The diversity and coevolution of Rubisco, plastids, pyrenoids, and chloroplast- based CO2-concentrating mechanisms in algae
1
1998
... 当前, 表层海水中CO2的浓度为10-20 µmol·L-1 (Badger et al., 1998), 低于绝大多数浮游植物中催化固碳反应的关键酶——核酮糖-1,5-二磷酸羧化酶/加氧酶(RubisCO)的半饱和常数.因此, 大多数浮游植物依赖于耗能的CO2浓缩机制(CO2-concentrating mechanism, CCM)以提高胞内RubisCO周围CO2的浓度(Reinfelder, 2011).已有研究表明, 酸化条件下, CO2浓度升高可使CCM下调, 节省能量, 从而对部分浮游植物的生长具有不同程度的促进作用(Riebesell et al., 1993; Mackey et al., 2015).除了CO2浓度升高, 随之降低的海水pH则同样可能对浮游植物产生直接或间接的影响.例如, 近期我国学者对海洋优势固氮蓝藻束毛藻(Trichodesmium)开展的研究表明, 海水pH下降导致其固氮酶效率降低, 干扰胞内pH稳态, 影响细胞产能, 且该负效应大于CO2浓度升高所产生的正效应, 故酸化的净效应为抑制束毛藻的固氮作用(Hong et al., 2017).此外, 海水pH下降还可能改变浮游植物对铁、磷等关键营养元素的利用(Shi et al., 2010; McQuaid et al., 2018), 从而间接影响浮游植物生长.除了光合作用和固氮作用外, 海水酸化还会对颗石藻、有孔虫类、珊瑚等钙化生物的钙化作用产生负面影响(Riebesell et al., 2000; Cao & Caldeira, 2008; Hofmann et al., 2010; Kroeker et al., 2013).对于海洋软体动物、棘皮动物、甲壳动物及鱼类等生物而言, 海洋酸化不仅会降低其幼体发育速度和个体大小(Ross et al., 2011), 还会降低部分生物类群的呼吸速率, 扰乱生物体内酸碱平衡, 影响其生长与繁殖(Ishimatsu et al., 2004; Pörtner & Langenbuch, 2005).海洋酸化对生物个体的影响会进一步体现在对海洋生态系统和海洋生物地球化学循环的影响.Riebesell等(2007)通过开展中尺度生态系统(mesocosm) CO2加富实验发现, 与350 mg·kg-1相比, 1 050 mg·kg-1 CO2浓度下的浮游植物群落在12天内多消耗了39%的无机碳, 且浮游植物的C:N从6升高到8, 这一方面将有利于提高生物碳泵的效率, 但另一方面C:N的增加会降低有机物的营养价值, 影响细菌降解与浮游植物繁殖, 改变海洋生态系统的动力学. 此外, 酸化与光辐射(Suggett et al., 2013; Shi et al., 2015; Kleynhans et al., 2016)以及酸化与暖化(Pörtner, 2008; Mayor et al., 2012; Kroeker et al., 2013)之间的耦合作用, 也会从个体、种群、群落等不同水平上对海洋生态系统产生影响. ...
Interannual variability of oceanic CO2 and biogeochemical properties in the Western North Atlantic subtropical gyre
1
2001
... 对全球气候变化的研究历史最早可以追溯到19世纪.早在19世纪20年代, 科学家就已经发现了几种温室气体, 但并未引起广泛的关注.直到19世纪30年代, 科学家意识到燃烧化石燃料对气候的影响后, 便开展了一系列研究工作.之后随着科学技术的发展, 全球气候变化对生态系统影响的研究于20世纪90年代迎来了黄金期(Weart, 2003), 其中就包括对海洋生态系统影响的研究.以全球气候变化导致的海洋酸化为例, 早在1906年, 针对海洋酸碱性, Moore等(1906)就已经开展了酸碱性对海胆受精卵的生长和细胞分裂的影响研究; 1951年, Rubey总结了早期酸化的研究工作, 为后人的研究提供了理论基础; 1957年, Revelle和Suess首次提出海洋吸收人类排放的CO2会降低海洋的缓冲能力, 引起了科学界对人类活动影响海洋生态系统的重视; 1966年, Broecker和Takahashi首次报道了海水CO32-的浓度, 并提出其将随着海水CO2浓度的升高而降低; 1978年, Brewer提出利用总碱度和溶解无机碳的浓度校正海水中的CO2系统; 1993年硼同位素等古海洋示踪剂开始被拓展应用于估算海洋化学参数的变化(Spivack et al., 1993); 2001年, Bates第一次利用时间序列数据记录了北大西洋西部海水碳酸盐体系的变化,主要包括海水的CO2浓度、CO2分压以及碱度的变动; 近年来, 利用受控实验评估生物体对海水碳酸盐体系变化响应的论文大量涌现, 与此同时科学家开始通过构建数值模型来预测未来海水碳酸盐系统的变化以及海洋酸化对海洋生物地球化学循环的影响. ...
The science of hypoxia in the Northern Gulf of Mexico: a review
1
2010
... 海水富营养化会刺激浮游植物和大型藻类的生长, 增加海洋CO2的捕获量, 但过高的浮游植物生物量, 会降低光的穿透性, 影响水下生物的生长.此外, 营养盐比例的不平衡会改变浮游植物群落组成, 可能由此促进有毒藻类大量繁殖, 导致藻华的爆发(Hallegraeff, 2010; Flynn et al., 2015).有害藻类的大量繁殖不仅会导致鱼类和贝类等中毒死亡, 还会扩大海水的缺氧区(Rosenthal, 1985; Naylor et al., 2000; Rabalais et al., 2010).有研究表明, 1969-2009年期间, 由于人类活动的干扰, 大洋缺氧区的面积大小呈现递增趋势(Rabalais et al., 2010).以墨西哥北部湾缺氧区为例, 其面积从1985年的40 km2左右扩大到2015年的22 000 km2 (Sen Gupta et al., 1996; Bianchi et al., 2010; Turner & Rabalais, 2017).Neumann等(2012)的模型预测显示, 大洋季节性缺氧的持续时间将在21世纪末增加一倍.低氧会阻碍氮的硝化过程, 影响浮游植物对氮营养盐的吸收, 从而改变海洋初级生产力(Codispoti et al., 2001; Gruber, 2004), 并且不完全的硝化过程会导致温室气体N2O产量增多, 加快全球气候变暖的进程(Codispoti et al., 2001; Jin & Gruber, 2003; Nevison et al., 2003).此外, 低氧会导致有氧呼吸生物向外迁徙, 降低系统的生物多样性, 对系统物质循环与能量的流动产生影响, 破坏海洋生态系统的稳定性(Breitburg et al., 2009). ...
Hypoxia, nitrogen, and fisheries: integrating effects across local and global landscapes
1
2009
... 海水富营养化会刺激浮游植物和大型藻类的生长, 增加海洋CO2的捕获量, 但过高的浮游植物生物量, 会降低光的穿透性, 影响水下生物的生长.此外, 营养盐比例的不平衡会改变浮游植物群落组成, 可能由此促进有毒藻类大量繁殖, 导致藻华的爆发(Hallegraeff, 2010; Flynn et al., 2015).有害藻类的大量繁殖不仅会导致鱼类和贝类等中毒死亡, 还会扩大海水的缺氧区(Rosenthal, 1985; Naylor et al., 2000; Rabalais et al., 2010).有研究表明, 1969-2009年期间, 由于人类活动的干扰, 大洋缺氧区的面积大小呈现递增趋势(Rabalais et al., 2010).以墨西哥北部湾缺氧区为例, 其面积从1985年的40 km2左右扩大到2015年的22 000 km2 (Sen Gupta et al., 1996; Bianchi et al., 2010; Turner & Rabalais, 2017).Neumann等(2012)的模型预测显示, 大洋季节性缺氧的持续时间将在21世纪末增加一倍.低氧会阻碍氮的硝化过程, 影响浮游植物对氮营养盐的吸收, 从而改变海洋初级生产力(Codispoti et al., 2001; Gruber, 2004), 并且不完全的硝化过程会导致温室气体N2O产量增多, 加快全球气候变暖的进程(Codispoti et al., 2001; Jin & Gruber, 2003; Nevison et al., 2003).此外, 低氧会导致有氧呼吸生物向外迁徙, 降低系统的生物多样性, 对系统物质循环与能量的流动产生影响, 破坏海洋生态系统的稳定性(Breitburg et al., 2009). ...
Direct observation of the oceanic CO2 increase
1
1978
... 对全球气候变化的研究历史最早可以追溯到19世纪.早在19世纪20年代, 科学家就已经发现了几种温室气体, 但并未引起广泛的关注.直到19世纪30年代, 科学家意识到燃烧化石燃料对气候的影响后, 便开展了一系列研究工作.之后随着科学技术的发展, 全球气候变化对生态系统影响的研究于20世纪90年代迎来了黄金期(Weart, 2003), 其中就包括对海洋生态系统影响的研究.以全球气候变化导致的海洋酸化为例, 早在1906年, 针对海洋酸碱性, Moore等(1906)就已经开展了酸碱性对海胆受精卵的生长和细胞分裂的影响研究; 1951年, Rubey总结了早期酸化的研究工作, 为后人的研究提供了理论基础; 1957年, Revelle和Suess首次提出海洋吸收人类排放的CO2会降低海洋的缓冲能力, 引起了科学界对人类活动影响海洋生态系统的重视; 1966年, Broecker和Takahashi首次报道了海水CO32-的浓度, 并提出其将随着海水CO2浓度的升高而降低; 1978年, Brewer提出利用总碱度和溶解无机碳的浓度校正海水中的CO2系统; 1993年硼同位素等古海洋示踪剂开始被拓展应用于估算海洋化学参数的变化(Spivack et al., 1993); 2001年, Bates第一次利用时间序列数据记录了北大西洋西部海水碳酸盐体系的变化,主要包括海水的CO2浓度、CO2分压以及碱度的变动; 近年来, 利用受控实验评估生物体对海水碳酸盐体系变化响应的论文大量涌现, 与此同时科学家开始通过构建数值模型来预测未来海水碳酸盐系统的变化以及海洋酸化对海洋生物地球化学循环的影响. ...
Calcium carbonate precipitation on the Bahama Banks
1
1966
... 对全球气候变化的研究历史最早可以追溯到19世纪.早在19世纪20年代, 科学家就已经发现了几种温室气体, 但并未引起广泛的关注.直到19世纪30年代, 科学家意识到燃烧化石燃料对气候的影响后, 便开展了一系列研究工作.之后随着科学技术的发展, 全球气候变化对生态系统影响的研究于20世纪90年代迎来了黄金期(Weart, 2003), 其中就包括对海洋生态系统影响的研究.以全球气候变化导致的海洋酸化为例, 早在1906年, 针对海洋酸碱性, Moore等(1906)就已经开展了酸碱性对海胆受精卵的生长和细胞分裂的影响研究; 1951年, Rubey总结了早期酸化的研究工作, 为后人的研究提供了理论基础; 1957年, Revelle和Suess首次提出海洋吸收人类排放的CO2会降低海洋的缓冲能力, 引起了科学界对人类活动影响海洋生态系统的重视; 1966年, Broecker和Takahashi首次报道了海水CO32-的浓度, 并提出其将随着海水CO2浓度的升高而降低; 1978年, Brewer提出利用总碱度和溶解无机碳的浓度校正海水中的CO2系统; 1993年硼同位素等古海洋示踪剂开始被拓展应用于估算海洋化学参数的变化(Spivack et al., 1993); 2001年, Bates第一次利用时间序列数据记录了北大西洋西部海水碳酸盐体系的变化,主要包括海水的CO2浓度、CO2分压以及碱度的变动; 近年来, 利用受控实验评估生物体对海水碳酸盐体系变化响应的论文大量涌现, 与此同时科学家开始通过构建数值模型来预测未来海水碳酸盐系统的变化以及海洋酸化对海洋生物地球化学循环的影响. ...
Inclusion of facilitation into ecological theory
1
2003
... 暖化的海水对生物的影响因种而异.对于大多数浮游植物而言, 在适温范围内, 海水暖化会促进其生长(Eppley, 1972), 但不同浮游植物对温度变化的响应程度不同, 因此全球暖化下浮游植物的生态位将发生改变(Xiao et al., 2018).然而, 对于大多数海洋动物, 暖化将在增加其体内代谢活性的同时加快其能量的消耗, 从而可能导致海洋生物自身生长与繁殖能力的下降(Pörtner & Knust, 2007).这些个体生理生化过程的变化会进一步改变种群的大小、时空分布格局与营养级水平, 最终引起群落结构与功能的改变(Bruno et al., 2003; Hyrenbach & Veit, 2003).从海洋生态系统的层面上来看, 全球变暖会改变海洋的初级生产力, 影响生态系统的稳定性.近期我国科学家采用模型估算发现, 1948-2007年, 全球气候变暖导致海洋水体层化加剧, 使得北大西洋、北太平洋和印度洋初级生产力分别降低了40%、24%和25% (Wang et al., 2017).倘若暖化现象一直持续, 到2100年, 海洋初级生产力将可能进一步减少20% (Steinacher et al., 2010).此外, 已有研究表明, 海水温度升高1 ℃, 珊瑚即会出现白化现象(Hoegh- Guldberg et al., 2007), 这会导致珊瑚礁生态系统多样性降低, 严重时会引发珊瑚礁系统的崩溃(Donner, 2009).海水暖化除了通过温度增加而对海洋生态系统产生直接影响外, 还会造成海洋层化加剧、缺氧区扩增等, 从而间接地对海洋生态系统产生一系列影响(Keeling et al., 2010; Wang et al., 2017). ...
Anthropogenic carbon and ocean pH
1
2003
... 由于空气和海水之间持续的气体交换, 工业革命以来大气CO2浓度的急剧上升引起表层海水碳酸盐系统发生显著改变, 包括海水中的CO2、H+ (pH下降)和HCO3-浓度的升高以及CO32-浓度的下降, 即“海洋酸化” (Caldeira & Wickett, 2003; Doney et al., 2009).迄今为止, 表层海水的pH较工业革命之前(约8.21)已经降低了约0.1个单位, 预计到2100年其将继续下降到约7.7 (Doney et al., 2009).该酸化速率高于过去3亿年间的任何一个时期(Hönisch et al., 2012), 必将对海洋生态系统产生影响.因此, 海洋酸化不仅引起科学界的广泛关注, 政府间海洋学委员会、海洋研究科学委员会等国际组织也已将其列为国际合作的重点领域. ...
Atmospheric CO2 stabilization and ocean acidification
1
2008
... 当前, 表层海水中CO2的浓度为10-20 µmol·L-1 (Badger et al., 1998), 低于绝大多数浮游植物中催化固碳反应的关键酶——核酮糖-1,5-二磷酸羧化酶/加氧酶(RubisCO)的半饱和常数.因此, 大多数浮游植物依赖于耗能的CO2浓缩机制(CO2-concentrating mechanism, CCM)以提高胞内RubisCO周围CO2的浓度(Reinfelder, 2011).已有研究表明, 酸化条件下, CO2浓度升高可使CCM下调, 节省能量, 从而对部分浮游植物的生长具有不同程度的促进作用(Riebesell et al., 1993; Mackey et al., 2015).除了CO2浓度升高, 随之降低的海水pH则同样可能对浮游植物产生直接或间接的影响.例如, 近期我国学者对海洋优势固氮蓝藻束毛藻(Trichodesmium)开展的研究表明, 海水pH下降导致其固氮酶效率降低, 干扰胞内pH稳态, 影响细胞产能, 且该负效应大于CO2浓度升高所产生的正效应, 故酸化的净效应为抑制束毛藻的固氮作用(Hong et al., 2017).此外, 海水pH下降还可能改变浮游植物对铁、磷等关键营养元素的利用(Shi et al., 2010; McQuaid et al., 2018), 从而间接影响浮游植物生长.除了光合作用和固氮作用外, 海水酸化还会对颗石藻、有孔虫类、珊瑚等钙化生物的钙化作用产生负面影响(Riebesell et al., 2000; Cao & Caldeira, 2008; Hofmann et al., 2010; Kroeker et al., 2013).对于海洋软体动物、棘皮动物、甲壳动物及鱼类等生物而言, 海洋酸化不仅会降低其幼体发育速度和个体大小(Ross et al., 2011), 还会降低部分生物类群的呼吸速率, 扰乱生物体内酸碱平衡, 影响其生长与繁殖(Ishimatsu et al., 2004; Pörtner & Langenbuch, 2005).海洋酸化对生物个体的影响会进一步体现在对海洋生态系统和海洋生物地球化学循环的影响.Riebesell等(2007)通过开展中尺度生态系统(mesocosm) CO2加富实验发现, 与350 mg·kg-1相比, 1 050 mg·kg-1 CO2浓度下的浮游植物群落在12天内多消耗了39%的无机碳, 且浮游植物的C:N从6升高到8, 这一方面将有利于提高生物碳泵的效率, 但另一方面C:N的增加会降低有机物的营养价值, 影响细菌降解与浮游植物繁殖, 改变海洋生态系统的动力学. 此外, 酸化与光辐射(Suggett et al., 2013; Shi et al., 2015; Kleynhans et al., 2016)以及酸化与暖化(Pörtner, 2008; Mayor et al., 2012; Kroeker et al., 2013)之间的耦合作用, 也会从个体、种群、群落等不同水平上对海洋生态系统产生影响. ...
Our evolving conceptual model of the coastal eutrophication problem
1
2001
... 由于人类社会的快速发展, 污水排放的增多对海洋生态系统产生了一系列的影响.以工农业污水排放为例, 大量的氮磷营养盐向海水排放, 引起了沿海水域的富营养化现象, 导致生物营养结构变化、藻华频发以及低氧区的形成(Diaz & Rosenberg, 2008).这些变化相互作用, 共同影响着海洋生态系统的结构与功能(Cloern, 2001). ...
The oceanic fixed nitrogen and nitrous oxide budgets: moving targets as we enter the anthropocene?
2
2001
... 海水富营养化会刺激浮游植物和大型藻类的生长, 增加海洋CO2的捕获量, 但过高的浮游植物生物量, 会降低光的穿透性, 影响水下生物的生长.此外, 营养盐比例的不平衡会改变浮游植物群落组成, 可能由此促进有毒藻类大量繁殖, 导致藻华的爆发(Hallegraeff, 2010; Flynn et al., 2015).有害藻类的大量繁殖不仅会导致鱼类和贝类等中毒死亡, 还会扩大海水的缺氧区(Rosenthal, 1985; Naylor et al., 2000; Rabalais et al., 2010).有研究表明, 1969-2009年期间, 由于人类活动的干扰, 大洋缺氧区的面积大小呈现递增趋势(Rabalais et al., 2010).以墨西哥北部湾缺氧区为例, 其面积从1985年的40 km2左右扩大到2015年的22 000 km2 (Sen Gupta et al., 1996; Bianchi et al., 2010; Turner & Rabalais, 2017).Neumann等(2012)的模型预测显示, 大洋季节性缺氧的持续时间将在21世纪末增加一倍.低氧会阻碍氮的硝化过程, 影响浮游植物对氮营养盐的吸收, 从而改变海洋初级生产力(Codispoti et al., 2001; Gruber, 2004), 并且不完全的硝化过程会导致温室气体N2O产量增多, 加快全球气候变暖的进程(Codispoti et al., 2001; Jin & Gruber, 2003; Nevison et al., 2003).此外, 低氧会导致有氧呼吸生物向外迁徙, 降低系统的生物多样性, 对系统物质循环与能量的流动产生影响, 破坏海洋生态系统的稳定性(Breitburg et al., 2009). ...
... O产量增多, 加快全球气候变暖的进程(Codispoti et al., 2001; Jin & Gruber, 2003; Nevison et al., 2003).此外, 低氧会导致有氧呼吸生物向外迁徙, 降低系统的生物多样性, 对系统物质循环与能量的流动产生影响, 破坏海洋生态系统的稳定性(Breitburg et al., 2009). ...
Spreading dead zones and consequences for marine ecosystems
1
2008
... 由于人类社会的快速发展, 污水排放的增多对海洋生态系统产生了一系列的影响.以工农业污水排放为例, 大量的氮磷营养盐向海水排放, 引起了沿海水域的富营养化现象, 导致生物营养结构变化、藻华频发以及低氧区的形成(Diaz & Rosenberg, 2008).这些变化相互作用, 共同影响着海洋生态系统的结构与功能(Cloern, 2001). ...
Ocean acidification: the other CO2 problem
3
2009
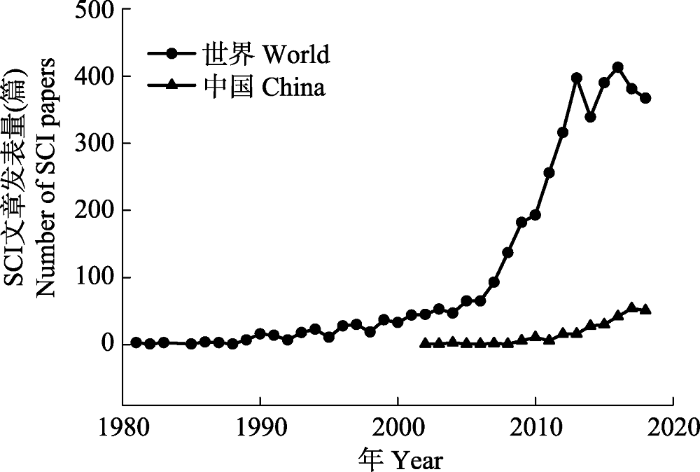
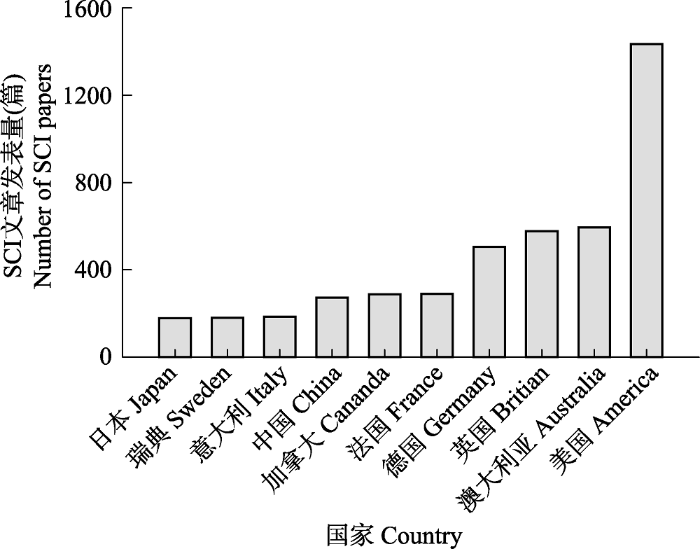
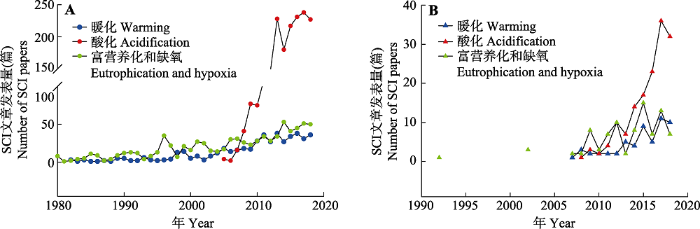
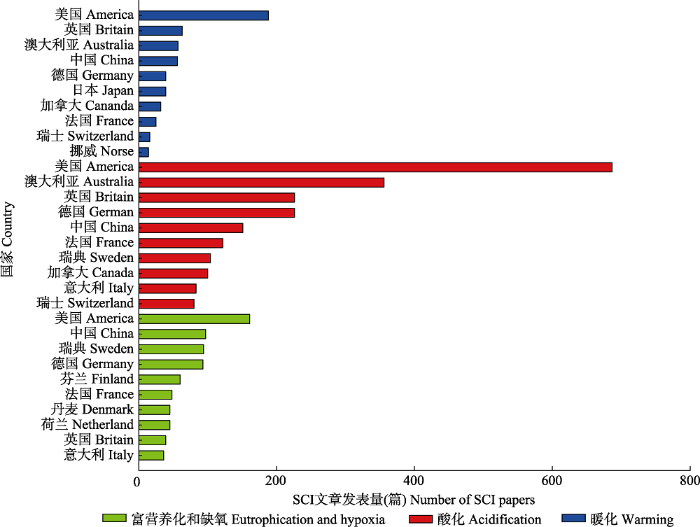
... 海洋生态系统是由海洋生物群落及其环境相互作用所构成的自然系统, 其服务功能及生态价值既是地球生命系统的重要支撑组成, 也是社会与环境可持续发展的一项基本要素.海洋生态系统的稳定主要依托于海洋中复杂的生物过程, 而这些过程往往易受海洋环境变化的影响.近半个世纪以来, 海洋和陆地一样面临着前所未有的生态压力, 不断加剧的人类活动正对海洋生态系统产生深远的影响, 例如, 大气二氧化碳(CO2)浓度的上升就是一个关键的问题.CO2浓度的上升会导致海洋暖化和酸化(Doney et al., 2009).暖化会引起海水层化加剧、海洋缺氧区扩增以及环流模式改变等(Keeling et al., 2010), 而酸化则会导致海水碳酸盐系统改变等化学变化, 进而影响生物的生理活动以及群落的结构功能等, 最终对海洋生态系统产生一系列的影响.此外, 人类活动干扰导致的海水富营养化以及由此引发的赤潮及海洋缺氧等问题也给海洋生态系统带来了严峻的挑战.迄今为止, 围绕气候变化和人类活动对海洋生态系统的影响, 国际上开展了大量的研究工作并取得了一定的进展.本文将通过文献计量的方法分析国内外的研究现状, 简要回顾该研究领域的发展简史, 聚焦海洋暖化、海洋酸化和富营养化与缺氧这三个核心研究方向, 重点阐述其对海洋生态系统初级生产的关键过程的影响, 总结已取得的重要进展以及存在的主要问题, 最后提出前沿展望. ...
... 由于空气和海水之间持续的气体交换, 工业革命以来大气CO2浓度的急剧上升引起表层海水碳酸盐系统发生显著改变, 包括海水中的CO2、H+ (pH下降)和HCO3-浓度的升高以及CO32-浓度的下降, 即“海洋酸化” (Caldeira & Wickett, 2003; Doney et al., 2009).迄今为止, 表层海水的pH较工业革命之前(约8.21)已经降低了约0.1个单位, 预计到2100年其将继续下降到约7.7 (Doney et al., 2009).该酸化速率高于过去3亿年间的任何一个时期(Hönisch et al., 2012), 必将对海洋生态系统产生影响.因此, 海洋酸化不仅引起科学界的广泛关注, 政府间海洋学委员会、海洋研究科学委员会等国际组织也已将其列为国际合作的重点领域. ...
... ).迄今为止, 表层海水的pH较工业革命之前(约8.21)已经降低了约0.1个单位, 预计到2100年其将继续下降到约7.7 (Doney et al., 2009).该酸化速率高于过去3亿年间的任何一个时期(Hönisch et al., 2012), 必将对海洋生态系统产生影响.因此, 海洋酸化不仅引起科学界的广泛关注, 政府间海洋学委员会、海洋研究科学委员会等国际组织也已将其列为国际合作的重点领域. ...
Coping with commitment: projected thermal stress on coral reefs under different future scenarios
1
2009
... 暖化的海水对生物的影响因种而异.对于大多数浮游植物而言, 在适温范围内, 海水暖化会促进其生长(Eppley, 1972), 但不同浮游植物对温度变化的响应程度不同, 因此全球暖化下浮游植物的生态位将发生改变(Xiao et al., 2018).然而, 对于大多数海洋动物, 暖化将在增加其体内代谢活性的同时加快其能量的消耗, 从而可能导致海洋生物自身生长与繁殖能力的下降(Pörtner & Knust, 2007).这些个体生理生化过程的变化会进一步改变种群的大小、时空分布格局与营养级水平, 最终引起群落结构与功能的改变(Bruno et al., 2003; Hyrenbach & Veit, 2003).从海洋生态系统的层面上来看, 全球变暖会改变海洋的初级生产力, 影响生态系统的稳定性.近期我国科学家采用模型估算发现, 1948-2007年, 全球气候变暖导致海洋水体层化加剧, 使得北大西洋、北太平洋和印度洋初级生产力分别降低了40%、24%和25% (Wang et al., 2017).倘若暖化现象一直持续, 到2100年, 海洋初级生产力将可能进一步减少20% (Steinacher et al., 2010).此外, 已有研究表明, 海水温度升高1 ℃, 珊瑚即会出现白化现象(Hoegh- Guldberg et al., 2007), 这会导致珊瑚礁生态系统多样性降低, 严重时会引发珊瑚礁系统的崩溃(Donner, 2009).海水暖化除了通过温度增加而对海洋生态系统产生直接影响外, 还会造成海洋层化加剧、缺氧区扩增等, 从而间接地对海洋生态系统产生一系列影响(Keeling et al., 2010; Wang et al., 2017). ...
Temperature and phytoplankton growth in the sea
1
1972
... 暖化的海水对生物的影响因种而异.对于大多数浮游植物而言, 在适温范围内, 海水暖化会促进其生长(Eppley, 1972), 但不同浮游植物对温度变化的响应程度不同, 因此全球暖化下浮游植物的生态位将发生改变(Xiao et al., 2018).然而, 对于大多数海洋动物, 暖化将在增加其体内代谢活性的同时加快其能量的消耗, 从而可能导致海洋生物自身生长与繁殖能力的下降(Pörtner & Knust, 2007).这些个体生理生化过程的变化会进一步改变种群的大小、时空分布格局与营养级水平, 最终引起群落结构与功能的改变(Bruno et al., 2003; Hyrenbach & Veit, 2003).从海洋生态系统的层面上来看, 全球变暖会改变海洋的初级生产力, 影响生态系统的稳定性.近期我国科学家采用模型估算发现, 1948-2007年, 全球气候变暖导致海洋水体层化加剧, 使得北大西洋、北太平洋和印度洋初级生产力分别降低了40%、24%和25% (Wang et al., 2017).倘若暖化现象一直持续, 到2100年, 海洋初级生产力将可能进一步减少20% (Steinacher et al., 2010).此外, 已有研究表明, 海水温度升高1 ℃, 珊瑚即会出现白化现象(Hoegh- Guldberg et al., 2007), 这会导致珊瑚礁生态系统多样性降低, 严重时会引发珊瑚礁系统的崩溃(Donner, 2009).海水暖化除了通过温度增加而对海洋生态系统产生直接影响外, 还会造成海洋层化加剧、缺氧区扩增等, 从而间接地对海洋生态系统产生一系列影响(Keeling et al., 2010; Wang et al., 2017). ...
Ocean acidification with (de)eutrophication will alter future phytoplankton growth and succession
1
2015
... 海水富营养化会刺激浮游植物和大型藻类的生长, 增加海洋CO2的捕获量, 但过高的浮游植物生物量, 会降低光的穿透性, 影响水下生物的生长.此外, 营养盐比例的不平衡会改变浮游植物群落组成, 可能由此促进有毒藻类大量繁殖, 导致藻华的爆发(Hallegraeff, 2010; Flynn et al., 2015).有害藻类的大量繁殖不仅会导致鱼类和贝类等中毒死亡, 还会扩大海水的缺氧区(Rosenthal, 1985; Naylor et al., 2000; Rabalais et al., 2010).有研究表明, 1969-2009年期间, 由于人类活动的干扰, 大洋缺氧区的面积大小呈现递增趋势(Rabalais et al., 2010).以墨西哥北部湾缺氧区为例, 其面积从1985年的40 km2左右扩大到2015年的22 000 km2 (Sen Gupta et al., 1996; Bianchi et al., 2010; Turner & Rabalais, 2017).Neumann等(2012)的模型预测显示, 大洋季节性缺氧的持续时间将在21世纪末增加一倍.低氧会阻碍氮的硝化过程, 影响浮游植物对氮营养盐的吸收, 从而改变海洋初级生产力(Codispoti et al., 2001; Gruber, 2004), 并且不完全的硝化过程会导致温室气体N2O产量增多, 加快全球气候变暖的进程(Codispoti et al., 2001; Jin & Gruber, 2003; Nevison et al., 2003).此外, 低氧会导致有氧呼吸生物向外迁徙, 降低系统的生物多样性, 对系统物质循环与能量的流动产生影响, 破坏海洋生态系统的稳定性(Breitburg et al., 2009). ...
The Ocean Carbon Cycle and Climate
1
2004
... 海水富营养化会刺激浮游植物和大型藻类的生长, 增加海洋CO2的捕获量, 但过高的浮游植物生物量, 会降低光的穿透性, 影响水下生物的生长.此外, 营养盐比例的不平衡会改变浮游植物群落组成, 可能由此促进有毒藻类大量繁殖, 导致藻华的爆发(Hallegraeff, 2010; Flynn et al., 2015).有害藻类的大量繁殖不仅会导致鱼类和贝类等中毒死亡, 还会扩大海水的缺氧区(Rosenthal, 1985; Naylor et al., 2000; Rabalais et al., 2010).有研究表明, 1969-2009年期间, 由于人类活动的干扰, 大洋缺氧区的面积大小呈现递增趋势(Rabalais et al., 2010).以墨西哥北部湾缺氧区为例, 其面积从1985年的40 km2左右扩大到2015年的22 000 km2 (Sen Gupta et al., 1996; Bianchi et al., 2010; Turner & Rabalais, 2017).Neumann等(2012)的模型预测显示, 大洋季节性缺氧的持续时间将在21世纪末增加一倍.低氧会阻碍氮的硝化过程, 影响浮游植物对氮营养盐的吸收, 从而改变海洋初级生产力(Codispoti et al., 2001; Gruber, 2004), 并且不完全的硝化过程会导致温室气体N2O产量增多, 加快全球气候变暖的进程(Codispoti et al., 2001; Jin & Gruber, 2003; Nevison et al., 2003).此外, 低氧会导致有氧呼吸生物向外迁徙, 降低系统的生物多样性, 对系统物质循环与能量的流动产生影响, 破坏海洋生态系统的稳定性(Breitburg et al., 2009). ...
Ocean climate change, phytoplankton community responses, and harmful algal blooms: a formidable predictive challenge
1
2010
... 海水富营养化会刺激浮游植物和大型藻类的生长, 增加海洋CO2的捕获量, 但过高的浮游植物生物量, 会降低光的穿透性, 影响水下生物的生长.此外, 营养盐比例的不平衡会改变浮游植物群落组成, 可能由此促进有毒藻类大量繁殖, 导致藻华的爆发(Hallegraeff, 2010; Flynn et al., 2015).有害藻类的大量繁殖不仅会导致鱼类和贝类等中毒死亡, 还会扩大海水的缺氧区(Rosenthal, 1985; Naylor et al., 2000; Rabalais et al., 2010).有研究表明, 1969-2009年期间, 由于人类活动的干扰, 大洋缺氧区的面积大小呈现递增趋势(Rabalais et al., 2010).以墨西哥北部湾缺氧区为例, 其面积从1985年的40 km2左右扩大到2015年的22 000 km2 (Sen Gupta et al., 1996; Bianchi et al., 2010; Turner & Rabalais, 2017).Neumann等(2012)的模型预测显示, 大洋季节性缺氧的持续时间将在21世纪末增加一倍.低氧会阻碍氮的硝化过程, 影响浮游植物对氮营养盐的吸收, 从而改变海洋初级生产力(Codispoti et al., 2001; Gruber, 2004), 并且不完全的硝化过程会导致温室气体N2O产量增多, 加快全球气候变暖的进程(Codispoti et al., 2001; Jin & Gruber, 2003; Nevison et al., 2003).此外, 低氧会导致有氧呼吸生物向外迁徙, 降低系统的生物多样性, 对系统物质循环与能量的流动产生影响, 破坏海洋生态系统的稳定性(Breitburg et al., 2009). ...
The impact of climate change on the world’s marine ecosystems
1
2010
... 工业革命以来, 人类活动导致大气中CO2浓度急剧上升, 大气温度升高了近0.6 ℃, 海水表层的平均温度在一个世纪内也相应地上升了约0.6 ℃ (Hoegh-Guldberg & Bruno, 2010).由于海洋生物的大多数生理生化过程都与温度密切相关, 因此海水暖化将直接影响到海洋生态系统和生物地球化学过程. ...
Coral reefs under rapid climate change and ocean acidification
1
2007
... 暖化的海水对生物的影响因种而异.对于大多数浮游植物而言, 在适温范围内, 海水暖化会促进其生长(Eppley, 1972), 但不同浮游植物对温度变化的响应程度不同, 因此全球暖化下浮游植物的生态位将发生改变(Xiao et al., 2018).然而, 对于大多数海洋动物, 暖化将在增加其体内代谢活性的同时加快其能量的消耗, 从而可能导致海洋生物自身生长与繁殖能力的下降(Pörtner & Knust, 2007).这些个体生理生化过程的变化会进一步改变种群的大小、时空分布格局与营养级水平, 最终引起群落结构与功能的改变(Bruno et al., 2003; Hyrenbach & Veit, 2003).从海洋生态系统的层面上来看, 全球变暖会改变海洋的初级生产力, 影响生态系统的稳定性.近期我国科学家采用模型估算发现, 1948-2007年, 全球气候变暖导致海洋水体层化加剧, 使得北大西洋、北太平洋和印度洋初级生产力分别降低了40%、24%和25% (Wang et al., 2017).倘若暖化现象一直持续, 到2100年, 海洋初级生产力将可能进一步减少20% (Steinacher et al., 2010).此外, 已有研究表明, 海水温度升高1 ℃, 珊瑚即会出现白化现象(Hoegh- Guldberg et al., 2007), 这会导致珊瑚礁生态系统多样性降低, 严重时会引发珊瑚礁系统的崩溃(Donner, 2009).海水暖化除了通过温度增加而对海洋生态系统产生直接影响外, 还会造成海洋层化加剧、缺氧区扩增等, 从而间接地对海洋生态系统产生一系列影响(Keeling et al., 2010; Wang et al., 2017). ...
The effect of ocean acidification on calcifying organisms in marine ecosystems: an organism-to-ecosystem perspective
1
2010
... 当前, 表层海水中CO2的浓度为10-20 µmol·L-1 (Badger et al., 1998), 低于绝大多数浮游植物中催化固碳反应的关键酶——核酮糖-1,5-二磷酸羧化酶/加氧酶(RubisCO)的半饱和常数.因此, 大多数浮游植物依赖于耗能的CO2浓缩机制(CO2-concentrating mechanism, CCM)以提高胞内RubisCO周围CO2的浓度(Reinfelder, 2011).已有研究表明, 酸化条件下, CO2浓度升高可使CCM下调, 节省能量, 从而对部分浮游植物的生长具有不同程度的促进作用(Riebesell et al., 1993; Mackey et al., 2015).除了CO2浓度升高, 随之降低的海水pH则同样可能对浮游植物产生直接或间接的影响.例如, 近期我国学者对海洋优势固氮蓝藻束毛藻(Trichodesmium)开展的研究表明, 海水pH下降导致其固氮酶效率降低, 干扰胞内pH稳态, 影响细胞产能, 且该负效应大于CO2浓度升高所产生的正效应, 故酸化的净效应为抑制束毛藻的固氮作用(Hong et al., 2017).此外, 海水pH下降还可能改变浮游植物对铁、磷等关键营养元素的利用(Shi et al., 2010; McQuaid et al., 2018), 从而间接影响浮游植物生长.除了光合作用和固氮作用外, 海水酸化还会对颗石藻、有孔虫类、珊瑚等钙化生物的钙化作用产生负面影响(Riebesell et al., 2000; Cao & Caldeira, 2008; Hofmann et al., 2010; Kroeker et al., 2013).对于海洋软体动物、棘皮动物、甲壳动物及鱼类等生物而言, 海洋酸化不仅会降低其幼体发育速度和个体大小(Ross et al., 2011), 还会降低部分生物类群的呼吸速率, 扰乱生物体内酸碱平衡, 影响其生长与繁殖(Ishimatsu et al., 2004; Pörtner & Langenbuch, 2005).海洋酸化对生物个体的影响会进一步体现在对海洋生态系统和海洋生物地球化学循环的影响.Riebesell等(2007)通过开展中尺度生态系统(mesocosm) CO2加富实验发现, 与350 mg·kg-1相比, 1 050 mg·kg-1 CO2浓度下的浮游植物群落在12天内多消耗了39%的无机碳, 且浮游植物的C:N从6升高到8, 这一方面将有利于提高生物碳泵的效率, 但另一方面C:N的增加会降低有机物的营养价值, 影响细菌降解与浮游植物繁殖, 改变海洋生态系统的动力学. 此外, 酸化与光辐射(Suggett et al., 2013; Shi et al., 2015; Kleynhans et al., 2016)以及酸化与暖化(Pörtner, 2008; Mayor et al., 2012; Kroeker et al., 2013)之间的耦合作用, 也会从个体、种群、群落等不同水平上对海洋生态系统产生影响. ...
The complex effects of ocean acidification on the prominent N2-fixing cyanobacterium Trichodesmium
1
2017
... 当前, 表层海水中CO2的浓度为10-20 µmol·L-1 (Badger et al., 1998), 低于绝大多数浮游植物中催化固碳反应的关键酶——核酮糖-1,5-二磷酸羧化酶/加氧酶(RubisCO)的半饱和常数.因此, 大多数浮游植物依赖于耗能的CO2浓缩机制(CO2-concentrating mechanism, CCM)以提高胞内RubisCO周围CO2的浓度(Reinfelder, 2011).已有研究表明, 酸化条件下, CO2浓度升高可使CCM下调, 节省能量, 从而对部分浮游植物的生长具有不同程度的促进作用(Riebesell et al., 1993; Mackey et al., 2015).除了CO2浓度升高, 随之降低的海水pH则同样可能对浮游植物产生直接或间接的影响.例如, 近期我国学者对海洋优势固氮蓝藻束毛藻(Trichodesmium)开展的研究表明, 海水pH下降导致其固氮酶效率降低, 干扰胞内pH稳态, 影响细胞产能, 且该负效应大于CO2浓度升高所产生的正效应, 故酸化的净效应为抑制束毛藻的固氮作用(Hong et al., 2017).此外, 海水pH下降还可能改变浮游植物对铁、磷等关键营养元素的利用(Shi et al., 2010; McQuaid et al., 2018), 从而间接影响浮游植物生长.除了光合作用和固氮作用外, 海水酸化还会对颗石藻、有孔虫类、珊瑚等钙化生物的钙化作用产生负面影响(Riebesell et al., 2000; Cao & Caldeira, 2008; Hofmann et al., 2010; Kroeker et al., 2013).对于海洋软体动物、棘皮动物、甲壳动物及鱼类等生物而言, 海洋酸化不仅会降低其幼体发育速度和个体大小(Ross et al., 2011), 还会降低部分生物类群的呼吸速率, 扰乱生物体内酸碱平衡, 影响其生长与繁殖(Ishimatsu et al., 2004; Pörtner & Langenbuch, 2005).海洋酸化对生物个体的影响会进一步体现在对海洋生态系统和海洋生物地球化学循环的影响.Riebesell等(2007)通过开展中尺度生态系统(mesocosm) CO2加富实验发现, 与350 mg·kg-1相比, 1 050 mg·kg-1 CO2浓度下的浮游植物群落在12天内多消耗了39%的无机碳, 且浮游植物的C:N从6升高到8, 这一方面将有利于提高生物碳泵的效率, 但另一方面C:N的增加会降低有机物的营养价值, 影响细菌降解与浮游植物繁殖, 改变海洋生态系统的动力学. 此外, 酸化与光辐射(Suggett et al., 2013; Shi et al., 2015; Kleynhans et al., 2016)以及酸化与暖化(Pörtner, 2008; Mayor et al., 2012; Kroeker et al., 2013)之间的耦合作用, 也会从个体、种群、群落等不同水平上对海洋生态系统产生影响. ...
The geological record of ocean acidification
1
2012
... 由于空气和海水之间持续的气体交换, 工业革命以来大气CO2浓度的急剧上升引起表层海水碳酸盐系统发生显著改变, 包括海水中的CO2、H+ (pH下降)和HCO3-浓度的升高以及CO32-浓度的下降, 即“海洋酸化” (Caldeira & Wickett, 2003; Doney et al., 2009).迄今为止, 表层海水的pH较工业革命之前(约8.21)已经降低了约0.1个单位, 预计到2100年其将继续下降到约7.7 (Doney et al., 2009).该酸化速率高于过去3亿年间的任何一个时期(Hönisch et al., 2012), 必将对海洋生态系统产生影响.因此, 海洋酸化不仅引起科学界的广泛关注, 政府间海洋学委员会、海洋研究科学委员会等国际组织也已将其列为国际合作的重点领域. ...
Marine phytoplankton and the changing ocean iron cycle
1
2016
... 目前国际上对海洋酸化的研究取得了一定的进展, 尤其是近年来越来越多的研究开始关注温度、光照、营养盐条件等其他环境因子的变化与海洋酸化耦合的影响(Shi et al., 2015; Hutchins & Boyd, 2016).然而, 以下几个关键科学问题仍然有待各个国家、学术组织之中的科学家通过加强合作交流以及在个体-种群-群落-生态系统等尺度上开展整合研究而予以阐明. ...
Ocean warming and seabird communities of the southern California Current System (1987-98): response at multiple temporal scales
1
2003
... 暖化的海水对生物的影响因种而异.对于大多数浮游植物而言, 在适温范围内, 海水暖化会促进其生长(Eppley, 1972), 但不同浮游植物对温度变化的响应程度不同, 因此全球暖化下浮游植物的生态位将发生改变(Xiao et al., 2018).然而, 对于大多数海洋动物, 暖化将在增加其体内代谢活性的同时加快其能量的消耗, 从而可能导致海洋生物自身生长与繁殖能力的下降(Pörtner & Knust, 2007).这些个体生理生化过程的变化会进一步改变种群的大小、时空分布格局与营养级水平, 最终引起群落结构与功能的改变(Bruno et al., 2003; Hyrenbach & Veit, 2003).从海洋生态系统的层面上来看, 全球变暖会改变海洋的初级生产力, 影响生态系统的稳定性.近期我国科学家采用模型估算发现, 1948-2007年, 全球气候变暖导致海洋水体层化加剧, 使得北大西洋、北太平洋和印度洋初级生产力分别降低了40%、24%和25% (Wang et al., 2017).倘若暖化现象一直持续, 到2100年, 海洋初级生产力将可能进一步减少20% (Steinacher et al., 2010).此外, 已有研究表明, 海水温度升高1 ℃, 珊瑚即会出现白化现象(Hoegh- Guldberg et al., 2007), 这会导致珊瑚礁生态系统多样性降低, 严重时会引发珊瑚礁系统的崩溃(Donner, 2009).海水暖化除了通过温度增加而对海洋生态系统产生直接影响外, 还会造成海洋层化加剧、缺氧区扩增等, 从而间接地对海洋生态系统产生一系列影响(Keeling et al., 2010; Wang et al., 2017). ...
Effects of CO2 on marine fish: larvae and adults
1
2004
... 当前, 表层海水中CO2的浓度为10-20 µmol·L-1 (Badger et al., 1998), 低于绝大多数浮游植物中催化固碳反应的关键酶——核酮糖-1,5-二磷酸羧化酶/加氧酶(RubisCO)的半饱和常数.因此, 大多数浮游植物依赖于耗能的CO2浓缩机制(CO2-concentrating mechanism, CCM)以提高胞内RubisCO周围CO2的浓度(Reinfelder, 2011).已有研究表明, 酸化条件下, CO2浓度升高可使CCM下调, 节省能量, 从而对部分浮游植物的生长具有不同程度的促进作用(Riebesell et al., 1993; Mackey et al., 2015).除了CO2浓度升高, 随之降低的海水pH则同样可能对浮游植物产生直接或间接的影响.例如, 近期我国学者对海洋优势固氮蓝藻束毛藻(Trichodesmium)开展的研究表明, 海水pH下降导致其固氮酶效率降低, 干扰胞内pH稳态, 影响细胞产能, 且该负效应大于CO2浓度升高所产生的正效应, 故酸化的净效应为抑制束毛藻的固氮作用(Hong et al., 2017).此外, 海水pH下降还可能改变浮游植物对铁、磷等关键营养元素的利用(Shi et al., 2010; McQuaid et al., 2018), 从而间接影响浮游植物生长.除了光合作用和固氮作用外, 海水酸化还会对颗石藻、有孔虫类、珊瑚等钙化生物的钙化作用产生负面影响(Riebesell et al., 2000; Cao & Caldeira, 2008; Hofmann et al., 2010; Kroeker et al., 2013).对于海洋软体动物、棘皮动物、甲壳动物及鱼类等生物而言, 海洋酸化不仅会降低其幼体发育速度和个体大小(Ross et al., 2011), 还会降低部分生物类群的呼吸速率, 扰乱生物体内酸碱平衡, 影响其生长与繁殖(Ishimatsu et al., 2004; Pörtner & Langenbuch, 2005).海洋酸化对生物个体的影响会进一步体现在对海洋生态系统和海洋生物地球化学循环的影响.Riebesell等(2007)通过开展中尺度生态系统(mesocosm) CO2加富实验发现, 与350 mg·kg-1相比, 1 050 mg·kg-1 CO2浓度下的浮游植物群落在12天内多消耗了39%的无机碳, 且浮游植物的C:N从6升高到8, 这一方面将有利于提高生物碳泵的效率, 但另一方面C:N的增加会降低有机物的营养价值, 影响细菌降解与浮游植物繁殖, 改变海洋生态系统的动力学. 此外, 酸化与光辐射(Suggett et al., 2013; Shi et al., 2015; Kleynhans et al., 2016)以及酸化与暖化(Pörtner, 2008; Mayor et al., 2012; Kroeker et al., 2013)之间的耦合作用, 也会从个体、种群、群落等不同水平上对海洋生态系统产生影响. ...
Offsetting the radiative benefit of ocean iron fertilization by enhancing N2O emissions
1
2003
... 海水富营养化会刺激浮游植物和大型藻类的生长, 增加海洋CO2的捕获量, 但过高的浮游植物生物量, 会降低光的穿透性, 影响水下生物的生长.此外, 营养盐比例的不平衡会改变浮游植物群落组成, 可能由此促进有毒藻类大量繁殖, 导致藻华的爆发(Hallegraeff, 2010; Flynn et al., 2015).有害藻类的大量繁殖不仅会导致鱼类和贝类等中毒死亡, 还会扩大海水的缺氧区(Rosenthal, 1985; Naylor et al., 2000; Rabalais et al., 2010).有研究表明, 1969-2009年期间, 由于人类活动的干扰, 大洋缺氧区的面积大小呈现递增趋势(Rabalais et al., 2010).以墨西哥北部湾缺氧区为例, 其面积从1985年的40 km2左右扩大到2015年的22 000 km2 (Sen Gupta et al., 1996; Bianchi et al., 2010; Turner & Rabalais, 2017).Neumann等(2012)的模型预测显示, 大洋季节性缺氧的持续时间将在21世纪末增加一倍.低氧会阻碍氮的硝化过程, 影响浮游植物对氮营养盐的吸收, 从而改变海洋初级生产力(Codispoti et al., 2001; Gruber, 2004), 并且不完全的硝化过程会导致温室气体N2O产量增多, 加快全球气候变暖的进程(Codispoti et al., 2001; Jin & Gruber, 2003; Nevison et al., 2003).此外, 低氧会导致有氧呼吸生物向外迁徙, 降低系统的生物多样性, 对系统物质循环与能量的流动产生影响, 破坏海洋生态系统的稳定性(Breitburg et al., 2009). ...
Ocean deoxygenation in a warming world
2
2010
... 海洋生态系统是由海洋生物群落及其环境相互作用所构成的自然系统, 其服务功能及生态价值既是地球生命系统的重要支撑组成, 也是社会与环境可持续发展的一项基本要素.海洋生态系统的稳定主要依托于海洋中复杂的生物过程, 而这些过程往往易受海洋环境变化的影响.近半个世纪以来, 海洋和陆地一样面临着前所未有的生态压力, 不断加剧的人类活动正对海洋生态系统产生深远的影响, 例如, 大气二氧化碳(CO2)浓度的上升就是一个关键的问题.CO2浓度的上升会导致海洋暖化和酸化(Doney et al., 2009).暖化会引起海水层化加剧、海洋缺氧区扩增以及环流模式改变等(Keeling et al., 2010), 而酸化则会导致海水碳酸盐系统改变等化学变化, 进而影响生物的生理活动以及群落的结构功能等, 最终对海洋生态系统产生一系列的影响.此外, 人类活动干扰导致的海水富营养化以及由此引发的赤潮及海洋缺氧等问题也给海洋生态系统带来了严峻的挑战.迄今为止, 围绕气候变化和人类活动对海洋生态系统的影响, 国际上开展了大量的研究工作并取得了一定的进展.本文将通过文献计量的方法分析国内外的研究现状, 简要回顾该研究领域的发展简史, 聚焦海洋暖化、海洋酸化和富营养化与缺氧这三个核心研究方向, 重点阐述其对海洋生态系统初级生产的关键过程的影响, 总结已取得的重要进展以及存在的主要问题, 最后提出前沿展望. ...
... 暖化的海水对生物的影响因种而异.对于大多数浮游植物而言, 在适温范围内, 海水暖化会促进其生长(Eppley, 1972), 但不同浮游植物对温度变化的响应程度不同, 因此全球暖化下浮游植物的生态位将发生改变(Xiao et al., 2018).然而, 对于大多数海洋动物, 暖化将在增加其体内代谢活性的同时加快其能量的消耗, 从而可能导致海洋生物自身生长与繁殖能力的下降(Pörtner & Knust, 2007).这些个体生理生化过程的变化会进一步改变种群的大小、时空分布格局与营养级水平, 最终引起群落结构与功能的改变(Bruno et al., 2003; Hyrenbach & Veit, 2003).从海洋生态系统的层面上来看, 全球变暖会改变海洋的初级生产力, 影响生态系统的稳定性.近期我国科学家采用模型估算发现, 1948-2007年, 全球气候变暖导致海洋水体层化加剧, 使得北大西洋、北太平洋和印度洋初级生产力分别降低了40%、24%和25% (Wang et al., 2017).倘若暖化现象一直持续, 到2100年, 海洋初级生产力将可能进一步减少20% (Steinacher et al., 2010).此外, 已有研究表明, 海水温度升高1 ℃, 珊瑚即会出现白化现象(Hoegh- Guldberg et al., 2007), 这会导致珊瑚礁生态系统多样性降低, 严重时会引发珊瑚礁系统的崩溃(Donner, 2009).海水暖化除了通过温度增加而对海洋生态系统产生直接影响外, 还会造成海洋层化加剧、缺氧区扩增等, 从而间接地对海洋生态系统产生一系列影响(Keeling et al., 2010; Wang et al., 2017). ...
Adaptation to elevated CO2 in different biodiversity contexts
1
2016
... 当前, 表层海水中CO2的浓度为10-20 µmol·L-1 (Badger et al., 1998), 低于绝大多数浮游植物中催化固碳反应的关键酶——核酮糖-1,5-二磷酸羧化酶/加氧酶(RubisCO)的半饱和常数.因此, 大多数浮游植物依赖于耗能的CO2浓缩机制(CO2-concentrating mechanism, CCM)以提高胞内RubisCO周围CO2的浓度(Reinfelder, 2011).已有研究表明, 酸化条件下, CO2浓度升高可使CCM下调, 节省能量, 从而对部分浮游植物的生长具有不同程度的促进作用(Riebesell et al., 1993; Mackey et al., 2015).除了CO2浓度升高, 随之降低的海水pH则同样可能对浮游植物产生直接或间接的影响.例如, 近期我国学者对海洋优势固氮蓝藻束毛藻(Trichodesmium)开展的研究表明, 海水pH下降导致其固氮酶效率降低, 干扰胞内pH稳态, 影响细胞产能, 且该负效应大于CO2浓度升高所产生的正效应, 故酸化的净效应为抑制束毛藻的固氮作用(Hong et al., 2017).此外, 海水pH下降还可能改变浮游植物对铁、磷等关键营养元素的利用(Shi et al., 2010; McQuaid et al., 2018), 从而间接影响浮游植物生长.除了光合作用和固氮作用外, 海水酸化还会对颗石藻、有孔虫类、珊瑚等钙化生物的钙化作用产生负面影响(Riebesell et al., 2000; Cao & Caldeira, 2008; Hofmann et al., 2010; Kroeker et al., 2013).对于海洋软体动物、棘皮动物、甲壳动物及鱼类等生物而言, 海洋酸化不仅会降低其幼体发育速度和个体大小(Ross et al., 2011), 还会降低部分生物类群的呼吸速率, 扰乱生物体内酸碱平衡, 影响其生长与繁殖(Ishimatsu et al., 2004; Pörtner & Langenbuch, 2005).海洋酸化对生物个体的影响会进一步体现在对海洋生态系统和海洋生物地球化学循环的影响.Riebesell等(2007)通过开展中尺度生态系统(mesocosm) CO2加富实验发现, 与350 mg·kg-1相比, 1 050 mg·kg-1 CO2浓度下的浮游植物群落在12天内多消耗了39%的无机碳, 且浮游植物的C:N从6升高到8, 这一方面将有利于提高生物碳泵的效率, 但另一方面C:N的增加会降低有机物的营养价值, 影响细菌降解与浮游植物繁殖, 改变海洋生态系统的动力学. 此外, 酸化与光辐射(Suggett et al., 2013; Shi et al., 2015; Kleynhans et al., 2016)以及酸化与暖化(Pörtner, 2008; Mayor et al., 2012; Kroeker et al., 2013)之间的耦合作用, 也会从个体、种群、群落等不同水平上对海洋生态系统产生影响. ...
Impacts of ocean acidification on marine organisms: quantifying sensitivities and interaction with warming
2
2013
... 当前, 表层海水中CO2的浓度为10-20 µmol·L-1 (Badger et al., 1998), 低于绝大多数浮游植物中催化固碳反应的关键酶——核酮糖-1,5-二磷酸羧化酶/加氧酶(RubisCO)的半饱和常数.因此, 大多数浮游植物依赖于耗能的CO2浓缩机制(CO2-concentrating mechanism, CCM)以提高胞内RubisCO周围CO2的浓度(Reinfelder, 2011).已有研究表明, 酸化条件下, CO2浓度升高可使CCM下调, 节省能量, 从而对部分浮游植物的生长具有不同程度的促进作用(Riebesell et al., 1993; Mackey et al., 2015).除了CO2浓度升高, 随之降低的海水pH则同样可能对浮游植物产生直接或间接的影响.例如, 近期我国学者对海洋优势固氮蓝藻束毛藻(Trichodesmium)开展的研究表明, 海水pH下降导致其固氮酶效率降低, 干扰胞内pH稳态, 影响细胞产能, 且该负效应大于CO2浓度升高所产生的正效应, 故酸化的净效应为抑制束毛藻的固氮作用(Hong et al., 2017).此外, 海水pH下降还可能改变浮游植物对铁、磷等关键营养元素的利用(Shi et al., 2010; McQuaid et al., 2018), 从而间接影响浮游植物生长.除了光合作用和固氮作用外, 海水酸化还会对颗石藻、有孔虫类、珊瑚等钙化生物的钙化作用产生负面影响(Riebesell et al., 2000; Cao & Caldeira, 2008; Hofmann et al., 2010; Kroeker et al., 2013).对于海洋软体动物、棘皮动物、甲壳动物及鱼类等生物而言, 海洋酸化不仅会降低其幼体发育速度和个体大小(Ross et al., 2011), 还会降低部分生物类群的呼吸速率, 扰乱生物体内酸碱平衡, 影响其生长与繁殖(Ishimatsu et al., 2004; Pörtner & Langenbuch, 2005).海洋酸化对生物个体的影响会进一步体现在对海洋生态系统和海洋生物地球化学循环的影响.Riebesell等(2007)通过开展中尺度生态系统(mesocosm) CO2加富实验发现, 与350 mg·kg-1相比, 1 050 mg·kg-1 CO2浓度下的浮游植物群落在12天内多消耗了39%的无机碳, 且浮游植物的C:N从6升高到8, 这一方面将有利于提高生物碳泵的效率, 但另一方面C:N的增加会降低有机物的营养价值, 影响细菌降解与浮游植物繁殖, 改变海洋生态系统的动力学. 此外, 酸化与光辐射(Suggett et al., 2013; Shi et al., 2015; Kleynhans et al., 2016)以及酸化与暖化(Pörtner, 2008; Mayor et al., 2012; Kroeker et al., 2013)之间的耦合作用, 也会从个体、种群、群落等不同水平上对海洋生态系统产生影响. ...
... ; Kroeker et al., 2013)之间的耦合作用, 也会从个体、种群、群落等不同水平上对海洋生态系统产生影响. ...
Response of photosynthesis to ocean acidification
1
2015
... 当前, 表层海水中CO2的浓度为10-20 µmol·L-1 (Badger et al., 1998), 低于绝大多数浮游植物中催化固碳反应的关键酶——核酮糖-1,5-二磷酸羧化酶/加氧酶(RubisCO)的半饱和常数.因此, 大多数浮游植物依赖于耗能的CO2浓缩机制(CO2-concentrating mechanism, CCM)以提高胞内RubisCO周围CO2的浓度(Reinfelder, 2011).已有研究表明, 酸化条件下, CO2浓度升高可使CCM下调, 节省能量, 从而对部分浮游植物的生长具有不同程度的促进作用(Riebesell et al., 1993; Mackey et al., 2015).除了CO2浓度升高, 随之降低的海水pH则同样可能对浮游植物产生直接或间接的影响.例如, 近期我国学者对海洋优势固氮蓝藻束毛藻(Trichodesmium)开展的研究表明, 海水pH下降导致其固氮酶效率降低, 干扰胞内pH稳态, 影响细胞产能, 且该负效应大于CO2浓度升高所产生的正效应, 故酸化的净效应为抑制束毛藻的固氮作用(Hong et al., 2017).此外, 海水pH下降还可能改变浮游植物对铁、磷等关键营养元素的利用(Shi et al., 2010; McQuaid et al., 2018), 从而间接影响浮游植物生长.除了光合作用和固氮作用外, 海水酸化还会对颗石藻、有孔虫类、珊瑚等钙化生物的钙化作用产生负面影响(Riebesell et al., 2000; Cao & Caldeira, 2008; Hofmann et al., 2010; Kroeker et al., 2013).对于海洋软体动物、棘皮动物、甲壳动物及鱼类等生物而言, 海洋酸化不仅会降低其幼体发育速度和个体大小(Ross et al., 2011), 还会降低部分生物类群的呼吸速率, 扰乱生物体内酸碱平衡, 影响其生长与繁殖(Ishimatsu et al., 2004; Pörtner & Langenbuch, 2005).海洋酸化对生物个体的影响会进一步体现在对海洋生态系统和海洋生物地球化学循环的影响.Riebesell等(2007)通过开展中尺度生态系统(mesocosm) CO2加富实验发现, 与350 mg·kg-1相比, 1 050 mg·kg-1 CO2浓度下的浮游植物群落在12天内多消耗了39%的无机碳, 且浮游植物的C:N从6升高到8, 这一方面将有利于提高生物碳泵的效率, 但另一方面C:N的增加会降低有机物的营养价值, 影响细菌降解与浮游植物繁殖, 改变海洋生态系统的动力学. 此外, 酸化与光辐射(Suggett et al., 2013; Shi et al., 2015; Kleynhans et al., 2016)以及酸化与暖化(Pörtner, 2008; Mayor et al., 2012; Kroeker et al., 2013)之间的耦合作用, 也会从个体、种群、群落等不同水平上对海洋生态系统产生影响. ...
End of century ocean warming and acidification effects on reproductive success in a temperate marine copepod
1
2012
... 当前, 表层海水中CO2的浓度为10-20 µmol·L-1 (Badger et al., 1998), 低于绝大多数浮游植物中催化固碳反应的关键酶——核酮糖-1,5-二磷酸羧化酶/加氧酶(RubisCO)的半饱和常数.因此, 大多数浮游植物依赖于耗能的CO2浓缩机制(CO2-concentrating mechanism, CCM)以提高胞内RubisCO周围CO2的浓度(Reinfelder, 2011).已有研究表明, 酸化条件下, CO2浓度升高可使CCM下调, 节省能量, 从而对部分浮游植物的生长具有不同程度的促进作用(Riebesell et al., 1993; Mackey et al., 2015).除了CO2浓度升高, 随之降低的海水pH则同样可能对浮游植物产生直接或间接的影响.例如, 近期我国学者对海洋优势固氮蓝藻束毛藻(Trichodesmium)开展的研究表明, 海水pH下降导致其固氮酶效率降低, 干扰胞内pH稳态, 影响细胞产能, 且该负效应大于CO2浓度升高所产生的正效应, 故酸化的净效应为抑制束毛藻的固氮作用(Hong et al., 2017).此外, 海水pH下降还可能改变浮游植物对铁、磷等关键营养元素的利用(Shi et al., 2010; McQuaid et al., 2018), 从而间接影响浮游植物生长.除了光合作用和固氮作用外, 海水酸化还会对颗石藻、有孔虫类、珊瑚等钙化生物的钙化作用产生负面影响(Riebesell et al., 2000; Cao & Caldeira, 2008; Hofmann et al., 2010; Kroeker et al., 2013).对于海洋软体动物、棘皮动物、甲壳动物及鱼类等生物而言, 海洋酸化不仅会降低其幼体发育速度和个体大小(Ross et al., 2011), 还会降低部分生物类群的呼吸速率, 扰乱生物体内酸碱平衡, 影响其生长与繁殖(Ishimatsu et al., 2004; Pörtner & Langenbuch, 2005).海洋酸化对生物个体的影响会进一步体现在对海洋生态系统和海洋生物地球化学循环的影响.Riebesell等(2007)通过开展中尺度生态系统(mesocosm) CO2加富实验发现, 与350 mg·kg-1相比, 1 050 mg·kg-1 CO2浓度下的浮游植物群落在12天内多消耗了39%的无机碳, 且浮游植物的C:N从6升高到8, 这一方面将有利于提高生物碳泵的效率, 但另一方面C:N的增加会降低有机物的营养价值, 影响细菌降解与浮游植物繁殖, 改变海洋生态系统的动力学. 此外, 酸化与光辐射(Suggett et al., 2013; Shi et al., 2015; Kleynhans et al., 2016)以及酸化与暖化(Pörtner, 2008; Mayor et al., 2012; Kroeker et al., 2013)之间的耦合作用, 也会从个体、种群、群落等不同水平上对海洋生态系统产生影响. ...
Carbonate-sensitive phytotransferrin controls high-affinity iron uptake in diatoms
1
2018
... 当前, 表层海水中CO2的浓度为10-20 µmol·L-1 (Badger et al., 1998), 低于绝大多数浮游植物中催化固碳反应的关键酶——核酮糖-1,5-二磷酸羧化酶/加氧酶(RubisCO)的半饱和常数.因此, 大多数浮游植物依赖于耗能的CO2浓缩机制(CO2-concentrating mechanism, CCM)以提高胞内RubisCO周围CO2的浓度(Reinfelder, 2011).已有研究表明, 酸化条件下, CO2浓度升高可使CCM下调, 节省能量, 从而对部分浮游植物的生长具有不同程度的促进作用(Riebesell et al., 1993; Mackey et al., 2015).除了CO2浓度升高, 随之降低的海水pH则同样可能对浮游植物产生直接或间接的影响.例如, 近期我国学者对海洋优势固氮蓝藻束毛藻(Trichodesmium)开展的研究表明, 海水pH下降导致其固氮酶效率降低, 干扰胞内pH稳态, 影响细胞产能, 且该负效应大于CO2浓度升高所产生的正效应, 故酸化的净效应为抑制束毛藻的固氮作用(Hong et al., 2017).此外, 海水pH下降还可能改变浮游植物对铁、磷等关键营养元素的利用(Shi et al., 2010; McQuaid et al., 2018), 从而间接影响浮游植物生长.除了光合作用和固氮作用外, 海水酸化还会对颗石藻、有孔虫类、珊瑚等钙化生物的钙化作用产生负面影响(Riebesell et al., 2000; Cao & Caldeira, 2008; Hofmann et al., 2010; Kroeker et al., 2013).对于海洋软体动物、棘皮动物、甲壳动物及鱼类等生物而言, 海洋酸化不仅会降低其幼体发育速度和个体大小(Ross et al., 2011), 还会降低部分生物类群的呼吸速率, 扰乱生物体内酸碱平衡, 影响其生长与繁殖(Ishimatsu et al., 2004; Pörtner & Langenbuch, 2005).海洋酸化对生物个体的影响会进一步体现在对海洋生态系统和海洋生物地球化学循环的影响.Riebesell等(2007)通过开展中尺度生态系统(mesocosm) CO2加富实验发现, 与350 mg·kg-1相比, 1 050 mg·kg-1 CO2浓度下的浮游植物群落在12天内多消耗了39%的无机碳, 且浮游植物的C:N从6升高到8, 这一方面将有利于提高生物碳泵的效率, 但另一方面C:N的增加会降低有机物的营养价值, 影响细菌降解与浮游植物繁殖, 改变海洋生态系统的动力学. 此外, 酸化与光辐射(Suggett et al., 2013; Shi et al., 2015; Kleynhans et al., 2016)以及酸化与暖化(Pörtner, 2008; Mayor et al., 2012; Kroeker et al., 2013)之间的耦合作用, 也会从个体、种群、群落等不同水平上对海洋生态系统产生影响. ...
On the effects of alkalies and acids, and of alkaline and acid salts, upon growth and cell division in the fertilized eggs of Echinus esculentus —A study in relationship to the causation of malignant disease
1
1906
... 对全球气候变化的研究历史最早可以追溯到19世纪.早在19世纪20年代, 科学家就已经发现了几种温室气体, 但并未引起广泛的关注.直到19世纪30年代, 科学家意识到燃烧化石燃料对气候的影响后, 便开展了一系列研究工作.之后随着科学技术的发展, 全球气候变化对生态系统影响的研究于20世纪90年代迎来了黄金期(Weart, 2003), 其中就包括对海洋生态系统影响的研究.以全球气候变化导致的海洋酸化为例, 早在1906年, 针对海洋酸碱性, Moore等(1906)就已经开展了酸碱性对海胆受精卵的生长和细胞分裂的影响研究; 1951年, Rubey总结了早期酸化的研究工作, 为后人的研究提供了理论基础; 1957年, Revelle和Suess首次提出海洋吸收人类排放的CO2会降低海洋的缓冲能力, 引起了科学界对人类活动影响海洋生态系统的重视; 1966年, Broecker和Takahashi首次报道了海水CO32-的浓度, 并提出其将随着海水CO2浓度的升高而降低; 1978年, Brewer提出利用总碱度和溶解无机碳的浓度校正海水中的CO2系统; 1993年硼同位素等古海洋示踪剂开始被拓展应用于估算海洋化学参数的变化(Spivack et al., 1993); 2001年, Bates第一次利用时间序列数据记录了北大西洋西部海水碳酸盐体系的变化,主要包括海水的CO2浓度、CO2分压以及碱度的变动; 近年来, 利用受控实验评估生物体对海水碳酸盐体系变化响应的论文大量涌现, 与此同时科学家开始通过构建数值模型来预测未来海水碳酸盐系统的变化以及海洋酸化对海洋生物地球化学循环的影响. ...
Effect of aquaculture on world fish supplies
1
2000
... 海水富营养化会刺激浮游植物和大型藻类的生长, 增加海洋CO2的捕获量, 但过高的浮游植物生物量, 会降低光的穿透性, 影响水下生物的生长.此外, 营养盐比例的不平衡会改变浮游植物群落组成, 可能由此促进有毒藻类大量繁殖, 导致藻华的爆发(Hallegraeff, 2010; Flynn et al., 2015).有害藻类的大量繁殖不仅会导致鱼类和贝类等中毒死亡, 还会扩大海水的缺氧区(Rosenthal, 1985; Naylor et al., 2000; Rabalais et al., 2010).有研究表明, 1969-2009年期间, 由于人类活动的干扰, 大洋缺氧区的面积大小呈现递增趋势(Rabalais et al., 2010).以墨西哥北部湾缺氧区为例, 其面积从1985年的40 km2左右扩大到2015年的22 000 km2 (Sen Gupta et al., 1996; Bianchi et al., 2010; Turner & Rabalais, 2017).Neumann等(2012)的模型预测显示, 大洋季节性缺氧的持续时间将在21世纪末增加一倍.低氧会阻碍氮的硝化过程, 影响浮游植物对氮营养盐的吸收, 从而改变海洋初级生产力(Codispoti et al., 2001; Gruber, 2004), 并且不完全的硝化过程会导致温室气体N2O产量增多, 加快全球气候变暖的进程(Codispoti et al., 2001; Jin & Gruber, 2003; Nevison et al., 2003).此外, 低氧会导致有氧呼吸生物向外迁徙, 降低系统的生物多样性, 对系统物质循环与能量的流动产生影响, 破坏海洋生态系统的稳定性(Breitburg et al., 2009). ...
Extremes of temperature, oxygen and blooms in the Baltic Sea in a changing climate
1
2012
... 海水富营养化会刺激浮游植物和大型藻类的生长, 增加海洋CO2的捕获量, 但过高的浮游植物生物量, 会降低光的穿透性, 影响水下生物的生长.此外, 营养盐比例的不平衡会改变浮游植物群落组成, 可能由此促进有毒藻类大量繁殖, 导致藻华的爆发(Hallegraeff, 2010; Flynn et al., 2015).有害藻类的大量繁殖不仅会导致鱼类和贝类等中毒死亡, 还会扩大海水的缺氧区(Rosenthal, 1985; Naylor et al., 2000; Rabalais et al., 2010).有研究表明, 1969-2009年期间, 由于人类活动的干扰, 大洋缺氧区的面积大小呈现递增趋势(Rabalais et al., 2010).以墨西哥北部湾缺氧区为例, 其面积从1985年的40 km2左右扩大到2015年的22 000 km2 (Sen Gupta et al., 1996; Bianchi et al., 2010; Turner & Rabalais, 2017).Neumann等(2012)的模型预测显示, 大洋季节性缺氧的持续时间将在21世纪末增加一倍.低氧会阻碍氮的硝化过程, 影响浮游植物对氮营养盐的吸收, 从而改变海洋初级生产力(Codispoti et al., 2001; Gruber, 2004), 并且不完全的硝化过程会导致温室气体N2O产量增多, 加快全球气候变暖的进程(Codispoti et al., 2001; Jin & Gruber, 2003; Nevison et al., 2003).此外, 低氧会导致有氧呼吸生物向外迁徙, 降低系统的生物多样性, 对系统物质循环与能量的流动产生影响, 破坏海洋生态系统的稳定性(Breitburg et al., 2009). ...
Global distribution of N2O and the ΔN2O-AOU yield in the subsurface ocean
1
2003
... 海水富营养化会刺激浮游植物和大型藻类的生长, 增加海洋CO2的捕获量, 但过高的浮游植物生物量, 会降低光的穿透性, 影响水下生物的生长.此外, 营养盐比例的不平衡会改变浮游植物群落组成, 可能由此促进有毒藻类大量繁殖, 导致藻华的爆发(Hallegraeff, 2010; Flynn et al., 2015).有害藻类的大量繁殖不仅会导致鱼类和贝类等中毒死亡, 还会扩大海水的缺氧区(Rosenthal, 1985; Naylor et al., 2000; Rabalais et al., 2010).有研究表明, 1969-2009年期间, 由于人类活动的干扰, 大洋缺氧区的面积大小呈现递增趋势(Rabalais et al., 2010).以墨西哥北部湾缺氧区为例, 其面积从1985年的40 km2左右扩大到2015年的22 000 km2 (Sen Gupta et al., 1996; Bianchi et al., 2010; Turner & Rabalais, 2017).Neumann等(2012)的模型预测显示, 大洋季节性缺氧的持续时间将在21世纪末增加一倍.低氧会阻碍氮的硝化过程, 影响浮游植物对氮营养盐的吸收, 从而改变海洋初级生产力(Codispoti et al., 2001; Gruber, 2004), 并且不完全的硝化过程会导致温室气体N2O产量增多, 加快全球气候变暖的进程(Codispoti et al., 2001; Jin & Gruber, 2003; Nevison et al., 2003).此外, 低氧会导致有氧呼吸生物向外迁徙, 降低系统的生物多样性, 对系统物质循环与能量的流动产生影响, 破坏海洋生态系统的稳定性(Breitburg et al., 2009). ...
Ecosystem effects of ocean acidification in times of ocean warming: a physiologist’s view
1
2008
... 当前, 表层海水中CO2的浓度为10-20 µmol·L-1 (Badger et al., 1998), 低于绝大多数浮游植物中催化固碳反应的关键酶——核酮糖-1,5-二磷酸羧化酶/加氧酶(RubisCO)的半饱和常数.因此, 大多数浮游植物依赖于耗能的CO2浓缩机制(CO2-concentrating mechanism, CCM)以提高胞内RubisCO周围CO2的浓度(Reinfelder, 2011).已有研究表明, 酸化条件下, CO2浓度升高可使CCM下调, 节省能量, 从而对部分浮游植物的生长具有不同程度的促进作用(Riebesell et al., 1993; Mackey et al., 2015).除了CO2浓度升高, 随之降低的海水pH则同样可能对浮游植物产生直接或间接的影响.例如, 近期我国学者对海洋优势固氮蓝藻束毛藻(Trichodesmium)开展的研究表明, 海水pH下降导致其固氮酶效率降低, 干扰胞内pH稳态, 影响细胞产能, 且该负效应大于CO2浓度升高所产生的正效应, 故酸化的净效应为抑制束毛藻的固氮作用(Hong et al., 2017).此外, 海水pH下降还可能改变浮游植物对铁、磷等关键营养元素的利用(Shi et al., 2010; McQuaid et al., 2018), 从而间接影响浮游植物生长.除了光合作用和固氮作用外, 海水酸化还会对颗石藻、有孔虫类、珊瑚等钙化生物的钙化作用产生负面影响(Riebesell et al., 2000; Cao & Caldeira, 2008; Hofmann et al., 2010; Kroeker et al., 2013).对于海洋软体动物、棘皮动物、甲壳动物及鱼类等生物而言, 海洋酸化不仅会降低其幼体发育速度和个体大小(Ross et al., 2011), 还会降低部分生物类群的呼吸速率, 扰乱生物体内酸碱平衡, 影响其生长与繁殖(Ishimatsu et al., 2004; Pörtner & Langenbuch, 2005).海洋酸化对生物个体的影响会进一步体现在对海洋生态系统和海洋生物地球化学循环的影响.Riebesell等(2007)通过开展中尺度生态系统(mesocosm) CO2加富实验发现, 与350 mg·kg-1相比, 1 050 mg·kg-1 CO2浓度下的浮游植物群落在12天内多消耗了39%的无机碳, 且浮游植物的C:N从6升高到8, 这一方面将有利于提高生物碳泵的效率, 但另一方面C:N的增加会降低有机物的营养价值, 影响细菌降解与浮游植物繁殖, 改变海洋生态系统的动力学. 此外, 酸化与光辐射(Suggett et al., 2013; Shi et al., 2015; Kleynhans et al., 2016)以及酸化与暖化(Pörtner, 2008; Mayor et al., 2012; Kroeker et al., 2013)之间的耦合作用, 也会从个体、种群、群落等不同水平上对海洋生态系统产生影响. ...
Climate change affects marine fishes through the oxygen limitation of thermal tolerance
1
2007
... 暖化的海水对生物的影响因种而异.对于大多数浮游植物而言, 在适温范围内, 海水暖化会促进其生长(Eppley, 1972), 但不同浮游植物对温度变化的响应程度不同, 因此全球暖化下浮游植物的生态位将发生改变(Xiao et al., 2018).然而, 对于大多数海洋动物, 暖化将在增加其体内代谢活性的同时加快其能量的消耗, 从而可能导致海洋生物自身生长与繁殖能力的下降(Pörtner & Knust, 2007).这些个体生理生化过程的变化会进一步改变种群的大小、时空分布格局与营养级水平, 最终引起群落结构与功能的改变(Bruno et al., 2003; Hyrenbach & Veit, 2003).从海洋生态系统的层面上来看, 全球变暖会改变海洋的初级生产力, 影响生态系统的稳定性.近期我国科学家采用模型估算发现, 1948-2007年, 全球气候变暖导致海洋水体层化加剧, 使得北大西洋、北太平洋和印度洋初级生产力分别降低了40%、24%和25% (Wang et al., 2017).倘若暖化现象一直持续, 到2100年, 海洋初级生产力将可能进一步减少20% (Steinacher et al., 2010).此外, 已有研究表明, 海水温度升高1 ℃, 珊瑚即会出现白化现象(Hoegh- Guldberg et al., 2007), 这会导致珊瑚礁生态系统多样性降低, 严重时会引发珊瑚礁系统的崩溃(Donner, 2009).海水暖化除了通过温度增加而对海洋生态系统产生直接影响外, 还会造成海洋层化加剧、缺氧区扩增等, 从而间接地对海洋生态系统产生一系列影响(Keeling et al., 2010; Wang et al., 2017). ...
Synergistic effects of temperature extremes, hypoxia, and increases in CO2 on marine animals: from earth history to global change
1
2005
... 当前, 表层海水中CO2的浓度为10-20 µmol·L-1 (Badger et al., 1998), 低于绝大多数浮游植物中催化固碳反应的关键酶——核酮糖-1,5-二磷酸羧化酶/加氧酶(RubisCO)的半饱和常数.因此, 大多数浮游植物依赖于耗能的CO2浓缩机制(CO2-concentrating mechanism, CCM)以提高胞内RubisCO周围CO2的浓度(Reinfelder, 2011).已有研究表明, 酸化条件下, CO2浓度升高可使CCM下调, 节省能量, 从而对部分浮游植物的生长具有不同程度的促进作用(Riebesell et al., 1993; Mackey et al., 2015).除了CO2浓度升高, 随之降低的海水pH则同样可能对浮游植物产生直接或间接的影响.例如, 近期我国学者对海洋优势固氮蓝藻束毛藻(Trichodesmium)开展的研究表明, 海水pH下降导致其固氮酶效率降低, 干扰胞内pH稳态, 影响细胞产能, 且该负效应大于CO2浓度升高所产生的正效应, 故酸化的净效应为抑制束毛藻的固氮作用(Hong et al., 2017).此外, 海水pH下降还可能改变浮游植物对铁、磷等关键营养元素的利用(Shi et al., 2010; McQuaid et al., 2018), 从而间接影响浮游植物生长.除了光合作用和固氮作用外, 海水酸化还会对颗石藻、有孔虫类、珊瑚等钙化生物的钙化作用产生负面影响(Riebesell et al., 2000; Cao & Caldeira, 2008; Hofmann et al., 2010; Kroeker et al., 2013).对于海洋软体动物、棘皮动物、甲壳动物及鱼类等生物而言, 海洋酸化不仅会降低其幼体发育速度和个体大小(Ross et al., 2011), 还会降低部分生物类群的呼吸速率, 扰乱生物体内酸碱平衡, 影响其生长与繁殖(Ishimatsu et al., 2004; Pörtner & Langenbuch, 2005).海洋酸化对生物个体的影响会进一步体现在对海洋生态系统和海洋生物地球化学循环的影响.Riebesell等(2007)通过开展中尺度生态系统(mesocosm) CO2加富实验发现, 与350 mg·kg-1相比, 1 050 mg·kg-1 CO2浓度下的浮游植物群落在12天内多消耗了39%的无机碳, 且浮游植物的C:N从6升高到8, 这一方面将有利于提高生物碳泵的效率, 但另一方面C:N的增加会降低有机物的营养价值, 影响细菌降解与浮游植物繁殖, 改变海洋生态系统的动力学. 此外, 酸化与光辐射(Suggett et al., 2013; Shi et al., 2015; Kleynhans et al., 2016)以及酸化与暖化(Pörtner, 2008; Mayor et al., 2012; Kroeker et al., 2013)之间的耦合作用, 也会从个体、种群、群落等不同水平上对海洋生态系统产生影响. ...
Dynamics and distribution of natural and human- caused hypoxia
2
2010
... 海水富营养化会刺激浮游植物和大型藻类的生长, 增加海洋CO2的捕获量, 但过高的浮游植物生物量, 会降低光的穿透性, 影响水下生物的生长.此外, 营养盐比例的不平衡会改变浮游植物群落组成, 可能由此促进有毒藻类大量繁殖, 导致藻华的爆发(Hallegraeff, 2010; Flynn et al., 2015).有害藻类的大量繁殖不仅会导致鱼类和贝类等中毒死亡, 还会扩大海水的缺氧区(Rosenthal, 1985; Naylor et al., 2000; Rabalais et al., 2010).有研究表明, 1969-2009年期间, 由于人类活动的干扰, 大洋缺氧区的面积大小呈现递增趋势(Rabalais et al., 2010).以墨西哥北部湾缺氧区为例, 其面积从1985年的40 km2左右扩大到2015年的22 000 km2 (Sen Gupta et al., 1996; Bianchi et al., 2010; Turner & Rabalais, 2017).Neumann等(2012)的模型预测显示, 大洋季节性缺氧的持续时间将在21世纪末增加一倍.低氧会阻碍氮的硝化过程, 影响浮游植物对氮营养盐的吸收, 从而改变海洋初级生产力(Codispoti et al., 2001; Gruber, 2004), 并且不完全的硝化过程会导致温室气体N2O产量增多, 加快全球气候变暖的进程(Codispoti et al., 2001; Jin & Gruber, 2003; Nevison et al., 2003).此外, 低氧会导致有氧呼吸生物向外迁徙, 降低系统的生物多样性, 对系统物质循环与能量的流动产生影响, 破坏海洋生态系统的稳定性(Breitburg et al., 2009). ...
... ).有研究表明, 1969-2009年期间, 由于人类活动的干扰, 大洋缺氧区的面积大小呈现递增趋势(Rabalais et al., 2010).以墨西哥北部湾缺氧区为例, 其面积从1985年的40 km2左右扩大到2015年的22 000 km2 (Sen Gupta et al., 1996; Bianchi et al., 2010; Turner & Rabalais, 2017).Neumann等(2012)的模型预测显示, 大洋季节性缺氧的持续时间将在21世纪末增加一倍.低氧会阻碍氮的硝化过程, 影响浮游植物对氮营养盐的吸收, 从而改变海洋初级生产力(Codispoti et al., 2001; Gruber, 2004), 并且不完全的硝化过程会导致温室气体N2O产量增多, 加快全球气候变暖的进程(Codispoti et al., 2001; Jin & Gruber, 2003; Nevison et al., 2003).此外, 低氧会导致有氧呼吸生物向外迁徙, 降低系统的生物多样性, 对系统物质循环与能量的流动产生影响, 破坏海洋生态系统的稳定性(Breitburg et al., 2009). ...
Carbon concentrating mechanisms in eukaryotic marine phytoplankton
1
2011
... 当前, 表层海水中CO2的浓度为10-20 µmol·L-1 (Badger et al., 1998), 低于绝大多数浮游植物中催化固碳反应的关键酶——核酮糖-1,5-二磷酸羧化酶/加氧酶(RubisCO)的半饱和常数.因此, 大多数浮游植物依赖于耗能的CO2浓缩机制(CO2-concentrating mechanism, CCM)以提高胞内RubisCO周围CO2的浓度(Reinfelder, 2011).已有研究表明, 酸化条件下, CO2浓度升高可使CCM下调, 节省能量, 从而对部分浮游植物的生长具有不同程度的促进作用(Riebesell et al., 1993; Mackey et al., 2015).除了CO2浓度升高, 随之降低的海水pH则同样可能对浮游植物产生直接或间接的影响.例如, 近期我国学者对海洋优势固氮蓝藻束毛藻(Trichodesmium)开展的研究表明, 海水pH下降导致其固氮酶效率降低, 干扰胞内pH稳态, 影响细胞产能, 且该负效应大于CO2浓度升高所产生的正效应, 故酸化的净效应为抑制束毛藻的固氮作用(Hong et al., 2017).此外, 海水pH下降还可能改变浮游植物对铁、磷等关键营养元素的利用(Shi et al., 2010; McQuaid et al., 2018), 从而间接影响浮游植物生长.除了光合作用和固氮作用外, 海水酸化还会对颗石藻、有孔虫类、珊瑚等钙化生物的钙化作用产生负面影响(Riebesell et al., 2000; Cao & Caldeira, 2008; Hofmann et al., 2010; Kroeker et al., 2013).对于海洋软体动物、棘皮动物、甲壳动物及鱼类等生物而言, 海洋酸化不仅会降低其幼体发育速度和个体大小(Ross et al., 2011), 还会降低部分生物类群的呼吸速率, 扰乱生物体内酸碱平衡, 影响其生长与繁殖(Ishimatsu et al., 2004; Pörtner & Langenbuch, 2005).海洋酸化对生物个体的影响会进一步体现在对海洋生态系统和海洋生物地球化学循环的影响.Riebesell等(2007)通过开展中尺度生态系统(mesocosm) CO2加富实验发现, 与350 mg·kg-1相比, 1 050 mg·kg-1 CO2浓度下的浮游植物群落在12天内多消耗了39%的无机碳, 且浮游植物的C:N从6升高到8, 这一方面将有利于提高生物碳泵的效率, 但另一方面C:N的增加会降低有机物的营养价值, 影响细菌降解与浮游植物繁殖, 改变海洋生态系统的动力学. 此外, 酸化与光辐射(Suggett et al., 2013; Shi et al., 2015; Kleynhans et al., 2016)以及酸化与暖化(Pörtner, 2008; Mayor et al., 2012; Kroeker et al., 2013)之间的耦合作用, 也会从个体、种群、群落等不同水平上对海洋生态系统产生影响. ...
Carbon dioxide exchange between atmosphere and ocean and the question of an increase of atmospheric CO2 during the past decades
1
1957
... 对全球气候变化的研究历史最早可以追溯到19世纪.早在19世纪20年代, 科学家就已经发现了几种温室气体, 但并未引起广泛的关注.直到19世纪30年代, 科学家意识到燃烧化石燃料对气候的影响后, 便开展了一系列研究工作.之后随着科学技术的发展, 全球气候变化对生态系统影响的研究于20世纪90年代迎来了黄金期(Weart, 2003), 其中就包括对海洋生态系统影响的研究.以全球气候变化导致的海洋酸化为例, 早在1906年, 针对海洋酸碱性, Moore等(1906)就已经开展了酸碱性对海胆受精卵的生长和细胞分裂的影响研究; 1951年, Rubey总结了早期酸化的研究工作, 为后人的研究提供了理论基础; 1957年, Revelle和Suess首次提出海洋吸收人类排放的CO2会降低海洋的缓冲能力, 引起了科学界对人类活动影响海洋生态系统的重视; 1966年, Broecker和Takahashi首次报道了海水CO32-的浓度, 并提出其将随着海水CO2浓度的升高而降低; 1978年, Brewer提出利用总碱度和溶解无机碳的浓度校正海水中的CO2系统; 1993年硼同位素等古海洋示踪剂开始被拓展应用于估算海洋化学参数的变化(Spivack et al., 1993); 2001年, Bates第一次利用时间序列数据记录了北大西洋西部海水碳酸盐体系的变化,主要包括海水的CO2浓度、CO2分压以及碱度的变动; 近年来, 利用受控实验评估生物体对海水碳酸盐体系变化响应的论文大量涌现, 与此同时科学家开始通过构建数值模型来预测未来海水碳酸盐系统的变化以及海洋酸化对海洋生物地球化学循环的影响. ...
Enhanced biological carbon consumption in a high CO2 ocean
1
2007
... 当前, 表层海水中CO2的浓度为10-20 µmol·L-1 (Badger et al., 1998), 低于绝大多数浮游植物中催化固碳反应的关键酶——核酮糖-1,5-二磷酸羧化酶/加氧酶(RubisCO)的半饱和常数.因此, 大多数浮游植物依赖于耗能的CO2浓缩机制(CO2-concentrating mechanism, CCM)以提高胞内RubisCO周围CO2的浓度(Reinfelder, 2011).已有研究表明, 酸化条件下, CO2浓度升高可使CCM下调, 节省能量, 从而对部分浮游植物的生长具有不同程度的促进作用(Riebesell et al., 1993; Mackey et al., 2015).除了CO2浓度升高, 随之降低的海水pH则同样可能对浮游植物产生直接或间接的影响.例如, 近期我国学者对海洋优势固氮蓝藻束毛藻(Trichodesmium)开展的研究表明, 海水pH下降导致其固氮酶效率降低, 干扰胞内pH稳态, 影响细胞产能, 且该负效应大于CO2浓度升高所产生的正效应, 故酸化的净效应为抑制束毛藻的固氮作用(Hong et al., 2017).此外, 海水pH下降还可能改变浮游植物对铁、磷等关键营养元素的利用(Shi et al., 2010; McQuaid et al., 2018), 从而间接影响浮游植物生长.除了光合作用和固氮作用外, 海水酸化还会对颗石藻、有孔虫类、珊瑚等钙化生物的钙化作用产生负面影响(Riebesell et al., 2000; Cao & Caldeira, 2008; Hofmann et al., 2010; Kroeker et al., 2013).对于海洋软体动物、棘皮动物、甲壳动物及鱼类等生物而言, 海洋酸化不仅会降低其幼体发育速度和个体大小(Ross et al., 2011), 还会降低部分生物类群的呼吸速率, 扰乱生物体内酸碱平衡, 影响其生长与繁殖(Ishimatsu et al., 2004; Pörtner & Langenbuch, 2005).海洋酸化对生物个体的影响会进一步体现在对海洋生态系统和海洋生物地球化学循环的影响.Riebesell等(2007)通过开展中尺度生态系统(mesocosm) CO2加富实验发现, 与350 mg·kg-1相比, 1 050 mg·kg-1 CO2浓度下的浮游植物群落在12天内多消耗了39%的无机碳, 且浮游植物的C:N从6升高到8, 这一方面将有利于提高生物碳泵的效率, 但另一方面C:N的增加会降低有机物的营养价值, 影响细菌降解与浮游植物繁殖, 改变海洋生态系统的动力学. 此外, 酸化与光辐射(Suggett et al., 2013; Shi et al., 2015; Kleynhans et al., 2016)以及酸化与暖化(Pörtner, 2008; Mayor et al., 2012; Kroeker et al., 2013)之间的耦合作用, 也会从个体、种群、群落等不同水平上对海洋生态系统产生影响. ...
Carbon dioxide limitation of marine phytoplankton growth rates
1
1993
... 当前, 表层海水中CO2的浓度为10-20 µmol·L-1 (Badger et al., 1998), 低于绝大多数浮游植物中催化固碳反应的关键酶——核酮糖-1,5-二磷酸羧化酶/加氧酶(RubisCO)的半饱和常数.因此, 大多数浮游植物依赖于耗能的CO2浓缩机制(CO2-concentrating mechanism, CCM)以提高胞内RubisCO周围CO2的浓度(Reinfelder, 2011).已有研究表明, 酸化条件下, CO2浓度升高可使CCM下调, 节省能量, 从而对部分浮游植物的生长具有不同程度的促进作用(Riebesell et al., 1993; Mackey et al., 2015).除了CO2浓度升高, 随之降低的海水pH则同样可能对浮游植物产生直接或间接的影响.例如, 近期我国学者对海洋优势固氮蓝藻束毛藻(Trichodesmium)开展的研究表明, 海水pH下降导致其固氮酶效率降低, 干扰胞内pH稳态, 影响细胞产能, 且该负效应大于CO2浓度升高所产生的正效应, 故酸化的净效应为抑制束毛藻的固氮作用(Hong et al., 2017).此外, 海水pH下降还可能改变浮游植物对铁、磷等关键营养元素的利用(Shi et al., 2010; McQuaid et al., 2018), 从而间接影响浮游植物生长.除了光合作用和固氮作用外, 海水酸化还会对颗石藻、有孔虫类、珊瑚等钙化生物的钙化作用产生负面影响(Riebesell et al., 2000; Cao & Caldeira, 2008; Hofmann et al., 2010; Kroeker et al., 2013).对于海洋软体动物、棘皮动物、甲壳动物及鱼类等生物而言, 海洋酸化不仅会降低其幼体发育速度和个体大小(Ross et al., 2011), 还会降低部分生物类群的呼吸速率, 扰乱生物体内酸碱平衡, 影响其生长与繁殖(Ishimatsu et al., 2004; Pörtner & Langenbuch, 2005).海洋酸化对生物个体的影响会进一步体现在对海洋生态系统和海洋生物地球化学循环的影响.Riebesell等(2007)通过开展中尺度生态系统(mesocosm) CO2加富实验发现, 与350 mg·kg-1相比, 1 050 mg·kg-1 CO2浓度下的浮游植物群落在12天内多消耗了39%的无机碳, 且浮游植物的C:N从6升高到8, 这一方面将有利于提高生物碳泵的效率, 但另一方面C:N的增加会降低有机物的营养价值, 影响细菌降解与浮游植物繁殖, 改变海洋生态系统的动力学. 此外, 酸化与光辐射(Suggett et al., 2013; Shi et al., 2015; Kleynhans et al., 2016)以及酸化与暖化(Pörtner, 2008; Mayor et al., 2012; Kroeker et al., 2013)之间的耦合作用, 也会从个体、种群、群落等不同水平上对海洋生态系统产生影响. ...
Reduced calcification of marine plankton in response to increased atmospheric CO2
1
2000
... 当前, 表层海水中CO2的浓度为10-20 µmol·L-1 (Badger et al., 1998), 低于绝大多数浮游植物中催化固碳反应的关键酶——核酮糖-1,5-二磷酸羧化酶/加氧酶(RubisCO)的半饱和常数.因此, 大多数浮游植物依赖于耗能的CO2浓缩机制(CO2-concentrating mechanism, CCM)以提高胞内RubisCO周围CO2的浓度(Reinfelder, 2011).已有研究表明, 酸化条件下, CO2浓度升高可使CCM下调, 节省能量, 从而对部分浮游植物的生长具有不同程度的促进作用(Riebesell et al., 1993; Mackey et al., 2015).除了CO2浓度升高, 随之降低的海水pH则同样可能对浮游植物产生直接或间接的影响.例如, 近期我国学者对海洋优势固氮蓝藻束毛藻(Trichodesmium)开展的研究表明, 海水pH下降导致其固氮酶效率降低, 干扰胞内pH稳态, 影响细胞产能, 且该负效应大于CO2浓度升高所产生的正效应, 故酸化的净效应为抑制束毛藻的固氮作用(Hong et al., 2017).此外, 海水pH下降还可能改变浮游植物对铁、磷等关键营养元素的利用(Shi et al., 2010; McQuaid et al., 2018), 从而间接影响浮游植物生长.除了光合作用和固氮作用外, 海水酸化还会对颗石藻、有孔虫类、珊瑚等钙化生物的钙化作用产生负面影响(Riebesell et al., 2000; Cao & Caldeira, 2008; Hofmann et al., 2010; Kroeker et al., 2013).对于海洋软体动物、棘皮动物、甲壳动物及鱼类等生物而言, 海洋酸化不仅会降低其幼体发育速度和个体大小(Ross et al., 2011), 还会降低部分生物类群的呼吸速率, 扰乱生物体内酸碱平衡, 影响其生长与繁殖(Ishimatsu et al., 2004; Pörtner & Langenbuch, 2005).海洋酸化对生物个体的影响会进一步体现在对海洋生态系统和海洋生物地球化学循环的影响.Riebesell等(2007)通过开展中尺度生态系统(mesocosm) CO2加富实验发现, 与350 mg·kg-1相比, 1 050 mg·kg-1 CO2浓度下的浮游植物群落在12天内多消耗了39%的无机碳, 且浮游植物的C:N从6升高到8, 这一方面将有利于提高生物碳泵的效率, 但另一方面C:N的增加会降低有机物的营养价值, 影响细菌降解与浮游植物繁殖, 改变海洋生态系统的动力学. 此外, 酸化与光辐射(Suggett et al., 2013; Shi et al., 2015; Kleynhans et al., 2016)以及酸化与暖化(Pörtner, 2008; Mayor et al., 2012; Kroeker et al., 2013)之间的耦合作用, 也会从个体、种群、群落等不同水平上对海洋生态系统产生影响. ...
Constraints and perspectives in aquaculture development
1
1985
... 海水富营养化会刺激浮游植物和大型藻类的生长, 增加海洋CO2的捕获量, 但过高的浮游植物生物量, 会降低光的穿透性, 影响水下生物的生长.此外, 营养盐比例的不平衡会改变浮游植物群落组成, 可能由此促进有毒藻类大量繁殖, 导致藻华的爆发(Hallegraeff, 2010; Flynn et al., 2015).有害藻类的大量繁殖不仅会导致鱼类和贝类等中毒死亡, 还会扩大海水的缺氧区(Rosenthal, 1985; Naylor et al., 2000; Rabalais et al., 2010).有研究表明, 1969-2009年期间, 由于人类活动的干扰, 大洋缺氧区的面积大小呈现递增趋势(Rabalais et al., 2010).以墨西哥北部湾缺氧区为例, 其面积从1985年的40 km2左右扩大到2015年的22 000 km2 (Sen Gupta et al., 1996; Bianchi et al., 2010; Turner & Rabalais, 2017).Neumann等(2012)的模型预测显示, 大洋季节性缺氧的持续时间将在21世纪末增加一倍.低氧会阻碍氮的硝化过程, 影响浮游植物对氮营养盐的吸收, 从而改变海洋初级生产力(Codispoti et al., 2001; Gruber, 2004), 并且不完全的硝化过程会导致温室气体N2O产量增多, 加快全球气候变暖的进程(Codispoti et al., 2001; Jin & Gruber, 2003; Nevison et al., 2003).此外, 低氧会导致有氧呼吸生物向外迁徙, 降低系统的生物多样性, 对系统物质循环与能量的流动产生影响, 破坏海洋生态系统的稳定性(Breitburg et al., 2009). ...
The impact of ocean acidification on reproduction, early development and settlement of marine organisms
1
2011
... 当前, 表层海水中CO2的浓度为10-20 µmol·L-1 (Badger et al., 1998), 低于绝大多数浮游植物中催化固碳反应的关键酶——核酮糖-1,5-二磷酸羧化酶/加氧酶(RubisCO)的半饱和常数.因此, 大多数浮游植物依赖于耗能的CO2浓缩机制(CO2-concentrating mechanism, CCM)以提高胞内RubisCO周围CO2的浓度(Reinfelder, 2011).已有研究表明, 酸化条件下, CO2浓度升高可使CCM下调, 节省能量, 从而对部分浮游植物的生长具有不同程度的促进作用(Riebesell et al., 1993; Mackey et al., 2015).除了CO2浓度升高, 随之降低的海水pH则同样可能对浮游植物产生直接或间接的影响.例如, 近期我国学者对海洋优势固氮蓝藻束毛藻(Trichodesmium)开展的研究表明, 海水pH下降导致其固氮酶效率降低, 干扰胞内pH稳态, 影响细胞产能, 且该负效应大于CO2浓度升高所产生的正效应, 故酸化的净效应为抑制束毛藻的固氮作用(Hong et al., 2017).此外, 海水pH下降还可能改变浮游植物对铁、磷等关键营养元素的利用(Shi et al., 2010; McQuaid et al., 2018), 从而间接影响浮游植物生长.除了光合作用和固氮作用外, 海水酸化还会对颗石藻、有孔虫类、珊瑚等钙化生物的钙化作用产生负面影响(Riebesell et al., 2000; Cao & Caldeira, 2008; Hofmann et al., 2010; Kroeker et al., 2013).对于海洋软体动物、棘皮动物、甲壳动物及鱼类等生物而言, 海洋酸化不仅会降低其幼体发育速度和个体大小(Ross et al., 2011), 还会降低部分生物类群的呼吸速率, 扰乱生物体内酸碱平衡, 影响其生长与繁殖(Ishimatsu et al., 2004; Pörtner & Langenbuch, 2005).海洋酸化对生物个体的影响会进一步体现在对海洋生态系统和海洋生物地球化学循环的影响.Riebesell等(2007)通过开展中尺度生态系统(mesocosm) CO2加富实验发现, 与350 mg·kg-1相比, 1 050 mg·kg-1 CO2浓度下的浮游植物群落在12天内多消耗了39%的无机碳, 且浮游植物的C:N从6升高到8, 这一方面将有利于提高生物碳泵的效率, 但另一方面C:N的增加会降低有机物的营养价值, 影响细菌降解与浮游植物繁殖, 改变海洋生态系统的动力学. 此外, 酸化与光辐射(Suggett et al., 2013; Shi et al., 2015; Kleynhans et al., 2016)以及酸化与暖化(Pörtner, 2008; Mayor et al., 2012; Kroeker et al., 2013)之间的耦合作用, 也会从个体、种群、群落等不同水平上对海洋生态系统产生影响. ...
Geologic history of sea water: an attempt to state the problem
1
1951
... 对全球气候变化的研究历史最早可以追溯到19世纪.早在19世纪20年代, 科学家就已经发现了几种温室气体, 但并未引起广泛的关注.直到19世纪30年代, 科学家意识到燃烧化石燃料对气候的影响后, 便开展了一系列研究工作.之后随着科学技术的发展, 全球气候变化对生态系统影响的研究于20世纪90年代迎来了黄金期(Weart, 2003), 其中就包括对海洋生态系统影响的研究.以全球气候变化导致的海洋酸化为例, 早在1906年, 针对海洋酸碱性, Moore等(1906)就已经开展了酸碱性对海胆受精卵的生长和细胞分裂的影响研究; 1951年, Rubey总结了早期酸化的研究工作, 为后人的研究提供了理论基础; 1957年, Revelle和Suess首次提出海洋吸收人类排放的CO2会降低海洋的缓冲能力, 引起了科学界对人类活动影响海洋生态系统的重视; 1966年, Broecker和Takahashi首次报道了海水CO32-的浓度, 并提出其将随着海水CO2浓度的升高而降低; 1978年, Brewer提出利用总碱度和溶解无机碳的浓度校正海水中的CO2系统; 1993年硼同位素等古海洋示踪剂开始被拓展应用于估算海洋化学参数的变化(Spivack et al., 1993); 2001年, Bates第一次利用时间序列数据记录了北大西洋西部海水碳酸盐体系的变化,主要包括海水的CO2浓度、CO2分压以及碱度的变动; 近年来, 利用受控实验评估生物体对海水碳酸盐体系变化响应的论文大量涌现, 与此同时科学家开始通过构建数值模型来预测未来海水碳酸盐系统的变化以及海洋酸化对海洋生物地球化学循环的影响. ...
Seasonal oxygen depletion in the continental-shelf waters of Louisiana: historical record of benthic foraminifers
1
1996
... 海水富营养化会刺激浮游植物和大型藻类的生长, 增加海洋CO2的捕获量, 但过高的浮游植物生物量, 会降低光的穿透性, 影响水下生物的生长.此外, 营养盐比例的不平衡会改变浮游植物群落组成, 可能由此促进有毒藻类大量繁殖, 导致藻华的爆发(Hallegraeff, 2010; Flynn et al., 2015).有害藻类的大量繁殖不仅会导致鱼类和贝类等中毒死亡, 还会扩大海水的缺氧区(Rosenthal, 1985; Naylor et al., 2000; Rabalais et al., 2010).有研究表明, 1969-2009年期间, 由于人类活动的干扰, 大洋缺氧区的面积大小呈现递增趋势(Rabalais et al., 2010).以墨西哥北部湾缺氧区为例, 其面积从1985年的40 km2左右扩大到2015年的22 000 km2 (Sen Gupta et al., 1996; Bianchi et al., 2010; Turner & Rabalais, 2017).Neumann等(2012)的模型预测显示, 大洋季节性缺氧的持续时间将在21世纪末增加一倍.低氧会阻碍氮的硝化过程, 影响浮游植物对氮营养盐的吸收, 从而改变海洋初级生产力(Codispoti et al., 2001; Gruber, 2004), 并且不完全的硝化过程会导致温室气体N2O产量增多, 加快全球气候变暖的进程(Codispoti et al., 2001; Jin & Gruber, 2003; Nevison et al., 2003).此外, 低氧会导致有氧呼吸生物向外迁徙, 降低系统的生物多样性, 对系统物质循环与能量的流动产生影响, 破坏海洋生态系统的稳定性(Breitburg et al., 2009). ...
Interactive effects of light, nitrogen source, and carbon dioxide on energy metabolism in the diatom Thalassiosira pseudonana
2
2015
... 当前, 表层海水中CO2的浓度为10-20 µmol·L-1 (Badger et al., 1998), 低于绝大多数浮游植物中催化固碳反应的关键酶——核酮糖-1,5-二磷酸羧化酶/加氧酶(RubisCO)的半饱和常数.因此, 大多数浮游植物依赖于耗能的CO2浓缩机制(CO2-concentrating mechanism, CCM)以提高胞内RubisCO周围CO2的浓度(Reinfelder, 2011).已有研究表明, 酸化条件下, CO2浓度升高可使CCM下调, 节省能量, 从而对部分浮游植物的生长具有不同程度的促进作用(Riebesell et al., 1993; Mackey et al., 2015).除了CO2浓度升高, 随之降低的海水pH则同样可能对浮游植物产生直接或间接的影响.例如, 近期我国学者对海洋优势固氮蓝藻束毛藻(Trichodesmium)开展的研究表明, 海水pH下降导致其固氮酶效率降低, 干扰胞内pH稳态, 影响细胞产能, 且该负效应大于CO2浓度升高所产生的正效应, 故酸化的净效应为抑制束毛藻的固氮作用(Hong et al., 2017).此外, 海水pH下降还可能改变浮游植物对铁、磷等关键营养元素的利用(Shi et al., 2010; McQuaid et al., 2018), 从而间接影响浮游植物生长.除了光合作用和固氮作用外, 海水酸化还会对颗石藻、有孔虫类、珊瑚等钙化生物的钙化作用产生负面影响(Riebesell et al., 2000; Cao & Caldeira, 2008; Hofmann et al., 2010; Kroeker et al., 2013).对于海洋软体动物、棘皮动物、甲壳动物及鱼类等生物而言, 海洋酸化不仅会降低其幼体发育速度和个体大小(Ross et al., 2011), 还会降低部分生物类群的呼吸速率, 扰乱生物体内酸碱平衡, 影响其生长与繁殖(Ishimatsu et al., 2004; Pörtner & Langenbuch, 2005).海洋酸化对生物个体的影响会进一步体现在对海洋生态系统和海洋生物地球化学循环的影响.Riebesell等(2007)通过开展中尺度生态系统(mesocosm) CO2加富实验发现, 与350 mg·kg-1相比, 1 050 mg·kg-1 CO2浓度下的浮游植物群落在12天内多消耗了39%的无机碳, 且浮游植物的C:N从6升高到8, 这一方面将有利于提高生物碳泵的效率, 但另一方面C:N的增加会降低有机物的营养价值, 影响细菌降解与浮游植物繁殖, 改变海洋生态系统的动力学. 此外, 酸化与光辐射(Suggett et al., 2013; Shi et al., 2015; Kleynhans et al., 2016)以及酸化与暖化(Pörtner, 2008; Mayor et al., 2012; Kroeker et al., 2013)之间的耦合作用, 也会从个体、种群、群落等不同水平上对海洋生态系统产生影响. ...
... 目前国际上对海洋酸化的研究取得了一定的进展, 尤其是近年来越来越多的研究开始关注温度、光照、营养盐条件等其他环境因子的变化与海洋酸化耦合的影响(Shi et al., 2015; Hutchins & Boyd, 2016).然而, 以下几个关键科学问题仍然有待各个国家、学术组织之中的科学家通过加强合作交流以及在个体-种群-群落-生态系统等尺度上开展整合研究而予以阐明. ...
Effect of ocean acidification on iron availability to marine phytoplankton
1
2010
... 当前, 表层海水中CO2的浓度为10-20 µmol·L-1 (Badger et al., 1998), 低于绝大多数浮游植物中催化固碳反应的关键酶——核酮糖-1,5-二磷酸羧化酶/加氧酶(RubisCO)的半饱和常数.因此, 大多数浮游植物依赖于耗能的CO2浓缩机制(CO2-concentrating mechanism, CCM)以提高胞内RubisCO周围CO2的浓度(Reinfelder, 2011).已有研究表明, 酸化条件下, CO2浓度升高可使CCM下调, 节省能量, 从而对部分浮游植物的生长具有不同程度的促进作用(Riebesell et al., 1993; Mackey et al., 2015).除了CO2浓度升高, 随之降低的海水pH则同样可能对浮游植物产生直接或间接的影响.例如, 近期我国学者对海洋优势固氮蓝藻束毛藻(Trichodesmium)开展的研究表明, 海水pH下降导致其固氮酶效率降低, 干扰胞内pH稳态, 影响细胞产能, 且该负效应大于CO2浓度升高所产生的正效应, 故酸化的净效应为抑制束毛藻的固氮作用(Hong et al., 2017).此外, 海水pH下降还可能改变浮游植物对铁、磷等关键营养元素的利用(Shi et al., 2010; McQuaid et al., 2018), 从而间接影响浮游植物生长.除了光合作用和固氮作用外, 海水酸化还会对颗石藻、有孔虫类、珊瑚等钙化生物的钙化作用产生负面影响(Riebesell et al., 2000; Cao & Caldeira, 2008; Hofmann et al., 2010; Kroeker et al., 2013).对于海洋软体动物、棘皮动物、甲壳动物及鱼类等生物而言, 海洋酸化不仅会降低其幼体发育速度和个体大小(Ross et al., 2011), 还会降低部分生物类群的呼吸速率, 扰乱生物体内酸碱平衡, 影响其生长与繁殖(Ishimatsu et al., 2004; Pörtner & Langenbuch, 2005).海洋酸化对生物个体的影响会进一步体现在对海洋生态系统和海洋生物地球化学循环的影响.Riebesell等(2007)通过开展中尺度生态系统(mesocosm) CO2加富实验发现, 与350 mg·kg-1相比, 1 050 mg·kg-1 CO2浓度下的浮游植物群落在12天内多消耗了39%的无机碳, 且浮游植物的C:N从6升高到8, 这一方面将有利于提高生物碳泵的效率, 但另一方面C:N的增加会降低有机物的营养价值, 影响细菌降解与浮游植物繁殖, 改变海洋生态系统的动力学. 此外, 酸化与光辐射(Suggett et al., 2013; Shi et al., 2015; Kleynhans et al., 2016)以及酸化与暖化(Pörtner, 2008; Mayor et al., 2012; Kroeker et al., 2013)之间的耦合作用, 也会从个体、种群、群落等不同水平上对海洋生态系统产生影响. ...
Foraminiferal boron isotope ratios as a proxy for surface ocean pH over the past 21 Myr
1
1993
... 对全球气候变化的研究历史最早可以追溯到19世纪.早在19世纪20年代, 科学家就已经发现了几种温室气体, 但并未引起广泛的关注.直到19世纪30年代, 科学家意识到燃烧化石燃料对气候的影响后, 便开展了一系列研究工作.之后随着科学技术的发展, 全球气候变化对生态系统影响的研究于20世纪90年代迎来了黄金期(Weart, 2003), 其中就包括对海洋生态系统影响的研究.以全球气候变化导致的海洋酸化为例, 早在1906年, 针对海洋酸碱性, Moore等(1906)就已经开展了酸碱性对海胆受精卵的生长和细胞分裂的影响研究; 1951年, Rubey总结了早期酸化的研究工作, 为后人的研究提供了理论基础; 1957年, Revelle和Suess首次提出海洋吸收人类排放的CO2会降低海洋的缓冲能力, 引起了科学界对人类活动影响海洋生态系统的重视; 1966年, Broecker和Takahashi首次报道了海水CO32-的浓度, 并提出其将随着海水CO2浓度的升高而降低; 1978年, Brewer提出利用总碱度和溶解无机碳的浓度校正海水中的CO2系统; 1993年硼同位素等古海洋示踪剂开始被拓展应用于估算海洋化学参数的变化(Spivack et al., 1993); 2001年, Bates第一次利用时间序列数据记录了北大西洋西部海水碳酸盐体系的变化,主要包括海水的CO2浓度、CO2分压以及碱度的变动; 近年来, 利用受控实验评估生物体对海水碳酸盐体系变化响应的论文大量涌现, 与此同时科学家开始通过构建数值模型来预测未来海水碳酸盐系统的变化以及海洋酸化对海洋生物地球化学循环的影响. ...
Projected 21st century decrease in marine productivity: a multi-model analysis
1
2010
... 暖化的海水对生物的影响因种而异.对于大多数浮游植物而言, 在适温范围内, 海水暖化会促进其生长(Eppley, 1972), 但不同浮游植物对温度变化的响应程度不同, 因此全球暖化下浮游植物的生态位将发生改变(Xiao et al., 2018).然而, 对于大多数海洋动物, 暖化将在增加其体内代谢活性的同时加快其能量的消耗, 从而可能导致海洋生物自身生长与繁殖能力的下降(Pörtner & Knust, 2007).这些个体生理生化过程的变化会进一步改变种群的大小、时空分布格局与营养级水平, 最终引起群落结构与功能的改变(Bruno et al., 2003; Hyrenbach & Veit, 2003).从海洋生态系统的层面上来看, 全球变暖会改变海洋的初级生产力, 影响生态系统的稳定性.近期我国科学家采用模型估算发现, 1948-2007年, 全球气候变暖导致海洋水体层化加剧, 使得北大西洋、北太平洋和印度洋初级生产力分别降低了40%、24%和25% (Wang et al., 2017).倘若暖化现象一直持续, 到2100年, 海洋初级生产力将可能进一步减少20% (Steinacher et al., 2010).此外, 已有研究表明, 海水温度升高1 ℃, 珊瑚即会出现白化现象(Hoegh- Guldberg et al., 2007), 这会导致珊瑚礁生态系统多样性降低, 严重时会引发珊瑚礁系统的崩溃(Donner, 2009).海水暖化除了通过温度增加而对海洋生态系统产生直接影响外, 还会造成海洋层化加剧、缺氧区扩增等, 从而间接地对海洋生态系统产生一系列影响(Keeling et al., 2010; Wang et al., 2017). ...
Light availability determines susceptibility of reef building corals to ocean acidification
1
2013
... 当前, 表层海水中CO2的浓度为10-20 µmol·L-1 (Badger et al., 1998), 低于绝大多数浮游植物中催化固碳反应的关键酶——核酮糖-1,5-二磷酸羧化酶/加氧酶(RubisCO)的半饱和常数.因此, 大多数浮游植物依赖于耗能的CO2浓缩机制(CO2-concentrating mechanism, CCM)以提高胞内RubisCO周围CO2的浓度(Reinfelder, 2011).已有研究表明, 酸化条件下, CO2浓度升高可使CCM下调, 节省能量, 从而对部分浮游植物的生长具有不同程度的促进作用(Riebesell et al., 1993; Mackey et al., 2015).除了CO2浓度升高, 随之降低的海水pH则同样可能对浮游植物产生直接或间接的影响.例如, 近期我国学者对海洋优势固氮蓝藻束毛藻(Trichodesmium)开展的研究表明, 海水pH下降导致其固氮酶效率降低, 干扰胞内pH稳态, 影响细胞产能, 且该负效应大于CO2浓度升高所产生的正效应, 故酸化的净效应为抑制束毛藻的固氮作用(Hong et al., 2017).此外, 海水pH下降还可能改变浮游植物对铁、磷等关键营养元素的利用(Shi et al., 2010; McQuaid et al., 2018), 从而间接影响浮游植物生长.除了光合作用和固氮作用外, 海水酸化还会对颗石藻、有孔虫类、珊瑚等钙化生物的钙化作用产生负面影响(Riebesell et al., 2000; Cao & Caldeira, 2008; Hofmann et al., 2010; Kroeker et al., 2013).对于海洋软体动物、棘皮动物、甲壳动物及鱼类等生物而言, 海洋酸化不仅会降低其幼体发育速度和个体大小(Ross et al., 2011), 还会降低部分生物类群的呼吸速率, 扰乱生物体内酸碱平衡, 影响其生长与繁殖(Ishimatsu et al., 2004; Pörtner & Langenbuch, 2005).海洋酸化对生物个体的影响会进一步体现在对海洋生态系统和海洋生物地球化学循环的影响.Riebesell等(2007)通过开展中尺度生态系统(mesocosm) CO2加富实验发现, 与350 mg·kg-1相比, 1 050 mg·kg-1 CO2浓度下的浮游植物群落在12天内多消耗了39%的无机碳, 且浮游植物的C:N从6升高到8, 这一方面将有利于提高生物碳泵的效率, 但另一方面C:N的增加会降低有机物的营养价值, 影响细菌降解与浮游植物繁殖, 改变海洋生态系统的动力学. 此外, 酸化与光辐射(Suggett et al., 2013; Shi et al., 2015; Kleynhans et al., 2016)以及酸化与暖化(Pörtner, 2008; Mayor et al., 2012; Kroeker et al., 2013)之间的耦合作用, 也会从个体、种群、群落等不同水平上对海洋生态系统产生影响. ...
1
2017
... 海水富营养化会刺激浮游植物和大型藻类的生长, 增加海洋CO2的捕获量, 但过高的浮游植物生物量, 会降低光的穿透性, 影响水下生物的生长.此外, 营养盐比例的不平衡会改变浮游植物群落组成, 可能由此促进有毒藻类大量繁殖, 导致藻华的爆发(Hallegraeff, 2010; Flynn et al., 2015).有害藻类的大量繁殖不仅会导致鱼类和贝类等中毒死亡, 还会扩大海水的缺氧区(Rosenthal, 1985; Naylor et al., 2000; Rabalais et al., 2010).有研究表明, 1969-2009年期间, 由于人类活动的干扰, 大洋缺氧区的面积大小呈现递增趋势(Rabalais et al., 2010).以墨西哥北部湾缺氧区为例, 其面积从1985年的40 km2左右扩大到2015年的22 000 km2 (Sen Gupta et al., 1996; Bianchi et al., 2010; Turner & Rabalais, 2017).Neumann等(2012)的模型预测显示, 大洋季节性缺氧的持续时间将在21世纪末增加一倍.低氧会阻碍氮的硝化过程, 影响浮游植物对氮营养盐的吸收, 从而改变海洋初级生产力(Codispoti et al., 2001; Gruber, 2004), 并且不完全的硝化过程会导致温室气体N2O产量增多, 加快全球气候变暖的进程(Codispoti et al., 2001; Jin & Gruber, 2003; Nevison et al., 2003).此外, 低氧会导致有氧呼吸生物向外迁徙, 降低系统的生物多样性, 对系统物质循环与能量的流动产生影响, 破坏海洋生态系统的稳定性(Breitburg et al., 2009). ...
Modern dust aerosol availability in northwestern China
2
2017
... 暖化的海水对生物的影响因种而异.对于大多数浮游植物而言, 在适温范围内, 海水暖化会促进其生长(Eppley, 1972), 但不同浮游植物对温度变化的响应程度不同, 因此全球暖化下浮游植物的生态位将发生改变(Xiao et al., 2018).然而, 对于大多数海洋动物, 暖化将在增加其体内代谢活性的同时加快其能量的消耗, 从而可能导致海洋生物自身生长与繁殖能力的下降(Pörtner & Knust, 2007).这些个体生理生化过程的变化会进一步改变种群的大小、时空分布格局与营养级水平, 最终引起群落结构与功能的改变(Bruno et al., 2003; Hyrenbach & Veit, 2003).从海洋生态系统的层面上来看, 全球变暖会改变海洋的初级生产力, 影响生态系统的稳定性.近期我国科学家采用模型估算发现, 1948-2007年, 全球气候变暖导致海洋水体层化加剧, 使得北大西洋、北太平洋和印度洋初级生产力分别降低了40%、24%和25% (Wang et al., 2017).倘若暖化现象一直持续, 到2100年, 海洋初级生产力将可能进一步减少20% (Steinacher et al., 2010).此外, 已有研究表明, 海水温度升高1 ℃, 珊瑚即会出现白化现象(Hoegh- Guldberg et al., 2007), 这会导致珊瑚礁生态系统多样性降低, 严重时会引发珊瑚礁系统的崩溃(Donner, 2009).海水暖化除了通过温度增加而对海洋生态系统产生直接影响外, 还会造成海洋层化加剧、缺氧区扩增等, 从而间接地对海洋生态系统产生一系列影响(Keeling et al., 2010; Wang et al., 2017). ...
... ; Wang et al., 2017). ...
1
2003
... 对全球气候变化的研究历史最早可以追溯到19世纪.早在19世纪20年代, 科学家就已经发现了几种温室气体, 但并未引起广泛的关注.直到19世纪30年代, 科学家意识到燃烧化石燃料对气候的影响后, 便开展了一系列研究工作.之后随着科学技术的发展, 全球气候变化对生态系统影响的研究于20世纪90年代迎来了黄金期(Weart, 2003), 其中就包括对海洋生态系统影响的研究.以全球气候变化导致的海洋酸化为例, 早在1906年, 针对海洋酸碱性, Moore等(1906)就已经开展了酸碱性对海胆受精卵的生长和细胞分裂的影响研究; 1951年, Rubey总结了早期酸化的研究工作, 为后人的研究提供了理论基础; 1957年, Revelle和Suess首次提出海洋吸收人类排放的CO2会降低海洋的缓冲能力, 引起了科学界对人类活动影响海洋生态系统的重视; 1966年, Broecker和Takahashi首次报道了海水CO32-的浓度, 并提出其将随着海水CO2浓度的升高而降低; 1978年, Brewer提出利用总碱度和溶解无机碳的浓度校正海水中的CO2系统; 1993年硼同位素等古海洋示踪剂开始被拓展应用于估算海洋化学参数的变化(Spivack et al., 1993); 2001年, Bates第一次利用时间序列数据记录了北大西洋西部海水碳酸盐体系的变化,主要包括海水的CO2浓度、CO2分压以及碱度的变动; 近年来, 利用受控实验评估生物体对海水碳酸盐体系变化响应的论文大量涌现, 与此同时科学家开始通过构建数值模型来预测未来海水碳酸盐系统的变化以及海洋酸化对海洋生物地球化学循环的影响. ...
Realized niches explain spatial gradients in seasonal abundance of phytoplankton groups in the South China Sea
1
2018
... 暖化的海水对生物的影响因种而异.对于大多数浮游植物而言, 在适温范围内, 海水暖化会促进其生长(Eppley, 1972), 但不同浮游植物对温度变化的响应程度不同, 因此全球暖化下浮游植物的生态位将发生改变(Xiao et al., 2018).然而, 对于大多数海洋动物, 暖化将在增加其体内代谢活性的同时加快其能量的消耗, 从而可能导致海洋生物自身生长与繁殖能力的下降(Pörtner & Knust, 2007).这些个体生理生化过程的变化会进一步改变种群的大小、时空分布格局与营养级水平, 最终引起群落结构与功能的改变(Bruno et al., 2003; Hyrenbach & Veit, 2003).从海洋生态系统的层面上来看, 全球变暖会改变海洋的初级生产力, 影响生态系统的稳定性.近期我国科学家采用模型估算发现, 1948-2007年, 全球气候变暖导致海洋水体层化加剧, 使得北大西洋、北太平洋和印度洋初级生产力分别降低了40%、24%和25% (Wang et al., 2017).倘若暖化现象一直持续, 到2100年, 海洋初级生产力将可能进一步减少20% (Steinacher et al., 2010).此外, 已有研究表明, 海水温度升高1 ℃, 珊瑚即会出现白化现象(Hoegh- Guldberg et al., 2007), 这会导致珊瑚礁生态系统多样性降低, 严重时会引发珊瑚礁系统的崩溃(Donner, 2009).海水暖化除了通过温度增加而对海洋生态系统产生直接影响外, 还会造成海洋层化加剧、缺氧区扩增等, 从而间接地对海洋生态系统产生一系列影响(Keeling et al., 2010; Wang et al., 2017). ...






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}