


灌丛是陆地生态系统重要的组成部分, 在森林恢复、水土保持和生态平衡维持中起着非常重要的作用(谢宗强和唐志尧, 2017; 张建华等, 2017)。全球气候变暖和人为干扰会不断影响灌丛的分布范围和物种组成(Sturm et al., 2001), 进而影响群落的生产力和生物多样性(彭海英等, 2013)。因此, 明确灌丛的分布规律和物种组成特征对指导植被恢复和进行物种多样性保护具有重要的意义。我国灌丛面积近2 × 108 hm2, 占陆地总面积的1/5, 是全国现存森林面积的近2倍(侯学煜, 1982; 胡会峰等, 2006), 是我国陆地生态系统中重要的植被类型。近年来, 由于退化植被的恢复及重建, 我国灌丛植被的分布面积增长迅速, 已成为国内植被生态学家关注的重点。不少学者已经对我国灌丛生物量的分布及其与环境的关系(杨弦等, 2017; 张蔷等, 2017), 灌丛生态系统的碳、氮、磷储量(郭焱培等, 2017; 李家湘等, 2017b)等方面做了详细研究。还有一些研究报道了我国华南地区典型亚热带灌丛的物种组成特征(张亚茹等, 2013), 分析了常绿阔叶林区域内灌丛维管植物生活型组成, 并探讨了亚热带灌丛生活型谱特征及其与水热因子间的关系(李家湘等, 2017a)。然而, 关于我国北方灌丛群落当前的分布情况和物种组成的本底资料仍然十分缺乏。华北地区属暖温带半湿润-半干旱气候, 沿海比较湿润, 其地带性植被为落叶阔叶林(王荷生, 1999), 由于人类活动的干扰, 该地区的森林常发生逆行演替, 退化为草地和灌木林地, 并且在本地区广泛分布, 使植被的环境效应发生很大的变化。同时, 显著的降水梯度和温度梯度使得该地区灌丛类型多样, 这对分析和理解灌丛群落的物种共存和群落构建机制有重要作用。
群落内物种的共存模式是物种的进化历史和其他生态过程共同作用的结果(Webb, 2000; 牛克昌等, 2009; Rosindell et al., 2011; 朱璧如和张大勇, 2011), 分析共存物种的亲缘关系可以从系统发育角度探讨群落物种组成的现状和原因(黄建雄等, 2010), 有助于更准确地理解群落的构建机制。基于系统发育推断群落构建机制的前提假设是物种的生态特征在进化上是保守的, 即物种间的亲缘关系越近, 生态特征越相似, 因而可以用谱系距离来代表生态特征距离。目前, 基于生态位理论的环境过滤和竞争排斥被认为是群落构建过程中的主要驱动力。一般而言, 环境过滤作用会使得相同生境出现亲缘关系较近、适应能力相似的物种, 群落结构会表现为系统发育的聚集(牛红玉等, 2011); 相反, 竞争排斥作用会使亲缘关系较近、生态位相似的物种无法共存于同一环境, 群落内的物种表现为系统发育的发散(Webb et al., 2002; 牛红玉等, 2011)。当然, 如果更多的远缘物种由于种间竞争被剔除, 群落的系统发育结构也会表现为汇聚(Mayfield & Levine, 2010)。若群落构建为随机的中性过程, 则根据中性理论(Hubbell, 2001), 由于随机的生态漂变、扩散限制等过程, 群落内物种组成会表现为系统发育随机。
一个群落的系统发育结构模式可能受多种生态因子的影响。近年来, 区域尺度上对群落系统发育结构的研究逐渐证明了气候因子、空间尺度、干扰、地形及土壤因子对森林或草地群落系统发育结构的影响(黄建雄等, 2010; 闫帮国等, 2010; 宋凯等, 2011; 卢孟孟等, 2014; 柴永福和岳明, 2016)。比如, Michalski等(2017)和Whitfeld等(2012)证明了环境过滤在热带和亚热带森林演替过程中系统发育构建的作用。在我国, Qian等(2014)和Huang等(2014)分别对中国长白山温带森林和南部亚热带森林的研究也表明, 环境因子是温带和亚热带森林群落系统发育结构变化的主要驱动因素。另外, 由于群落内垂直结构上物种组成的差异, 不同垂直结构层物种的系统发育结构也有差异(Xu et al., 2017), 且不同结构层的系统发育结构对环境的响应也不同, 如长白山温带森林中环境因子对木本层群落的影响要强于对草本层群落的影响(Qian et al., 2014)。此外, 对于同一种植被类型, 由于不同建群种对环境适应策略的差异会导致不同群系的分布范围和物种组成有显著不同, 这种不同是否会体现在群落的谱系结构上仍然不清楚。因此, 在区域尺度上探讨不同群落类型的系统发育结构特征以及其环境驱动因子, 有利于进一步认识影响群落系统发育结构的生态学过程。
以往对群落系统发育结构的研究多集中在森林或草地群落, 很少关注灌丛群落系统发育结构的变化模式及影响因子。本研究基于华北地区自然植物群落资源综合考察数据库的灌丛调查数据, 分析我国华北地区灌丛群落的分布、物种组成及系统发育结构特征, 同时结合WorldClim数据库提取的环境因子分析其环境驱动机制。具体分析如下问题: (1)华北地区主要典型灌丛群落的分布格局及物种组成特征。(2)华北地区主要典型灌丛群落的系统发育结构特征, 并对比不同群落类型、不同垂直结构层的系统发育结构特征。(3)灌丛群落系统发育结构变化的环境驱动因子及其与群落类型的关系。
1 材料和方法
1.1 样方建立和植物群落调查
本研究所有数据源于华北地区自然植物群落资源综合考察数据库。采用分层随机抽样方法来布设采样点, 以1:100万中国植被图为依据, 以群系为设计单元, 兼顾植被面积及其复杂程度。具体操作为: 1)将各县域与植被图叠加, 获取各县分布的主要植物群落类型; 2)以群系类型为基础, 在各省域以适当的面积比例设置样方数量; 在山地, 要按海拔高度和植被类型设置样地。本研究所调查的行政区包括北京市(BEJ)、天津市(TIJ)、河北省(HEB)、河南省(HEN)、陕西省(SHX)、山西省(SAX)、山东省(SHD)、宁夏回族自治区(NIX)、内蒙古自治区(NMG)和甘肃省(GSU)。
本研究进行数据分析所用的灌丛样方数为2 331个(图1), 每个灌丛样方调查面积为100 m2 (划分为4个5 m × 5 m的样格); 如果群落类型较破碎, 则利用4个相邻不超过50 m的5 m × 5 m的样格。按照《华北地区自然植物群落资源综合考察规范》记录样方的坡度、坡向和海拔等环境特征并对灌木层和草本层分别进行常规群落学调查。对于灌木层物种记录所有物种的种名、平均高度、平均基径、盖度和多度;对于草本层物种记录种名、平均高度、盖度和多度等级。
图1
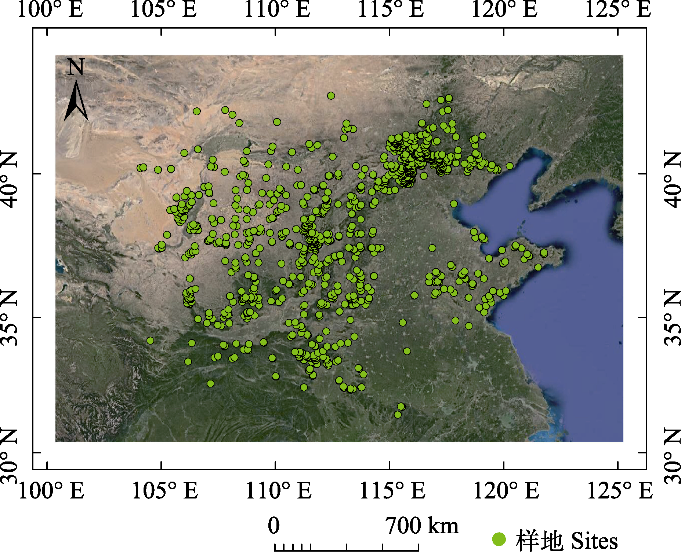
图1
华北地区灌丛调查样方分布图。
Fig. 1
Distribution of survey plots for shrubland in North China.
1.2 系统发育结构特征
以2 331个样方中调查的所有灌木和草本物种为物种库, 以APGIII系统为大框架, 利用Phylomatic (http://www.phylodiversity.net)基于Zanne等(2014)构建的进化树骨架生成带有枝长的进化树。
净亲缘关系指数(NRI)和最近种间亲缘关系指数(NTI)是表征群落系统发育结构最常用的两个指数。NRI侧重于从整体上描述群落中物种形成的谱系结构, 而NTI着重描述近缘物种间构成的群落谱系格局(Swenson et al., 2007)。本文侧重分析群落整体的谱系结构, 因此仅用NRI表征群落的系统发育结构(Webb, 2000), 具体计算在R软件中的Picante包中进行(Webb et al., 2002)。计算过程如下: 1)基于所构建的进化树分别计算每个样方中灌木层和草本层物种的平均成对谱系距离(MPD); 2)保持物种丰富度和发生频率, 用独立置换的方法作为零模型生成随机群落矩阵(Gotelli, 2000), 运行999次从而获得该样方中物种在随机零模型下的MPD的分布, 最后利用随机分布结果将观察值标准化, 从而获得NRI。由于灌木和草本物种多度量化上标准不统一, 所以在计算过程中均不进行多度加权。
NRI计算公式如下:
$NRI=\frac{MPD_{observed}-MPD_{randomized}}{ sdMPD_{randomized}}$
MPDobserved为观察值, MPDrandomized为随机零模型期望值(n = 999), sdMPDrandomized为零模型分布的标准偏差。NRI < 0说明MPD低于期望值, 表示系统发育的汇聚; NRI > 0表示系统发育发散; NRI = 0表示系统发育结构为随机。NRI绝对值大于1.96认为群落的系统发育结构显著偏离零模型。
1.3 气候因子
气候基准数据来源于WorldClim数据库(http:// www.worldclim.org), 该数据库收集了1950-2000年全球各地气象站的每月气象数据, 将2000年作为基准年采用插值法生成全球气候栅格数据。本文利用R软件Raster软件包提取了华北地区的19个气候因子(Bio1-Bio19)(表1), 空间分辨率为1 km。
表1 华北地区灌丛研究中采用的环境因子
Table 1
| 编码 Code | 环境因子 Environmental variables | 缩写 Abbreviation |
|---|---|---|
| Bio1 | 年平均气温 Annual mean air temperature | AMT |
| Bio2 | 每月最高气温与最低气温差值的平均值 Mean diurnal air temperature range | MDTR |
| Bio3 | 等温性 Isothermality | ISO |
| Bio4 | 季节性气温变异 Air temperature seasonality | TS |
| Bio5 | 最热月的最高气温 Max temperature of the warmest month | MTWM |
| Bio6 | 最冷月的最低气温 Min temperature of the coldest month | MTCM |
| Bio7 | 气温的年较差 Temperature annual range | TAR |
| Bio8 | 最湿季度的平均气温 Mean temperature of the wettest quarter | MTWEQ |
| Bio9 | 最干季度的平均气温 Mean temperature of the driest quarter | MTDQ |
| Bio10 | 最热季度的平均气温 Mean temperature of the warmest quarter | MTWAQ |
| Bio11 | 最冷季度的平均气温 Mean temperature of the coldest quarter | MTCQ |
| Bio12 | 年降水量 Annual precipitation | AP |
| Bio13 | 最湿月的降水量 Precipitation of the wettest month | PWM |
| Bio14 | 最干月的降水量 Precipitation of the driest month | PDM |
| Bio15 | 降水季节性变化 Precipitation seasonality | PS |
| Bio16 | 最湿季度的降水量 Precipitation of the wettest quarter | PWEQ |
| Bio17 | 最干季度的降水量 Precipitation of the driest quarter | PDQ |
| Bio18 | 最热季度的降水量 Precipitation of the warmest quarter | PWAQ |
| Bio19 | 最冷季度的降水量 Precipitation of the coldest quarter | PCQ |
| Alt | 海拔 Altitude | Alt |
1.4 数据分析
依据陈灵芝等(2014)所著的《中国植物区系与植被地理》对所有样方划分植被型和群系类型, 并统计其数量特征, 确定华北灌丛的主要植被型和群系类型。按照不同群系类型在各地区的样方分布情况确定灌丛群落的分布特点。按科、属分别统计物种数量, 并计算木本物种和草本物种的相对频度(RF), RF = (该种的频度/所有种的频度总和) × 100 (叶万辉等, 2008)。
对所有灌丛群落的木本物种和草本物种分别计算物种丰富度和NRI, 并用t检验比较木本物种和草本物种在物种丰富度和系统发育结构上的差异, 同时用单因素方差分析比较同一结构层不同植被型间物种丰富度和系统发育结构的差异。用双尾t检验分析木本物种和草本物种的NRI平均值与0的差异, 若均值显著大于或小于0表示系统发育结构汇聚或 发散。
利用向前逐步回归法分析并筛选影响灌丛木本物种和草本物种系统发育结构变化的主要环境因子, 并用一般线性模型分析这些环境因子与植被型和群落类型的交互作用, 验证不同群系类型系统发育结构对环境的响应。数据统计相对频度在R软件(R Development Core Team, 2014))中完成, t检验在Statistic 6.0软件中完成。
2 结果
2.1 灌丛群落物种组成特征及分布情况
依据《中国植物区系与植被地理》将灌丛群落划分为5种植被型, 分别为落叶灌丛、常绿针叶灌丛、常绿革叶灌丛、灌丛荒漠和竹灌丛, 共包括195个群系类型。结果显示, 落叶灌丛为主要植被型, 分布最广, 且包含的群系类型最多(图2A)。调查样方数最多的群系类型为虎榛子(Ostryopsis davidiana)灌丛、荆条(Vitex negundo var. heterophylla)灌丛、荆条+酸枣(Ziziphus jujuba)灌丛、沙棘(Hippophae rhamnoides)灌丛(图2B)。按地区分类后, 各群系类型的分布表现出明显的地域特点: 落叶灌丛在各地区都有分布, 而陕西、山西、山东、北京、河北、河南等地居多(图3); 常绿革叶灌丛在陕西、山西和河南的高山和亚高山地区有少量分布, 主要以杜鹃(Rhodoclendron simsii)灌丛为主; 灌丛荒漠主要分布于内蒙古和宁夏。
图2
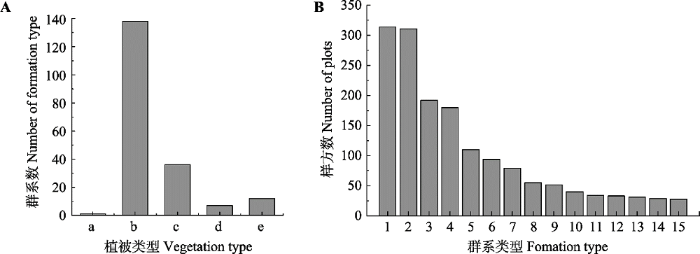
图2
华北地区灌丛的主要植被类型(A)和群系类型(B)。植被类型: a, 竹灌丛; b, 落叶灌丛; c, 灌丛荒漠; d, 常绿针叶灌丛; e, 常绿革叶灌丛。群系类型: 1, 虎榛子灌丛; 2, 荆条灌丛; 3, 荆条+酸枣灌丛; 4, 沙棘灌丛; 5, 山杏灌丛; 6, 黄刺玫灌丛; 7, 绣线菊灌丛; 8, 锦鸡儿灌丛; 9, 柠条锦鸡儿灌丛; 10, 酸枣灌丛; 11, 红砂荒漠; 12, 柽柳灌丛; 13, 野皂荚灌丛; 14, 白刺花灌丛; 15, 黄蔷薇灌丛。
Fig. 2
Major vegetation types (A) and formation types (B) of shrubland in North China. Vegetation types: a, bamboo thickets; b, deciduous thickets; c, desert thickets; d, evergreen needle-leaved thickets; e, evergreen leather-leaved thickets. Formation types: 1, Ostryopsis davidiana shrub; 2, Vitex negundo var. heterophylla shrub; 3, Vitex negundo var. heterophylla + Ziziphus jujube shrub; 4, Hippophae rhamnoides shrub; 5, Armeniaca sibirica shrub; 6, Rosa xanthine shrub; 7, Spiraea salicifolia shrub; 8, Caragana sinica shrub; 9, Caragana korshinskii shrub; 10, Ziziphus jujube shrub; 11, Reaumuria soongarica shrub; 12, Tamarix chinensis shrub; 13, Gleditsia microphylla shrub; 14, Sophora davidii shrub; 15, Rosa hugonis shrub.
图3
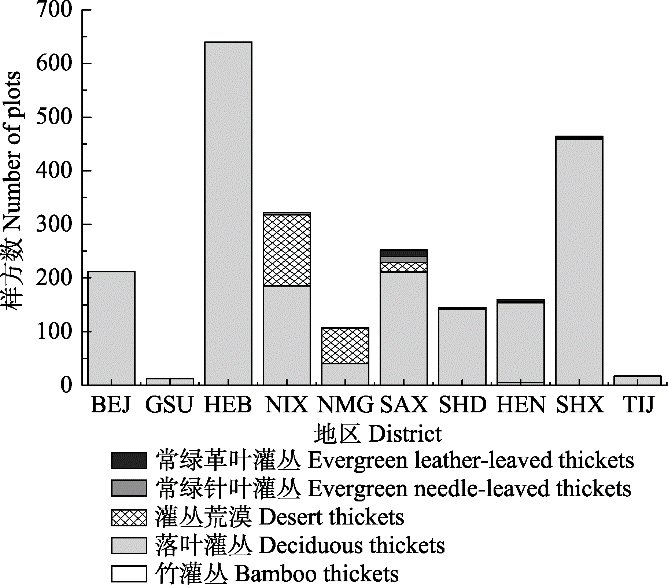
图3
华北地区灌丛主要植被型在各地区的分布。BEJ, 北京; TIJ, 天津; HEB, 河北; HEN, 河南; SHX, 陕西; SAX, 山西; SHD, 山东; NIX, 宁夏; NMG, 内蒙古; GSU, 甘肃。
Fig. 3
Distribution of major vegetation types of shrubland in North China. BEJ, Beijing; TIJ, Tianjin; HEB, Hebei; HEN, Henan; SHX, Shaanxi; SAX, Shanxi; SHD, Shandong; NIX, Ningxia; NMG, Nei Mongol; GSU, Gansu.
经统计, 共记录维管植物1 791种(不包括未鉴定到属的物种), 其中灌木层木本物种(包括乔木、灌木、亚灌木和木质藤本) 570种, 分属75科, 207个属, 占总种数的31.82%; 草本物种(草质藤本和草本) 1 221种, 分属99科, 491属, 占总种数的68.18%。木本物种主要的科有蔷薇科、豆科、忍冬科、鼠李科、木犀科、杨柳科、壳斗科、卫矛科、榆科和小檗科(表2)。草本物种主要的科有菊科、禾本科、豆科、毛茛科、唇形科、蔷薇科、苋科、伞形科、十字花科和蓼科。荆条、酸枣和虎榛子是频度最高的灌木物种, 白莲蒿(Artemisia sacrorum)、大披针薹草(Carex lanceolata)和狗尾草(Setaria viridis)是频度最高的草本物种(表3)。草本物种的丰富度显著高于木本植物, 不同植被型间物种丰富度差异不显著(图4)。
表2 华北地区主要灌丛群落中物种数前10的科
Table 2
| 序号 Rank | 木本植物 Woody species | 草本植物 Herbaceous species | ||
|---|---|---|---|---|
| 科名 Family | 物种数 No. of species | 科名 Family | 物种数 No. of species | |
| 1 | 蔷薇科 Rosaceae | 101 | 菊科 Asteraceae | 219 |
| 2 | 豆科 Fabaceae | 62 | 禾本科 Poaceae | 138 |
| 3 | 忍冬科 Caprifoliaceae | 24 | 豆科 Fabaceae | 101 |
| 4 | 鼠李科 Rhamnaceae | 23 | 毛茛科 Ranunculaceae | 60 |
| 5 | 木犀科 Oleaceae | 20 | 唇形科 Lamiaceae | 55 |
| 6 | 杨柳科 Salicaceae | 17 | 蔷薇科 Rosaceae | 48 |
| 7 | 壳斗科 Fagaceae | 16 | 苋科 Amaranthaceae | 40 |
| 8 | 卫矛科 Celastraceae | 14 | 伞形科 Apiaceae | 37 |
| 9 | 榆科 Ulmaceae | 12 | 十字花科 Brassicaceae | 27 |
| 10 | 小檗科 Berberidaceae | 11 | 蓼科 Polygonaceae | 26 |
| 合计 Total | 300 | 合计 Total | 751 | |
表3 华北地区主要灌丛群落中物种频度前10的物种
Table 3
| 序号 Rank | 木本植物 Woody species | 草本植物 Herbaceous species | ||
|---|---|---|---|---|
| 种名 Species | 相对频度 Relative frequency | 种名 Species | 相对频度 Relative frequency | |
| 1 | 荆条 Vitex negundo var. heterophylla | 9.53 | 白莲蒿 Artemisia sacrorum | 2.35 |
| 2 | 酸枣 Ziziphus jujuba var. spinosa | 4.76 | 大披针薹草 Carex lanceolata | 2.24 |
| 3 | 虎榛子 Ostryopsis davidiana | 3.98 | 狗尾草 Setaria viridis | 1.74 |
| 4 | 山杏 Armeniaca sibirica | 3.12 | 北京隐子草 Cleistogenes hancei | 1.52 |
| 5 | 土庄绣线菊 Spiraea pubescens | 2.78 | 委陵菜 Potentilla chinensis | 1.52 |
| 6 | 沙棘 Hippophae rhamnoides | 2.76 | 小红菊 Dendranthema chanetii | 1.45 |
| 7 | 三裂绣线菊 Spiraea trilobata | 2.71 | 阿尔泰狗娃花 Heteropappus altaicus | 1.37 |
| 8 | 黄刺玫 Rosa xanthina | 2.50 | 茜草 Rubia cordifolia | 1.31 |
| 9 | 二色胡枝子 Lespedeza bicolor | 2.16 | 山蒿 Artemisia brachyloba | 1.31 |
| 10 | 绣线菊 Spiraea salicifolia | 1.83 | 野青茅 Deyeuxia arundinacea | 1.29 |
| 合计 Total | 36.13 | 合计 Total | 16.10 | |
图4
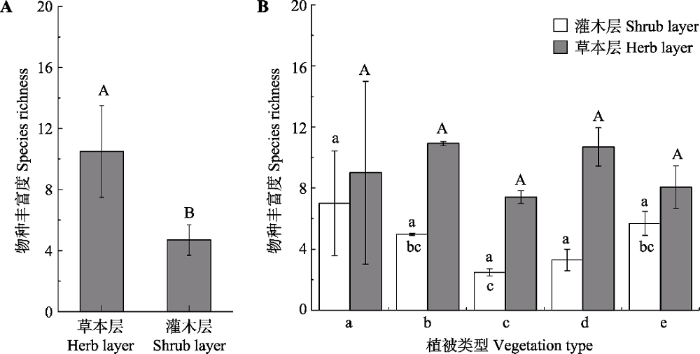
图4
华北地区灌丛灌木层和草本层的物种丰富度(平均值±标准误差)。A, 所有群落草本层和灌木层物种丰富度之间的差异。B, 同一结构层不同植被型之间的差异。用t检验分析整体的差异, 用单因素方差分析检验不同植被型之间的差异。柱形上方不同字母代表相应处理之间在5%水平有显著性差异。a, 竹灌丛; b, 落叶灌丛; c, 灌丛荒漠; d, 常绿针叶灌丛; e, 常绿革叶灌丛。
Fig. 4
Species richness of shrub species and herb species of shrubland in North China (mean ± SE). A, Differences of species richness between herb and shrub species for all communities. B, Differences among vegetation types. Differences between herb and shrub species for all communities was texted by t test and differences among vegetation types were tested using a one-way ANOVA with a Tukey post hoc test of significance. Significance different at p < 0.05 was indicated by different letters. a, bamboo thickets; b, deciduous thickets; c, desert thickets; d, evergreen needle-leaved thickets; e, evergreen leather-leaved thickets.
2.2 不同植被型和不同垂直结构层群落的系统发育结构
图5
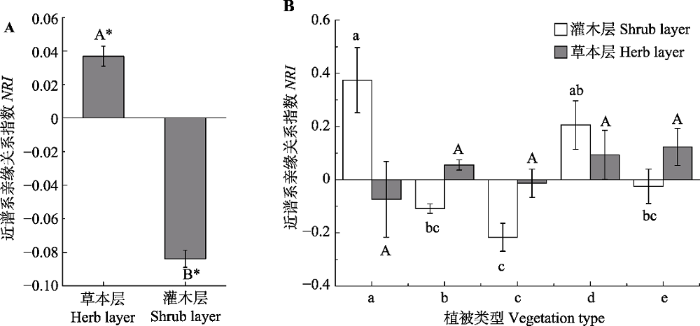
图5
华北地区灌丛灌木层和草本层物种的净谱系亲缘关系指数(NRI)(平均值±标准误差)。A, 所有群落草本层和灌木层物种丰富度之间的差异。B, 同一结构层不同植被型之间的差异。用t检验分析整体的差异, 用单因素方差分析检验不同植被型之间的差异。柱形上方不同字母代表相应处理之间在5%水平有显著性差异。*表示NRI的均值和零值有显著差异(p < 0.05)。a, 竹灌丛; b, 落叶灌丛; c, 灌丛荒漠; d, 常绿针叶灌丛; e, 常绿革叶灌丛。
Fig. 5
Net relatedness index (NRI) of shrub species and herb species of shrubland in North China (mean ± SE). A, Differences of species richness between herb and shrub species for all communities. B, Differences among vegetation types. Differences between herb and shrub species for all communities was texted by t test and differences among vegetation types were tested using a one-way ANOVA with a Tukey post hoc test of significance. Significance different at p < 0.05 was indicated by different letters. * indicates that the mean of NRI was different with zero. a, bamboo thickets; b, deciduous thickets; c, desert thickets; d, evergreen needle-leaved thickets; e, evergreen leather-leaved thickets.
2.3 气候因子对灌丛系统发育结构的影响
表4 环境因子对华北地区灌丛群落灌木层和草本层物种净谱系亲缘关系指数(NRI)的影响
Table 4
| 结构层 Structural layer | R2 | p | 显著环境因子 Significant environmental factors | 标准化回归系数 Beta regression coefficient | t | p |
|---|---|---|---|---|---|---|
| 灌木层 Shrub layer | 0.18 | <0.05 | MTWEQ | 6.81 | 7.91 | <0.01 |
| PWEQ | 5.08 | 5.16 | <0.01 | |||
| MDTR | 2.39 | 2.19 | 0.03 | |||
| MTCQ | 1.74 | 2.73 | 0.01 | |||
| Alt | 0.22 | 2.44 | 0.01 | |||
| ISO | -0.89 | -2.71 | 0.01 | |||
| PWAQ | -5.45 | -4.64 | <0.01 | |||
| MTWM | -9.54 | -8.32 | <0.01 | |||
| 草本层 Herb layer | 0.05 | <0.05 | TS | 0.12 | 2.46 | 0.01 |
| MTWEQ | 0.12 | 2.56 | 0.01 | |||
| PDM | 0.12 | 2.84 | <0.01 | |||
| PS | 0.08 | 2.39 | 0.02 |
See
环境因子同
表5 华北地区植被型(VT)和环境因子对净谱系亲缘关系指数的交互 作用
Table 5
| 交互作用 | 灌木 Shrub | ||
|---|---|---|---|
| d.f. | F | p | |
| MDTR × VT | 4 | 39.15 | <0.01 |
| ISO × VT | 4 | 36.33 | <0.01 |
| MTWM × VT | 4 | 35.02 | <0.01 |
| MTWEQ × VT | 4 | 36.63 | <0.01 |
| MTCQ × VT | 4 | 42.16 | <0.01 |
| PWEQ × VT | 4 | 28.76 | <0.01 |
| PWAQ × VT | 4 | 28.70 | <0.01 |
| Alt × VT | 4 | 39.59 | <0.01 |
| TS × VT | 4 | 5.69 | <0.01 |
| MTWEQ × VT | 4 | 4.33 | <0.01 |
| PDM × VT | 4 | 1.86 | <0.01 |
| PS × VT | 4 | 12.6 | <0.01 |
See
环境因子同
表6 华北地区群系类型(FT)和环境因子对净谱系亲缘关系指数的交互作用
Table 6
| 交互作用 | 灌木 Shrub | ||
|---|---|---|---|
| d.f. | F | p | |
| MDTR × FT | 194 | 4.15 | <0.01 |
| ISO × FT | 194 | 4.12 | <0.01 |
| MTWM × FT | 194 | 4.12 | <0.01 |
| MTWEQ × FT | 194 | 4.09 | <0.01 |
| MTCQ × FT | 194 | 4.02 | <0.01 |
| PWEQ × FT | 194 | 3.95 | <0.01 |
| PWAQ × FT | 194 | 3.94 | <0.01 |
| Alt × FT | 194 | 4.17 | <0.01 |
| TS × FT | 194 | 2.60 | <0.01 |
| MTWEQ × FT | 194 | 2.54 | <0.01 |
| PDM × FT | 194 | 2.33 | <0.01 |
| PS × FT | 194 | 2.63 | <0.01 |
See
环境因子同
3 讨论
3.1 华北灌丛的分布及物种组成
华北地区地形、地质的特点, 为我国温带灌丛的分布奠定了基础。本研究表明, 华北地区自然灌丛群系类型丰富, 各植被型的分布表现出明显的地带性特点。落叶灌丛为该区的主要植被类型, 在各地区均有分布, 包含的群系类型最多。华北地区分布最广的灌丛群系有虎榛子灌丛、荆条灌丛和沙棘灌丛, 且这些灌丛的海拔分布范围也较广, 分别为612-1 965、70-1 497和1 907-2 645 m, 表明这些灌丛的建群种对温度和水分变化有较强的适应能力。常绿革叶灌丛类型相对较少, 主要分布于河南、山西和陕西等地的高山地带和亚高山地带, 主要类型有: 金背杜鹃(Rhododendron clementinae)灌丛、迎红杜鹃(Rhododendron mucronulatum)灌丛和头花杜鹃(Rhododendron capitatum)灌丛。受温度和降水的影响, 灌丛荒漠主要分布于内蒙古、甘肃北部、宁夏等地, 主要类型有油蒿(Artemisia ordosica)荒漠、盐爪爪(Kalidium foliatum)荒漠、珍珠猪毛菜(Salsola passerina)荒漠等。
物种组成分析结果显示, 草本层的物种丰富度高于灌木层。灌木层物种以蔷薇科最为丰富, 有101种, 反映了蔷薇科植物在该区群落物种组成中的重要地位, 也证明了蔷薇科以温带成分为主的特征(吴征镒, 1991)。已有研究分析了我国新疆(惠俊爱等, 2003)、西藏(俞德浚等, 1984)和云南(徐廷志, 2001)等地蔷薇科植物的区系特点和地理分布, 证明了蔷薇科植物的重要地位。对各地不同群落类型物种组成的研究也表明蔷薇科物种在灌丛群落物种组成中的优势地位, 如桂西灌丛(申文辉等, 2016), 云南哀牢山中山湿性常绿阔叶林(温韩东等, 2018), 青海高寒灌丛(杨路存等, 2018)。华北地区灌丛群落中的草本物种以菊科和禾本科的物种丰富度最高, 这和国内其他地区灌丛群落物种组成分析的结果相同(如:杜焰玲, 2013; 申文辉等, 2016; 刘梦, 2018; 杨路存等, 2018)。在华北地区菊科中以蒿属植物居多, 这和华北地区的温带气候特点有关(窦燕平等, 1985)。从物种的相对频度上看, 荆条、酸枣和虎榛子在我国北方地区广为分布, 是北方干旱山区阳坡、半阳坡灌丛的主要优势种或伴生种, 因此是该区相对频度最高的灌木物种。白莲蒿、大披针薹草和狗尾草等是多数灌丛草本层的主要优势种或伴生种, 因此在草本植物中频度最高。
3.2 华北灌丛群落不同垂直结构层的系统发育 结构
基于生态位理论, 生境过滤和竞争排斥是影响群落结构的两个主要过程。生境过滤使共存的物种利用相似的资源, 使得物种间的系统发育结构相对聚集; 竞争排斥则是不同的物种竞争同一资源, 使得物种的系统发育结构相对发散(牛红玉等, 2011)。本文零模型分析结果表明, 华北地区灌丛群落灌木层和草本层的系统发育结构均未超出随机阈值范围。这说明在区域尺度上, 灌丛群落物种的系统发育结构受到较大的中性过程的作用(如扩散限制)。中性过程在草本层群落构建过程中的作用和最近一项关于温带森林草本层群落系统发育结构的研究结果相同(赵鸣飞等, 2017)。这可能与草本植物的自身特点有关, 其扩散能力强, 种群具有较大的时空波动性(Donoghue, 2008)。另外, 竞争排斥与生境过滤作用相互平衡的群落也能引起系统发育结构的随机模式(Helmus et al., 2007)。
虽然单个样地中并未发现显著偏离零模型的谱系结构, 但NRI在灌木层整体表现为系统发育的汇聚而草本层表现为系统发育的发散, 说明灌丛群落的灌木层和草本层仍然有不同的群落构建规则。换言之, 生境过滤和种间竞争等确定性过程在灌木层和草本层物种构建过程中的相对重要性不同(Kembel & Hubbell, 2006)。已有研究表明, 热带森林中林下草本植物对森林物种丰富度的维持有决定性贡献, 而且由于空间和环境异质性(如地形、土壤、光照等)的影响, 草本层和木本植物的物种组成及空间分布会表现出不同, 甚至相反的构建机制(Murphy et al., 2016), 而且这种不同并不是由于生活史的不同造成的。本研究中, 灌木是群落的主要优势种和建群种, 这些物种的分布明显受大尺度气候、土壤环境变化的影响, 所以环境过滤的作用最终形成了灌木物种系统发育的聚集现象。相反, 草本层的物种组成不仅受气候和土壤的影响, 还受上层灌木物种的影响, 这种种间相互作用虽然是相对的, 但是对草本层物种组成的影响可能要强于对灌木层物种组成的影响。另外, 草本层群落的物种丰富度一般高于灌木层, 这更容易增强物种间对同一资源的竞争, 从而影响群落内物种间的亲缘关系。这些原因都有可能导致草本层物种系统发育结构的发散。
不同植被型灌丛系统发育结构的不同主要体现在灌木层, 且这种不同主要由建群种的生态特性和系统发育地位造成。常绿针叶灌丛和竹灌丛的灌木层表现为系统发育结构的发散, 而其他类型表现为系统发育结构的汇聚。竹灌丛和革叶灌丛的系统发育的发散是由于其建群种一般为禾本科和松柏科植物, 这些物种一般和伴生种亲缘关系较远, 因此会导致整个灌木层系统发育结构的离散。
3.3 气候因子对华北灌丛群落系统发育结构的 影响
大尺度上环境因子对群落系统发育结构的影响已被大量研究证实, 如温带和热带森林由于降水和光照的不同而表现出了不同的群落构建机制, 热带森林群落构建过程中扩散限制占优势, 而在温带森林的群落构建过程中环境过滤占优势(Myers et al., 2013)。而且, 同样是热带森林, 雨林和干旱森林群落构建机制变化是完全不一样的。因为在雨林中光是群落演替的主要驱动因子, 而在干旱森林中水分是主要驱动因子(Bhaskar et al., 2014)。另外海拔的不同也会影响群落的构建机制, 一般而言高海拔生境下的植物群落更容易表现为系统发育聚集, 低海拔生境的群落则为系统发育发散(Kembel & Hubbell, 2006; 赵鸣飞等, 2017)。总之, 不同的生境可能形成不同的构建机制, 但是目前并没有统一的规律来说明一种生境一定会对应一种特定的构建机制。我们的研究结果也证明, 大尺度上温度和降水的变化对华北灌丛群落的系统发育结构有显著的影响。最热季度降水量的增加会导致灌木层物种系统发育结构的发散, 而最干月降水量的增加会促进草本层物种系统发育结构的离散(表4)。这可能是因为降水量高的地方物种丰富度高, 物种间相互作用增加(代爽等, 2013), 因此灌木层和草本层物种的系统发育结构都会表现为离散, 而降水量小, 气候干旱地区的物种丰富度相对较低, 且多数为适应此气候条件的近缘物种, 因此其系统发育结构会趋于汇聚。最热月的最高温的增加会导致灌木层物种系统发育结构的汇聚, 说明温度会作为一种过滤因子促使灌木层近缘物种的共存。季节性温度变异和最湿季度平均温度的变化与草本层物种系统发育结构的变化密切相关, 表明草本层和灌木层物种系统发育结构对温度的响应模式不同。这些结果也证明了严酷生境下群落的系统发育结构更容易汇聚的假说。
本文分析结果虽然表明环境因子对灌木层和草本层的系统发育结构都有明显的影响, 但对灌木层的影响明显强于对草本层的影响。这可能和草本物种分布更容易受微生境的异质性的影响有关(Gilliam, 2007; Neufeld & Young, 2014)。同时, 我们的结果也表明, 气候因子不是影响灌丛系统发育结构的唯一因子, 因为气候因子解释了灌木层系统发育结构变异的18%, 而仅仅解释了草本层系统发育结构变异的5%。这说明, 除了气候因子, 其他的非生物因子(比如土壤营养元素含量、酸碱度)和生物因子(物种间的相互作用)也影响着群落的系统发育结构(牛红玉等, 2011)。本研究虽然没有量化种间的相互作用, 但分析了气候因子和植被型及群系类型的交互作用。结果表明, 气候因子和植被型及群落类型有明显的交互作用。理论上每一种植被型或群系类型都有自己特有的外貌特征和分布范围, 其系统发育结构的变化应是环境过滤和物种间相互作用共同驱动的结果。区域尺度上不同群落类型的地理分布主要取决于建群种对气候的响应(宋创业和郭柯, 2007), 而在局域尺度上群落的分布可能随地形、土壤及物种间相互作用而变化。气候因子和植被型及群落类型有明显的交互作用, 表明大尺度上温度和降水的变化对不同的灌丛植被型和群系类型的系统发育结构有不同的影响。因此, 气候环境对群落系统发育结构的影响可能和群落建群种的分布有关。

致谢
感谢华北地区自然植物群落资源考察组提供的原始数据, 感谢项目进行中所有参与单位的老师和同学们的辛勤付出。数据分析过程中得到嘉道理农场暨植物园张金龙老师的帮助。在此一并感谢。
参考文献
Community assembly and functional diversity along succession post-management
Research advances in plant community assembly mechanisms
植物群落构建机制研究进展
Relationship between shrub species richness and climate across central Inner Mongolia, China
内蒙古灌木群落物种丰富度与气候的关系
Aphylogenetic perspective on the distribution of plant diversity
Organization of Artemisia in Xinjiang
新疆蒿属植物的整理
Study on the Community Diversity and Regeneration Characteristic of Typical Secondary Shrub Community in Midpiece of Qinling Mountains.
秦岭中段典型灌丛群落物种多样性及更新特征研究
The ecological significance of the herbaceous layer in temperate forest ecosystems
Null model analysis of species co-occurrence patterns
Storage of carbon, nitrogen and phosphorus in temperate shrubland ecosystems across northern China
中国北方温带灌丛生态系统碳、氮、磷储量
Separating the determinants of phylogenetic community structure
Vegetation carbon storage of major shrublands in China
中国主要灌丛植被碳储量
Different relationships between temporal phylogenetic turnover and phylogenetic similarity and in two forests were detected by a new null model
Influence of environmental factors on phylogenetic structure at multiple spatial scales in an evergreen broad-leaved forest of China
不同尺度上环境因子对常绿阔叶林群落的谱系结构的影响
The district characteristics and geographical distribution of the Rosaceae in Xinjiang
新疆蔷薇科植物的区系特点和地理分布
The phylogenetic structure of a neotropical forest tree community
Composition of plant life forms of subtropical shrubland in China and its correlation with temperature and precipitation
中国亚热带灌丛植物生活型组成及其与水热因子的相关性
Leaf nitrogen and phosphorus concentration and the empirical regulations in dominant woody plants of shrublands across southern China
中国南方灌丛优势木本植物叶的氮、磷含量及其影响因素
Species biodiversity of seven typical shrub communities in the middle of Guangxi Zhuang Autonomous Region
广西中部7种典型灌丛群落的物种多样性特征
Phylogenetic community structure of subtropical forests along elevational gradients in Ailao Mountains of southwest China
沿海拔梯度变化的哀牢山亚热带森林群落系统发育结构
Opposing effects of competitive exclusion on the phylogenetic structure of communities
Phylogenetic turnover during subtropical forest succession across environmental and phylogenetic scales
Higher β-diversity observed for herbs over woody plants is driven by stronger habitat filtering in a tropical understory
Beta-diversity in temperate and tropical forests reflects dissimilar mechanisms of community assembly
Ecophysiology of the herbaceous layer in temperate deciduous forests
New progress in community assembly: Community phylogenetic structure combining evolution and ecology
群落构建研究的新进展: 进化和生态相结合的群落谱系结构研究
Community assembly: The relative importance of neutral theory and niche theory
群落构建的中性理论和生态位理论
Effects of shrub encroachment on biomass and biodiversity in the typical steppe of Inner Mongolia
内蒙古典型草原灌丛化对生物量和生物多样性的影响
Phylogenetic structure and phylogenetic diversity of angiosperm assemblages in forests along an elevational gradient in Changbaishan, China
The unified neutral theory of biodiversity and biogeography at age ten
Species composition and diversity of different shrub communities in west Guangxi
桂西不同灌丛植物群落物种组成及其多样性
Relationship between plant community and soil on the inter-dune lowland in the middle of Otingdag sand land
浑善达克沙地中部丘间低地植物群落分布与土壤环境关系
Variation in phylogenetic structure of forest communities along a human disturbance gradient in Gutianshan forest, China
不同程度人为干扰对古田山森林群落谱系结构的影响
Increasing shrub abundance in the Arctic
The influence of spatial and size scale on phylogenetic relatedness in tropical forest communities
The evolution and sources of North China’s flora
华北植物区系的演变和来源
Exploring the phylogenetic structure of ecological communities: An example for rain forest trees
Phylogenies and community ecology
Species composition and community structure of a 20 hm 2 plot of mid-mountain moist evergreen broad-leaved forest on the Mts. Ailaoshan, Yunnan Province, China
云南哀牢山中山湿性常绿阔叶林20 hm 2动态样地的物种组成与群落结构
Change in community phylogenetic structure during tropical forest succession: Evidence from New Guinea
The areal-types of Chinese genera of seed plants
中国种子植物属的分布区类型
Studies on carbon storage of shrubland ecosystems in China
中国灌丛生态系统碳储量的研究
Using phylogeny and functional traits for assessing community assembly along environmental gradients: A deterministic process driven by elevation
The floristic characteristics and geographical distribution of Rosaceae in Yunnan
云南蔷薇科植物的区系特征和地理分布
Plant community assembly rules across a subalpine grazing gradient in western Sichuan, China
放牧干扰梯度下川西亚高山植物群落的组合机理
Species diversity, biomass, and their relationship in the alpine shrubberies of Qinghai Province
青海省高寒灌丛物种多样性、生物量及其关系
Distribution of biomass in relation to environments in shrublands of temperate China
中国北方温带灌丛生物量的分布及其与环境的关系
Community structure of a 20 hm 2 lower subtropical evergreen broadleaved forest plot in Dinghushan, China
鼎湖山南亚热带常绿阔叶林20公顷样地群落特征研究
The floristic characteristics and phytogeographic distribution of the Rosaceae of Xizang
西藏蔷薇科植物的区系特点和地理分布
Three keys to the radiation of angiosperms into freezing environments
Responses of growth and litterfall production to nitrogen addition treatments from common shrublands in Mt. Dongling, Beijing, China
北京东灵山地区常见灌丛生长及凋落物生产对氮添加的响应
Estimation of biomass allocation and carbon density of Rhododendron simsii shrubland in the subtropical mountainous areas of China
中国亚热带山地杜鹃灌丛生物量分配及其碳密度估算
Shrub community characteristics and quantitative calculation of theirs biomass in southern China
我国南亚热带灌丛群落特征及生物量的定量计算
Phylogenetic structure and diversity of herbaceous communities in the conifer forests along an elevational gradient in Luya Mountain, Shanxi, China
山西芦芽山针叶林草本层群落谱系结构与多样性的海拔格局

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}