




放牧是全世界最广泛的人为土地利用方式(Díaz et al., 2007; Michaels et al., 2021)。全球超1/2的陆地均为放牧地, 放牧生态系统不仅为人类提供1/2的食用肉类和1/3以上的奶类(张扬建等, 2020), 同时还具备调节气候、水土保持、维持生物多样性和碳固存等功能, 具有重要的生态、经济和社会效益(García et al., 2012; Wu et al., 2020)。然而, 无序和不当的放牧管理所导致的植被退化和服务功能下降会损害生态系统的可持续发展(García et al., 2012; 王斌等, 2012; Zhang et al., 2018)。放牧对生态系统的结构和功能的影响一直是生态学研究的核心问题(Škornik et al., 2010)。
物种组成和群落结构作为植被的两个最基本特征, 可反映群落种间关系和环境对物种生存与生长的影响, 在揭示和探讨群落构建与生态系统功能等多个方面具有重要作用(刘长成等, 2009; 赵广东等, 2021)。放牧会改变植被的物种组成和群落结构, 随着放牧强度的增加, 优势种(功能群)发生更替(杨浩等, 2009), 或虽未发生更替, 但表现出特征值(如重要值)的变化(赵生龙等, 2020; Michaels et al., 2021); 物种丰富度表现为增加、减少或中性(苗福泓等, 2016; Herrero-Jáuregui & Oesterheld, 2018; Gao & Carmel, 2020); 植被结构(如高度、盖度等)随之减小, 多呈现矮小化趋势(李西良等, 2014, 2015), 进而导致生物量的减少(赵彬彬等, 2009; 赵生龙等, 2020), 但在不同层次(植物群落的分层)或不同植被类型以及不同尺度上有所区别(Michaels et al., 2021)。上述研究结果的差异一方面源于放牧地环境差异, 如植被类型、地形、气候条件、研究尺度、植被进化史等, 另一方面源于放牧者的管理差异, 如放牧的强度(载畜率)、频率、对象(牲畜类型)、范围、季节等, 但上述因素中的主导因素与相对重要性仍需进一步研究(Öllerer et al., 2019)。
喀斯特生态系统受碳酸盐岩溶蚀性强、水文二元结构明显以及区域雨热分布不均等因素的综合影响, 其小生境(石沟、石缝等)的多样组合和浅薄且不连续的土被形成的高度异质性生境成为植被发育的基础(曾馥平等, 2007; 文丽等, 2015; Geekiyanage et al., 2019)。我国西南喀斯特生态系统脆弱, 近几十年人类活动的持续干扰导致其退化明显, 石漠化问题较为突出(王世杰等, 2003)。喀斯特山地现存植被多为经历干扰后恢复形成的次生草丛、灌草丛、灌丛、灌乔丛等植被类型(蒋建生等, 2008; 文丽等, 2015; 张芳等, 2020; 张忠华等, 2022)。恢复喀斯特植被是解决石漠化问题的关键所在(宋同清等, 2010; 刘玉国等, 2011)。近年来, 以植被恢复为主的喀斯特生态修复工程已取得重要成效(蒋忠诚等, 2009; Tong et al., 2018; Wu et al., 2020), 但因喀斯特生态系统脆弱性和人类活动的不合理干扰等因素, “返漠”现象仍时有发生(宋同清等, 2014)。因此, 我国西南喀斯特地区的植被恢复依然任重道远。
喀斯特生境特征、物种生物学特征、种内种间关系以及人为或自然干扰等因素共同影响着喀斯特植被结构与动态(郭柯等, 2011; 文丽等, 2015; Geekiyanage et al., 2019; Wang et al., 2019)。不同干扰方式对植被自然恢复的影响不同, 对植被破坏越大, 恢复越慢(曾馥平等, 2007)。喀斯特灌草丛群落处于植被遭严重破坏后恢复的初始阶段, 群落结构相对单一, 稳定性较差, 易受外界干扰活动影响, 干扰力超过其承受阈值时, 群落甚至会发生逆向演替(刘玉国等, 2011; 文丽等, 2015; Wang et al., 2019)。我国西南喀斯特地貌区有限的自然资源使得放牧成为喀斯特山区人们生存和经济发展的重要途径, 但放牧在改善社会经济的同时, 也存在因过度放牧而导致石漠化演变进程加快的风险。尖锐的人地矛盾是喀斯特地上植被恢复与重建工作的一大障碍(文丽等, 2015)。一直以来, 国外学者对放牧生态系统的研究主要集中在全球典型草地分布区(Díaz et al., 2007), 国内相关研究主要集中在内蒙古(温带草原和温带荒漠)和青藏高原(高寒草甸和高寒草原)(张扬建等, 2020), 而对同样拥有丰富草地资源和放牧史的喀斯特地区, 相关研究仍极为匮乏。放牧对脆弱的喀斯特生态系统会产生何种影响? 是否会阻碍喀斯特植被的恢复? 对这些科学问题的理解尚浅。
本研究选取广西马山县喀斯特山地的灌草丛植被, 根据放牧强度(重牧、中牧、轻牧、无放牧)设置样方并开展群落调查, 从而分析植被物种组成与群落结构等植被特征变化。本研究旨在探讨如下问题: 1)喀斯特山地灌草丛的物种组成如何响应放牧干扰的强度变化? 2)放牧强度的增加如何影响灌草丛群落的高度、盖度和地上生物量? 探究喀斯特山地植被对放牧干扰的响应格局, 对进一步揭示喀斯特生态系统的退化机理以及指导植被恢复和重建具有重要意义。
1 材料和方法
1.1 研究区概况
研究地点位于广西马山县(107.71°-108.50° E, 23.40°-24.04° N)的古零镇和永州镇。该县位于广西中北部, 是典型的喀斯特峰丛洼地地貌区, 其喀斯特面积为16.3万hm2, 占全县土地面积的69.5% (邹知明等, 2012)。马山县属南亚热带湿润性季风气候,平均年日照时间1 461 h, 年降水量1 722 mm, 降水多集中在5-8月, 年平均气温21.80 ℃。土壤类型以棕色石灰土为主。马山县石漠化、潜在石漠化和非石漠化土地面积分别占马山县总喀斯特面积的21.9%、45.2%和32.9% (邓必玉等, 2012)。原生性植被为土壤顶极常绿落叶阔叶混交林, 但在人为干扰、自然灾害等多重因素影响下, 现存植被多为稀树灌丛、灌草丛和人工林。喀斯特灌草丛面积占全县天然次生草地面积的87.59%, 占全县土地总面积的49.67% (邹知明等, 2012), 灌草丛资源丰富。灌草丛常见种包括黄荆(Vitex negundo)、芒(Miscanthus sinensis)、雀梅藤(Sageretia thea)、红背山麻杆(Alchornea trewioides)、白茅(Imperata cylindrica)、荩草(Arthraxon hispidus)等。
马山县享有“中国黑山羊之乡”的美称, 马山黑山羊为国家地理标志产品, 山羊饲养已成为当地群众主要的经济来源。
1.2 样方选取与调查
考虑到短期控制实验(如2-3年)难以反映放牧对生态系统影响的积累效应, 以及控制实验难以模拟自然状态下的放牧活动, 为分析自然状态下放牧对喀斯特植被的影响, 本研究于2020-2021年植物生长旺盛季节(7-8月), 在马山县内严格选取具有代表性的喀斯特石山坡地作为放牧研究样地。综合考虑研究地周边散养山羊的养殖规模、山羊活动范围和放牧干扰历史等因素, 选取了7处放牧点。所选放牧点均为山羊养殖数量在100只以上, 放牧干扰达5年以上且无火烧和开垦历史的喀斯特坡地。在7处放牧点, 选取距离羊舍较近(300 m内), 植物明显有山羊长期啃食和踩踏痕迹以及地表粪便积累量大的地点设置重度放牧(HG)样方; 选取距离羊舍偏远(300-500 m), 植物受啃食和踩踏程度较重, 地表山羊粪便积累量较多的地点设置中度放牧(MG)样方; 选取距离羊舍较远(500 m以上), 植被受啃食和践踏痕迹较少且山羊粪便量少的地点设置轻度放牧(LG)样方; 无放牧(CK)样方在放牧点周边无放牧和其他人为干扰, 且生境类型与放牧点相似的地方设置。在每种放牧强度设置4-5个5 m × 5 m样方, 每个样方间隔50 m以上, 7个放牧点共设置116个样方。测定所有样方内灌木层和草本层植物的种名、高度、盖度等指标, 同时记录岩石裸露率、海拔、坡向、坡度等环境指标。
1.3 土壤理化性质的测定
按照梅花五点法采集土壤样品。在每个样方中去除表层凋落物后使用环刀(100 cm3)取表层土壤, 同时在环刀周围取100 g左右的土壤样品, 并将五点土壤混合均匀后带回实验室。称取环刀土样鲜质量, 带回实验室后对其进行烘干、称量, 计算土壤含水量(SWC)、土壤密度(Bd)、总孔隙度(Tp)。鲜土在实验室风干后过筛, 用于测定土壤化学性质。测定指标及其测定方法为: 土壤有机质(SOM)含量采用重铬酸钾容量法-外加热法测定; 全氮(TN)含量采用半微量凯氏法测定; 全磷(TP)含量采用NaOH熔融-钼锑抗比色法测定; 全钾(TK)含量采用NaOH熔融-火焰光度法测定; 全钙(TCa)和全镁(TMg)含量用四酸消煮-原子吸收火焰光谱法测定; 碱解氮(AN)含量采用碱解扩散法测定; 速效磷(AP)含量采用NaHCO3浸提-钼锑抗比色法测定; 速效钾(AK)含量采用乙酸铵浸提-火焰光度法测定; pH采用电极电位法测定。
1.4 地上生物量的测定
为估测植被地上生物量, 在每个样方临近且放牧干扰一致的地点随机设置3个1 m × 1 m的小样方, 用收获法齐地刈割小样方内植被地上部分并就地称取鲜质量, 带回实验室在80 ℃下烘干至恒质量并称取干质量, 将3个小样方内的地上生物量取平均值用于估算每个样方的地上生物量。
1.5 数据分析
式中, 相对高度= (某物种(科/属下物种)高度(和)/全部物种的高度之和) × 100, 相对盖度= (某物种(科/属下物种)盖度(和)/全部物种的盖度之和) × 100, 相对频度= (某物种频度/全部物种的频度之和) × 100, 相对多样性= (某科(属)下物种的物种数/全部物种数) × 100。
采用非度量多维尺度分析(NMDS)排序揭示放牧干扰下灌木层、草本层以及群落整体的物种组成变化及其影响因子。首先运用物种盖度计算Bray- Curtis指数, 而后基于该指数进行NMDS排序, 使用envfit函数拟合添加环境因子, 并得到环境因子与NMDS排序前两轴的相关性系数, 使用“bonferroni”法校正p值。根据协强系数(Stress)判断排序拟合结果, Stress ≤ 0.2表明拟合可接受; Stress > 0.2表明拟合较差, 解释力较弱, 需考虑添加排序轴数量(Kruskal, 1964; 邓建明等, 2016; 山丹等, 2019)。环境因子包括放牧干扰强度(自无牧区至重牧区分别采用1-4表示)、地形因子(岩石裸露率、海拔、坡向、坡度)和土壤因子(Bd、SWC、Tp、SOM含量、TN含量、TP含量、TK含量、TCa含量、TMg含量、AN含量、AP含量、AK含量和pH), 共计18个环境因子。其中坡向数据经过坡向转换指数(TRASP) = (1 - cos((π/180)×(坡向-30)))/2转换为0-1表示(余敏等, 2013)。NMDS排序图中, 样点用圆点表示, 点越近相似程度越大; 排序椭圆越小表示该放牧强度内的物种组成越相似。环境因子用带箭头的线段表示, 线段长短对应环境因子与样点的相关性大小, 线段与排序轴或线段之间的夹角越小, 表明该环境因子与排序轴或另一环境因子的相关性越大, 当夹角为钝角时, 表明相关性为负(徐远杰等, 2017)。上述数据分析均在“vegan”包(Oksanen et al., 2022)中完成。
采用单因素方差分析和非参数检验分析不同放牧强度对群落高度、盖度和地上生物量的影响。首先检验数据的正态性和方差齐性, 如数据满足分析条件则采用单因素方差分析, 使用最小显著差异(LSD)法进行显著性水平(α = 0.05)检验。不符合分析条件的数据, 采用Kruskal-Wails非参数检验进行差异性比较, 对差异显著的数据进行成对比较。上述数据分析及绘图均在R 4.0.3软件中完成。
2 结果
2.1 放牧对灌草丛物种组成的影响
调查表明, 样方内有维管植物272种, 隶属77科200属, 其中灌木层97种, 隶属39科74属, 草本层234种, 隶属68科180属。灌木层科、属、种数量随放牧强度增加而明显锐减(图1A)。从重要值看, 灌木层优势科分别为马鞭草科、豆科和鼠李科等(附录I), 优势属为牡荆属(Vitex)、雀梅藤属(Sageretia)和酸藤子属(Embelia)等(附录II), 优势种包括黄荆、雀梅藤和柘(Maclura tricuspidata)等(附录III)。黄荆及其所在科属(马鞭草科牡荆属)的重要值均随放牧强度的增加而增加, 大戟科、报春花科, 酸藤子属、杜茎山属(Maesa)、紫珠属(Callicarpa)重要值表现为单峰变化, 轻牧区达峰值。
图1
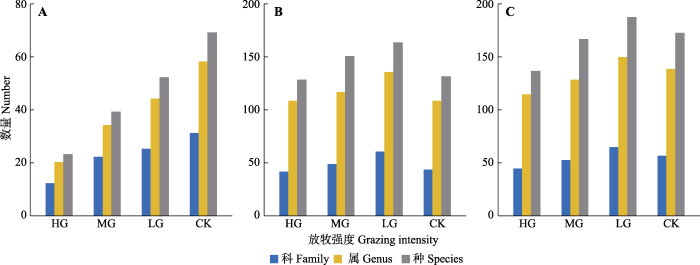
图1
中国亚热带喀斯特山地灌草丛不同放牧强度下的植物科、属、种总数。A, 灌木层。B, 草本层。C, 群落整体。CK, 无放牧; HG, 重度放牧; LG, 轻度放牧; MG, 中度放牧。
Fig. 1
Total numbers of plant families, genera and species of shrub tussock under different grazing intensities in subtropical karst mountains in southwest China. A, Shrub layer. B, Herb layer. C, Total community. CK, none grazing; HG, heavy grazing; LG, light grazing; MG, moderate grazing.
草本层科、属、种数量在放牧影响下呈单峰变化趋势, 峰值出现在轻牧区(图1B)。科水平上, 禾本科(22属27种)、菊科(20属27种)、豆科(17属26种)植物物种数最多。属水平上, 榕属(Ficus, 7种)物种数最多, 其次是山蚂蟥属(Desmodium, 5种)和蒿属(Artemisia, 4种)等。从重要值看, 优势科为禾本科、菊科、肾蕨科和豆科(附录I), 优势属为荩草属(Arthraxon)、莠竹属(Microstegium)、泽兰属(Eupatorium)等(附录II), 优势种为矛叶荩草(Arthraxon lanceolatus)、肾蕨(Nephrolepis cordifolia)、蔓生莠竹(Microstegium fasciculatum)和飞机草(Chromolaena odorata)等(附录III)。菊科、豆科和马鞭草科及其下的属(如鸡眼草属(Kummerowia)、蒿属等)和种(如艾(Artemisia argyi)、白莲蒿(Artemisia stechmanniana)和鸡眼草(Kummerowia striata)等)重要值随放牧强度增加而增加。大戟科、桑科、叶下珠科, 莠竹属、芒属(Miscanthus)、榕属, 芒、野桐(Mallotus tenuifolius)和红背山麻杆等的重要值随放牧强度增加而减小。
群落整体的科属种数量在放牧影响下同草本层一样呈单峰变化, 轻牧区达峰值(图1C)。物种水平上排名靠前的科和属及其变化趋势也同草本层较为一致。科、属、种水平上排名前10的重要值之和变化呈偏“U”形, 自轻牧区到重牧区, 增幅均小于11% (附录I-III)。
NMDS排序结果(图2)表明, 灌木层、草本层和群落整体的排序拟合效果均相对理想(Stress < 0.2)。多数样方间的物种组成趋于相似, 但同一放牧强度中的物种组成也存在较大差异。各放牧强度中距离置信区间最远的样点主要来自7个放牧点中的2-3个放牧点。不同层次中无牧区的物种组成均较放牧区相似, 其中以灌木层的相似程度最高。随着放牧强度增加, 灌木层中物种相似程度增加, 以重牧区最为集中, 而草本层中重牧区的物种组成差异大于中牧区和轻牧区。
图2
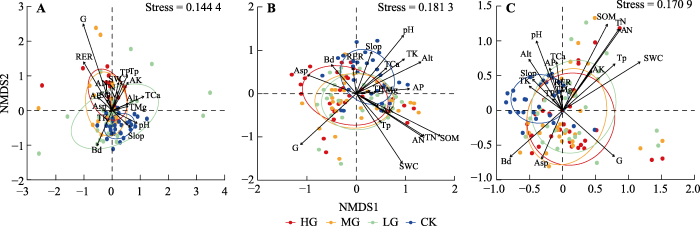
图2
中国亚热带喀斯特山地灌草丛样方与环境因子的非度量多维尺度排序(NMDS)。A, 灌木层。B, 草本层。C, 群落整体。CK, 无放牧; HG, 重度放牧; LG, 轻度放牧; MG, 中度放牧。AK, 速效钾含量; Alt, 海拔; AN, 碱解氮含量; AP, 速效磷含量; Asp, 坡向; Bd, 土壤密度; G, 放牧强度; RER, 岩石裸露率; Slop, 坡度; SOM, 土壤有机质含量; Stress, 协强系数; SWC, 土壤含水量; TCa, 全钙含量; TK, 全钾含量; TMg, 全镁含量; TN, 全氮含量; TP, 全磷含量; Tp, 土壤总孔隙度。图中椭圆为60%置信区间。
Fig. 2
Non-metric multi-dimensional scaling (NMDS) ordination of shrub tussock plots and environmental factors in subtropical karst mountains of China. A, Shrub layer. B, Herb layer. C, Total community. CK, none grazing; HG, heavy grazing; LG, light grazing; MG, moderate grazing. AK, available potassium content; Alt, altitude; AN, alkali-hydrolyzable nitrogen content; AP, available phosphorus content; Asp, slope aspect; Bd, soil density; G, grazing intensity; RER, rock exposure rate; Slop, slope; SOM, soil organic matter content; SWC, soil water content; TCa, total calcium content; TK, total potassium content; TMg, total magnesium content; TN, total nitrogen content; TP, total phosphorus content; Tp, total soil porosity. The ellipse in the figure is the 60% confidence interval.
2.2 环境因子对灌草丛群落的影响
对于灌木层(图2A), 放牧强度和岩石裸露率与NMDS排序轴有显著相关关系(表1), 表明这2个因子对灌木层物种组成有着较强的解释能力。其余环境因子的线段较短, 且与排序轴无显著相关关系, 表明它们的解释率较低。对于草本层(图2B)和群落整体(图2C), 放牧强度、SOM含量、AN含量、TN含量、SWC和pH均与NMDS排序轴具有显著相关关系(表1), 表明它们对草本层乃至群落整体的物种组成有着较强的解释能力。此外, 草本层和群落整体还分别受到海拔和TK含量、Bd和Tp的显著影响。由群落整体来看, 随着放牧强度的增加, 海拔、土壤pH、TK含量逐渐减小, 无牧区的植物群落主要分布在海拔较高、坡度较陡、土壤TK含量、TCa含量、AP含量和pH较高的区域, 海拔和土壤pH是无牧区群落分布主要制约因素。放牧区物种大多分布在放牧强度主导、坡向较大、Bd较大、土壤养分含量较低的区域。
表1 中国亚热带喀斯特山地灌草丛非度量多维尺度排序(NMDS)排序轴与环境因子的相关性系数
Table 1
| 因子 Factor | 灌木层 Shrub layer | 草本层 Herb layer | 群落整体 Total community | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NMDS1 | NMDS2 | R2 | p | NMDS1 | NMDS2 | R2 | p | NMDS1 | NMDS2 | R2 | p | |
| G | -0.387 | 0.922 | 0.330 | 0.001 | -0.692 | -0.722 | 0.151 | 0.001 | 0.764 | -0.646 | 0.156 | 0.001 |
| RER | -0.587 | 0.810 | 0.146 | 0.001 | -0.111 | 0.994 | 0.021 | 0.303 | -0.059 | 0.998 | 0.013 | 0.492 |
| Alt | 0.948 | 0.319 | 0.018 | 0.349 | 0.874 | 0.486 | 0.129 | 0.001 | -0.572 | 0.821 | 0.118 | 0.002 |
| Slop | 0.797 | -0.604 | 0.032 | 0.177 | 0.310 | 0.951 | 0.059 | 0.049 | -0.713 | 0.701 | 0.061 | 0.037 |
| Asp | -1.000 | -0.003 | 0.004 | 0.795 | -0.916 | 0.401 | 0.066 | 0.028 | -0.414 | -0.910 | 0.087 | 0.008 |
| SOM | -0.521 | 0.853 | 0.006 | 0.695 | 0.868 | -0.496 | 0.220 | 0.001 | 0.481 | 0.877 | 0.295 | 0.001 |
| TN | -0.329 | 0.944 | 0.015 | 0.409 | 0.816 | -0.578 | 0.163 | 0.001 | 0.590 | 0.808 | 0.309 | 0.001 |
| TP | 0.386 | 0.923 | 0.038 | 0.104 | 0.976 | 0.218 | 0.005 | 0.732 | -0.681 | 0.732 | 0.008 | 0.670 |
| TK | -0.939 | -0.345 | 0.001 | 0.974 | 0.774 | 0.634 | 0.091 | 0.006 | -0.824 | 0.566 | 0.056 | 0.042 |
| AN | -0.384 | 0.923 | 0.018 | 0.367 | 0.804 | -0.594 | 0.160 | 0.001 | 0.601 | 0.799 | 0.315 | 0.001 |
| AP | -0.794 | 0.608 | 0.019 | 0.340 | 0.995 | 0.101 | 0.068 | 0.023 | -0.308 | 0.951 | 0.060 | 0.029 |
| AK | 0.574 | 0.819 | 0.049 | 0.055 | 0.834 | -0.551 | 0.017 | 0.377 | 0.615 | 0.789 | 0.077 | 0.010 |
| pH | 0.924 | -0.383 | 0.045 | 0.073 | 0.581 | 0.814 | 0.156 | 0.001 | -0.371 | 0.929 | 0.170 | 0.001 |
| TCa | 0.941 | 0.337 | 0.070 | 0.020 | 0.722 | 0.692 | 0.042 | 0.076 | -0.132 | 0.991 | 0.073 | 0.009 |
| TMg | 0.978 | 0.208 | 0.016 | 0.352 | 0.996 | 0.094 | 0.010 | 0.583 | -0.125 | 0.992 | 0.007 | 0.681 |
| Bd | -0.469 | -0.883 | 0.065 | 0.031 | -0.602 | 0.798 | 0.040 | 0.085 | -0.765 | -0.644 | 0.160 | 0.001 |
| Tp | 0.471 | 0.882 | 0.065 | 0.025 | 0.606 | -0.795 | 0.040 | 0.090 | 0.766 | 0.643 | 0.159 | 0.001 |
| SWC | 0.269 | 0.963 | 0.034 | 0.171 | 0.504 | -0.864 | 0.196 | 0.001 | 0.860 | 0.510 | 0.276 | 0.001 |
p, 环境因子与NMDS排序轴相关系数的显著度, 加粗数字表示p值经过“bonferroni”校正后仍保有显著性; R2, 多元回归的决定系数。其他因子含义见
p, significant correlation between the environmental factors and NMDS axis, and bold figure indicates that the p value remains significant after “bonferroni” correction; R2, the regression determination coefficient of multiple regression. Other factors see
2.3 放牧对灌草丛群落结构的影响
随放牧强度的增加, 群落高度、盖度和地上生物量均呈明显降低趋势(图3)。从群落高度看, 灌木层中除轻牧区和无牧区间无显著差异外, 其余牧区间均呈现显著差异; 草本层中仅重牧区分别同轻牧区和无牧区、中牧区同无牧区存在显著差异。从盖度变化看, 除群落整体和灌木层中无牧和轻牧区群落盖度显著大于中牧和重牧区, 草本层中轻牧区群落盖度显著大于重牧区外, 其余不同层次中的各牧区间无显著差异。从生物量变化来看, 无牧区的地上生物量显著高于重牧和中牧区, 放牧区之间以及轻牧和无牧区间的差异则并不显著, 地上生物量整体随放牧强度降低呈单调上升趋势。
图3
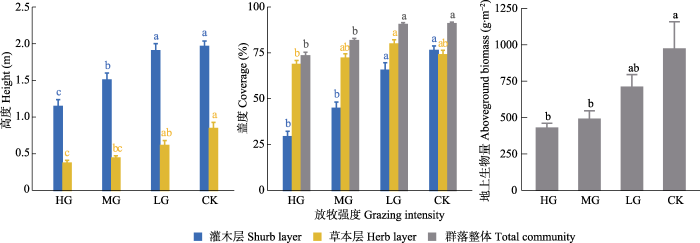
图3
中国亚热带喀斯特山地灌草丛不同放牧强度下的群落高度、盖度和地上生物量变化(平均值±标准误)。CK, 无放牧; HG, 重度放牧; LG, 轻度放牧; MG, 中度放牧。不同小写字母表明不同放牧强度间差异显著(p < 0.05)。
Fig. 3
Changes of community height, coverage and aboveground biomass of shrub tussock under different grazing intensities in subtropical karst mountains of China (mean ± SE). CK, none grazing; HG, heavy grazing; LG, light grazing; MG, moderate grazing. Different lowercase letters show significant difference between different grazing intensities (p < 0.05).
3 讨论
3.1 放牧对喀斯特山地灌草丛物种组成的影响
就科、属、种数量(图1)而言, 随着放牧强度增加, 灌木层明显锐减, 而草本层和群落整体均呈单峰变化趋势, 峰值出现在轻牧区。上述差异的形成很可能是由山羊的选择性采食所决定的。有研究发现, 山羊食性广, 尤为喜食灌木鲜嫩枝叶, 灌木植物采食量一般占全部采食量的85%左右(林嘉等, 1994)。李洪曙等(2002)在贵州喀斯特灌草丛进行为期3年的放牧研究同样表明山羊对灌木择食性更大。结合NMDS分析结果(图2A), 表明放牧强度对灌木层的影响最显著, 验证山羊偏好采食灌木植物。山羊“食谱”中草本植物占比较少, 灌草采食比例虽受资源的可获得性、空间距离、放牧季节等因素影响而发生改变(万里强等, 2003; 董全民等, 2011), 但仍以灌木层植物为主。在长期高强度放牧条件下, 灌草丛内的资源空间配置发生重组, 灌木层所释放的生态位为草本植物的快速繁殖生长以及种库内植物和入侵种定植提供契机(杨利民等, 1999a; 万里强等, 2012), 从而表现为禾本科以及外来入侵种飞机草和鬼针草重要值随放牧强度增加而增加(附录I, 附录III)。Souther等(2019)在高海拔半干旱草原的研究同样发现, 高强度放牧使群落物种组成更大比例地向一年生物种和外来物种转移。多数放牧研究结果均发现适度放牧有助于物种丰富度和初级生产力的提高, 支持中度干扰假说(董全民等, 2011; 李文怀等, 2014)。本研究中轻牧区的物种丰富度较其他牧区高, 且主要由草本层贡献, 表明轻度放牧为喀斯特山地灌草丛的“中度干扰水平”。李洪曙等(2002)和Škornik等(2010)分别在喀斯特灌丛草地和牧场开展的放牧实验也表明, 轻牧能较好地维持植被的多样性, 促进植被良性发展。
针对我国北方草地的放牧研究表明, 群落优势种(优势类群)会随放牧强度的变化而发生更迭(杨浩等, 2009), 但也有研究认为, 放牧干扰主要影响物种的空间分布和群落的高度、盖度、丰度等, 而非群落优势种(优势类群)组成(赵生龙等, 2020; Michaels et al., 2021)。本研究中主要优势物种乃至次优势种(如黄荆、矛叶荩草)在各放牧强度中均有分布, 其差别仅在于重要值的不同(附录III), 即优势度不同, 由此导致不同放牧强度间的物种组成趋于相似。一方面, 放牧本身有着“环境滤筛”作用(Abbas et al., 2019), 山羊的选择性采食是各牧区物种组成差异的关键。另一方面, 各牧区物种组成存在趋于相似情况或主要源于放牧压力未达到群落发生“质变”的阈值。首先, 山羊主要采食灌木鲜嫩枝叶, 灌木植物能在较短时间内抽叶生长(万里强等, 2003), 而建群种黄荆作为先锋种, 其抗干扰能力强, 本研究中即便是重度放牧也不足以改变其优势地位。其次, 在实地考察中发现, 多数牧民具备生态保护意识, 会因地制宜维持一定的养殖数量, 并实行多地点轮牧, 灌木可获得更多恢复生长时间。此外, 区域种库同样影响物种组成。区域种库由地史过程(如构造运动、冰期、地质年代)和区域过程(物种的形成、迁移扩散以及消亡)决定(方精云等, 2009), 喀斯特山地生境破碎, 高生境异质性决定其区域种库相对丰富且复杂, 本研究中放牧干扰下的实际种库(即调查群落中的物种)虽有变化, 但仍在研究区所在区域种库范围内(张大勇和姜新华, 1997; Herrero-Jáuregui & Oesterheld, 2018)。当然, 放牧干扰不足以在短期内引起灌草丛发生质变, 即群落优势种被其他物种所取代, 群落整体物种组成趋于简单, 饲用植物品质发生劣化等(刘洪来等, 2011), 这可能是放牧强度、植物性状、牲畜活动偏好等诸多因素耦合作用的结果(毛志宏和朱教君, 2006; 王岭, 2010; Michaels et al., 2021), 未来仍需进一步研究。
本研究表明, 放牧促进了喀斯特山地灌木层植物组成的同质化。灌木层放牧区排序点较无牧区分散, 但其相似程度随放牧强度增加而增加(图2A), 可见放牧的滤筛作用使灌木层物种组成随放牧强度增加趋于简单, 支持放牧导致主要采食植物类群发生植被同质化的观点(Kouba et al., 2021)。放牧区排序点较无牧区分散, 主要是因为放牧作为一种连续的外界干扰过程, 该过程一方面抑制优势种(优势类群)的竞争, 另一方面也会使群落断层形成的频率加快(杨利民等, 1999a), 加之山羊活动会加快种子传播速率并扩大传播范围, 长期放牧作用下使群落组成差异增加, 然而上述两个方面的作用实际上受放牧强度的调控。无牧区的灌木层生长良好, 物种丰富, 样方间的共有种数量也因此增加, 由此导致其物种组成更为相似。草本层中同一放牧强度下物种组成差异更大, 这与多数草本植物自身体型更小, 传播、繁殖和生长能力更强, 物质能量贮存更低, 适应性更强(杨利民等, 1999b)的特性密切相关。草本植物的上述特性一方面决定其所需生态位较小, 这在一定程度上促进了草本层生态位分化, 另一方面保证草本植物能在群落断层出现时更快占据合适的资源空间, 但这同时也意味着群落内部种内、种间竞争更激烈(Török et al., 2018; 张芳等, 2020), 而山羊排泄物可通过促进土壤养分的再分配加剧竞争(Škornik et al., 2010), 这些无疑会进一步增大草本层物种组成差异, 且差异随放牧强度增加而加大。此外, 喀斯特生境自身立地环境差异以及由放牧行为所造成的高度生境异质性同样可能导致植被组成差异增加(王岭, 2010; Geekiyanage et al., 2019)。Niu等(2016)发现放牧会消耗土壤养分, 长期高强度放牧会加剧土壤贫瘠。图2中相对离散的样点主要集中在2-3个放牧点, 群落整体(图2C)中牧区的样点相较于无牧区多分布在环境较干热、土壤密度较高且相对贫瘠的区域, 表明生境差异可能是造成物种组成差异的原因之一。
3.2 放牧对喀斯特山地灌草丛群落结构的影响
有研究表明, 植被结构对放牧强度的反应更明显(Riesch et al., 2020)。同周华坤等(2004)和赵生龙等(2020)的研究结果类似, 本研究中的植被高度受山羊采食活动的直接影响, 随放牧强度增加呈矮化趋势, 然而由于山羊采食偏好影响, 灌木层和草本层的高度变化速率并不一致, 前者主要受采食活动影响, 后者则主要受践踏活动影响。灌木层盖度同高度变化趋势一致, 但草本层盖度变幅并不剧烈。Michaels等(2021)发现与放牧相关的物种丰富度增加与主要优势种盖度的减少有关。在重牧和中牧区, 灌木层植被冠层稀疏, 地表草本植被所获光热、空间、土壤养分增加(Celaya et al., 2010), 生态位得以释放, 被山羊厌食且耐践踏的草本植物更易定植生长, 这也是禾本科在重牧区重要值占比最大以及重牧区草本层物种组成差异大的主要原因, 与Bakhshi等(2020)发现禾本科植物盖度会随放牧强度增加而增加类似。同时, 山羊频繁的采食、践踏活动可能促使部分草本植物超补偿生长(侯向阳, 2016), 且粪便富集区更易形成草本植物密集生长斑块(Milchunas & Lauenroth, 1993; Škornik et al., 2010), 因而在重牧和中牧条件下的草本植物种类和群落盖度能维持较高水平。在轻牧和无牧区, 草本层受干扰较小甚至无干扰, 生长状况保持良好, 灌木层因被采食而出现缺口, 草本层植物所获光热资源增加(Celaya et al., 2010), 光竞争也愈发激烈(Škornik et al., 2010), 草本植物可能会采取增加叶面积等一系列权衡措施来提高光捕获能力(崔海军等, 2015; 李西良等, 2015), 同时山羊采食活动也可能将更适应该区域环境的植物种子带入并定植, 从而表现为轻牧区草本层植物的盖度较无牧区高。山羊的适度活动是维持群落资源竞争状态的主导因子。关于植被应对山羊放牧干扰究竟采取哪些权衡措施予以响应, 未来尚需进一步研究。
地上生物量是地表植被生产力的重要量化指标(袁文平等, 2014), 是群落组成、群落高度和盖度等群落特征共同作用的结果, 也是放牧干扰下植被载畜能力和生产潜力的判断依据(赵生龙等, 2020)。本研究中, 在山羊采食活动影响下, 地上生物量随群落物种数、群落高度和盖度的减小而逐渐减小, 与Škornik等(2010)和赵彬彬等(2009)在喀斯特牧场和高寒草甸的研究结果相似。但赵彬彬等(2009)也发现毒草和杂类草的平均个体生物量并未发生变化, 部分甚至在放牧区增加, 本研究发现不在山羊“食谱”核心的草本植物盖度与重要值比重在重牧和中牧条件下能维持较高水平与之类似。植物光合作用面积随着叶片被大量采食、破坏而减小, 导致植物物质能量积累减少, 从而影响植物的正常生长发育和繁殖, 随着高产灌木的减少, 最终表现为植被生产力的下降(孙海群等, 1999; 赵生龙等, 2020)。如在重牧区, 群落分层现象逐渐弱化, 群落矮小化明显, 草本层植物重要值大于灌木层, 地上生物量的主要贡献者也逐渐由木本植物转向草本植物, 由此导致该区域地上生物量仅为422.32 g·m-2, 不到无牧区(969.79 g·m-2)的一半, 说明过高的放牧强度会导致喀斯特灌草丛生产潜力下降, 进而影响其载畜能力, 长期作用下可能增加植被退化风险。
3.3 放牧是否会阻碍喀斯特山地植被的自然恢复?
植被的同质化是生物贫化的一种重要形式, 在一定程度上降低了生态系统对长期大规模干扰的植被恢复力(Salgado-Luarte et al., 2019)。本研究的灌木层植被趋于同质化, 表明长期山羊放牧会降低喀斯特灌草丛的恢复力, 而入侵种的增加同样加剧了同质化趋势。物种多样性阻抗假说(Elton, 1958)认为群落的入侵抵抗性与群落物种丰富度成正比。本研究中, 一方面, 山羊放牧导致灌草丛物种丰富度下降, 群落的入侵抵抗性随之下降, 放牧形成的空余生态位为外来入侵种的入侵、定植与扩散提供机会, 从而表现为外来入侵种飞机草和鬼针草的重要值随放牧强度增加而增加。另一方面, 外来入侵种会迅速抢占空间资源, 如多年生的飞机草的繁殖力和环境适应性极强, 还可通过化感作用抑制本地植物的生长(刘金海等, 2006), 这些入侵种甚至可以通过土壤遗留效应影响本地种的生长, 进而加剧物种多样性下降, 植被趋于同质化。可见, 放牧以及入侵种的增加致使灌草丛物种组成简单化, 优质饲用植物类群减产, 生态系统结构与功能发生退化。
黄荆是喀斯特退化山地的先锋种, 也是本研究中重要值占比最大的优势种, 其重要值随着放牧强度的增加而增加(附录III), 说明其耐牧性与适应性强, 同时也表明山羊放牧会导致喀斯特灌草丛的退化加剧, 加大石漠化风险(曾馥平等, 2007; 张芳等, 2020)。菊科同黄荆重要值变化趋势一致, 或因菊科本身具刺激性气味、口感不佳等生理特性(万里强等, 2000), 致使其受采食较少。因此, 黄荆与部分菊科物种(如入侵种飞机草和鬼针草)的分布及其多度或可作为喀斯特地区放牧干扰强弱与生态系统退化的判断依据。总的来看, 在山羊放牧干扰下, 喀斯特灌草丛物种丰富度和地上生物量明显下降, 岩石裸露率增加, 植被物种组成和群落结构趋向简单, 出现结构稀疏化、矮小化趋势, 进而导致植被生产
力下降, 表明放牧可促使灌草丛逆向演替, 植被发生退化, 长此以往势必会阻碍喀斯特山地植被的自然恢复, 增加石漠化的风险。放牧对生态系统的负面影响多源自于放牧的长期无序与不当管理(García et al., 2012), 因而, 对于生态系统脆弱的喀斯特山地, 应倡导牧民控制牲畜数量, 坚持轮牧, 出现植被退化倾向的区域应鼓励圈养, 必要时可实行放养与圈养交替进行。
附录I 中国亚热带喀斯特山地灌草丛灌木层、草本层和群落整体中排名前10科的重要值
Supplement I Important values of top 10 families in shrub layer, herb layer and entire community of shrub tussock in subtropical karst mountains of China
附录II 中国亚热带喀斯特山地灌草丛灌木层、草本层和群落整体中排名前10属的重要值
Supplement II Important values of top 10 genera in shrub layer, herb layer and entire community of shrub tussock in subtropical karst mountains of China
附录III 中国亚热带喀斯特山地灌草丛灌木层、草本层和群落整体中排名前10种的重要值
Supplement III Important values of top 10 species in shrub layer, herb layer and entire community of shrub tussock in subtropical karst mountains of China
致谢
感谢广西师范大学谢强副教授在植物种类鉴定工作中给予的帮助。
参考文献
The accumulation of species and recovery of species composition along a 70 year succession in a tropical secondary forest
DOI:10.1016/j.ecolind. 2019.105524 [本文引用: 1]
Study on the effects of different levels of grazing and exclosure on vegetation and soil properties in semi-arid rangelands of Iran
DOI:10.1016/j.chnaes.2019.07.003 URL [本文引用: 1]
Changes in heathland vegetation under goat grazing: effects of breed and stocking rate
DOI:10.1111/j.1654-109X.2009.01054.x URL [本文引用: 2]
Variation in community characteristics and its influencing factors of alpine grasslands along an elevation gradient
高山草地植物群落数量特征沿海拔梯度变化及其影响因素
Dynamic changes and cause analysis of rocky desertification in karst areas of Mashan County in Guangxi
广西马山县岩溶土地石漠化动态变化情况及变化原因分析
Application of NMDS to analysis of phytoplankton community: a case study of Qinshui River
非度量多维标度在亲水河浮游植物群落分析中的应用
Plant trait responses to grazing—A global synthesis
DOI:10.1111/j.1365-2486.2006.01288.x URL [本文引用: 2]
Effects of yak grazing intensity on quantitative characteristics of plant community in a two- seasonal rotation pasture in Kobresia parva meadow
牦牛放牧强度对小嵩草草甸两季轮牧草场植物群落数量特征的影响
Local and regional processes control species richness of plant communities: the species pool hypothesis
DOI:10.3724/SP.J.1003.2009.09141 URL [本文引用: 1]
局域和区域过程共同控制着群落的物种多样性: 种库假说
DOI:10.3724/SP.J.1003.2009.09141
[本文引用: 1]

解释群落的物种多样性大小是生态学研究的一个重要的理论和实践问题。人们提出了群落物种多样性的多种假说, Zobel等人提出的种库假说(species pool hypothesis)是生物多样性理论研究的重要发展。该假说认为, 一个群落的物种多样性不仅与环境条件和生态过程(ecological process)(如竞争、捕食)有关, 也受区域种库(regional species pool)的限制。区域种库是指一个地区可进入某一群落的潜在物种数量, 它由地史过程(如冰期、地质年代)和区域过程(物种形成、迁移扩散以及消亡)所决定。按照种库假说, 某一生境类型的面积越大, 地质年代越古老, 物种形成的机会也就越多, 因而能适应和分布于该生境的物种也就越多, 实际群落中的物种丰富度也就越高。种库在空间上主要有两个层次: 区域种库和实际种库, 前者指某一生境所拥有的潜在物种数量, 主要由生物地理过程(biogeographic processes)所决定; 后者则为调查的群落中实际出现的物种数量, 主要由竞争等生态过程和区域种库共同决定。本文对种库假说的基本概念、主要内容、种库确定方法等作了介绍, 并阐述了作者对这些问题的理解和认识。
A global meta-analysis of grazing effects on plant richness
DOI:10.1016/j.agee.2020.107072 [本文引用: 1]
Goat grazing, its interactions with other herbivores and biodiversity conservation issues
DOI:10.1016/j.smallrumres.2012.03.021 URL [本文引用: 3]
Plant ecology of tropical and subtropical karst ecosystems
DOI:10.1111/btp.12696
[本文引用: 3]
Substantial areas of tropical forests, including those within nine tropical biodiversity hotspots, contain karst landscapes that have developed on soluble carbonate rocks. Here, we review how the ecology of karst forest trees is influenced by hydrological, edaphic, and topographic factors that exhibit fine spatial heterogeneity. Comparative analysis of drought tolerance traits including wood density contributes to the assessment of whether karst tree species are more drought-tolerant compared to non-karst trees. Although karst ecosystems are generally considered to have low phosphorus availability, foliar nitrogen-to-phosphorus ratios exhibit wide variation across karst regions without a clear difference from non-karst ecosystems. According to the analyses of leaf phenology, stem water storage, and isotopic signatures from xylem sap, water use strategies of karst trees can be classified into five types: (a) soil water dependent, (b) epikarst water dependent (mainly use water stored in fine pores and gaps within the epikarst rock during the dry season), (c) groundwater dependent, (d) fog water dependent, and (e) drought-deciduous (shed leaves during the dry season). Overall, published data suggest that only a subset of karst tree species are exclusively distributed within karst hilltops where water availability is limited. The diverse resource acquisition and utilization strategies of karst plants across edaphic habitats must be considered when developing effective strategies to conserve and restore biodiversity in karst landscapes, which are under increasing anthropogenic pressure.
Ecological adaptation of plants and control of rocky-desertification on karst region of Southwest China
DOI:10.3724/SP.J.1258.2011.00991
[本文引用: 1]
Karst region of Southwest China is ecologically very fragile. It has been suffering from severe rock-desertification and its vegetation has been damaging heavily. The restoration or reconstruction of the vegetation is extremely difficult. In recent years, a lot of pure and application-oriented basic researches have been performed in order to scientifically and technologically support the management of the rock-desertification. In this paper, we summarize habitat characteristics of the karst region and review the progress in ecological researches on plant adaptation, plant population, plant community and ecosystem in the region. In addition, as for current situation in management of the rock-desertification, we propose, particularly from angle of plant ecology, expectations for further researches in the region.
我国西南喀斯特植物生态适应性与石漠化治理
DOI:10.3724/SP.J.1258.2011.00991
[本文引用: 1]
我国西南喀斯特地区生态脆弱, 石漠化问题严重, 植被恢复/重建的难度极大。为此, 近年来开展了许多基础性研究, 以求为石漠化治理提供科技支撑。该文概略介绍了该地区喀斯特生境的特点, 回顾和评述了喀斯特生境中植物适应性、植物种群、植物群落和生态系统生态学方面取得的主要研究进展, 并结合石漠化综合治理的现状, 提出了喀斯特植物生态学研究的几点期望。
Effects of grazing intensity on plant richness and diversity, a meta-analysis
DOI:10.1111/oik.04893 URL [本文引用: 2]
Advances and prospects of grassland plant basic biology research
草原植物基础生物学研究进展与展望
Succession of shrubby grassland of rocky mountain and its improvement and utilization in karst areas
岩溶地区石山灌草丛草地演替及其培育和利用
Study of fragile ecosystem reconstruction technology in the karst peak-cluster mountain
岩溶峰丛山地脆弱生态系统重建技术研究
Plant community composition and structure under short-term grazing exclusion in steppic arid rangelands
DOI:10.1016/j.ecolind. 2020.106910 [本文引用: 1]
Nonmetric multidimensional scaling: a numerical method
DOI:10.1007/BF02289694 URL [本文引用: 1]
Rational utilization of shrubby grassland in karst
喀斯特灌丛草场的合理放牧利用
Effects of grazing intensity and topography on species abundance distribution in a typical steppe of Inner Mongolia
DOI:10.3724/SP.J.1258.2014.00016 URL [本文引用: 1]
放牧强度和地形对内蒙古典型草原物种多度分布的影响
DOI:10.3724/SP.J.1258.2014.00016
[本文引用: 1]
为了更好地理解放牧对草原生态系统物种多度分布格局的影响, 以及常见种和稀有种对维持群落多样性的作用, 以内蒙古典型草原为研究对象, 基于长期放牧控制实验平台(包括7个载畜率水平(0、1.5、3.0、4.5、6.0、7.5、9.0 sheep·hm<sup>-2</sup>)和两种地形系统(平地和坡地)), 研究了群落内全部物种、常见种和稀有种的丰富度和多度对放牧强度的响应规律, 并选取对数正态模型、对数级数模型和幂分割模型, 对物种多度数据进行拟合。结果表明: 1)平地系统中, 物种丰富度和多度在低放牧强度下(1.5、3.0 sheep·hm<sup>-2</sup>)增加, 而在中、高度放牧强度下(4.5-9.0 sheep·hm<sup>-2</sup>)降低, 全部物种的多度分布在大多数放牧强度下符合幂分割模型, 在高放牧强度下也符合对数正态模型; 坡地系统中, 物种丰富度和多度随着放牧强度增加而显著降低, 全部物种的多度分布在各个放牧强度下, 均符合幂分割模型和对数正态模型。2)随着放牧强度增加, 常见种的多度响应趋势与全部物种的响应趋势一致, 其多度分布均符合幂分割模型和对数正态模型; 稀有种的丰富度响应趋势与全部物种的响应趋势一致, 其多度分布符合幂分割模型, 同时也部分符合对数正态和对数级数模型。总之, 适宜的载畜率有利于生物多样性和初级生产力的提高, 平地系统中物种多度的响应在一定程度上支持放牧优化假说; 而坡地系统中不同物种多度的响应差异说明: 确定最佳载畜率时, 还需要考虑地形因素的影响。此外, 模型的拟合结果表明: 生态位分化机制对内蒙古典型草原物种多度分布起着主要作用, 常见种和稀有种通过不同的响应方式共同维持着草原生态系统的物种多样性。
Plastic responses of stem and leaf functional traits in Leymus chinensis to long-term grazing in a meadow steppe
DOI:10.3724/SP.J.1258.2014.00040 URL [本文引用: 1]
草甸草原羊草茎叶功能性状对长期过度放牧的可塑性响应
DOI:10.3724/SP.J.1258.2014.00040
[本文引用: 1]
植物对不同功能性状进行权衡, 通过表型可塑性达到对异质生境的适应是植物的一种生态对策。羊草(Leymus chinensis)是欧亚温带草原东缘的主要优势植物, 研究其对放牧的表型反应对揭示草原生态系统的放牧响应机制具有代表意义。该文以内蒙古呼伦贝尔草甸草原为例, 通过设置不同放牧压力与围封的长期试验, 研究了羊草茎叶功能性状对放牧的可塑性响应模式。结果表明: 1)与长期围封相比, 长期放牧导致羊草茎叶性状显著小型化, 其中, 株高和个体地上生物量分别降低76.82%和89.88%, 但3年短期围封对茎性状影响不显著, 说明羊草表型矮小化现象具有一定的保守性; 2)通过排序构建羊草性状可塑性变化谱, 发现茎质量、总质量、茎高、株高、叶面积等为对放牧响应的敏感性状, 而叶片数、茎粗、叶宽等较为稳定, 为惰性性状; 3)放牧干扰下, 羊草性状可塑性程度与其变异性之间符合y = y<sub>0</sub> + ae<sup>bx</sup>拟合关系, 随着植物性状的响应强度增大, 其变异性增强; 4)偏最小二乘法分析发现茎长、株高、叶面积、叶长等性状的投影重要性指标大于1, 对地上生物量变化的解释率为68.6%, 是导致长期放牧下羊草个体生物量降低的主要因子。研究认为, 矮化型变是羊草的避牧适应对策, 在亚稳态下, 通过不同性状的权衡, 充分利用环境资源完成其生活史。
Plant functional traits and their trade-offs in response to grazing: a review
DOI:10.3724/SP.J.1259.2015.00159 URL [本文引用: 2]
放牧对草原植物功能性状及其权衡关系的调控
DOI:10.3724/SP.J.1259.2015.00159
[本文引用: 2]
放牧是草原生态系统利用与管理的主要方式; 而过度放牧导致的草原退化和生产力衰减问题已成为当前生产及理论上的一个棘手难题。近年来, 植物功能性状及其权衡等新理论与方法的引入, 为解析放牧对草原的影响机制提供了新工具, 成为草原生态学的一个新兴领域, 但目前尚缺乏系统的回顾总结。该文综述了近年来国内外关于草原植物功能性状对放牧的响应及其权衡关系的研究进展, 综合分析了植物性状权衡变化的主要特征、成因与机制, 针对学界关于放牧下植物矮化型变机制的“避牧适应假说”与“生长受阻假说”的争论, 提出了下一步需重点关注的科学问题。
Study on daily food consumption of grazing goats in Zhejiang Province
浙江山羊放牧日食量测定的研究
Biomass of canopy and shrub layers of karst forests in Pudding, Guizhou, China
贵州普定喀斯特次生林乔灌层地上生物量
DOI:10.3773/j.issn.1005-264x.2009.04.008
[本文引用: 1]
生物量是植物群落最重要的特征之一, 也是研究生态系统基本过程和功能的重要参数。我国西南喀斯特山地次生林是退化生态系统恢复过程中的重要阶段, 其生境特殊, 乔灌层种类繁杂且生长形态多样, 物种间体积质量密度差异悬殊, 生物量测定极为困难。所以, 有关其生物量的基础资料极为缺乏。该文根据对普定喀斯特生态站的窄叶石栎(Lithocarpus confinis)+云南鼠刺(Itea yunnanensis)林、圆果化香(Platycarya longipes)+云南鼠刺林、圆果化香+槲栎(Quercus aliena)林3个主要次生森林类型的样地调查资料, 采用样木回归模型法和收获法, 研究了群落乔灌层地上生物量及其分配格局。利用172株样木数据建立了圆果化香、窄叶石栎、云南鼠刺、刺楸(Kalopanax sep-temlobus)、安顺润楠(Machilus cavaleriei)、槲栎、香叶树(Lindera communis)、川钓樟(Lindera pulcherrima var. hemsleyana)、异叶鼠李(Rhamnus heterophylla)、倒卵叶旌节花(Stachyurus obovatus)、薄叶鼠李(Rhamnus leptophylla)、贵州花椒(Zanthoxylum esquirolii)、竹叶椒(Zanthoxylum planispinum)、铁仔(Myrsine africana)和刺异叶花椒(Zanthoxylum dimorphophyllum var. spinifolium)15个主要树种的分种生物量回归模型。同时, 利用这些样木建立了3个不同胸径(D)组(D5 cm)的生物量回归模型。通过进一步分种和分组的计算得出: 这3个次生乔木林的乔灌层地上生物量分别为85.6×103、65.3×103和115.2×103 kg·hm–2; 层次分配上, 3个样地的乔木层地上生物量占绝对优势, 分别约占乔灌层地上生物量的98.5%、96.6%和99.0%; 径级分配上, 3个样地的生物量主要集中在大径级(D≥10 cm)的个体上; 物种分配上, 3个样地的生物量排序前10位的物种分别约占乔灌层地上生物量的99.3%、97.3%和99.0%, 并集中分配在少量优势树种中。
Research progress of grassland degraded succession and diagnosis
草地退化演替过程及诊断研究进展
DOI:10.11733/j.issn.1007-0435.2011.05.026
[本文引用: 1]
随着人口的不断增加和现代工农业文明的冲击,草地生态系统出现不同程度的退化,导致草地生态系统组成、结构、过程和功能等方面发生量和质的变化;对草地退化的系统理解是进行草地恢复与重建的基础与前提。本文综述了草地退化的内涵、驱动力、退化过程与特点,以及草地生态系统退化程度诊断等一系列问题,绘制了描述草地退化程度的概念模型;建议从生态学角度研究草地退化程度诊断时选择相应的气候顶级系统或受干扰程度较轻的"自然生态系统";从草地经营学角度研究草地退化程度诊断时选择适度放牧草地作为参照系统。归纳了退化草地生态系统诊断的生物途径、生境途径和功能服务途径;分析了草地退化程度诊断的方法与可选择的指标体系,并对草地退化程度诊断及草地恢复中应注意的事项进行了讨论。
The damage and control measures of Eupatorium odoratum
飞机草的危害及防治措施简介
Species composition and community structure at different vegetation successional stages in Puding, Guizhou Province, China
贵州省普定县不同植被演替阶段的物种组成与群落结构特征
DOI:10.3724/SP.J.1258.2011.01009
[本文引用: 2]
通过对贵州省普定县喀斯特地区不同植被演替阶段群落的调查, 研究了植被演替过程中群落物种组成和群落结构的变化。结果表明, 该地区的植被主要处于5个演替阶段, 即次生乔木林、乔灌过渡林、藤刺灌丛、稀灌草丛以及火烧干扰后的蕨类植物群落。本次调查共记录到植物365种, 隶属89科218属。其中, 蕨类植物31种, 隶属14科23属; 种子植物334种, 隶属75科195属。物种分布较多的科主要有蔷薇科、菊科、禾本科、百合科、忍冬科、唇形科、莎草科、樟科、葡萄科和水龙骨科。随着正向演替的推进, 物种丰富度增加, 群落结构趋于复杂化。藤刺灌丛与乔灌过渡林群落层次不明显, 次生乔木林分层明显。从藤刺灌丛向次生乔木林演替的过程中, 小径级个体所占比例明显降低, 高于1.3 m植物的总密度、乔木密度和藤本密度都先升高后降低, 而灌木密度呈逐渐降低的趋势。对喀斯特地区植被的恢复提出了参考措施。
Effects of disturbances on species composition and diversity of plant communities
干扰对植物群落物种组成及多样性的影响
Influence of yak grazing on plant niche characteristics in alpine meadow communities at the northeastern edge of the Qinghai- Tibetan Plateau
青藏高原东北边缘高寒草甸植物种群生态位特征对牦牛放牧的响应
Grazing affects vegetation diversity and heterogeneity in California vernal pools
DOI:10.1002/ecy.3295 [本文引用: 6]
Quantitative effects of grazing on vegetation and soils over a global range of environments
DOI:10.2307/2937150 URL [本文引用: 1]
Grazing increases functional richness but not functional divergence in Tibetan alpine meadow plant communities
DOI:10.1007/s10531-015-0960-2 URL [本文引用: 1]
Beyond the obvious impact of domestic livestock grazing on temperate forest vegetation—A global review
DOI:10.1016/j.biocon.2019.07.007 URL [本文引用: 1]
Grazing by wild red deer maintains characteristic vegetation of semi-natural open habitats: evidence from a three-year exclusion experiment
DOI:10.1111/avsc.12505 URL [本文引用: 1]
Goat grazing reduces diversity and leads to functional, taxonomic, and phylogenetic homogenization in an arid shrubland
DOI:10.1002/ldr.3208 URL [本文引用: 1]
Differentiation of vegetation types and biodiversity in the southern edge of Hulunbuir grassland
呼伦贝尔草原南缘植被类型分异及生物多样性特征
The effect of grazing pressure on species richness, composition and productivity in North Adriatic karst pastures
DOI:10.1080/11263501003750250 URL [本文引用: 6]
Occurrence, spatial-temporal dynamics and regulation strategies of karst rocky desertification in southwest China
中国西南喀斯特石漠化时空演变特征、发生机制与调控对策
Community composition and biodiversity characteristics of forests in karst cluster-peak- depression region
DOI:10.3724/SP.J.1003.2010.355 URL [本文引用: 1]
喀斯特峰丛洼地不同类型森林群落的组成与生物多样性特征
DOI:10.3724/SP.J.1003.2010.355
[本文引用: 1]
不同退化程度石漠化生态重建的关键是恢复植被, 提高土地生产力。本文基于动态监测样地(200 m × 40 m)植被的全面调查, 研究了喀斯特峰丛洼地人工林、次生林和原生林3类典型森林群落木本植物的组成与生物多样性特征。结果表明, 3类森林的物种组成分别为26科52属65种、33科68属100种和43科91属123种, 常绿物种分别占41.54%、47.00%和52.85%; 科、属、种和生活型组成复杂, 优势科或种明显, TWINSPAN分类第3级水平上可分别划分为8、9和8个群落类型。原生林多样性和结构性指标均高于人工林和次生林, 人工林的Shannon-Wiener指数、Simpson指数、均匀度、冠幅、胸径和树高均高于次生林, 而种类、密度和盖度则低于次生林。不同类型森林群落特征不同, 应采取相应的经营策略。
Complex response of vegetation to grazing suggests need for coordinated, landscape-level approaches to grazing management
DOI:10.1016/j.gecco.2019.e00770 [本文引用: 1]
Progress on grassland degenerated succession
草地退化演替研究进展
Increased vegetation growth and carbon stock in China karst via ecological engineering
DOI:10.1038/s41893-017-0004-x URL [本文引用: 1]
Vegetation type and grazing intensity jointly shape grazing effects on grassland biodiversity
DOI:10.1002/ece3.4508
PMID:30397469
[本文引用: 1]
In the Palaearctic steppe zone, overgrazing was identified as one of the key drivers of declining grassland biodiversity, which underlines the necessity of the functional evaluation of increased grazing pressure on grassland vegetation. We tested the following hypotheses: (a) The effect of grazing intensity on species and functional diversity is strongly dependent on grassland type. (b) The magnitude of diet selectivity of grazers decreases with increasing grazing intensity. (c) Increasing grazing intensity increases evenness and functional evenness of the subjected grasslands. We analyzed vegetation patterns in four types of grasslands (Dry alkali short-grass steppes, Dry loess steppes, Non-alkali wet and Alkali wet grasslands) along an intensity gradient of beef cattle grazing at 73 sites in Hungary. Species richness, Shannon diversity, evenness, and four leaf traits were analyzed. We calculated community-weighted means for each single trait, and multi-trait functional richness, functional evenness, and divergence for all leaf traits. All species and functional diversity metrics were significantly affected by the grassland type, except leaf dry matter content. The effect of interaction between grazing intensity and grassland type was also significant for functional richness, functional evenness, community-weighted means of leaf area, and for species richness and evenness. An upward trend of specific leaf area was detected in all grasslands with the highest scores for the overgrazed sites, but the change was also grassland type dependent. The detected trend suggests that with increased intensity the overall selectivity of grazing decreased. We found that evenness was affected but functional evenness was not affected by grazing intensity. Functional evenness scores were more related to the grassland type than to changes in grazing intensity, and displayed a high variability. We stress that one-size-fits-all strategies cannot be recommended and actions should be fine-tuned at least at the level of grassland type.
Effects of different stocking rates on vegetation traits of cultivated pastures in southern China
不同放牧强度对南方人工草地植被特征的影响
Study on dietary composition and preference of goats on a shrubland in the Three Gorges Region of Yangtz River
三峡地区灌丛草地放牧山羊日粮组成及其喜食性
DOI:10.11733/j.issn.1007-0435.2000.03.005
[本文引用: 1]
在三峡地区灌丛草地研究放牧山羊的日粮组成及其喜食性。结果表明,在山羊日粮中50%以上由灌木植物组成。山羊对灌木的喜食性指数极显著地高于禾草和杂类草(P<0.01),而对禾草和杂类草喜食性的差异不明显。在灌木中,山羊对马桑和杉木等的喜食程度较高,其中马桑的喜食性指数为63%,而麻栎、水锦树和柳杉仅为15%、13%和12%。对于禾草,山羊明显喜食芒和黄背草(P<0.05),喜食性指数分别为16%和12%。对杂类草中的竹节草喜食性指数为13%。
Advance in studies on grazing and utilization of shrubby grasslands by goats
灌丛草地山羊放牧利用研究进展
Impacts of the rangeland degradation on CO2 flux and the underlying mechanisms in the Three-River Source Region on the Qinghai-Tibetan Plateau
草地退化对三江源区高寒草甸生态系统CO2通量的影响及其原因
Changes of vegetation features of rocky desertification process in karst area of Guizhou
贵州喀斯特区石漠化过程中植被特征的变化
Karst landscapes of China: patterns, ecosystem processes and services
DOI:10.1007/s10980-019-00912-w
[本文引用: 2]
Context The karst region of southwestern China, one of the largest continuous karsts in the world, is known for its unique landscapes and rich biodiversity. This region has suffered severe environmental degradation (e.g., vegetation cover loss, soil erosion and biodiversity loss). In recent decades, Chinese governments at different levels have initiated several ecological programs (e.g., Green for Grain, Mountain Closure) to restore the degraded environment and to alleviate poverty. Objectives This study summarizes landscape studies of karst landscapes patterns, their dynamics and interactions among landscape pattern, hydrological processes and ecosystem services (ES). Methods We conducted a systematic literature review of science and land use policy to identify knowledge gaps and recommend future research and policy directions. Results Karst landscapes have experienced rapid turnover in recent decades due largely to the overlap of intense human activity on the fragile karst ecosystems. Many studies have comprehensively examined hydrology, soil processes and ecosystem services (ES) and their relationships with landscape pattern. Most of these studies have found that karst ecosystems recover with improved ES. However, the importance of epikarst in hydrological and soil processes, intense anthropogenic disturbance and landscape heterogeneity in landscape models remains elusive. Conclusions Future research should focus on in-depth examination and modelling of karst specific hydrological and soil processes, investigating relationships between climatic change, landscape change, ecological processes, and region-specific ES assessments. Results from such research should provide the necessary scientific support for a comprehensive, national karst rocky desertification treatment project (Stage II) and poverty alleviation initiatives.
Karst rocky desertification: formation background, evolution and comprehensive taming
喀斯特石漠化的形成背景、演化与治理
The succession characteristics and its driving mechanism of plant community in karst region, Southwest China
中国西南喀斯特植物群落演替特征及驱动机制
Climate change weakens the positive effect of human activities on karst vegetation productivity restoration in southern China
DOI:10.1016/j.ecolind. 2020.106392 [本文引用: 2]
Spatial heterogeneity and its causes in evergreen broad-leaved forests in the Ailao Mountains, Yunnan Province
DOI:10.17520/biods.2016199
[本文引用: 1]
Understanding the variation of species richness over spatial scales, and elucidating the response of plant community composition to habitat heterogeneity has been one of the major topics in the study of community ecology. Niche differentiation related to habitat heterogeneity plays an important role in shaping diversity levels and species distribution patterns of plant communities. Based on a survey of 42 plots of four types of evergreen broad-leaved forests in the Ailao Mountains of Yunnan Province, this paper explored how community distribution and tree species richness of such forests changed along topographic and edaphic gradients. We found significant differences in community composition among the four types of evergreen broad-leaved forests. Tree species richness of monsoon evergreen broad-leaved forests was significantly higher than that of the other forest types. Tree species richness of mid-montane moist evergreen broad-leaved forests was significantly higher than that of semi-humid evergreen broad-leaved forests, whereas no significant difference was observed among the other forest types. Non-metric multidimensional scaling (NMDS) revealed that nine environmental gradients, including elevation, slope, soil water content, soil organic matter, were the determinants of forest community composition and tree distribution patterns. As the best predictors of tree species richness, soil available boron, elevation, soil total potassium and convexity together explained 63.2% of the variance of tree species richness. The combination of topographic and edaphic factors explained most of the variance of community composition and tree species richness of the evergreen broad leaved forests in the Ailao Mountains.
云南哀牢山常绿阔叶林的空间分异及其影响因素
DOI:10.17520/biods.2016199
[本文引用: 1]
理解物种丰富度在空间上的变化, 阐明植物群落组成对生境异质性的响应是群落生态学研究的核心议题之一。与生境异质性有关的生态位分化对植物群落的多样性格局和物种分布具有重要影响。本文以分布在云南哀牢山的4种常绿阔叶林为研究对象, 通过对42个森林样地的调查取样, 探讨了常绿阔叶林的群落分布和树种丰富度在地形和土壤环境梯度上的变异。结果表明: 4种常绿阔叶林的树种组成有显著的差异。季风常绿阔叶林的树种丰富度极显著地高于其他3种类型; 中山湿性常绿阔叶林极显著地高于半湿润常绿阔叶林; 其他类型之间的差异不显著。非度量多维尺度分析显示, 海拔、坡度、土壤含水率及有机质等9个环境梯度是控制森林群落结构和树种分布的主要因素。土壤有效硼、海拔、全钾和凹凸度为树种丰富度的最佳预测变量, 累积解释了树种丰富度63.2%的变异。地形和土壤因子一起能够解释哀牢山常绿阔叶林的群落组成和树种丰富度的大部分变异。
Response of plant species composition and community structure to long-term grazing in typical steppe of Inner Mongolia
内蒙古典型草原物种组成和群落结构对长期放牧的响应
DOI:10.3773/j.issn.1005-264x.2009.03.009
[本文引用: 2]
该文比较研究了内蒙古羊草草原和大针茅草原放牧演替系列经过20 a (1985~2005年)放牧, 利用群落组成与结构的变化分析了这两个放牧演替系列上15个植物群落的变化趋势, 并依此探讨长期放牧对草原生态系统结构和功能的影响。结果表明: 在大针茅(Stipa grandis)牧压梯度系列上, 群落仍以大针茅群系为主, 冷蒿(Artemisia frigida)为建群种的群落消失; 在羊草(Leymus chinensis)牧压梯度系列上, 羊草和冷蒿不再是建群种, 被西伯利亚羽茅(Achnatherum sibiricum)、大针茅、冰草(Agropyron cristatum)和糙隐子草(Cleistogenes squarrosa)所替代。综合两个牧压梯度系列的研究结果得出以下基本结论: 长期过度放牧进一步加速了草原群落的退化进程, 但退化草原具有较高的恢复弹性, 控制放牧使部分群落得到一定程度的恢复。同一植物群落时间变化, 或不同演替阶段的植物群落在空间序列上的位移程度主要取决于放牧史和利用强度。过去20 a中, 特别是2003年以来草原管理政策的改变并没有导致该区域草地的整体退化, 而是在一定程度上有利于草地的恢复。
β-diversity of grassland communities along gradient of grazing disturbance
草地群落放牧干扰梯度β多样性研究
Effect of grazing disturbance gradient on plant diversity of main steppe grassland in the Songnen Plain
松嫩平原主要草原群落放牧干扰梯度对植物多样性的影响
Gradient analysis and environmental interpretation of understory herb-layer communities in Xiaoshegou of Lingkong Mountain, Shanxi, China
DOI:10.3724/SP.J.1258.2013.00039
[本文引用: 1]
Aims Many past practices in afforestation and forest management were instrumented for addressing the issues of tree species selection, planting regimes and development of overstory structure, but neglected understory vegetation. Our objective was to determine the controlling factors of herb-layer plant distribution and the importance of topography in determining local-scale spatial patterns of herbaceous plants.Methods The occurrence and distribution of herb-layer plants were investigated on 26 plots in the Xiaoshegou catchment of Lingkong Mountain, Shanxi Province, China. Community types were classified using two-way indicator species analysis (TWINSPAN) and the relationship between the distribution and abundance of herb-layer species and environmental gradients was analyzed using the canonical correspondence analysis (CCA) ordination method. Forward selection and Monte Carlo permutation test were used to select the factors important in determining the herb-layer plant distribution. Partial CCA (PCCA) was also performed to partition the variance that was explainable by categorical habitat and biotic factors studied.Important findings The 26 survey plots were classified into six groups characterized by the dominant overstory tree species. The results of CCA ordination reflected the relationship between herb-layer community structure and selective environmental variables. The classification of 26 plots in CCA ordination was consistent with the result of TWINSPAN. Forward selection and Monte Carlo test suggested that stand type, soil nutrients and slope position were the most important factors determining understory plant distribution. PCCA revealed that habitat and biotic factors together explained 42.9% of variance in the distribution of understory herbaceous plants. Habitat factors alone explained 31.8% of the variance, biotic factors alone explained 7.8% of the variance and interaction between habitat and biotic factors explained 3.2% of the variance. The partitioning of variance using the PCCA helped with identifying the important habitat factors regulating understory herbaceous plant distribution at the study site. However, the fact that more than half of the variance was unaccounted for by the factors studied suggests that other factors we did not measure could also play a role in determining the occurrence and distribution of herbaceous plants on the forest floor, e.g., human activities and random events. Our study demonstrates the importance of both topography and overstory tree species in determining the occurrence and distribution of herb-layer plant species in temperate forest of mountainous areas.
山西灵空山小蛇沟林下草本层植物群落梯度分析及环境解释
DOI:10.3724/SP.J.1258.2013.00039
[本文引用: 1]
通过对山西灵空山小蛇沟集水区的林下草本层植物群落进行调查和多元分析——TWINSPAN分类、典范对应分析(CCA)与生境、生物因素变量分离, 探讨林分水平上草本层物种分布与环境因子之间的关系。结果如下: 1) TWINSPAN将26个调查样方划分为6种群落类型: 以辽东栎(Quercus wutaishanica)为主的辽东栎-油松(Pinus tabulaeformis)林型、辽东栎杂木林型、辽东栎林型、华北落叶松(Larix principis-rupprechtii)林型、油松林和阔叶油松林型、油松-辽东栎均匀混交林型, 体现了该地区地带性植被类型为暖温带森林的特点。2)群落类型的划分与CCA的结果相吻合, 主要反映了CCA排序第一、二轴的环境梯度, CCA排序轴第一轴突出反映了林分类型与土壤养分梯度, 第二排序轴与坡度、坡位显著相关。Monte Carlo检验结果表明, 林分类型、土壤养分和坡度是影响小蛇沟集水区内林下草本物种分异的最主要的环境因子。3)生境因子与生物因子解释了物种格局变化的42.9%, 其中生境因子占31.8%, 生物因子占7.9%, 生境因子与生物因子交互作用解释部分占3.2%。良好的环境解释反映了调查取样和环境因子选取的合理性。对于50%以上未能被解释的变异部分, 可能归咎于未被选取的因子如干扰或者随机过程。4)在海拔梯度较小的山区, 坡向等小地形因子能较好地指示局部生境的小气候条件, 对林下植物的分布有较好的解释力。
Satellite-based vegetation production models of terrestrial ecosystem: an overview
DOI:10.11867/j.issn.1001-8166.2014.05.0541
[本文引用: 1]
Vegetation,as the principal component of terrestrial ecosystem,plays an important role in sustaining global substance and energy cycle,adjusting carbon balance and alleviating the rise of atmospheric CO2 concentration and global climate change. Vegetation production of terrestrial ecosystem has been one of the major subjects for the research on global change. The satellite-based model of vegetation productivity has undergone several stages of development,including the initial simple statistical model,the later process model based on light use efficiency principle. Based on remote sensing vegetation data with spatially and temporally continuous distribution,statistical model is crucial in estimating vegetation productivity on the regional and global scale. Statistical model can be classified into two categories: one is direct establishment of the correlation between vegetation index and vegetation productivity,based on which regional estimation is possible; the other is the establishment of regression parameter vector for regional applications,which is realized through the integrated utilization of vegetation indices and other environmental factors and using regression tree,neural network and other complex statistical methods. Light use efficiency model is the major approach to estimating vegetation productivity based on remote sensing data. However,there are large differences on the calculations of the fraction of absorbed photosynthetically active radiation,environmental stress factors,and the model performance also need improve. Future studies should continue to improve model ability,develop multiple model ensemble algorithms and provide simulation uncertainties.
陆地生态系统植被生产力遥感模型研究进展
DOI:10.11867/j.issn.1001-8166.2014.05.0541
[本文引用: 1]
陆地生态系统植被生产力一直是全球变化领域内的研究热点,对其模拟的准确与否直接决定了后续碳循环要素的模拟精度,也关系到能否准确评估陆地生态系统对人类社会可持续发展的支持能力。遥感数据因其能够提供时空连续的植被变化信息,在区域植被生产力的模拟中扮演了不可替代的角色。目前遥感模型可以分为统计模型和过程模型2类。前者主要基于植被指数等与观测值的统计关系,从最初的线性关系发展到利用回归树等多变量的统计模型。后者则是基于光能利用率原理,借助于遥感数据的时空连续性实现对区域和全球植被生产力的准确评估。然而,这些模型在计算植物冠层吸收的光合有效辐射比例、环境对最大光能利用率的限制等诸多方面存在显著的差异,对于一些关键的生态系统过程描述不完善,总体而言模拟能力仍然有待提高。此外,遥感数据也被广泛地应用于动态植被模型的发展和应用中,为模拟提供植被类型、叶面积指数等关键的输入数据。后续的研究应该进一步改进模型公式,发展集合预估算法,并应考虑由于输入数据和参数的不确定性而导致的区域模拟误差,以提高对区域植被生产力的模拟精度。
Changes in vegetation after 22 years’ natural restoration in the karst disturbed area in Northwest Guangxi
DOI:10.1016/S1872-2032(08)60016-5 URL [本文引用: 3]
桂西北喀斯特人为干扰区植被自然恢复22年后群落特征
A hypothesis for the origin and maintenance of within-community species diversity
DOI:10.17520/biods.1997026 URL [本文引用: 1]
群落内物种多样性发生与维持的一个假说
Changes of woody community structure and diversity in karst peak- cluster depressions in southwest China
西南喀斯特峰丛洼地木本植物群落结构与多样性变化
Changes in multiple ecosystem services between 2000 and 2013 and their driving factors in the Grazing Withdrawal Program, China
DOI:10.1016/j.ecoleng.2018.02.028 URL [本文引用: 1]
Research progress on the effects of grazing on grassland ecosystem
DOI:10.17521/cjpe.2019.0314 URL [本文引用: 2]
放牧对草地生态系统影响的研究进展
DOI:10.17521/cjpe.2019.0314
[本文引用: 2]
全球草地占据30%左右的陆地面积,其在全球气候变化、碳氮及养分循环、保持水土、调节畜牧业生产等方面具有重要的作用。目前草地的主要利用方式之一就是放牧,不同的牲畜种类、放牧强度、年限、历史和制度等,会影响草地植物群落、生物多样性及土壤微生物,进而影响草地生态系统结构、功能和过程。本研究围绕放牧对草地生态系统结构、功能和过程的影响,1)回顾了20世纪50年代到现在各个历史阶段放牧对草地生态系统影响研究;2)利用文献计量分析的方法,剖析了放牧对草地影响研究的热点内容、重要区域和关键词等;3)阐明了放牧对草地植物生长、群落特征、碳氮及养分循环、生产力及土壤质量等的各方面的影响研究进展及国内相关研究的优势、存在的主要问题和不足;4)基于上述分析,从草地放牧精准管理、经典假说验证、放牧和全球变化研究相结合等方面,提出未来研究的前沿方向和优先领域。本文系统在总结放牧对草地生态系统影响的研究进展、研究优势及存在问题的基础上,提出未来的研究应与全球变化相结合,为我国的草地放牧生态学研究、适应性管理和可持续利用等提供科学基础。
Species composition and community structure of a north subtropical karst secondary forest in central Guizhou Province, China
黔中北亚热带喀斯特次生林动态监测样地: 物种组成与群落结构
The effect of grazing on above-ground biomass allocation of 27 plant species in an alpine meadow plant community in Qinghai-Tibetan Plateau
放牧对青藏高原东缘高寒草甸群落27种植物地上生物量分配的影响
Species composition and community structure of Miyaluo subalpine dark coniferous forest dynamic plot in western Sichuan, China
川西米亚罗亚高山暗针叶林动态样地物种组成与群落结构
Response of relationship between community species diversity and aboveground biomass to grazing intensity in the Urat desert steppe in north China
乌拉特荒漠草原群落物种多样性和生物量关系对放牧强度的响应
Effect of long-term grazing on alpine shrub vegetation in Qinghai-Tibet Plateau
长期放牧对青藏高原高寒灌丛植被的影响
Take full advantage of shrubby grassland resources in rocky mountain for the development of goats competitive industries in Mashan County of Guangxi
充分利用石山灌草丛草地资源发展广西马山县山羊优势产业

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}