
植物生态学报 ›› 2017, Vol. 41 ›› Issue (11): 1190-1198.DOI: 10.17521/cjpe.2017.0153
李巧峡*( ), 黄小霞, 陈纹, 王镛, 孙坤
), 黄小霞, 陈纹, 王镛, 孙坤
收稿日期:2017-06-14
接受日期:2017-11-20
出版日期:2017-11-10
发布日期:2017-11-10
通讯作者:
李巧峡
基金资助:
Qiao-Xia LI*(), Xiao-Xia HUANG, Wen CHEN, Yong WANG, Kun SUN
Received:2017-06-14
Accepted:2017-11-20
Online:2017-11-10
Published:2017-11-10
Contact:
Qiao-Xia LI
摘要:
紫花地丁(Viola philippica)是具典型开放花与闭锁花混合繁育系统的植物, 其两型花的形态差异为花器官发育研究提供了极好的模型。该文以不同光周期下生长的或在不同光周期间调换的紫花地丁植株为研究对象, 基于形态解剖学方法, 研究了过渡闭锁花的形态结构、开放花到完全闭锁花或完全闭锁花到开放花的变化式样, 以及完全闭锁花与过渡闭锁花中不发育雄蕊与花瓣的位置效应。结果显示: 该物种在短日照与中日照下均有开放花与过渡闭锁花形成, 短日照下大多数花芽为开放花, 少数为过渡闭锁花, 中日照下大多数花芽为过渡闭锁花, 少数为开放花; 长日照下全为完全闭锁花。在过渡闭锁花芽中, 存在着一系列雄蕊与花瓣数目不同的过渡类型, 既有偏向开放花的5雄3瓣的过渡类型, 也有偏向完全闭锁花的2雄1瓣的过渡类型。其中, 前一种类型在短日照下的过渡闭锁花芽中所占比例较大, 而后一种类型在中日照下所占比例较大。另外, 过渡闭锁花与完全闭锁花的雄蕊与花瓣发育程度均存在一定的位置效应, 其花芽腹侧的下花瓣(对应于开放花的下花瓣)与相邻的2枚雄蕊普遍发育最好, 而后花瓣(相对于前花瓣)的发育与完全闭锁花的基本一样, 为器官原基状, 与后花瓣相邻的2枚雄蕊也普遍最小, 而且也最容易发育为无小孢子发生的膜质状结构或原基状结构。同时, 将具完全闭锁花的植株置于短日照下或将具开放花的植株置于长日照下一段时间后, 重新诱导的花芽经历一系列过渡闭锁花类型后发生花型的相互转变。因此, 不同光周期对紫花地丁两型花发育的影响是渐进的, 长日照会抑制部分花瓣与雄蕊的发育, 短日照能拮抗并破除长日照对雄蕊与花瓣的抑制。
李巧峡, 黄小霞, 陈纹, 王镛, 孙坤. 紫花地丁两型花相互转变过程中花部形态结构的变化式样研究. 植物生态学报, 2017, 41(11): 1190-1198. DOI: 10.17521/cjpe.2017.0153
Qiao-Xia LI, Xiao-Xia HUANG, Wen CHEN, Yong WANG, Kun SUN. Patterns of flower morphology and structural changes during interconversion between chasmogamous and cleistogamous flowers in Viola philippica. Chinese Journal of Plant Ecology, 2017, 41(11): 1190-1198. DOI: 10.17521/cjpe.2017.0153

图1 紫花地丁两型花的表型变异。A, 开放花。B, 完全闭锁花。C-J, 过渡闭锁花。K, 开放花花芽的横切面。L, 完全闭锁花花芽的横切面。M-O, 过渡闭锁花花芽的横切面。ca, 心皮; fi, 花丝; pe, 花瓣; se, 花萼; st, 雄蕊。A-J, 比例尺为500 μm; K, 比例尺为200 μm; L-O, 比例尺为100 μm。
Fig. 1 The phenotype variation of dimorphic flower in Viola philippica. A, Chasmogamous flower. B, Cleistogamous flower. C-J, Intermediate cleistogamous flower. K, The cross section of chasmogamous flower. L, The cross section of cleistogamous flower. M-O, The cross section of intermediate cleistogamous flower. ca, carpel; fi, filament; pe, petal; se, sepal; st, stamen. A-J, bar = 500 μm; K, bar = 200 μm; L-O, bar = 100 μm.

图2 紫花地丁两型花中雄蕊形态与数量的变异。A, 开放花中的5枚雄蕊。B-L, 过渡闭锁花中不同数目的雄蕊。M, 完全闭锁花中的2枚雄蕊。an, 花药; fi, 花丝; sc, 附属结构-雄蕊帽。比例尺为500 μm。
Fig. 2 The morphological and number variation of stamens in dimorphic flower of Viola philippica. A, The five stamens in chasmogamous flower. B-L, The different number of stamens in intermediate cleistogamous flower. M, The two stamens in cleistogamous flower. an, anther; fi, filament; sc, stamen cap, the membranous appendage structure. Bar = 500 μm.
| 光周期 Photoperiod (light/dark) | 开放花植株比率 The ratio of plant with CH flowers (%) | 5枚或4枚雄蕊、3个花瓣过渡闭锁花植株比率 The ratio of plant with inCL flowers of 5 or 4 stamens and 3 petals (%) | 3枚雄蕊、3个花瓣过渡闭锁花植株比率 The ratio of plant with inCL flowers of 3 stamens and 3 petals (%) | 2枚雄蕊、3个花瓣过渡闭锁花植株比率 The ratio of plant with inCL flowers of 2 stamens and 3 petals (%) | 2枚雄蕊、1个花瓣过渡闭锁花植株比率 The ratio of plant with inCL flowers of 2 stamens and 1 petal (%) | 完全闭锁花植株比率 The ratio of plant with CL flowers (%) |
|---|---|---|---|---|---|---|
| 10 h/14 h | 81.93 ± 0.016d | 6.40± 0.011c | 4.73 ± 0.012c | 4.87 ± 0.006c | 2.10 ± 0.008b | 0.00 ± 0.000a |
| 12 h/12 h | 3.50 ± 0.004b | 4.70 ± 0.014b | 9.27 ± 0.009c | 23.80 ± 0.032d | 59.57 ± 0.008e | 0.00 ± 0.000a |
| 16 h/8 h | 0.00 ± 0.000a | 0.00 ± 0.000a | 0.00 ± 0.000a | 0.00 ± 0.000a | 0.00 ± 0.000a | 100.00 ± 0.000b |
表1 紫花地丁不同光周期对过渡闭锁花形态结构的影响(平均值±标准误差)
Table 1 The effect of different photoperiod on the morphological structure of intermediate cleistogamous flowers of Viola philippica (mean ± SE)
| 光周期 Photoperiod (light/dark) | 开放花植株比率 The ratio of plant with CH flowers (%) | 5枚或4枚雄蕊、3个花瓣过渡闭锁花植株比率 The ratio of plant with inCL flowers of 5 or 4 stamens and 3 petals (%) | 3枚雄蕊、3个花瓣过渡闭锁花植株比率 The ratio of plant with inCL flowers of 3 stamens and 3 petals (%) | 2枚雄蕊、3个花瓣过渡闭锁花植株比率 The ratio of plant with inCL flowers of 2 stamens and 3 petals (%) | 2枚雄蕊、1个花瓣过渡闭锁花植株比率 The ratio of plant with inCL flowers of 2 stamens and 1 petal (%) | 完全闭锁花植株比率 The ratio of plant with CL flowers (%) |
|---|---|---|---|---|---|---|
| 10 h/14 h | 81.93 ± 0.016d | 6.40± 0.011c | 4.73 ± 0.012c | 4.87 ± 0.006c | 2.10 ± 0.008b | 0.00 ± 0.000a |
| 12 h/12 h | 3.50 ± 0.004b | 4.70 ± 0.014b | 9.27 ± 0.009c | 23.80 ± 0.032d | 59.57 ± 0.008e | 0.00 ± 0.000a |
| 16 h/8 h | 0.00 ± 0.000a | 0.00 ± 0.000a | 0.00 ± 0.000a | 0.00 ± 0.000a | 0.00 ± 0.000a | 100.00 ± 0.000b |
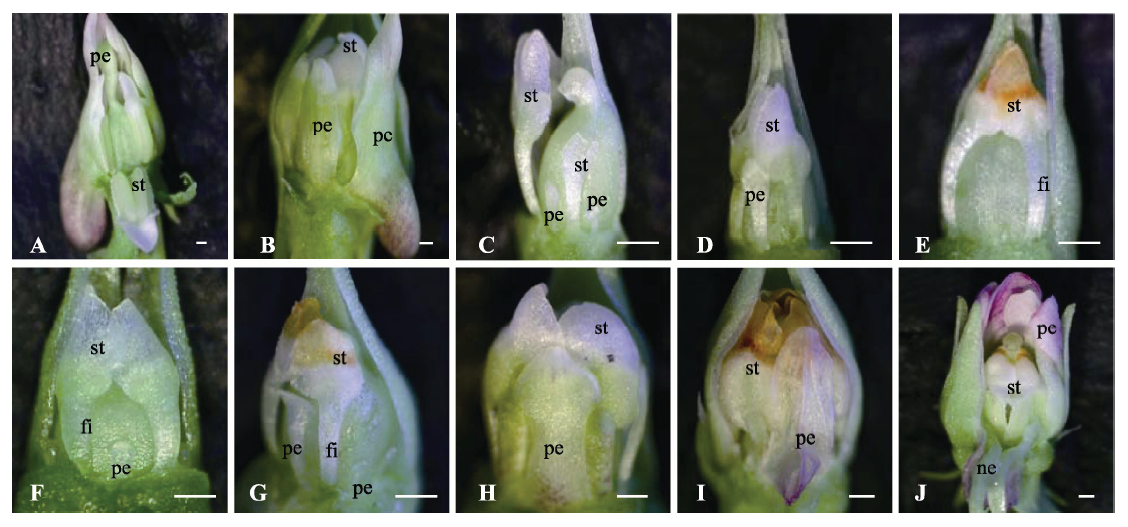
图3 紫花地丁开放花与完全闭锁花植株在一定光周期下的花型变化趋势。A-E, 具有开放花的植株置于16 h光照时间后花芽形态的变化趋势。A, 0天时的花芽形态。B, 10天时的花芽形态。C, 20天时的花芽形态。D, 30天时的花芽形态。E, 40天时的花芽形态。F-J, 具有完全闭锁花的植株置于10 h光照时间后花芽形态的变化趋势。F, 0天时的花芽形态。G, 20天时的花芽形态。H, 40天时的花芽形态。I, 60天时的花芽形态。J, 80天时的花芽形态。fi, 花丝; ne, 蜜腺体; pe, 花瓣; st, 雄蕊。比例尺500 μm。
Fig. 3 The variation trends of flowers type of the plants with chasmogamous or cleistogamous flowers under different photoperiod in Viola philippica. A-E, The variation trends of flowers type of the plants with chasmogamous flowers under 16 h daylight. A, The morphological structure of flowers at 0 days. B, The morphological structure of flowers at 10 days. C, The morphological structure of flowers at 20 days. D, The morphological structure of flowers at 30 days. E, The morphological structure of flowers at 40 days. F-J, The variation trends of flowers type of the plants with cleistogamous flowers under 10 h daylight. F, The morphological structure of flowers at 0 days. G, The morphological structure of flowers at 20 days. H, The morphological structure of flowers at 40 days. I, The morphological structure of flowers at 60 days. J, The morphological structure of flowers at 80 days. fi, filament; ne, nectar; pe, petal; st, stamen. Bar = 500 μm.
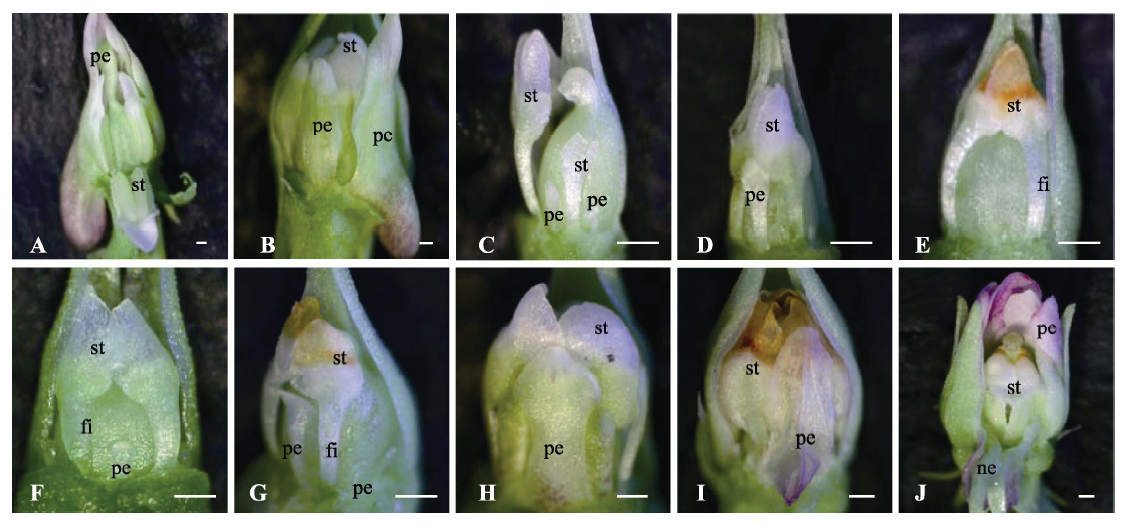
图4 不同光周期下紫花地丁两型花花器官大小的变化趋势。A, B, 开放花植株在16 h日照下新诱导花芽花器官大小的变化趋势。C, D, 完全闭锁花植株在10 h日照下新诱导花芽花器官大小的变化趋势。AnL, 花药的长度; FiL, 花丝的长度; LpeL, 下花瓣的长度; LpeW, 下花瓣的宽度。不同小写字母表示不同时间段下花器官大小之间存在显著差异(p < 0.05)。
Fig. 4 The variation trends of floral organ size in dimorphic flowers of Viola philippica under different photoperiod. A, B, The variation trends of flowers organ size in newly developed floral buds of the plants with chasmogamous flowers under 16 h daylight. C, D, the variation trends of flowers organ size in newly developed floral buds of the plants with cleistogamous flowers under 10 h daylight. AnL, the length of anther; FiL, the length of filament; LpeL, the length of lower petal; LpeW, the width of lower petal. The different lowercase letters indicated that there were significant differences in the size of floral organs as the time went on (p < 0.05).
| [1] | Ainsworth C, Crossley S, Buchanan-Wollaston V, Thangavelu M, Parke J (1995). Male and female flowers of the dioecious plant Sorrel show different patterns of MADS box gene expression.The Plant Cell, 7, 1583-1598. |
| [2] |
Auge H, Neuffer B, Erlinghagen F, Grupe R, Brandl R (2001). Demographic and random amplified polymorphic DNA analyses reveal high levels of genetic diversity in a clonal violet.Molecular Ecology, 10, 1811-1819.
DOI URL PMID |
| [3] |
Beattie AJ (1976). Plant dispersion, pollination and gene flow inViola. Oecologia, 25, 291-300.
DOI URL PMID |
| [4] | Campbell CS (1982). Cleistogamy inAndropogon. American Journal of Botany, 69, 1625-1635. |
| [5] |
Campbell CS, Quinn JA, Cheplick GP, Bell TJ (1983). Cleistogamy in grasses.Annual Review of Ecology and Systematic, 14, 411-441.
DOI URL |
| [6] |
Culley TM, Klooster MR (2007). The cleistogamous breeding system: A review of its frequency, evolution, and ecology in angiosperms. Botanical Review, 73, 1-30.
DOI URL |
| [7] |
Culley TM, Wolfe A (2001). Population genetic structure of the cleistogamous plant speciesViola pubescens Aiton (Violaceae), as indicated by allozyme and ISSR molecular markers. Heredity, 86, 545-556.
DOI URL PMID |
| [8] |
Hardenack S, Ye D, Saedler H, Grant S (1994). Comparison of MADS box gene expression in developing male and female flowers of the dioecious plant white campion.The Plant Cell, 6, 1775-1787.
DOI URL PMID |
| [9] |
Kater MM, Franken J, Carney KJ, Colombo L, Angenent GC (2001). Sex determination in the monoecious species cucumber is confined to specific floral whorls.The Plant Cell, 13, 481-493.
DOI URL PMID |
| [10] |
Lange MJP, Knop N, Lange T (2012). Stamen-derived bioactive gibberellin is essential for male flower development of Cucurbita maxima L. Journal of Experimental Botany, 63, 2681-2691.
DOI URL |
| [11] |
Lange MJP, Lange T (2016). Ovary-derived precursor gibberellin A9 is essential for female flower development in cucumber.Development, 143, 4425-4429.
DOI URL PMID |
| [12] | Lee CW, Erickson HT, Janick J (1978). Chasmogamous and cleistogamous pollination inSalpiglossis sinuata. Physiologia Plantarum, 43, 225-230. |
| [13] |
Li QX, Huo QD, Wang J, Zhao J, Sun K, He CY (2016). Expression of B-class MADS-box genes in response to variations in photoperiod is associated with chasmogamous and cleistogamous flower development inViola philippica. BMC Plant Biology, 16, 151.
DOI URL PMID |
| [14] |
Li Z, Liu G, Zhang J, Lu S, Yi S, Bao M (2012). Cloning and characterization of paleoAP3-like MADS-box gene in London plane tree. Biologia Plantarum, 56, 585-589.
DOI URL |
| [15] |
Liu QL, Liu XW, Liu WW, Liao WJ (2006). Comparison between cleistogamy and chasmogamy in perennial herb Viola philippica (Violaceae). Journal of Beijing Normal University (Science Edition), 42, 605-609. (in Chinese with English abstract)[刘绮丽, 刘香梅, 刘薇薇, 廖万金 (2006). 紫花地丁开放花和闭锁花繁殖特征的研究. 北京师范大学学报(自然科学版), 42, 606-610.]
DOI URL |
| [16] |
Lord EM (1979). Physiological controlson the production of cleistogamous and chasmogamous flowers in Lamium amplexicaule L.(Labiatae). Annals of Botany, 44, 757-766.
DOI URL |
| [17] | Lord EM (1981). Cleistogamy: A tool for the study of floral morphogenesis function and evolution.Botanical Review, 47, 421-450. |
| [18] | Malobecki A, Marcussen T, Bohdanowicz J, Migdalek G, Slomka A, Kuta E (2016). Cleistogamy and phylogenetic position ofViola uliginosa(Violaceae) re-examined. Botanical Journal of the Linnean Society, 182, 180-194. |
| [19] |
Mayers AM, Lord EM (1983). Comparative flower development in the cleistogamous speciesViola odorata. I. A growth rate study. American Journal of Botany, 70, 1548-1555.
DOI URL |
| [20] | Minter TC, Lord EM (1983). Effects of water stress, abscisicacid, and gibberellic acid on flower production and differentiation in the cleistogamous speciesCollomia grandiflora Dougl. ex Lindl.(Polemoniaceae). American Journal of Botany, 70, 618-624. |
| [21] |
Morinaga SI, Nagano AJ, Miyazaki S, Kubo M, Demura T, Fukuda H, Sakai S, Hasebe M (2008). Ecogenomics of cleistogamous and chasmogamous flowering: Genome- wide gene expression patterns from cross-species microarrayanalysis inCardamine kokaiensis(Brassicaceae). Journal of Ecology, 96, 1086-1097.
DOI URL |
| [22] |
Munguía-Rosas MA, Campos-Navarrete MJ, Parra-Tabla V (2013). The effect of pollen source vs. flower type on progeny performance and seed predation under contrasting light environments in a cleistogamous herb.PLOS ONE, 8, e80934. doi: 10.1371/journal.pone.0080934.
DOI URL PMID |
| [23] |
Paoletti C, Holsinger E (1999). Spatial patterns of polygenic variation in Impatiens capensis, a species with an environmentally controlled mixed mating system. Journal of Evolutionary Biology, 12, 689-696.
DOI URL |
| [24] |
Schemske DW (1978). Evolution of reproductive characteristics inImpatiens(Balsaminaceae): The significance of cleistogamy and chasmogamy. Ecology, 59, 596-613.
DOI URL |
| [25] |
Schoen DJ, Lloyd DG (1984). The selection of cleistogamy and heteromorphic diaspores.Biological Journal of the Linnean Society, 23, 303-322.
DOI URL |
| [26] | Sigrist MR, Sazima M (2002). Ruellia brevifolia (Pohl) Ezcurra (Acanthaceae): Flowering phenology, pollination biology and reproduction. Brazilian Journal of Botany, 25, 35-42. |
| [27] |
Solbrig OT (1976). On the relative advantages of cross- and self-fertilization.Annals of the Missouri Botanical Garden, 63, 262-276.
DOI URL |
| [28] |
Stebbins GL (1957). Self fertilization and population variability in the higher plants.The American Naturalist, 91, 337-354.
DOI URL |
| [29] | Teaching and Research Section of Plant Microscopy Set in Huazhong Agricultural College (1984). Love’s hematoxylin dyeing as a whole and red-solid double drop staining in the application of paraffin section.Chinese Bulletin of Botany, 3, 56.[华中农学院植物教研室植物显微技术组 (1984). 爱氏苏木精整体染色及番红-固绿双重染色滴染法在石蜡切片中的运用. 植物学通报, 3, 56.] |
| [30] | Uphof JCT (1938). Cleistogamic flowers. Botanical Review, 4, 21-49. |
| [31] |
Wahlert GA, Marcussen T, Paula-Souza JD, Feng M, Ballard HE (2014). A phylogeny of the Violaceae (Malpighiales) inferred from plastid DNA sequences: Implications for generic diversity and intrafamilial classification.Systematic Botany, 39, 239-252.
DOI URL |
| [32] |
Waller DM (1979). The relative costs of self- and cross-fertilized seeds in Impatiens capensis(Balsaminaceae). American Journal of Botany, 66, 313-320.
DOI URL |
| [33] |
Waller DM (1984). Differences in fitness between seedlings derived from cleistogamous and chasmogamous flowers inImpatiens capensis. Evolution, 38, 427-440.
DOI URL PMID |
| [34] |
Wang Y, Ballard HE, McNally RR, Wyatt SE (2013). Gibberellins are involved but not sufficient to trigger a shift between chasmogamous-cleistogamous flower types inViola pubescens. Journal of the Torrey Botanical Society, 140, 1-8.
DOI URL |
| [35] |
Wang Y, Li QX, Sun K, Chen W (2017). The study on dimorphic flower development and the soluble sugar and starch content inViola philippica. Acta Horticulturae Sinica, 44, 323-329. (in Chinese with English abstract)[王镛, 李巧峡, 孙坤, 陈纹 (2017). 紫花地丁开放花与闭锁花的发育及可溶性糖与淀粉含量的研究. 园艺学报, 44, 323-329.]
DOI URL |
| [1] | 徐申林, 刘文哲. 喜树花序内性别表达及性别分配[J]. 植物生态学报, 2011, 35(12): 1290-1299. |
| [2] | 谢田朋, 杜国祯, 张格非, 赵志刚. 黄帚橐吾种子生产的花序位置效应及其对幼苗建植的影响[J]. 植物生态学报, 2010, 34(4): 418-426. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19