellular mechanisms of potassium transport in plants
.Physiologia Plantarum 133 , 637 -650 .
[本文引用: 1]
[2]
Chao DY Dilkes B Luo H Douqlas A Yakubova E Lahner B Salt DE 2013 ). Polyploids exhibit higher potassium uptake and salinity tolerance in Arabidopsis
. Science , 341 , 658 -659 .
[本文引用: 1]
[3]
Chen GP Wang HZ Shi NN Chen SY 2006 ). Na+/H+ antiporter and its relationship with plant salt tolerance
.China Biotechnology 26 (5 ), 101 -106 . (in Chinese with English abstract)[陈观平 , 王慧中 , 施农农 , 陈受宜 (2006 ). Na+/H+逆向转运蛋白与植物耐盐性的关系研究进展
. 中国生物工程杂志 , 26 (5 ), 101 -106 .]
[本文引用: 1]
[4]
Chen Z Newman I Zhou M Mendham N Zhang G Shabala S 2005 ). Screening plants for salt tolerance by measuring K+ flux: A case study for barely
.Plant, Cell & Environment 28 , 1230 -1246 .
[本文引用: 1]
[5]
Chen Z Pottosin II Cuin TA Fuglsang AT Tester M Jha D Zepeda-Jazo I Zhou M Palmgren MG Newman IA Shabala S 2007 a). Root plasma membrane transporters controlling K+/Na+ homeostasis in salt-stressed barley
.Plant Physiology 145 , 1714 -1725 .
[本文引用: 3]
[6]
Chen Z Shabala S Mendham N Newman I Zhang GP Zhou MX 2008 ). Combining ability of salinity tolerance on the basis of NaCl-induced K flux from roots of barley
.Crop Science 48 , 1382 -1388 .
[本文引用: 1]
[7]
Chen Z Zhou M Newman IA Mendham NJ Zhang GP Shabala S 2007 b). Potassium and sodium relations in salinised barley tissues as a basis of differential salt tolerance
.Functional Plant Biology 34 , 150 -162 .
[本文引用: 1]
[8]
Coskun D Britto DT Jean YK Kabir I Tolay I Torun AA Kronzucker HJ 2013 ). K+ efflux and retention in response to NaCl stress do not predict salt tolerance in contrasting genotypes of rice (Oryza sativa L.)
. PLOS ONE , 8 (2 ), e57767 . doi: 10.1371/journal.pone.0057767 .
[本文引用: 1]
[9]
Cuin TA Betts SA Chalmandrier R Shabala S 2008 ). A root’s ability to retain K+ correlates with salt tolerance in wheat
.Journal of Experimental Botany 59 , 2697 -2706 .
[本文引用: 2]
[10]
Cuin TA Bose J Stefano G Jha D Tester M Mancuso S Shabala S 2011 ). Assessing the role of root plasma membrane and tonoplast Na+/H+ exchangers in salinity tolerance in wheat: In planta quantification methods
.Plant, Cell & Environment 34 , 947 -961 .
[11]
Cuin TA Zhou M Parsons D Shabala S 2012 ). Genetic behaviour of physiological traits conferring cytosolic K+/Na+ homeostasis in wheat
.Plant Biology 14 , 438 -446 .
[本文引用: 1]
[12]
Demidchik V Maathuis FJM 2007 ). Physiological roles of nonselective cation channels in plants: From salt stress to signaling and development
.New Phytologist 175 , 384 -404 .
[本文引用: 1]
[13]
Feki K Quintero FJ Khoudi H Leidi EO Masmoudi K Pardo JM Brini F 2014 ). A constitutively active form of a durum wheat Na+/H+ antiporter SOS1 confers high salt tolerance to transgenic Arabidopsis
. Plant Cell Reportsl Reports , 33 , 277 -288 .
[本文引用: 2]
[14]
Gouiaa S Khoudi H Leidi EO Pardo JM Masmoudi K 2012 ). Expression of wheat Na+/H+ antiporter TNHXS1 and H+-pyrophosphatase TVP1 genes in tobacco from a bicistronic transcriptional unit improves salt tolerance
. Plant Molecular Biology , 79 , 137 -155 .
[本文引用: 1]
[15]
Guan WK Xu N 2012 ). Research situation and resources utilization of Elaeagnus angustifolia
. Anhui Agricultural Science Bulletin , 18 (19 ), 119 -121 . (in Chinese with English abstract)[管文轲 , 徐娜 (2012 ). 沙枣资源利用研究与开发现状述评
. 安徽农学通报 , 18 (19 ), 119 -121 .]
[本文引用: 1]
[16]
Guo LJ Wang YT 2008 ). Conservation research and prospects of Elaeagnus germplasm resources and utilization values
. Chinese Wild Plant Resources , 27 (5 ), 32 -34 . (in Chinese with English abstract)[郭丽君 , 王玉涛 (2008 ). 沙枣种质资源特性及利用价值
. 中国野生植物资源 , 27 (5 ), 32 -34 .]
[本文引用: 1]
[17]
Kong X Luo Z Dong H Eneji AE Li W 2012 ). Effects of non-uniform root zone salinity on water use, Na+ recirculation, and Na+ and H+ flux in cotton
.Journal of Experimental Botany 63 , 2105 -2116 .
[本文引用: 3]
[18]
Liu BY 2007 ). Study on Ecophysiological Response and Ion Distribution of Elaeagnus angustifolia to Salt Stress
. Master degree dissertation, Tianjin Normal University , Tianjin . 27 -53 . (in Chinese with English abstract)[刘宝玉 (2007 ). 盐胁迫下沙枣生理生态响应与离子分配研究
. 硕士学位论文, 天津师范大学, 天津 . 27 -53 .]
[本文引用: 1]
[19]
Liu ZX 2013 ). Physiological Mechanism of Heterogeneous Responses of Elaeagnus angustifolia to NaCl and Na2 SO4 Stress
. PhD dissertation, Chinese Academy of Forestry , Beijing . 100 -104 . (in Chinese with English abstract)[刘正祥 (2013 ). 沙枣对氯化钠和硫酸钠胁迫异质性响应的生理机制
. 博士学位论文, 中国林业科学研究院, 北京 . 100 -104 .]
[本文引用: 3]
[20]
Liu ZX Zhang HX Yang XY Liu T Di WB 2014 ). Growth, and cationic absorption, transportation and allocation of Elaeagnus angustifolia seedlings under NaCl stress
. Acta Ecologica Sinica , 34 , 326 -336 . (in Chinese with English abstract)[刘正祥 , 张华新 , 杨秀艳 , 刘涛 , 狄文彬 (2014 ). NaCl胁迫下沙枣幼苗生长和阳离子吸收、运输与分配特性
. 生态学报 , 34 , 326 -336 .]
[本文引用: 1]
[21]
Ma Q Bao AK Wu GQ Wang SM 2011 ). Plasma membrane Na+/H+ antiporter is involved in plant salt tolerance
.Chinese Bulletin of Botany 46 , 206 -215 . (in Chinese with English abstract)[马清 , 包爱科 , 伍国强 , 王锁民 (2011 ). 质膜Na+/H+逆向转运蛋白与植物耐盐性
. 植物学报 , 46 , 206 -215 .]
[本文引用: 2]
[22]
Maathuis FJM 2006 ). The role of monovalent cation transporters in plant responses to salinity
.Journal of Experimental Botany 57 , 1137 -1147 .
[本文引用: 1]
[23]
Maathuis FJM Amtmann A 1999 ). K+ nutrition and Na+ toxicity: The basis of cellular K+/Na+ ratios
.Annals of Botany 84 , 123 -133 .
[本文引用: 1]
[24]
Olias R Eljakaoui Z Li JU Morales PD Marin-manzano MC Pardo JM Belver A 2009 ). The plasma membrane Na+/H+ antiporter SOS1 is essential for salt tolerance in tomato and affects the partitioning of Na+ between plant organs
.Plant, Cell & Environment 32 , 904 -916 .
[25]
Rausch T Kirsch M Löw R Lehr A Viereck R An ZG 1996 ). Salt stress responses of higher plants: The role of proton pumps and Na+/H+-antiporters
.Journal of Plant Physiology 148 , 425 -433 .
[本文引用: 1]
[26]
Shabala S Cuin TA 2008 ). Potassium transport and plant salt tolerance
.Physiologia Plantarum 133 , 651 -669 .
[本文引用: 2]
[27]
Shi HZ Lee B Wu SJ Zhu JK 2003 ). Overexpression of a plasma membrane Na+/H+ antiporter gene improves salt tolerance in Arabidopsis thaliana
. Nature Biotechnology , 21 , 81 -85 .
[本文引用: 1]
[28]
Shi HZ Ishitani M Kim C Zhu JK 2000 ). The Arabidopsis thaliana salt tolerance gene SOS1 encodes a putative Na+/H+ antiporter
. Proceedings of the National Academy of Sciences of the United States of America , 97 , 6896 -6901 .
[本文引用: 1]
[29]
Sun J 2011 ). Signaling Network in the Perception of Salt Stress and Ionic Homeostasis Regulation in Populus euphratica
. PhD dissertation, Beijing Forestry University , Beijing . 26 -28 . (in Chinese with English abstract)[孙健 (2011 ). 胡杨响应盐胁迫与离子平衡调控信号网络研究
. 博士学位论文, 北京林业大学, 北京 . 26 -28 .]
[本文引用: 4]
[30]
Sun J Chen S Dai S Wang R Li N Shen X Zhou X Lu C Zheng X Hu Z Zhang Z Song J Xu Y 2009 ). NaCl-induced alternations of cellular and tissue ion fluxes in roots of salt-resistant and salt-sensitive poplar species
.Plant Physiology 149 , 1141 -1153 .
[本文引用: 2]
[31]
Wegner LH Stefano G Shabala L Rossi M Mancuso M Shabala S 2011 ). Sequential depolarization of root cortical and stelar cells induced by an acute salt shock—Implications for Na+ and K+ transport into xylem vessels
. Plant, Cell & Environment, 34, 859-869.
[本文引用: 1]
[32]
Yang Q Chen ZZ Zhou XF Yin HB Li X Xin XF Hong XH Zhu JK Gong Z 2009 ). Overexpression of SOS (salt overly sensitive) genes increases salt tolerance in transgenic Arabidopsis
. Molecular Plant , 2 , 22 -31 .
[本文引用: 1]
[33]
Yang S Liu T Zhang HX Li HY Zhang L 2014 ). Growth and physiological characteristics of Elaeagnus angustifolia L. under salt stress
. Journal of Fujian College of Forestry , 34 (1 ), 64 -70 . (in Chinese with English abstract)[杨升 , 刘涛 , 张华新 , 李焕勇 , 张丽 (2014 ). 盐胁迫下沙枣幼苗的生长表现和生理特性
. 福建林学院学报 , 34 (1 ), 64 -70 .]
[本文引用: 2]
[34]
Yang S Zhang HX Liu T Wu HW Yang XY Ni JW Chen QX 2016 ). Study on ion metabolism characteristics of Elaeagnus angustifolia L. seedlings under NaCl stress
. Forest Research , 29 (1 ), 140 -146 . (in Chinese with English abstract)[杨升 , 张华新 , 刘涛 , 武海雯 , 杨秀艳 , 倪建伟 , 陈秋夏 (2016 ). NaCl胁迫下沙枣幼苗的离子代谢特性
. 林业科学研究 , 29 (1 ), 140 -146 .]
[本文引用: 2]
[35]
Yang S Zhang HX Yang XY Chen QX Wu HW 2015 ). Differential growth performance ofElaeagnus angustifolia L. provenances under NaCl stress
. Scientia Silvae Sinicae , 51 (9 ), 51 -58 . (in Chinese with English abstract)[杨升 , 张华新 , 杨秀艳 , 陈秋夏 , 武海雯 (2015 ). NaCl胁迫下不同种源沙枣的生长表现差异
. 林业科学 , 51 (9 ), 51 -58 .]
[本文引用: 1]
[36]
Yue Y Zhang M Zhang J Duan L Li Z 2012 ). SOS1 gene overexpression increased salt tolerance in transgenic tobacco by maintaining a higher K+/Na+ ratio
.Journal of Plant Physiology 169 , 255 -261 .
[本文引用: 1]
[37]
Zhang BZ Cao ZY Zhao KF 1992 ). A study on some physiological properties ofElaeagnus angustifolia under salt stress condition
. Scientia Silvae Sinicae , 28 (2 ), 187 -189 . (in Chinese with English abstract)[张宝泽 , 曹子谊 , 赵可夫 (1992 ). 盐分胁迫下沙枣某些生理特性的研究
. 林业科学 , 28 (2 ), 187 -189 .]
[本文引用: 1]
Cellular mechanisms of potassium transport in plants
1
2008
... 维持体内K+ /Na+ 平衡是植物适应盐胁迫环境的一个关键手段, 利用先进技术手段和转基因技术, 人们对其机制开展了大量有益的探索研究, 发现植物一方面通过限制Na+ 的进入和增加Na+ 的外排或区隔化, 以减少体内或细胞质中的Na+ 浓度, 另一方面限制体内K+ 流失, 从而实现体内K+ /Na+ 平衡(Chao et al ., 2013 ; Feki et al ., 2014 ).早期的研究结果显示细胞质中的Na+ 外排主要由质膜上的Na+ /H+ 逆向转运蛋白(SOS1)负责, 并由H+ -ATPase酶负责提供驱动力, 同时引起细胞膜电位重新极化, 减少Na+ 的流入(Shi et al ., 2000 , 2003 ; Maathuis, 2006 ).另外, K+ 的吸收会受到Na+ 的竞争性抑制, 并且诱导通过外向K+ 通道而外流的K+ 增加(Demidchik & Maathuis, 2007 ; Britto & Kronzucker, 2008 ; Shabala & Cuin, 2008 ).同时, 有研究显示在盐胁迫下大麦(Hordeum vulgare )和小麦(Triticum aestivum )耐盐型品种根系的K+ 外流量均小于盐敏感型品种(Chen et al ., 2005 ; Cuin et al ., 2008 ).因此, 了解植物根系区域Na+ 和K+ 离子流情况, 对于理解植物耐盐能力非常重要. ...
Polyploids exhibit higher potassium uptake and salinity tolerance in Arabidopsis
1
2013
... 维持体内K+ /Na+ 平衡是植物适应盐胁迫环境的一个关键手段, 利用先进技术手段和转基因技术, 人们对其机制开展了大量有益的探索研究, 发现植物一方面通过限制Na+ 的进入和增加Na+ 的外排或区隔化, 以减少体内或细胞质中的Na+ 浓度, 另一方面限制体内K+ 流失, 从而实现体内K+ /Na+ 平衡(Chao et al ., 2013 ; Feki et al ., 2014 ).早期的研究结果显示细胞质中的Na+ 外排主要由质膜上的Na+ /H+ 逆向转运蛋白(SOS1)负责, 并由H+ -ATPase酶负责提供驱动力, 同时引起细胞膜电位重新极化, 减少Na+ 的流入(Shi et al ., 2000 , 2003 ; Maathuis, 2006 ).另外, K+ 的吸收会受到Na+ 的竞争性抑制, 并且诱导通过外向K+ 通道而外流的K+ 增加(Demidchik & Maathuis, 2007 ; Britto & Kronzucker, 2008 ; Shabala & Cuin, 2008 ).同时, 有研究显示在盐胁迫下大麦(Hordeum vulgare )和小麦(Triticum aestivum )耐盐型品种根系的K+ 外流量均小于盐敏感型品种(Chen et al ., 2005 ; Cuin et al ., 2008 ).因此, 了解植物根系区域Na+ 和K+ 离子流情况, 对于理解植物耐盐能力非常重要. ...
Na+/H+逆向转运蛋白与植物耐盐性的关系研究进展
1
2006
... 在高等植物中, 质膜Na+ /H+ 逆向转运蛋白除了负责调节根部Na+ 外排和体内Na+ 长距离运输外, 还可以调控细胞内的K+ 平衡、pH值和Ca2+ 信号转导 (马清等, 2011 ; 孙健, 2011 ).质膜Na+ /H+ 逆向转运体在盐胁迫下被诱导合成或者激活, 进而将Na+ 从细胞质外排到细胞外, 但是排Na+ 需要质膜两侧产生的H+ 电化学势梯度来完成, 而质膜H+ -ATPase利用水解ATP产生的能量将H+ 泵出细胞质能形成H+ 电化学势梯度(Rausch et al ., 1996 ; 陈观平等, 2006 ; 马清等, 2011 ).同时, Olias等(2009)对番茄(Solanum lycopersicum )研究发现质膜Na+ /H+ 逆向转运体(SlSOS1)不仅在NaCl胁迫下维持离子平衡起重要作用, 而且对于Na+ 在各器官间的区隔化也发挥关键作用.Cuin等(2011)研究认为Na+ /H+ 逆向转运体可以作为小麦耐盐性筛选的一个量化指标.大量转基因试验证实质膜Na+ /H+ 逆向转运体的转基因植物也都表现出更强的耐盐能力(Yang et al ., 2009 ; Yue et al ., 2012 ; Feki et al ., 2014 ).在本试验中, NaCl胁迫下, 沙枣幼苗根系Na+ 外排显著增加, 耐盐型沙枣种源的Na+ 外流速率大于盐敏感型种源, 并且瞬时盐胁迫下, H+ 由内流变为外流, 耐盐型种源沙枣幼苗根系的H+ 外流量明显大于盐敏感型种源, 说明耐盐型种源沙枣幼苗在盐胁迫下具有更强的Na+ /H+ 逆向转运体和质膜H+ -ATPase活性.本研究的药理学试验显示, Na+ /H+ 逆向转运体抑制剂Amiloride显著减少Na+ 的外排, 并且阿拉尔种源在NaCl和NaCl+ Amiloride处理下Na+ 净外排量均显著高于银川种源, 这进一步表明质膜Na+ /H+ 逆向转运蛋白在沙枣根尖外排Na+ 过程中起着非常重要的作用, 但是两个种源在抑制量间无显著差异(449 pmol·cm-2 ·s-1 和 429 pmol·cm-2 ·s-1 ), 并且在NaCl + TEA处理后两个种源根尖的Na+ 净流量也无明显差异, 这可能与抑制剂浓度, 处理时间和试验材料有关, 也可能沙枣的排Na+ 机制有其未知的特殊性,有待进一步研究探讨. ...
Screening plants for salt tolerance by measuring K+ flux: A case study for barely
1
2005
... 维持体内K+ /Na+ 平衡是植物适应盐胁迫环境的一个关键手段, 利用先进技术手段和转基因技术, 人们对其机制开展了大量有益的探索研究, 发现植物一方面通过限制Na+ 的进入和增加Na+ 的外排或区隔化, 以减少体内或细胞质中的Na+ 浓度, 另一方面限制体内K+ 流失, 从而实现体内K+ /Na+ 平衡(Chao et al ., 2013 ; Feki et al ., 2014 ).早期的研究结果显示细胞质中的Na+ 外排主要由质膜上的Na+ /H+ 逆向转运蛋白(SOS1)负责, 并由H+ -ATPase酶负责提供驱动力, 同时引起细胞膜电位重新极化, 减少Na+ 的流入(Shi et al ., 2000 , 2003 ; Maathuis, 2006 ).另外, K+ 的吸收会受到Na+ 的竞争性抑制, 并且诱导通过外向K+ 通道而外流的K+ 增加(Demidchik & Maathuis, 2007 ; Britto & Kronzucker, 2008 ; Shabala & Cuin, 2008 ).同时, 有研究显示在盐胁迫下大麦(Hordeum vulgare )和小麦(Triticum aestivum )耐盐型品种根系的K+ 外流量均小于盐敏感型品种(Chen et al ., 2005 ; Cuin et al ., 2008 ).因此, 了解植物根系区域Na+ 和K+ 离子流情况, 对于理解植物耐盐能力非常重要. ...
Root plasma membrane transporters controlling K+/Na+ homeostasis in salt-stressed barley
3
2007
... 植物维持细胞溶质中较高的K+ /Na+ 是评判植物耐盐性的一个重要因素, 植物主要通过限制Na+ 吸收, 增加Na+ 外排和区隔化, 以及控制K+ 外流量来实现较高的K+ /Na+ (Maathuis & Amtmann, 1999 ; Chen et al ., 2007a ; Cuin et al ., 2012 ).有研究发现, 耐盐性较强的品种在盐胁迫下具有更强的外排Na+ 能力或保留K+ 能力(Chen et al ., 2007b , 2008 ; Cuin et al ., 2008 ; Sun et al ., 2009 ).但是也有研究显示K+ 外流不能预测不同基因型水稻(Oryza sativa )的耐盐性(Coskun et al ., 2013 ).在本试验中, NaCl胁迫后, 沙枣幼苗根系的稳态Na+ 和K+ 外流量均显著增加, 并且耐盐型沙枣种源的Na+ 净流量明显大于盐敏感型种源, 而K+ 外流的净流量小于盐敏感型种源.这表明NaCl胁迫引起沙枣幼苗Na+ 积累而K+ 流失, 但耐盐型沙枣种源幼苗根系具有更好的Na+ 外排能力和保留K+ 能力. ...
... 另外, 在NaCl胁迫下, 细胞去极化程度会加强, 由此促使通过外向K+ 通道和非选择性阳离子通道外流的K+ 增加(Chen et al ., 2007a ; Shabala & Cuin, 2008 ; Sun et al ., 2009 ).在本试验中, 在NaCl胁迫 24 h和瞬时NaCl胁迫后, K+ 外流量显著增加, 但受到K+ 通道抑制剂TEA的限制, 这与对大麦(Chen et al ., 2007a )的研究结论相同.另外, 在TEA处理后, 两个种源间的K+ 外流差异不明显, 由此推测两个种源的K+ 外流在其他通道途径间无差异, 经K+ 通道而发生的K+ 外流是引起种源间K+ 外流和保持K+ /Na+ 平衡能力差异的一个主要原因. ...
... 通道抑制剂TEA的限制, 这与对大麦(Chen et al ., 2007a )的研究结论相同.另外, 在TEA处理后, 两个种源间的K+ 外流差异不明显, 由此推测两个种源的K+ 外流在其他通道途径间无差异, 经K+ 通道而发生的K+ 外流是引起种源间K+ 外流和保持K+ /Na+ 平衡能力差异的一个主要原因. ...
Combining ability of salinity tolerance on the basis of NaCl-induced K flux from roots of barley
1
2008
... 植物维持细胞溶质中较高的K+ /Na+ 是评判植物耐盐性的一个重要因素, 植物主要通过限制Na+ 吸收, 增加Na+ 外排和区隔化, 以及控制K+ 外流量来实现较高的K+ /Na+ (Maathuis & Amtmann, 1999 ; Chen et al ., 2007a ; Cuin et al ., 2012 ).有研究发现, 耐盐性较强的品种在盐胁迫下具有更强的外排Na+ 能力或保留K+ 能力(Chen et al ., 2007b , 2008 ; Cuin et al ., 2008 ; Sun et al ., 2009 ).但是也有研究显示K+ 外流不能预测不同基因型水稻(Oryza sativa )的耐盐性(Coskun et al ., 2013 ).在本试验中, NaCl胁迫后, 沙枣幼苗根系的稳态Na+ 和K+ 外流量均显著增加, 并且耐盐型沙枣种源的Na+ 净流量明显大于盐敏感型种源, 而K+ 外流的净流量小于盐敏感型种源.这表明NaCl胁迫引起沙枣幼苗Na+ 积累而K+ 流失, 但耐盐型沙枣种源幼苗根系具有更好的Na+ 外排能力和保留K+ 能力. ...
Potassium and sodium relations in salinised barley tissues as a basis of differential salt tolerance
1
2007
... 植物维持细胞溶质中较高的K+ /Na+ 是评判植物耐盐性的一个重要因素, 植物主要通过限制Na+ 吸收, 增加Na+ 外排和区隔化, 以及控制K+ 外流量来实现较高的K+ /Na+ (Maathuis & Amtmann, 1999 ; Chen et al ., 2007a ; Cuin et al ., 2012 ).有研究发现, 耐盐性较强的品种在盐胁迫下具有更强的外排Na+ 能力或保留K+ 能力(Chen et al ., 2007b , 2008 ; Cuin et al ., 2008 ; Sun et al ., 2009 ).但是也有研究显示K+ 外流不能预测不同基因型水稻(Oryza sativa )的耐盐性(Coskun et al ., 2013 ).在本试验中, NaCl胁迫后, 沙枣幼苗根系的稳态Na+ 和K+ 外流量均显著增加, 并且耐盐型沙枣种源的Na+ 净流量明显大于盐敏感型种源, 而K+ 外流的净流量小于盐敏感型种源.这表明NaCl胁迫引起沙枣幼苗Na+ 积累而K+ 流失, 但耐盐型沙枣种源幼苗根系具有更好的Na+ 外排能力和保留K+ 能力. ...
K+ efflux and retention in response to NaCl stress do not predict salt tolerance in contrasting genotypes of rice (Oryza sativa L.)
1
2013
... 植物维持细胞溶质中较高的K+ /Na+ 是评判植物耐盐性的一个重要因素, 植物主要通过限制Na+ 吸收, 增加Na+ 外排和区隔化, 以及控制K+ 外流量来实现较高的K+ /Na+ (Maathuis & Amtmann, 1999 ; Chen et al ., 2007a ; Cuin et al ., 2012 ).有研究发现, 耐盐性较强的品种在盐胁迫下具有更强的外排Na+ 能力或保留K+ 能力(Chen et al ., 2007b , 2008 ; Cuin et al ., 2008 ; Sun et al ., 2009 ).但是也有研究显示K+ 外流不能预测不同基因型水稻(Oryza sativa )的耐盐性(Coskun et al ., 2013 ).在本试验中, NaCl胁迫后, 沙枣幼苗根系的稳态Na+ 和K+ 外流量均显著增加, 并且耐盐型沙枣种源的Na+ 净流量明显大于盐敏感型种源, 而K+ 外流的净流量小于盐敏感型种源.这表明NaCl胁迫引起沙枣幼苗Na+ 积累而K+ 流失, 但耐盐型沙枣种源幼苗根系具有更好的Na+ 外排能力和保留K+ 能力. ...
A root’s ability to retain K+ correlates with salt tolerance in wheat
2
2008
... 维持体内K+ /Na+ 平衡是植物适应盐胁迫环境的一个关键手段, 利用先进技术手段和转基因技术, 人们对其机制开展了大量有益的探索研究, 发现植物一方面通过限制Na+ 的进入和增加Na+ 的外排或区隔化, 以减少体内或细胞质中的Na+ 浓度, 另一方面限制体内K+ 流失, 从而实现体内K+ /Na+ 平衡(Chao et al ., 2013 ; Feki et al ., 2014 ).早期的研究结果显示细胞质中的Na+ 外排主要由质膜上的Na+ /H+ 逆向转运蛋白(SOS1)负责, 并由H+ -ATPase酶负责提供驱动力, 同时引起细胞膜电位重新极化, 减少Na+ 的流入(Shi et al ., 2000 , 2003 ; Maathuis, 2006 ).另外, K+ 的吸收会受到Na+ 的竞争性抑制, 并且诱导通过外向K+ 通道而外流的K+ 增加(Demidchik & Maathuis, 2007 ; Britto & Kronzucker, 2008 ; Shabala & Cuin, 2008 ).同时, 有研究显示在盐胁迫下大麦(Hordeum vulgare )和小麦(Triticum aestivum )耐盐型品种根系的K+ 外流量均小于盐敏感型品种(Chen et al ., 2005 ; Cuin et al ., 2008 ).因此, 了解植物根系区域Na+ 和K+ 离子流情况, 对于理解植物耐盐能力非常重要. ...
... 植物维持细胞溶质中较高的K+ /Na+ 是评判植物耐盐性的一个重要因素, 植物主要通过限制Na+ 吸收, 增加Na+ 外排和区隔化, 以及控制K+ 外流量来实现较高的K+ /Na+ (Maathuis & Amtmann, 1999 ; Chen et al ., 2007a ; Cuin et al ., 2012 ).有研究发现, 耐盐性较强的品种在盐胁迫下具有更强的外排Na+ 能力或保留K+ 能力(Chen et al ., 2007b , 2008 ; Cuin et al ., 2008 ; Sun et al ., 2009 ).但是也有研究显示K+ 外流不能预测不同基因型水稻(Oryza sativa )的耐盐性(Coskun et al ., 2013 ).在本试验中, NaCl胁迫后, 沙枣幼苗根系的稳态Na+ 和K+ 外流量均显著增加, 并且耐盐型沙枣种源的Na+ 净流量明显大于盐敏感型种源, 而K+ 外流的净流量小于盐敏感型种源.这表明NaCl胁迫引起沙枣幼苗Na+ 积累而K+ 流失, 但耐盐型沙枣种源幼苗根系具有更好的Na+ 外排能力和保留K+ 能力. ...
Assessing the role of root plasma membrane and tonoplast Na+/H+ exchangers in salinity tolerance in wheat: In planta quantification methods
2011
Genetic behaviour of physiological traits conferring cytosolic K+/Na+ homeostasis in wheat
1
2012
... 植物维持细胞溶质中较高的K+ /Na+ 是评判植物耐盐性的一个重要因素, 植物主要通过限制Na+ 吸收, 增加Na+ 外排和区隔化, 以及控制K+ 外流量来实现较高的K+ /Na+ (Maathuis & Amtmann, 1999 ; Chen et al ., 2007a ; Cuin et al ., 2012 ).有研究发现, 耐盐性较强的品种在盐胁迫下具有更强的外排Na+ 能力或保留K+ 能力(Chen et al ., 2007b , 2008 ; Cuin et al ., 2008 ; Sun et al ., 2009 ).但是也有研究显示K+ 外流不能预测不同基因型水稻(Oryza sativa )的耐盐性(Coskun et al ., 2013 ).在本试验中, NaCl胁迫后, 沙枣幼苗根系的稳态Na+ 和K+ 外流量均显著增加, 并且耐盐型沙枣种源的Na+ 净流量明显大于盐敏感型种源, 而K+ 外流的净流量小于盐敏感型种源.这表明NaCl胁迫引起沙枣幼苗Na+ 积累而K+ 流失, 但耐盐型沙枣种源幼苗根系具有更好的Na+ 外排能力和保留K+ 能力. ...
Physiological roles of nonselective cation channels in plants: From salt stress to signaling and development
1
2007
... 维持体内K+ /Na+ 平衡是植物适应盐胁迫环境的一个关键手段, 利用先进技术手段和转基因技术, 人们对其机制开展了大量有益的探索研究, 发现植物一方面通过限制Na+ 的进入和增加Na+ 的外排或区隔化, 以减少体内或细胞质中的Na+ 浓度, 另一方面限制体内K+ 流失, 从而实现体内K+ /Na+ 平衡(Chao et al ., 2013 ; Feki et al ., 2014 ).早期的研究结果显示细胞质中的Na+ 外排主要由质膜上的Na+ /H+ 逆向转运蛋白(SOS1)负责, 并由H+ -ATPase酶负责提供驱动力, 同时引起细胞膜电位重新极化, 减少Na+ 的流入(Shi et al ., 2000 , 2003 ; Maathuis, 2006 ).另外, K+ 的吸收会受到Na+ 的竞争性抑制, 并且诱导通过外向K+ 通道而外流的K+ 增加(Demidchik & Maathuis, 2007 ; Britto & Kronzucker, 2008 ; Shabala & Cuin, 2008 ).同时, 有研究显示在盐胁迫下大麦(Hordeum vulgare )和小麦(Triticum aestivum )耐盐型品种根系的K+ 外流量均小于盐敏感型品种(Chen et al ., 2005 ; Cuin et al ., 2008 ).因此, 了解植物根系区域Na+ 和K+ 离子流情况, 对于理解植物耐盐能力非常重要. ...
A constitutively active form of a durum wheat Na+/H+ antiporter SOS1 confers high salt tolerance to transgenic Arabidopsis
2
2014
... 维持体内K+ /Na+ 平衡是植物适应盐胁迫环境的一个关键手段, 利用先进技术手段和转基因技术, 人们对其机制开展了大量有益的探索研究, 发现植物一方面通过限制Na+ 的进入和增加Na+ 的外排或区隔化, 以减少体内或细胞质中的Na+ 浓度, 另一方面限制体内K+ 流失, 从而实现体内K+ /Na+ 平衡(Chao et al ., 2013 ; Feki et al ., 2014 ).早期的研究结果显示细胞质中的Na+ 外排主要由质膜上的Na+ /H+ 逆向转运蛋白(SOS1)负责, 并由H+ -ATPase酶负责提供驱动力, 同时引起细胞膜电位重新极化, 减少Na+ 的流入(Shi et al ., 2000 , 2003 ; Maathuis, 2006 ).另外, K+ 的吸收会受到Na+ 的竞争性抑制, 并且诱导通过外向K+ 通道而外流的K+ 增加(Demidchik & Maathuis, 2007 ; Britto & Kronzucker, 2008 ; Shabala & Cuin, 2008 ).同时, 有研究显示在盐胁迫下大麦(Hordeum vulgare )和小麦(Triticum aestivum )耐盐型品种根系的K+ 外流量均小于盐敏感型品种(Chen et al ., 2005 ; Cuin et al ., 2008 ).因此, 了解植物根系区域Na+ 和K+ 离子流情况, 对于理解植物耐盐能力非常重要. ...
... 在高等植物中, 质膜Na+ /H+ 逆向转运蛋白除了负责调节根部Na+ 外排和体内Na+ 长距离运输外, 还可以调控细胞内的K+ 平衡、pH值和Ca2+ 信号转导 (马清等, 2011 ; 孙健, 2011 ).质膜Na+ /H+ 逆向转运体在盐胁迫下被诱导合成或者激活, 进而将Na+ 从细胞质外排到细胞外, 但是排Na+ 需要质膜两侧产生的H+ 电化学势梯度来完成, 而质膜H+ -ATPase利用水解ATP产生的能量将H+ 泵出细胞质能形成H+ 电化学势梯度(Rausch et al ., 1996 ; 陈观平等, 2006 ; 马清等, 2011 ).同时, Olias等(2009)对番茄(Solanum lycopersicum )研究发现质膜Na+ /H+ 逆向转运体(SlSOS1)不仅在NaCl胁迫下维持离子平衡起重要作用, 而且对于Na+ 在各器官间的区隔化也发挥关键作用.Cuin等(2011)研究认为Na+ /H+ 逆向转运体可以作为小麦耐盐性筛选的一个量化指标.大量转基因试验证实质膜Na+ /H+ 逆向转运体的转基因植物也都表现出更强的耐盐能力(Yang et al ., 2009 ; Yue et al ., 2012 ; Feki et al ., 2014 ).在本试验中, NaCl胁迫下, 沙枣幼苗根系Na+ 外排显著增加, 耐盐型沙枣种源的Na+ 外流速率大于盐敏感型种源, 并且瞬时盐胁迫下, H+ 由内流变为外流, 耐盐型种源沙枣幼苗根系的H+ 外流量明显大于盐敏感型种源, 说明耐盐型种源沙枣幼苗在盐胁迫下具有更强的Na+ /H+ 逆向转运体和质膜H+ -ATPase活性.本研究的药理学试验显示, Na+ /H+ 逆向转运体抑制剂Amiloride显著减少Na+ 的外排, 并且阿拉尔种源在NaCl和NaCl+ Amiloride处理下Na+ 净外排量均显著高于银川种源, 这进一步表明质膜Na+ /H+ 逆向转运蛋白在沙枣根尖外排Na+ 过程中起着非常重要的作用, 但是两个种源在抑制量间无显著差异(449 pmol·cm-2 ·s-1 和 429 pmol·cm-2 ·s-1 ), 并且在NaCl + TEA处理后两个种源根尖的Na+ 净流量也无明显差异, 这可能与抑制剂浓度, 处理时间和试验材料有关, 也可能沙枣的排Na+ 机制有其未知的特殊性,有待进一步研究探讨. ...
Expression of wheat Na+/H+ antiporter TNHXS1 and H+-pyrophosphatase TVP1 genes in tobacco from a bicistronic transcriptional unit improves salt tolerance
1
2012
... K+ 是植物代谢过程中的重要物质, 限制体内Na+ 积累, 减少体内K+ 流失, 可以提高植物耐盐性(Gouiaa et al ., 2012 ).如图2 所示, 在对照条件下, 银川和阿拉尔两个种源沙枣根系在对照条件下K+ 外流的净流量仅分别为180和159 pmol·cm-2 ·s-1 , 而在150 mmol·L-1 NaCl胁迫下, 两个种源沙枣根系K+ 外流的净流量显著增大, 分别为469和360 pmol·cm-2 ·s-1 ,并且银川种源沙枣幼苗K+ 外流净流量明显高于阿拉尔种源, 这表明NaCl胁迫能够激发沙枣根系中K+ 外流, 并且银川种源沙枣幼苗根系保留K+ 能力弱于阿拉尔种源. ...
沙枣资源利用研究与开发现状述评
1
2012
... 沙枣(Elaeagnus angustifolia )是胡颓子科胡颓子属的落叶小乔木, 主要分布于我国西北五省, 在北京、天津和山东等地也开展了引种栽培, 具有耐旱、耐盐碱和抗风沙的特性, 能提高土壤肥力, 改善土壤环境, 已经被列为半干旱地区和盐土地防护林带的重要树种(郭丽君和王玉涛, 2008 ; 管文轲和徐娜, 2012 ; 杨升等, 2014 ).大量的研究结果显示, 沙枣幼苗在盐胁迫下, 根、茎和叶片中Na+ 含量增加, 而K+ 含量减少(张宝泽等, 1992 ; 刘宝玉, 2007 ; 刘正祥等, 2014 ).前期研究也发现, 沙枣幼苗随着NaCl浓度的增加, 根、茎和叶中的Na+ 含量显著增加, 而K+ 含量显著减少, 并且阿拉尔种源沙枣幼苗与银川种源幼苗间存在显著性差异, 阿拉尔种源沙枣幼苗能积累更多的Na+ 在根中, 而叶片中Na+ 含量较少, K+ 含量在NaCl胁迫下的降幅也低于银川种源(杨升等, 2014 , 2016 ).但是这些结果都是植株在试验中积累下来的一个总体含量, 是静态的, 而研究探讨盐胁迫下Na+ 和K+ 在沙枣体内具有怎样的动态运移机制, 并且比较不同种源间的差异, 有利于进一步理解沙枣的耐盐机制.因此, 本研究选取沙枣银川种源(YC, 盐敏感型)和阿拉尔种源(ALE, 耐盐型)水培苗为试验材料(杨升等, 2015 , 2016 ), 采用非损伤微测技术(non-invasive micro-test technology, NMT)研究: (1)在150 mmol·L-1 NaCl胁迫24 h后, 沙枣幼苗根系Na+ 和K+ 的稳态离子流; (2) 150 mmol·L-1 NaCl瞬时处理, K+ 和H+ 离子流的动态特征; (3)盐胁迫下, 分别用阿米洛利(Na+ /H+ 逆向转运体抑制剂, Amiloride) 和氯化四乙胺(K+ 通道抑制剂, TEA)处理后, Na+ 和K+ 的离子流特性, 以期从离子动态转运方面探讨沙枣耐盐机制和沙枣种源间K+ /Na+ 平衡的调控差异, 为进一步发掘优良耐盐沙枣种质资源提供理论参考依据. ...
沙枣种质资源特性及利用价值
1
2008
... 沙枣(Elaeagnus angustifolia )是胡颓子科胡颓子属的落叶小乔木, 主要分布于我国西北五省, 在北京、天津和山东等地也开展了引种栽培, 具有耐旱、耐盐碱和抗风沙的特性, 能提高土壤肥力, 改善土壤环境, 已经被列为半干旱地区和盐土地防护林带的重要树种(郭丽君和王玉涛, 2008 ; 管文轲和徐娜, 2012 ; 杨升等, 2014 ).大量的研究结果显示, 沙枣幼苗在盐胁迫下, 根、茎和叶片中Na+ 含量增加, 而K+ 含量减少(张宝泽等, 1992 ; 刘宝玉, 2007 ; 刘正祥等, 2014 ).前期研究也发现, 沙枣幼苗随着NaCl浓度的增加, 根、茎和叶中的Na+ 含量显著增加, 而K+ 含量显著减少, 并且阿拉尔种源沙枣幼苗与银川种源幼苗间存在显著性差异, 阿拉尔种源沙枣幼苗能积累更多的Na+ 在根中, 而叶片中Na+ 含量较少, K+ 含量在NaCl胁迫下的降幅也低于银川种源(杨升等, 2014 , 2016 ).但是这些结果都是植株在试验中积累下来的一个总体含量, 是静态的, 而研究探讨盐胁迫下Na+ 和K+ 在沙枣体内具有怎样的动态运移机制, 并且比较不同种源间的差异, 有利于进一步理解沙枣的耐盐机制.因此, 本研究选取沙枣银川种源(YC, 盐敏感型)和阿拉尔种源(ALE, 耐盐型)水培苗为试验材料(杨升等, 2015 , 2016 ), 采用非损伤微测技术(non-invasive micro-test technology, NMT)研究: (1)在150 mmol·L-1 NaCl胁迫24 h后, 沙枣幼苗根系Na+ 和K+ 的稳态离子流; (2) 150 mmol·L-1 NaCl瞬时处理, K+ 和H+ 离子流的动态特征; (3)盐胁迫下, 分别用阿米洛利(Na+ /H+ 逆向转运体抑制剂, Amiloride) 和氯化四乙胺(K+ 通道抑制剂, TEA)处理后, Na+ 和K+ 的离子流特性, 以期从离子动态转运方面探讨沙枣耐盐机制和沙枣种源间K+ /Na+ 平衡的调控差异, 为进一步发掘优良耐盐沙枣种质资源提供理论参考依据. ...
Effects of non-uniform root zone salinity on water use, Na+ recirculation, and Na+ and H+ flux in cotton
3
2012
... 水培20天后, 每个种源选取大小、长势基本一致的沙枣幼苗32株, 其中, 每个种源中24株用含 150 mmol·L-1 NaCl的1/4 Hoagland营养液培养24 h, 8株作为对照.分别用100 µmol·L-1 Amiloride (Kong et al ., 2012 )和20 mmol·L-1 TEA (Wegner et al ., 2011 )处理NaCl胁迫24 h后的根系30 min, 再测定根尖的Na+ 和K+ 的离子流. ...
... 在美国扬格(旭月北京)测试中心利用NMT (NMT100 Series, Younger USA LLC, Amherst, USA)测定盐胁迫下根尖Na+ 、K+ 和H+ 的离子流特性(孙健, 2011 ; Kong et al ., 2012 ).所用测试液: 0.1 mmol·L-1 KCl, 0.1 mmol·L-1 MgCl2 , 0.1 mmol·L-1 CaCl2 , 0.5 mmol·L-1 NaCl, 0.2 mmol·L-1 Na2 SO4 , 0.3 mmol·L-1 2-N-吗啉代乙烷磺酸二水(MES), pH 6.0.前期试验确定了沙枣幼苗根系的最佳扫描位置距根尖顶端为600 μm (对照)和450 μm (150 mmol·L-1 NaCl)(刘正祥, 2013 ). ...
... 离子流药理学(Kong et al ., 2012 ): 选取大小、部位基本一致的嫩根, 剪取3-4 cm的完整根尖.先冲洗干净, 然后, 对照和盐胁迫离子流测定用基本测试液平衡30 min, 而用于药理学离子流测定的根尖用含Amiloride或TEA的基本测试液平衡30 min.平衡后, 固定在培养皿中, 测定根尖相应位置(对照为600 μm, NaCl处理为450 μm) Na+ 和K+ 的离子流, 持续测定10 min. ...
盐胁迫下沙枣生理生态响应与离子分配研究
1
2007
... 沙枣(Elaeagnus angustifolia )是胡颓子科胡颓子属的落叶小乔木, 主要分布于我国西北五省, 在北京、天津和山东等地也开展了引种栽培, 具有耐旱、耐盐碱和抗风沙的特性, 能提高土壤肥力, 改善土壤环境, 已经被列为半干旱地区和盐土地防护林带的重要树种(郭丽君和王玉涛, 2008 ; 管文轲和徐娜, 2012 ; 杨升等, 2014 ).大量的研究结果显示, 沙枣幼苗在盐胁迫下, 根、茎和叶片中Na+ 含量增加, 而K+ 含量减少(张宝泽等, 1992 ; 刘宝玉, 2007 ; 刘正祥等, 2014 ).前期研究也发现, 沙枣幼苗随着NaCl浓度的增加, 根、茎和叶中的Na+ 含量显著增加, 而K+ 含量显著减少, 并且阿拉尔种源沙枣幼苗与银川种源幼苗间存在显著性差异, 阿拉尔种源沙枣幼苗能积累更多的Na+ 在根中, 而叶片中Na+ 含量较少, K+ 含量在NaCl胁迫下的降幅也低于银川种源(杨升等, 2014 , 2016 ).但是这些结果都是植株在试验中积累下来的一个总体含量, 是静态的, 而研究探讨盐胁迫下Na+ 和K+ 在沙枣体内具有怎样的动态运移机制, 并且比较不同种源间的差异, 有利于进一步理解沙枣的耐盐机制.因此, 本研究选取沙枣银川种源(YC, 盐敏感型)和阿拉尔种源(ALE, 耐盐型)水培苗为试验材料(杨升等, 2015 , 2016 ), 采用非损伤微测技术(non-invasive micro-test technology, NMT)研究: (1)在150 mmol·L-1 NaCl胁迫24 h后, 沙枣幼苗根系Na+ 和K+ 的稳态离子流; (2) 150 mmol·L-1 NaCl瞬时处理, K+ 和H+ 离子流的动态特征; (3)盐胁迫下, 分别用阿米洛利(Na+ /H+ 逆向转运体抑制剂, Amiloride) 和氯化四乙胺(K+ 通道抑制剂, TEA)处理后, Na+ 和K+ 的离子流特性, 以期从离子动态转运方面探讨沙枣耐盐机制和沙枣种源间K+ /Na+ 平衡的调控差异, 为进一步发掘优良耐盐沙枣种质资源提供理论参考依据. ...
沙枣对氯化钠和硫酸钠胁迫异质性响应的生理机制
3
2013
... 在美国扬格(旭月北京)测试中心利用NMT (NMT100 Series, Younger USA LLC, Amherst, USA)测定盐胁迫下根尖Na+ 、K+ 和H+ 的离子流特性(孙健, 2011 ; Kong et al ., 2012 ).所用测试液: 0.1 mmol·L-1 KCl, 0.1 mmol·L-1 MgCl2 , 0.1 mmol·L-1 CaCl2 , 0.5 mmol·L-1 NaCl, 0.2 mmol·L-1 Na2 SO4 , 0.3 mmol·L-1 2-N-吗啉代乙烷磺酸二水(MES), pH 6.0.前期试验确定了沙枣幼苗根系的最佳扫描位置距根尖顶端为600 μm (对照)和450 μm (150 mmol·L-1 NaCl)(刘正祥, 2013 ). ...
... 稳态离子流测定(孙健, 2011 ; 刘正祥, 2013 ): 选取大小、部位基本一致的嫩根, 剪取3-4 cm的完整根尖, 先用去离子水冲洗干净, 再在测试液中平衡30 min.然后, 固定在培养皿中, 定点测量根尖Na+ 和K+ 的离子流, 待离子流基本稳定后, 测量 10 min, 每处理水平测量6条根尖(即6次重复). ...
... 离子动力学测定(孙健, 2011 ; 刘正祥, 2013 ): 剪取3-4 cm的完整根尖, 先用去离子水冲洗干净, 再在测试液中平衡30 min.然后, 固定在培养皿中, 测量K+ 和H+ 的离子流, 待稳定测量5-6 min后, 加入一定量的NaCl溶液(pH 6.0), 使测试液中NaCl浓度为150 mmol·L-1 , 再定点测量K+ 和H+ 的离子流30-35 min, 每个种源测量6条根尖.由于扩散效应, 加入NaCl后的前3 min数据给予剔除. ...
NaCl胁迫下沙枣幼苗生长和阳离子吸收、运输与分配特性
1
2014
... 沙枣(Elaeagnus angustifolia )是胡颓子科胡颓子属的落叶小乔木, 主要分布于我国西北五省, 在北京、天津和山东等地也开展了引种栽培, 具有耐旱、耐盐碱和抗风沙的特性, 能提高土壤肥力, 改善土壤环境, 已经被列为半干旱地区和盐土地防护林带的重要树种(郭丽君和王玉涛, 2008 ; 管文轲和徐娜, 2012 ; 杨升等, 2014 ).大量的研究结果显示, 沙枣幼苗在盐胁迫下, 根、茎和叶片中Na+ 含量增加, 而K+ 含量减少(张宝泽等, 1992 ; 刘宝玉, 2007 ; 刘正祥等, 2014 ).前期研究也发现, 沙枣幼苗随着NaCl浓度的增加, 根、茎和叶中的Na+ 含量显著增加, 而K+ 含量显著减少, 并且阿拉尔种源沙枣幼苗与银川种源幼苗间存在显著性差异, 阿拉尔种源沙枣幼苗能积累更多的Na+ 在根中, 而叶片中Na+ 含量较少, K+ 含量在NaCl胁迫下的降幅也低于银川种源(杨升等, 2014 , 2016 ).但是这些结果都是植株在试验中积累下来的一个总体含量, 是静态的, 而研究探讨盐胁迫下Na+ 和K+ 在沙枣体内具有怎样的动态运移机制, 并且比较不同种源间的差异, 有利于进一步理解沙枣的耐盐机制.因此, 本研究选取沙枣银川种源(YC, 盐敏感型)和阿拉尔种源(ALE, 耐盐型)水培苗为试验材料(杨升等, 2015 , 2016 ), 采用非损伤微测技术(non-invasive micro-test technology, NMT)研究: (1)在150 mmol·L-1 NaCl胁迫24 h后, 沙枣幼苗根系Na+ 和K+ 的稳态离子流; (2) 150 mmol·L-1 NaCl瞬时处理, K+ 和H+ 离子流的动态特征; (3)盐胁迫下, 分别用阿米洛利(Na+ /H+ 逆向转运体抑制剂, Amiloride) 和氯化四乙胺(K+ 通道抑制剂, TEA)处理后, Na+ 和K+ 的离子流特性, 以期从离子动态转运方面探讨沙枣耐盐机制和沙枣种源间K+ /Na+ 平衡的调控差异, 为进一步发掘优良耐盐沙枣种质资源提供理论参考依据. ...
质膜Na+/H+逆向转运蛋白与植物耐盐性
2
2011
... 在高等植物中, 质膜Na+ /H+ 逆向转运蛋白除了负责调节根部Na+ 外排和体内Na+ 长距离运输外, 还可以调控细胞内的K+ 平衡、pH值和Ca2+ 信号转导 (马清等, 2011 ; 孙健, 2011 ).质膜Na+ /H+ 逆向转运体在盐胁迫下被诱导合成或者激活, 进而将Na+ 从细胞质外排到细胞外, 但是排Na+ 需要质膜两侧产生的H+ 电化学势梯度来完成, 而质膜H+ -ATPase利用水解ATP产生的能量将H+ 泵出细胞质能形成H+ 电化学势梯度(Rausch et al ., 1996 ; 陈观平等, 2006 ; 马清等, 2011 ).同时, Olias等(2009)对番茄(Solanum lycopersicum )研究发现质膜Na+ /H+ 逆向转运体(SlSOS1)不仅在NaCl胁迫下维持离子平衡起重要作用, 而且对于Na+ 在各器官间的区隔化也发挥关键作用.Cuin等(2011)研究认为Na+ /H+ 逆向转运体可以作为小麦耐盐性筛选的一个量化指标.大量转基因试验证实质膜Na+ /H+ 逆向转运体的转基因植物也都表现出更强的耐盐能力(Yang et al ., 2009 ; Yue et al ., 2012 ; Feki et al ., 2014 ).在本试验中, NaCl胁迫下, 沙枣幼苗根系Na+ 外排显著增加, 耐盐型沙枣种源的Na+ 外流速率大于盐敏感型种源, 并且瞬时盐胁迫下, H+ 由内流变为外流, 耐盐型种源沙枣幼苗根系的H+ 外流量明显大于盐敏感型种源, 说明耐盐型种源沙枣幼苗在盐胁迫下具有更强的Na+ /H+ 逆向转运体和质膜H+ -ATPase活性.本研究的药理学试验显示, Na+ /H+ 逆向转运体抑制剂Amiloride显著减少Na+ 的外排, 并且阿拉尔种源在NaCl和NaCl+ Amiloride处理下Na+ 净外排量均显著高于银川种源, 这进一步表明质膜Na+ /H+ 逆向转运蛋白在沙枣根尖外排Na+ 过程中起着非常重要的作用, 但是两个种源在抑制量间无显著差异(449 pmol·cm-2 ·s-1 和 429 pmol·cm-2 ·s-1 ), 并且在NaCl + TEA处理后两个种源根尖的Na+ 净流量也无明显差异, 这可能与抑制剂浓度, 处理时间和试验材料有关, 也可能沙枣的排Na+ 机制有其未知的特殊性,有待进一步研究探讨. ...
... ; 马清等, 2011 ).同时, Olias等(2009)对番茄(Solanum lycopersicum )研究发现质膜Na+ /H+ 逆向转运体(SlSOS1)不仅在NaCl胁迫下维持离子平衡起重要作用, 而且对于Na+ 在各器官间的区隔化也发挥关键作用.Cuin等(2011)研究认为Na+ /H+ 逆向转运体可以作为小麦耐盐性筛选的一个量化指标.大量转基因试验证实质膜Na+ /H+ 逆向转运体的转基因植物也都表现出更强的耐盐能力(Yang et al ., 2009 ; Yue et al ., 2012 ; Feki et al ., 2014 ).在本试验中, NaCl胁迫下, 沙枣幼苗根系Na+ 外排显著增加, 耐盐型沙枣种源的Na+ 外流速率大于盐敏感型种源, 并且瞬时盐胁迫下, H+ 由内流变为外流, 耐盐型种源沙枣幼苗根系的H+ 外流量明显大于盐敏感型种源, 说明耐盐型种源沙枣幼苗在盐胁迫下具有更强的Na+ /H+ 逆向转运体和质膜H+ -ATPase活性.本研究的药理学试验显示, Na+ /H+ 逆向转运体抑制剂Amiloride显著减少Na+ 的外排, 并且阿拉尔种源在NaCl和NaCl+ Amiloride处理下Na+ 净外排量均显著高于银川种源, 这进一步表明质膜Na+ /H+ 逆向转运蛋白在沙枣根尖外排Na+ 过程中起着非常重要的作用, 但是两个种源在抑制量间无显著差异(449 pmol·cm-2 ·s-1 和 429 pmol·cm-2 ·s-1 ), 并且在NaCl + TEA处理后两个种源根尖的Na+ 净流量也无明显差异, 这可能与抑制剂浓度, 处理时间和试验材料有关, 也可能沙枣的排Na+ 机制有其未知的特殊性,有待进一步研究探讨. ...
The role of monovalent cation transporters in plant responses to salinity
1
2006
... 维持体内K+ /Na+ 平衡是植物适应盐胁迫环境的一个关键手段, 利用先进技术手段和转基因技术, 人们对其机制开展了大量有益的探索研究, 发现植物一方面通过限制Na+ 的进入和增加Na+ 的外排或区隔化, 以减少体内或细胞质中的Na+ 浓度, 另一方面限制体内K+ 流失, 从而实现体内K+ /Na+ 平衡(Chao et al ., 2013 ; Feki et al ., 2014 ).早期的研究结果显示细胞质中的Na+ 外排主要由质膜上的Na+ /H+ 逆向转运蛋白(SOS1)负责, 并由H+ -ATPase酶负责提供驱动力, 同时引起细胞膜电位重新极化, 减少Na+ 的流入(Shi et al ., 2000 , 2003 ; Maathuis, 2006 ).另外, K+ 的吸收会受到Na+ 的竞争性抑制, 并且诱导通过外向K+ 通道而外流的K+ 增加(Demidchik & Maathuis, 2007 ; Britto & Kronzucker, 2008 ; Shabala & Cuin, 2008 ).同时, 有研究显示在盐胁迫下大麦(Hordeum vulgare )和小麦(Triticum aestivum )耐盐型品种根系的K+ 外流量均小于盐敏感型品种(Chen et al ., 2005 ; Cuin et al ., 2008 ).因此, 了解植物根系区域Na+ 和K+ 离子流情况, 对于理解植物耐盐能力非常重要. ...
K+ nutrition and Na+ toxicity: The basis of cellular K+/Na+ ratios
1
1999
... 植物维持细胞溶质中较高的K+ /Na+ 是评判植物耐盐性的一个重要因素, 植物主要通过限制Na+ 吸收, 增加Na+ 外排和区隔化, 以及控制K+ 外流量来实现较高的K+ /Na+ (Maathuis & Amtmann, 1999 ; Chen et al ., 2007a ; Cuin et al ., 2012 ).有研究发现, 耐盐性较强的品种在盐胁迫下具有更强的外排Na+ 能力或保留K+ 能力(Chen et al ., 2007b , 2008 ; Cuin et al ., 2008 ; Sun et al ., 2009 ).但是也有研究显示K+ 外流不能预测不同基因型水稻(Oryza sativa )的耐盐性(Coskun et al ., 2013 ).在本试验中, NaCl胁迫后, 沙枣幼苗根系的稳态Na+ 和K+ 外流量均显著增加, 并且耐盐型沙枣种源的Na+ 净流量明显大于盐敏感型种源, 而K+ 外流的净流量小于盐敏感型种源.这表明NaCl胁迫引起沙枣幼苗Na+ 积累而K+ 流失, 但耐盐型沙枣种源幼苗根系具有更好的Na+ 外排能力和保留K+ 能力. ...
The plasma membrane Na+/H+ antiporter SOS1 is essential for salt tolerance in tomato and affects the partitioning of Na+ between plant organs
2009
Salt stress responses of higher plants: The role of proton pumps and Na+/H+-antiporters
1
1996
... 在高等植物中, 质膜Na+ /H+ 逆向转运蛋白除了负责调节根部Na+ 外排和体内Na+ 长距离运输外, 还可以调控细胞内的K+ 平衡、pH值和Ca2+ 信号转导 (马清等, 2011 ; 孙健, 2011 ).质膜Na+ /H+ 逆向转运体在盐胁迫下被诱导合成或者激活, 进而将Na+ 从细胞质外排到细胞外, 但是排Na+ 需要质膜两侧产生的H+ 电化学势梯度来完成, 而质膜H+ -ATPase利用水解ATP产生的能量将H+ 泵出细胞质能形成H+ 电化学势梯度(Rausch et al ., 1996 ; 陈观平等, 2006 ; 马清等, 2011 ).同时, Olias等(2009)对番茄(Solanum lycopersicum )研究发现质膜Na+ /H+ 逆向转运体(SlSOS1)不仅在NaCl胁迫下维持离子平衡起重要作用, 而且对于Na+ 在各器官间的区隔化也发挥关键作用.Cuin等(2011)研究认为Na+ /H+ 逆向转运体可以作为小麦耐盐性筛选的一个量化指标.大量转基因试验证实质膜Na+ /H+ 逆向转运体的转基因植物也都表现出更强的耐盐能力(Yang et al ., 2009 ; Yue et al ., 2012 ; Feki et al ., 2014 ).在本试验中, NaCl胁迫下, 沙枣幼苗根系Na+ 外排显著增加, 耐盐型沙枣种源的Na+ 外流速率大于盐敏感型种源, 并且瞬时盐胁迫下, H+ 由内流变为外流, 耐盐型种源沙枣幼苗根系的H+ 外流量明显大于盐敏感型种源, 说明耐盐型种源沙枣幼苗在盐胁迫下具有更强的Na+ /H+ 逆向转运体和质膜H+ -ATPase活性.本研究的药理学试验显示, Na+ /H+ 逆向转运体抑制剂Amiloride显著减少Na+ 的外排, 并且阿拉尔种源在NaCl和NaCl+ Amiloride处理下Na+ 净外排量均显著高于银川种源, 这进一步表明质膜Na+ /H+ 逆向转运蛋白在沙枣根尖外排Na+ 过程中起着非常重要的作用, 但是两个种源在抑制量间无显著差异(449 pmol·cm-2 ·s-1 和 429 pmol·cm-2 ·s-1 ), 并且在NaCl + TEA处理后两个种源根尖的Na+ 净流量也无明显差异, 这可能与抑制剂浓度, 处理时间和试验材料有关, 也可能沙枣的排Na+ 机制有其未知的特殊性,有待进一步研究探讨. ...
Potassium transport and plant salt tolerance
2
2008
... 维持体内K+ /Na+ 平衡是植物适应盐胁迫环境的一个关键手段, 利用先进技术手段和转基因技术, 人们对其机制开展了大量有益的探索研究, 发现植物一方面通过限制Na+ 的进入和增加Na+ 的外排或区隔化, 以减少体内或细胞质中的Na+ 浓度, 另一方面限制体内K+ 流失, 从而实现体内K+ /Na+ 平衡(Chao et al ., 2013 ; Feki et al ., 2014 ).早期的研究结果显示细胞质中的Na+ 外排主要由质膜上的Na+ /H+ 逆向转运蛋白(SOS1)负责, 并由H+ -ATPase酶负责提供驱动力, 同时引起细胞膜电位重新极化, 减少Na+ 的流入(Shi et al ., 2000 , 2003 ; Maathuis, 2006 ).另外, K+ 的吸收会受到Na+ 的竞争性抑制, 并且诱导通过外向K+ 通道而外流的K+ 增加(Demidchik & Maathuis, 2007 ; Britto & Kronzucker, 2008 ; Shabala & Cuin, 2008 ).同时, 有研究显示在盐胁迫下大麦(Hordeum vulgare )和小麦(Triticum aestivum )耐盐型品种根系的K+ 外流量均小于盐敏感型品种(Chen et al ., 2005 ; Cuin et al ., 2008 ).因此, 了解植物根系区域Na+ 和K+ 离子流情况, 对于理解植物耐盐能力非常重要. ...
... 另外, 在NaCl胁迫下, 细胞去极化程度会加强, 由此促使通过外向K+ 通道和非选择性阳离子通道外流的K+ 增加(Chen et al ., 2007a ; Shabala & Cuin, 2008 ; Sun et al ., 2009 ).在本试验中, 在NaCl胁迫 24 h和瞬时NaCl胁迫后, K+ 外流量显著增加, 但受到K+ 通道抑制剂TEA的限制, 这与对大麦(Chen et al ., 2007a )的研究结论相同.另外, 在TEA处理后, 两个种源间的K+ 外流差异不明显, 由此推测两个种源的K+ 外流在其他通道途径间无差异, 经K+ 通道而发生的K+ 外流是引起种源间K+ 外流和保持K+ /Na+ 平衡能力差异的一个主要原因. ...
Overexpression of a plasma membrane Na+/H+ antiporter gene improves salt tolerance in Arabidopsis thaliana
1
2003
... 维持体内K+ /Na+ 平衡是植物适应盐胁迫环境的一个关键手段, 利用先进技术手段和转基因技术, 人们对其机制开展了大量有益的探索研究, 发现植物一方面通过限制Na+ 的进入和增加Na+ 的外排或区隔化, 以减少体内或细胞质中的Na+ 浓度, 另一方面限制体内K+ 流失, 从而实现体内K+ /Na+ 平衡(Chao et al ., 2013 ; Feki et al ., 2014 ).早期的研究结果显示细胞质中的Na+ 外排主要由质膜上的Na+ /H+ 逆向转运蛋白(SOS1)负责, 并由H+ -ATPase酶负责提供驱动力, 同时引起细胞膜电位重新极化, 减少Na+ 的流入(Shi et al ., 2000 , 2003 ; Maathuis, 2006 ).另外, K+ 的吸收会受到Na+ 的竞争性抑制, 并且诱导通过外向K+ 通道而外流的K+ 增加(Demidchik & Maathuis, 2007 ; Britto & Kronzucker, 2008 ; Shabala & Cuin, 2008 ).同时, 有研究显示在盐胁迫下大麦(Hordeum vulgare )和小麦(Triticum aestivum )耐盐型品种根系的K+ 外流量均小于盐敏感型品种(Chen et al ., 2005 ; Cuin et al ., 2008 ).因此, 了解植物根系区域Na+ 和K+ 离子流情况, 对于理解植物耐盐能力非常重要. ...
The Arabidopsis thaliana salt tolerance gene SOS1 encodes a putative Na+/H+ antiporter
1
2000
... 维持体内K+ /Na+ 平衡是植物适应盐胁迫环境的一个关键手段, 利用先进技术手段和转基因技术, 人们对其机制开展了大量有益的探索研究, 发现植物一方面通过限制Na+ 的进入和增加Na+ 的外排或区隔化, 以减少体内或细胞质中的Na+ 浓度, 另一方面限制体内K+ 流失, 从而实现体内K+ /Na+ 平衡(Chao et al ., 2013 ; Feki et al ., 2014 ).早期的研究结果显示细胞质中的Na+ 外排主要由质膜上的Na+ /H+ 逆向转运蛋白(SOS1)负责, 并由H+ -ATPase酶负责提供驱动力, 同时引起细胞膜电位重新极化, 减少Na+ 的流入(Shi et al ., 2000 , 2003 ; Maathuis, 2006 ).另外, K+ 的吸收会受到Na+ 的竞争性抑制, 并且诱导通过外向K+ 通道而外流的K+ 增加(Demidchik & Maathuis, 2007 ; Britto & Kronzucker, 2008 ; Shabala & Cuin, 2008 ).同时, 有研究显示在盐胁迫下大麦(Hordeum vulgare )和小麦(Triticum aestivum )耐盐型品种根系的K+ 外流量均小于盐敏感型品种(Chen et al ., 2005 ; Cuin et al ., 2008 ).因此, 了解植物根系区域Na+ 和K+ 离子流情况, 对于理解植物耐盐能力非常重要. ...
胡杨响应盐胁迫与离子平衡调控信号网络研究
4
2011
... 在美国扬格(旭月北京)测试中心利用NMT (NMT100 Series, Younger USA LLC, Amherst, USA)测定盐胁迫下根尖Na+ 、K+ 和H+ 的离子流特性(孙健, 2011 ; Kong et al ., 2012 ).所用测试液: 0.1 mmol·L-1 KCl, 0.1 mmol·L-1 MgCl2 , 0.1 mmol·L-1 CaCl2 , 0.5 mmol·L-1 NaCl, 0.2 mmol·L-1 Na2 SO4 , 0.3 mmol·L-1 2-N-吗啉代乙烷磺酸二水(MES), pH 6.0.前期试验确定了沙枣幼苗根系的最佳扫描位置距根尖顶端为600 μm (对照)和450 μm (150 mmol·L-1 NaCl)(刘正祥, 2013 ). ...
... 稳态离子流测定(孙健, 2011 ; 刘正祥, 2013 ): 选取大小、部位基本一致的嫩根, 剪取3-4 cm的完整根尖, 先用去离子水冲洗干净, 再在测试液中平衡30 min.然后, 固定在培养皿中, 定点测量根尖Na+ 和K+ 的离子流, 待离子流基本稳定后, 测量 10 min, 每处理水平测量6条根尖(即6次重复). ...
... 离子动力学测定(孙健, 2011 ; 刘正祥, 2013 ): 剪取3-4 cm的完整根尖, 先用去离子水冲洗干净, 再在测试液中平衡30 min.然后, 固定在培养皿中, 测量K+ 和H+ 的离子流, 待稳定测量5-6 min后, 加入一定量的NaCl溶液(pH 6.0), 使测试液中NaCl浓度为150 mmol·L-1 , 再定点测量K+ 和H+ 的离子流30-35 min, 每个种源测量6条根尖.由于扩散效应, 加入NaCl后的前3 min数据给予剔除. ...
... 在高等植物中, 质膜Na+ /H+ 逆向转运蛋白除了负责调节根部Na+ 外排和体内Na+ 长距离运输外, 还可以调控细胞内的K+ 平衡、pH值和Ca2+ 信号转导 (马清等, 2011 ; 孙健, 2011 ).质膜Na+ /H+ 逆向转运体在盐胁迫下被诱导合成或者激活, 进而将Na+ 从细胞质外排到细胞外, 但是排Na+ 需要质膜两侧产生的H+ 电化学势梯度来完成, 而质膜H+ -ATPase利用水解ATP产生的能量将H+ 泵出细胞质能形成H+ 电化学势梯度(Rausch et al ., 1996 ; 陈观平等, 2006 ; 马清等, 2011 ).同时, Olias等(2009)对番茄(Solanum lycopersicum )研究发现质膜Na+ /H+ 逆向转运体(SlSOS1)不仅在NaCl胁迫下维持离子平衡起重要作用, 而且对于Na+ 在各器官间的区隔化也发挥关键作用.Cuin等(2011)研究认为Na+ /H+ 逆向转运体可以作为小麦耐盐性筛选的一个量化指标.大量转基因试验证实质膜Na+ /H+ 逆向转运体的转基因植物也都表现出更强的耐盐能力(Yang et al ., 2009 ; Yue et al ., 2012 ; Feki et al ., 2014 ).在本试验中, NaCl胁迫下, 沙枣幼苗根系Na+ 外排显著增加, 耐盐型沙枣种源的Na+ 外流速率大于盐敏感型种源, 并且瞬时盐胁迫下, H+ 由内流变为外流, 耐盐型种源沙枣幼苗根系的H+ 外流量明显大于盐敏感型种源, 说明耐盐型种源沙枣幼苗在盐胁迫下具有更强的Na+ /H+ 逆向转运体和质膜H+ -ATPase活性.本研究的药理学试验显示, Na+ /H+ 逆向转运体抑制剂Amiloride显著减少Na+ 的外排, 并且阿拉尔种源在NaCl和NaCl+ Amiloride处理下Na+ 净外排量均显著高于银川种源, 这进一步表明质膜Na+ /H+ 逆向转运蛋白在沙枣根尖外排Na+ 过程中起着非常重要的作用, 但是两个种源在抑制量间无显著差异(449 pmol·cm-2 ·s-1 和 429 pmol·cm-2 ·s-1 ), 并且在NaCl + TEA处理后两个种源根尖的Na+ 净流量也无明显差异, 这可能与抑制剂浓度, 处理时间和试验材料有关, 也可能沙枣的排Na+ 机制有其未知的特殊性,有待进一步研究探讨. ...
NaCl-induced alternations of cellular and tissue ion fluxes in roots of salt-resistant and salt-sensitive poplar species
2
2009
... 植物维持细胞溶质中较高的K+ /Na+ 是评判植物耐盐性的一个重要因素, 植物主要通过限制Na+ 吸收, 增加Na+ 外排和区隔化, 以及控制K+ 外流量来实现较高的K+ /Na+ (Maathuis & Amtmann, 1999 ; Chen et al ., 2007a ; Cuin et al ., 2012 ).有研究发现, 耐盐性较强的品种在盐胁迫下具有更强的外排Na+ 能力或保留K+ 能力(Chen et al ., 2007b , 2008 ; Cuin et al ., 2008 ; Sun et al ., 2009 ).但是也有研究显示K+ 外流不能预测不同基因型水稻(Oryza sativa )的耐盐性(Coskun et al ., 2013 ).在本试验中, NaCl胁迫后, 沙枣幼苗根系的稳态Na+ 和K+ 外流量均显著增加, 并且耐盐型沙枣种源的Na+ 净流量明显大于盐敏感型种源, 而K+ 外流的净流量小于盐敏感型种源.这表明NaCl胁迫引起沙枣幼苗Na+ 积累而K+ 流失, 但耐盐型沙枣种源幼苗根系具有更好的Na+ 外排能力和保留K+ 能力. ...
... 另外, 在NaCl胁迫下, 细胞去极化程度会加强, 由此促使通过外向K+ 通道和非选择性阳离子通道外流的K+ 增加(Chen et al ., 2007a ; Shabala & Cuin, 2008 ; Sun et al ., 2009 ).在本试验中, 在NaCl胁迫 24 h和瞬时NaCl胁迫后, K+ 外流量显著增加, 但受到K+ 通道抑制剂TEA的限制, 这与对大麦(Chen et al ., 2007a )的研究结论相同.另外, 在TEA处理后, 两个种源间的K+ 外流差异不明显, 由此推测两个种源的K+ 外流在其他通道途径间无差异, 经K+ 通道而发生的K+ 外流是引起种源间K+ 外流和保持K+ /Na+ 平衡能力差异的一个主要原因. ...
Sequential depolarization of root cortical and stelar cells induced by an acute salt shock—Implications for Na+ and K+ transport into xylem vessels
1
2011
... 水培20天后, 每个种源选取大小、长势基本一致的沙枣幼苗32株, 其中, 每个种源中24株用含 150 mmol·L-1 NaCl的1/4 Hoagland营养液培养24 h, 8株作为对照.分别用100 µmol·L-1 Amiloride (Kong et al ., 2012 )和20 mmol·L-1 TEA (Wegner et al ., 2011 )处理NaCl胁迫24 h后的根系30 min, 再测定根尖的Na+ 和K+ 的离子流. ...
Overexpression of SOS (salt overly sensitive) genes increases salt tolerance in transgenic Arabidopsis
1
2009
... 在高等植物中, 质膜Na+ /H+ 逆向转运蛋白除了负责调节根部Na+ 外排和体内Na+ 长距离运输外, 还可以调控细胞内的K+ 平衡、pH值和Ca2+ 信号转导 (马清等, 2011 ; 孙健, 2011 ).质膜Na+ /H+ 逆向转运体在盐胁迫下被诱导合成或者激活, 进而将Na+ 从细胞质外排到细胞外, 但是排Na+ 需要质膜两侧产生的H+ 电化学势梯度来完成, 而质膜H+ -ATPase利用水解ATP产生的能量将H+ 泵出细胞质能形成H+ 电化学势梯度(Rausch et al ., 1996 ; 陈观平等, 2006 ; 马清等, 2011 ).同时, Olias等(2009)对番茄(Solanum lycopersicum )研究发现质膜Na+ /H+ 逆向转运体(SlSOS1)不仅在NaCl胁迫下维持离子平衡起重要作用, 而且对于Na+ 在各器官间的区隔化也发挥关键作用.Cuin等(2011)研究认为Na+ /H+ 逆向转运体可以作为小麦耐盐性筛选的一个量化指标.大量转基因试验证实质膜Na+ /H+ 逆向转运体的转基因植物也都表现出更强的耐盐能力(Yang et al ., 2009 ; Yue et al ., 2012 ; Feki et al ., 2014 ).在本试验中, NaCl胁迫下, 沙枣幼苗根系Na+ 外排显著增加, 耐盐型沙枣种源的Na+ 外流速率大于盐敏感型种源, 并且瞬时盐胁迫下, H+ 由内流变为外流, 耐盐型种源沙枣幼苗根系的H+ 外流量明显大于盐敏感型种源, 说明耐盐型种源沙枣幼苗在盐胁迫下具有更强的Na+ /H+ 逆向转运体和质膜H+ -ATPase活性.本研究的药理学试验显示, Na+ /H+ 逆向转运体抑制剂Amiloride显著减少Na+ 的外排, 并且阿拉尔种源在NaCl和NaCl+ Amiloride处理下Na+ 净外排量均显著高于银川种源, 这进一步表明质膜Na+ /H+ 逆向转运蛋白在沙枣根尖外排Na+ 过程中起着非常重要的作用, 但是两个种源在抑制量间无显著差异(449 pmol·cm-2 ·s-1 和 429 pmol·cm-2 ·s-1 ), 并且在NaCl + TEA处理后两个种源根尖的Na+ 净流量也无明显差异, 这可能与抑制剂浓度, 处理时间和试验材料有关, 也可能沙枣的排Na+ 机制有其未知的特殊性,有待进一步研究探讨. ...
盐胁迫下沙枣幼苗的生长表现和生理特性
2
2014
... 沙枣(Elaeagnus angustifolia )是胡颓子科胡颓子属的落叶小乔木, 主要分布于我国西北五省, 在北京、天津和山东等地也开展了引种栽培, 具有耐旱、耐盐碱和抗风沙的特性, 能提高土壤肥力, 改善土壤环境, 已经被列为半干旱地区和盐土地防护林带的重要树种(郭丽君和王玉涛, 2008 ; 管文轲和徐娜, 2012 ; 杨升等, 2014 ).大量的研究结果显示, 沙枣幼苗在盐胁迫下, 根、茎和叶片中Na+ 含量增加, 而K+ 含量减少(张宝泽等, 1992 ; 刘宝玉, 2007 ; 刘正祥等, 2014 ).前期研究也发现, 沙枣幼苗随着NaCl浓度的增加, 根、茎和叶中的Na+ 含量显著增加, 而K+ 含量显著减少, 并且阿拉尔种源沙枣幼苗与银川种源幼苗间存在显著性差异, 阿拉尔种源沙枣幼苗能积累更多的Na+ 在根中, 而叶片中Na+ 含量较少, K+ 含量在NaCl胁迫下的降幅也低于银川种源(杨升等, 2014 , 2016 ).但是这些结果都是植株在试验中积累下来的一个总体含量, 是静态的, 而研究探讨盐胁迫下Na+ 和K+ 在沙枣体内具有怎样的动态运移机制, 并且比较不同种源间的差异, 有利于进一步理解沙枣的耐盐机制.因此, 本研究选取沙枣银川种源(YC, 盐敏感型)和阿拉尔种源(ALE, 耐盐型)水培苗为试验材料(杨升等, 2015 , 2016 ), 采用非损伤微测技术(non-invasive micro-test technology, NMT)研究: (1)在150 mmol·L-1 NaCl胁迫24 h后, 沙枣幼苗根系Na+ 和K+ 的稳态离子流; (2) 150 mmol·L-1 NaCl瞬时处理, K+ 和H+ 离子流的动态特征; (3)盐胁迫下, 分别用阿米洛利(Na+ /H+ 逆向转运体抑制剂, Amiloride) 和氯化四乙胺(K+ 通道抑制剂, TEA)处理后, Na+ 和K+ 的离子流特性, 以期从离子动态转运方面探讨沙枣耐盐机制和沙枣种源间K+ /Na+ 平衡的调控差异, 为进一步发掘优良耐盐沙枣种质资源提供理论参考依据. ...
... 含量在NaCl胁迫下的降幅也低于银川种源(杨升等, 2014 , 2016 ).但是这些结果都是植株在试验中积累下来的一个总体含量, 是静态的, 而研究探讨盐胁迫下Na+ 和K+ 在沙枣体内具有怎样的动态运移机制, 并且比较不同种源间的差异, 有利于进一步理解沙枣的耐盐机制.因此, 本研究选取沙枣银川种源(YC, 盐敏感型)和阿拉尔种源(ALE, 耐盐型)水培苗为试验材料(杨升等, 2015 , 2016 ), 采用非损伤微测技术(non-invasive micro-test technology, NMT)研究: (1)在150 mmol·L-1 NaCl胁迫24 h后, 沙枣幼苗根系Na+ 和K+ 的稳态离子流; (2) 150 mmol·L-1 NaCl瞬时处理, K+ 和H+ 离子流的动态特征; (3)盐胁迫下, 分别用阿米洛利(Na+ /H+ 逆向转运体抑制剂, Amiloride) 和氯化四乙胺(K+ 通道抑制剂, TEA)处理后, Na+ 和K+ 的离子流特性, 以期从离子动态转运方面探讨沙枣耐盐机制和沙枣种源间K+ /Na+ 平衡的调控差异, 为进一步发掘优良耐盐沙枣种质资源提供理论参考依据. ...
NaCl胁迫下沙枣幼苗的离子代谢特性
2
2016
... 沙枣(Elaeagnus angustifolia )是胡颓子科胡颓子属的落叶小乔木, 主要分布于我国西北五省, 在北京、天津和山东等地也开展了引种栽培, 具有耐旱、耐盐碱和抗风沙的特性, 能提高土壤肥力, 改善土壤环境, 已经被列为半干旱地区和盐土地防护林带的重要树种(郭丽君和王玉涛, 2008 ; 管文轲和徐娜, 2012 ; 杨升等, 2014 ).大量的研究结果显示, 沙枣幼苗在盐胁迫下, 根、茎和叶片中Na+ 含量增加, 而K+ 含量减少(张宝泽等, 1992 ; 刘宝玉, 2007 ; 刘正祥等, 2014 ).前期研究也发现, 沙枣幼苗随着NaCl浓度的增加, 根、茎和叶中的Na+ 含量显著增加, 而K+ 含量显著减少, 并且阿拉尔种源沙枣幼苗与银川种源幼苗间存在显著性差异, 阿拉尔种源沙枣幼苗能积累更多的Na+ 在根中, 而叶片中Na+ 含量较少, K+ 含量在NaCl胁迫下的降幅也低于银川种源(杨升等, 2014 , 2016 ).但是这些结果都是植株在试验中积累下来的一个总体含量, 是静态的, 而研究探讨盐胁迫下Na+ 和K+ 在沙枣体内具有怎样的动态运移机制, 并且比较不同种源间的差异, 有利于进一步理解沙枣的耐盐机制.因此, 本研究选取沙枣银川种源(YC, 盐敏感型)和阿拉尔种源(ALE, 耐盐型)水培苗为试验材料(杨升等, 2015 , 2016 ), 采用非损伤微测技术(non-invasive micro-test technology, NMT)研究: (1)在150 mmol·L-1 NaCl胁迫24 h后, 沙枣幼苗根系Na+ 和K+ 的稳态离子流; (2) 150 mmol·L-1 NaCl瞬时处理, K+ 和H+ 离子流的动态特征; (3)盐胁迫下, 分别用阿米洛利(Na+ /H+ 逆向转运体抑制剂, Amiloride) 和氯化四乙胺(K+ 通道抑制剂, TEA)处理后, Na+ 和K+ 的离子流特性, 以期从离子动态转运方面探讨沙枣耐盐机制和沙枣种源间K+ /Na+ 平衡的调控差异, 为进一步发掘优良耐盐沙枣种质资源提供理论参考依据. ...
... , 2016 ), 采用非损伤微测技术(non-invasive micro-test technology, NMT)研究: (1)在150 mmol·L-1 NaCl胁迫24 h后, 沙枣幼苗根系Na+ 和K+ 的稳态离子流; (2) 150 mmol·L-1 NaCl瞬时处理, K+ 和H+ 离子流的动态特征; (3)盐胁迫下, 分别用阿米洛利(Na+ /H+ 逆向转运体抑制剂, Amiloride) 和氯化四乙胺(K+ 通道抑制剂, TEA)处理后, Na+ 和K+ 的离子流特性, 以期从离子动态转运方面探讨沙枣耐盐机制和沙枣种源间K+ /Na+ 平衡的调控差异, 为进一步发掘优良耐盐沙枣种质资源提供理论参考依据. ...
NaCl胁迫下不同种源沙枣的生长表现差异
1
2015
... 沙枣(Elaeagnus angustifolia )是胡颓子科胡颓子属的落叶小乔木, 主要分布于我国西北五省, 在北京、天津和山东等地也开展了引种栽培, 具有耐旱、耐盐碱和抗风沙的特性, 能提高土壤肥力, 改善土壤环境, 已经被列为半干旱地区和盐土地防护林带的重要树种(郭丽君和王玉涛, 2008 ; 管文轲和徐娜, 2012 ; 杨升等, 2014 ).大量的研究结果显示, 沙枣幼苗在盐胁迫下, 根、茎和叶片中Na+ 含量增加, 而K+ 含量减少(张宝泽等, 1992 ; 刘宝玉, 2007 ; 刘正祥等, 2014 ).前期研究也发现, 沙枣幼苗随着NaCl浓度的增加, 根、茎和叶中的Na+ 含量显著增加, 而K+ 含量显著减少, 并且阿拉尔种源沙枣幼苗与银川种源幼苗间存在显著性差异, 阿拉尔种源沙枣幼苗能积累更多的Na+ 在根中, 而叶片中Na+ 含量较少, K+ 含量在NaCl胁迫下的降幅也低于银川种源(杨升等, 2014 , 2016 ).但是这些结果都是植株在试验中积累下来的一个总体含量, 是静态的, 而研究探讨盐胁迫下Na+ 和K+ 在沙枣体内具有怎样的动态运移机制, 并且比较不同种源间的差异, 有利于进一步理解沙枣的耐盐机制.因此, 本研究选取沙枣银川种源(YC, 盐敏感型)和阿拉尔种源(ALE, 耐盐型)水培苗为试验材料(杨升等, 2015 , 2016 ), 采用非损伤微测技术(non-invasive micro-test technology, NMT)研究: (1)在150 mmol·L-1 NaCl胁迫24 h后, 沙枣幼苗根系Na+ 和K+ 的稳态离子流; (2) 150 mmol·L-1 NaCl瞬时处理, K+ 和H+ 离子流的动态特征; (3)盐胁迫下, 分别用阿米洛利(Na+ /H+ 逆向转运体抑制剂, Amiloride) 和氯化四乙胺(K+ 通道抑制剂, TEA)处理后, Na+ 和K+ 的离子流特性, 以期从离子动态转运方面探讨沙枣耐盐机制和沙枣种源间K+ /Na+ 平衡的调控差异, 为进一步发掘优良耐盐沙枣种质资源提供理论参考依据. ...
SOS1 gene overexpression increased salt tolerance in transgenic tobacco by maintaining a higher K+/Na+ ratio
1
2012
... 在高等植物中, 质膜Na+ /H+ 逆向转运蛋白除了负责调节根部Na+ 外排和体内Na+ 长距离运输外, 还可以调控细胞内的K+ 平衡、pH值和Ca2+ 信号转导 (马清等, 2011 ; 孙健, 2011 ).质膜Na+ /H+ 逆向转运体在盐胁迫下被诱导合成或者激活, 进而将Na+ 从细胞质外排到细胞外, 但是排Na+ 需要质膜两侧产生的H+ 电化学势梯度来完成, 而质膜H+ -ATPase利用水解ATP产生的能量将H+ 泵出细胞质能形成H+ 电化学势梯度(Rausch et al ., 1996 ; 陈观平等, 2006 ; 马清等, 2011 ).同时, Olias等(2009)对番茄(Solanum lycopersicum )研究发现质膜Na+ /H+ 逆向转运体(SlSOS1)不仅在NaCl胁迫下维持离子平衡起重要作用, 而且对于Na+ 在各器官间的区隔化也发挥关键作用.Cuin等(2011)研究认为Na+ /H+ 逆向转运体可以作为小麦耐盐性筛选的一个量化指标.大量转基因试验证实质膜Na+ /H+ 逆向转运体的转基因植物也都表现出更强的耐盐能力(Yang et al ., 2009 ; Yue et al ., 2012 ; Feki et al ., 2014 ).在本试验中, NaCl胁迫下, 沙枣幼苗根系Na+ 外排显著增加, 耐盐型沙枣种源的Na+ 外流速率大于盐敏感型种源, 并且瞬时盐胁迫下, H+ 由内流变为外流, 耐盐型种源沙枣幼苗根系的H+ 外流量明显大于盐敏感型种源, 说明耐盐型种源沙枣幼苗在盐胁迫下具有更强的Na+ /H+ 逆向转运体和质膜H+ -ATPase活性.本研究的药理学试验显示, Na+ /H+ 逆向转运体抑制剂Amiloride显著减少Na+ 的外排, 并且阿拉尔种源在NaCl和NaCl+ Amiloride处理下Na+ 净外排量均显著高于银川种源, 这进一步表明质膜Na+ /H+ 逆向转运蛋白在沙枣根尖外排Na+ 过程中起着非常重要的作用, 但是两个种源在抑制量间无显著差异(449 pmol·cm-2 ·s-1 和 429 pmol·cm-2 ·s-1 ), 并且在NaCl + TEA处理后两个种源根尖的Na+ 净流量也无明显差异, 这可能与抑制剂浓度, 处理时间和试验材料有关, 也可能沙枣的排Na+ 机制有其未知的特殊性,有待进一步研究探讨. ...
盐分胁迫下沙枣某些生理特性的研究
1
1992
... 沙枣(Elaeagnus angustifolia )是胡颓子科胡颓子属的落叶小乔木, 主要分布于我国西北五省, 在北京、天津和山东等地也开展了引种栽培, 具有耐旱、耐盐碱和抗风沙的特性, 能提高土壤肥力, 改善土壤环境, 已经被列为半干旱地区和盐土地防护林带的重要树种(郭丽君和王玉涛, 2008 ; 管文轲和徐娜, 2012 ; 杨升等, 2014 ).大量的研究结果显示, 沙枣幼苗在盐胁迫下, 根、茎和叶片中Na+ 含量增加, 而K+ 含量减少(张宝泽等, 1992 ; 刘宝玉, 2007 ; 刘正祥等, 2014 ).前期研究也发现, 沙枣幼苗随着NaCl浓度的增加, 根、茎和叶中的Na+ 含量显著增加, 而K+ 含量显著减少, 并且阿拉尔种源沙枣幼苗与银川种源幼苗间存在显著性差异, 阿拉尔种源沙枣幼苗能积累更多的Na+ 在根中, 而叶片中Na+ 含量较少, K+ 含量在NaCl胁迫下的降幅也低于银川种源(杨升等, 2014 , 2016 ).但是这些结果都是植株在试验中积累下来的一个总体含量, 是静态的, 而研究探讨盐胁迫下Na+ 和K+ 在沙枣体内具有怎样的动态运移机制, 并且比较不同种源间的差异, 有利于进一步理解沙枣的耐盐机制.因此, 本研究选取沙枣银川种源(YC, 盐敏感型)和阿拉尔种源(ALE, 耐盐型)水培苗为试验材料(杨升等, 2015 , 2016 ), 采用非损伤微测技术(non-invasive micro-test technology, NMT)研究: (1)在150 mmol·L-1 NaCl胁迫24 h后, 沙枣幼苗根系Na+ 和K+ 的稳态离子流; (2) 150 mmol·L-1 NaCl瞬时处理, K+ 和H+ 离子流的动态特征; (3)盐胁迫下, 分别用阿米洛利(Na+ /H+ 逆向转运体抑制剂, Amiloride) 和氯化四乙胺(K+ 通道抑制剂, TEA)处理后, Na+ 和K+ 的离子流特性, 以期从离子动态转运方面探讨沙枣耐盐机制和沙枣种源间K+ /Na+ 平衡的调控差异, 为进一步发掘优良耐盐沙枣种质资源提供理论参考依据. ...


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}