


据统计, 全球大多数地区处于干旱、半干旱状态, 只有不足10%的土地较适合种植农作物, 而我国有近48%的陆地处于干旱、半干旱地区(李合生, 2012)。干旱胁迫是植物较为频繁遭受的一种逆境。植物在整个生命周期中, 常常会遭遇来自大气或者土壤的水分亏缺(Wright et al., 2004)。干旱胁迫会干扰植物体内的许多生理生化过程, 造成活性氧(ROS)增多、膜稳定性降低、蛋白质变性增加、离子平衡变化、新陈代谢紊乱及物理损伤等(Taiz & Zeiger, 2010), 致使植物的生产力降低(Blackman et al., 2010), 而极端干旱可能威胁到植物的存活(Tyree & Zimmermann, 2002; Breshears et al., 2005)。
在遭遇极端环境时, 植物会采取合适的应对机制以避免或减轻伤害, 如植物会合成抗氧化剂或诱导抗氧化酶活性升高, 以清除ROS自由基, 避免或减轻细胞受到的氧化损伤(Taiz & Zeiger, 2010)。不过, 过强的环境胁迫通常会引起ROS产生和清除机制的失衡, 导致植物细胞遭受明显氧化损伤。植物还会通过在体内积累溶质, 采用增强渗透调节的方式来应对干旱胁迫(Taiz & Zeiger, 2010; 李合生, 2012; 潘瑞炽, 2012)。
氮是植物必需的矿质元素, 植物通过氮代谢会合成少量的含氮化合物于生活的细胞中, 它们起着重要的生理生化作用。在这些化合物中有组成原生质的结构蛋白以及催化生物化学过程的酶。在植物处于水分亏缺状态时, 氮明显影响到植物的生长发育、生理代谢和水分利用效率(Lawlor & Cornic, 2002)。因此, 氮对植物的抗旱性很可能有一定影响。
桢楠(Phoebe zhennan)是亚热带常绿阔叶高大乔木, 生长于海拔1β100 m以下的地区, 为国家二级保护植物, 是我国最著名的一种用材树种(方文培, 1981)。近年来, 国家及地方政府倡导大力发展和保护珍稀种质资源, 四川在多地营造了桢楠人工林, 并在城镇园林建设和四旁绿化中大量栽植。然而, 在一些季节性干旱地区, 普遍存在着桢楠幼树因土壤缺水而生长不良甚至枯死的情况, 这严重制约了桢楠人工林的发展。在应对干旱缺水问题时, 除采取必要的给水措施外, 充分了解树木的生态学特性, 采用科学的培育措施(如施肥), 发挥和利用其自身的抗旱潜力也是一个重要的途径。因此, 本研究以二年生桢楠幼树为研究对象, 一方面分析其抗性生理对干旱胁迫的响应, 另一方面探讨施氮能否缓解干旱胁迫对其幼树的不利影响, 以期为水资源短缺地区和季节性干旱地区桢楠幼树的科学管理提供一定的理论依据。
1 材料和方法
1.1 试验地概况
试验地设在四川农业大学教学科研基地, 地理位置为103.86° E, 30.71° N。属亚热带湿润气候区, 全年无霜期282天, 四季分明, 气候温和, 雨量充沛, 多年平均降水量896.1 mm, 但季节分配很不均匀。雨季主要集中在7至9月, 年平均相对湿度达84%, 多年年平均气温16.4 ℃, 平均年日照时间1β104.5 h。试验期间, 为避免降雨对盆内土壤含水量造成影响, 试验在透明且四周通风透气的人工遮雨棚下进行。
1.2 试验材料及苗木培育
1)供试苗木: 桢楠幼树, 树龄2.3年, 平均株高(57 ± 7) cm, 平均地径(0.9 ± 0.1) cm。
2)栽植容器: 聚乙烯塑料盆(口径29 cm, 底径18 cm, 高30 cm)。
3)栽植土壤: 农耕土(沙壤), 其主要化学性质为: pH值7.85, 有机质24.38 g·kg-1, 全氮0.66 g·kg-1, 有效磷128.63 mg·kg-1, 全钾5.81 g·kg-1, 速效钾15.27 mg·kg-1。每盆干土质量约为11.7 kg。
于2015年2月选取同一种源、生长健壮、形态相近的一年生桢楠幼树, 移栽至盆中, 每盆1株, 浇适量水, 进行常规田间管理, 保持土壤含水量在28%左右(约为田间持水量的80%), 培育1.3年, 让其冠幅和根系充分生长。
1.3 试验设计
试验分以下两个阶段:
第一阶段: 干旱梯度设置。参照国家气象中心制定的国家标准《气象干旱等级标准》(GB/T 20481- 2006), 并依据Hisao等(1973), Misra和Tyler (1999)的方法设置4个土壤含水量水平: 80%田间持水量(FC)、50% FC、30% FC和15% FC, 分别相当于体积含水量28.8%、18.0%、10.8%和5.4% (经测定, 本试验用土的饱和体积含水量约为36%)。干旱处理于2016年4月开始进行。干旱前对全部拟处理植株浇水至土壤田间持水量, 然后使其自然干旱, 每天用HH2土壤水分测定仪(ML2x, DELTA-T, Cambridge, UK)测定各盆土的体积含水量, 采用称量法并结合HH2调整土壤含水量至各个处理设定值, 每天18:00称量各盆总质量并测定土壤含水量, 然后补充水分, 使土壤含水量维持在设定的水平上(表1)。
表1 不同干旱水平下容器内土壤体积含水量及盆栽总质量
Table 1
| 土壤含水量(处理水平) Soil water content % FC (The level of drought) | 体积含水量 Soil volumetric water content (%) | 盆栽总质量(平均值±标准偏差) Pot’s mass (mean ± SD) (g) |
|---|---|---|
| 80% FC (水分充足 Sufficient water) | 28.8 | 13β696 ± 97 |
| 50% FC (轻度干旱 Mild drought) | 18.0 | 12β979 ± 112 |
| 30% FC (中度干旱 Moderate drought) | 10.8 | 12β357 ± 106 |
| 15%FC (重度干旱 Severe drought) | 5.4 | 12β070 ± 84 |
FC, field water holding capacity.
FC, 田间持水量。
待设定的土壤含水量梯度形成一周后(5月), 采集各植株新梢上相同部位的功能叶, 以测定活性氧代谢与渗透调节相关生理指标。同时对植株进行施氮处理。
第二阶段: 施氮设计。维持稳定的干旱胁迫水平并进行施氮。刘祖琪和张石诚(1994)以90 kg N·hm-2作为一年生苗木的全年施肥标准, 本试验中二年生桢楠幼树以此标准的3倍施入。由于供试栽植盆盆口面积约为0.049 m2, 以年施氮量约1.35 g·pot-1作为中氮(MN)水平, 年施氮量2.70 g·pot-1作为高氮(HN)水平。氮源为含氮量46.7%的尿素, 按设定的施氮量平均分成4份, 于干旱梯度形成后第7、14、21和28天分别施入, 同时设置不施氮(N0)的处理(表2), 共设12个处理, 每个处理5个重复, 共60盆。待最后一次施肥完成后30天(使植株充分吸收氮素并用于其生理过程后)再次测定植株各项生理指标。
表2 干旱胁迫与对应的施氮水平
Table 2
| 试验代号 Test code | 土壤水分含量 Soil moisture content (% FC) | 氮水平 N level (g·pot-1) |
|---|---|---|
| 15N0 | 15 | 0 |
| 15MN | 15 | 1.35 |
| 15HN | 15 | 2.70 |
| 30N0 | 30 | 0 |
| 30MN | 30 | 1.35 |
| 30HN | 30 | 2.70 |
| 50N0 | 50 | 0 |
| 50MN | 50 | 1.35 |
| 50HN | 50 | 2.70 |
| 80N0 | 80 | 0 |
| 80MN | 80 | 1.35 |
| 80HN | 80 | 2.70 |
FC, field water holding capacity.
FC, 田间持水量。
1.4 测定指标及测定方法
渗透调节物质: 游离脯氨酸(Pro)含量采用磺基水杨酸提取, 酸性茚三酮显色法测定(李合生等, 2000); 可溶性糖(SS)含量采用蒽酮比色法(李合生等, 2000); 可溶性蛋白(SP)含量采用考马斯亮蓝G-250染色法(熊庆娥, 2003)。
活性氧和抗氧化代谢系统: 过氧化氢(H2O2)含量测定参考刘俊等(2000)的方法; 超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)采用同一提取液测定。提取液制备方法为: 取叶片0.1 g, 置预冷的研钵中, 加入8 mL pH值为7.0的磷酸缓冲液研磨成匀浆, 然后2-4 ℃ 15β000 r·min-1离心15 min, 上清液即为酶提取液。SOD活性测定采用硝基四氮唑蓝(NBT)还原法(Giannopolitis & Ries, 1977; 高俊凤, 2006); POD活性测定采用愈创木酚法(李合生等, 2000; 熊庆娥, 2003)。CAT活性测定采用紫外分光光度法(Aebi, 1984; 李合生等, 2000)。
膜脂过氧化产物: 丙二醛(MDA)含量采用硫代巴比妥酸(TBA)加热比色法测定(熊庆娥, 2003)。
1.5 数据处理与分析
采用SPSS 19.0统计分析软件对试验数据进行单因素方差分析, 以双因素方差分析总体评估水分处理、施氮主效应及二者交互效应, 邓肯新复极差法(SSR法)进行各处理间差异显著性检验, 并进行多重比较(显著性水平α = 0.05)。用Microsoft Excel 2010制作图表。
2 结果和分析
2.1 干旱胁迫下桢楠幼树叶片渗透调节的变化及施氮的影响
2.1.1 Pro含量
图1
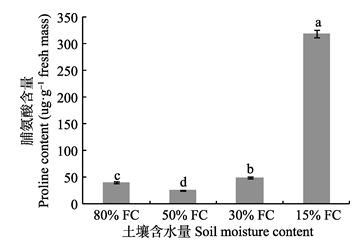
图1
干旱梯度形成后7天桢楠幼树脯氨酸的含量(平均值±标准偏差)。不同小写字母表示不同含水量之间差异显著(p < 0.05)。FC, 田间持水量。
Fig. 1
Proline content in Phoebe zhennan seedlings seven days after the formation of drought gradient (mean ± SD). Different lowercase letters indicate significant differences between treatments (p < 0.05). FC, field water holding capacity.
表3 施氮结束后30天桢楠幼树脯氨酸的含量(平均值±标准偏差)
Table 3
| 处理 Treatment | 土壤含水量 Soil moisture content | |||||
|---|---|---|---|---|---|---|
| 80% FC | 50% FC | 30% FC | 15% FC | 平均值 Mean | ||
| 氮水平 N level | N0 | 3.35 ± 0.11Cb | 2.23 ± 0.25Cc | 2.60 ± 0.03Cbc | 8.33 ± 0.27Ca | 4.13 ± 2.57C |
| MN | 5.77 ± 0.55Bc | 5.97 ± 0.16Bc | 12.93 ± 0.33Bb | 18.48 ± 0.18Ba | 10.79 ± 5.54B | |
| HN | 10.88 ± 0.09Ac | 7.43 ± 0.20Ad | 26.90 ± 1.85Ab | 37.74 ± 0.17Aa | 20.74 ± 12.83A | |
| 平均值 Average value | 6.67 ± 3.34c | 5.21 ± 2.33d | 14.14 ± 10.60b | 21.52 ± 12.94a | ||
| F(SW×SN) | 332.84** | |||||
| F(SW) | 11β481.98** | |||||
| F(SN) | 2β442.89** | |||||
N0, MN, HN, N input 0, 1.35, 2.70 g·pot-1. In the same column of the table, different capital letters indicate significant difference at the probability level of α = 0.05 among N treatments; in the same line of the table, different lowercase letters indicate significant difference at the level of α = 0.05 among drought treatments. The main effects of soil moisture, N input and their interaction were denoted by F(SW), F(SN) and F(SW×SN), respectively. The symbol ** shows significance at 0.01 probability level. FC, field water holding capacity.
N0、MN、HN, 施氮量分别为0、1.35、2.70 g·pot-1。同一列(即在同一土壤含水量下不同施氮水平间)不同大写字母表示在α = 0.05水平上差异显著; 同一行(即在同一施氮水平下不同土壤含水量间)不同的小写字母表示在α = 0.05水平上差异显著; F(SW)、F(SN)和F(SW×SN)分别代表经两因素方差分析所得水分处理主效应、施氮主效应及两者交互效应的F值。**表示显著性达到0.01显著水平。FC, 田间持水量。
2.1.2 SS含量由图2可知, 干旱梯度形成7天, 随着干旱胁迫强度增大, 桢楠幼树叶片SS含量显著增加。表明桢楠幼树为抵御逐渐增强的干旱胁迫, 会产生更多的SS参与到渗透调节中。
图2
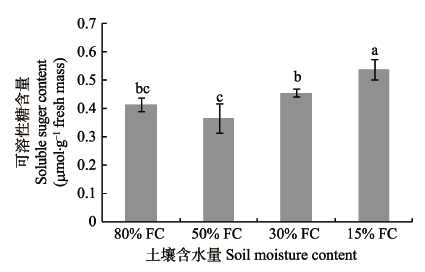
图2
干旱梯度形成后7天桢楠幼树可溶性糖的含量(平均值±标准偏差)。不同小写字母表示不同含水量之间差异显著(p < 0.05)。FC, 田间持水量。
Fig. 2
Soluble sugar content in Phoebe zhennan seedlings seven days after the formation of drought gradient (mean ± SD). Different lowercase letters indicate significant differences between treatments (p < 0.05). FC, field water holding capacity.
表4则显示, 水分充足或轻度干旱状态下施氮, SS含量均表现为MN > HN > N0; 中度干旱下施氮, 表现为N0 > HN > MN; 重度干旱下施氮, 表现为HN > N0 > MN。表明在水分充足或轻度干旱下, 适度的施氮可以显著提高桢楠幼树叶片SS含量, 使其渗透调节能力增强, 过量则作用相反; 处于中度和重度干旱胁迫时, 氮的添加不利于SS的合成。同时方差分析表明, 干旱胁迫与施氮对SS含量具有显著的互作效应。
表4 施氮结束后30天桢楠幼树可溶性糖的含量(平均值±标准偏差)
Table 4
| 处理 Treatment | 土壤含水量 Soil moisture content | |||||
|---|---|---|---|---|---|---|
| 80% FC | 50% FC | 30% FC | 15% FC | 平均值 Mean | ||
| 氮水平 N level | N0 | 0.516 ± 0.007Bb | 0.536 ± 0.009Bb | 0.605 ± 0.008Aa | 0.612 ± 0.010ABa | 0.568 ± 0.045A |
| MN | 0.548 ± 0.005Ab | 0.564 ± 0.011Ab | 0.569 ± 0.009Bb | 0.595 ± 0.013Ba | 0.569 ± 0.020A | |
| HN | 0.523 ± 0.009ABc | 0.548 ±0.010 ABc | 0.574 ± 0.008Bb | 0.633 ± 0.036Aa | 0.569 ± 0.046A | |
| 平均值 Mean | 0.529 ± 0.015d | 0.549 ± 0.015c | 0.583 ± 0.018b | 0.614 ± 0.026a | ||
| F(SW×SN) | 6.560** | |||||
| F(SW) | 67.479** | |||||
| F(SN) | 0.062 | |||||
Notes see
表注同
图3
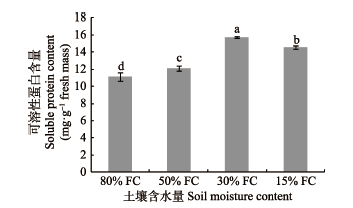
图3
干旱梯度形成后7天可桢楠幼树可溶性蛋白的含量(平均值±标准偏差)。不同小写字母表示不同含水量之间差异显著(p < 0.05)。FC, 田间持水量。
Fig. 3
Soluble protein content in Phoebe zhennan seedlings seven days after the formation of drought gradient (mean ± SD). Different lowercase letters indicate significant differences between treatments (p < 0.05). FC, field water holding capacity.
表5 施氮结束后30天桢楠幼树可溶蛋白的含量(平均值±标准偏差)
Table 5
| 处理 Treatment | 土壤含水量 Soil moisture content | |||||
|---|---|---|---|---|---|---|
| 80% FC | 50% FC | 30% FC | 15% FC | 平均值 Mean | ||
| 氮水平 N level | N0 | 7.57 ± 0.06Aa | 6.86 ± 0.13Ab | 4.61 ± 0.16Cd | 5.53 ± 0.10Cc | 6.14 ± 1.20B |
| MN | 6.65 ± 0.05Bb | 5.84 ± 0.13Bc | 5.34 ± 0.48Bd | 7.16 ± 0.06Aa | 6.25 ± 0.77B | |
| HN | 5.79 ± 0.03Cc | 6.16 ± 0.38Bb | 7.72 ± 0.14Aa | 6.37 ± 0.21Bb | 6.51 ± 0.79A | |
| 平均值 Mean | 6.67 ± 0.77a | 6.28 ± 0.50b | 5.89 ± 1.43b | 6.35 ± 0.71c | ||
| F(SW×SN) | 99.28** | |||||
| F(SW) | 21.61** | |||||
| F(SN) | 10.15** | |||||
Notes see
表注同
2.2 干旱胁迫下桢楠幼树叶片活性氧代谢的变化及施氮的影响
图4
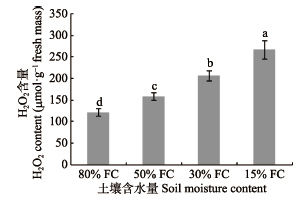
图4
干旱梯度形成后7天桢楠幼树过氧化氢(H2O2)的含量(平均值±标准偏差)。不同小写字母表示不同含水量之间差异显著(p < 0.05)。FC, 田间持水量。
Fig. 4
H2O2 content in Phoebe zhennan seedlings seven days after the formation of drought gradient (mean ± SD). Different lowercase letters indicate significant differences between treatments (p < 0.05). FC, field water holding capacity.
表6 施氮结束后30天桢楠幼树过氧化氢的含量(平均值±标准偏差)
Table 6
| 处理 Treatment | 土壤含水量 Soil moisture content | |||||
|---|---|---|---|---|---|---|
| 80% FC | 50% FC | 30% FC | 15% FC | 平均值 Mean | ||
| 氮水平 N level | N0 | 127.49 ± 6.40Ab | 141.71 ± 2.68Aa | 132.09 ± 2.59Ab | 123.57 ± 7.03Ab | 131.21 ± 8.30C |
| MN | 112.22 ± 10.23Bb | 130.13 ± 3.08Ba | 110.92 ± 3.12Cb | 101.57 ± 6.07Ac | 113.71 ± 12.07B | |
| HN | 116.72 ± 0.08Bab | 115.01 ± 1.02Cb | 122.61 ± 0.96Bab | 125.21 ± 3.07Ba | 119.89 ± 4.60A | |
| 平均值 Mean | 118.81 ± 9.10bc | 128.95 ± 11.79a | 121.88 ± 9.42b | 116.78 ± 12.43c | ||
| F(SW×SN) | 212.35** | |||||
| F(SW) | 255.08** | |||||
| F(SN) | 945.69** | |||||
Notes see
表注同
2.2.2 抗氧化酶活性由图5可知, 干旱梯度形成7天时, 其叶片的抗氧化酶(SOD和CAT)活性随着干旱胁迫强度的增大而显著升高, 而POD活性表现为先升后降。施氮结束后第30天时(表7), 在相同施氮水平下, 3种酶活性随着土壤含水量的降低, 整体呈现出先升高后降低的趋势, 可见随着干旱时间的延长和干旱胁迫强度的增大, 抗氧化酶的活性降低, 导致酶促清除系统对ROS的清除能力下降或丧失。由表7还可见, 在相同的土壤含水量下, 施氮对3种酶活性有显著的影响。其中, 在中度和重度干旱胁迫下, 桢楠幼树叶片SOD活性均表现为施氮处理显著高于不施氮处理, 而在正常水分与轻度胁迫下, 各施氮处理之间的差异不明显; POD活性总体上表现为MN < N0, 而HN的缓解效应总体上减弱; CAT活性在水分充足和轻度胁迫下表现为N0 > HN > MN, 而在中度和重度胁迫下则为HN > MN ≈ N0, 表明高氮处理对重度胁迫的缓解作用不明显。干旱胁迫与施氮对SOD、CAT和POD活性的影响表现出显著的互作。
图5
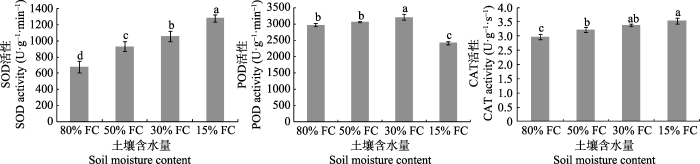
图5
干旱梯度形成后7天桢楠幼树抗氧化酶的活性(平均值±标准偏差)。不同小写字母表示不同含水量之间差异显著(p < 0.05)。FC, 田间持水量。CAT, 过氧化氢酶; POD, 过氧化物酶; SOD, 超氧化物歧化酶。
Fig. 5
Antioxidant enzyme activity in Phoebe zhennan seedlings seven days after the formation of drought gradient (mean ± SD). Different lowercase letters indicate significant differences between treatments (p < 0.05). FC, field water holding capacity. CAT, catalase; POD, peroxidase; SOD, superoxide dismutase.
表7 施氮结束后30天桢楠幼树抗氧化酶的活性(平均值±标准偏差)
Table 7
| 指标 Index | 处理 Treatment | 土壤含水量 Soil moisture content | 平均值 Mean | F | ||||
|---|---|---|---|---|---|---|---|---|
| 80% FC | 50% FC | 30% FC | 15% FC | |||||
| SOD活性 SOD activity (U·g-1·min-1) | 氮水平 N level | N0 | 1β251.53 ± 2.25Ab | 1β337.66 ± 24.89Aa | 1β041.02 ± 53.00Cc | 897.28 ± 42.24Bd | 1β131.87 ± 183.48B | F(SW×SN) = 36.74** F(SW) = 161.34** F(SN) = 58.74** |
| MN | 1β284.91 ± 1.15Aa | 1β306.73 ± 6.73ABa | 1β143.10 ± 33.87Bb | 1β136.76 ± 16.73Ab | 1β217.87 ± 83.47C | |||
| HN | 1β258.43 ± 4.21Ab | 1β278.82 ± 18.54Bb | 1β336.30 ± 37.39Aa | 1β113.63 ± 17.59Ac | 1β246.80 ± 87.83A | |||
| 平均值 Mean | 1β264.96 ± 15.46b | 1β307.74 ± 30.03a | 1β173.47 ± 134.93c | 1β049.22 ± 116.96d | ||||
| POD活性 POD activity (U·g-1·min-1) | 氮水平 N level | N0 | 2β153.65 ± 1.49Bab | 2β043.13 ± 27.43Ab | 2β200.82 ± 39.94Ba | 2β073.00 ± 61.27Bb | 2β117.65 ± 73.54B | F(SW×SN) = 36.19** F(SW) = 24.11** F(SN) = 123.95** |
| MN | 1β888.18 ± 39.09Ca | 1β960.08 ± 30.45Aa | 1β749.85 ± 141.03Cb | 1β673.58 ± 17.14Cb | 1β817.92 ± 133.86C | |||
| HN | 2β347.99 ± 134.53Ab | 1β720.76 ± 73.13Bc | 2β545.87 ± 62.54Aa | 2β412.04 ± 48.07Ab | 2β256.67 ± 339.69A | |||
| 平均值 Mean | 2β129.94 ± 211.82a | 1β907.99 ± 150.89c | 2β165.51 ± 354.77a | 2β052.87 ± 322.59b | ||||
| CAT活性 CAT activity (U·g-1·s-1) | 氮水平 N level | N0 | 2.90 ± 0.06Ab | 3.15 ± 0.05Aa | 3.10 ± 0.05ABa | 2.60 ± 0.04Bc | 2.93 ± 0.23A | F(SW×SN) = 8.39** F(SW) = 28.39** F(SN) = 21.69** |
| MN | 2.52 ± 0.15Bc | 2.74 ± 0.09Cb | 3.02 ± 0.02Ba | 2.75 ± 0.09Bb | 2.75 ± 0.20B | |||
| HN | 2.74 ± 0.11Ad | 2.94 ± 0.10Bcd | 3.26 ± 0.09Aa | 3.04 ± 0.10Abc | 3.01 ± 0.21A | |||
| 平均值 Mean | 2.74 ± 0.21c | 2.94 ± 0.19b | 3.12 ± 0.12a | 2.79 ± 0.21c | ||||
CAT, catalase; POD, peroxidase; SOD, superoxide dismutase. Notes see
CAT, 过氧化氢酶; POD, 过氧化物酶; SOD, 超氧化物歧化酶。表注同
2.3 干旱胁迫下桢楠幼树叶片细胞膜脂过氧化和膜透性的变化及施氮的影响
2.3.1 MDA含量由图6可知, 干旱胁迫加重了桢楠幼树叶片细胞生物膜脂过氧化, 从而产生更多的MDA。而施氮后30天时(表8), 其MDA含量随着土壤含水量的降低, 整体呈现出先上升后降低的趋势, 表明随着干旱时间的延长和干旱胁迫的增强, 植物可能会采取某种机制降低其体内MDA的含量。且由表8可知, 在相同的土壤含水量下, 除重度干旱外, 桢楠幼树叶片MDA含量均表现为施氮组低于不施氮组。其中水分充足时, HN处理下的叶片MDA含量低于其他施氮水平; 轻度和中度干旱胁迫水平下, MN的MDA含量最低。而在重度干旱水平下, 其MDA含量则表现为HN > MN > N0。这表明, 除重度干旱胁迫外, 适量施氮可以显著减轻桢楠幼树膜脂过氧化程度, 总体上以MN效果最佳。分析表明, 干旱胁迫与施氮对MDA含量具有极显著的交互效应。
图6
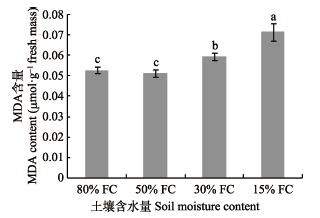
图6
干旱梯度形成后7天桢楠幼树丙二醛(MDA)的含量(平均值±标准偏差)。不同小写字母表示不同含水量之间差异显著(p < 0.05)。FC, 田间持水量。
Fig. 6
Malondialdehyde (MDA) content in Phoebe zhennan seedlings seven days after the formation of drought gradient (mean ± SD). Different lowercase letters indicate significant differences between treatments (p < 0.05). FC, field water holding capacity.
表8 施氮结束后30天桢楠幼树丙二醛的含量(平均值±标准偏差)
Table 8
| 处理 Treatment | 土壤含水量 Soil moisture content | |||||
|---|---|---|---|---|---|---|
| 80% FC | 50% FC | 30% FC | 15% FC | 平均值 Mean | ||
| 氮水平 N level | N0 | 0.070β2 ± 0.000β5Ac | 0.074β8 ± 0.001β5Ab | 0.078β3 ± 0.001β8Aa | 0.067β0 ± 0.001β8Bd | 0.072β6 ± 0.004β7A |
| MN | 0.067β9 ± 0.002β3ABb | 0.069β8 ± 0.000β8Bb | 0.073β0 ± 0.001β2Ba | 0.069β9 ± 0.000β6Ab | 0.070β2 ± 0.002β2B | |
| HN | 0.066β8 ± 0.001β3Bc | 0.071β7 ± 0.001β3Bb | 0.077β0 ± 0.002β5Aa | 0.071β0 ± 0.001β8Ab | 0.071β6 ± 0.004β1A | |
| 平均值 Mean | 0.068β3 ± 0.002β0c | 0.072β1 ± 0.002β4b | 0.076β1 ± 0.002β9a | 0.069β3 ± 0.002β3c | ||
| F(SW×SN) | 6.133β2** | |||||
| F(SW) | 43.294β3** | |||||
| F(SN) | 7.046β1** | |||||
Notes see
表注同
2.3.2 REC
由图7可知, 干旱梯度形成7天时, 桢楠幼树REC随着干旱胁迫强度的增大, 显著下降后上升, 当处于重度干旱时显著高于水分充足时, 表明在干旱胁迫强度较弱时, 桢楠幼树叶片细胞膜受害不严重, 膜透性未发生显著改变; 当干旱胁迫增大到重度干旱时, 细胞膜透性迅速增大, 稳定性降低, 细胞膜严重受损, 大量离子外渗, 因而其REC表现出升高的趋势。而施氮后30天(表9), 同一氮水平下其REC随着土壤含水量的降低, 大体呈现出先上升后降的趋势。而在相同的土壤含水量状态下, 其REC表现为施氮组高于不施氮组(重度干旱胁迫除外), 表明通过施N能够促进桢楠幼树细胞产生更多的电解质。此外, 干旱胁迫与施氮对相对电导率的影响具有显著的交互效应。
图7
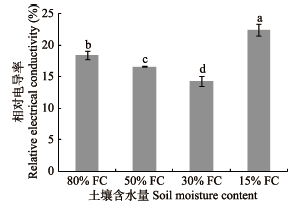
图7
干旱梯度形成后7天桢楠幼树叶片的相对电导率(平均值±标准偏差)。不同小写字母表示不同含水量之间差异显著(p < 0.05)。FC, 田间持水量。
Fig. 7
Relative electrical conductivity in Phoebe zhennan seedlings seven days after the formation of drought gradient (mean ± SD). Different lowercase letters indicate significant differences between treatments (p < 0.05). FC, field water holding capacity.
表9 施氮结束后30天桢楠幼树叶片的相对电导率(平均值±标准偏差)
Table 9
| 处理 Treatment | 土壤含水量 Soil moisture content | |||||
|---|---|---|---|---|---|---|
| 80% FC | 50% FC | 30% FC | 15% FC | 平均值 Mean | ||
| 氮水平 N level | N0 | 31.67 ± 0.31Bb | 37.46 ± 0.58Ba | 33.24 ± 0.92Bb | 32.73 ± 2.04Ab | 33.77 ± 2.50B |
| MN | 39.52 ± 3.95Ab | 45.54 ± 1.56Aa | 35.56 ± 0.62Bc | 32.57 ± 2.75Ac | 38.30 ± 5.52A | |
| HN | 41.54 ± 1.64Aa | 37.30 ± 0.61Bb | 41.28 ± 0.72Aa | 33.01 ± 0.04Ac | 38.28 ± 3.72A | |
| 平均值 Mean | 37.58 ± 5.00b | 40.10 ± 4.18a | 36.69 ± 3.64b | 32.77 ± 1.72c | ||
| F(SW×SN) | 13.48** | |||||
| F(SW) | 28.52** | |||||
| F(SN) | 27.93** | |||||
Notes see
表注同
3 讨论和结论
3.1 施氮对干旱胁迫下桢楠幼树叶片渗透调节的影响
Hasio (1973)认为, 渗透调节是植物忍耐和抵御干旱逆境的一种重要适应机制。植物通过渗透调节, 保持膨压, 有利于保持细胞各种生理过程的正常进行(Taylor, 1996)。已有研究发现, 当植物遭受干旱胁迫时, 在细胞中合成的Pro、SS和甜菜碱等有机溶质起着重要的生理作用, 不仅可以调节细胞质的渗透势, 对酶、蛋白质、生物膜也能起保护作用(Hanson et al., 1979; Patakas et al., 2002; Caprioli et al., 2004)。还有研究表明, 在干旱条件下植物的氮代谢会发生变化, 这是Pro大量积累的原因(Hason, 1980; Dashek & Erickson, 1981), 本试验在一定程度上证实了这一结论。不过, 试验中桢楠幼树在严重缺水时虽然积累了大量Pro, 但此时细胞膜系统已经受到了一定程度的破坏, 渗透调节能力可能较细胞膜完整时的情况大为降低。
本试验中, 随着干旱胁迫的加重, 桢楠幼树叶片SS含量逐渐上升, 这与Kuhns和Gjerstad (1988)、李永洁等(2014)的研究结果相似, 可能是淀粉酶活性的升高加剧了淀粉水解(陈立松和刘星辉, 2001)的结果, 而糖浓度的增加可提高细胞渗透势, 降低细胞水势, 减少干旱下的水分损失(杨德光等, 2015)。同时, 本研究中施氮处理整体增加了桢楠幼树叶片的可溶性糖积累。生产实践中, 在一定程度的干旱胁迫范围内, 通过适度施氮可以促使桢楠幼树产生更多的可溶性糖, 从而达到提高植株渗透调节能力的目的。
有研究表明, 干旱胁迫下植物蛋白质合成受阻(Hulbert et al., 1988)。然而, Heikkala等(1984)在研究玉米(Zea mays)时发现, 干旱胁迫下植物蛋白质含量增加, 甚至可能合成一些新的蛋白质。本试验中,桢楠幼树叶片SP含量随干旱胁迫强度的增加表现为先升高后降低的趋势, 这与刘珊等(2016)和李洁(2015)的研究结果相近。可能在干旱胁迫初期, 植物刚刚萎蔫时, 还存在蛋白质(如酶和胁迫诱导蛋白)的合成(颜华等, 2002), 致使SP含量升高; 此外, 也可能在干旱胁迫下, 植物细胞内合成酶活性下降或水解酶活性增强, 促使结构蛋白转变成可溶性蛋白, 造成SP含量升高(潘瑞炽, 2012)。但随着水分胁迫的
加剧, 植物脱水严重, 蛋白质的合成基本停滞, 可溶性蛋白也开始分解(吴旭红和郑桂萍, 1995), 此时SP含量降低。另外, 桢楠幼树处于轻微干旱胁迫条件时, 施氮可使SP含量显著减少, 但在中度和重度干旱胁迫下则相反, 可能随着胁迫时间的延长和胁迫强度的增加, 桢楠幼树已经遭到一定程度的损伤, 蛋白质的合成由分解过程所代替, 此时其抗旱性更弱, 而通过施氮可在较大程度上增加其体内SP含量, 提高其抗旱性。
3.2 施氮对干旱胁迫下桢楠幼树叶片活性氧代谢和抗氧化酶活性的影响
在桢楠幼树受到干旱胁迫时, 其体内SOD和CAT活性明显增强, 这是植株为减轻干旱胁迫伤害所做的应对, 但POD活性变化不显著, 表明桢楠幼树POD所发挥的抗氧化作用相对其他两种酶更小。当干旱胁迫状态维持一段时间后, POD活性相比施氮前变化更加明显, 且与H2O2含量的变化规律更为相似, 这可能主要是由于随着胁迫时间延长, POD参与到抗氧化胁迫中。Zhang和Kirkham (1994)也曾报道, POD在逆境或衰老后期更为活跃, 可能是植物衰老到一定阶段的产物。
随着干旱胁迫的增强, 桢楠幼树叶片细胞中活性氧的产生逐渐增多, 其膜脂过氧化也会更加严重。当胁迫强度过强时, 抗氧化酶的活性会降低, 可以推测此时桢楠幼树机体已经遭受了很大程度的破坏, ROS的酶促清除系统等抵御干旱胁迫的适应调控机制已逐渐减弱。这与Fridovich (1975)提出的生物自由基伤害学说相一致, 认为当胁迫强度增大到一定程度时, 细胞膜系统会受到严重破坏, 植物机体也会受到很大程度的损伤。
3.3 施氮对干旱胁迫下桢楠幼树叶片细胞膜脂过氧化和膜透性的影响
MDA是膜脂过氧化的主要产物, 对膜和细胞中的许多生物功能分子如蛋白质、核酸和酶等均有很强的破坏作用, 并破坏生物膜的结构和功能, 其含量高低和细胞膜透性变化是反映细胞膜脂过氧化作用强弱和质膜破坏程度的重要指标(陈少裕, 1991; Delhaize & Ryan, 1995)。本研究中, 随着干旱胁迫强度的增大, 桢楠叶片MDA含量升高, 但胁迫强度过大时其MDA含量降低, 这与李娟等(2015)、李娜(2014)的研究结果基本一致。研究还发现, 干旱胁迫初期, 轻度干旱胁迫中的桢楠幼树MDA含量显著低于水分充足的桢楠幼树, 这可能与其具有较高的SOD活性有关(左应梅等, 2016)。同时由试验结果可见, 在受到轻度和中度干旱胁迫时, 中量施氮更有利于降低膜脂过氧化的程度, 在重度干旱胁迫下施氮反而会使膜脂过氧化的程度加重, 这可能与过量施氮使土壤溶液盐浓度过高, 土壤水势降低, 导致植物吸水困难, 产生渗透胁迫, 使其遭受盐害有关(李合生, 2012)。
一般认为, 植物在受到干旱胁迫时, 其细胞膜透性的增加会导致组织电解质的外渗(Kocheva et al., 2014)。本研究发现, 在干旱梯度刚形成时, 桢楠幼树叶片相对电导率会随着干旱胁迫强度的增大呈现出先降低后升高的趋势, 这与以往的研究结果相似(孙明等, 2010; 代英超等, 2015)。但也有研究表明, 随着干旱胁迫的增强, 叶片的相对电导率逐渐升高(吕娥娥等, 2016; 杨传宝等, 2016), 这可能与供试植物不同及植物所遭受的干旱胁迫强度有关。另外, 在相同的干旱胁迫强度下, 桢楠叶片电解质相对电导率随着氮的施入而升高, 可能施肥提供了更多的氮源, 使桢楠幼树能够利用更多的氮, 参与到细胞赖以生存的结构构建或功能成分的合成中, 如嘌呤、碱基、酶、维生素、激素、核酸、糖苷等(周云龙和刘全儒, 2016), 所以施氮组桢楠幼树体内会产生更多的电解质, 以致植物细胞浸提液的电导率增大。
综上所述, 为处于干旱胁迫状态的桢楠幼树提供适量的氮, 可提高其渗透调节能力, 同时也可降低其活性氧的产生, 从而在一定程度上减轻干旱胁迫的伤害。但过量的氮会造成相反的结果, 因此对受干旱胁迫的桢楠幼树施氮需控制在适当水平, 切忌过量。对二年生桢楠幼树而言, 每株施氮1.35 g (折合尿素2.89 g)对轻、中度干旱胁迫具有一定的缓解作用, 但施氮达到2.7 g·株-1 (折合尿素5.78 g)以上时会对植株造成不利影响。

参考文献
Catalase in vitro.
DOI:10.1016/S0076-6879(84)05016-3 URL [本文引用: 1]
Leaf hydraulics and drought stress: Response, recovery and survivorship in four woody temperate plant species
Regional vegetation die-off in response to global-change-type drought
DOI:10.1073/pnas.0505734102
URL
PMID:16217022
[本文引用: 1]

Future drought is projected to occur under warmer temperature conditions as climate change progresses, referred to here as globalchange-type drought yet quantitative assessments of the triggers and potential extent of drought-induced vegetation die-off remain pivotal uncertainties in assessing climate-change impacts. Of particular concern is regional-scale mortality of overstory trees, which rapidly alters ecosystem type, associated ecosystem properties, and land surface conditions for decades. Here, we quantify regional-scale vegetation die-off across southwestern North American woodlands in 2002-2003 in response to drought and associated bark beetle infestations. At an intensively studied site within the region, we quantified that after 15 months of depleted soil water content, >90% of the dominant, overstory tree species (Pinus edulis, a$pi\tilde{n}on$) died. The die-off was reflected in changes in a remotely sensed index of vegetation greenness (Normalized Difference Vegetation Index), not only at the intensively studied site but also across the region, extending over 12,000 km2or more; aerial and field surveys confirmed the general extent of the die-off. Notably, the recent drought was warmer than the previous subcontinental drought of the 1950s. The limited, available observations suggest that die-off from the recent drought was more extensive than that from the previous drought, extending into wetter sites within the tree species' distribution. Our results quantify a trigger leading to rapid, drought-induced die-off of overstory woody plants at subcontinental scale and highlight the potential for such die-off to be more severe and extensive for future global-change-type drought under warmer conditions.
Trehalose in desiccated rotifers: A comparison between a bdelloid and a monogonont species
DOI:10.1016/j.cbpb.2004.10.019
URL
PMID:15596399
[本文引用: 1]
In response to drought bdelloid and monogonont rotifers undergo anhydrobiosis and are assumed to synthesize protective chemicals, which are commonly sugars. In contrast to most anhydrobionts, bdelloids have earlier been shown to lack trehalose as protective chemical, and more importantly to lack trehalose synthase (tps) genes. It remains to be assessed if the absence of trehalose is a characteristic common to the entire taxon Rotifera, or if it is limited to bdelloids, or is peculiar to the two bdelloid species investigated so far. In this study, anhydrobiotic adults of a bdelloid species ( Macrotrachela quadricornifera) and resting eggs of a monogonont species ( Brachionus plicatilis) were analysed by thin layer chromatography and gas chromatography to detect the presence of trehalose. No trehalose was detected in the bdelloid, while the anhydrobiotic resting egg of the monogonont rotifer contained about 0.35% trehalose of its dry weight. Although very little, the presence of trehalose in B. plicatilis suggests that the trehalose synthase genes, absent in bdelloid rotifers, are present in non-bdelloid rotifers.
Effects of water stress on cell wall H +-ATPase activity in leaves of
水分胁迫对抗旱性不同的荔枝叶片细胞壁H +-ATPase活性的影响
Injury of membrane lipid peroxidation to plant cell
膜脂过氧化对植物细胞的伤害
Physiological responses of the rare and endangered Ardisia violaceaMyrsinaceae) seedlings to progressive drought stress.
DOI:10.5846/stxb201308302175
URL
[本文引用: 1]
采用盆栽控水法,研究了珍稀濒危植物堇叶紫金牛(Ardisia violacea)在持续干旱条件下的生理响应.随着持续干旱时间的延长,堇叶紫金牛应对持续干旱的阶段可分为适应期、轻度干旱期、中度干旱期和重度干旱 期.在适应期和轻度干旱期,堇叶紫金牛叶片游离脯氨酸和可溶性糖含量稳定在一个较低水平,可溶性蛋白质含量先下降后快速上升,细胞膜系统和抗氧化酶系统能 主动进行生理调节;中度干旱期,丙二醛(MDA)含量和质膜相对透性迅速升高,细胞膜系统受损加剧,游离脯氨酸、可溶性糖含量均急剧增加,对抵御干旱起到 重要的渗透调节作用.在轻度干旱期和中度干旱期,光合色素中叶绿素a和叶绿素b含量显著提高,以抵抗干旱胁迫.重度干旱期,细胞膜系统、抗氧化酶SOD、 游离脯氨酸和可溶性糖含量上升,但MDA略微下降,这时可能达到植物耐受干旱的极限,不再发生膜脂过氧化作用.综上表明,堇叶紫金牛具有较强的耐旱 性,RWC为49.94%是细胞膜系统、抗氧化酶系统和渗透调节物质含量变化的拐点,渗透调节和抗氧化酶系统的主动适应是其耐旱的主要机制.
珍稀濒危植物堇叶紫金牛对持续干旱的生理响应
DOI:10.5846/stxb201308302175
URL
[本文引用: 1]
采用盆栽控水法,研究了珍稀濒危植物堇叶紫金牛(Ardisia violacea)在持续干旱条件下的生理响应.随着持续干旱时间的延长,堇叶紫金牛应对持续干旱的阶段可分为适应期、轻度干旱期、中度干旱期和重度干旱 期.在适应期和轻度干旱期,堇叶紫金牛叶片游离脯氨酸和可溶性糖含量稳定在一个较低水平,可溶性蛋白质含量先下降后快速上升,细胞膜系统和抗氧化酶系统能 主动进行生理调节;中度干旱期,丙二醛(MDA)含量和质膜相对透性迅速升高,细胞膜系统受损加剧,游离脯氨酸、可溶性糖含量均急剧增加,对抵御干旱起到 重要的渗透调节作用.在轻度干旱期和中度干旱期,光合色素中叶绿素a和叶绿素b含量显著提高,以抵抗干旱胁迫.重度干旱期,细胞膜系统、抗氧化酶SOD、 游离脯氨酸和可溶性糖含量上升,但MDA略微下降,这时可能达到植物耐受干旱的极限,不再发生膜脂过氧化作用.综上表明,堇叶紫金牛具有较强的耐旱 性,RWC为49.94%是细胞膜系统、抗氧化酶系统和渗透调节物质含量变化的拐点,渗透调节和抗氧化酶系统的主动适应是其耐旱的主要机制.
Isolation, assay, biosynthesis, metabolism, uptake and cranslocation and function of proline in plant cells and tissues
DOI:10.1007/BF02860578
URL
[本文引用: 1]
The amino acid L-proline has been the subject of intensive research during the past ten to fifteen years. This stems from the observations that it incorporates into peptide linkage thereby serving as a precursor to peptidyl-bound L-hydroxyproline, a constituent of "extensin," and that it accumulates when some plants are exposed to diverse biological and environmental stresses. The contents of selected papers which have been published during the last quarter of a century regarding the isolation, assay, biosynthesis, metabolism, transport and function of L-proline within various plant tissues and their cells are both interpreted and summarized in this review. Occasionally, relevant information from animal and bacterial systems concerning these topics is included. Hydroxyproline-containing proteins are not considered. L-proline was reported to be a constituent of leaves as early as the 1950's. Since then, it and its analogues have been extracted from the organs of a variety of plants. The analogues include: methyl-hydroxylproline; 4-methylene-DL-proline; L-azetidine-2-carboxylic acid; 2,3,cis-3,4-trans-dihydroxy-L-proline; L-pipecolic acid and 4-trans-hydroxyproline. L-proline can be both detected and quantified by colorimetric, combined fluorometric-amino acid analyzer and gas chromatographic procedures. L-proline may be synthesized from L-glutamic acid via the following biosynthetic pathway: L-glutamic acid $@>\gamma \text{-glutamic acid kinase}>>$ γ-glutamyl phosphate $@>\gamma \text{-glutamyl phosphate reductase}>>$ γ-glutamyl semialdehyde $@>\text{spontaneous cyclization}>>$ Δ′-pyrroline-5-carboxylate (P5C) $@>{\rm P}5{\rm C}\ \text{reductase}>>$ L-proline. Proline can also originate from L-arginine and L-ornithine. Biosynthesis from the latter compound proceeds either through the γ-glutamyl semialdehyde and pyrroline-5-carboxylate pathway or alternatively a α-keto-δ-aminovaleric and pyrroline-2-carboxylate pathway. The metabolism of L-proline most likely involves the reverse of the biosynthetic pathway with an initial prolyl dehydrogenase- or prolyl oxidase-mediated conversion of L-proline to Δ′-pyrroline-5-carboxylate. The metabolism of L-proline has been demonstrated to occur in excised tissues and cell free extracts, cell suspension cultures and reproductive structures. Little is known about the mechanism by which L-proline is taken up by cultured plant cells and excised tissues. Once within the plant L-proline can be translocated through the phloem at velocities similar to those for carbon dioxide assimilates. In addition to serving as a substrate for peptidyl-bound hydroxyproline, L-proline may function as an adaptation to diverse biological and environmental stresses, a cryoprotectant, a nitrogen pool, a precursor for chlorophyll synthesis upon relief of stress, a regulator together with L-histidine of fertility and sterility and/or a substrate for respiration. /// Depuis ces quinze dernières années, l'amino-acide L-proline a été l'objet de recherches très intensives parce que l'on a observé que L-proline s'incorpores dans les liaisons peptidiques, et par ce moyen se sers de précurseur à la fraction peptidique L-hydroxyproline, un des constituants de "l'extensine," et s'accumule lorsque les plantes sont soumise à des contraintes environmentales et biologique diverses. Le contenu d'un certain nombre d'articles qui furent publiés ce dernier quart de siècle traite de l'isolement, les essais en laboratoire, la biosynthèse, le métabolisme, le transport et les fonctions de la L-proline dans les cellules et les tissus variés de plante sont interprétés et résumé dans cette revue. Occasionnellement, des informations complémentaires relevant des systèmes animaux et bactériens jugés utiles ont été introduites. Les protéines contenant de l'hydroxyproline n'est pas envisagé ici. Depuis le début des années mille neuf cent cinquante, la L-proline fut reconnue d'être un des constituants dans les feuilles. Depuis lors, L-proline ont été extraite, ainsi que ses analogues, d'organes de diverses sortes de plantes. Ses analogues comportent: la methyle-hydroxyle-proline; la 4-methylene DL-proline; l'acide L-azetidine-2-carboxylique; la 2,3, cis-3, 4-trans-dihydroxy-L-proline; l'acide L-pipécolique et l'hydroxyproline 4-trans. La L-proline peut être détectée et quantifié par la colorimétrie à l'analyse fluorométrique des acides aminés et par la chromatographie en phase gazeuse. La L-proline peut être synthétisée de l'acide L-glutamique par les étapes biosynthétiques suivantes: acide L-glutamique $@>\text{acide}\ \gamma \text{-glutamique kinase}>>$ γ glutamyl phosphate $@>\gamma \text{-glutamyl phosphate r}\acute{{\rm e}}\text{ductase}>>$ γ glutamyl semialdéhyde $@>\text{clyclisation spontan}\acute{{\rm e}}\text{e}>>$ Δ′-pyrroline-5-carboxylate (P5C) $@>{\rm P}5{\rm C}\ \text{r}\acute{{\rm e}}\text{ductase}>>$ L-proline. La proline peut aussi avoir son origine de L-arginine ou de L-ornithine. La biosynthèse de cette dernière a lieu soit par voie du γ glutamyl semi-aldehyde et P5C, soit par celui des réaction biosynthétiques de l'acide γ-keto δ amino valérique et du pyrroline-2-carboxylate. Le métabolisme de la L-proline probablement inclut les étapes inverses de la biosynthèse avec une conversion initiale de la L-proline dans Δ′-pyrroline-5-carboxylate gr09ce à la prolyl déhydrogenase ou la prolyl oxidase. Le métabolisme de la L-proline a été démontré de survenir dans des tissus excisés, des extraits sans cellules, des cultures de suspension cellulaires et des structures reproductives. On ne sait que très peu du mécanisme par lequel la L-proline est incorporée dans les cultures cellulaires des plantes et des tissus excisés. Une fois dans la plante la L-proline peut être transportée dans le phloème à des vélocités semblable de cells des assimilats d'oxyde de carbone. Outre son utilité comme substrat pour la hydroxyproline, bondé avec la peptidyle la L-proline peut fonctionner comme une adaptation à des contraintes biologique et environmentales diverses, un cryoprotecteur, une reserve d'azote, un précurseur de la synthèse chlorophyllienne lorsque la contrainte est levée; un régulateur, avec la L-histidine, de la fertilité et de la sterilité et/ou un substrat respiratoire.
Aluminum toxicity and tolerance in plants
DOI:10.1104/pp.107.2.315
URL
PMID:12228360
[本文引用: 1]
ABSTRACT Aluminum (Al) is the most abundant metal in the earths crust, comprising about 7% of its mass. Since many plant species are sensitive to micromolar concentrations of Al, the potential for soils to be A1 toxic is considerable. Fortu- nately, most of the A1 is bound by ligands or occurs in other nonphytotoxic forms such as aluminosilicates and precip- itates. However, solubilization of this A1 is enhanced by low pH and A1 toxicity is a major factor limiting plant production on acid soils. Soil acidification can develop naturally when basic cations are leached from soils, but it can be accelerated by some farming practices and by acid rain (Kennedy, 1986). Strategies to maintain production on these soils include the application of lime to raise the soil pH and the use of plants that are tolerant of acid soils. Although A1 toxicity has been identified as a problem of acid soils for over 70 years, our knowledge about the primary sites of toxicity and the chain of events that finally affects plant growth remains largely speculative. In this paper we review recent progress that has been made in our understanding of A1 toxicity and the mechanisms of A1 tolerance in plants.
Superoxide dismutase
DOI:10.1146/annurev.bi.44.070175.001051 URL
Superoxide dismutases: I. Occurrence in higher plants
DOI:10.1104/pp.59.2.309
URL
PMID:16659839
[本文引用: 1]
Shoots, roots, and seeds of corn (Zea mays L., cv. Michigan 500), oats (Avena sativa L., cv. Au Sable), and peas (Pisum sativum L., cv. Wando) were analyzed for their superoxide dismutase content using a photochemical assay system consisting of methionine, riboflavin, and p-nitro blue tetrazolium. The enzyme is present in the shoots, roots, and seeds of the three species. On a dry weight basis, shoots contain more enzyme than roots. In seeds, the enzyme is present in both the embryo and the storage tissue. Electrophoresis indicated a total of 10 distinct forms of the enzyme. Corn contained seven of these forms and oats three. Peas contained one of the corn and two of the oat enzymes. Nine of the enzyme activities were eliminated with cyanide treatment suggesting that they may be cupro-zinc enzymes, whereas one was cyanide-resistant and may be a manganese enzyme. Some of the leaf superoxide dismutases were found primarily in mitochondria or chloroplasts. Peroxidases at high concentrations interfere with the assay. In test tube assays of crude extracts from seedlings, the interference was negligible. On gels, however, peroxidases may account for two of the 10 superoxide dismutase forms.
Capacity for proline accumulation during water stress in barley and its implications for breeding for drought resistance
DOI:10.2135/cropsci1979.0011183X001900040015x
URL
[本文引用: 1]
During controlled water stress applied at the three-leaf stage, the barley (Hordeum vulgare L.) cultivars 'Proctor' and 'Excelsior' showed different rates of free proline accumulation and leaf-firing. Both processes were faster in Proctor (a drought-susceptible cultivar) than in Excelsior (a drought-resistant cultivar). The different rates of proline accumulation during stress in Proctor, in Excelsior, and in a wild barley (H. spontaneum C. Koch) could be attributed to differences in the rate of decline of leaf water potential ( psi leaf); at the same psi leaf value, all three genotypes contained similar amounts of free proline. After non-destructive screening of the F2 generation from a Proctor X Excelsior cross for proline-accumulating capacity under controlled water stress conditions, F3 seed from plants selected for high and low proline accumulation was obtained. When F3 palnts were tested for proline accumulation during controlled stress, the selection for proline accumulation was found to have been effective. Reselection among F3 plants and subsequent testing of the F4 generation confirmed this result. Among individual plants of the F3 generation (which was segregating for the proline accumulation trait) there was a positive association between the amount of free proline accumulated during stress and the severity of leaf-firing. Massive proline accumulation is a symptom of severe internal water stress in barley and apparently has no survival value during drought. Although there is heritable variation in the amount of proline accumulated during stress, it is not of practical value in breeding for drought resistance.ADDITIONAL ABSTRACT:During controlled water stress, rates of proline accumulation and leaf firing were greater in Proctor (drought-susceptible) than in Excelsior (drought-resistant) seedlings. The different rates of proline accumulation among the above varieties and Hordeum spontaneum (intermediate resistance) were attributable to differences in the rate of decline of leaf water potential. At a given value of the latter, similar concentrations of free proline were present in the leaves of the three genotypes. F2 plants derived from a Proctor X Excelsior cross were selected for high or low proline accumulation. F3 and subsequent F4 progenies (obtained by F3 reselection) retained the proline accumulation characteristics of the F2 selections. Both high proline concentration and intensity of leaf firing are symptomatic rather than causative of drought susceptibility. Leaf firing would be a more readily measurable selection criterion.ADDITIONAL ABSTRACT:Studies were carried out determine if high proline accumulation was associated with susceptibility or with resistance to water stress. During controlled water stress applied at the 3-leaf stage the drought-susceptible barley cv. Proctor showed higher rates of free proline accumulation and leaf firing [scorch] than did the drought-resistant cv. Excelsior. The different rates of proline accumulation during stress in Proctor, Excelsior and a wild barley were attributed to differences in the rate of decline of leaf water potential; at the same leaf water potential, all 3 genotypes contained similar amounts of free proline. Progeny from a Proctor X Excelsior cross selected for high and low proline accumulation showed a positive association between the amount of free proline accumulated during stress and the severity of leaf firing. Massive proline accumulation is a symptom of severe internal water stress in barley but has apparently no survival value during drought
Interpreting the metabolic response of plants to water stress
Biological adaptation by-plants to drying (dessication) or drought-prone environments is analyzed. The metabolic components of adaptation and the metabolic symptoms of stress injury are reviewed. Breeding programs for selection of drought-resistance in plants are suggested.
Induction of heat shock protein messenger RNA in maize mesocotyls by water stress, abscisci acid and wounding
DOI:10.1104/pp.76.1.270
URL
PMID:16663813
Exposure of the excised growing region of the mesocotyl of young corn seedlings to heat shock stimulated the production of specific heat shock proteins and the intensification of synthesis of two proteins with a molecular weight of approximately 70,000. Water stress and abscisic acid also stimulated synthesis of these 70,000-dalton proteins, and other unique proteins distinct from those induced by heat shock. Growing tissues of intact corn mesocotyls exposed to heat shock, water stress, or abscisic acid accumulated mRNA species homologous to a cloned genomic probe of the 5鈥 end of the 70,000-dalton Drosophila heat shock protein gene. Since cut segments of the mesocotyl under unstressed conditions produced a similar mRNA, we suggest that the hsp 70 gene is activated in corn by a variety of diverse stresses. Production of the mRNA is rapid, but transient, being induced within 3 hours of the imposition of the stress, but declining after reaching a maximum at 9 hours.
Plants response to water stress
DOI:10.1146/annurev.pp.24.060173.002511 URL
Inhibition of protein synthesis in loblolly pine hypocotyls by mannitol-induced water stress
DOI:10.1093/treephys/4.1.19
URL
PMID:14972832
[本文引用: 1]
Abstract The relationship between mannitol-induced water stress and protein synthesis was investigated in hypocotyl slices of loblolly pine (Pinus taeda L.). Mannitol-induced water stress inhibited the incorporation of l-[35S]methionine into protein. As the water potential decreased, incorporation of label into protein decreased in both the soluble- and membrane-protein fractions. There were no significant differences in response to water stress among seed sources from four different geographical regions.
An improvement of the diffusion model for assessment of drought stress in plant tissues
DOI:10.1111/ppl.12074
URL
PMID:23724871
[本文引用: 1]
The article discusses an improvement of a previously developed method for assessment of ion leakage from plant tissues as a gauge of membrane and cell wall performance under stressful environment. It employs conductometric measurements of the ion efflux from leaves and their quantitative interpretation by a theoretical model based on the laws of diffusion. Experimental data are readily fit with the model and results are in accordance with relative water content of dehydrated barley (Hordeum vulgare) seedlings of two distinct cultivars. Some new parameters obtained from fitting are proposed as reliable indicators of the leaf status. They appear to be helpful in further distinguishing the behavior of two separate cellular structures with respect to their electrolyte permeability. It is concluded that the established method based on the kinetics of ion leakage is adequate for evaluation of contrasting genotypes under normal and stress conditions. Furthermore, it could be used as a simple and powerful tool for routine analysis and screening for drought tolerance in crops.
Photosynthate allocation in loblolly pine (Pinus taeda) seedlings as affected by moisture stress
Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants
DOI:10.1046/j.0016-8025.2001.00814.x
URL
PMID:11841670
[本文引用: 1]
Summary Experimental studies on CO 2 assimilation of mesophytic C3 plants in relation to relative water content (RWC) are discussed. Decreasing RWC slows the actual rate of photosynthetic CO 2 assimilation (A) and decreases the potential rate (A pot ). Generally, as RWC falls from c . 100 to c. 75%, the stomatal conductance (g s ) decreases, and with it A. However, there are two general types of relation of A pot to RWC, which are called Type 1 and Type 2. Type 1 has two main phases. As RWC decreases from 100 to c. 75%, A pot is unaffected, but decreasing stomatal conductance (g s ) results in smaller A, and lower CO 2 concentration inside the leaf (C i ) and in the chloroplast (C c ), the latter falling possibly to the compensation point. Down-regulation of electron transport occurs by energy quenching mechanisms, and changes in carbohydrate and nitrogen metabolism are considered acclimatory, caused by low C i and reversible by elevated CO 2 . Below 75% RWC, there is metabolic inhibition of A pot , inhibition of A then being partly (but progressively less) reversible by elevated CO 2 ; g s regulates A progressively less, and C i and CO 2 compensation point, 螕 rise. It is suggested that this is the true stress phase, where the decrease in A pot is caused by decreased ATP synthesis and a consequent decreased synthesis of RuBP. In the Type 2 response, A pot decreases progressively at RWC 100 to 75%, with A being progressively less restored to the unstressed value by elevated CO 2 . Decreased g s leads to a lower C i and C c but they probably do not reach compensation point: g s becomes progressively less important and metabolic limitations more important as RWC falls. The primary effect of low RWC on A pot is most probably caused by limited RuBP synthesis, as a result of decreased ATP synthesis, either through inhibition of Coupling Factor activity or amount due to increased ion concentration. Carbohydrate synthesis and accumulation decrease. Type 2 response is considered equivalent to Type 1 at RWC below c. 75%, with A pot inhibited by limited ATP and RuBP synthesis, respiratory metabolism dominates and C i and 螕 rise. The importance of inhibited ATP synthesis as a primary cause of decreasing A pot is discussed. Factors determining the Type 1 and Type 2 responses are unknown. Electron transport is maintained (but down-regulated) in Types 1 and 2 over a wide range of RWC, and a large reduced/oxidized adenylate ratio results. Metabolic imbalance results in amino acid accumulation and decreased and altered protein synthesis. These conditions profoundly affect cell functions and ultimately cause cell death. Type 1 and 2 responses may reflect differences in g s and in sensitivity of metabolism to decreasing RWC.
Effects of drought stress on soluble proteins of
DOI:10.15889/j.issn.1002-1302.2015.12.038
URL
采用液体培养法,研究了不同浓度聚乙二醇(PEG)模拟干旱处理对青稞幼苗叶片可溶性蛋白含量的影响,并利用SDS-PAGE电泳检测干旱对供试青稞品种叶片可溶性蛋白的变化及其与干旱胁迫的关系.结果表明:在短时间干旱胁迫下,旱地紫青稞、大麻青稞叶片可溶性蛋白含量均随着干旱胁迫增强呈先降低、后升高的趋势;但在长时间干旱胁迫下,2份材料的可溶性蛋白含量随着干旱胁迫增强逐渐降低;对干旱胁迫4d的可溶性蛋白进行SDS-PAGE分析,发现在55 ku处有1个蛋白或1组蛋白为干旱诱导蛋白.
干旱胁迫对青稞幼苗可溶性蛋白的影响
DOI:10.15889/j.issn.1002-1302.2015.12.038
URL
采用液体培养法,研究了不同浓度聚乙二醇(PEG)模拟干旱处理对青稞幼苗叶片可溶性蛋白含量的影响,并利用SDS-PAGE电泳检测干旱对供试青稞品种叶片可溶性蛋白的变化及其与干旱胁迫的关系.结果表明:在短时间干旱胁迫下,旱地紫青稞、大麻青稞叶片可溶性蛋白含量均随着干旱胁迫增强呈先降低、后升高的趋势;但在长时间干旱胁迫下,2份材料的可溶性蛋白含量随着干旱胁迫增强逐渐降低;对干旱胁迫4d的可溶性蛋白进行SDS-PAGE分析,发现在55 ku处有1个蛋白或1组蛋白为干旱诱导蛋白.
Physiological response of two
DOI:10.16213/j.cnki.scjas.2015.03.027
URL
为提高杜鹃的抗旱性,通过对西鹃和毛鹃干旱胁迫过程中叶片含水量、细胞膜透性、叶绿素含量、过氧化物酶(POD)、过氧化氢酶(CAT)活性等生理指标的测定,研究土壤干旱胁迫下杜鹃的生理响应机制并利用隶属函数法综合评价杜鹃的抗旱性。结果表明:西鹃和毛鹃的叶片含水量随着干旱胁迫时间的延长而逐渐降低,细胞膜透性逐渐上升,叶绿素、脯氨酸、可溶性蛋白质随着干旱胁迫时间的增加呈先增加后减小的趋势,过氧化物酶(POD)活性呈上升趋势,过氧化氢酶(CAT)活性呈先增加后减小的趋势;西鹃丙二醛含量上升,而毛鹃丙二醛含量先上升后下降。利用隶属函数法综合评价得出西鹃的抗旱性大于毛鹃。
2种杜鹃对干旱胁迫的生理响应及抗旱性评价
DOI:10.16213/j.cnki.scjas.2015.03.027
URL
为提高杜鹃的抗旱性,通过对西鹃和毛鹃干旱胁迫过程中叶片含水量、细胞膜透性、叶绿素含量、过氧化物酶(POD)、过氧化氢酶(CAT)活性等生理指标的测定,研究土壤干旱胁迫下杜鹃的生理响应机制并利用隶属函数法综合评价杜鹃的抗旱性。结果表明:西鹃和毛鹃的叶片含水量随着干旱胁迫时间的延长而逐渐降低,细胞膜透性逐渐上升,叶绿素、脯氨酸、可溶性蛋白质随着干旱胁迫时间的增加呈先增加后减小的趋势,过氧化物酶(POD)活性呈上升趋势,过氧化氢酶(CAT)活性呈先增加后减小的趋势;西鹃丙二醛含量上升,而毛鹃丙二醛含量先上升后下降。利用隶属函数法综合评价得出西鹃的抗旱性大于毛鹃。
Physiological and Ecological Response of Larix gmelinii Seedlings under Soil Drought Stress and Different Nitrogen Levels
.随着全球气候与环境变化的加剧,地球表面平均温度升高、水资源日趋短缺、土壤有效含水量逐年减少以及大气氮沉降已成为重要的全球性环境问题。土壤氮素和水分是许多陆地生态系统中植物生长的主要限制因子,显著影响植物的分布和生长发育。全球气候导致的干旱导致我国落叶松林北移,加之我国大气氮沉降持续增加,导致森林土壤中可利用氮的含量不断增加。因此,探究干旱与氮素综合作用下落叶松幼苗的生长特性、生理特性以及适应性机制,对改善干旱系统植被条件、了解北方森林生态系统功能及格局的改变具有重要意义,也为落叶松人工林繁育过程中合理调节水肥管理、优化培育措施提供理论参考。本研究以1年生落叶松(Larix)为研究对象,采用大棚盆栽方法人为控制土壤水分和施N控制试验,设置三个水分梯度:CK(正常供水,土壤水分含量占田间持水量的80%-70%,下同)、MS(中度干旱胁迫,60%-50%)、HS(严重干旱胁迫,40%-30%);四种施氮水平:NO(不施氮,Og NH4NO3-N·m-2·a-1)、LN(低氮,5g NH4NO3-N·m-2·a-1、MN(中氮,10g NH4NO3-N·m-2·a-1)、HN(高氮,15gNH4NO3-NN·m-2·a-1),从植株生长、生理指标入手,系统的研究了落叶松幼苗对干旱胁迫和施N的生理生态响应,并揭示了落叶松幼苗的耐旱性机制。研究结果表明: (1)随着干旱胁迫程度的增加,不同氮处理下的落叶松幼苗株高、基径生长量呈下降趋势,并对对落叶松幼苗的生物量积累和生物量分配格局具有明显的影响,干旱胁迫越严重,幼苗叶面积、总生物量的积累越少,地上生物量越来越小,地下生物量积累越来越多,R/S升高,SLA和LAR增逐渐加,并产生极显著影响(P0.001)。在干旱胁迫与施氮的交互作用下,施氮处理可在一定程度上改善落叶松幼苗株高、基径、RGR、叶面积、总生物量,促进生长,但是不同水分条件下的氮素效应不同。正常水分供应条件下,施氮水平的提高增加了幼苗生长指标和生物量的积累。而干旱胁迫条件下,施加了过量的氮后幼苗生长量、生物量积累反而降下降,植株通过降低叶面积、降低叶片分配比例来降低自身的水分消耗状况,并增加茎和根的分配比例,将有限的资源较多的生物量分配给地下部分和茎生物量。 (2)不同干旱胁迫处理下,落叶松幼苗光合作用明显受到抑制,随干旱程度增加在同一光强时幼苗Pn逐渐降低,光饱和点逐渐降低。而不同干旱胁迫条件下适量的施氮能在一定程度上提高落叶松幼苗的光合能力。低氮处理下在严重干旱时对植株光合能力有明显的促进作用,而高氮处理下的促进作用减弱并转向抑制。严重干旱情况下,幼苗Pmax在HN水平下表现出最低值,在LN水平下表现出最大值。此时氮处理水平下的Pn和Ci都表现出相同的变化趋势,Pn的下降是非气孔因素引起的。 不同干旱胁迫处理下,落叶松幼苗茎的呼吸作用也明显受到抑制。严重干旱情况下的高氮处理下幼苗的呼吸值最小,说明过量增加氮肥抑制了植物的生长,幼苗的光合作用低的同时,所能固定的光合产物也随之减少,用以为幼苗提供生命活动所需的能量就少。而正常水分供应条件下的高氮浓度下的幼苗呼吸值,和中度干旱情况下的中氮处理下幼苗的呼吸值明显高于同一供水条件下的其他施氮水平,说明在适当的氮处理水平下,落叶松幼苗的生长代谢速度最快,需要通过呼吸作用为其生命活动提供能量。 干旱胁迫使得落叶松幼苗叶片中的叶绿素a、叶绿素b、总叶绿素以及叶绿素a/b上升。但是同一水分处理下各个氮处理水平之间的变化趋势不同,适量的氮素水平能显著提高落叶松幼苗的叶绿素a、叶绿素b、总叶绿素含量,且变化趋势一致。MS和HS条件下下,HN水平都抑制了光合色素的积累,以HN处理下幼苗的Chla、Chlb和Ch1含量最低。但在MS条件下,MN水平下表现出较高的光合色素水平。而在HS条件下,LN水平下表现出较高的光合色素水平。 (3)落叶松幼苗叶片相对含水量(LRWC)随干旱胁迫程度的增加而降低,同一水分处理下不同氮处理水平下的LRWC表现出来的变化也各不相同。中度干旱胁迫下,LRWC随施氮水平的增高表现出先升高后降低的趋势,在MN水平下幼苗LRWC最高,显著高于其他施氮水平(P0.05)。严重干旱胁迫下,LRWC表现出与中度干旱胁迫下相同的变化趋势,但LRWC在LN水平下表现出最高值,且HN水平下表现出最低值。 (4)幼苗总SS含量均随干旱胁迫程度的增加而降低,且同一水分处理下不同氮处理水平之间的变化趋势也显示为一致的趋势,只是变化幅度不同。在HS条件下,叶片、茎、根SS含量随施氮量的增加表现出先增加后降低的趋势,且LN处理表现出SS含量最高,HN处理表现出叶片含量最低。而随着干旱胁迫的加剧,各个施氮水平的幼苗Pro、MDA含量同对照相比整体呈上升趋势。其中,中度干旱胁迫下,各个施氮水平的幼苗Pro含量、MDA含量同对照相比整体呈小幅度上升趋势,但在MN水平下升上幅度最小;而严重干旱胁迫下,各个施氮水平的幼苗Pro含量、MDA含量同正常水分条件下相比上升幅度较大,并以HN水平下的含量最高,LN水平下的含量最低。
落叶松幼苗对干旱胁迫及氮添加的生理生态响应
随着全球气候与环境变化的加剧,地球表面平均温度升高、水资源日趋短缺、土壤有效含水量逐年减少以及大气氮沉降已成为重要的全球性环境问题。土壤氮素和水分是许多陆地生态系统中植物生长的主要限制因子,显著影响植物的分布和生长发育。全球气候导致的干旱导致我国落叶松林北移,加之我国大气氮沉降持续增加,导致森林土壤中可利用氮的含量不断增加。因此,探究干旱与氮素综合作用下落叶松幼苗的生长特性、生理特性以及适应性机制,对改善干旱系统植被条件、了解北方森林生态系统功能及格局的改变具有重要意义,也为落叶松人工林繁育过程中合理调节水肥管理、优化培育措施提供理论参考。本研究以1年生落叶松(Larix)为研究对象,采用大棚盆栽方法人为控制土壤水分和施N控制试验,设置三个水分梯度:CK(正常供水,土壤水分含量占田间持水量的80%-70%,下同)、MS(中度干旱胁迫,60%-50%)、HS(严重干旱胁迫,40%-30%);四种施氮水平:NO(不施氮,Og NH4NO3-N·m-2·a-1)、LN(低氮,5g NH4NO3-N·m-2·a-1、MN(中氮,10g NH4NO3-N·m-2·a-1)、HN(高氮,15gNH4NO3-NN·m-2·a-1),从植株生长、生理指标入手,系统的研究了落叶松幼苗对干旱胁迫和施N的生理生态响应,并揭示了落叶松幼苗的耐旱性机制。研究结果表明: (1)随着干旱胁迫程度的增加,不同氮处理下的落叶松幼苗株高、基径生长量呈下降趋势,并对对落叶松幼苗的生物量积累和生物量分配格局具有明显的影响,干旱胁迫越严重,幼苗叶面积、总生物量的积累越少,地上生物量越来越小,地下生物量积累越来越多,R/S升高,SLA和LAR增逐渐加,并产生极显著影响(P0.001)。在干旱胁迫与施氮的交互作用下,施氮处理可在一定程度上改善落叶松幼苗株高、基径、RGR、叶面积、总生物量,促进生长,但是不同水分条件下的氮素效应不同。正常水分供应条件下,施氮水平的提高增加了幼苗生长指标和生物量的积累。而干旱胁迫条件下,施加了过量的氮后幼苗生长量、生物量积累反而降下降,植株通过降低叶面积、降低叶片分配比例来降低自身的水分消耗状况,并增加茎和根的分配比例,将有限的资源较多的生物量分配给地下部分和茎生物量。 (2)不同干旱胁迫处理下,落叶松幼苗光合作用明显受到抑制,随干旱程度增加在同一光强时幼苗Pn逐渐降低,光饱和点逐渐降低。而不同干旱胁迫条件下适量的施氮能在一定程度上提高落叶松幼苗的光合能力。低氮处理下在严重干旱时对植株光合能力有明显的促进作用,而高氮处理下的促进作用减弱并转向抑制。严重干旱情况下,幼苗Pmax在HN水平下表现出最低值,在LN水平下表现出最大值。此时氮处理水平下的Pn和Ci都表现出相同的变化趋势,Pn的下降是非气孔因素引起的。 不同干旱胁迫处理下,落叶松幼苗茎的呼吸作用也明显受到抑制。严重干旱情况下的高氮处理下幼苗的呼吸值最小,说明过量增加氮肥抑制了植物的生长,幼苗的光合作用低的同时,所能固定的光合产物也随之减少,用以为幼苗提供生命活动所需的能量就少。而正常水分供应条件下的高氮浓度下的幼苗呼吸值,和中度干旱情况下的中氮处理下幼苗的呼吸值明显高于同一供水条件下的其他施氮水平,说明在适当的氮处理水平下,落叶松幼苗的生长代谢速度最快,需要通过呼吸作用为其生命活动提供能量。 干旱胁迫使得落叶松幼苗叶片中的叶绿素a、叶绿素b、总叶绿素以及叶绿素a/b上升。但是同一水分处理下各个氮处理水平之间的变化趋势不同,适量的氮素水平能显著提高落叶松幼苗的叶绿素a、叶绿素b、总叶绿素含量,且变化趋势一致。MS和HS条件下下,HN水平都抑制了光合色素的积累,以HN处理下幼苗的Chla、Chlb和Ch1含量最低。但在MS条件下,MN水平下表现出较高的光合色素水平。而在HS条件下,LN水平下表现出较高的光合色素水平。 (3)落叶松幼苗叶片相对含水量(LRWC)随干旱胁迫程度的增加而降低,同一水分处理下不同氮处理水平下的LRWC表现出来的变化也各不相同。中度干旱胁迫下,LRWC随施氮水平的增高表现出先升高后降低的趋势,在MN水平下幼苗LRWC最高,显著高于其他施氮水平(P0.05)。严重干旱胁迫下,LRWC表现出与中度干旱胁迫下相同的变化趋势,但LRWC在LN水平下表现出最高值,且HN水平下表现出最低值。 (4)幼苗总SS含量均随干旱胁迫程度的增加而降低,且同一水分处理下不同氮处理水平之间的变化趋势也显示为一致的趋势,只是变化幅度不同。在HS条件下,叶片、茎、根SS含量随施氮量的增加表现出先增加后降低的趋势,且LN处理表现出SS含量最高,HN处理表现出叶片含量最低。而随着干旱胁迫的加剧,各个施氮水平的幼苗Pro、MDA含量同对照相比整体呈上升趋势。其中,中度干旱胁迫下,各个施氮水平的幼苗Pro含量、MDA含量同对照相比整体呈小幅度上升趋势,但在MN水平下升上幅度最小;而严重干旱胁迫下,各个施氮水平的幼苗Pro含量、MDA含量同正常水分条件下相比上升幅度较大,并以HN水平下的含量最高,LN水平下的含量最低。
Physiological Responds and Adaptation of Miscanthus sacchariflorus and Miscanthus sinensis to Drought Stress. PhD dissertation, Northeast Forestry University
水分是影响植物生长的主要环境因子之一。近年来,随着全球气候变暖,我国水资源的匮乏日益严重,尤其是东北的广大地区,这一问题尤为突出。干旱胁迫已经成为制约东北地区植物生长发育和城市绿化的重要生态问题。荻和芒是理想的第二代能源植物,具有广泛的应用价值,但东北地区水资源匮乏的状况制约了荻和芒在该地区的推广和应用。因此研究干旱胁迫下荻和芒的生长、光合特性和生理特性的变化规律,对提高它的水分管理水平,促进荻和芒在东北地区的可持续发展具有重要意义。 本研究选用芒属植物荻(Miscanthus sacchariflorus)和芒(Misconstrues sinensis)为实验材料,采用聚乙二醇(PEG)模拟干旱法对其种子进行4个PEG浓度水平的胁迫处理(5%、10%、15%、20%),研究荻和芒种子在PEG干旱胁迫下的萌发特性;采用盆栽控水的方法分别对荻和芒的实生苗进行4种土壤水分处理即:正常供水(土壤含水量为最大田间持水量80%,下同)、轻度土壤干旱胁迫(65%)、中度土壤干旱胁迫(50%)和重度土壤干旱胁迫(35%),分别测定了不同土壤干旱胁迫下荻和芒光合、生理和生长指标,研究荻和芒对土壤干旱胁迫的适应性;同时调查了东北地区荻和芒的分布和生境特征,主要研究结果如下: (1)东北野生荻芒分布和生境特征 荻和芒在东北地区属点状分布,喜低海拔、土壤湿润,光照充足的生境;不同生境下荻和芒的生长性状存在差异,随着生境内土壤含水量的上升和光照条件的改善,荻和芒的生长性状和生物量提高,各生境间部分性状差异显著;荻和芒的叶形指数(叶长/叶宽),株高、地下茎粗、分布深度等表型性状受环境因素影响小,相对稳定;荻和芒的株高、茎粗等茎部性状与生物量显著相关。 (2)PEG胁迫对荻和芒种子萌发特性的影响 低浓度的PEG(5-20%)对芒和荻种子的发芽率没有明显的影响,但对芒种子的发芽指数、活力指数、胚根长度、胚根鲜重以及荻种子胚芽的长度有一定的促进作用;高浓度的PEG(15~20%),对芒和荻种子的发芽率、发芽指数、活力指数、胚根胚芽长度以及鲜重都有明显的抑制作用;荻种子的半致死浓度为15%,致死浓度为20%,而芒种子的半致死浓度为20%;随着PEG浓度的升高,芒和荻的萌发抗旱指数逐渐下降,芒在20%PEG处理时表现为中间型,而荻在15%PEG处理时就表现为不抗旱。 (3)土壤干旱胁迫对荻和芒光合特性的影响 土壤干旱胁迫下,荻和芒叶片的相对含水量(RWC)、叶绿素含量(Chl)、净光合速率(Pn)、最大净光合速率(Pmax)、表观量子效率(AQE)、暗呼吸速率(Rd)、光饱和点(LSP)、蒸腾速率(Tr)、最大羧化速率(Vcmax)和电子传递速(Jmax)均随着土壤干旱胁迫程度的加剧而下降;光补偿点(LCP)和水分利用效率(WUE)随土壤干旱胁迫程度的加剧而上升。荻和芒的气孔导度(Gs)随着土壤干旱胁迫程度的加剧而下降;胞间C02浓度(Ci)随着土壤干旱胁迫程度的加剧呈先下降后上升的趋势;气孔阻力值(Ls)随着土壤干旱胁迫程度的加剧呈先上升后下降的趋势。 荻在轻度土壤干旱胁迫下、芒在轻度和中度土壤干旱胁迫下,对光合作用的抑制由气孔限制所引起;而荻在中度、重度土壤干旱胁迫下,芒在重度土壤干旱胁迫下对光合作用的抑制由非气孔限制所引起。 (4)土壤干旱胁迫对荻和芒的生理特性的影响 在土壤干旱胁迫下荻和芒的细胞膜相对透性,丙二醛(MDA)和过氧化氢(H202)含量随着土壤干旱胁迫程度的加剧而显著增加。 在轻度干旱胁迫下,荻的超氧化物歧化酶(SOD)、过氧化氢酶(CAT)和过氧化物酶(POD)活性随着干旱胁迫时间的延长而逐渐上升;而在中度和重度干旱胁迫下SOD、CAT和POD活性随着干旱胁迫时间的延长呈先上升后下降趋势。在不同程度土壤干旱胁迫下,芒的SOD和CAT活性随着干旱胁迫时间的延长而逐渐上升;而POD活性则在不同干旱胁迫处理下无明显变化。 在不同程度干旱胁迫下芒叶片的抗坏血酸过氧化物酶(APX)、甘肽还原酶(GR)活性和还原型抗坏血酸(AsA)、还原型谷胱甘肽(GSH)含量始终保持上升趋势;而荻在轻度干旱胁迫下也可以保持较高的抗坏血酸合成和代谢活性,但在中度和重度干旱胁迫下,随着干旱胁迫的加剧和胁迫时间的延长荻的APX、GR活性和AsA、GSH含量开始下降。 (5)土壤干旱胁迫对荻和芒的生长和生物量的影响 荻和芒的株高、地径、茎节间长度、叶面积和比叶重随着土壤干旱胁迫程度的加剧而下降。轻度干旱胁迫下,芒的地下茎粗和长度均高于正常供水处理,表明适宜的土壤干旱胁迫对芒地下茎的生长有一定的促进作用;而在荻和芒在中、重度干旱胁迫下,地下茎粗、长度和分蘖数量均随着土壤干旱胁迫程度的加剧而显著下降。 土壤干旱胁迫下,除芒在轻度干旱胁迫下地下部分生物量增加外,其余胁迫处理中荻和芒的单株总生物量、地上部分生物量、地下部分生物量和冠根比均随土壤干旱胁迫程度的加剧而下降,胁迫越重,生长速度越慢,生长量越小,生物量越低。
荻和芒对干旱胁迫的生理响应和适应性
水分是影响植物生长的主要环境因子之一。近年来,随着全球气候变暖,我国水资源的匮乏日益严重,尤其是东北的广大地区,这一问题尤为突出。干旱胁迫已经成为制约东北地区植物生长发育和城市绿化的重要生态问题。荻和芒是理想的第二代能源植物,具有广泛的应用价值,但东北地区水资源匮乏的状况制约了荻和芒在该地区的推广和应用。因此研究干旱胁迫下荻和芒的生长、光合特性和生理特性的变化规律,对提高它的水分管理水平,促进荻和芒在东北地区的可持续发展具有重要意义。 本研究选用芒属植物荻(Miscanthus sacchariflorus)和芒(Misconstrues sinensis)为实验材料,采用聚乙二醇(PEG)模拟干旱法对其种子进行4个PEG浓度水平的胁迫处理(5%、10%、15%、20%),研究荻和芒种子在PEG干旱胁迫下的萌发特性;采用盆栽控水的方法分别对荻和芒的实生苗进行4种土壤水分处理即:正常供水(土壤含水量为最大田间持水量80%,下同)、轻度土壤干旱胁迫(65%)、中度土壤干旱胁迫(50%)和重度土壤干旱胁迫(35%),分别测定了不同土壤干旱胁迫下荻和芒光合、生理和生长指标,研究荻和芒对土壤干旱胁迫的适应性;同时调查了东北地区荻和芒的分布和生境特征,主要研究结果如下: (1)东北野生荻芒分布和生境特征 荻和芒在东北地区属点状分布,喜低海拔、土壤湿润,光照充足的生境;不同生境下荻和芒的生长性状存在差异,随着生境内土壤含水量的上升和光照条件的改善,荻和芒的生长性状和生物量提高,各生境间部分性状差异显著;荻和芒的叶形指数(叶长/叶宽),株高、地下茎粗、分布深度等表型性状受环境因素影响小,相对稳定;荻和芒的株高、茎粗等茎部性状与生物量显著相关。 (2)PEG胁迫对荻和芒种子萌发特性的影响 低浓度的PEG(5-20%)对芒和荻种子的发芽率没有明显的影响,但对芒种子的发芽指数、活力指数、胚根长度、胚根鲜重以及荻种子胚芽的长度有一定的促进作用;高浓度的PEG(15~20%),对芒和荻种子的发芽率、发芽指数、活力指数、胚根胚芽长度以及鲜重都有明显的抑制作用;荻种子的半致死浓度为15%,致死浓度为20%,而芒种子的半致死浓度为20%;随着PEG浓度的升高,芒和荻的萌发抗旱指数逐渐下降,芒在20%PEG处理时表现为中间型,而荻在15%PEG处理时就表现为不抗旱。 (3)土壤干旱胁迫对荻和芒光合特性的影响 土壤干旱胁迫下,荻和芒叶片的相对含水量(RWC)、叶绿素含量(Chl)、净光合速率(Pn)、最大净光合速率(Pmax)、表观量子效率(AQE)、暗呼吸速率(Rd)、光饱和点(LSP)、蒸腾速率(Tr)、最大羧化速率(Vcmax)和电子传递速(Jmax)均随着土壤干旱胁迫程度的加剧而下降;光补偿点(LCP)和水分利用效率(WUE)随土壤干旱胁迫程度的加剧而上升。荻和芒的气孔导度(Gs)随着土壤干旱胁迫程度的加剧而下降;胞间C02浓度(Ci)随着土壤干旱胁迫程度的加剧呈先下降后上升的趋势;气孔阻力值(Ls)随着土壤干旱胁迫程度的加剧呈先上升后下降的趋势。 荻在轻度土壤干旱胁迫下、芒在轻度和中度土壤干旱胁迫下,对光合作用的抑制由气孔限制所引起;而荻在中度、重度土壤干旱胁迫下,芒在重度土壤干旱胁迫下对光合作用的抑制由非气孔限制所引起。 (4)土壤干旱胁迫对荻和芒的生理特性的影响 在土壤干旱胁迫下荻和芒的细胞膜相对透性,丙二醛(MDA)和过氧化氢(H202)含量随着土壤干旱胁迫程度的加剧而显著增加。 在轻度干旱胁迫下,荻的超氧化物歧化酶(SOD)、过氧化氢酶(CAT)和过氧化物酶(POD)活性随着干旱胁迫时间的延长而逐渐上升;而在中度和重度干旱胁迫下SOD、CAT和POD活性随着干旱胁迫时间的延长呈先上升后下降趋势。在不同程度土壤干旱胁迫下,芒的SOD和CAT活性随着干旱胁迫时间的延长而逐渐上升;而POD活性则在不同干旱胁迫处理下无明显变化。 在不同程度干旱胁迫下芒叶片的抗坏血酸过氧化物酶(APX)、甘肽还原酶(GR)活性和还原型抗坏血酸(AsA)、还原型谷胱甘肽(GSH)含量始终保持上升趋势;而荻在轻度干旱胁迫下也可以保持较高的抗坏血酸合成和代谢活性,但在中度和重度干旱胁迫下,随着干旱胁迫的加剧和胁迫时间的延长荻的APX、GR活性和AsA、GSH含量开始下降。 (5)土壤干旱胁迫对荻和芒的生长和生物量的影响 荻和芒的株高、地径、茎节间长度、叶面积和比叶重随着土壤干旱胁迫程度的加剧而下降。轻度干旱胁迫下,芒的地下茎粗和长度均高于正常供水处理,表明适宜的土壤干旱胁迫对芒地下茎的生长有一定的促进作用;而在荻和芒在中、重度干旱胁迫下,地下茎粗、长度和分蘖数量均随着土壤干旱胁迫程度的加剧而显著下降。 土壤干旱胁迫下,除芒在轻度干旱胁迫下地下部分生物量增加外,其余胁迫处理中荻和芒的单株总生物量、地上部分生物量、地下部分生物量和冠根比均随土壤干旱胁迫程度的加剧而下降,胁迫越重,生长速度越慢,生长量越小,生物量越低。
Physiological responses of Lycium ruthenicum Murr. seedlings to drought stress.
黑果枸杞幼苗对干旱胁迫的生理响应
An improved method for the determination of hydrogen peroxide in leaves
DOI:10.3321/j.issn:1000-3282.2000.05.025
URL
Ti(Ⅳ)-H2O2比色法因背景物质干扰而测得的植物叶片内H2O2含量偏高,5%三氯乙酸抽提,活性炭脱色,Ti(Ⅳ)-4-(2-吡啶偶氮)间苯二酚(PAR)比色法测得的H2O2含量偏低。萃取法有效地脱去丙酮提液中的色素,且H2O2的回收率在95%以上。用过氧化氢酶(CAT)处理作空白对照,利用H2O2与Ti(Ⅳ)-PAR的显色反应,建立了一种简便、快速、准确的植物叶片内的H2O2含量测定方法,H2
植物叶片中过氧化氢含量测定方法的改进
DOI:10.3321/j.issn:1000-3282.2000.05.025
URL
Ti(Ⅳ)-H2O2比色法因背景物质干扰而测得的植物叶片内H2O2含量偏高,5%三氯乙酸抽提,活性炭脱色,Ti(Ⅳ)-4-(2-吡啶偶氮)间苯二酚(PAR)比色法测得的H2O2含量偏低。萃取法有效地脱去丙酮提液中的色素,且H2O2的回收率在95%以上。用过氧化氢酶(CAT)处理作空白对照,利用H2O2与Ti(Ⅳ)-PAR的显色反应,建立了一种简便、快速、准确的植物叶片内的H2O2含量测定方法,H2
Physiological responses of the limestone endemic plant Triadica rotundifolia seedlings to drought stress.
石漠化树种圆叶乌桕对干旱胁迫的生理响应
Physiological responses of the desert shrub
DOI:10.11686/cyxb2015274
URL
Magsci
[本文引用: 1]
采用盆栽实验,研究了不同程度干旱胁迫(重度、中度、轻度、对照,分别为15%、30%、50%、70%田间最大持水量)对荒漠灌木蒙古岩黄芪幼苗光合特性、抗氧化特性和渗透调节物质的影响。结果显示,在中度和重度干旱胁迫下,除胞间CO<sub>2</sub> 浓度外,其余各光合指标均显著下降;在轻度干旱胁迫下,蒙古岩黄芪幼苗净光合速率、PSⅡ的潜在活性与对照相比上升了20.7%和5.7%。中度和重度胁迫下蒙古岩黄芪叶片的丙二醛(MDA)含量、细胞相对质膜透性、超氧化物歧化酶(SOD)、过氧化氢酶(CAT)以及过氧化物酶(POD)活性显著高于对照,而在轻度干旱胁迫下无明显变化。叶片中游离脯氨酸含量和可溶性糖含量以及K<sup>+</sup>含量随干旱胁迫的加剧而显著增加;根、茎、叶中的Na<sup>+</sup>含量在干旱胁迫处理中均显著下降。蒙古岩黄芪在干旱胁迫下采取协同升高叶片中SOD、 CAT、POD活性,提高脯氨酸和可溶性糖含量,同时选择性吸收K<sup>+</sup>的策略来减小干旱对植株所造成的伤害。实验结果可为蒙古岩黄芪适应干旱胁迫的策略提供理论依据,为其保护利用以及干旱与半干旱区的植被恢复提供参考。
荒漠灌木蒙古岩黄芪对干旱胁迫的生理响应
DOI:10.11686/cyxb2015274
URL
Magsci
[本文引用: 1]
采用盆栽实验,研究了不同程度干旱胁迫(重度、中度、轻度、对照,分别为15%、30%、50%、70%田间最大持水量)对荒漠灌木蒙古岩黄芪幼苗光合特性、抗氧化特性和渗透调节物质的影响。结果显示,在中度和重度干旱胁迫下,除胞间CO<sub>2</sub> 浓度外,其余各光合指标均显著下降;在轻度干旱胁迫下,蒙古岩黄芪幼苗净光合速率、PSⅡ的潜在活性与对照相比上升了20.7%和5.7%。中度和重度胁迫下蒙古岩黄芪叶片的丙二醛(MDA)含量、细胞相对质膜透性、超氧化物歧化酶(SOD)、过氧化氢酶(CAT)以及过氧化物酶(POD)活性显著高于对照,而在轻度干旱胁迫下无明显变化。叶片中游离脯氨酸含量和可溶性糖含量以及K<sup>+</sup>含量随干旱胁迫的加剧而显著增加;根、茎、叶中的Na<sup>+</sup>含量在干旱胁迫处理中均显著下降。蒙古岩黄芪在干旱胁迫下采取协同升高叶片中SOD、 CAT、POD活性,提高脯氨酸和可溶性糖含量,同时选择性吸收K<sup>+</sup>的策略来减小干旱对植株所造成的伤害。实验结果可为蒙古岩黄芪适应干旱胁迫的策略提供理论依据,为其保护利用以及干旱与半干旱区的植被恢复提供参考。
Influence of soil moisture on soil solution chemistry and concentrations of minerals in the calcicoles Phleum phleoides and Veronica spicata grown on a limestone soil.
The role of organic solute and ion accumulation in osmotic adjustment in drought-stressed grapevines
DOI:10.1016/S0168-9452(02)00140-1
URL
[本文引用: 1]
Water relations, gas exchange as well as organic solute and ion accumulation were studied in the leaves of 2-year-old grapevines ( Vitis vinifera L, cv. Savatiano) grown under well watered (control) and water stress conditions. Both osmotic potential at full turgor ( 螤 100) and at turgor loss point ( 螤 0) decreased significantly in stressed plants compared with the control. Photosynthetic rate, and stomatal conductance were also significantly lower in stressed plants. Starch concentration decreased almost threefold in stressed plants, while there were no significant differences in sugar accumulation between the two treatments. Total inorganic ion concentration increased rapidly in stressed plants and seems to be the major component of osmotic adjustment in stressed grapevines. Thus, the energetic cost of osmotic adjustment in grapevines using inorganic ions would be expected to be much lower than for those species using organic solutes.
Effect of water stress and nitrogen application on morphological and physiological character of Zoysia japonica cv. Shanghai.
分析了氮、水分与上海结缕草(Zoysia japonica cv. Shanghai)生长之间的相互关系,为上海结缕草草坪科学施肥和灌溉提供依据。试验选择干旱胁迫和施氮2个因素,各设置3个水平,结果表明:轻度干旱胁迫+中氮组合处理的草坪品质非常好,草坪密度和地上生物量高于或接近无干旱胁迫+中氮组合处理,且显著好于轻度干旱胁迫的其他处理和重度干旱胁迫的全部处理(P0.05);高氮处理的结缕草株高在同一干旱水平中均明显高于其他处理(P0.05)。在相同干旱胁迫下,施氮后第19天,结缕草叶片的相对电导率、游离脯氨酸含量和可溶性糖含量均随施氮量的增加而呈现升高的趋势;与此相反,施氮后第41天时,则呈现出随施氮量的增加而降低的趋势;随着干旱胁迫程度的增加,叶片丙二醛含量呈现增加的趋势。研究结果表明,适度的干旱胁迫和适量的氮肥有助于维持上海结缕草较好的坪用品质。
干旱胁迫和施氮对结缕草种群特征和生理特性的影响
分析了氮、水分与上海结缕草(Zoysia japonica cv. Shanghai)生长之间的相互关系,为上海结缕草草坪科学施肥和灌溉提供依据。试验选择干旱胁迫和施氮2个因素,各设置3个水平,结果表明:轻度干旱胁迫+中氮组合处理的草坪品质非常好,草坪密度和地上生物量高于或接近无干旱胁迫+中氮组合处理,且显著好于轻度干旱胁迫的其他处理和重度干旱胁迫的全部处理(P0.05);高氮处理的结缕草株高在同一干旱水平中均明显高于其他处理(P0.05)。在相同干旱胁迫下,施氮后第19天,结缕草叶片的相对电导率、游离脯氨酸含量和可溶性糖含量均随施氮量的增加而呈现升高的趋势;与此相反,施氮后第41天时,则呈现出随施氮量的增加而降低的趋势;随着干旱胁迫程度的增加,叶片丙二醛含量呈现增加的趋势。研究结果表明,适度的干旱胁迫和适量的氮肥有助于维持上海结缕草较好的坪用品质。
Proline and water deficit: Ups, downs, ins and outs
DOI:10.1105/tpc.8.8.1221 URL [本文引用: 1]
The worldwide leaf economics spectrum
DOI:10.1038/nature02403 URL [本文引用: 1]
Effect of the moisture content coercive on morphological and physiological process of crops
水分胁迫对作物形态和生理过程的影响
Research progress of plant water stress inducible proteins
DOI:10.3969/j.issn.1000-1336.2002.02.028
URL
[本文引用: 1]
主要介绍了植物水分胁迫诱导蛋白的表达模式、特征、分类、功能及其诱导过程中的信号转导及诱导机制。认为胁迫诱导蛋白的产生是植物对逆境胁迫的一种适应性反应 ,诱导蛋白从多方面保护植物避免或减少胁迫所造成的伤害。植物通过多种途径感受并转导干旱胁迫信号 ,诱导出多种基因表达产物 ,从而尽可能地增强对逆境的抗性
植物水分胁迫诱导蛋白的研究进展
.
DOI:10.3969/j.issn.1000-1336.2002.02.028
URL
[本文引用: 1]
主要介绍了植物水分胁迫诱导蛋白的表达模式、特征、分类、功能及其诱导过程中的信号转导及诱导机制。认为胁迫诱导蛋白的产生是植物对逆境胁迫的一种适应性反应 ,诱导蛋白从多方面保护植物避免或减少胁迫所造成的伤害。植物通过多种途径感受并转导干旱胁迫信号 ,诱导出多种基因表达产物 ,从而尽可能地增强对逆境的抗性
Growth and physiological responses to drought stress and comprehensive evaluation on drought tolerance in
白杨派无性系苗期对干旱胁迫的生长生理响应及抗旱性综合评价
Progress on crops osmotic adjustment and genetic engineering of osmotic stress resistance
作物渗透调节及抗渗透胁迫基因工程研究进展
Drought-stress-induced changes in activities of superoxide dismutase, catalase, and peroxidase in wheat species
DOI:10.1016/S0143-4004(05)80424-2
URL
ABSTRACT Activities of superoxide dismutase (SOD), catalase (CAT), and peroxidase (POD), as well as malondialdehyde (MDA) contents and solute potentials, were studied in seedlings of seven wheat ( Triticum ) species (nine genotypes representing three ploidy levels: hexaploid, tetraploid, diploid) subjected to water stress for 4, 8, and 12 days by withholding water. Solute potentials of all genotypes were lowered by water stress. In most species, SOD and CAT activities showed an increase or maintenance in the early phase of drought and then a decrease with further increase in magnitude of water stress. On the contrary, POD activities and MDA contents greatly increased in response to water stress. Enzymatic activities partly recovered and MDA contents decreased with rewatering. Under drought, hexaploid wheats had higher POD activities and MDA contents than tetraploid and diploid wheats; solute potentials and activities of SOD and CAT, however, were similar among the three groups. These results suggest that water stress alters the equilibrium between free radical production and enzymatic defense reactions in wheat species and that hexaploid wheats have less efficient antioxidant systems (e.g., the ascorbate-glutathione cycle and the nonenzymatic system) than tetraploid and diploid wheats.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}