由枝和叶所构成的树冠是植物进行光合作用、呼吸作用、蒸腾作用等一系列生理活动的主要场所, 其大小、结构、形状及相互间的协变规律直接决定了植物个体内的生物量分配、生长活力和生产力。枝条作为植株的重要结构单元, 不仅是植株运输水分和营养物质的重要通道, 也是支撑树冠的主要结构并直接影响树冠的形状和大小, 同时它还是叶片的支撑体, 能通过其长度、结构、分布方式等变化来调整冠层内叶片的排布方式, 将叶伸展到适宜的空间形成一个合理的光拦截体系, 提高光合效率(黎云祥和刘玉成, 1996 ; Day et al ., 2002 ; Thomas & Winner, 2002 ; 张运春等, 2005 )。叶片是植物进行光合作用的主要器官, 叶片的数量、面积以及展叶效率等空间结构特征的变化直接影响着植株的光拦截效率和光合效率。因此, 枝条的结构特征及其与叶性状之间的协变规律反映植物对光照、空间等资源的利用和适应策略(Borchert & Slade, 1981 ; Bicknell, 1982 ; Boojh & Ramakrishnan, 1982 ; Bell, 1984 ; 陈波等, 2002 )。
在木本植物中, 枝条的结构可以用枝条的茎长度、直径以及纤细率(枝条长度与直径比)来表示。对于用来着生和支撑叶、花和果实等器官的小枝而言, 其长度、纤细率以及展叶效率等性状的变化将直接影响植株的机械稳定性、光捕获效率和空间拓展能力, 进而影响到植株的光合和竞争能力的各个方面(Honda & Fisher, 1978 ; Niklas & Kerchner, 1984 )。例如, 树冠内叶片的分布模式是木本植物生产力的一个重要决定因素(Horn, 1971 ; Yagi & Kikuzawa, 1999 ), 而它的变化主要受小枝的结构变化模式、小枝在树冠内的空间排布方式以及小枝的个体大小等与枝条结构密切相关的因素限制(Koike, 1989 ; Yagi & Kikuzawa, 1999 )。在一些具有长短枝分化的物种中, 短枝的茎较短, 叶片排布更紧密, 长枝的茎较长, 叶片可以分散排布在较长的节间上(Yagi & Kikuzawa, 1999 )。尽管短枝上的总叶面积比长枝小(Gregory, 1980 ; Yagi & Kikuzawa, 1999 ), 但单位枝长基础上的叶面积比长枝大(Caesar & Macdonald, 1984 ; Yagi & Kikuzawa, 1999 )。小枝通过这种结构变化模式可以调控一个特定大小的小枝上的叶片数量(Maruyama, 1983 ; Koike, 1989 ; Takenaka, 1997 ; Yagi & Kikuzawa, 1999 )和展叶效率, 影响到植株的生产力。然而, 近年来, 关于植物小枝上茎-叶关系的研究多集中于茎直径维度的叶大小-小枝大小之间的关系研究(Westoby et al ., 2002 ; Pickup et al ., 2005 ; Wright et al ., 2007 ; Li et al ., 2008 ; Xiang et al ., 2009 ; Yang et al ., 2009 ; 杨冬梅等, 2012 ), 对于茎长维度的茎结构的另一维度——茎长度和纤细率对小枝展叶效率及相应的适应策略的研究则相对不足。小枝是个悬臂结构, 小枝茎长度不仅会影响其支撑叶片的排布, 还与小枝的机械安全密切相关。根据恒定胁迫模型(the constant-stress model)理论预测, 小枝茎横截面外缘所能承受的最大弯曲胁迫(M )主要由枝条长度(L )和枝条所承受的力(F )共同决定, 即M = L × F 。对于一个无花、果等其他附属物的小枝而言, 枝条所承受的力主要来自风对枝条的压力和拉拽力所形成的风压胁迫, 而风压胁迫(F )又是叶面积(A )和风压(W )的乘积, 即F = A × W (Dean & Long, 1986 )。由此可知, 在风压一定的情况下, 小枝茎基部所能承受的最大弯曲胁迫是由枝条长度和叶面积决定的, 并且两者之间是负相关关系。所以, 我们预测小枝的展叶效率将随着小枝茎长度的增加而下降。
同时, 对于生活在同一群落里的常绿和落叶物种而言, 由于展叶物候和生物学习性的不同, 它们的功能特征通常也会有很大的差异。例如, 相比于落叶物种, 常绿物种通常具有长的叶寿命、低的比叶面积(SLA )、低的光合速率和高的养分含量(Reich et al ., 1992 ; Westoby et al ., 2002 ; Wright et al ., 2004 ; Li et al ., 2008 ); 另外, 由于常绿物种为了确保在冬季低温等不利环境下叶片不凋落, 需要水分运输系统保持畅通, 确保水分运输安全, 其木质部需要比落叶物种具有更强的抵抗冰冻引起栓塞的能力(Cavender-Bares & Holbrook, 2001 ; Cavender-Bares et al ., 2005 ), 致使其投入在支撑结构上的成本增加, 表现出在相同的茎投入时, 常绿物种通常比落叶物种具有更小的总叶面积和单叶面积(Sun et al ., 2006 ; Yang et al ., 2009 )。这种不同的成本-收益关系将有可能导致常绿和落叶物种采取不同的枝条伸展策略以改变小枝上的叶排布方式, 使两类物种都能够捕获足够的光能以保持相对稳定的碳收益。因此, 两类物种可能会具有不同的茎结构与展叶效率之间的变化关系。
在本研究中, 我们以当年生小枝作为研究对象, 以小枝茎长度、纤细率表征茎的结构; 以单位小枝茎干质量的总叶面积(即叶面积比)、叶密度(即单位茎长度的叶数量)和叶茎生物量比(即单位小枝茎干质量的总叶干质量)表征展叶效率。通过对清凉峰的常绿和落叶阔叶物种的小枝茎结构与展叶效率之间关系的研究, 检测当年生小枝茎结构对展叶效率的影响, 及其在两类不同生活型物种间的差异, 探讨茎长结构变化与展叶效率之间的协变关系在植物生活史策略中的重要意义, 以进一步加深我们对植物生活史对策的理解。
1 材料和方法
1.1 研究区自然概况
研究区位于浙江省清凉峰自然保护区, 地理位置118.87°-119.18° E, 30.08°-30.28° N, 主峰海拔 1β787.4 m。保护区地处中亚热带北缘, 气候复杂, 横跨亚热带和温带两个季风带, 具有明显的亚热带中山山地季风特征。主峰清凉峰则更具有典型的高山气候特点, 立体气候明显, 从山麓到山顶年平均气温从15.3 ℃降至7.8 ℃, 7月为最热月, 历年平均气温28.10 ℃, 1月为最冷月, 历年平均气温3.40 ℃。极端最高气温为41.90 ℃ (1996年8月6日), 极端最低气温为-13.30 ℃ (1967年1月16日)。全年降水量 1β500-1β900 mm, 相对湿度在78%-82%之间。全山区年日照时间1β500-2 000 h, 年日照百分率为35%- 39%。海拔800-1 100 m区域年日照时间1 500-1 600 h (翁东明等, 2009 ; 程丽媛等, 2016 )。
保护区内植被带景观垂直层次分明, 群落内乔木层植物平均密度为2 062株·hm-2 , 灌木层平均密度为17 163株·hm-2 , 草本层平均密度为5 976株·hm-2 。灌木层植株密度基本是乔木层植株密度的8.3倍, 是草本层植株密度的2.9倍(翁东明等, 2009 ; 程丽媛等, 2016 )。
1.2 植物采样和测量
本研究于2011年7-8月, 当枝条生长和叶片展开完成时, 采集了60种落叶和25种常绿阔叶木本植物的小枝为研究对象。本研究中定义“小枝”为末端无分枝的当年生小枝, 即从最末端到最后一个末梢分枝处, 所以通常是不分枝的, 且无花和果实, 包括终端节间部分和其上所有叶。对每一种所研究的木本植物随机选择3-5个个体, 且从植物外冠层对每一个个体随机选取没有明显叶面积损失的3-5个小枝。以小枝为单位, 记录其上所有叶数量、叶面积(通过扫描叶片, 并用MapInfo软件进行计算)、茎长度、茎直径等。将茎、叶分别在70 ℃烘箱中烘干48 h至恒质量并记录干质量。单叶面积(ILA )由总叶面积和叶数量计算得到(ILA = LA /LN ); 单叶干质量(ILM )由总叶干质量和叶数量计算获得(ILM = LM /LN ); 以小枝茎长度和茎纤细率表征小枝茎长维度的结构变化, 其中小枝茎纤细率(SSR )由小枝茎长度与茎直径的比值表示(SSR = SL /SD ); 以叶面积比、叶茎质量比和叶密度表征展叶效率, 其中叶面积比(LAR )由当年生小枝上总叶面积与茎干质量的比值表示(LAR = LA /SM ); 叶茎质量比(LSR )由当年生小枝上总叶干质量与茎干质量的比值表示(LMR = LM /SM ); 叶密度(DLN )由当年生小枝上的总叶数与小枝茎长度的比值表示(DLN = LN /SL ); 比叶面积(SLA )由总叶面积和总叶干质量的比值表示(SLA = LA /LM ), 用以表征叶的结构变化。
1.3 数据分析
对于植物构型性状以物种平均值进行分析。在对枝条结构与展叶效率的关系分析前, 首先将各参数的物种平均值进行对数转换(以10为底), 使之符合正态分布, 然后用关系方程y = bxa 来描述, 对数转换后为log(y ) = log(b ) + a log(x ), 其中x 和y 表示两个性状变量, 在双对数直角坐标系中, b 表示性状关系的截距, a 表示斜率, 即异速生长的指数(a = 1时为等速生长, 大于或小于1时为异速生长)(Harvey & Pagel, 1991 )。异速生长方程的参数估计采用标准化主轴估计(SMA)方法(Warton et al ., 2006 ), 由软件(S)MATR Version 2.0 (Falster et al ., 2006 )计算完成。回归斜率的置信区间根据(Pitman, 1939 )方法计算。
在异速生长分析中, 对于常绿和落叶物种组的斜率采用Warton和Weber (2002)的方法进行异质性检验, 如果检验结果没有异质性(p > 0.05), 就给出一个共同斜率。共同斜率下的截距差异(y 轴向)以及沿共同主轴方向位移的差异通过t 检验或方差分析检验(Wright et al ., 2001 ; Warton et al ., 2006 )进行分析。
此外, 落叶和常绿物种组的各构型性状间的比较采用独立样本的t 检验。所有数据分析均在SPSS 18.0下进行。
2 结果
2.1 小枝茎结构与展叶效率的关系
2.1.1 小枝茎结构与叶面积比(LAR )的关系
在本研究中, LAR 与SL 、小枝SSR 在落叶和常绿物种组中均具有显著的负相关关系(图1 ; 表1 ), 其共同斜率分别为-0.961 (p = 0.773, 95%置信区间-1.097到-0.843)和-0.844 (p = 0.546, 95%置信区间-0.970到-0.736), 表明小枝上的叶面积比随小枝茎长度和茎纤细率的增加而逐渐递减。同时, 落叶物种组的叶面积比与小枝茎长度、茎纤细率相关关系共同斜率的截距显著大于常绿物种组(图1 , p 值均小于0.001), 表明在某一给定的茎长结构下, 落叶物种比常绿物种具有更高的叶面积比。
图1 叶面积比与当年生小枝茎长度(A)、茎纤细率(B)之间的关系。
Fig. 1 Relationship between leaf area ratio and current- year twig stem length (A) and stem slender ratio (B).
2.1.2 小枝茎结构与叶密度的关系
无论在落叶和常绿物种组内, 小枝茎长度均与其上的叶数量呈显著的正相关关系(图2 ; 表1 ), 其共同斜率为0.495 (p = 0.653, 95%置信区间0.407到0.602), 显著小于1.0 (p < 0.000β1), 即存在显著的小于1的异速生长关系, 表明叶数量的增加速度没有小枝茎长度的增加速度快, 长的枝条相对于短枝具有低的出叶强度。此外, 落叶物种的小枝茎长度与叶数量的相关关系共同斜率的截距显著低于常绿物种组(图2 , p < 0.001), 表明在某一给定的茎长度下, 常绿物种比落叶物种具有更多的叶数量。
图2 小枝上的叶数量与当年生小枝茎长度之间的关系。
Fig. 2 Relationship between leaf number per twig and current-year twig stem length.
小枝茎纤细率和茎长度在落叶和常绿物种组中均与叶密度呈显著的负相关关系(图3 ; 表1 ), 其共同斜率分别为-0.766 (p = 0.377, 95%置信区间-0.873到-0.677)和-0.885 (p = 0.622, 95%置信区间-0.988到-0.793), 表明随着小枝茎纤细率和长度的增加, 叶密度逐渐减小。此外, 落叶物种的小枝结构性状与叶密度相关关系共同斜率的截距显著低于常绿物种组(图3 , p 值均小于0.001), 表明在某一给定的小枝茎纤细率和茎长度下, 常绿物种比落叶物种具有更高的叶密度。
图3 叶密度与小枝茎纤细率(A)和小枝茎长度(B)的关系。
Fig. 3 Relationship between the density of leaf number and stem slender ratio (A) and stem length (B).
2.1.3 小枝茎结构与叶茎生物量比的关系
小枝的茎纤细率和茎长度在落叶和常绿物种组中均与叶茎生物量比呈显著的负相关关系(图4 ; 表1 ), 且在两组关系中, 同时表现出落叶物种关系斜率显著低于常绿物种(p 值均小于0.02), 表明在小枝茎纤细率和长度增加相同的情况下, 落叶物种的小枝叶生物量分配比例比常绿物种下降得更快。
图4 叶茎质量比与小枝茎纤细率(A)和小枝茎长度(B)的关系。
Fig. 4 Relationship between the leaf/stem mass ratio and stem slender ratio (A) and stem length (B).
2.2 常绿和落叶物种展叶效率及各功能性状的比较
在本研究区域, 常绿物种和落叶物种在展叶效率和部分茎、叶功能性状上表现出显著的差异(表2 )。落叶物种比常绿物种具有更高的单叶面积和当年生小枝上的总叶面积(p ≤ 0.001); 具有更低的叶密度和叶面积比(p < 0.05)。然而, 两个物种组在茎长度、直径、叶干质量、茎干质量及叶茎质量比和茎纤细率上无显著差异(p > 0.05)。
3 讨论
3.1 枝条结构对展叶效率的影响
本研究中, 无论是落叶还是常绿物种, 用以表征展叶效率的叶面积比、叶密度、叶茎生物量比, 均与表征当年生小枝茎结构的小枝茎长度和茎纤细率呈显著的负相关关系(图1 , 图3 , 图4 ), 即随着当年生小枝茎长度和纤细率的增加, 小枝的展叶效率逐渐降低。这一结果与Takenaka和Yagi等的研究结果(Takenaka, 1997 ; Yagi & Kikuzawa, 1999 ; Yagi, 2000 , 2004 )类似, 他们在多个木本植物的种内研究发现, 较长的当年生枝条具有更小的单位枝长基础上的叶面积。而叶密度可能是小枝展叶效率的一个主要调节因素。我们的结果显示, 无论是在常绿还是落叶物种中, 不仅小枝茎长度和茎纤细率均与叶密度(叶数量/茎长度)呈显著的负相关关系(图3 ), 并且每个小枝上的叶数量也直接与小枝茎长度呈斜率显著小于1.0 (p < 0.000β1)的正的异速生长关系(图2 ; 表1 ), 表明叶数量的增加速度没有小枝茎长度的增加速度快, 长的枝条相对于短枝具有更低的出叶强度。Yagi (2004)的研究结果也表明, 长的当年生小枝比短的当年生小枝具有更小的叶数量与枝长度的比值, 即单位枝长度的叶片数更少。在我们以前的研究中也发现, 小枝茎长度和单位茎长度的叶片数
量不仅在每一个物种(或生境)组中(R 2 从0.315到0.814, p < 0.05), 而且在整个综合数据库中(R 2 = 0.615, p < 0.001)均存在显著的负相关关系(杨冬梅等, 2012 )。这种现象被归因于长枝比短枝具有更强的顶端优势, 侧芽更容易被抑制, 使得叶密度减小(Xiang et al ., 2009 ; Yagi & Kikuzawa, 1999 ; Yagi, 2000 )。
从机械力学的角度看, 展叶效率随小枝茎长度和茎纤细率的增加而下降可能是小枝在面对机械载重和风压胁迫时, 确保小枝安全的需要。作为起支撑作用的小枝, 首先要能够承受其自身的质量, 避免折断、撕裂或其他形式的机械损伤(Mcmahon, 1973 ; King & Loucks, 1978 ; King, 1981 , 1986 ; Long et al ., 1981 ; Givnish, 1986 ; Gartner, 1995 ), 必须能够抵抗来自茎及其上叶片质量所带来的静态和动态的压力和拉力胁迫。恒定胁迫模型(The constant-stress model)理论认为, 令植物茎横截面外缘弯曲所需要做的功(即弯曲矩, the bending moment) M 主要是由力F 和臂长(即茎长) L 共同决定, 即M = F × L 。对于植物而言, 茎所承受的力是叶面积A 与风压W 的乘积, 即F = A × W (Dean & Long, 1986 )。所以, 随着小枝茎长度的增加, 在小枝上相同叶面积对小枝茎基部的机械负载会相应地增加, 特别是在有风吹动摇摆的情况下, 长枝将相应地减少枝条上的叶片面积或干质量来降低机械载重和风压胁迫, 导致展叶效率随小枝茎长度和茎纤细率的增加而下降。其次, 从植株树冠构建的角度看, 枝条结构与展叶效率之间的负相关关系将有利于植株更有效地捕获光能, 提高整体光合效率。因为相互遮阴是当年生小枝结构变化的一个重要决定因素, 更大的展叶密度将会导致当年生小枝内叶片之间相互荫蔽的程度增加(Niklas, 1988 ; Yagi & Kikuzawa, 1999 )。另外, 在总的生物量一定的情况下, 当植物将更多的生物量分配给茎以使枝条生长得更长时, 将有利于植株将更多的叶片伸展到冠层外部, 同时可以让一定数量的叶片分散在更长的枝条范围内, 降低同一枝条上叶片之间相互荫蔽的程度, 以利于捕获更多的光能, 提高植株的光合效率。综上所述, 枝条结构与展叶效率之间的负相关关系具有重要的生态学意义, 有利于保持小枝的机械安全, 并提高整个植株的光合效率。
3.2 常绿和落叶物种间的比较
常绿与落叶是植物通过长期进化适应形成的两种不同的生活史对策, 它们在叶寿命和叶片结构(如比叶面积)(Reich et al ., 1992 ; Westoby et al ., 2002 ; Wright et al ., 2004 ; Li et al ., 2008 )、茎木质部抗栓塞能力(Cavender-Bares & Holbrook, 2001 ; Cavender- Bares et al ., 2005 )等许多方面都显著不同。在本研究中, 虽然当年生小枝茎结构性状与表征展叶效率的性状之间的负相关关系斜率在常绿和落叶物种间无显著差异, 但截距差异显著。在3个表征展叶效率的性状中, 尽管常绿物种叶密度与小枝茎结构性状之间的关系截距显著大于落叶物种, 在某一给定的茎长或茎纤细率下, 常绿物种比落叶物种具有更多的叶数量, 即具有较高的出叶强度, 与以前相关研究发现的叶大小与叶数量之间是相互权衡的结果一致(Corner, 1949 ; Kleiman & Aarssen, 2007 ; Yang et al ., 2008 )。但由于常绿物种的单叶面积16.21 cm2 显著小于落叶物种的单叶面积34.66 cm2 (p < 0.001, 表2 ), 导致其叶面积比(单位茎质量的叶面积)和叶茎生物量比(单位茎质量的叶质量)与小枝茎结构性状之间的关系截距反而显著小于落叶物种, 即随着小枝茎长度和纤细率的增加, 常绿物种每单位质量的茎生物量投资获得的总叶面积和总叶质量都比落叶物种少, 常绿物种的实际展叶效率比落叶物种低。对数据结果进一步分析表明, 常绿物种更低展叶效率的主要原因是其比叶面积显著小于落叶物种 (p < 0.001, 表2 ), 因为我们的结果显示, 在不考虑小枝茎长结构影响的情况下, 茎叶生物量比在常绿和落叶物种中没有显著差异(p = 0.335, 表2 ), 表明总生物量在叶和茎之间的分配比例在常绿和落叶物种中是没有显著差异的, 可是落叶物种的单叶面积(p < 0.001, 表2 )和总叶面积(p = 0.001, 表2 )都要显著大于常绿物种, 并且常绿物种茎干质量与总叶面积相关关系共同斜率下的截距显著小于落叶物种(p ˂ 0.05, 表1 ), 即在某一给定的茎干质量下, 常绿物种比落叶物种具有更小的叶面积。许多研究也都发现常绿物种通常比落叶物种具有更高的比叶质量(即比叶面积的倒数)(Reich et al ., 1992 ; Westoby et al ., 2002 ; Wright et al ., 2004 ; 潘少安等, 2015 )。并且, Reich等(1998)发现比叶面积较低的植物常形成厚度较大而面积较小的叶片, 而刘艳芳(2015)在贡嘎山阔叶木本植物叶片解剖结构特征及其环境适应研究中发现的常绿植物的叶片厚度、栅栏组织厚度、海绵组织厚度、上表皮厚度和栅栏细胞层数都显著大于落叶植物的结果, 支持了Reich等(1998)的发现。
常绿物种较低的比叶面积继而导致较低的单位茎结构基础上的展叶效率, 可能与其具有较长的叶寿命有关。一般而言, 叶寿命越长, 叶片越厚, 单位叶面积的建成消耗也越大, 叶子需要在足够长的时间内通过自身的光合生产所形成的有机碳来补偿初始的建成消耗(包括抵抗草食动物的防御构造), 并维持自身生存所需的物质消耗(Coley, 1988 ; Kikuzawa, 1991 ; Schulze et al ., 1994 ; Kikuzawa, 1995 ; 张林和罗天祥, 2004 )。所以叶寿命与比叶面积成反比(Reich et al ., 1992 ; Reich & Bowman, 1999 ; Ryser & Urbas, 2000 ; 张林和罗天祥, 2004 )。比叶面积较低的植物, 叶片中很大一部分物质用于构建保卫构造(防虫食、防止过度失水等)或者增加叶肉细胞密度以抵御环境胁迫(Reich et al ., 1998 ; 张林和罗天祥, 2004 )。与落叶物种相比, 常绿物种叶片在冬季不凋落, 需经受较长时间的冬季低温等不利生长环境, 面对冬季的选择压力, 常绿植物可能采取高消耗慢收益的保守型生存策略(Cornelissen, 1999 ; 刘艳芳, 2015 ), 通过革质的叶表皮和叶肉海绵组织分层等特点来增加抗寒力(董丽等, 2003 ; 刘艳芳, 2015 )。而落叶植物的叶寿命相对较短, 需要在相对短的生长时间内积累足够的光合产物以满足植株生长需要, 薄的叶片组织能够缩短CO2 从气孔扩散到叶绿体的路径, 也能增加气体交换的速率, 进而提高光合作用速率(Diemer, 1998 ; 刘艳芳, 2015 ), 所以, 较高的SLA 是落叶物种所采取的一种低投入快收益的积极型生存策略。
4 结论
我们的研究结果表明, 无论是在常绿物种还是在落叶物种中, 小枝茎长度和茎纤细率均与表征展叶效率的叶面积比、叶茎生物量比和叶密度显著负相关, 表明随着小枝茎长度和茎纤细率的增加, 小枝的展叶效率逐渐下降。其中, 叶密度是小枝展叶效率的一个主要调节因素。展叶效率随小枝茎长度和茎纤细率的增加而下降, 既有利于保持小枝的机械稳定性和确保小枝安全, 也可以让植株在总生物量一定的情况下, 将更多的生物量用于枝长生长以便将更多的叶片伸展到冠层外部, 并且让叶片分散排布在更长的枝长范围内而减少叶片之间的相互荫蔽程度, 从而提高植株整体的光捕获效率和光合效率。同时, 我们的研究也发现, 尽管当年生小枝茎结构性状与展叶效率间的负相关关系斜率在常绿和落叶物种间无显著差异, 并且常绿物种叶密度与小枝茎结构性状之间的关系截距显著大于落叶物种, 但由于单叶面积显著小于落叶物种, 导致其叶面积比和叶茎生物量比与小枝茎结构性状之间的关系截距显著小于落叶物种, 即随着小枝茎长度和纤细率的增加, 常绿物种每单位质量的茎生物量投资获得的总叶面积和总叶质量都比落叶物种少, 常绿物种的实际展叶效率比落叶物种低。对数据的进一步分析表明, 常绿物种较低的展叶效率主要归因于其叶片的比叶面积比落叶物种低。因为常绿物种的叶寿命较长, 冬季叶片不凋落, 需面对冬季选择压力, 所以采取高消耗慢收益的保守型生存策略, 形成小而厚的叶片增强抗寒力, 确保安全度过冬季低温期。而落叶物种叶寿命相对较短, 需在短时间内完成植株生长所需要的物质积累, 故采取高比叶面积、高展叶效率的低投入快收益的积极型生存策略。因此, 小枝茎长维度上的茎结构性状与展叶效率之间的关系能够反映植物对环境的适应策略, 深入探讨这一关系在不同物种、不同环境条件下的变化将有利于我们更好地理解植物的生活史对策。
致谢 浙江省清凉峰自然保护区张宏伟工程师、浙江师范大学化学与生命科学学院陈建华教授为本研究中的物种鉴定提供了大量的帮助; 在研究过程中, 得到了浙江师范大学行知学院章佳佳、钱敏杰、郑瑶、周丹和金灵妙等同学的大力协助; 浙江省清凉峰自然保护区为本研究的野外采样提供了便利条件, 在此一并致谢。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Bell AD (1984). Dynamic morphology: A contribution to plant population ecology. In: Dirzo R ed. Perspectives on Plant Population Ecology . Sinaner, Sunderland, USA .
[本文引用: 1]
[2]
Bicknell SH 1982 ). Development of canopy stratification during early succession in northern hardwoods.
Forest Eco- logy & Management, 4 , 41 -51 .
[本文引用: 1]
[3]
Boojh R Ramakrishnan PS 1982 ). Growth strategy of trees related to successional status. I. Architecture and extension growth.
Forest Ecology & Management, 4 , 359 -374 .
[本文引用: 1]
[4]
Borchert R Slade NA 1981 ). Bifurcation ratios and the adaptive geometry of trees.
Botanical Gazette, 142 , 394 -401 .
[本文引用: 1]
[5]
Caesar JC Macdonald AD 1984 ). Shoot development in Betula papyrifera . IV. Comparisons between growth characteristics and expression of vegetative long and short shoots
. Canadian Journal of Botany, 62 , 446 -453 .
[本文引用: 1]
[6]
Cavender-Bares J Cortes P Rambal S Joffre R Miles B Rocheteau A 2005 ). Summer and winter sensitivity of leaves and xylem to minimum freezing temperatures: A comparison of co-occurring Mediterranean oaks that differ in leaf lifespan.
New Phytologist, 168 , 597 -612 .
[本文引用: 2]
[7]
Cavender-Bares J Holbrook NM 2001 ). Hydraulic properties and freezing-induced cavitation in sympatric evergreen and deciduous oaks with contrasting habitats.
Plant, Cell & Environment, 24 , 1243 -1256 .
[本文引用: 2]
[8]
Chen B Song YC Da LJ 2002 ). Woody plant architecture and its research in plant ecology.
Chinese Journal of Ecology, 21 , 52 -56 . (in Chinese with English abstract)[ 陈波 , 宋永昌 , 达良俊 (2002 ). 木本植物的构型及其在植物生态学研究的进展
. 生态学杂志 , 21 , 52 -56 .]
[本文引用: 1]
[9]
Cheng LY Cao T Zhang HW Yu J 2016 ). Studies on flora of bryophytes in Qingliangfeng National Reserve, Zhejiang Province.
Acta Botanica Boreali-Occidentalia Sinica, 23 , 398 -403 . (in Chinese with English abstract)[ 程丽媛 , 曹同 , 张宏伟 , 于晶 (2016 ). 浙江省清凉峰自然保护区苔藓植物区系成分研究
. 西北植物学报 , 36 , 398 -403 .]
[本文引用: 2]
[10]
Coley PD 1988 ). Effects of plant growth rate and leaf lifetime on the amount and type of anti-herbivore defense.
Oecologia, 74 , 531 -536 .
[本文引用: 1]
[11]
Cornelissen JHC 1999 ). A triangular relationship between leaf size and seed size among woody species: Allometry, ontogeny, ecology and taxonomy.
Oecologia, 118 , 248 -255 .
[本文引用: 1]
[12]
Corner EJH 1949 ). The durian theory or the origin of the modern tree.
Annals of Botany, 13 , 367 -414 .
[本文引用: 1]
[13]
Day ME Greenwood MS Diazsala C 2002 ). Age- and size- related trends in woody plant shoot development: Regulatory pathways and evidence for genetic control.
Tree Physiology, 22 , 507 -513 .
[本文引用: 1]
[14]
Dean TJ Long JN 1986 ). Validity of constant-stress and elastic-instability principles of stem formation in
Pinus contorta and Trifolium pratense . Annals of Botany, 58 , 833 -840 .
[本文引用: 2]
[15]
Diemer M 1998 ). Life span and dynamics of leaves of herbaceous perennials in high-elevation environments: “News from the elephant’s leg”.
Functional Ecology, 12 , 413 -425 .
[本文引用: 1]
[16]
Dong L Jia GX Su XH 2003 ). Change of the leaf tissue structure of evergreen broad-leaf plants during overwintering.
Acta Horticulture Sinica, 30 , 59 -64 . (in Chinese with English abstract)[ 董丽 , 贾桂霞 , 苏雪痕 (2003 ). 常绿阔叶植物越冬期间叶片组织结构的适应性变化
. 园艺学报 , 30 , 59 -64 .]
[本文引用: 1]
[17]
Falster DS Warton DI Wright IJ
URL
[本文引用: 1]
[18]
Gartner BL 1995).Plant Stems:Physiology and Functional Morphology . Academic Press, San Diego, USA .
[本文引用: 2]
[19]
Givnish TJ 1986 ). Biomechanical constraints on self-thinning in plant populations.
Journal of Theoretical Biology, 119 , 139 -146 .
[本文引用: 1]
[20]
Gregory RA 1980 ). Annual cycle of shoot development in sugar maple.
Canadian Journal of Forest Research, 10 , 316 -326 .
[21]
Harvey PH Pagel MD The Comparative Method in Evolutionary Biology . Oxford University Press , Oxford, UK . 239 .
[本文引用: 2]
[22]
Honda H Fisher JB 1978 ). Tree branch angle: Maximizing effective leaf area.
Science, 199 , 888 -890 .
[本文引用: 1]
[23]
Horn HS 1971 ). The adaptive geometry of trees.
Botanical Gazette, 142 , 394 -401 .
[本文引用: 1]
[24]
Kikuzawa K 1991 ). A cost-benefit analysis of leaf habit and leaf longevity of trees and their geographical pattern.
The American Naturalist, 138 , 1250 -1263 .
[本文引用: 1]
[25]
Kikuzawa K 1995 ). Leaf phenology as an optimal strategy for carbon gain in plants.
Canadian Journal Botany, 73 , 158 -163 .
[本文引用: 1]
[26]
King D 1981 ). Tree dimensions: Maximizing the rate of height growth in dense stands.
Oecologia, 51 , 351 -356 .
[本文引用: 1]
[27]
King D Loucks OL 1978 ). The theory of tree bole and branch form.
Radiation and Environmental Biophysics, 15 , 141 -165 .
[本文引用: 1]
[28]
King DA 1986 ). Tree form, height growth, and susceptibility to wind damage in
Acer saccharum . Ecology, 67 , 980 -990 .
[本文引用: 1]
[29]
Kleiman D Aarssen LW 2007 ). The leaf size/number trade-off in trees.
Journal of Ecology, 95 , 376 -382 .
[本文引用: 2]
[30]
Koike F 1989 ). Foliage-crown development and interaction in
Quercus gilva and Q. acuta . Journal of Ecology, 77 , 92 -111 .
[本文引用: 3]
[31]
Li G Yang D Sun S 2008 ). Allometric relationships between lamina area, lamina mass and petiole mass of 93 temperate woody species vary with leaf habit, leaf form and altitude.
Functional Ecology, 22 , 557 -564 .
[本文引用: 1]
[32]
Li YX Liu YC 1996 ). The neighbourhood modular interference effects of seedlings of Gordonia acuminata .
Journal of Southwest China Normal University (Natural Science ) , 21 , 271 -275 . (in Chinese with English abstract)[ 黎云祥 , 刘玉成 (1996 ). 四川大头茶苗木构件水平的邻体干扰效应
. 西南师范大学学报自然科学版 , 21 , 271 -275 .]
[本文引用: 3]
[33]
Liu YF 2015 ). The Study on Anatomical Structures Character and Its Environmental Adaptation of Leaves from Broad- Leaf Woody Plants in Gongga Mountain . Master degree dissertation, Southwest University, Chongqing.
(in Chinese with English abstract)[ 刘艳芳 (2015 ). 贡嘎山阔叶木本植物叶片解剖结构特征及其环境适应研究
. 硕士学位论文, 西南大学, 重庆.]
[本文引用: 1]
[34]
Long JN Smith FW Scott DRM 1981 ). The role of Douglas- fir stem sapwood and heartwood in the mechanical and physiological support of crowns and development of stem form.
Canadian Journal of Forest Research, 11 , 459 -464 .
[本文引用: 1]
[35]
Maruyama K 1983 ). Shoot characteristics as a function of the bud length on Japanese beech trees.
Journal of the Japanese Forestry Society, 65 , 43 -51 .
[本文引用: 1]
[36]
Mcmahon T 1973 ). Size and shape in biology.
Science, 179 , 1201 -1204 .
[本文引用: 1]
[37]
Niklas KJ Kerchner V 1984 ). Mechanical and photosynthetic constraints on the evolution of plant shape.
Paleobiology, 10 , 79 -101 .
[本文引用: 1]
[38]
Niklas KJ 1988 ). The role of phyllotactic pattern as a “developmental constraint” on the interception of light by leaf surfaces.
Evolution, 42 , 1 -16 .
[本文引用: 1]
[39]
Pan SA Peng GQ Yang DM 2015 ). Biomass allocation strategies within a leaf: Implication for leaf size optimization.
Chinese Journal of Plant Ecology, 39 , 971 -979 . (in Chinese with English abstract)[ 潘少安 , 彭国全 , 杨冬梅 (2015 ). 从叶内生物量分配策略的角度理解叶大小的优化
. 植物生态学报 , 39 , 971 -979 .]
[本文引用: 1]
[40]
Pickup M Westoby M Basden A 2005 ). Dry mass costs of deploying leaf area in relation to leaf size.
Functional Ecology, 19 , 88 -97 .
[本文引用: 1]
[41]
Pitman E 1939 ). A note on normal correlation.
Biometrika, 31 , 9 -12 .
[本文引用: 4]
[42]
Reich P Walters M Ellsworth D 1992 ). Leaf life-span in relation to leaf, plant, and stand characteristics among diverse ecosystems.
Ecological Monographs, 62 , 365 -392 .
[本文引用: 1]
[43]
Reich PB Bowman WD 1999 ). Generality of leaf trait relationships: A test across six biomes.
Ecology, 80 , 1955 -1969 .
[本文引用: 1]
[44]
Reich PB Walters MB Ellsworth DS Vose JM Volin JC Gresham C Bowman WD 1998 ). Relationships of leaf dark respiration to leaf nitrogen, specific leaf area and leaf life-span: A test across biomes and functional groups.
Oecologia, 114 , 471 -482 .
[本文引用: 1]
[45]
Ryser P Urbas P 2000 ). Ecological significance of leaf life span among Central European grass species.
Oikos, 91 , 41 -50 .
[本文引用: 1]
[46]
Schulze E Kelliher FM Korner C Lloyd J Leuning R 1994 ). Relationships among maximum stomatal conductance, ecosystem surface conductance, carbon assimilation rate, and plant nitrogen nutrition: A global ecology scaling exercise.
Ecology, Evolution, and Systematics, 25 , 629 -662 .
[本文引用: 1]
[47]
Sun S Jin D Shi P 2006 ). The leaf size-twig size spectrum of temperate woody species along an altitudinal gradient: An invariant allometric scaling relationship.
Annals of Botany, 97 , 97 -107 .
[本文引用: 2]
[48]
Takenaka A 1997 ). Structural variation in current-year shoots of broad-leaved evergreen tree saplings under forest canopies in warm temperate Japan.
Tree Physiology, 17 , 205 -210 .
[本文引用: 1]
[49]
Thomas SC Winner WE 2002 ). Photosynthetic differences between saplings and adult trees: An integration of field results by meta-analysis.
Tree Physiology, 22 , 117 -127 .
[50]
Warton DI Weber NC 2002 ). Common slope tests for bivariate errors-in-variables models.
Biometrical Journal, 44 , 161 -174 .
[本文引用: 2]
[51]
Warton DI Wright IJ Falster DS Westoby M 2006 ). Bivariate line-fitting methods for allometry.
Biological Reviews, 81 , 259 -291 .
[本文引用: 2]
[52]
Weng DM Zhang L Chen XD Shen GC Zhang HW Zhang FG Yu MJ 2009 ). Species diversity of Fagus hayatae community in Qingliangfeng National Nature Reserve.
Journal of Zhejiang Forestry Science and Technology, 29 (4 ), 1 -6 .[翁东明 , 张磊 , 陈晓栋 , 沈国春 , 张宏伟 , 张方钢 , 于明坚 (2009 ). 清凉峰自然保护区台湾水青冈群落物种多样性研究
. 浙江林业科技 , 29 (4 ), 1 -6 .]
[本文引用: 4]
[53]
Westoby M Falster DS Moles AT Vesk PA Wright IJ 2002 ). Plant ecological strategies: Some leading dimensions of variation between species.
Annual Review of Ecology and Systematics, 33 , 125 -159 .
[本文引用: 1]
[54]
Wright IJ Ackerly DD Bongers F Harms KE Ibarra- Manriquez G Martinez-Ramos M Mazer SJ Muller- Landau HC Paz H Pitman NCA Poorter L Silman MR Vriesendorp CF Webb CO Westoby M Wright SJ 2007 ). Relationships among ecologically important dimensions of plant trait variation in seven neotropical forests.
Annals of Botany, 99 , 1003 -1015 .
[本文引用: 1]
[55]
Wright IJ Reich PB Westoby M 2001 ) Strategy shifts in leaf physiology, structure and nutrient content between species of high- and low-rainfall and high- and low-nutrient habitats.
Functional Ecology, 15 , 423 -434 .
[本文引用: 3]
[56]
Wright IJ Reich PB Westoby M Ackerly DD Baruch Z Bongers F Cavender-Bares J Chapin FS Cornelissen JHC Diemer M 2004 ). The leaf economics spectrum worldwide.
Nature, 428 , 821 -827 .
[本文引用: 2]
[57]
Xiang SA Liu YL Fang F Wu N Sun SC 2009 ). Stem architectural effect on leaf size, leaf number, and leaf mass fraction in plant twigs of woody species.
International Journal of Plant Sciences, 170 , 999 -1008 .
[本文引用: 2]
[58]
Yagi T 2000 ). Morphology and biomass allocation of current-year shoots of ten tall tree species in cool temperate Japan.
Journal of Plant Research, 113 , 171 -183 .
[本文引用: 1]
[59]
Yagi T 2004 ). Within-tree variations in shoot differentiation patterns of 10 tall tree species in a Japanese cool-temperate forest.
Canadian Journal of Botany, 82 , 228 -243 .
[本文引用: 9]
[60]
Yagi T Kikuzawa K 1999 ). Patterns in size-related variations in current-year shoot structure in eight deciduous tree species.
Journal of Plant Research, 112 , 343 -352 .
[本文引用: 1]
[61]
Yang DM Li GY Sun SC 2008 ). The generality of leaf size versus number trade-off in temperate woody species.
Annals of Botany, 102 , 623 -629 .
[本文引用: 2]
[62]
Yang DM Li GY Sun SC 2009 ). The effects of leaf size, leaf habit, and leaf form on leaf/stem relationships in plant twigs of temperate woody species.
Journal of Vegetation Science, 20 , 359 -366 .
[本文引用: 2]
[63]
Yang DM Zhan F Zhang HW 2012 ). Trade-off between leaf size and number in current-year twigs of deciduous broad-leaved woody species at different altidudes on Qingliang Mountain, southeastern China.
Chinese Journal of Plant Ecology, 36 , 281 -291 . (in Chinese with English abstract)[ 杨冬梅 , 占峰 , 张宏伟 (2012 ). 清凉峰不同海拔木本植物小枝内叶大小-数量权衡关系
. 植物生态学报 , 36 , 281 -291 .]
[本文引用: 3]
[64]
Zhang L Luo TX 2004 ). Advances in ecological studies on leaf lifespan and associated leaf traits.
Acta Phytoecologica Sinica, 28 , 844 -852 . (in Chinese with English abstract)[ 张林 , 罗天祥 (2004 ). 植物叶寿命及其相关叶性状的生态学研究进展
. 植物生态学报 , 28 , 844 -852 .]
[本文引用: 1]
[65]
Zhang YC Du XJ Zhang QY Gao XM Su ZX 2005 ). Functions of branches of the clonal tree
Symplocos laurina . Acta Phytoecologica Sinica, 29 , 799 -806 . (in Chinese with English abstract)[ 张运春 , 杜晓军 , 张桥英 , 高贤明 , 苏智先 (2005 ). 克隆乔木黄牛奶树枝条的功能特征
. 植物生态学报 , 29 , 799 -806 .]
1
... 由枝和叶所构成的树冠是植物进行光合作用、呼吸作用、蒸腾作用等一系列生理活动的主要场所, 其大小、结构、形状及相互间的协变规律直接决定了植物个体内的生物量分配、生长活力和生产力.枝条作为植株的重要结构单元, 不仅是植株运输水分和营养物质的重要通道, 也是支撑树冠的主要结构并直接影响树冠的形状和大小, 同时它还是叶片的支撑体, 能通过其长度、结构、分布方式等变化来调整冠层内叶片的排布方式, 将叶伸展到适宜的空间形成一个合理的光拦截体系, 提高光合效率(黎云祥和刘玉成, 1996 ; Day et al ., 2002 ; Thomas & Winner, 2002 ; 张运春等, 2005 ).叶片是植物进行光合作用的主要器官, 叶片的数量、面积以及展叶效率等空间结构特征的变化直接影响着植株的光拦截效率和光合效率.因此, 枝条的结构特征及其与叶性状之间的协变规律反映植物对光照、空间等资源的利用和适应策略(Borchert & Slade, 1981 ; Bicknell, 1982 ; Boojh & Ramakrishnan, 1982 ; Bell, 1984 ; 陈波等, 2002 ). ...
Development of canopy stratification during early succession in northern hardwoods.
1
1982
... 由枝和叶所构成的树冠是植物进行光合作用、呼吸作用、蒸腾作用等一系列生理活动的主要场所, 其大小、结构、形状及相互间的协变规律直接决定了植物个体内的生物量分配、生长活力和生产力.枝条作为植株的重要结构单元, 不仅是植株运输水分和营养物质的重要通道, 也是支撑树冠的主要结构并直接影响树冠的形状和大小, 同时它还是叶片的支撑体, 能通过其长度、结构、分布方式等变化来调整冠层内叶片的排布方式, 将叶伸展到适宜的空间形成一个合理的光拦截体系, 提高光合效率(黎云祥和刘玉成, 1996 ; Day et al ., 2002 ; Thomas & Winner, 2002 ; 张运春等, 2005 ).叶片是植物进行光合作用的主要器官, 叶片的数量、面积以及展叶效率等空间结构特征的变化直接影响着植株的光拦截效率和光合效率.因此, 枝条的结构特征及其与叶性状之间的协变规律反映植物对光照、空间等资源的利用和适应策略(Borchert & Slade, 1981 ; Bicknell, 1982 ; Boojh & Ramakrishnan, 1982 ; Bell, 1984 ; 陈波等, 2002 ). ...
Growth strategy of trees related to successional status. I. Architecture and extension growth.
1
1982
... 由枝和叶所构成的树冠是植物进行光合作用、呼吸作用、蒸腾作用等一系列生理活动的主要场所, 其大小、结构、形状及相互间的协变规律直接决定了植物个体内的生物量分配、生长活力和生产力.枝条作为植株的重要结构单元, 不仅是植株运输水分和营养物质的重要通道, 也是支撑树冠的主要结构并直接影响树冠的形状和大小, 同时它还是叶片的支撑体, 能通过其长度、结构、分布方式等变化来调整冠层内叶片的排布方式, 将叶伸展到适宜的空间形成一个合理的光拦截体系, 提高光合效率(黎云祥和刘玉成, 1996 ; Day et al ., 2002 ; Thomas & Winner, 2002 ; 张运春等, 2005 ).叶片是植物进行光合作用的主要器官, 叶片的数量、面积以及展叶效率等空间结构特征的变化直接影响着植株的光拦截效率和光合效率.因此, 枝条的结构特征及其与叶性状之间的协变规律反映植物对光照、空间等资源的利用和适应策略(Borchert & Slade, 1981 ; Bicknell, 1982 ; Boojh & Ramakrishnan, 1982 ; Bell, 1984 ; 陈波等, 2002 ). ...
Bifurcation ratios and the adaptive geometry of trees.
1
1981
... 由枝和叶所构成的树冠是植物进行光合作用、呼吸作用、蒸腾作用等一系列生理活动的主要场所, 其大小、结构、形状及相互间的协变规律直接决定了植物个体内的生物量分配、生长活力和生产力.枝条作为植株的重要结构单元, 不仅是植株运输水分和营养物质的重要通道, 也是支撑树冠的主要结构并直接影响树冠的形状和大小, 同时它还是叶片的支撑体, 能通过其长度、结构、分布方式等变化来调整冠层内叶片的排布方式, 将叶伸展到适宜的空间形成一个合理的光拦截体系, 提高光合效率(黎云祥和刘玉成, 1996 ; Day et al ., 2002 ; Thomas & Winner, 2002 ; 张运春等, 2005 ).叶片是植物进行光合作用的主要器官, 叶片的数量、面积以及展叶效率等空间结构特征的变化直接影响着植株的光拦截效率和光合效率.因此, 枝条的结构特征及其与叶性状之间的协变规律反映植物对光照、空间等资源的利用和适应策略(Borchert & Slade, 1981 ; Bicknell, 1982 ; Boojh & Ramakrishnan, 1982 ; Bell, 1984 ; 陈波等, 2002 ). ...
Shoot development in Betula papyrifera . IV. Comparisons between growth characteristics and expression of vegetative long and short shoots
1
1984
... 在木本植物中, 枝条的结构可以用枝条的茎长度、直径以及纤细率(枝条长度与直径比)来表示.对于用来着生和支撑叶、花和果实等器官的小枝而言, 其长度、纤细率以及展叶效率等性状的变化将直接影响植株的机械稳定性、光捕获效率和空间拓展能力, 进而影响到植株的光合和竞争能力的各个方面(Honda & Fisher, 1978 ; Niklas & Kerchner, 1984 ).例如, 树冠内叶片的分布模式是木本植物生产力的一个重要决定因素(Horn, 1971 ; Yagi & Kikuzawa, 1999 ), 而它的变化主要受小枝的结构变化模式、小枝在树冠内的空间排布方式以及小枝的个体大小等与枝条结构密切相关的因素限制(Koike, 1989 ; Yagi & Kikuzawa, 1999 ).在一些具有长短枝分化的物种中, 短枝的茎较短, 叶片排布更紧密, 长枝的茎较长, 叶片可以分散排布在较长的节间上(Yagi & Kikuzawa, 1999 ).尽管短枝上的总叶面积比长枝小(Gregory, 1980 ; Yagi & Kikuzawa, 1999 ), 但单位枝长基础上的叶面积比长枝大(Caesar & Macdonald, 1984 ; Yagi & Kikuzawa, 1999 ).小枝通过这种结构变化模式可以调控一个特定大小的小枝上的叶片数量(Maruyama, 1983 ; Koike, 1989 ; Takenaka, 1997 ; Yagi & Kikuzawa, 1999 )和展叶效率, 影响到植株的生产力.然而, 近年来, 关于植物小枝上茎-叶关系的研究多集中于茎直径维度的叶大小-小枝大小之间的关系研究(Westoby et al ., 2002 ; Pickup et al ., 2005 ; Wright et al ., 2007 ; Li et al ., 2008 ; Xiang et al ., 2009 ; Yang et al ., 2009 ; 杨冬梅等, 2012 ), 对于茎长维度的茎结构的另一维度——茎长度和纤细率对小枝展叶效率及相应的适应策略的研究则相对不足.小枝是个悬臂结构, 小枝茎长度不仅会影响其支撑叶片的排布, 还与小枝的机械安全密切相关.根据恒定胁迫模型(the constant-stress model)理论预测, 小枝茎横截面外缘所能承受的最大弯曲胁迫(M )主要由枝条长度(L )和枝条所承受的力(F )共同决定, 即M = L × F .对于一个无花、果等其他附属物的小枝而言, 枝条所承受的力主要来自风对枝条的压力和拉拽力所形成的风压胁迫, 而风压胁迫(F )又是叶面积(A )和风压(W )的乘积, 即F = A × W (Dean & Long, 1986 ).由此可知, 在风压一定的情况下, 小枝茎基部所能承受的最大弯曲胁迫是由枝条长度和叶面积决定的, 并且两者之间是负相关关系.所以, 我们预测小枝的展叶效率将随着小枝茎长度的增加而下降. ...
Summer and winter sensitivity of leaves and xylem to minimum freezing temperatures: A comparison of co-occurring Mediterranean oaks that differ in leaf lifespan.
2
2005
... 同时, 对于生活在同一群落里的常绿和落叶物种而言, 由于展叶物候和生物学习性的不同, 它们的功能特征通常也会有很大的差异.例如, 相比于落叶物种, 常绿物种通常具有长的叶寿命、低的比叶面积(SLA )、低的光合速率和高的养分含量(Reich et al ., 1992 ; Westoby et al ., 2002 ; Wright et al ., 2004 ; Li et al ., 2008 ); 另外, 由于常绿物种为了确保在冬季低温等不利环境下叶片不凋落, 需要水分运输系统保持畅通, 确保水分运输安全, 其木质部需要比落叶物种具有更强的抵抗冰冻引起栓塞的能力(Cavender-Bares & Holbrook, 2001 ; Cavender-Bares et al ., 2005 ), 致使其投入在支撑结构上的成本增加, 表现出在相同的茎投入时, 常绿物种通常比落叶物种具有更小的总叶面积和单叶面积(Sun et al ., 2006 ; Yang et al ., 2009 ).这种不同的成本-收益关系将有可能导致常绿和落叶物种采取不同的枝条伸展策略以改变小枝上的叶排布方式, 使两类物种都能够捕获足够的光能以保持相对稳定的碳收益.因此, 两类物种可能会具有不同的茎结构与展叶效率之间的变化关系. ...
... 常绿与落叶是植物通过长期进化适应形成的两种不同的生活史对策, 它们在叶寿命和叶片结构(如比叶面积)(Reich et al ., 1992 ; Westoby et al ., 2002 ; Wright et al ., 2004 ; Li et al ., 2008 )、茎木质部抗栓塞能力(Cavender-Bares & Holbrook, 2001 ; Cavender- Bares et al ., 2005 )等许多方面都显著不同.在本研究中, 虽然当年生小枝茎结构性状与表征展叶效率的性状之间的负相关关系斜率在常绿和落叶物种间无显著差异, 但截距差异显著.在3个表征展叶效率的性状中, 尽管常绿物种叶密度与小枝茎结构性状之间的关系截距显著大于落叶物种, 在某一给定的茎长或茎纤细率下, 常绿物种比落叶物种具有更多的叶数量, 即具有较高的出叶强度, 与以前相关研究发现的叶大小与叶数量之间是相互权衡的结果一致(Corner, 1949 ; Kleiman & Aarssen, 2007 ; Yang et al ., 2008 ).但由于常绿物种的单叶面积16.21 cm2 显著小于落叶物种的单叶面积34.66 cm2 (p < 0.001, 表2 ), 导致其叶面积比(单位茎质量的叶面积)和叶茎生物量比(单位茎质量的叶质量)与小枝茎结构性状之间的关系截距反而显著小于落叶物种, 即随着小枝茎长度和纤细率的增加, 常绿物种每单位质量的茎生物量投资获得的总叶面积和总叶质量都比落叶物种少, 常绿物种的实际展叶效率比落叶物种低.对数据结果进一步分析表明, 常绿物种更低展叶效率的主要原因是其比叶面积显著小于落叶物种 (p < 0.001, 表2 ), 因为我们的结果显示, 在不考虑小枝茎长结构影响的情况下, 茎叶生物量比在常绿和落叶物种中没有显著差异(p = 0.335, 表2 ), 表明总生物量在叶和茎之间的分配比例在常绿和落叶物种中是没有显著差异的, 可是落叶物种的单叶面积(p < 0.001, 表2 )和总叶面积(p = 0.001, 表2 )都要显著大于常绿物种, 并且常绿物种茎干质量与总叶面积相关关系共同斜率下的截距显著小于落叶物种(p ˂ 0.05, 表1 ), 即在某一给定的茎干质量下, 常绿物种比落叶物种具有更小的叶面积.许多研究也都发现常绿物种通常比落叶物种具有更高的比叶质量(即比叶面积的倒数)(Reich et al ., 1992 ; Westoby et al ., 2002 ; Wright et al ., 2004 ; 潘少安等, 2015 ).并且, Reich等(1998)发现比叶面积较低的植物常形成厚度较大而面积较小的叶片, 而刘艳芳(2015)在贡嘎山阔叶木本植物叶片解剖结构特征及其环境适应研究中发现的常绿植物的叶片厚度、栅栏组织厚度、海绵组织厚度、上表皮厚度和栅栏细胞层数都显著大于落叶植物的结果, 支持了Reich等(1998)的发现. ...
Hydraulic properties and freezing-induced cavitation in sympatric evergreen and deciduous oaks with contrasting habitats.
2
2001
... 同时, 对于生活在同一群落里的常绿和落叶物种而言, 由于展叶物候和生物学习性的不同, 它们的功能特征通常也会有很大的差异.例如, 相比于落叶物种, 常绿物种通常具有长的叶寿命、低的比叶面积(SLA )、低的光合速率和高的养分含量(Reich et al ., 1992 ; Westoby et al ., 2002 ; Wright et al ., 2004 ; Li et al ., 2008 ); 另外, 由于常绿物种为了确保在冬季低温等不利环境下叶片不凋落, 需要水分运输系统保持畅通, 确保水分运输安全, 其木质部需要比落叶物种具有更强的抵抗冰冻引起栓塞的能力(Cavender-Bares & Holbrook, 2001 ; Cavender-Bares et al ., 2005 ), 致使其投入在支撑结构上的成本增加, 表现出在相同的茎投入时, 常绿物种通常比落叶物种具有更小的总叶面积和单叶面积(Sun et al ., 2006 ; Yang et al ., 2009 ).这种不同的成本-收益关系将有可能导致常绿和落叶物种采取不同的枝条伸展策略以改变小枝上的叶排布方式, 使两类物种都能够捕获足够的光能以保持相对稳定的碳收益.因此, 两类物种可能会具有不同的茎结构与展叶效率之间的变化关系. ...
... 常绿与落叶是植物通过长期进化适应形成的两种不同的生活史对策, 它们在叶寿命和叶片结构(如比叶面积)(Reich et al ., 1992 ; Westoby et al ., 2002 ; Wright et al ., 2004 ; Li et al ., 2008 )、茎木质部抗栓塞能力(Cavender-Bares & Holbrook, 2001 ; Cavender- Bares et al ., 2005 )等许多方面都显著不同.在本研究中, 虽然当年生小枝茎结构性状与表征展叶效率的性状之间的负相关关系斜率在常绿和落叶物种间无显著差异, 但截距差异显著.在3个表征展叶效率的性状中, 尽管常绿物种叶密度与小枝茎结构性状之间的关系截距显著大于落叶物种, 在某一给定的茎长或茎纤细率下, 常绿物种比落叶物种具有更多的叶数量, 即具有较高的出叶强度, 与以前相关研究发现的叶大小与叶数量之间是相互权衡的结果一致(Corner, 1949 ; Kleiman & Aarssen, 2007 ; Yang et al ., 2008 ).但由于常绿物种的单叶面积16.21 cm2 显著小于落叶物种的单叶面积34.66 cm2 (p < 0.001, 表2 ), 导致其叶面积比(单位茎质量的叶面积)和叶茎生物量比(单位茎质量的叶质量)与小枝茎结构性状之间的关系截距反而显著小于落叶物种, 即随着小枝茎长度和纤细率的增加, 常绿物种每单位质量的茎生物量投资获得的总叶面积和总叶质量都比落叶物种少, 常绿物种的实际展叶效率比落叶物种低.对数据结果进一步分析表明, 常绿物种更低展叶效率的主要原因是其比叶面积显著小于落叶物种 (p < 0.001, 表2 ), 因为我们的结果显示, 在不考虑小枝茎长结构影响的情况下, 茎叶生物量比在常绿和落叶物种中没有显著差异(p = 0.335, 表2 ), 表明总生物量在叶和茎之间的分配比例在常绿和落叶物种中是没有显著差异的, 可是落叶物种的单叶面积(p < 0.001, 表2 )和总叶面积(p = 0.001, 表2 )都要显著大于常绿物种, 并且常绿物种茎干质量与总叶面积相关关系共同斜率下的截距显著小于落叶物种(p ˂ 0.05, 表1 ), 即在某一给定的茎干质量下, 常绿物种比落叶物种具有更小的叶面积.许多研究也都发现常绿物种通常比落叶物种具有更高的比叶质量(即比叶面积的倒数)(Reich et al ., 1992 ; Westoby et al ., 2002 ; Wright et al ., 2004 ; 潘少安等, 2015 ).并且, Reich等(1998)发现比叶面积较低的植物常形成厚度较大而面积较小的叶片, 而刘艳芳(2015)在贡嘎山阔叶木本植物叶片解剖结构特征及其环境适应研究中发现的常绿植物的叶片厚度、栅栏组织厚度、海绵组织厚度、上表皮厚度和栅栏细胞层数都显著大于落叶植物的结果, 支持了Reich等(1998)的发现. ...
木本植物的构型及其在植物生态学研究的进展
1
2002
... 由枝和叶所构成的树冠是植物进行光合作用、呼吸作用、蒸腾作用等一系列生理活动的主要场所, 其大小、结构、形状及相互间的协变规律直接决定了植物个体内的生物量分配、生长活力和生产力.枝条作为植株的重要结构单元, 不仅是植株运输水分和营养物质的重要通道, 也是支撑树冠的主要结构并直接影响树冠的形状和大小, 同时它还是叶片的支撑体, 能通过其长度、结构、分布方式等变化来调整冠层内叶片的排布方式, 将叶伸展到适宜的空间形成一个合理的光拦截体系, 提高光合效率(黎云祥和刘玉成, 1996 ; Day et al ., 2002 ; Thomas & Winner, 2002 ; 张运春等, 2005 ).叶片是植物进行光合作用的主要器官, 叶片的数量、面积以及展叶效率等空间结构特征的变化直接影响着植株的光拦截效率和光合效率.因此, 枝条的结构特征及其与叶性状之间的协变规律反映植物对光照、空间等资源的利用和适应策略(Borchert & Slade, 1981 ; Bicknell, 1982 ; Boojh & Ramakrishnan, 1982 ; Bell, 1984 ; 陈波等, 2002 ). ...
浙江省清凉峰自然保护区苔藓植物区系成分研究
2
2016
... 研究区位于浙江省清凉峰自然保护区, 地理位置118.87°-119.18° E, 30.08°-30.28° N, 主峰海拔 1β787.4 m.保护区地处中亚热带北缘, 气候复杂, 横跨亚热带和温带两个季风带, 具有明显的亚热带中山山地季风特征.主峰清凉峰则更具有典型的高山气候特点, 立体气候明显, 从山麓到山顶年平均气温从15.3 ℃降至7.8 ℃, 7月为最热月, 历年平均气温28.10 ℃, 1月为最冷月, 历年平均气温3.40 ℃.极端最高气温为41.90 ℃ (1996年8月6日), 极端最低气温为-13.30 ℃ (1967年1月16日).全年降水量 1β500-1β900 mm, 相对湿度在78%-82%之间.全山区年日照时间1β500-2 000 h, 年日照百分率为35%- 39%.海拔800-1 100 m区域年日照时间1 500-1 600 h (翁东明等, 2009 ; 程丽媛等, 2016 ). ...
... 保护区内植被带景观垂直层次分明, 群落内乔木层植物平均密度为2 062株·hm-2 , 灌木层平均密度为17 163株·hm-2 , 草本层平均密度为5 976株·hm-2 .灌木层植株密度基本是乔木层植株密度的8.3倍, 是草本层植株密度的2.9倍(翁东明等, 2009 ; 程丽媛等, 2016 ). ...
Effects of plant growth rate and leaf lifetime on the amount and type of anti-herbivore defense.
1
1988
... 常绿物种较低的比叶面积继而导致较低的单位茎结构基础上的展叶效率, 可能与其具有较长的叶寿命有关.一般而言, 叶寿命越长, 叶片越厚, 单位叶面积的建成消耗也越大, 叶子需要在足够长的时间内通过自身的光合生产所形成的有机碳来补偿初始的建成消耗(包括抵抗草食动物的防御构造), 并维持自身生存所需的物质消耗(Coley, 1988 ; Kikuzawa, 1991 ; Schulze et al ., 1994 ; Kikuzawa, 1995 ; 张林和罗天祥, 2004 ).所以叶寿命与比叶面积成反比(Reich et al ., 1992 ; Reich & Bowman, 1999 ; Ryser & Urbas, 2000 ; 张林和罗天祥, 2004 ).比叶面积较低的植物, 叶片中很大一部分物质用于构建保卫构造(防虫食、防止过度失水等)或者增加叶肉细胞密度以抵御环境胁迫(Reich et al ., 1998 ; 张林和罗天祥, 2004 ).与落叶物种相比, 常绿物种叶片在冬季不凋落, 需经受较长时间的冬季低温等不利生长环境, 面对冬季的选择压力, 常绿植物可能采取高消耗慢收益的保守型生存策略(Cornelissen, 1999 ; 刘艳芳, 2015 ), 通过革质的叶表皮和叶肉海绵组织分层等特点来增加抗寒力(董丽等, 2003 ; 刘艳芳, 2015 ).而落叶植物的叶寿命相对较短, 需要在相对短的生长时间内积累足够的光合产物以满足植株生长需要, 薄的叶片组织能够缩短CO2 从气孔扩散到叶绿体的路径, 也能增加气体交换的速率, 进而提高光合作用速率(Diemer, 1998 ; 刘艳芳, 2015 ), 所以, 较高的SLA 是落叶物种所采取的一种低投入快收益的积极型生存策略. ...
A triangular relationship between leaf size and seed size among woody species: Allometry, ontogeny, ecology and taxonomy.
1
1999
... 常绿物种较低的比叶面积继而导致较低的单位茎结构基础上的展叶效率, 可能与其具有较长的叶寿命有关.一般而言, 叶寿命越长, 叶片越厚, 单位叶面积的建成消耗也越大, 叶子需要在足够长的时间内通过自身的光合生产所形成的有机碳来补偿初始的建成消耗(包括抵抗草食动物的防御构造), 并维持自身生存所需的物质消耗(Coley, 1988 ; Kikuzawa, 1991 ; Schulze et al ., 1994 ; Kikuzawa, 1995 ; 张林和罗天祥, 2004 ).所以叶寿命与比叶面积成反比(Reich et al ., 1992 ; Reich & Bowman, 1999 ; Ryser & Urbas, 2000 ; 张林和罗天祥, 2004 ).比叶面积较低的植物, 叶片中很大一部分物质用于构建保卫构造(防虫食、防止过度失水等)或者增加叶肉细胞密度以抵御环境胁迫(Reich et al ., 1998 ; 张林和罗天祥, 2004 ).与落叶物种相比, 常绿物种叶片在冬季不凋落, 需经受较长时间的冬季低温等不利生长环境, 面对冬季的选择压力, 常绿植物可能采取高消耗慢收益的保守型生存策略(Cornelissen, 1999 ; 刘艳芳, 2015 ), 通过革质的叶表皮和叶肉海绵组织分层等特点来增加抗寒力(董丽等, 2003 ; 刘艳芳, 2015 ).而落叶植物的叶寿命相对较短, 需要在相对短的生长时间内积累足够的光合产物以满足植株生长需要, 薄的叶片组织能够缩短CO2 从气孔扩散到叶绿体的路径, 也能增加气体交换的速率, 进而提高光合作用速率(Diemer, 1998 ; 刘艳芳, 2015 ), 所以, 较高的SLA 是落叶物种所采取的一种低投入快收益的积极型生存策略. ...
The durian theory or the origin of the modern tree.
1
1949
... 常绿与落叶是植物通过长期进化适应形成的两种不同的生活史对策, 它们在叶寿命和叶片结构(如比叶面积)(Reich et al ., 1992 ; Westoby et al ., 2002 ; Wright et al ., 2004 ; Li et al ., 2008 )、茎木质部抗栓塞能力(Cavender-Bares & Holbrook, 2001 ; Cavender- Bares et al ., 2005 )等许多方面都显著不同.在本研究中, 虽然当年生小枝茎结构性状与表征展叶效率的性状之间的负相关关系斜率在常绿和落叶物种间无显著差异, 但截距差异显著.在3个表征展叶效率的性状中, 尽管常绿物种叶密度与小枝茎结构性状之间的关系截距显著大于落叶物种, 在某一给定的茎长或茎纤细率下, 常绿物种比落叶物种具有更多的叶数量, 即具有较高的出叶强度, 与以前相关研究发现的叶大小与叶数量之间是相互权衡的结果一致(Corner, 1949 ; Kleiman & Aarssen, 2007 ; Yang et al ., 2008 ).但由于常绿物种的单叶面积16.21 cm2 显著小于落叶物种的单叶面积34.66 cm2 (p < 0.001, 表2 ), 导致其叶面积比(单位茎质量的叶面积)和叶茎生物量比(单位茎质量的叶质量)与小枝茎结构性状之间的关系截距反而显著小于落叶物种, 即随着小枝茎长度和纤细率的增加, 常绿物种每单位质量的茎生物量投资获得的总叶面积和总叶质量都比落叶物种少, 常绿物种的实际展叶效率比落叶物种低.对数据结果进一步分析表明, 常绿物种更低展叶效率的主要原因是其比叶面积显著小于落叶物种 (p < 0.001, 表2 ), 因为我们的结果显示, 在不考虑小枝茎长结构影响的情况下, 茎叶生物量比在常绿和落叶物种中没有显著差异(p = 0.335, 表2 ), 表明总生物量在叶和茎之间的分配比例在常绿和落叶物种中是没有显著差异的, 可是落叶物种的单叶面积(p < 0.001, 表2 )和总叶面积(p = 0.001, 表2 )都要显著大于常绿物种, 并且常绿物种茎干质量与总叶面积相关关系共同斜率下的截距显著小于落叶物种(p ˂ 0.05, 表1 ), 即在某一给定的茎干质量下, 常绿物种比落叶物种具有更小的叶面积.许多研究也都发现常绿物种通常比落叶物种具有更高的比叶质量(即比叶面积的倒数)(Reich et al ., 1992 ; Westoby et al ., 2002 ; Wright et al ., 2004 ; 潘少安等, 2015 ).并且, Reich等(1998)发现比叶面积较低的植物常形成厚度较大而面积较小的叶片, 而刘艳芳(2015)在贡嘎山阔叶木本植物叶片解剖结构特征及其环境适应研究中发现的常绿植物的叶片厚度、栅栏组织厚度、海绵组织厚度、上表皮厚度和栅栏细胞层数都显著大于落叶植物的结果, 支持了Reich等(1998)的发现. ...
Age- and size- related trends in woody plant shoot development: Regulatory pathways and evidence for genetic control.
1
2002
... 由枝和叶所构成的树冠是植物进行光合作用、呼吸作用、蒸腾作用等一系列生理活动的主要场所, 其大小、结构、形状及相互间的协变规律直接决定了植物个体内的生物量分配、生长活力和生产力.枝条作为植株的重要结构单元, 不仅是植株运输水分和营养物质的重要通道, 也是支撑树冠的主要结构并直接影响树冠的形状和大小, 同时它还是叶片的支撑体, 能通过其长度、结构、分布方式等变化来调整冠层内叶片的排布方式, 将叶伸展到适宜的空间形成一个合理的光拦截体系, 提高光合效率(黎云祥和刘玉成, 1996 ; Day et al ., 2002 ; Thomas & Winner, 2002 ; 张运春等, 2005 ).叶片是植物进行光合作用的主要器官, 叶片的数量、面积以及展叶效率等空间结构特征的变化直接影响着植株的光拦截效率和光合效率.因此, 枝条的结构特征及其与叶性状之间的协变规律反映植物对光照、空间等资源的利用和适应策略(Borchert & Slade, 1981 ; Bicknell, 1982 ; Boojh & Ramakrishnan, 1982 ; Bell, 1984 ; 陈波等, 2002 ). ...
Validity of constant-stress and elastic-instability principles of stem formation in
2
1986
... 在木本植物中, 枝条的结构可以用枝条的茎长度、直径以及纤细率(枝条长度与直径比)来表示.对于用来着生和支撑叶、花和果实等器官的小枝而言, 其长度、纤细率以及展叶效率等性状的变化将直接影响植株的机械稳定性、光捕获效率和空间拓展能力, 进而影响到植株的光合和竞争能力的各个方面(Honda & Fisher, 1978 ; Niklas & Kerchner, 1984 ).例如, 树冠内叶片的分布模式是木本植物生产力的一个重要决定因素(Horn, 1971 ; Yagi & Kikuzawa, 1999 ), 而它的变化主要受小枝的结构变化模式、小枝在树冠内的空间排布方式以及小枝的个体大小等与枝条结构密切相关的因素限制(Koike, 1989 ; Yagi & Kikuzawa, 1999 ).在一些具有长短枝分化的物种中, 短枝的茎较短, 叶片排布更紧密, 长枝的茎较长, 叶片可以分散排布在较长的节间上(Yagi & Kikuzawa, 1999 ).尽管短枝上的总叶面积比长枝小(Gregory, 1980 ; Yagi & Kikuzawa, 1999 ), 但单位枝长基础上的叶面积比长枝大(Caesar & Macdonald, 1984 ; Yagi & Kikuzawa, 1999 ).小枝通过这种结构变化模式可以调控一个特定大小的小枝上的叶片数量(Maruyama, 1983 ; Koike, 1989 ; Takenaka, 1997 ; Yagi & Kikuzawa, 1999 )和展叶效率, 影响到植株的生产力.然而, 近年来, 关于植物小枝上茎-叶关系的研究多集中于茎直径维度的叶大小-小枝大小之间的关系研究(Westoby et al ., 2002 ; Pickup et al ., 2005 ; Wright et al ., 2007 ; Li et al ., 2008 ; Xiang et al ., 2009 ; Yang et al ., 2009 ; 杨冬梅等, 2012 ), 对于茎长维度的茎结构的另一维度——茎长度和纤细率对小枝展叶效率及相应的适应策略的研究则相对不足.小枝是个悬臂结构, 小枝茎长度不仅会影响其支撑叶片的排布, 还与小枝的机械安全密切相关.根据恒定胁迫模型(the constant-stress model)理论预测, 小枝茎横截面外缘所能承受的最大弯曲胁迫(M )主要由枝条长度(L )和枝条所承受的力(F )共同决定, 即M = L × F .对于一个无花、果等其他附属物的小枝而言, 枝条所承受的力主要来自风对枝条的压力和拉拽力所形成的风压胁迫, 而风压胁迫(F )又是叶面积(A )和风压(W )的乘积, 即F = A × W (Dean & Long, 1986 ).由此可知, 在风压一定的情况下, 小枝茎基部所能承受的最大弯曲胁迫是由枝条长度和叶面积决定的, 并且两者之间是负相关关系.所以, 我们预测小枝的展叶效率将随着小枝茎长度的增加而下降. ...
... 从机械力学的角度看, 展叶效率随小枝茎长度和茎纤细率的增加而下降可能是小枝在面对机械载重和风压胁迫时, 确保小枝安全的需要.作为起支撑作用的小枝, 首先要能够承受其自身的质量, 避免折断、撕裂或其他形式的机械损伤(Mcmahon, 1973 ; King & Loucks, 1978 ; King, 1981 , 1986 ; Long et al ., 1981 ; Givnish, 1986 ; Gartner, 1995 ), 必须能够抵抗来自茎及其上叶片质量所带来的静态和动态的压力和拉力胁迫.恒定胁迫模型(The constant-stress model)理论认为, 令植物茎横截面外缘弯曲所需要做的功(即弯曲矩, the bending moment) M 主要是由力F 和臂长(即茎长) L 共同决定, 即M = F × L .对于植物而言, 茎所承受的力是叶面积A 与风压W 的乘积, 即F = A × W (Dean & Long, 1986 ).所以, 随着小枝茎长度的增加, 在小枝上相同叶面积对小枝茎基部的机械负载会相应地增加, 特别是在有风吹动摇摆的情况下, 长枝将相应地减少枝条上的叶片面积或干质量来降低机械载重和风压胁迫, 导致展叶效率随小枝茎长度和茎纤细率的增加而下降.其次, 从植株树冠构建的角度看, 枝条结构与展叶效率之间的负相关关系将有利于植株更有效地捕获光能, 提高整体光合效率.因为相互遮阴是当年生小枝结构变化的一个重要决定因素, 更大的展叶密度将会导致当年生小枝内叶片之间相互荫蔽的程度增加(Niklas, 1988 ; Yagi & Kikuzawa, 1999 ).另外, 在总的生物量一定的情况下, 当植物将更多的生物量分配给茎以使枝条生长得更长时, 将有利于植株将更多的叶片伸展到冠层外部, 同时可以让一定数量的叶片分散在更长的枝条范围内, 降低同一枝条上叶片之间相互荫蔽的程度, 以利于捕获更多的光能, 提高植株的光合效率.综上所述, 枝条结构与展叶效率之间的负相关关系具有重要的生态学意义, 有利于保持小枝的机械安全, 并提高整个植株的光合效率. ...
Life span and dynamics of leaves of herbaceous perennials in high-elevation environments: “News from the elephant’s leg”.
1
1998
... 常绿物种较低的比叶面积继而导致较低的单位茎结构基础上的展叶效率, 可能与其具有较长的叶寿命有关.一般而言, 叶寿命越长, 叶片越厚, 单位叶面积的建成消耗也越大, 叶子需要在足够长的时间内通过自身的光合生产所形成的有机碳来补偿初始的建成消耗(包括抵抗草食动物的防御构造), 并维持自身生存所需的物质消耗(Coley, 1988 ; Kikuzawa, 1991 ; Schulze et al ., 1994 ; Kikuzawa, 1995 ; 张林和罗天祥, 2004 ).所以叶寿命与比叶面积成反比(Reich et al ., 1992 ; Reich & Bowman, 1999 ; Ryser & Urbas, 2000 ; 张林和罗天祥, 2004 ).比叶面积较低的植物, 叶片中很大一部分物质用于构建保卫构造(防虫食、防止过度失水等)或者增加叶肉细胞密度以抵御环境胁迫(Reich et al ., 1998 ; 张林和罗天祥, 2004 ).与落叶物种相比, 常绿物种叶片在冬季不凋落, 需经受较长时间的冬季低温等不利生长环境, 面对冬季的选择压力, 常绿植物可能采取高消耗慢收益的保守型生存策略(Cornelissen, 1999 ; 刘艳芳, 2015 ), 通过革质的叶表皮和叶肉海绵组织分层等特点来增加抗寒力(董丽等, 2003 ; 刘艳芳, 2015 ).而落叶植物的叶寿命相对较短, 需要在相对短的生长时间内积累足够的光合产物以满足植株生长需要, 薄的叶片组织能够缩短CO2 从气孔扩散到叶绿体的路径, 也能增加气体交换的速率, 进而提高光合作用速率(Diemer, 1998 ; 刘艳芳, 2015 ), 所以, 较高的SLA 是落叶物种所采取的一种低投入快收益的积极型生存策略. ...
常绿阔叶植物越冬期间叶片组织结构的适应性变化
1
2003
... 常绿物种较低的比叶面积继而导致较低的单位茎结构基础上的展叶效率, 可能与其具有较长的叶寿命有关.一般而言, 叶寿命越长, 叶片越厚, 单位叶面积的建成消耗也越大, 叶子需要在足够长的时间内通过自身的光合生产所形成的有机碳来补偿初始的建成消耗(包括抵抗草食动物的防御构造), 并维持自身生存所需的物质消耗(Coley, 1988 ; Kikuzawa, 1991 ; Schulze et al ., 1994 ; Kikuzawa, 1995 ; 张林和罗天祥, 2004 ).所以叶寿命与比叶面积成反比(Reich et al ., 1992 ; Reich & Bowman, 1999 ; Ryser & Urbas, 2000 ; 张林和罗天祥, 2004 ).比叶面积较低的植物, 叶片中很大一部分物质用于构建保卫构造(防虫食、防止过度失水等)或者增加叶肉细胞密度以抵御环境胁迫(Reich et al ., 1998 ; 张林和罗天祥, 2004 ).与落叶物种相比, 常绿物种叶片在冬季不凋落, 需经受较长时间的冬季低温等不利生长环境, 面对冬季的选择压力, 常绿植物可能采取高消耗慢收益的保守型生存策略(Cornelissen, 1999 ; 刘艳芳, 2015 ), 通过革质的叶表皮和叶肉海绵组织分层等特点来增加抗寒力(董丽等, 2003 ; 刘艳芳, 2015 ).而落叶植物的叶寿命相对较短, 需要在相对短的生长时间内积累足够的光合产物以满足植株生长需要, 薄的叶片组织能够缩短CO2 从气孔扩散到叶绿体的路径, 也能增加气体交换的速率, 进而提高光合作用速率(Diemer, 1998 ; 刘艳芳, 2015 ), 所以, 较高的SLA 是落叶物种所采取的一种低投入快收益的积极型生存策略. ...
1
... 对于植物构型性状以物种平均值进行分析.在对枝条结构与展叶效率的关系分析前, 首先将各参数的物种平均值进行对数转换(以10为底), 使之符合正态分布, 然后用关系方程y = bxa 来描述, 对数转换后为log(y ) = log(b ) + a log(x ), 其中x 和y 表示两个性状变量, 在双对数直角坐标系中, b 表示性状关系的截距, a 表示斜率, 即异速生长的指数(a = 1时为等速生长, 大于或小于1时为异速生长)(Harvey & Pagel, 1991 ).异速生长方程的参数估计采用标准化主轴估计(SMA)方法(Warton et al ., 2006 ), 由软件(S)MATR Version 2.0 (Falster et al ., 2006 )计算完成.回归斜率的置信区间根据(Pitman, 1939 )方法计算. ...
2
... 从机械力学的角度看, 展叶效率随小枝茎长度和茎纤细率的增加而下降可能是小枝在面对机械载重和风压胁迫时, 确保小枝安全的需要.作为起支撑作用的小枝, 首先要能够承受其自身的质量, 避免折断、撕裂或其他形式的机械损伤(Mcmahon, 1973 ; King & Loucks, 1978 ; King, 1981 , 1986 ; Long et al ., 1981 ; Givnish, 1986 ; Gartner, 1995 ), 必须能够抵抗来自茎及其上叶片质量所带来的静态和动态的压力和拉力胁迫.恒定胁迫模型(The constant-stress model)理论认为, 令植物茎横截面外缘弯曲所需要做的功(即弯曲矩, the bending moment) M 主要是由力F 和臂长(即茎长) L 共同决定, 即M = F × L .对于植物而言, 茎所承受的力是叶面积A 与风压W 的乘积, 即F = A × W (Dean & Long, 1986 ).所以, 随着小枝茎长度的增加, 在小枝上相同叶面积对小枝茎基部的机械负载会相应地增加, 特别是在有风吹动摇摆的情况下, 长枝将相应地减少枝条上的叶片面积或干质量来降低机械载重和风压胁迫, 导致展叶效率随小枝茎长度和茎纤细率的增加而下降.其次, 从植株树冠构建的角度看, 枝条结构与展叶效率之间的负相关关系将有利于植株更有效地捕获光能, 提高整体光合效率.因为相互遮阴是当年生小枝结构变化的一个重要决定因素, 更大的展叶密度将会导致当年生小枝内叶片之间相互荫蔽的程度增加(Niklas, 1988 ; Yagi & Kikuzawa, 1999 ).另外, 在总的生物量一定的情况下, 当植物将更多的生物量分配给茎以使枝条生长得更长时, 将有利于植株将更多的叶片伸展到冠层外部, 同时可以让一定数量的叶片分散在更长的枝条范围内, 降低同一枝条上叶片之间相互荫蔽的程度, 以利于捕获更多的光能, 提高植株的光合效率.综上所述, 枝条结构与展叶效率之间的负相关关系具有重要的生态学意义, 有利于保持小枝的机械安全, 并提高整个植株的光合效率. ...
... ; Gartner, 1995 ), 必须能够抵抗来自茎及其上叶片质量所带来的静态和动态的压力和拉力胁迫.恒定胁迫模型(The constant-stress model)理论认为, 令植物茎横截面外缘弯曲所需要做的功(即弯曲矩, the bending moment) M 主要是由力F 和臂长(即茎长) L 共同决定, 即M = F × L .对于植物而言, 茎所承受的力是叶面积A 与风压W 的乘积, 即F = A × W (Dean & Long, 1986 ).所以, 随着小枝茎长度的增加, 在小枝上相同叶面积对小枝茎基部的机械负载会相应地增加, 特别是在有风吹动摇摆的情况下, 长枝将相应地减少枝条上的叶片面积或干质量来降低机械载重和风压胁迫, 导致展叶效率随小枝茎长度和茎纤细率的增加而下降.其次, 从植株树冠构建的角度看, 枝条结构与展叶效率之间的负相关关系将有利于植株更有效地捕获光能, 提高整体光合效率.因为相互遮阴是当年生小枝结构变化的一个重要决定因素, 更大的展叶密度将会导致当年生小枝内叶片之间相互荫蔽的程度增加(Niklas, 1988 ; Yagi & Kikuzawa, 1999 ).另外, 在总的生物量一定的情况下, 当植物将更多的生物量分配给茎以使枝条生长得更长时, 将有利于植株将更多的叶片伸展到冠层外部, 同时可以让一定数量的叶片分散在更长的枝条范围内, 降低同一枝条上叶片之间相互荫蔽的程度, 以利于捕获更多的光能, 提高植株的光合效率.综上所述, 枝条结构与展叶效率之间的负相关关系具有重要的生态学意义, 有利于保持小枝的机械安全, 并提高整个植株的光合效率. ...
Biomechanical constraints on self-thinning in plant populations.
1
1986
... 在木本植物中, 枝条的结构可以用枝条的茎长度、直径以及纤细率(枝条长度与直径比)来表示.对于用来着生和支撑叶、花和果实等器官的小枝而言, 其长度、纤细率以及展叶效率等性状的变化将直接影响植株的机械稳定性、光捕获效率和空间拓展能力, 进而影响到植株的光合和竞争能力的各个方面(Honda & Fisher, 1978 ; Niklas & Kerchner, 1984 ).例如, 树冠内叶片的分布模式是木本植物生产力的一个重要决定因素(Horn, 1971 ; Yagi & Kikuzawa, 1999 ), 而它的变化主要受小枝的结构变化模式、小枝在树冠内的空间排布方式以及小枝的个体大小等与枝条结构密切相关的因素限制(Koike, 1989 ; Yagi & Kikuzawa, 1999 ).在一些具有长短枝分化的物种中, 短枝的茎较短, 叶片排布更紧密, 长枝的茎较长, 叶片可以分散排布在较长的节间上(Yagi & Kikuzawa, 1999 ).尽管短枝上的总叶面积比长枝小(Gregory, 1980 ; Yagi & Kikuzawa, 1999 ), 但单位枝长基础上的叶面积比长枝大(Caesar & Macdonald, 1984 ; Yagi & Kikuzawa, 1999 ).小枝通过这种结构变化模式可以调控一个特定大小的小枝上的叶片数量(Maruyama, 1983 ; Koike, 1989 ; Takenaka, 1997 ; Yagi & Kikuzawa, 1999 )和展叶效率, 影响到植株的生产力.然而, 近年来, 关于植物小枝上茎-叶关系的研究多集中于茎直径维度的叶大小-小枝大小之间的关系研究(Westoby et al ., 2002 ; Pickup et al ., 2005 ; Wright et al ., 2007 ; Li et al ., 2008 ; Xiang et al ., 2009 ; Yang et al ., 2009 ; 杨冬梅等, 2012 ), 对于茎长维度的茎结构的另一维度——茎长度和纤细率对小枝展叶效率及相应的适应策略的研究则相对不足.小枝是个悬臂结构, 小枝茎长度不仅会影响其支撑叶片的排布, 还与小枝的机械安全密切相关.根据恒定胁迫模型(the constant-stress model)理论预测, 小枝茎横截面外缘所能承受的最大弯曲胁迫(M )主要由枝条长度(L )和枝条所承受的力(F )共同决定, 即M = L × F .对于一个无花、果等其他附属物的小枝而言, 枝条所承受的力主要来自风对枝条的压力和拉拽力所形成的风压胁迫, 而风压胁迫(F )又是叶面积(A )和风压(W )的乘积, 即F = A × W (Dean & Long, 1986 ).由此可知, 在风压一定的情况下, 小枝茎基部所能承受的最大弯曲胁迫是由枝条长度和叶面积决定的, 并且两者之间是负相关关系.所以, 我们预测小枝的展叶效率将随着小枝茎长度的增加而下降. ...
Annual cycle of shoot development in sugar maple.
1980
2
... 在木本植物中, 枝条的结构可以用枝条的茎长度、直径以及纤细率(枝条长度与直径比)来表示.对于用来着生和支撑叶、花和果实等器官的小枝而言, 其长度、纤细率以及展叶效率等性状的变化将直接影响植株的机械稳定性、光捕获效率和空间拓展能力, 进而影响到植株的光合和竞争能力的各个方面(Honda & Fisher, 1978 ; Niklas & Kerchner, 1984 ).例如, 树冠内叶片的分布模式是木本植物生产力的一个重要决定因素(Horn, 1971 ; Yagi & Kikuzawa, 1999 ), 而它的变化主要受小枝的结构变化模式、小枝在树冠内的空间排布方式以及小枝的个体大小等与枝条结构密切相关的因素限制(Koike, 1989 ; Yagi & Kikuzawa, 1999 ).在一些具有长短枝分化的物种中, 短枝的茎较短, 叶片排布更紧密, 长枝的茎较长, 叶片可以分散排布在较长的节间上(Yagi & Kikuzawa, 1999 ).尽管短枝上的总叶面积比长枝小(Gregory, 1980 ; Yagi & Kikuzawa, 1999 ), 但单位枝长基础上的叶面积比长枝大(Caesar & Macdonald, 1984 ; Yagi & Kikuzawa, 1999 ).小枝通过这种结构变化模式可以调控一个特定大小的小枝上的叶片数量(Maruyama, 1983 ; Koike, 1989 ; Takenaka, 1997 ; Yagi & Kikuzawa, 1999 )和展叶效率, 影响到植株的生产力.然而, 近年来, 关于植物小枝上茎-叶关系的研究多集中于茎直径维度的叶大小-小枝大小之间的关系研究(Westoby et al ., 2002 ; Pickup et al ., 2005 ; Wright et al ., 2007 ; Li et al ., 2008 ; Xiang et al ., 2009 ; Yang et al ., 2009 ; 杨冬梅等, 2012 ), 对于茎长维度的茎结构的另一维度——茎长度和纤细率对小枝展叶效率及相应的适应策略的研究则相对不足.小枝是个悬臂结构, 小枝茎长度不仅会影响其支撑叶片的排布, 还与小枝的机械安全密切相关.根据恒定胁迫模型(the constant-stress model)理论预测, 小枝茎横截面外缘所能承受的最大弯曲胁迫(M )主要由枝条长度(L )和枝条所承受的力(F )共同决定, 即M = L × F .对于一个无花、果等其他附属物的小枝而言, 枝条所承受的力主要来自风对枝条的压力和拉拽力所形成的风压胁迫, 而风压胁迫(F )又是叶面积(A )和风压(W )的乘积, 即F = A × W (Dean & Long, 1986 ).由此可知, 在风压一定的情况下, 小枝茎基部所能承受的最大弯曲胁迫是由枝条长度和叶面积决定的, 并且两者之间是负相关关系.所以, 我们预测小枝的展叶效率将随着小枝茎长度的增加而下降. ...
... 对于植物构型性状以物种平均值进行分析.在对枝条结构与展叶效率的关系分析前, 首先将各参数的物种平均值进行对数转换(以10为底), 使之符合正态分布, 然后用关系方程y = bxa 来描述, 对数转换后为log(y ) = log(b ) + a log(x ), 其中x 和y 表示两个性状变量, 在双对数直角坐标系中, b 表示性状关系的截距, a 表示斜率, 即异速生长的指数(a = 1时为等速生长, 大于或小于1时为异速生长)(Harvey & Pagel, 1991 ).异速生长方程的参数估计采用标准化主轴估计(SMA)方法(Warton et al ., 2006 ), 由软件(S)MATR Version 2.0 (Falster et al ., 2006 )计算完成.回归斜率的置信区间根据(Pitman, 1939 )方法计算. ...
Tree branch angle: Maximizing effective leaf area.
1
1978
... 在木本植物中, 枝条的结构可以用枝条的茎长度、直径以及纤细率(枝条长度与直径比)来表示.对于用来着生和支撑叶、花和果实等器官的小枝而言, 其长度、纤细率以及展叶效率等性状的变化将直接影响植株的机械稳定性、光捕获效率和空间拓展能力, 进而影响到植株的光合和竞争能力的各个方面(Honda & Fisher, 1978 ; Niklas & Kerchner, 1984 ).例如, 树冠内叶片的分布模式是木本植物生产力的一个重要决定因素(Horn, 1971 ; Yagi & Kikuzawa, 1999 ), 而它的变化主要受小枝的结构变化模式、小枝在树冠内的空间排布方式以及小枝的个体大小等与枝条结构密切相关的因素限制(Koike, 1989 ; Yagi & Kikuzawa, 1999 ).在一些具有长短枝分化的物种中, 短枝的茎较短, 叶片排布更紧密, 长枝的茎较长, 叶片可以分散排布在较长的节间上(Yagi & Kikuzawa, 1999 ).尽管短枝上的总叶面积比长枝小(Gregory, 1980 ; Yagi & Kikuzawa, 1999 ), 但单位枝长基础上的叶面积比长枝大(Caesar & Macdonald, 1984 ; Yagi & Kikuzawa, 1999 ).小枝通过这种结构变化模式可以调控一个特定大小的小枝上的叶片数量(Maruyama, 1983 ; Koike, 1989 ; Takenaka, 1997 ; Yagi & Kikuzawa, 1999 )和展叶效率, 影响到植株的生产力.然而, 近年来, 关于植物小枝上茎-叶关系的研究多集中于茎直径维度的叶大小-小枝大小之间的关系研究(Westoby et al ., 2002 ; Pickup et al ., 2005 ; Wright et al ., 2007 ; Li et al ., 2008 ; Xiang et al ., 2009 ; Yang et al ., 2009 ; 杨冬梅等, 2012 ), 对于茎长维度的茎结构的另一维度——茎长度和纤细率对小枝展叶效率及相应的适应策略的研究则相对不足.小枝是个悬臂结构, 小枝茎长度不仅会影响其支撑叶片的排布, 还与小枝的机械安全密切相关.根据恒定胁迫模型(the constant-stress model)理论预测, 小枝茎横截面外缘所能承受的最大弯曲胁迫(M )主要由枝条长度(L )和枝条所承受的力(F )共同决定, 即M = L × F .对于一个无花、果等其他附属物的小枝而言, 枝条所承受的力主要来自风对枝条的压力和拉拽力所形成的风压胁迫, 而风压胁迫(F )又是叶面积(A )和风压(W )的乘积, 即F = A × W (Dean & Long, 1986 ).由此可知, 在风压一定的情况下, 小枝茎基部所能承受的最大弯曲胁迫是由枝条长度和叶面积决定的, 并且两者之间是负相关关系.所以, 我们预测小枝的展叶效率将随着小枝茎长度的增加而下降. ...
The adaptive geometry of trees.
1
1971
... 常绿物种较低的比叶面积继而导致较低的单位茎结构基础上的展叶效率, 可能与其具有较长的叶寿命有关.一般而言, 叶寿命越长, 叶片越厚, 单位叶面积的建成消耗也越大, 叶子需要在足够长的时间内通过自身的光合生产所形成的有机碳来补偿初始的建成消耗(包括抵抗草食动物的防御构造), 并维持自身生存所需的物质消耗(Coley, 1988 ; Kikuzawa, 1991 ; Schulze et al ., 1994 ; Kikuzawa, 1995 ; 张林和罗天祥, 2004 ).所以叶寿命与比叶面积成反比(Reich et al ., 1992 ; Reich & Bowman, 1999 ; Ryser & Urbas, 2000 ; 张林和罗天祥, 2004 ).比叶面积较低的植物, 叶片中很大一部分物质用于构建保卫构造(防虫食、防止过度失水等)或者增加叶肉细胞密度以抵御环境胁迫(Reich et al ., 1998 ; 张林和罗天祥, 2004 ).与落叶物种相比, 常绿物种叶片在冬季不凋落, 需经受较长时间的冬季低温等不利生长环境, 面对冬季的选择压力, 常绿植物可能采取高消耗慢收益的保守型生存策略(Cornelissen, 1999 ; 刘艳芳, 2015 ), 通过革质的叶表皮和叶肉海绵组织分层等特点来增加抗寒力(董丽等, 2003 ; 刘艳芳, 2015 ).而落叶植物的叶寿命相对较短, 需要在相对短的生长时间内积累足够的光合产物以满足植株生长需要, 薄的叶片组织能够缩短CO2 从气孔扩散到叶绿体的路径, 也能增加气体交换的速率, 进而提高光合作用速率(Diemer, 1998 ; 刘艳芳, 2015 ), 所以, 较高的SLA 是落叶物种所采取的一种低投入快收益的积极型生存策略. ...
A cost-benefit analysis of leaf habit and leaf longevity of trees and their geographical pattern.
1
1991
... 常绿物种较低的比叶面积继而导致较低的单位茎结构基础上的展叶效率, 可能与其具有较长的叶寿命有关.一般而言, 叶寿命越长, 叶片越厚, 单位叶面积的建成消耗也越大, 叶子需要在足够长的时间内通过自身的光合生产所形成的有机碳来补偿初始的建成消耗(包括抵抗草食动物的防御构造), 并维持自身生存所需的物质消耗(Coley, 1988 ; Kikuzawa, 1991 ; Schulze et al ., 1994 ; Kikuzawa, 1995 ; 张林和罗天祥, 2004 ).所以叶寿命与比叶面积成反比(Reich et al ., 1992 ; Reich & Bowman, 1999 ; Ryser & Urbas, 2000 ; 张林和罗天祥, 2004 ).比叶面积较低的植物, 叶片中很大一部分物质用于构建保卫构造(防虫食、防止过度失水等)或者增加叶肉细胞密度以抵御环境胁迫(Reich et al ., 1998 ; 张林和罗天祥, 2004 ).与落叶物种相比, 常绿物种叶片在冬季不凋落, 需经受较长时间的冬季低温等不利生长环境, 面对冬季的选择压力, 常绿植物可能采取高消耗慢收益的保守型生存策略(Cornelissen, 1999 ; 刘艳芳, 2015 ), 通过革质的叶表皮和叶肉海绵组织分层等特点来增加抗寒力(董丽等, 2003 ; 刘艳芳, 2015 ).而落叶植物的叶寿命相对较短, 需要在相对短的生长时间内积累足够的光合产物以满足植株生长需要, 薄的叶片组织能够缩短CO2 从气孔扩散到叶绿体的路径, 也能增加气体交换的速率, 进而提高光合作用速率(Diemer, 1998 ; 刘艳芳, 2015 ), 所以, 较高的SLA 是落叶物种所采取的一种低投入快收益的积极型生存策略. ...
Leaf phenology as an optimal strategy for carbon gain in plants.
1
1995
... 从机械力学的角度看, 展叶效率随小枝茎长度和茎纤细率的增加而下降可能是小枝在面对机械载重和风压胁迫时, 确保小枝安全的需要.作为起支撑作用的小枝, 首先要能够承受其自身的质量, 避免折断、撕裂或其他形式的机械损伤(Mcmahon, 1973 ; King & Loucks, 1978 ; King, 1981 , 1986 ; Long et al ., 1981 ; Givnish, 1986 ; Gartner, 1995 ), 必须能够抵抗来自茎及其上叶片质量所带来的静态和动态的压力和拉力胁迫.恒定胁迫模型(The constant-stress model)理论认为, 令植物茎横截面外缘弯曲所需要做的功(即弯曲矩, the bending moment) M 主要是由力F 和臂长(即茎长) L 共同决定, 即M = F × L .对于植物而言, 茎所承受的力是叶面积A 与风压W 的乘积, 即F = A × W (Dean & Long, 1986 ).所以, 随着小枝茎长度的增加, 在小枝上相同叶面积对小枝茎基部的机械负载会相应地增加, 特别是在有风吹动摇摆的情况下, 长枝将相应地减少枝条上的叶片面积或干质量来降低机械载重和风压胁迫, 导致展叶效率随小枝茎长度和茎纤细率的增加而下降.其次, 从植株树冠构建的角度看, 枝条结构与展叶效率之间的负相关关系将有利于植株更有效地捕获光能, 提高整体光合效率.因为相互遮阴是当年生小枝结构变化的一个重要决定因素, 更大的展叶密度将会导致当年生小枝内叶片之间相互荫蔽的程度增加(Niklas, 1988 ; Yagi & Kikuzawa, 1999 ).另外, 在总的生物量一定的情况下, 当植物将更多的生物量分配给茎以使枝条生长得更长时, 将有利于植株将更多的叶片伸展到冠层外部, 同时可以让一定数量的叶片分散在更长的枝条范围内, 降低同一枝条上叶片之间相互荫蔽的程度, 以利于捕获更多的光能, 提高植株的光合效率.综上所述, 枝条结构与展叶效率之间的负相关关系具有重要的生态学意义, 有利于保持小枝的机械安全, 并提高整个植株的光合效率. ...
Tree dimensions: Maximizing the rate of height growth in dense stands.
1
1981
... 从机械力学的角度看, 展叶效率随小枝茎长度和茎纤细率的增加而下降可能是小枝在面对机械载重和风压胁迫时, 确保小枝安全的需要.作为起支撑作用的小枝, 首先要能够承受其自身的质量, 避免折断、撕裂或其他形式的机械损伤(Mcmahon, 1973 ; King & Loucks, 1978 ; King, 1981 , 1986 ; Long et al ., 1981 ; Givnish, 1986 ; Gartner, 1995 ), 必须能够抵抗来自茎及其上叶片质量所带来的静态和动态的压力和拉力胁迫.恒定胁迫模型(The constant-stress model)理论认为, 令植物茎横截面外缘弯曲所需要做的功(即弯曲矩, the bending moment) M 主要是由力F 和臂长(即茎长) L 共同决定, 即M = F × L .对于植物而言, 茎所承受的力是叶面积A 与风压W 的乘积, 即F = A × W (Dean & Long, 1986 ).所以, 随着小枝茎长度的增加, 在小枝上相同叶面积对小枝茎基部的机械负载会相应地增加, 特别是在有风吹动摇摆的情况下, 长枝将相应地减少枝条上的叶片面积或干质量来降低机械载重和风压胁迫, 导致展叶效率随小枝茎长度和茎纤细率的增加而下降.其次, 从植株树冠构建的角度看, 枝条结构与展叶效率之间的负相关关系将有利于植株更有效地捕获光能, 提高整体光合效率.因为相互遮阴是当年生小枝结构变化的一个重要决定因素, 更大的展叶密度将会导致当年生小枝内叶片之间相互荫蔽的程度增加(Niklas, 1988 ; Yagi & Kikuzawa, 1999 ).另外, 在总的生物量一定的情况下, 当植物将更多的生物量分配给茎以使枝条生长得更长时, 将有利于植株将更多的叶片伸展到冠层外部, 同时可以让一定数量的叶片分散在更长的枝条范围内, 降低同一枝条上叶片之间相互荫蔽的程度, 以利于捕获更多的光能, 提高植株的光合效率.综上所述, 枝条结构与展叶效率之间的负相关关系具有重要的生态学意义, 有利于保持小枝的机械安全, 并提高整个植株的光合效率. ...
The theory of tree bole and branch form.
1
1978
... 从机械力学的角度看, 展叶效率随小枝茎长度和茎纤细率的增加而下降可能是小枝在面对机械载重和风压胁迫时, 确保小枝安全的需要.作为起支撑作用的小枝, 首先要能够承受其自身的质量, 避免折断、撕裂或其他形式的机械损伤(Mcmahon, 1973 ; King & Loucks, 1978 ; King, 1981 , 1986 ; Long et al ., 1981 ; Givnish, 1986 ; Gartner, 1995 ), 必须能够抵抗来自茎及其上叶片质量所带来的静态和动态的压力和拉力胁迫.恒定胁迫模型(The constant-stress model)理论认为, 令植物茎横截面外缘弯曲所需要做的功(即弯曲矩, the bending moment) M 主要是由力F 和臂长(即茎长) L 共同决定, 即M = F × L .对于植物而言, 茎所承受的力是叶面积A 与风压W 的乘积, 即F = A × W (Dean & Long, 1986 ).所以, 随着小枝茎长度的增加, 在小枝上相同叶面积对小枝茎基部的机械负载会相应地增加, 特别是在有风吹动摇摆的情况下, 长枝将相应地减少枝条上的叶片面积或干质量来降低机械载重和风压胁迫, 导致展叶效率随小枝茎长度和茎纤细率的增加而下降.其次, 从植株树冠构建的角度看, 枝条结构与展叶效率之间的负相关关系将有利于植株更有效地捕获光能, 提高整体光合效率.因为相互遮阴是当年生小枝结构变化的一个重要决定因素, 更大的展叶密度将会导致当年生小枝内叶片之间相互荫蔽的程度增加(Niklas, 1988 ; Yagi & Kikuzawa, 1999 ).另外, 在总的生物量一定的情况下, 当植物将更多的生物量分配给茎以使枝条生长得更长时, 将有利于植株将更多的叶片伸展到冠层外部, 同时可以让一定数量的叶片分散在更长的枝条范围内, 降低同一枝条上叶片之间相互荫蔽的程度, 以利于捕获更多的光能, 提高植株的光合效率.综上所述, 枝条结构与展叶效率之间的负相关关系具有重要的生态学意义, 有利于保持小枝的机械安全, 并提高整个植株的光合效率. ...
Tree form, height growth, and susceptibility to wind damage in
1
1986
... 常绿与落叶是植物通过长期进化适应形成的两种不同的生活史对策, 它们在叶寿命和叶片结构(如比叶面积)(Reich et al ., 1992 ; Westoby et al ., 2002 ; Wright et al ., 2004 ; Li et al ., 2008 )、茎木质部抗栓塞能力(Cavender-Bares & Holbrook, 2001 ; Cavender- Bares et al ., 2005 )等许多方面都显著不同.在本研究中, 虽然当年生小枝茎结构性状与表征展叶效率的性状之间的负相关关系斜率在常绿和落叶物种间无显著差异, 但截距差异显著.在3个表征展叶效率的性状中, 尽管常绿物种叶密度与小枝茎结构性状之间的关系截距显著大于落叶物种, 在某一给定的茎长或茎纤细率下, 常绿物种比落叶物种具有更多的叶数量, 即具有较高的出叶强度, 与以前相关研究发现的叶大小与叶数量之间是相互权衡的结果一致(Corner, 1949 ; Kleiman & Aarssen, 2007 ; Yang et al ., 2008 ).但由于常绿物种的单叶面积16.21 cm2 显著小于落叶物种的单叶面积34.66 cm2 (p < 0.001, 表2 ), 导致其叶面积比(单位茎质量的叶面积)和叶茎生物量比(单位茎质量的叶质量)与小枝茎结构性状之间的关系截距反而显著小于落叶物种, 即随着小枝茎长度和纤细率的增加, 常绿物种每单位质量的茎生物量投资获得的总叶面积和总叶质量都比落叶物种少, 常绿物种的实际展叶效率比落叶物种低.对数据结果进一步分析表明, 常绿物种更低展叶效率的主要原因是其比叶面积显著小于落叶物种 (p < 0.001, 表2 ), 因为我们的结果显示, 在不考虑小枝茎长结构影响的情况下, 茎叶生物量比在常绿和落叶物种中没有显著差异(p = 0.335, 表2 ), 表明总生物量在叶和茎之间的分配比例在常绿和落叶物种中是没有显著差异的, 可是落叶物种的单叶面积(p < 0.001, 表2 )和总叶面积(p = 0.001, 表2 )都要显著大于常绿物种, 并且常绿物种茎干质量与总叶面积相关关系共同斜率下的截距显著小于落叶物种(p ˂ 0.05, 表1 ), 即在某一给定的茎干质量下, 常绿物种比落叶物种具有更小的叶面积.许多研究也都发现常绿物种通常比落叶物种具有更高的比叶质量(即比叶面积的倒数)(Reich et al ., 1992 ; Westoby et al ., 2002 ; Wright et al ., 2004 ; 潘少安等, 2015 ).并且, Reich等(1998)发现比叶面积较低的植物常形成厚度较大而面积较小的叶片, 而刘艳芳(2015)在贡嘎山阔叶木本植物叶片解剖结构特征及其环境适应研究中发现的常绿植物的叶片厚度、栅栏组织厚度、海绵组织厚度、上表皮厚度和栅栏细胞层数都显著大于落叶植物的结果, 支持了Reich等(1998)的发现. ...
The leaf size/number trade-off in trees.
2
2007
... 在木本植物中, 枝条的结构可以用枝条的茎长度、直径以及纤细率(枝条长度与直径比)来表示.对于用来着生和支撑叶、花和果实等器官的小枝而言, 其长度、纤细率以及展叶效率等性状的变化将直接影响植株的机械稳定性、光捕获效率和空间拓展能力, 进而影响到植株的光合和竞争能力的各个方面(Honda & Fisher, 1978 ; Niklas & Kerchner, 1984 ).例如, 树冠内叶片的分布模式是木本植物生产力的一个重要决定因素(Horn, 1971 ; Yagi & Kikuzawa, 1999 ), 而它的变化主要受小枝的结构变化模式、小枝在树冠内的空间排布方式以及小枝的个体大小等与枝条结构密切相关的因素限制(Koike, 1989 ; Yagi & Kikuzawa, 1999 ).在一些具有长短枝分化的物种中, 短枝的茎较短, 叶片排布更紧密, 长枝的茎较长, 叶片可以分散排布在较长的节间上(Yagi & Kikuzawa, 1999 ).尽管短枝上的总叶面积比长枝小(Gregory, 1980 ; Yagi & Kikuzawa, 1999 ), 但单位枝长基础上的叶面积比长枝大(Caesar & Macdonald, 1984 ; Yagi & Kikuzawa, 1999 ).小枝通过这种结构变化模式可以调控一个特定大小的小枝上的叶片数量(Maruyama, 1983 ; Koike, 1989 ; Takenaka, 1997 ; Yagi & Kikuzawa, 1999 )和展叶效率, 影响到植株的生产力.然而, 近年来, 关于植物小枝上茎-叶关系的研究多集中于茎直径维度的叶大小-小枝大小之间的关系研究(Westoby et al ., 2002 ; Pickup et al ., 2005 ; Wright et al ., 2007 ; Li et al ., 2008 ; Xiang et al ., 2009 ; Yang et al ., 2009 ; 杨冬梅等, 2012 ), 对于茎长维度的茎结构的另一维度——茎长度和纤细率对小枝展叶效率及相应的适应策略的研究则相对不足.小枝是个悬臂结构, 小枝茎长度不仅会影响其支撑叶片的排布, 还与小枝的机械安全密切相关.根据恒定胁迫模型(the constant-stress model)理论预测, 小枝茎横截面外缘所能承受的最大弯曲胁迫(M )主要由枝条长度(L )和枝条所承受的力(F )共同决定, 即M = L × F .对于一个无花、果等其他附属物的小枝而言, 枝条所承受的力主要来自风对枝条的压力和拉拽力所形成的风压胁迫, 而风压胁迫(F )又是叶面积(A )和风压(W )的乘积, 即F = A × W (Dean & Long, 1986 ).由此可知, 在风压一定的情况下, 小枝茎基部所能承受的最大弯曲胁迫是由枝条长度和叶面积决定的, 并且两者之间是负相关关系.所以, 我们预测小枝的展叶效率将随着小枝茎长度的增加而下降. ...
... ; Koike, 1989 ; Takenaka, 1997 ; Yagi & Kikuzawa, 1999 )和展叶效率, 影响到植株的生产力.然而, 近年来, 关于植物小枝上茎-叶关系的研究多集中于茎直径维度的叶大小-小枝大小之间的关系研究(Westoby et al ., 2002 ; Pickup et al ., 2005 ; Wright et al ., 2007 ; Li et al ., 2008 ; Xiang et al ., 2009 ; Yang et al ., 2009 ; 杨冬梅等, 2012 ), 对于茎长维度的茎结构的另一维度——茎长度和纤细率对小枝展叶效率及相应的适应策略的研究则相对不足.小枝是个悬臂结构, 小枝茎长度不仅会影响其支撑叶片的排布, 还与小枝的机械安全密切相关.根据恒定胁迫模型(the constant-stress model)理论预测, 小枝茎横截面外缘所能承受的最大弯曲胁迫(M )主要由枝条长度(L )和枝条所承受的力(F )共同决定, 即M = L × F .对于一个无花、果等其他附属物的小枝而言, 枝条所承受的力主要来自风对枝条的压力和拉拽力所形成的风压胁迫, 而风压胁迫(F )又是叶面积(A )和风压(W )的乘积, 即F = A × W (Dean & Long, 1986 ).由此可知, 在风压一定的情况下, 小枝茎基部所能承受的最大弯曲胁迫是由枝条长度和叶面积决定的, 并且两者之间是负相关关系.所以, 我们预测小枝的展叶效率将随着小枝茎长度的增加而下降. ...
Foliage-crown development and interaction in
3
1989
... 在木本植物中, 枝条的结构可以用枝条的茎长度、直径以及纤细率(枝条长度与直径比)来表示.对于用来着生和支撑叶、花和果实等器官的小枝而言, 其长度、纤细率以及展叶效率等性状的变化将直接影响植株的机械稳定性、光捕获效率和空间拓展能力, 进而影响到植株的光合和竞争能力的各个方面(Honda & Fisher, 1978 ; Niklas & Kerchner, 1984 ).例如, 树冠内叶片的分布模式是木本植物生产力的一个重要决定因素(Horn, 1971 ; Yagi & Kikuzawa, 1999 ), 而它的变化主要受小枝的结构变化模式、小枝在树冠内的空间排布方式以及小枝的个体大小等与枝条结构密切相关的因素限制(Koike, 1989 ; Yagi & Kikuzawa, 1999 ).在一些具有长短枝分化的物种中, 短枝的茎较短, 叶片排布更紧密, 长枝的茎较长, 叶片可以分散排布在较长的节间上(Yagi & Kikuzawa, 1999 ).尽管短枝上的总叶面积比长枝小(Gregory, 1980 ; Yagi & Kikuzawa, 1999 ), 但单位枝长基础上的叶面积比长枝大(Caesar & Macdonald, 1984 ; Yagi & Kikuzawa, 1999 ).小枝通过这种结构变化模式可以调控一个特定大小的小枝上的叶片数量(Maruyama, 1983 ; Koike, 1989 ; Takenaka, 1997 ; Yagi & Kikuzawa, 1999 )和展叶效率, 影响到植株的生产力.然而, 近年来, 关于植物小枝上茎-叶关系的研究多集中于茎直径维度的叶大小-小枝大小之间的关系研究(Westoby et al ., 2002 ; Pickup et al ., 2005 ; Wright et al ., 2007 ; Li et al ., 2008 ; Xiang et al ., 2009 ; Yang et al ., 2009 ; 杨冬梅等, 2012 ), 对于茎长维度的茎结构的另一维度——茎长度和纤细率对小枝展叶效率及相应的适应策略的研究则相对不足.小枝是个悬臂结构, 小枝茎长度不仅会影响其支撑叶片的排布, 还与小枝的机械安全密切相关.根据恒定胁迫模型(the constant-stress model)理论预测, 小枝茎横截面外缘所能承受的最大弯曲胁迫(M )主要由枝条长度(L )和枝条所承受的力(F )共同决定, 即M = L × F .对于一个无花、果等其他附属物的小枝而言, 枝条所承受的力主要来自风对枝条的压力和拉拽力所形成的风压胁迫, 而风压胁迫(F )又是叶面积(A )和风压(W )的乘积, 即F = A × W (Dean & Long, 1986 ).由此可知, 在风压一定的情况下, 小枝茎基部所能承受的最大弯曲胁迫是由枝条长度和叶面积决定的, 并且两者之间是负相关关系.所以, 我们预测小枝的展叶效率将随着小枝茎长度的增加而下降. ...
... 同时, 对于生活在同一群落里的常绿和落叶物种而言, 由于展叶物候和生物学习性的不同, 它们的功能特征通常也会有很大的差异.例如, 相比于落叶物种, 常绿物种通常具有长的叶寿命、低的比叶面积(SLA )、低的光合速率和高的养分含量(Reich et al ., 1992 ; Westoby et al ., 2002 ; Wright et al ., 2004 ; Li et al ., 2008 ); 另外, 由于常绿物种为了确保在冬季低温等不利环境下叶片不凋落, 需要水分运输系统保持畅通, 确保水分运输安全, 其木质部需要比落叶物种具有更强的抵抗冰冻引起栓塞的能力(Cavender-Bares & Holbrook, 2001 ; Cavender-Bares et al ., 2005 ), 致使其投入在支撑结构上的成本增加, 表现出在相同的茎投入时, 常绿物种通常比落叶物种具有更小的总叶面积和单叶面积(Sun et al ., 2006 ; Yang et al ., 2009 ).这种不同的成本-收益关系将有可能导致常绿和落叶物种采取不同的枝条伸展策略以改变小枝上的叶排布方式, 使两类物种都能够捕获足够的光能以保持相对稳定的碳收益.因此, 两类物种可能会具有不同的茎结构与展叶效率之间的变化关系. ...
... 常绿与落叶是植物通过长期进化适应形成的两种不同的生活史对策, 它们在叶寿命和叶片结构(如比叶面积)(Reich et al ., 1992 ; Westoby et al ., 2002 ; Wright et al ., 2004 ; Li et al ., 2008 )、茎木质部抗栓塞能力(Cavender-Bares & Holbrook, 2001 ; Cavender- Bares et al ., 2005 )等许多方面都显著不同.在本研究中, 虽然当年生小枝茎结构性状与表征展叶效率的性状之间的负相关关系斜率在常绿和落叶物种间无显著差异, 但截距差异显著.在3个表征展叶效率的性状中, 尽管常绿物种叶密度与小枝茎结构性状之间的关系截距显著大于落叶物种, 在某一给定的茎长或茎纤细率下, 常绿物种比落叶物种具有更多的叶数量, 即具有较高的出叶强度, 与以前相关研究发现的叶大小与叶数量之间是相互权衡的结果一致(Corner, 1949 ; Kleiman & Aarssen, 2007 ; Yang et al ., 2008 ).但由于常绿物种的单叶面积16.21 cm2 显著小于落叶物种的单叶面积34.66 cm2 (p < 0.001, 表2 ), 导致其叶面积比(单位茎质量的叶面积)和叶茎生物量比(单位茎质量的叶质量)与小枝茎结构性状之间的关系截距反而显著小于落叶物种, 即随着小枝茎长度和纤细率的增加, 常绿物种每单位质量的茎生物量投资获得的总叶面积和总叶质量都比落叶物种少, 常绿物种的实际展叶效率比落叶物种低.对数据结果进一步分析表明, 常绿物种更低展叶效率的主要原因是其比叶面积显著小于落叶物种 (p < 0.001, 表2 ), 因为我们的结果显示, 在不考虑小枝茎长结构影响的情况下, 茎叶生物量比在常绿和落叶物种中没有显著差异(p = 0.335, 表2 ), 表明总生物量在叶和茎之间的分配比例在常绿和落叶物种中是没有显著差异的, 可是落叶物种的单叶面积(p < 0.001, 表2 )和总叶面积(p = 0.001, 表2 )都要显著大于常绿物种, 并且常绿物种茎干质量与总叶面积相关关系共同斜率下的截距显著小于落叶物种(p ˂ 0.05, 表1 ), 即在某一给定的茎干质量下, 常绿物种比落叶物种具有更小的叶面积.许多研究也都发现常绿物种通常比落叶物种具有更高的比叶质量(即比叶面积的倒数)(Reich et al ., 1992 ; Westoby et al ., 2002 ; Wright et al ., 2004 ; 潘少安等, 2015 ).并且, Reich等(1998)发现比叶面积较低的植物常形成厚度较大而面积较小的叶片, 而刘艳芳(2015)在贡嘎山阔叶木本植物叶片解剖结构特征及其环境适应研究中发现的常绿植物的叶片厚度、栅栏组织厚度、海绵组织厚度、上表皮厚度和栅栏细胞层数都显著大于落叶植物的结果, 支持了Reich等(1998)的发现. ...
Allometric relationships between lamina area, lamina mass and petiole mass of 93 temperate woody species vary with leaf habit, leaf form and altitude.
1
2008
... 由枝和叶所构成的树冠是植物进行光合作用、呼吸作用、蒸腾作用等一系列生理活动的主要场所, 其大小、结构、形状及相互间的协变规律直接决定了植物个体内的生物量分配、生长活力和生产力.枝条作为植株的重要结构单元, 不仅是植株运输水分和营养物质的重要通道, 也是支撑树冠的主要结构并直接影响树冠的形状和大小, 同时它还是叶片的支撑体, 能通过其长度、结构、分布方式等变化来调整冠层内叶片的排布方式, 将叶伸展到适宜的空间形成一个合理的光拦截体系, 提高光合效率(黎云祥和刘玉成, 1996 ; Day et al ., 2002 ; Thomas & Winner, 2002 ; 张运春等, 2005 ).叶片是植物进行光合作用的主要器官, 叶片的数量、面积以及展叶效率等空间结构特征的变化直接影响着植株的光拦截效率和光合效率.因此, 枝条的结构特征及其与叶性状之间的协变规律反映植物对光照、空间等资源的利用和适应策略(Borchert & Slade, 1981 ; Bicknell, 1982 ; Boojh & Ramakrishnan, 1982 ; Bell, 1984 ; 陈波等, 2002 ). ...
四川大头茶苗木构件水平的邻体干扰效应
3
1996
... 常绿物种较低的比叶面积继而导致较低的单位茎结构基础上的展叶效率, 可能与其具有较长的叶寿命有关.一般而言, 叶寿命越长, 叶片越厚, 单位叶面积的建成消耗也越大, 叶子需要在足够长的时间内通过自身的光合生产所形成的有机碳来补偿初始的建成消耗(包括抵抗草食动物的防御构造), 并维持自身生存所需的物质消耗(Coley, 1988 ; Kikuzawa, 1991 ; Schulze et al ., 1994 ; Kikuzawa, 1995 ; 张林和罗天祥, 2004 ).所以叶寿命与比叶面积成反比(Reich et al ., 1992 ; Reich & Bowman, 1999 ; Ryser & Urbas, 2000 ; 张林和罗天祥, 2004 ).比叶面积较低的植物, 叶片中很大一部分物质用于构建保卫构造(防虫食、防止过度失水等)或者增加叶肉细胞密度以抵御环境胁迫(Reich et al ., 1998 ; 张林和罗天祥, 2004 ).与落叶物种相比, 常绿物种叶片在冬季不凋落, 需经受较长时间的冬季低温等不利生长环境, 面对冬季的选择压力, 常绿植物可能采取高消耗慢收益的保守型生存策略(Cornelissen, 1999 ; 刘艳芳, 2015 ), 通过革质的叶表皮和叶肉海绵组织分层等特点来增加抗寒力(董丽等, 2003 ; 刘艳芳, 2015 ).而落叶植物的叶寿命相对较短, 需要在相对短的生长时间内积累足够的光合产物以满足植株生长需要, 薄的叶片组织能够缩短CO2 从气孔扩散到叶绿体的路径, 也能增加气体交换的速率, 进而提高光合作用速率(Diemer, 1998 ; 刘艳芳, 2015 ), 所以, 较高的SLA 是落叶物种所采取的一种低投入快收益的积极型生存策略. ...
... ; 刘艳芳, 2015 ).而落叶植物的叶寿命相对较短, 需要在相对短的生长时间内积累足够的光合产物以满足植株生长需要, 薄的叶片组织能够缩短CO2 从气孔扩散到叶绿体的路径, 也能增加气体交换的速率, 进而提高光合作用速率(Diemer, 1998 ; 刘艳芳, 2015 ), 所以, 较高的SLA 是落叶物种所采取的一种低投入快收益的积极型生存策略. ...
... ; 刘艳芳, 2015 ), 所以, 较高的SLA 是落叶物种所采取的一种低投入快收益的积极型生存策略. ...
贡嘎山阔叶木本植物叶片解剖结构特征及其环境适应研究
1
2015
... 从机械力学的角度看, 展叶效率随小枝茎长度和茎纤细率的增加而下降可能是小枝在面对机械载重和风压胁迫时, 确保小枝安全的需要.作为起支撑作用的小枝, 首先要能够承受其自身的质量, 避免折断、撕裂或其他形式的机械损伤(Mcmahon, 1973 ; King & Loucks, 1978 ; King, 1981 , 1986 ; Long et al ., 1981 ; Givnish, 1986 ; Gartner, 1995 ), 必须能够抵抗来自茎及其上叶片质量所带来的静态和动态的压力和拉力胁迫.恒定胁迫模型(The constant-stress model)理论认为, 令植物茎横截面外缘弯曲所需要做的功(即弯曲矩, the bending moment) M 主要是由力F 和臂长(即茎长) L 共同决定, 即M = F × L .对于植物而言, 茎所承受的力是叶面积A 与风压W 的乘积, 即F = A × W (Dean & Long, 1986 ).所以, 随着小枝茎长度的增加, 在小枝上相同叶面积对小枝茎基部的机械负载会相应地增加, 特别是在有风吹动摇摆的情况下, 长枝将相应地减少枝条上的叶片面积或干质量来降低机械载重和风压胁迫, 导致展叶效率随小枝茎长度和茎纤细率的增加而下降.其次, 从植株树冠构建的角度看, 枝条结构与展叶效率之间的负相关关系将有利于植株更有效地捕获光能, 提高整体光合效率.因为相互遮阴是当年生小枝结构变化的一个重要决定因素, 更大的展叶密度将会导致当年生小枝内叶片之间相互荫蔽的程度增加(Niklas, 1988 ; Yagi & Kikuzawa, 1999 ).另外, 在总的生物量一定的情况下, 当植物将更多的生物量分配给茎以使枝条生长得更长时, 将有利于植株将更多的叶片伸展到冠层外部, 同时可以让一定数量的叶片分散在更长的枝条范围内, 降低同一枝条上叶片之间相互荫蔽的程度, 以利于捕获更多的光能, 提高植株的光合效率.综上所述, 枝条结构与展叶效率之间的负相关关系具有重要的生态学意义, 有利于保持小枝的机械安全, 并提高整个植株的光合效率. ...
The role of Douglas- fir stem sapwood and heartwood in the mechanical and physiological support of crowns and development of stem form.
1
1981
... 在木本植物中, 枝条的结构可以用枝条的茎长度、直径以及纤细率(枝条长度与直径比)来表示.对于用来着生和支撑叶、花和果实等器官的小枝而言, 其长度、纤细率以及展叶效率等性状的变化将直接影响植株的机械稳定性、光捕获效率和空间拓展能力, 进而影响到植株的光合和竞争能力的各个方面(Honda & Fisher, 1978 ; Niklas & Kerchner, 1984 ).例如, 树冠内叶片的分布模式是木本植物生产力的一个重要决定因素(Horn, 1971 ; Yagi & Kikuzawa, 1999 ), 而它的变化主要受小枝的结构变化模式、小枝在树冠内的空间排布方式以及小枝的个体大小等与枝条结构密切相关的因素限制(Koike, 1989 ; Yagi & Kikuzawa, 1999 ).在一些具有长短枝分化的物种中, 短枝的茎较短, 叶片排布更紧密, 长枝的茎较长, 叶片可以分散排布在较长的节间上(Yagi & Kikuzawa, 1999 ).尽管短枝上的总叶面积比长枝小(Gregory, 1980 ; Yagi & Kikuzawa, 1999 ), 但单位枝长基础上的叶面积比长枝大(Caesar & Macdonald, 1984 ; Yagi & Kikuzawa, 1999 ).小枝通过这种结构变化模式可以调控一个特定大小的小枝上的叶片数量(Maruyama, 1983 ; Koike, 1989 ; Takenaka, 1997 ; Yagi & Kikuzawa, 1999 )和展叶效率, 影响到植株的生产力.然而, 近年来, 关于植物小枝上茎-叶关系的研究多集中于茎直径维度的叶大小-小枝大小之间的关系研究(Westoby et al ., 2002 ; Pickup et al ., 2005 ; Wright et al ., 2007 ; Li et al ., 2008 ; Xiang et al ., 2009 ; Yang et al ., 2009 ; 杨冬梅等, 2012 ), 对于茎长维度的茎结构的另一维度——茎长度和纤细率对小枝展叶效率及相应的适应策略的研究则相对不足.小枝是个悬臂结构, 小枝茎长度不仅会影响其支撑叶片的排布, 还与小枝的机械安全密切相关.根据恒定胁迫模型(the constant-stress model)理论预测, 小枝茎横截面外缘所能承受的最大弯曲胁迫(M )主要由枝条长度(L )和枝条所承受的力(F )共同决定, 即M = L × F .对于一个无花、果等其他附属物的小枝而言, 枝条所承受的力主要来自风对枝条的压力和拉拽力所形成的风压胁迫, 而风压胁迫(F )又是叶面积(A )和风压(W )的乘积, 即F = A × W (Dean & Long, 1986 ).由此可知, 在风压一定的情况下, 小枝茎基部所能承受的最大弯曲胁迫是由枝条长度和叶面积决定的, 并且两者之间是负相关关系.所以, 我们预测小枝的展叶效率将随着小枝茎长度的增加而下降. ...
Shoot characteristics as a function of the bud length on Japanese beech trees.
1
1983
... 从机械力学的角度看, 展叶效率随小枝茎长度和茎纤细率的增加而下降可能是小枝在面对机械载重和风压胁迫时, 确保小枝安全的需要.作为起支撑作用的小枝, 首先要能够承受其自身的质量, 避免折断、撕裂或其他形式的机械损伤(Mcmahon, 1973 ; King & Loucks, 1978 ; King, 1981 , 1986 ; Long et al ., 1981 ; Givnish, 1986 ; Gartner, 1995 ), 必须能够抵抗来自茎及其上叶片质量所带来的静态和动态的压力和拉力胁迫.恒定胁迫模型(The constant-stress model)理论认为, 令植物茎横截面外缘弯曲所需要做的功(即弯曲矩, the bending moment) M 主要是由力F 和臂长(即茎长) L 共同决定, 即M = F × L .对于植物而言, 茎所承受的力是叶面积A 与风压W 的乘积, 即F = A × W (Dean & Long, 1986 ).所以, 随着小枝茎长度的增加, 在小枝上相同叶面积对小枝茎基部的机械负载会相应地增加, 特别是在有风吹动摇摆的情况下, 长枝将相应地减少枝条上的叶片面积或干质量来降低机械载重和风压胁迫, 导致展叶效率随小枝茎长度和茎纤细率的增加而下降.其次, 从植株树冠构建的角度看, 枝条结构与展叶效率之间的负相关关系将有利于植株更有效地捕获光能, 提高整体光合效率.因为相互遮阴是当年生小枝结构变化的一个重要决定因素, 更大的展叶密度将会导致当年生小枝内叶片之间相互荫蔽的程度增加(Niklas, 1988 ; Yagi & Kikuzawa, 1999 ).另外, 在总的生物量一定的情况下, 当植物将更多的生物量分配给茎以使枝条生长得更长时, 将有利于植株将更多的叶片伸展到冠层外部, 同时可以让一定数量的叶片分散在更长的枝条范围内, 降低同一枝条上叶片之间相互荫蔽的程度, 以利于捕获更多的光能, 提高植株的光合效率.综上所述, 枝条结构与展叶效率之间的负相关关系具有重要的生态学意义, 有利于保持小枝的机械安全, 并提高整个植株的光合效率. ...
Size and shape in biology.
1
1973
... 在木本植物中, 枝条的结构可以用枝条的茎长度、直径以及纤细率(枝条长度与直径比)来表示.对于用来着生和支撑叶、花和果实等器官的小枝而言, 其长度、纤细率以及展叶效率等性状的变化将直接影响植株的机械稳定性、光捕获效率和空间拓展能力, 进而影响到植株的光合和竞争能力的各个方面(Honda & Fisher, 1978 ; Niklas & Kerchner, 1984 ).例如, 树冠内叶片的分布模式是木本植物生产力的一个重要决定因素(Horn, 1971 ; Yagi & Kikuzawa, 1999 ), 而它的变化主要受小枝的结构变化模式、小枝在树冠内的空间排布方式以及小枝的个体大小等与枝条结构密切相关的因素限制(Koike, 1989 ; Yagi & Kikuzawa, 1999 ).在一些具有长短枝分化的物种中, 短枝的茎较短, 叶片排布更紧密, 长枝的茎较长, 叶片可以分散排布在较长的节间上(Yagi & Kikuzawa, 1999 ).尽管短枝上的总叶面积比长枝小(Gregory, 1980 ; Yagi & Kikuzawa, 1999 ), 但单位枝长基础上的叶面积比长枝大(Caesar & Macdonald, 1984 ; Yagi & Kikuzawa, 1999 ).小枝通过这种结构变化模式可以调控一个特定大小的小枝上的叶片数量(Maruyama, 1983 ; Koike, 1989 ; Takenaka, 1997 ; Yagi & Kikuzawa, 1999 )和展叶效率, 影响到植株的生产力.然而, 近年来, 关于植物小枝上茎-叶关系的研究多集中于茎直径维度的叶大小-小枝大小之间的关系研究(Westoby et al ., 2002 ; Pickup et al ., 2005 ; Wright et al ., 2007 ; Li et al ., 2008 ; Xiang et al ., 2009 ; Yang et al ., 2009 ; 杨冬梅等, 2012 ), 对于茎长维度的茎结构的另一维度——茎长度和纤细率对小枝展叶效率及相应的适应策略的研究则相对不足.小枝是个悬臂结构, 小枝茎长度不仅会影响其支撑叶片的排布, 还与小枝的机械安全密切相关.根据恒定胁迫模型(the constant-stress model)理论预测, 小枝茎横截面外缘所能承受的最大弯曲胁迫(M )主要由枝条长度(L )和枝条所承受的力(F )共同决定, 即M = L × F .对于一个无花、果等其他附属物的小枝而言, 枝条所承受的力主要来自风对枝条的压力和拉拽力所形成的风压胁迫, 而风压胁迫(F )又是叶面积(A )和风压(W )的乘积, 即F = A × W (Dean & Long, 1986 ).由此可知, 在风压一定的情况下, 小枝茎基部所能承受的最大弯曲胁迫是由枝条长度和叶面积决定的, 并且两者之间是负相关关系.所以, 我们预测小枝的展叶效率将随着小枝茎长度的增加而下降. ...
Mechanical and photosynthetic constraints on the evolution of plant shape.
1
1984
... 从机械力学的角度看, 展叶效率随小枝茎长度和茎纤细率的增加而下降可能是小枝在面对机械载重和风压胁迫时, 确保小枝安全的需要.作为起支撑作用的小枝, 首先要能够承受其自身的质量, 避免折断、撕裂或其他形式的机械损伤(Mcmahon, 1973 ; King & Loucks, 1978 ; King, 1981 , 1986 ; Long et al ., 1981 ; Givnish, 1986 ; Gartner, 1995 ), 必须能够抵抗来自茎及其上叶片质量所带来的静态和动态的压力和拉力胁迫.恒定胁迫模型(The constant-stress model)理论认为, 令植物茎横截面外缘弯曲所需要做的功(即弯曲矩, the bending moment) M 主要是由力F 和臂长(即茎长) L 共同决定, 即M = F × L .对于植物而言, 茎所承受的力是叶面积A 与风压W 的乘积, 即F = A × W (Dean & Long, 1986 ).所以, 随着小枝茎长度的增加, 在小枝上相同叶面积对小枝茎基部的机械负载会相应地增加, 特别是在有风吹动摇摆的情况下, 长枝将相应地减少枝条上的叶片面积或干质量来降低机械载重和风压胁迫, 导致展叶效率随小枝茎长度和茎纤细率的增加而下降.其次, 从植株树冠构建的角度看, 枝条结构与展叶效率之间的负相关关系将有利于植株更有效地捕获光能, 提高整体光合效率.因为相互遮阴是当年生小枝结构变化的一个重要决定因素, 更大的展叶密度将会导致当年生小枝内叶片之间相互荫蔽的程度增加(Niklas, 1988 ; Yagi & Kikuzawa, 1999 ).另外, 在总的生物量一定的情况下, 当植物将更多的生物量分配给茎以使枝条生长得更长时, 将有利于植株将更多的叶片伸展到冠层外部, 同时可以让一定数量的叶片分散在更长的枝条范围内, 降低同一枝条上叶片之间相互荫蔽的程度, 以利于捕获更多的光能, 提高植株的光合效率.综上所述, 枝条结构与展叶效率之间的负相关关系具有重要的生态学意义, 有利于保持小枝的机械安全, 并提高整个植株的光合效率. ...
The role of phyllotactic pattern as a “developmental constraint” on the interception of light by leaf surfaces.
1
1988
... 常绿与落叶是植物通过长期进化适应形成的两种不同的生活史对策, 它们在叶寿命和叶片结构(如比叶面积)(Reich et al ., 1992 ; Westoby et al ., 2002 ; Wright et al ., 2004 ; Li et al ., 2008 )、茎木质部抗栓塞能力(Cavender-Bares & Holbrook, 2001 ; Cavender- Bares et al ., 2005 )等许多方面都显著不同.在本研究中, 虽然当年生小枝茎结构性状与表征展叶效率的性状之间的负相关关系斜率在常绿和落叶物种间无显著差异, 但截距差异显著.在3个表征展叶效率的性状中, 尽管常绿物种叶密度与小枝茎结构性状之间的关系截距显著大于落叶物种, 在某一给定的茎长或茎纤细率下, 常绿物种比落叶物种具有更多的叶数量, 即具有较高的出叶强度, 与以前相关研究发现的叶大小与叶数量之间是相互权衡的结果一致(Corner, 1949 ; Kleiman & Aarssen, 2007 ; Yang et al ., 2008 ).但由于常绿物种的单叶面积16.21 cm2 显著小于落叶物种的单叶面积34.66 cm2 (p < 0.001, 表2 ), 导致其叶面积比(单位茎质量的叶面积)和叶茎生物量比(单位茎质量的叶质量)与小枝茎结构性状之间的关系截距反而显著小于落叶物种, 即随着小枝茎长度和纤细率的增加, 常绿物种每单位质量的茎生物量投资获得的总叶面积和总叶质量都比落叶物种少, 常绿物种的实际展叶效率比落叶物种低.对数据结果进一步分析表明, 常绿物种更低展叶效率的主要原因是其比叶面积显著小于落叶物种 (p < 0.001, 表2 ), 因为我们的结果显示, 在不考虑小枝茎长结构影响的情况下, 茎叶生物量比在常绿和落叶物种中没有显著差异(p = 0.335, 表2 ), 表明总生物量在叶和茎之间的分配比例在常绿和落叶物种中是没有显著差异的, 可是落叶物种的单叶面积(p < 0.001, 表2 )和总叶面积(p = 0.001, 表2 )都要显著大于常绿物种, 并且常绿物种茎干质量与总叶面积相关关系共同斜率下的截距显著小于落叶物种(p ˂ 0.05, 表1 ), 即在某一给定的茎干质量下, 常绿物种比落叶物种具有更小的叶面积.许多研究也都发现常绿物种通常比落叶物种具有更高的比叶质量(即比叶面积的倒数)(Reich et al ., 1992 ; Westoby et al ., 2002 ; Wright et al ., 2004 ; 潘少安等, 2015 ).并且, Reich等(1998)发现比叶面积较低的植物常形成厚度较大而面积较小的叶片, 而刘艳芳(2015)在贡嘎山阔叶木本植物叶片解剖结构特征及其环境适应研究中发现的常绿植物的叶片厚度、栅栏组织厚度、海绵组织厚度、上表皮厚度和栅栏细胞层数都显著大于落叶植物的结果, 支持了Reich等(1998)的发现. ...
从叶内生物量分配策略的角度理解叶大小的优化
1
2015
... 在木本植物中, 枝条的结构可以用枝条的茎长度、直径以及纤细率(枝条长度与直径比)来表示.对于用来着生和支撑叶、花和果实等器官的小枝而言, 其长度、纤细率以及展叶效率等性状的变化将直接影响植株的机械稳定性、光捕获效率和空间拓展能力, 进而影响到植株的光合和竞争能力的各个方面(Honda & Fisher, 1978 ; Niklas & Kerchner, 1984 ).例如, 树冠内叶片的分布模式是木本植物生产力的一个重要决定因素(Horn, 1971 ; Yagi & Kikuzawa, 1999 ), 而它的变化主要受小枝的结构变化模式、小枝在树冠内的空间排布方式以及小枝的个体大小等与枝条结构密切相关的因素限制(Koike, 1989 ; Yagi & Kikuzawa, 1999 ).在一些具有长短枝分化的物种中, 短枝的茎较短, 叶片排布更紧密, 长枝的茎较长, 叶片可以分散排布在较长的节间上(Yagi & Kikuzawa, 1999 ).尽管短枝上的总叶面积比长枝小(Gregory, 1980 ; Yagi & Kikuzawa, 1999 ), 但单位枝长基础上的叶面积比长枝大(Caesar & Macdonald, 1984 ; Yagi & Kikuzawa, 1999 ).小枝通过这种结构变化模式可以调控一个特定大小的小枝上的叶片数量(Maruyama, 1983 ; Koike, 1989 ; Takenaka, 1997 ; Yagi & Kikuzawa, 1999 )和展叶效率, 影响到植株的生产力.然而, 近年来, 关于植物小枝上茎-叶关系的研究多集中于茎直径维度的叶大小-小枝大小之间的关系研究(Westoby et al ., 2002 ; Pickup et al ., 2005 ; Wright et al ., 2007 ; Li et al ., 2008 ; Xiang et al ., 2009 ; Yang et al ., 2009 ; 杨冬梅等, 2012 ), 对于茎长维度的茎结构的另一维度——茎长度和纤细率对小枝展叶效率及相应的适应策略的研究则相对不足.小枝是个悬臂结构, 小枝茎长度不仅会影响其支撑叶片的排布, 还与小枝的机械安全密切相关.根据恒定胁迫模型(the constant-stress model)理论预测, 小枝茎横截面外缘所能承受的最大弯曲胁迫(M )主要由枝条长度(L )和枝条所承受的力(F )共同决定, 即M = L × F .对于一个无花、果等其他附属物的小枝而言, 枝条所承受的力主要来自风对枝条的压力和拉拽力所形成的风压胁迫, 而风压胁迫(F )又是叶面积(A )和风压(W )的乘积, 即F = A × W (Dean & Long, 1986 ).由此可知, 在风压一定的情况下, 小枝茎基部所能承受的最大弯曲胁迫是由枝条长度和叶面积决定的, 并且两者之间是负相关关系.所以, 我们预测小枝的展叶效率将随着小枝茎长度的增加而下降. ...
Dry mass costs of deploying leaf area in relation to leaf size.
1
2005
... 对于植物构型性状以物种平均值进行分析.在对枝条结构与展叶效率的关系分析前, 首先将各参数的物种平均值进行对数转换(以10为底), 使之符合正态分布, 然后用关系方程y = bxa 来描述, 对数转换后为log(y ) = log(b ) + a log(x ), 其中x 和y 表示两个性状变量, 在双对数直角坐标系中, b 表示性状关系的截距, a 表示斜率, 即异速生长的指数(a = 1时为等速生长, 大于或小于1时为异速生长)(Harvey & Pagel, 1991 ).异速生长方程的参数估计采用标准化主轴估计(SMA)方法(Warton et al ., 2006 ), 由软件(S)MATR Version 2.0 (Falster et al ., 2006 )计算完成.回归斜率的置信区间根据(Pitman, 1939 )方法计算. ...
A note on normal correlation.
4
1939
... 同时, 对于生活在同一群落里的常绿和落叶物种而言, 由于展叶物候和生物学习性的不同, 它们的功能特征通常也会有很大的差异.例如, 相比于落叶物种, 常绿物种通常具有长的叶寿命、低的比叶面积(SLA )、低的光合速率和高的养分含量(Reich et al ., 1992 ; Westoby et al ., 2002 ; Wright et al ., 2004 ; Li et al ., 2008 ); 另外, 由于常绿物种为了确保在冬季低温等不利环境下叶片不凋落, 需要水分运输系统保持畅通, 确保水分运输安全, 其木质部需要比落叶物种具有更强的抵抗冰冻引起栓塞的能力(Cavender-Bares & Holbrook, 2001 ; Cavender-Bares et al ., 2005 ), 致使其投入在支撑结构上的成本增加, 表现出在相同的茎投入时, 常绿物种通常比落叶物种具有更小的总叶面积和单叶面积(Sun et al ., 2006 ; Yang et al ., 2009 ).这种不同的成本-收益关系将有可能导致常绿和落叶物种采取不同的枝条伸展策略以改变小枝上的叶排布方式, 使两类物种都能够捕获足够的光能以保持相对稳定的碳收益.因此, 两类物种可能会具有不同的茎结构与展叶效率之间的变化关系. ...
... 常绿与落叶是植物通过长期进化适应形成的两种不同的生活史对策, 它们在叶寿命和叶片结构(如比叶面积)(Reich et al ., 1992 ; Westoby et al ., 2002 ; Wright et al ., 2004 ; Li et al ., 2008 )、茎木质部抗栓塞能力(Cavender-Bares & Holbrook, 2001 ; Cavender- Bares et al ., 2005 )等许多方面都显著不同.在本研究中, 虽然当年生小枝茎结构性状与表征展叶效率的性状之间的负相关关系斜率在常绿和落叶物种间无显著差异, 但截距差异显著.在3个表征展叶效率的性状中, 尽管常绿物种叶密度与小枝茎结构性状之间的关系截距显著大于落叶物种, 在某一给定的茎长或茎纤细率下, 常绿物种比落叶物种具有更多的叶数量, 即具有较高的出叶强度, 与以前相关研究发现的叶大小与叶数量之间是相互权衡的结果一致(Corner, 1949 ; Kleiman & Aarssen, 2007 ; Yang et al ., 2008 ).但由于常绿物种的单叶面积16.21 cm2 显著小于落叶物种的单叶面积34.66 cm2 (p < 0.001, 表2 ), 导致其叶面积比(单位茎质量的叶面积)和叶茎生物量比(单位茎质量的叶质量)与小枝茎结构性状之间的关系截距反而显著小于落叶物种, 即随着小枝茎长度和纤细率的增加, 常绿物种每单位质量的茎生物量投资获得的总叶面积和总叶质量都比落叶物种少, 常绿物种的实际展叶效率比落叶物种低.对数据结果进一步分析表明, 常绿物种更低展叶效率的主要原因是其比叶面积显著小于落叶物种 (p < 0.001, 表2 ), 因为我们的结果显示, 在不考虑小枝茎长结构影响的情况下, 茎叶生物量比在常绿和落叶物种中没有显著差异(p = 0.335, 表2 ), 表明总生物量在叶和茎之间的分配比例在常绿和落叶物种中是没有显著差异的, 可是落叶物种的单叶面积(p < 0.001, 表2 )和总叶面积(p = 0.001, 表2 )都要显著大于常绿物种, 并且常绿物种茎干质量与总叶面积相关关系共同斜率下的截距显著小于落叶物种(p ˂ 0.05, 表1 ), 即在某一给定的茎干质量下, 常绿物种比落叶物种具有更小的叶面积.许多研究也都发现常绿物种通常比落叶物种具有更高的比叶质量(即比叶面积的倒数)(Reich et al ., 1992 ; Westoby et al ., 2002 ; Wright et al ., 2004 ; 潘少安等, 2015 ).并且, Reich等(1998)发现比叶面积较低的植物常形成厚度较大而面积较小的叶片, 而刘艳芳(2015)在贡嘎山阔叶木本植物叶片解剖结构特征及其环境适应研究中发现的常绿植物的叶片厚度、栅栏组织厚度、海绵组织厚度、上表皮厚度和栅栏细胞层数都显著大于落叶植物的结果, 支持了Reich等(1998)的发现. ...
... ), 即在某一给定的茎干质量下, 常绿物种比落叶物种具有更小的叶面积.许多研究也都发现常绿物种通常比落叶物种具有更高的比叶质量(即比叶面积的倒数)(Reich et al ., 1992 ; Westoby et al ., 2002 ; Wright et al ., 2004 ; 潘少安等, 2015 ).并且, Reich等(1998)发现比叶面积较低的植物常形成厚度较大而面积较小的叶片, 而刘艳芳(2015)在贡嘎山阔叶木本植物叶片解剖结构特征及其环境适应研究中发现的常绿植物的叶片厚度、栅栏组织厚度、海绵组织厚度、上表皮厚度和栅栏细胞层数都显著大于落叶植物的结果, 支持了Reich等(1998)的发现. ...
... 常绿物种较低的比叶面积继而导致较低的单位茎结构基础上的展叶效率, 可能与其具有较长的叶寿命有关.一般而言, 叶寿命越长, 叶片越厚, 单位叶面积的建成消耗也越大, 叶子需要在足够长的时间内通过自身的光合生产所形成的有机碳来补偿初始的建成消耗(包括抵抗草食动物的防御构造), 并维持自身生存所需的物质消耗(Coley, 1988 ; Kikuzawa, 1991 ; Schulze et al ., 1994 ; Kikuzawa, 1995 ; 张林和罗天祥, 2004 ).所以叶寿命与比叶面积成反比(Reich et al ., 1992 ; Reich & Bowman, 1999 ; Ryser & Urbas, 2000 ; 张林和罗天祥, 2004 ).比叶面积较低的植物, 叶片中很大一部分物质用于构建保卫构造(防虫食、防止过度失水等)或者增加叶肉细胞密度以抵御环境胁迫(Reich et al ., 1998 ; 张林和罗天祥, 2004 ).与落叶物种相比, 常绿物种叶片在冬季不凋落, 需经受较长时间的冬季低温等不利生长环境, 面对冬季的选择压力, 常绿植物可能采取高消耗慢收益的保守型生存策略(Cornelissen, 1999 ; 刘艳芳, 2015 ), 通过革质的叶表皮和叶肉海绵组织分层等特点来增加抗寒力(董丽等, 2003 ; 刘艳芳, 2015 ).而落叶植物的叶寿命相对较短, 需要在相对短的生长时间内积累足够的光合产物以满足植株生长需要, 薄的叶片组织能够缩短CO2 从气孔扩散到叶绿体的路径, 也能增加气体交换的速率, 进而提高光合作用速率(Diemer, 1998 ; 刘艳芳, 2015 ), 所以, 较高的SLA 是落叶物种所采取的一种低投入快收益的积极型生存策略. ...
Leaf life-span in relation to leaf, plant, and stand characteristics among diverse ecosystems.
1
1992
... 常绿物种较低的比叶面积继而导致较低的单位茎结构基础上的展叶效率, 可能与其具有较长的叶寿命有关.一般而言, 叶寿命越长, 叶片越厚, 单位叶面积的建成消耗也越大, 叶子需要在足够长的时间内通过自身的光合生产所形成的有机碳来补偿初始的建成消耗(包括抵抗草食动物的防御构造), 并维持自身生存所需的物质消耗(Coley, 1988 ; Kikuzawa, 1991 ; Schulze et al ., 1994 ; Kikuzawa, 1995 ; 张林和罗天祥, 2004 ).所以叶寿命与比叶面积成反比(Reich et al ., 1992 ; Reich & Bowman, 1999 ; Ryser & Urbas, 2000 ; 张林和罗天祥, 2004 ).比叶面积较低的植物, 叶片中很大一部分物质用于构建保卫构造(防虫食、防止过度失水等)或者增加叶肉细胞密度以抵御环境胁迫(Reich et al ., 1998 ; 张林和罗天祥, 2004 ).与落叶物种相比, 常绿物种叶片在冬季不凋落, 需经受较长时间的冬季低温等不利生长环境, 面对冬季的选择压力, 常绿植物可能采取高消耗慢收益的保守型生存策略(Cornelissen, 1999 ; 刘艳芳, 2015 ), 通过革质的叶表皮和叶肉海绵组织分层等特点来增加抗寒力(董丽等, 2003 ; 刘艳芳, 2015 ).而落叶植物的叶寿命相对较短, 需要在相对短的生长时间内积累足够的光合产物以满足植株生长需要, 薄的叶片组织能够缩短CO2 从气孔扩散到叶绿体的路径, 也能增加气体交换的速率, 进而提高光合作用速率(Diemer, 1998 ; 刘艳芳, 2015 ), 所以, 较高的SLA 是落叶物种所采取的一种低投入快收益的积极型生存策略. ...
Generality of leaf trait relationships: A test across six biomes.
1
1999
... 常绿物种较低的比叶面积继而导致较低的单位茎结构基础上的展叶效率, 可能与其具有较长的叶寿命有关.一般而言, 叶寿命越长, 叶片越厚, 单位叶面积的建成消耗也越大, 叶子需要在足够长的时间内通过自身的光合生产所形成的有机碳来补偿初始的建成消耗(包括抵抗草食动物的防御构造), 并维持自身生存所需的物质消耗(Coley, 1988 ; Kikuzawa, 1991 ; Schulze et al ., 1994 ; Kikuzawa, 1995 ; 张林和罗天祥, 2004 ).所以叶寿命与比叶面积成反比(Reich et al ., 1992 ; Reich & Bowman, 1999 ; Ryser & Urbas, 2000 ; 张林和罗天祥, 2004 ).比叶面积较低的植物, 叶片中很大一部分物质用于构建保卫构造(防虫食、防止过度失水等)或者增加叶肉细胞密度以抵御环境胁迫(Reich et al ., 1998 ; 张林和罗天祥, 2004 ).与落叶物种相比, 常绿物种叶片在冬季不凋落, 需经受较长时间的冬季低温等不利生长环境, 面对冬季的选择压力, 常绿植物可能采取高消耗慢收益的保守型生存策略(Cornelissen, 1999 ; 刘艳芳, 2015 ), 通过革质的叶表皮和叶肉海绵组织分层等特点来增加抗寒力(董丽等, 2003 ; 刘艳芳, 2015 ).而落叶植物的叶寿命相对较短, 需要在相对短的生长时间内积累足够的光合产物以满足植株生长需要, 薄的叶片组织能够缩短CO2 从气孔扩散到叶绿体的路径, 也能增加气体交换的速率, 进而提高光合作用速率(Diemer, 1998 ; 刘艳芳, 2015 ), 所以, 较高的SLA 是落叶物种所采取的一种低投入快收益的积极型生存策略. ...
Relationships of leaf dark respiration to leaf nitrogen, specific leaf area and leaf life-span: A test across biomes and functional groups.
1
1998
... 常绿物种较低的比叶面积继而导致较低的单位茎结构基础上的展叶效率, 可能与其具有较长的叶寿命有关.一般而言, 叶寿命越长, 叶片越厚, 单位叶面积的建成消耗也越大, 叶子需要在足够长的时间内通过自身的光合生产所形成的有机碳来补偿初始的建成消耗(包括抵抗草食动物的防御构造), 并维持自身生存所需的物质消耗(Coley, 1988 ; Kikuzawa, 1991 ; Schulze et al ., 1994 ; Kikuzawa, 1995 ; 张林和罗天祥, 2004 ).所以叶寿命与比叶面积成反比(Reich et al ., 1992 ; Reich & Bowman, 1999 ; Ryser & Urbas, 2000 ; 张林和罗天祥, 2004 ).比叶面积较低的植物, 叶片中很大一部分物质用于构建保卫构造(防虫食、防止过度失水等)或者增加叶肉细胞密度以抵御环境胁迫(Reich et al ., 1998 ; 张林和罗天祥, 2004 ).与落叶物种相比, 常绿物种叶片在冬季不凋落, 需经受较长时间的冬季低温等不利生长环境, 面对冬季的选择压力, 常绿植物可能采取高消耗慢收益的保守型生存策略(Cornelissen, 1999 ; 刘艳芳, 2015 ), 通过革质的叶表皮和叶肉海绵组织分层等特点来增加抗寒力(董丽等, 2003 ; 刘艳芳, 2015 ).而落叶植物的叶寿命相对较短, 需要在相对短的生长时间内积累足够的光合产物以满足植株生长需要, 薄的叶片组织能够缩短CO2 从气孔扩散到叶绿体的路径, 也能增加气体交换的速率, 进而提高光合作用速率(Diemer, 1998 ; 刘艳芳, 2015 ), 所以, 较高的SLA 是落叶物种所采取的一种低投入快收益的积极型生存策略. ...
Ecological significance of leaf life span among Central European grass species.
1
2000
... 常绿物种较低的比叶面积继而导致较低的单位茎结构基础上的展叶效率, 可能与其具有较长的叶寿命有关.一般而言, 叶寿命越长, 叶片越厚, 单位叶面积的建成消耗也越大, 叶子需要在足够长的时间内通过自身的光合生产所形成的有机碳来补偿初始的建成消耗(包括抵抗草食动物的防御构造), 并维持自身生存所需的物质消耗(Coley, 1988 ; Kikuzawa, 1991 ; Schulze et al ., 1994 ; Kikuzawa, 1995 ; 张林和罗天祥, 2004 ).所以叶寿命与比叶面积成反比(Reich et al ., 1992 ; Reich & Bowman, 1999 ; Ryser & Urbas, 2000 ; 张林和罗天祥, 2004 ).比叶面积较低的植物, 叶片中很大一部分物质用于构建保卫构造(防虫食、防止过度失水等)或者增加叶肉细胞密度以抵御环境胁迫(Reich et al ., 1998 ; 张林和罗天祥, 2004 ).与落叶物种相比, 常绿物种叶片在冬季不凋落, 需经受较长时间的冬季低温等不利生长环境, 面对冬季的选择压力, 常绿植物可能采取高消耗慢收益的保守型生存策略(Cornelissen, 1999 ; 刘艳芳, 2015 ), 通过革质的叶表皮和叶肉海绵组织分层等特点来增加抗寒力(董丽等, 2003 ; 刘艳芳, 2015 ).而落叶植物的叶寿命相对较短, 需要在相对短的生长时间内积累足够的光合产物以满足植株生长需要, 薄的叶片组织能够缩短CO2 从气孔扩散到叶绿体的路径, 也能增加气体交换的速率, 进而提高光合作用速率(Diemer, 1998 ; 刘艳芳, 2015 ), 所以, 较高的SLA 是落叶物种所采取的一种低投入快收益的积极型生存策略. ...
Relationships among maximum stomatal conductance, ecosystem surface conductance, carbon assimilation rate, and plant nitrogen nutrition: A global ecology scaling exercise.
1
1994
... 同时, 对于生活在同一群落里的常绿和落叶物种而言, 由于展叶物候和生物学习性的不同, 它们的功能特征通常也会有很大的差异.例如, 相比于落叶物种, 常绿物种通常具有长的叶寿命、低的比叶面积(SLA )、低的光合速率和高的养分含量(Reich et al ., 1992 ; Westoby et al ., 2002 ; Wright et al ., 2004 ; Li et al ., 2008 ); 另外, 由于常绿物种为了确保在冬季低温等不利环境下叶片不凋落, 需要水分运输系统保持畅通, 确保水分运输安全, 其木质部需要比落叶物种具有更强的抵抗冰冻引起栓塞的能力(Cavender-Bares & Holbrook, 2001 ; Cavender-Bares et al ., 2005 ), 致使其投入在支撑结构上的成本增加, 表现出在相同的茎投入时, 常绿物种通常比落叶物种具有更小的总叶面积和单叶面积(Sun et al ., 2006 ; Yang et al ., 2009 ).这种不同的成本-收益关系将有可能导致常绿和落叶物种采取不同的枝条伸展策略以改变小枝上的叶排布方式, 使两类物种都能够捕获足够的光能以保持相对稳定的碳收益.因此, 两类物种可能会具有不同的茎结构与展叶效率之间的变化关系. ...
The leaf size-twig size spectrum of temperate woody species along an altitudinal gradient: An invariant allometric scaling relationship.
2
2006
... 在木本植物中, 枝条的结构可以用枝条的茎长度、直径以及纤细率(枝条长度与直径比)来表示.对于用来着生和支撑叶、花和果实等器官的小枝而言, 其长度、纤细率以及展叶效率等性状的变化将直接影响植株的机械稳定性、光捕获效率和空间拓展能力, 进而影响到植株的光合和竞争能力的各个方面(Honda & Fisher, 1978 ; Niklas & Kerchner, 1984 ).例如, 树冠内叶片的分布模式是木本植物生产力的一个重要决定因素(Horn, 1971 ; Yagi & Kikuzawa, 1999 ), 而它的变化主要受小枝的结构变化模式、小枝在树冠内的空间排布方式以及小枝的个体大小等与枝条结构密切相关的因素限制(Koike, 1989 ; Yagi & Kikuzawa, 1999 ).在一些具有长短枝分化的物种中, 短枝的茎较短, 叶片排布更紧密, 长枝的茎较长, 叶片可以分散排布在较长的节间上(Yagi & Kikuzawa, 1999 ).尽管短枝上的总叶面积比长枝小(Gregory, 1980 ; Yagi & Kikuzawa, 1999 ), 但单位枝长基础上的叶面积比长枝大(Caesar & Macdonald, 1984 ; Yagi & Kikuzawa, 1999 ).小枝通过这种结构变化模式可以调控一个特定大小的小枝上的叶片数量(Maruyama, 1983 ; Koike, 1989 ; Takenaka, 1997 ; Yagi & Kikuzawa, 1999 )和展叶效率, 影响到植株的生产力.然而, 近年来, 关于植物小枝上茎-叶关系的研究多集中于茎直径维度的叶大小-小枝大小之间的关系研究(Westoby et al ., 2002 ; Pickup et al ., 2005 ; Wright et al ., 2007 ; Li et al ., 2008 ; Xiang et al ., 2009 ; Yang et al ., 2009 ; 杨冬梅等, 2012 ), 对于茎长维度的茎结构的另一维度——茎长度和纤细率对小枝展叶效率及相应的适应策略的研究则相对不足.小枝是个悬臂结构, 小枝茎长度不仅会影响其支撑叶片的排布, 还与小枝的机械安全密切相关.根据恒定胁迫模型(the constant-stress model)理论预测, 小枝茎横截面外缘所能承受的最大弯曲胁迫(M )主要由枝条长度(L )和枝条所承受的力(F )共同决定, 即M = L × F .对于一个无花、果等其他附属物的小枝而言, 枝条所承受的力主要来自风对枝条的压力和拉拽力所形成的风压胁迫, 而风压胁迫(F )又是叶面积(A )和风压(W )的乘积, 即F = A × W (Dean & Long, 1986 ).由此可知, 在风压一定的情况下, 小枝茎基部所能承受的最大弯曲胁迫是由枝条长度和叶面积决定的, 并且两者之间是负相关关系.所以, 我们预测小枝的展叶效率将随着小枝茎长度的增加而下降. ...
... 本研究中, 无论是落叶还是常绿物种, 用以表征展叶效率的叶面积比、叶密度、叶茎生物量比, 均与表征当年生小枝茎结构的小枝茎长度和茎纤细率呈显著的负相关关系(图1 , 图3 , 图4 ), 即随着当年生小枝茎长度和纤细率的增加, 小枝的展叶效率逐渐降低.这一结果与Takenaka和Yagi等的研究结果(Takenaka, 1997 ; Yagi & Kikuzawa, 1999 ; Yagi, 2000 , 2004 )类似, 他们在多个木本植物的种内研究发现, 较长的当年生枝条具有更小的单位枝长基础上的叶面积.而叶密度可能是小枝展叶效率的一个主要调节因素.我们的结果显示, 无论是在常绿还是落叶物种中, 不仅小枝茎长度和茎纤细率均与叶密度(叶数量/茎长度)呈显著的负相关关系(图3 ), 并且每个小枝上的叶数量也直接与小枝茎长度呈斜率显著小于1.0 (p < 0.000β1)的正的异速生长关系(图2 ; 表1 ), 表明叶数量的增加速度没有小枝茎长度的增加速度快, 长的枝条相对于短枝具有更低的出叶强度.Yagi (2004)的研究结果也表明, 长的当年生小枝比短的当年生小枝具有更小的叶数量与枝长度的比值, 即单位枝长度的叶片数更少.在我们以前的研究中也发现, 小枝茎长度和单位茎长度的叶片数 ...
Structural variation in current-year shoots of broad-leaved evergreen tree saplings under forest canopies in warm temperate Japan.
1
1997
... 由枝和叶所构成的树冠是植物进行光合作用、呼吸作用、蒸腾作用等一系列生理活动的主要场所, 其大小、结构、形状及相互间的协变规律直接决定了植物个体内的生物量分配、生长活力和生产力.枝条作为植株的重要结构单元, 不仅是植株运输水分和营养物质的重要通道, 也是支撑树冠的主要结构并直接影响树冠的形状和大小, 同时它还是叶片的支撑体, 能通过其长度、结构、分布方式等变化来调整冠层内叶片的排布方式, 将叶伸展到适宜的空间形成一个合理的光拦截体系, 提高光合效率(黎云祥和刘玉成, 1996 ; Day et al ., 2002 ; Thomas & Winner, 2002 ; 张运春等, 2005 ).叶片是植物进行光合作用的主要器官, 叶片的数量、面积以及展叶效率等空间结构特征的变化直接影响着植株的光拦截效率和光合效率.因此, 枝条的结构特征及其与叶性状之间的协变规律反映植物对光照、空间等资源的利用和适应策略(Borchert & Slade, 1981 ; Bicknell, 1982 ; Boojh & Ramakrishnan, 1982 ; Bell, 1984 ; 陈波等, 2002 ). ...
Photosynthetic differences between saplings and adult trees: An integration of field results by meta-analysis.
2002
Common slope tests for bivariate errors-in-variables models.
2
2002
... 对于植物构型性状以物种平均值进行分析.在对枝条结构与展叶效率的关系分析前, 首先将各参数的物种平均值进行对数转换(以10为底), 使之符合正态分布, 然后用关系方程y = bxa 来描述, 对数转换后为log(y ) = log(b ) + a log(x ), 其中x 和y 表示两个性状变量, 在双对数直角坐标系中, b 表示性状关系的截距, a 表示斜率, 即异速生长的指数(a = 1时为等速生长, 大于或小于1时为异速生长)(Harvey & Pagel, 1991 ).异速生长方程的参数估计采用标准化主轴估计(SMA)方法(Warton et al ., 2006 ), 由软件(S)MATR Version 2.0 (Falster et al ., 2006 )计算完成.回归斜率的置信区间根据(Pitman, 1939 )方法计算. ...
... 在异速生长分析中, 对于常绿和落叶物种组的斜率采用Warton和Weber (2002)的方法进行异质性检验, 如果检验结果没有异质性(p > 0.05), 就给出一个共同斜率.共同斜率下的截距差异(y 轴向)以及沿共同主轴方向位移的差异通过t 检验或方差分析检验(Wright et al ., 2001 ; Warton et al ., 2006 )进行分析. ...
Bivariate line-fitting methods for allometry.
2
2006
... 研究区位于浙江省清凉峰自然保护区, 地理位置118.87°-119.18° E, 30.08°-30.28° N, 主峰海拔 1β787.4 m.保护区地处中亚热带北缘, 气候复杂, 横跨亚热带和温带两个季风带, 具有明显的亚热带中山山地季风特征.主峰清凉峰则更具有典型的高山气候特点, 立体气候明显, 从山麓到山顶年平均气温从15.3 ℃降至7.8 ℃, 7月为最热月, 历年平均气温28.10 ℃, 1月为最冷月, 历年平均气温3.40 ℃.极端最高气温为41.90 ℃ (1996年8月6日), 极端最低气温为-13.30 ℃ (1967年1月16日).全年降水量 1β500-1β900 mm, 相对湿度在78%-82%之间.全山区年日照时间1β500-2 000 h, 年日照百分率为35%- 39%.海拔800-1 100 m区域年日照时间1 500-1 600 h (翁东明等, 2009 ; 程丽媛等, 2016 ). ...
... 保护区内植被带景观垂直层次分明, 群落内乔木层植物平均密度为2 062株·hm-2 , 灌木层平均密度为17 163株·hm-2 , 草本层平均密度为5 976株·hm-2 .灌木层植株密度基本是乔木层植株密度的8.3倍, 是草本层植株密度的2.9倍(翁东明等, 2009 ; 程丽媛等, 2016 ). ...
清凉峰自然保护区台湾水青冈群落物种多样性研究
4
2009
... 在木本植物中, 枝条的结构可以用枝条的茎长度、直径以及纤细率(枝条长度与直径比)来表示.对于用来着生和支撑叶、花和果实等器官的小枝而言, 其长度、纤细率以及展叶效率等性状的变化将直接影响植株的机械稳定性、光捕获效率和空间拓展能力, 进而影响到植株的光合和竞争能力的各个方面(Honda & Fisher, 1978 ; Niklas & Kerchner, 1984 ).例如, 树冠内叶片的分布模式是木本植物生产力的一个重要决定因素(Horn, 1971 ; Yagi & Kikuzawa, 1999 ), 而它的变化主要受小枝的结构变化模式、小枝在树冠内的空间排布方式以及小枝的个体大小等与枝条结构密切相关的因素限制(Koike, 1989 ; Yagi & Kikuzawa, 1999 ).在一些具有长短枝分化的物种中, 短枝的茎较短, 叶片排布更紧密, 长枝的茎较长, 叶片可以分散排布在较长的节间上(Yagi & Kikuzawa, 1999 ).尽管短枝上的总叶面积比长枝小(Gregory, 1980 ; Yagi & Kikuzawa, 1999 ), 但单位枝长基础上的叶面积比长枝大(Caesar & Macdonald, 1984 ; Yagi & Kikuzawa, 1999 ).小枝通过这种结构变化模式可以调控一个特定大小的小枝上的叶片数量(Maruyama, 1983 ; Koike, 1989 ; Takenaka, 1997 ; Yagi & Kikuzawa, 1999 )和展叶效率, 影响到植株的生产力.然而, 近年来, 关于植物小枝上茎-叶关系的研究多集中于茎直径维度的叶大小-小枝大小之间的关系研究(Westoby et al ., 2002 ; Pickup et al ., 2005 ; Wright et al ., 2007 ; Li et al ., 2008 ; Xiang et al ., 2009 ; Yang et al ., 2009 ; 杨冬梅等, 2012 ), 对于茎长维度的茎结构的另一维度——茎长度和纤细率对小枝展叶效率及相应的适应策略的研究则相对不足.小枝是个悬臂结构, 小枝茎长度不仅会影响其支撑叶片的排布, 还与小枝的机械安全密切相关.根据恒定胁迫模型(the constant-stress model)理论预测, 小枝茎横截面外缘所能承受的最大弯曲胁迫(M )主要由枝条长度(L )和枝条所承受的力(F )共同决定, 即M = L × F .对于一个无花、果等其他附属物的小枝而言, 枝条所承受的力主要来自风对枝条的压力和拉拽力所形成的风压胁迫, 而风压胁迫(F )又是叶面积(A )和风压(W )的乘积, 即F = A × W (Dean & Long, 1986 ).由此可知, 在风压一定的情况下, 小枝茎基部所能承受的最大弯曲胁迫是由枝条长度和叶面积决定的, 并且两者之间是负相关关系.所以, 我们预测小枝的展叶效率将随着小枝茎长度的增加而下降. ...
... 同时, 对于生活在同一群落里的常绿和落叶物种而言, 由于展叶物候和生物学习性的不同, 它们的功能特征通常也会有很大的差异.例如, 相比于落叶物种, 常绿物种通常具有长的叶寿命、低的比叶面积(SLA )、低的光合速率和高的养分含量(Reich et al ., 1992 ; Westoby et al ., 2002 ; Wright et al ., 2004 ; Li et al ., 2008 ); 另外, 由于常绿物种为了确保在冬季低温等不利环境下叶片不凋落, 需要水分运输系统保持畅通, 确保水分运输安全, 其木质部需要比落叶物种具有更强的抵抗冰冻引起栓塞的能力(Cavender-Bares & Holbrook, 2001 ; Cavender-Bares et al ., 2005 ), 致使其投入在支撑结构上的成本增加, 表现出在相同的茎投入时, 常绿物种通常比落叶物种具有更小的总叶面积和单叶面积(Sun et al ., 2006 ; Yang et al ., 2009 ).这种不同的成本-收益关系将有可能导致常绿和落叶物种采取不同的枝条伸展策略以改变小枝上的叶排布方式, 使两类物种都能够捕获足够的光能以保持相对稳定的碳收益.因此, 两类物种可能会具有不同的茎结构与展叶效率之间的变化关系. ...
... 常绿与落叶是植物通过长期进化适应形成的两种不同的生活史对策, 它们在叶寿命和叶片结构(如比叶面积)(Reich et al ., 1992 ; Westoby et al ., 2002 ; Wright et al ., 2004 ; Li et al ., 2008 )、茎木质部抗栓塞能力(Cavender-Bares & Holbrook, 2001 ; Cavender- Bares et al ., 2005 )等许多方面都显著不同.在本研究中, 虽然当年生小枝茎结构性状与表征展叶效率的性状之间的负相关关系斜率在常绿和落叶物种间无显著差异, 但截距差异显著.在3个表征展叶效率的性状中, 尽管常绿物种叶密度与小枝茎结构性状之间的关系截距显著大于落叶物种, 在某一给定的茎长或茎纤细率下, 常绿物种比落叶物种具有更多的叶数量, 即具有较高的出叶强度, 与以前相关研究发现的叶大小与叶数量之间是相互权衡的结果一致(Corner, 1949 ; Kleiman & Aarssen, 2007 ; Yang et al ., 2008 ).但由于常绿物种的单叶面积16.21 cm2 显著小于落叶物种的单叶面积34.66 cm2 (p < 0.001, 表2 ), 导致其叶面积比(单位茎质量的叶面积)和叶茎生物量比(单位茎质量的叶质量)与小枝茎结构性状之间的关系截距反而显著小于落叶物种, 即随着小枝茎长度和纤细率的增加, 常绿物种每单位质量的茎生物量投资获得的总叶面积和总叶质量都比落叶物种少, 常绿物种的实际展叶效率比落叶物种低.对数据结果进一步分析表明, 常绿物种更低展叶效率的主要原因是其比叶面积显著小于落叶物种 (p < 0.001, 表2 ), 因为我们的结果显示, 在不考虑小枝茎长结构影响的情况下, 茎叶生物量比在常绿和落叶物种中没有显著差异(p = 0.335, 表2 ), 表明总生物量在叶和茎之间的分配比例在常绿和落叶物种中是没有显著差异的, 可是落叶物种的单叶面积(p < 0.001, 表2 )和总叶面积(p = 0.001, 表2 )都要显著大于常绿物种, 并且常绿物种茎干质量与总叶面积相关关系共同斜率下的截距显著小于落叶物种(p ˂ 0.05, 表1 ), 即在某一给定的茎干质量下, 常绿物种比落叶物种具有更小的叶面积.许多研究也都发现常绿物种通常比落叶物种具有更高的比叶质量(即比叶面积的倒数)(Reich et al ., 1992 ; Westoby et al ., 2002 ; Wright et al ., 2004 ; 潘少安等, 2015 ).并且, Reich等(1998)发现比叶面积较低的植物常形成厚度较大而面积较小的叶片, 而刘艳芳(2015)在贡嘎山阔叶木本植物叶片解剖结构特征及其环境适应研究中发现的常绿植物的叶片厚度、栅栏组织厚度、海绵组织厚度、上表皮厚度和栅栏细胞层数都显著大于落叶植物的结果, 支持了Reich等(1998)的发现. ...
... ; Westoby et al ., 2002 ; Wright et al ., 2004 ; 潘少安等, 2015 ).并且, Reich等(1998)发现比叶面积较低的植物常形成厚度较大而面积较小的叶片, 而刘艳芳(2015)在贡嘎山阔叶木本植物叶片解剖结构特征及其环境适应研究中发现的常绿植物的叶片厚度、栅栏组织厚度、海绵组织厚度、上表皮厚度和栅栏细胞层数都显著大于落叶植物的结果, 支持了Reich等(1998)的发现. ...
Plant ecological strategies: Some leading dimensions of variation between species.
1
2002
... 在木本植物中, 枝条的结构可以用枝条的茎长度、直径以及纤细率(枝条长度与直径比)来表示.对于用来着生和支撑叶、花和果实等器官的小枝而言, 其长度、纤细率以及展叶效率等性状的变化将直接影响植株的机械稳定性、光捕获效率和空间拓展能力, 进而影响到植株的光合和竞争能力的各个方面(Honda & Fisher, 1978 ; Niklas & Kerchner, 1984 ).例如, 树冠内叶片的分布模式是木本植物生产力的一个重要决定因素(Horn, 1971 ; Yagi & Kikuzawa, 1999 ), 而它的变化主要受小枝的结构变化模式、小枝在树冠内的空间排布方式以及小枝的个体大小等与枝条结构密切相关的因素限制(Koike, 1989 ; Yagi & Kikuzawa, 1999 ).在一些具有长短枝分化的物种中, 短枝的茎较短, 叶片排布更紧密, 长枝的茎较长, 叶片可以分散排布在较长的节间上(Yagi & Kikuzawa, 1999 ).尽管短枝上的总叶面积比长枝小(Gregory, 1980 ; Yagi & Kikuzawa, 1999 ), 但单位枝长基础上的叶面积比长枝大(Caesar & Macdonald, 1984 ; Yagi & Kikuzawa, 1999 ).小枝通过这种结构变化模式可以调控一个特定大小的小枝上的叶片数量(Maruyama, 1983 ; Koike, 1989 ; Takenaka, 1997 ; Yagi & Kikuzawa, 1999 )和展叶效率, 影响到植株的生产力.然而, 近年来, 关于植物小枝上茎-叶关系的研究多集中于茎直径维度的叶大小-小枝大小之间的关系研究(Westoby et al ., 2002 ; Pickup et al ., 2005 ; Wright et al ., 2007 ; Li et al ., 2008 ; Xiang et al ., 2009 ; Yang et al ., 2009 ; 杨冬梅等, 2012 ), 对于茎长维度的茎结构的另一维度——茎长度和纤细率对小枝展叶效率及相应的适应策略的研究则相对不足.小枝是个悬臂结构, 小枝茎长度不仅会影响其支撑叶片的排布, 还与小枝的机械安全密切相关.根据恒定胁迫模型(the constant-stress model)理论预测, 小枝茎横截面外缘所能承受的最大弯曲胁迫(M )主要由枝条长度(L )和枝条所承受的力(F )共同决定, 即M = L × F .对于一个无花、果等其他附属物的小枝而言, 枝条所承受的力主要来自风对枝条的压力和拉拽力所形成的风压胁迫, 而风压胁迫(F )又是叶面积(A )和风压(W )的乘积, 即F = A × W (Dean & Long, 1986 ).由此可知, 在风压一定的情况下, 小枝茎基部所能承受的最大弯曲胁迫是由枝条长度和叶面积决定的, 并且两者之间是负相关关系.所以, 我们预测小枝的展叶效率将随着小枝茎长度的增加而下降. ...
Relationships among ecologically important dimensions of plant trait variation in seven neotropical forests.
1
2007
... 在异速生长分析中, 对于常绿和落叶物种组的斜率采用Warton和Weber (2002)的方法进行异质性检验, 如果检验结果没有异质性(p > 0.05), 就给出一个共同斜率.共同斜率下的截距差异(y 轴向)以及沿共同主轴方向位移的差异通过t 检验或方差分析检验(Wright et al ., 2001 ; Warton et al ., 2006 )进行分析. ...
Strategy shifts in leaf physiology, structure and nutrient content between species of high- and low-rainfall and high- and low-nutrient habitats.
3
2001
... 同时, 对于生活在同一群落里的常绿和落叶物种而言, 由于展叶物候和生物学习性的不同, 它们的功能特征通常也会有很大的差异.例如, 相比于落叶物种, 常绿物种通常具有长的叶寿命、低的比叶面积(SLA )、低的光合速率和高的养分含量(Reich et al ., 1992 ; Westoby et al ., 2002 ; Wright et al ., 2004 ; Li et al ., 2008 ); 另外, 由于常绿物种为了确保在冬季低温等不利环境下叶片不凋落, 需要水分运输系统保持畅通, 确保水分运输安全, 其木质部需要比落叶物种具有更强的抵抗冰冻引起栓塞的能力(Cavender-Bares & Holbrook, 2001 ; Cavender-Bares et al ., 2005 ), 致使其投入在支撑结构上的成本增加, 表现出在相同的茎投入时, 常绿物种通常比落叶物种具有更小的总叶面积和单叶面积(Sun et al ., 2006 ; Yang et al ., 2009 ).这种不同的成本-收益关系将有可能导致常绿和落叶物种采取不同的枝条伸展策略以改变小枝上的叶排布方式, 使两类物种都能够捕获足够的光能以保持相对稳定的碳收益.因此, 两类物种可能会具有不同的茎结构与展叶效率之间的变化关系. ...
... 常绿与落叶是植物通过长期进化适应形成的两种不同的生活史对策, 它们在叶寿命和叶片结构(如比叶面积)(Reich et al ., 1992 ; Westoby et al ., 2002 ; Wright et al ., 2004 ; Li et al ., 2008 )、茎木质部抗栓塞能力(Cavender-Bares & Holbrook, 2001 ; Cavender- Bares et al ., 2005 )等许多方面都显著不同.在本研究中, 虽然当年生小枝茎结构性状与表征展叶效率的性状之间的负相关关系斜率在常绿和落叶物种间无显著差异, 但截距差异显著.在3个表征展叶效率的性状中, 尽管常绿物种叶密度与小枝茎结构性状之间的关系截距显著大于落叶物种, 在某一给定的茎长或茎纤细率下, 常绿物种比落叶物种具有更多的叶数量, 即具有较高的出叶强度, 与以前相关研究发现的叶大小与叶数量之间是相互权衡的结果一致(Corner, 1949 ; Kleiman & Aarssen, 2007 ; Yang et al ., 2008 ).但由于常绿物种的单叶面积16.21 cm2 显著小于落叶物种的单叶面积34.66 cm2 (p < 0.001, 表2 ), 导致其叶面积比(单位茎质量的叶面积)和叶茎生物量比(单位茎质量的叶质量)与小枝茎结构性状之间的关系截距反而显著小于落叶物种, 即随着小枝茎长度和纤细率的增加, 常绿物种每单位质量的茎生物量投资获得的总叶面积和总叶质量都比落叶物种少, 常绿物种的实际展叶效率比落叶物种低.对数据结果进一步分析表明, 常绿物种更低展叶效率的主要原因是其比叶面积显著小于落叶物种 (p < 0.001, 表2 ), 因为我们的结果显示, 在不考虑小枝茎长结构影响的情况下, 茎叶生物量比在常绿和落叶物种中没有显著差异(p = 0.335, 表2 ), 表明总生物量在叶和茎之间的分配比例在常绿和落叶物种中是没有显著差异的, 可是落叶物种的单叶面积(p < 0.001, 表2 )和总叶面积(p = 0.001, 表2 )都要显著大于常绿物种, 并且常绿物种茎干质量与总叶面积相关关系共同斜率下的截距显著小于落叶物种(p ˂ 0.05, 表1 ), 即在某一给定的茎干质量下, 常绿物种比落叶物种具有更小的叶面积.许多研究也都发现常绿物种通常比落叶物种具有更高的比叶质量(即比叶面积的倒数)(Reich et al ., 1992 ; Westoby et al ., 2002 ; Wright et al ., 2004 ; 潘少安等, 2015 ).并且, Reich等(1998)发现比叶面积较低的植物常形成厚度较大而面积较小的叶片, 而刘艳芳(2015)在贡嘎山阔叶木本植物叶片解剖结构特征及其环境适应研究中发现的常绿植物的叶片厚度、栅栏组织厚度、海绵组织厚度、上表皮厚度和栅栏细胞层数都显著大于落叶植物的结果, 支持了Reich等(1998)的发现. ...
... ; Wright et al ., 2004 ; 潘少安等, 2015 ).并且, Reich等(1998)发现比叶面积较低的植物常形成厚度较大而面积较小的叶片, 而刘艳芳(2015)在贡嘎山阔叶木本植物叶片解剖结构特征及其环境适应研究中发现的常绿植物的叶片厚度、栅栏组织厚度、海绵组织厚度、上表皮厚度和栅栏细胞层数都显著大于落叶植物的结果, 支持了Reich等(1998)的发现. ...
The leaf economics spectrum worldwide.
2
2004
... 在木本植物中, 枝条的结构可以用枝条的茎长度、直径以及纤细率(枝条长度与直径比)来表示.对于用来着生和支撑叶、花和果实等器官的小枝而言, 其长度、纤细率以及展叶效率等性状的变化将直接影响植株的机械稳定性、光捕获效率和空间拓展能力, 进而影响到植株的光合和竞争能力的各个方面(Honda & Fisher, 1978 ; Niklas & Kerchner, 1984 ).例如, 树冠内叶片的分布模式是木本植物生产力的一个重要决定因素(Horn, 1971 ; Yagi & Kikuzawa, 1999 ), 而它的变化主要受小枝的结构变化模式、小枝在树冠内的空间排布方式以及小枝的个体大小等与枝条结构密切相关的因素限制(Koike, 1989 ; Yagi & Kikuzawa, 1999 ).在一些具有长短枝分化的物种中, 短枝的茎较短, 叶片排布更紧密, 长枝的茎较长, 叶片可以分散排布在较长的节间上(Yagi & Kikuzawa, 1999 ).尽管短枝上的总叶面积比长枝小(Gregory, 1980 ; Yagi & Kikuzawa, 1999 ), 但单位枝长基础上的叶面积比长枝大(Caesar & Macdonald, 1984 ; Yagi & Kikuzawa, 1999 ).小枝通过这种结构变化模式可以调控一个特定大小的小枝上的叶片数量(Maruyama, 1983 ; Koike, 1989 ; Takenaka, 1997 ; Yagi & Kikuzawa, 1999 )和展叶效率, 影响到植株的生产力.然而, 近年来, 关于植物小枝上茎-叶关系的研究多集中于茎直径维度的叶大小-小枝大小之间的关系研究(Westoby et al ., 2002 ; Pickup et al ., 2005 ; Wright et al ., 2007 ; Li et al ., 2008 ; Xiang et al ., 2009 ; Yang et al ., 2009 ; 杨冬梅等, 2012 ), 对于茎长维度的茎结构的另一维度——茎长度和纤细率对小枝展叶效率及相应的适应策略的研究则相对不足.小枝是个悬臂结构, 小枝茎长度不仅会影响其支撑叶片的排布, 还与小枝的机械安全密切相关.根据恒定胁迫模型(the constant-stress model)理论预测, 小枝茎横截面外缘所能承受的最大弯曲胁迫(M )主要由枝条长度(L )和枝条所承受的力(F )共同决定, 即M = L × F .对于一个无花、果等其他附属物的小枝而言, 枝条所承受的力主要来自风对枝条的压力和拉拽力所形成的风压胁迫, 而风压胁迫(F )又是叶面积(A )和风压(W )的乘积, 即F = A × W (Dean & Long, 1986 ).由此可知, 在风压一定的情况下, 小枝茎基部所能承受的最大弯曲胁迫是由枝条长度和叶面积决定的, 并且两者之间是负相关关系.所以, 我们预测小枝的展叶效率将随着小枝茎长度的增加而下降. ...
... 量不仅在每一个物种(或生境)组中(R 2 从0.315到0.814, p < 0.05), 而且在整个综合数据库中(R 2 = 0.615, p < 0.001)均存在显著的负相关关系(杨冬梅等, 2012 ).这种现象被归因于长枝比短枝具有更强的顶端优势, 侧芽更容易被抑制, 使得叶密度减小(Xiang et al ., 2009 ; Yagi & Kikuzawa, 1999 ; Yagi, 2000 ). ...
Stem architectural effect on leaf size, leaf number, and leaf mass fraction in plant twigs of woody species.
2
2009
... 本研究中, 无论是落叶还是常绿物种, 用以表征展叶效率的叶面积比、叶密度、叶茎生物量比, 均与表征当年生小枝茎结构的小枝茎长度和茎纤细率呈显著的负相关关系(图1 , 图3 , 图4 ), 即随着当年生小枝茎长度和纤细率的增加, 小枝的展叶效率逐渐降低.这一结果与Takenaka和Yagi等的研究结果(Takenaka, 1997 ; Yagi & Kikuzawa, 1999 ; Yagi, 2000 , 2004 )类似, 他们在多个木本植物的种内研究发现, 较长的当年生枝条具有更小的单位枝长基础上的叶面积.而叶密度可能是小枝展叶效率的一个主要调节因素.我们的结果显示, 无论是在常绿还是落叶物种中, 不仅小枝茎长度和茎纤细率均与叶密度(叶数量/茎长度)呈显著的负相关关系(图3 ), 并且每个小枝上的叶数量也直接与小枝茎长度呈斜率显著小于1.0 (p < 0.000β1)的正的异速生长关系(图2 ; 表1 ), 表明叶数量的增加速度没有小枝茎长度的增加速度快, 长的枝条相对于短枝具有更低的出叶强度.Yagi (2004)的研究结果也表明, 长的当年生小枝比短的当年生小枝具有更小的叶数量与枝长度的比值, 即单位枝长度的叶片数更少.在我们以前的研究中也发现, 小枝茎长度和单位茎长度的叶片数 ...
... 量不仅在每一个物种(或生境)组中(R 2 从0.315到0.814, p < 0.05), 而且在整个综合数据库中(R 2 = 0.615, p < 0.001)均存在显著的负相关关系(杨冬梅等, 2012 ).这种现象被归因于长枝比短枝具有更强的顶端优势, 侧芽更容易被抑制, 使得叶密度减小(Xiang et al ., 2009 ; Yagi & Kikuzawa, 1999 ; Yagi, 2000 ). ...
Morphology and biomass allocation of current-year shoots of ten tall tree species in cool temperate Japan.
1
2000
... 本研究中, 无论是落叶还是常绿物种, 用以表征展叶效率的叶面积比、叶密度、叶茎生物量比, 均与表征当年生小枝茎结构的小枝茎长度和茎纤细率呈显著的负相关关系(图1 , 图3 , 图4 ), 即随着当年生小枝茎长度和纤细率的增加, 小枝的展叶效率逐渐降低.这一结果与Takenaka和Yagi等的研究结果(Takenaka, 1997 ; Yagi & Kikuzawa, 1999 ; Yagi, 2000 , 2004 )类似, 他们在多个木本植物的种内研究发现, 较长的当年生枝条具有更小的单位枝长基础上的叶面积.而叶密度可能是小枝展叶效率的一个主要调节因素.我们的结果显示, 无论是在常绿还是落叶物种中, 不仅小枝茎长度和茎纤细率均与叶密度(叶数量/茎长度)呈显著的负相关关系(图3 ), 并且每个小枝上的叶数量也直接与小枝茎长度呈斜率显著小于1.0 (p < 0.000β1)的正的异速生长关系(图2 ; 表1 ), 表明叶数量的增加速度没有小枝茎长度的增加速度快, 长的枝条相对于短枝具有更低的出叶强度.Yagi (2004)的研究结果也表明, 长的当年生小枝比短的当年生小枝具有更小的叶数量与枝长度的比值, 即单位枝长度的叶片数更少.在我们以前的研究中也发现, 小枝茎长度和单位茎长度的叶片数 ...
Within-tree variations in shoot differentiation patterns of 10 tall tree species in a Japanese cool-temperate forest.
9
2004
... 在木本植物中, 枝条的结构可以用枝条的茎长度、直径以及纤细率(枝条长度与直径比)来表示.对于用来着生和支撑叶、花和果实等器官的小枝而言, 其长度、纤细率以及展叶效率等性状的变化将直接影响植株的机械稳定性、光捕获效率和空间拓展能力, 进而影响到植株的光合和竞争能力的各个方面(Honda & Fisher, 1978 ; Niklas & Kerchner, 1984 ).例如, 树冠内叶片的分布模式是木本植物生产力的一个重要决定因素(Horn, 1971 ; Yagi & Kikuzawa, 1999 ), 而它的变化主要受小枝的结构变化模式、小枝在树冠内的空间排布方式以及小枝的个体大小等与枝条结构密切相关的因素限制(Koike, 1989 ; Yagi & Kikuzawa, 1999 ).在一些具有长短枝分化的物种中, 短枝的茎较短, 叶片排布更紧密, 长枝的茎较长, 叶片可以分散排布在较长的节间上(Yagi & Kikuzawa, 1999 ).尽管短枝上的总叶面积比长枝小(Gregory, 1980 ; Yagi & Kikuzawa, 1999 ), 但单位枝长基础上的叶面积比长枝大(Caesar & Macdonald, 1984 ; Yagi & Kikuzawa, 1999 ).小枝通过这种结构变化模式可以调控一个特定大小的小枝上的叶片数量(Maruyama, 1983 ; Koike, 1989 ; Takenaka, 1997 ; Yagi & Kikuzawa, 1999 )和展叶效率, 影响到植株的生产力.然而, 近年来, 关于植物小枝上茎-叶关系的研究多集中于茎直径维度的叶大小-小枝大小之间的关系研究(Westoby et al ., 2002 ; Pickup et al ., 2005 ; Wright et al ., 2007 ; Li et al ., 2008 ; Xiang et al ., 2009 ; Yang et al ., 2009 ; 杨冬梅等, 2012 ), 对于茎长维度的茎结构的另一维度——茎长度和纤细率对小枝展叶效率及相应的适应策略的研究则相对不足.小枝是个悬臂结构, 小枝茎长度不仅会影响其支撑叶片的排布, 还与小枝的机械安全密切相关.根据恒定胁迫模型(the constant-stress model)理论预测, 小枝茎横截面外缘所能承受的最大弯曲胁迫(M )主要由枝条长度(L )和枝条所承受的力(F )共同决定, 即M = L × F .对于一个无花、果等其他附属物的小枝而言, 枝条所承受的力主要来自风对枝条的压力和拉拽力所形成的风压胁迫, 而风压胁迫(F )又是叶面积(A )和风压(W )的乘积, 即F = A × W (Dean & Long, 1986 ).由此可知, 在风压一定的情况下, 小枝茎基部所能承受的最大弯曲胁迫是由枝条长度和叶面积决定的, 并且两者之间是负相关关系.所以, 我们预测小枝的展叶效率将随着小枝茎长度的增加而下降. ...
... ; Yagi & Kikuzawa, 1999 ).在一些具有长短枝分化的物种中, 短枝的茎较短, 叶片排布更紧密, 长枝的茎较长, 叶片可以分散排布在较长的节间上(Yagi & Kikuzawa, 1999 ).尽管短枝上的总叶面积比长枝小(Gregory, 1980 ; Yagi & Kikuzawa, 1999 ), 但单位枝长基础上的叶面积比长枝大(Caesar & Macdonald, 1984 ; Yagi & Kikuzawa, 1999 ).小枝通过这种结构变化模式可以调控一个特定大小的小枝上的叶片数量(Maruyama, 1983 ; Koike, 1989 ; Takenaka, 1997 ; Yagi & Kikuzawa, 1999 )和展叶效率, 影响到植株的生产力.然而, 近年来, 关于植物小枝上茎-叶关系的研究多集中于茎直径维度的叶大小-小枝大小之间的关系研究(Westoby et al ., 2002 ; Pickup et al ., 2005 ; Wright et al ., 2007 ; Li et al ., 2008 ; Xiang et al ., 2009 ; Yang et al ., 2009 ; 杨冬梅等, 2012 ), 对于茎长维度的茎结构的另一维度——茎长度和纤细率对小枝展叶效率及相应的适应策略的研究则相对不足.小枝是个悬臂结构, 小枝茎长度不仅会影响其支撑叶片的排布, 还与小枝的机械安全密切相关.根据恒定胁迫模型(the constant-stress model)理论预测, 小枝茎横截面外缘所能承受的最大弯曲胁迫(M )主要由枝条长度(L )和枝条所承受的力(F )共同决定, 即M = L × F .对于一个无花、果等其他附属物的小枝而言, 枝条所承受的力主要来自风对枝条的压力和拉拽力所形成的风压胁迫, 而风压胁迫(F )又是叶面积(A )和风压(W )的乘积, 即F = A × W (Dean & Long, 1986 ).由此可知, 在风压一定的情况下, 小枝茎基部所能承受的最大弯曲胁迫是由枝条长度和叶面积决定的, 并且两者之间是负相关关系.所以, 我们预测小枝的展叶效率将随着小枝茎长度的增加而下降. ...
... ).在一些具有长短枝分化的物种中, 短枝的茎较短, 叶片排布更紧密, 长枝的茎较长, 叶片可以分散排布在较长的节间上(Yagi & Kikuzawa, 1999 ).尽管短枝上的总叶面积比长枝小(Gregory, 1980 ; Yagi & Kikuzawa, 1999 ), 但单位枝长基础上的叶面积比长枝大(Caesar & Macdonald, 1984 ; Yagi & Kikuzawa, 1999 ).小枝通过这种结构变化模式可以调控一个特定大小的小枝上的叶片数量(Maruyama, 1983 ; Koike, 1989 ; Takenaka, 1997 ; Yagi & Kikuzawa, 1999 )和展叶效率, 影响到植株的生产力.然而, 近年来, 关于植物小枝上茎-叶关系的研究多集中于茎直径维度的叶大小-小枝大小之间的关系研究(Westoby et al ., 2002 ; Pickup et al ., 2005 ; Wright et al ., 2007 ; Li et al ., 2008 ; Xiang et al ., 2009 ; Yang et al ., 2009 ; 杨冬梅等, 2012 ), 对于茎长维度的茎结构的另一维度——茎长度和纤细率对小枝展叶效率及相应的适应策略的研究则相对不足.小枝是个悬臂结构, 小枝茎长度不仅会影响其支撑叶片的排布, 还与小枝的机械安全密切相关.根据恒定胁迫模型(the constant-stress model)理论预测, 小枝茎横截面外缘所能承受的最大弯曲胁迫(M )主要由枝条长度(L )和枝条所承受的力(F )共同决定, 即M = L × F .对于一个无花、果等其他附属物的小枝而言, 枝条所承受的力主要来自风对枝条的压力和拉拽力所形成的风压胁迫, 而风压胁迫(F )又是叶面积(A )和风压(W )的乘积, 即F = A × W (Dean & Long, 1986 ).由此可知, 在风压一定的情况下, 小枝茎基部所能承受的最大弯曲胁迫是由枝条长度和叶面积决定的, 并且两者之间是负相关关系.所以, 我们预测小枝的展叶效率将随着小枝茎长度的增加而下降. ...
... ; Yagi & Kikuzawa, 1999 ), 但单位枝长基础上的叶面积比长枝大(Caesar & Macdonald, 1984 ; Yagi & Kikuzawa, 1999 ).小枝通过这种结构变化模式可以调控一个特定大小的小枝上的叶片数量(Maruyama, 1983 ; Koike, 1989 ; Takenaka, 1997 ; Yagi & Kikuzawa, 1999 )和展叶效率, 影响到植株的生产力.然而, 近年来, 关于植物小枝上茎-叶关系的研究多集中于茎直径维度的叶大小-小枝大小之间的关系研究(Westoby et al ., 2002 ; Pickup et al ., 2005 ; Wright et al ., 2007 ; Li et al ., 2008 ; Xiang et al ., 2009 ; Yang et al ., 2009 ; 杨冬梅等, 2012 ), 对于茎长维度的茎结构的另一维度——茎长度和纤细率对小枝展叶效率及相应的适应策略的研究则相对不足.小枝是个悬臂结构, 小枝茎长度不仅会影响其支撑叶片的排布, 还与小枝的机械安全密切相关.根据恒定胁迫模型(the constant-stress model)理论预测, 小枝茎横截面外缘所能承受的最大弯曲胁迫(M )主要由枝条长度(L )和枝条所承受的力(F )共同决定, 即M = L × F .对于一个无花、果等其他附属物的小枝而言, 枝条所承受的力主要来自风对枝条的压力和拉拽力所形成的风压胁迫, 而风压胁迫(F )又是叶面积(A )和风压(W )的乘积, 即F = A × W (Dean & Long, 1986 ).由此可知, 在风压一定的情况下, 小枝茎基部所能承受的最大弯曲胁迫是由枝条长度和叶面积决定的, 并且两者之间是负相关关系.所以, 我们预测小枝的展叶效率将随着小枝茎长度的增加而下降. ...
... ; Yagi & Kikuzawa, 1999 ).小枝通过这种结构变化模式可以调控一个特定大小的小枝上的叶片数量(Maruyama, 1983 ; Koike, 1989 ; Takenaka, 1997 ; Yagi & Kikuzawa, 1999 )和展叶效率, 影响到植株的生产力.然而, 近年来, 关于植物小枝上茎-叶关系的研究多集中于茎直径维度的叶大小-小枝大小之间的关系研究(Westoby et al ., 2002 ; Pickup et al ., 2005 ; Wright et al ., 2007 ; Li et al ., 2008 ; Xiang et al ., 2009 ; Yang et al ., 2009 ; 杨冬梅等, 2012 ), 对于茎长维度的茎结构的另一维度——茎长度和纤细率对小枝展叶效率及相应的适应策略的研究则相对不足.小枝是个悬臂结构, 小枝茎长度不仅会影响其支撑叶片的排布, 还与小枝的机械安全密切相关.根据恒定胁迫模型(the constant-stress model)理论预测, 小枝茎横截面外缘所能承受的最大弯曲胁迫(M )主要由枝条长度(L )和枝条所承受的力(F )共同决定, 即M = L × F .对于一个无花、果等其他附属物的小枝而言, 枝条所承受的力主要来自风对枝条的压力和拉拽力所形成的风压胁迫, 而风压胁迫(F )又是叶面积(A )和风压(W )的乘积, 即F = A × W (Dean & Long, 1986 ).由此可知, 在风压一定的情况下, 小枝茎基部所能承受的最大弯曲胁迫是由枝条长度和叶面积决定的, 并且两者之间是负相关关系.所以, 我们预测小枝的展叶效率将随着小枝茎长度的增加而下降. ...
... ; Yagi & Kikuzawa, 1999 )和展叶效率, 影响到植株的生产力.然而, 近年来, 关于植物小枝上茎-叶关系的研究多集中于茎直径维度的叶大小-小枝大小之间的关系研究(Westoby et al ., 2002 ; Pickup et al ., 2005 ; Wright et al ., 2007 ; Li et al ., 2008 ; Xiang et al ., 2009 ; Yang et al ., 2009 ; 杨冬梅等, 2012 ), 对于茎长维度的茎结构的另一维度——茎长度和纤细率对小枝展叶效率及相应的适应策略的研究则相对不足.小枝是个悬臂结构, 小枝茎长度不仅会影响其支撑叶片的排布, 还与小枝的机械安全密切相关.根据恒定胁迫模型(the constant-stress model)理论预测, 小枝茎横截面外缘所能承受的最大弯曲胁迫(M )主要由枝条长度(L )和枝条所承受的力(F )共同决定, 即M = L × F .对于一个无花、果等其他附属物的小枝而言, 枝条所承受的力主要来自风对枝条的压力和拉拽力所形成的风压胁迫, 而风压胁迫(F )又是叶面积(A )和风压(W )的乘积, 即F = A × W (Dean & Long, 1986 ).由此可知, 在风压一定的情况下, 小枝茎基部所能承受的最大弯曲胁迫是由枝条长度和叶面积决定的, 并且两者之间是负相关关系.所以, 我们预测小枝的展叶效率将随着小枝茎长度的增加而下降. ...
... 本研究中, 无论是落叶还是常绿物种, 用以表征展叶效率的叶面积比、叶密度、叶茎生物量比, 均与表征当年生小枝茎结构的小枝茎长度和茎纤细率呈显著的负相关关系(图1 , 图3 , 图4 ), 即随着当年生小枝茎长度和纤细率的增加, 小枝的展叶效率逐渐降低.这一结果与Takenaka和Yagi等的研究结果(Takenaka, 1997 ; Yagi & Kikuzawa, 1999 ; Yagi, 2000 , 2004 )类似, 他们在多个木本植物的种内研究发现, 较长的当年生枝条具有更小的单位枝长基础上的叶面积.而叶密度可能是小枝展叶效率的一个主要调节因素.我们的结果显示, 无论是在常绿还是落叶物种中, 不仅小枝茎长度和茎纤细率均与叶密度(叶数量/茎长度)呈显著的负相关关系(图3 ), 并且每个小枝上的叶数量也直接与小枝茎长度呈斜率显著小于1.0 (p < 0.000β1)的正的异速生长关系(图2 ; 表1 ), 表明叶数量的增加速度没有小枝茎长度的增加速度快, 长的枝条相对于短枝具有更低的出叶强度.Yagi (2004)的研究结果也表明, 长的当年生小枝比短的当年生小枝具有更小的叶数量与枝长度的比值, 即单位枝长度的叶片数更少.在我们以前的研究中也发现, 小枝茎长度和单位茎长度的叶片数 ...
... 量不仅在每一个物种(或生境)组中(R 2 从0.315到0.814, p < 0.05), 而且在整个综合数据库中(R 2 = 0.615, p < 0.001)均存在显著的负相关关系(杨冬梅等, 2012 ).这种现象被归因于长枝比短枝具有更强的顶端优势, 侧芽更容易被抑制, 使得叶密度减小(Xiang et al ., 2009 ; Yagi & Kikuzawa, 1999 ; Yagi, 2000 ). ...
... 从机械力学的角度看, 展叶效率随小枝茎长度和茎纤细率的增加而下降可能是小枝在面对机械载重和风压胁迫时, 确保小枝安全的需要.作为起支撑作用的小枝, 首先要能够承受其自身的质量, 避免折断、撕裂或其他形式的机械损伤(Mcmahon, 1973 ; King & Loucks, 1978 ; King, 1981 , 1986 ; Long et al ., 1981 ; Givnish, 1986 ; Gartner, 1995 ), 必须能够抵抗来自茎及其上叶片质量所带来的静态和动态的压力和拉力胁迫.恒定胁迫模型(The constant-stress model)理论认为, 令植物茎横截面外缘弯曲所需要做的功(即弯曲矩, the bending moment) M 主要是由力F 和臂长(即茎长) L 共同决定, 即M = F × L .对于植物而言, 茎所承受的力是叶面积A 与风压W 的乘积, 即F = A × W (Dean & Long, 1986 ).所以, 随着小枝茎长度的增加, 在小枝上相同叶面积对小枝茎基部的机械负载会相应地增加, 特别是在有风吹动摇摆的情况下, 长枝将相应地减少枝条上的叶片面积或干质量来降低机械载重和风压胁迫, 导致展叶效率随小枝茎长度和茎纤细率的增加而下降.其次, 从植株树冠构建的角度看, 枝条结构与展叶效率之间的负相关关系将有利于植株更有效地捕获光能, 提高整体光合效率.因为相互遮阴是当年生小枝结构变化的一个重要决定因素, 更大的展叶密度将会导致当年生小枝内叶片之间相互荫蔽的程度增加(Niklas, 1988 ; Yagi & Kikuzawa, 1999 ).另外, 在总的生物量一定的情况下, 当植物将更多的生物量分配给茎以使枝条生长得更长时, 将有利于植株将更多的叶片伸展到冠层外部, 同时可以让一定数量的叶片分散在更长的枝条范围内, 降低同一枝条上叶片之间相互荫蔽的程度, 以利于捕获更多的光能, 提高植株的光合效率.综上所述, 枝条结构与展叶效率之间的负相关关系具有重要的生态学意义, 有利于保持小枝的机械安全, 并提高整个植株的光合效率. ...
Patterns in size-related variations in current-year shoot structure in eight deciduous tree species.
1
1999
... 常绿与落叶是植物通过长期进化适应形成的两种不同的生活史对策, 它们在叶寿命和叶片结构(如比叶面积)(Reich et al ., 1992 ; Westoby et al ., 2002 ; Wright et al ., 2004 ; Li et al ., 2008 )、茎木质部抗栓塞能力(Cavender-Bares & Holbrook, 2001 ; Cavender- Bares et al ., 2005 )等许多方面都显著不同.在本研究中, 虽然当年生小枝茎结构性状与表征展叶效率的性状之间的负相关关系斜率在常绿和落叶物种间无显著差异, 但截距差异显著.在3个表征展叶效率的性状中, 尽管常绿物种叶密度与小枝茎结构性状之间的关系截距显著大于落叶物种, 在某一给定的茎长或茎纤细率下, 常绿物种比落叶物种具有更多的叶数量, 即具有较高的出叶强度, 与以前相关研究发现的叶大小与叶数量之间是相互权衡的结果一致(Corner, 1949 ; Kleiman & Aarssen, 2007 ; Yang et al ., 2008 ).但由于常绿物种的单叶面积16.21 cm2 显著小于落叶物种的单叶面积34.66 cm2 (p < 0.001, 表2 ), 导致其叶面积比(单位茎质量的叶面积)和叶茎生物量比(单位茎质量的叶质量)与小枝茎结构性状之间的关系截距反而显著小于落叶物种, 即随着小枝茎长度和纤细率的增加, 常绿物种每单位质量的茎生物量投资获得的总叶面积和总叶质量都比落叶物种少, 常绿物种的实际展叶效率比落叶物种低.对数据结果进一步分析表明, 常绿物种更低展叶效率的主要原因是其比叶面积显著小于落叶物种 (p < 0.001, 表2 ), 因为我们的结果显示, 在不考虑小枝茎长结构影响的情况下, 茎叶生物量比在常绿和落叶物种中没有显著差异(p = 0.335, 表2 ), 表明总生物量在叶和茎之间的分配比例在常绿和落叶物种中是没有显著差异的, 可是落叶物种的单叶面积(p < 0.001, 表2 )和总叶面积(p = 0.001, 表2 )都要显著大于常绿物种, 并且常绿物种茎干质量与总叶面积相关关系共同斜率下的截距显著小于落叶物种(p ˂ 0.05, 表1 ), 即在某一给定的茎干质量下, 常绿物种比落叶物种具有更小的叶面积.许多研究也都发现常绿物种通常比落叶物种具有更高的比叶质量(即比叶面积的倒数)(Reich et al ., 1992 ; Westoby et al ., 2002 ; Wright et al ., 2004 ; 潘少安等, 2015 ).并且, Reich等(1998)发现比叶面积较低的植物常形成厚度较大而面积较小的叶片, 而刘艳芳(2015)在贡嘎山阔叶木本植物叶片解剖结构特征及其环境适应研究中发现的常绿植物的叶片厚度、栅栏组织厚度、海绵组织厚度、上表皮厚度和栅栏细胞层数都显著大于落叶植物的结果, 支持了Reich等(1998)的发现. ...
The generality of leaf size versus number trade-off in temperate woody species.
2
2008
... 在木本植物中, 枝条的结构可以用枝条的茎长度、直径以及纤细率(枝条长度与直径比)来表示.对于用来着生和支撑叶、花和果实等器官的小枝而言, 其长度、纤细率以及展叶效率等性状的变化将直接影响植株的机械稳定性、光捕获效率和空间拓展能力, 进而影响到植株的光合和竞争能力的各个方面(Honda & Fisher, 1978 ; Niklas & Kerchner, 1984 ).例如, 树冠内叶片的分布模式是木本植物生产力的一个重要决定因素(Horn, 1971 ; Yagi & Kikuzawa, 1999 ), 而它的变化主要受小枝的结构变化模式、小枝在树冠内的空间排布方式以及小枝的个体大小等与枝条结构密切相关的因素限制(Koike, 1989 ; Yagi & Kikuzawa, 1999 ).在一些具有长短枝分化的物种中, 短枝的茎较短, 叶片排布更紧密, 长枝的茎较长, 叶片可以分散排布在较长的节间上(Yagi & Kikuzawa, 1999 ).尽管短枝上的总叶面积比长枝小(Gregory, 1980 ; Yagi & Kikuzawa, 1999 ), 但单位枝长基础上的叶面积比长枝大(Caesar & Macdonald, 1984 ; Yagi & Kikuzawa, 1999 ).小枝通过这种结构变化模式可以调控一个特定大小的小枝上的叶片数量(Maruyama, 1983 ; Koike, 1989 ; Takenaka, 1997 ; Yagi & Kikuzawa, 1999 )和展叶效率, 影响到植株的生产力.然而, 近年来, 关于植物小枝上茎-叶关系的研究多集中于茎直径维度的叶大小-小枝大小之间的关系研究(Westoby et al ., 2002 ; Pickup et al ., 2005 ; Wright et al ., 2007 ; Li et al ., 2008 ; Xiang et al ., 2009 ; Yang et al ., 2009 ; 杨冬梅等, 2012 ), 对于茎长维度的茎结构的另一维度——茎长度和纤细率对小枝展叶效率及相应的适应策略的研究则相对不足.小枝是个悬臂结构, 小枝茎长度不仅会影响其支撑叶片的排布, 还与小枝的机械安全密切相关.根据恒定胁迫模型(the constant-stress model)理论预测, 小枝茎横截面外缘所能承受的最大弯曲胁迫(M )主要由枝条长度(L )和枝条所承受的力(F )共同决定, 即M = L × F .对于一个无花、果等其他附属物的小枝而言, 枝条所承受的力主要来自风对枝条的压力和拉拽力所形成的风压胁迫, 而风压胁迫(F )又是叶面积(A )和风压(W )的乘积, 即F = A × W (Dean & Long, 1986 ).由此可知, 在风压一定的情况下, 小枝茎基部所能承受的最大弯曲胁迫是由枝条长度和叶面积决定的, 并且两者之间是负相关关系.所以, 我们预测小枝的展叶效率将随着小枝茎长度的增加而下降. ...
... 同时, 对于生活在同一群落里的常绿和落叶物种而言, 由于展叶物候和生物学习性的不同, 它们的功能特征通常也会有很大的差异.例如, 相比于落叶物种, 常绿物种通常具有长的叶寿命、低的比叶面积(SLA )、低的光合速率和高的养分含量(Reich et al ., 1992 ; Westoby et al ., 2002 ; Wright et al ., 2004 ; Li et al ., 2008 ); 另外, 由于常绿物种为了确保在冬季低温等不利环境下叶片不凋落, 需要水分运输系统保持畅通, 确保水分运输安全, 其木质部需要比落叶物种具有更强的抵抗冰冻引起栓塞的能力(Cavender-Bares & Holbrook, 2001 ; Cavender-Bares et al ., 2005 ), 致使其投入在支撑结构上的成本增加, 表现出在相同的茎投入时, 常绿物种通常比落叶物种具有更小的总叶面积和单叶面积(Sun et al ., 2006 ; Yang et al ., 2009 ).这种不同的成本-收益关系将有可能导致常绿和落叶物种采取不同的枝条伸展策略以改变小枝上的叶排布方式, 使两类物种都能够捕获足够的光能以保持相对稳定的碳收益.因此, 两类物种可能会具有不同的茎结构与展叶效率之间的变化关系. ...
The effects of leaf size, leaf habit, and leaf form on leaf/stem relationships in plant twigs of temperate woody species.
2
2009
... 在木本植物中, 枝条的结构可以用枝条的茎长度、直径以及纤细率(枝条长度与直径比)来表示.对于用来着生和支撑叶、花和果实等器官的小枝而言, 其长度、纤细率以及展叶效率等性状的变化将直接影响植株的机械稳定性、光捕获效率和空间拓展能力, 进而影响到植株的光合和竞争能力的各个方面(Honda & Fisher, 1978 ; Niklas & Kerchner, 1984 ).例如, 树冠内叶片的分布模式是木本植物生产力的一个重要决定因素(Horn, 1971 ; Yagi & Kikuzawa, 1999 ), 而它的变化主要受小枝的结构变化模式、小枝在树冠内的空间排布方式以及小枝的个体大小等与枝条结构密切相关的因素限制(Koike, 1989 ; Yagi & Kikuzawa, 1999 ).在一些具有长短枝分化的物种中, 短枝的茎较短, 叶片排布更紧密, 长枝的茎较长, 叶片可以分散排布在较长的节间上(Yagi & Kikuzawa, 1999 ).尽管短枝上的总叶面积比长枝小(Gregory, 1980 ; Yagi & Kikuzawa, 1999 ), 但单位枝长基础上的叶面积比长枝大(Caesar & Macdonald, 1984 ; Yagi & Kikuzawa, 1999 ).小枝通过这种结构变化模式可以调控一个特定大小的小枝上的叶片数量(Maruyama, 1983 ; Koike, 1989 ; Takenaka, 1997 ; Yagi & Kikuzawa, 1999 )和展叶效率, 影响到植株的生产力.然而, 近年来, 关于植物小枝上茎-叶关系的研究多集中于茎直径维度的叶大小-小枝大小之间的关系研究(Westoby et al ., 2002 ; Pickup et al ., 2005 ; Wright et al ., 2007 ; Li et al ., 2008 ; Xiang et al ., 2009 ; Yang et al ., 2009 ; 杨冬梅等, 2012 ), 对于茎长维度的茎结构的另一维度——茎长度和纤细率对小枝展叶效率及相应的适应策略的研究则相对不足.小枝是个悬臂结构, 小枝茎长度不仅会影响其支撑叶片的排布, 还与小枝的机械安全密切相关.根据恒定胁迫模型(the constant-stress model)理论预测, 小枝茎横截面外缘所能承受的最大弯曲胁迫(M )主要由枝条长度(L )和枝条所承受的力(F )共同决定, 即M = L × F .对于一个无花、果等其他附属物的小枝而言, 枝条所承受的力主要来自风对枝条的压力和拉拽力所形成的风压胁迫, 而风压胁迫(F )又是叶面积(A )和风压(W )的乘积, 即F = A × W (Dean & Long, 1986 ).由此可知, 在风压一定的情况下, 小枝茎基部所能承受的最大弯曲胁迫是由枝条长度和叶面积决定的, 并且两者之间是负相关关系.所以, 我们预测小枝的展叶效率将随着小枝茎长度的增加而下降. ...
... 量不仅在每一个物种(或生境)组中(R 2 从0.315到0.814, p < 0.05), 而且在整个综合数据库中(R 2 = 0.615, p < 0.001)均存在显著的负相关关系(杨冬梅等, 2012 ).这种现象被归因于长枝比短枝具有更强的顶端优势, 侧芽更容易被抑制, 使得叶密度减小(Xiang et al ., 2009 ; Yagi & Kikuzawa, 1999 ; Yagi, 2000 ). ...
清凉峰不同海拔木本植物小枝内叶大小-数量权衡关系
3
2012
... 常绿物种较低的比叶面积继而导致较低的单位茎结构基础上的展叶效率, 可能与其具有较长的叶寿命有关.一般而言, 叶寿命越长, 叶片越厚, 单位叶面积的建成消耗也越大, 叶子需要在足够长的时间内通过自身的光合生产所形成的有机碳来补偿初始的建成消耗(包括抵抗草食动物的防御构造), 并维持自身生存所需的物质消耗(Coley, 1988 ; Kikuzawa, 1991 ; Schulze et al ., 1994 ; Kikuzawa, 1995 ; 张林和罗天祥, 2004 ).所以叶寿命与比叶面积成反比(Reich et al ., 1992 ; Reich & Bowman, 1999 ; Ryser & Urbas, 2000 ; 张林和罗天祥, 2004 ).比叶面积较低的植物, 叶片中很大一部分物质用于构建保卫构造(防虫食、防止过度失水等)或者增加叶肉细胞密度以抵御环境胁迫(Reich et al ., 1998 ; 张林和罗天祥, 2004 ).与落叶物种相比, 常绿物种叶片在冬季不凋落, 需经受较长时间的冬季低温等不利生长环境, 面对冬季的选择压力, 常绿植物可能采取高消耗慢收益的保守型生存策略(Cornelissen, 1999 ; 刘艳芳, 2015 ), 通过革质的叶表皮和叶肉海绵组织分层等特点来增加抗寒力(董丽等, 2003 ; 刘艳芳, 2015 ).而落叶植物的叶寿命相对较短, 需要在相对短的生长时间内积累足够的光合产物以满足植株生长需要, 薄的叶片组织能够缩短CO2 从气孔扩散到叶绿体的路径, 也能增加气体交换的速率, 进而提高光合作用速率(Diemer, 1998 ; 刘艳芳, 2015 ), 所以, 较高的SLA 是落叶物种所采取的一种低投入快收益的积极型生存策略. ...
... ; 张林和罗天祥, 2004 ).比叶面积较低的植物, 叶片中很大一部分物质用于构建保卫构造(防虫食、防止过度失水等)或者增加叶肉细胞密度以抵御环境胁迫(Reich et al ., 1998 ; 张林和罗天祥, 2004 ).与落叶物种相比, 常绿物种叶片在冬季不凋落, 需经受较长时间的冬季低温等不利生长环境, 面对冬季的选择压力, 常绿植物可能采取高消耗慢收益的保守型生存策略(Cornelissen, 1999 ; 刘艳芳, 2015 ), 通过革质的叶表皮和叶肉海绵组织分层等特点来增加抗寒力(董丽等, 2003 ; 刘艳芳, 2015 ).而落叶植物的叶寿命相对较短, 需要在相对短的生长时间内积累足够的光合产物以满足植株生长需要, 薄的叶片组织能够缩短CO2 从气孔扩散到叶绿体的路径, 也能增加气体交换的速率, 进而提高光合作用速率(Diemer, 1998 ; 刘艳芳, 2015 ), 所以, 较高的SLA 是落叶物种所采取的一种低投入快收益的积极型生存策略. ...
... ; 张林和罗天祥, 2004 ).与落叶物种相比, 常绿物种叶片在冬季不凋落, 需经受较长时间的冬季低温等不利生长环境, 面对冬季的选择压力, 常绿植物可能采取高消耗慢收益的保守型生存策略(Cornelissen, 1999 ; 刘艳芳, 2015 ), 通过革质的叶表皮和叶肉海绵组织分层等特点来增加抗寒力(董丽等, 2003 ; 刘艳芳, 2015 ).而落叶植物的叶寿命相对较短, 需要在相对短的生长时间内积累足够的光合产物以满足植株生长需要, 薄的叶片组织能够缩短CO2 从气孔扩散到叶绿体的路径, 也能增加气体交换的速率, 进而提高光合作用速率(Diemer, 1998 ; 刘艳芳, 2015 ), 所以, 较高的SLA 是落叶物种所采取的一种低投入快收益的积极型生存策略. ...
植物叶寿命及其相关叶性状的生态学研究进展
1
2004
... 由枝和叶所构成的树冠是植物进行光合作用、呼吸作用、蒸腾作用等一系列生理活动的主要场所, 其大小、结构、形状及相互间的协变规律直接决定了植物个体内的生物量分配、生长活力和生产力.枝条作为植株的重要结构单元, 不仅是植株运输水分和营养物质的重要通道, 也是支撑树冠的主要结构并直接影响树冠的形状和大小, 同时它还是叶片的支撑体, 能通过其长度、结构、分布方式等变化来调整冠层内叶片的排布方式, 将叶伸展到适宜的空间形成一个合理的光拦截体系, 提高光合效率(黎云祥和刘玉成, 1996 ; Day et al ., 2002 ; Thomas & Winner, 2002 ; 张运春等, 2005 ).叶片是植物进行光合作用的主要器官, 叶片的数量、面积以及展叶效率等空间结构特征的变化直接影响着植株的光拦截效率和光合效率.因此, 枝条的结构特征及其与叶性状之间的协变规律反映植物对光照、空间等资源的利用和适应策略(Borchert & Slade, 1981 ; Bicknell, 1982 ; Boojh & Ramakrishnan, 1982 ; Bell, 1984 ; 陈波等, 2002 ). ...


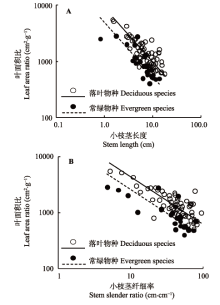
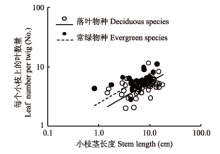


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}