
植物生态学报 ›› 2023, Vol. 47 ›› Issue (9): 1193-1210.DOI: 10.17521/cjpe.2022.0445
所属专题: 生物多样性
• 综述 • 下一篇
杨明威, 金晓芳*( )
)
收稿日期:2022-11-04
接受日期:2023-03-01
出版日期:2023-09-20
发布日期:2023-09-28
通讯作者:
* 金晓芳(基金资助:
YANG Ming-Wei, JIN Xiao-Fang*()
Received:2022-11-04
Accepted:2023-03-01
Online:2023-09-20
Published:2023-09-28
Contact:
* JIN Xiao-Fang(Supported by:摘要:
蜜距将花蜜隐藏在距的深处, 多样的蜜距形态和长度吸引各种喙长的传粉动物访花, 促进传粉生态位形成, 这对造就物种多样性起到了重要作用。因此蜜距被认为是促进物种形成的关键创新性状, 是研究被子植物花部结构演化、植物-传粉者协同进化的理想材料。系统地总结蜜距植物及其形态特征和传粉/盗蜜者, 有助于全面了解这一性状在被子植物中的系统发育分布和进化生态学意义。该文对国内外植物分类数据库(检索词“距”或“spur”)和已发表文章(检索词“有距植物属中文名+传粉”或“属拉丁名+ pollinator/nectar robber”)进行系统的梳理和总结, 收集蜜距植物、距长、蜜距形态、传粉者、盗蜜者等信息。整理出具有蜜距的植物分布在被子植物13个目23个科271个属, 共计3 427种。其中蜜距植物种数最多的科为兰科(1 536种)、罂粟科(487种)、毛茛科(351种)、凤仙花科(284种)、堇菜科(197种), 它们占所有蜜距植物种数的83.3%。根据蜜距的发育来源, 蜜距可分为6种类型: 花被片蜜距、花瓣蜜距、萼片蜜距、被丝托蜜距、花萼蜜距和花冠蜜距。花瓣蜜距分布于212个属, 显著高于其他类型。对2 546种植物的蜜距长度分析发现: 凤仙花科的蜜距平均长度最长, 达23.8 mm; 千屈菜科的蜜距最短, 平均仅1.6 mm。形态上, 大部分蜜距具有不同程度的弯曲, 这可能有助于筛选合适的传粉者, 提高访花者的传粉效率; 单花蜜距数量有1-6个, 大部分为1个, 多个蜜距可能增加传粉者的访花时间; 部分兰科和堇菜科植物蜜距内部具有可能分泌花蜜的附属物。蜜距植物的传粉者主要为膜翅目、鳞翅目、部分长喙双翅目昆虫及鸟类; 盗蜜者主要为膜翅目昆虫。蜜距的长度和传粉者的喙长普遍存在地理镶嵌的协同进化关系, 进化生态学家提出了“达尔文机械模型”的假说来解释这种现象; 而当传粉者喙长不变, 距长随着传粉者种类改变时则用“传粉者转变模型”来解释蜜距的伸长。未来在蜜距的进化生态学研究中, 不仅要关注蜜距植物的距长和传粉者喙长的形态学特征及传粉生态学研究, 还应借助发育解剖学和基因组学, 以及新的技术手段, 从细胞的分裂、伸长, 功能基因, 非生物因子等各个方面, 探索蜜距演化的驱动因素。
杨明威, 金晓芳. 被子植物蜜距的多样性及进化生态学研究. 植物生态学报, 2023, 47(9): 1193-1210. DOI: 10.17521/cjpe.2022.0445
YANG Ming-Wei, JIN Xiao-Fang. Diversity and evolutionary ecology of nectar spurs in angiosperms. Chinese Journal of Plant Ecology, 2023, 47(9): 1193-1210. DOI: 10.17521/cjpe.2022.0445

图1 被子植物蜜距的多样性。A, 二叶舌唇兰(兰科)。B, 木鱼坪淫羊藿(小檗科)。C, 中甸乌头(毛茛科)。D, 华北耧斗菜(毛茛科)。E, 早开堇菜(堇菜科)。F, 囊距紫堇(罂粟科)。G, 大花花锚(龙胆科)。H, 摩洛哥柳穿鱼(车前科)。I, 黄花狸藻(狸藻科)。J, 凤仙花属(凤仙花科)。K, 油点草(百合科)。L, 旱金莲(旱金莲科)。其中A-F为花瓣蜜距, G-I为花冠蜜距, J为萼片蜜距, K为花被片蜜距, L为被丝托蜜距。白色箭头所指为各种形态的蜜距。拍摄者: 金晓芳。
Fig. 1 Diversity of nectar spurs in angiosperms. A, Platanthera chlorantha (Orchidaceae). B, Epimedium franchetii (Berberidaceae). C, Aconitum piepunense (Ranunculaceae). D, Aquilegia yabeana (Ranunculaceae). E, Viola prionantha (Violaceae). F, Corydalis benecincta (Papaveraceae). G, Halenia elliptica var. grandiflora (Gentianaceae). H, Linaria maroccana (Plantaginaceae). I, Utricularia aurea (Lentibulariaceae). J, Impatiens sp. (Balsaminaceae). K, Tricyrtis macropoda (Liliaceae). L, Tropaeolum majus (Tropaeolaceae). A-F are petal spurs, G-I are corolla spurs, J is sepal spur, K is tepal spur, and L is hypanthium spur. The white arrows indicate the various forms of the nectar spurs. Photoed by JIN Xiao-Fang.
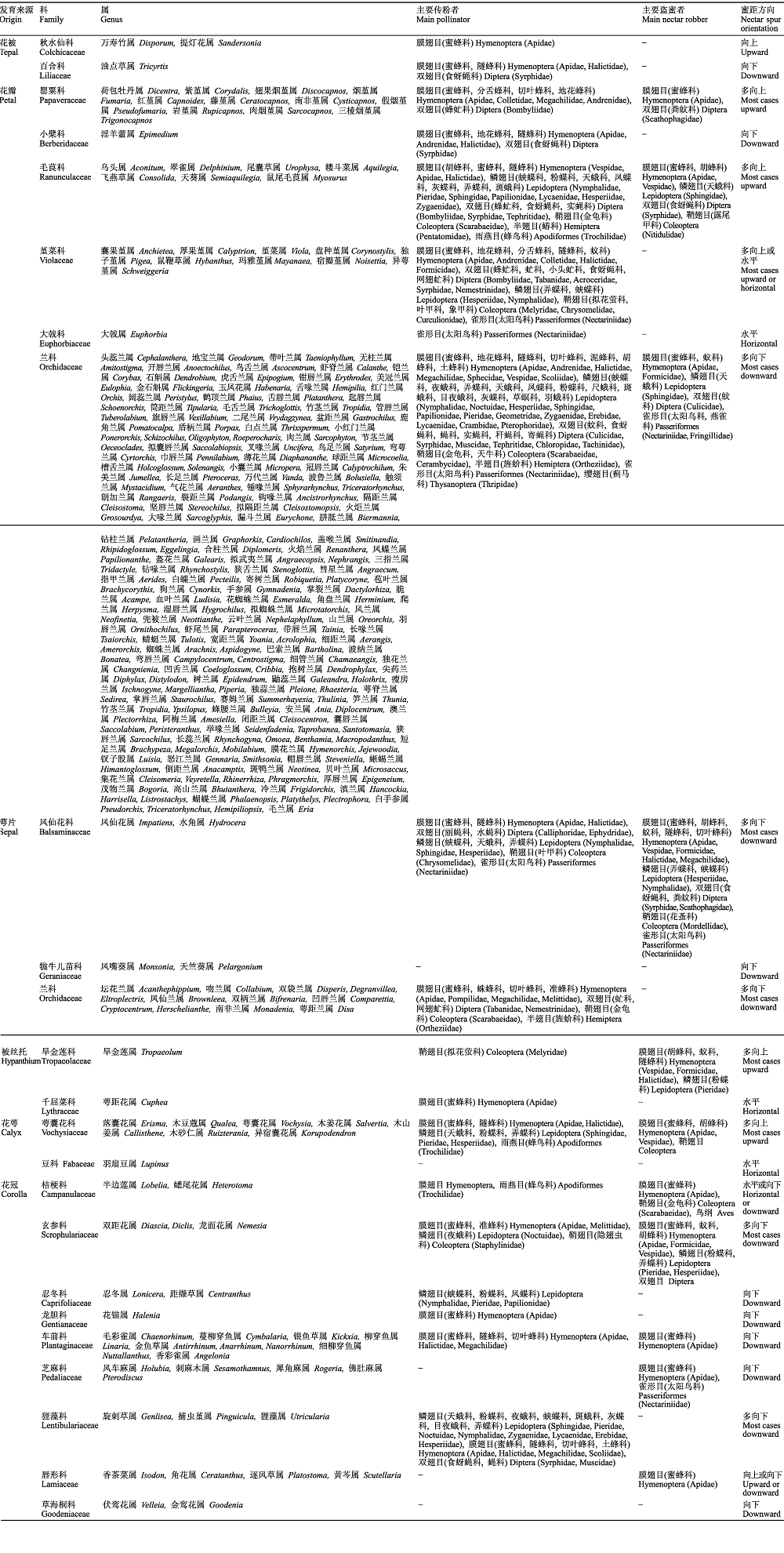 |
表1 被子植物蜜距的发育来源及其系统分布
Table 1 Origin and phylogenetic distribution of nectar spurs in angiosperms
|
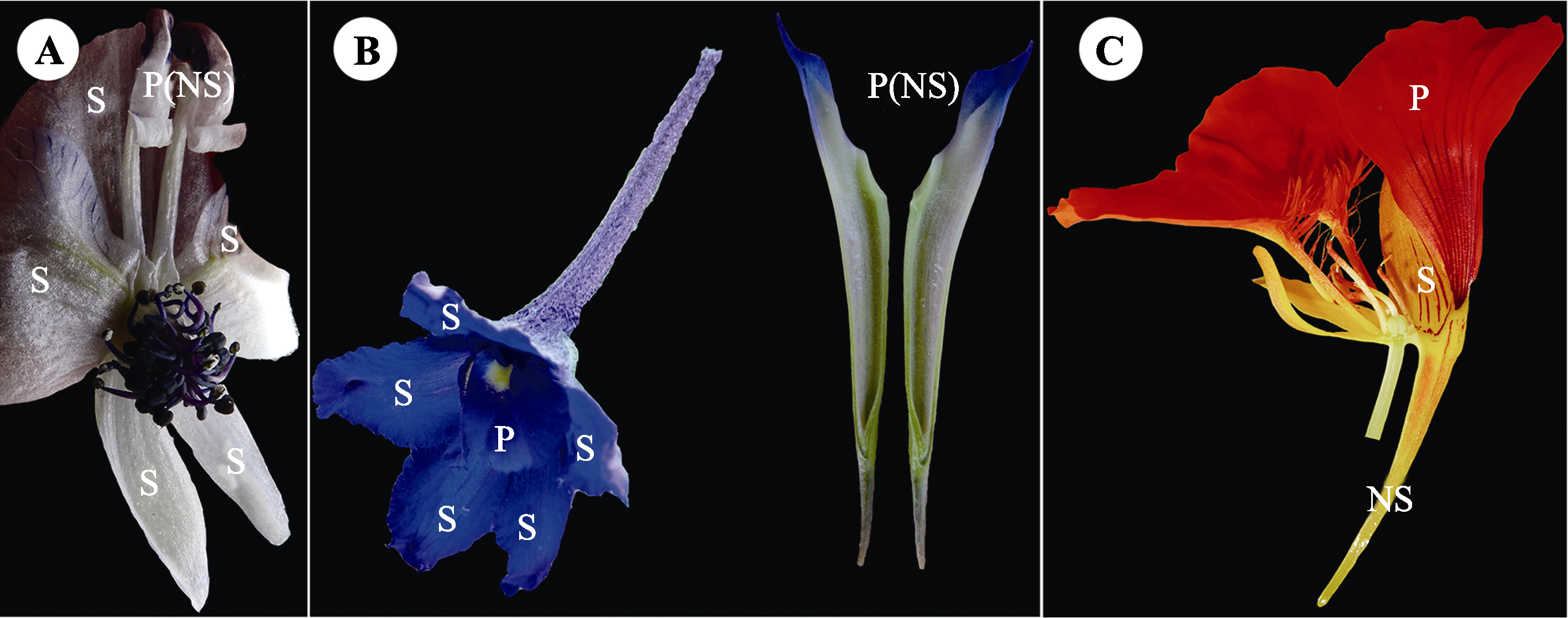
图2 花瓣卷缩形成的蜜距(翠雀属、乌头属)和被丝托蜜距的结构解剖。A, 中甸乌头的花瓣蜜距。B, 中甸翠雀花的花瓣蜜距(被包裹在萼距里面)。C, 旱金莲的被丝托蜜距。NS, 蜜距; P, 花瓣; S, 萼片。拍摄者: 金晓芳。
Fig. 2 Nectar spur formed by petal rolling (Aconitum, Delphinium) and hypanthium spur. A, Petal spur of A. piepunense. B, Petal spur of D. yuanum (encased in calyx spur). C, Hypanthium spur of Tropaeolum majus. NS, nectar spur; P, petal; S, sepal. Photoed by JIN Xiao-Fang.
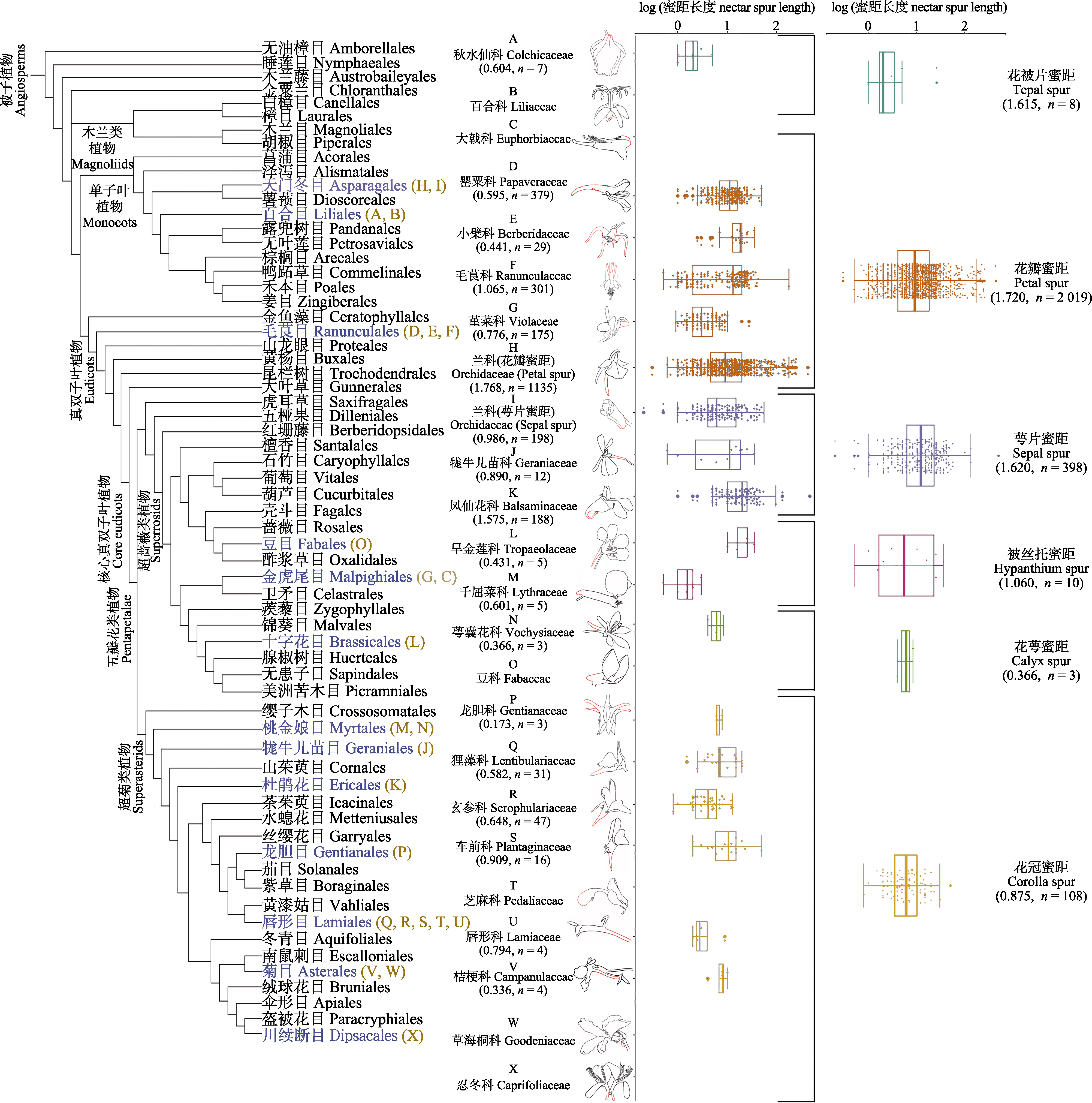
图3 被子植物蜜距在目一级的系统发育分布。系统树根据Angiosperm phylogeny poster (APP)—Flowering plant systematics, 2019绘制(Cole et al., 2019)。线描图红色部分为蜜距所在位置。植物科和蜜距类型所附括号内为距长变异系数和样本量。两列箱型图分别为17个科和6种蜜距类型的蜜距长度log值(平均值±标准差)。
Fig. 3 Distribution of nectar spur types across angiosperm orders. The phylogeny of angiosperm orders is drawn according to the Angiosperm phylogeny poster (APP)—Flowering plant systematics, 2019 (Cole et al., 2019). The red parts of the line diagrams show the location of the nectar spurs. Coefficient of variation of spur length and sample size are enclosed in parentheses for plant families and nectar spur types. The log values (mean ± SD) of spur length for 17 families and 6 types of nectar spur are shown in the two box plots.
| 标准差 Standard deviation | 平均值 Average value | 数据量 Amount of data | 传粉者属数 Genus number of pollinators | |||||
|---|---|---|---|---|---|---|---|---|
| r | p | r | p | r | p | r | p | |
| 蜜距类型 Type of nectar spur | 0.781 | 0.067 | 0.563 | 0.245 | 0.542 | 0.266 | 0.413 | 0.416 |
| 科 Family | 0.889 | <0.001 | 0.502 | 0.034 | 0.716 | <0.001 | 0.752 | <0.001 |
| 属 Genus | 0.699 | <0.001 | 0.371 | 0.107 | 0.099 | 0.679 | -0.079 | 0.808 |
表2 距长变异系数与距长其他特征的相关分析统计结果
Table 2 Statistical results of correlation analysis between the coefficient of variation of nectar spur length with other characteristics
| 标准差 Standard deviation | 平均值 Average value | 数据量 Amount of data | 传粉者属数 Genus number of pollinators | |||||
|---|---|---|---|---|---|---|---|---|
| r | p | r | p | r | p | r | p | |
| 蜜距类型 Type of nectar spur | 0.781 | 0.067 | 0.563 | 0.245 | 0.542 | 0.266 | 0.413 | 0.416 |
| 科 Family | 0.889 | <0.001 | 0.502 | 0.034 | 0.716 | <0.001 | 0.752 | <0.001 |
| 属 Genus | 0.699 | <0.001 | 0.371 | 0.107 | 0.099 | 0.679 | -0.079 | 0.808 |

图4 蜜距植物的主要访花者和盗蜜者。A, 南熊蜂访中甸翠雀花。B, 独居蜂访柔毛淫羊藿。C, 条蜂访刻叶紫堇。D, 熊蜂访椭圆叶花锚。E, 熊蜂访顶喙凤仙花。F, 条蜂访摩洛哥柳穿鱼。G, 条蜂盗蜜地锦苗。H, 黄胸木蜂盗蜜凤仙花。拍摄者: 金晓芳。
Fig. 4 Pollinators and nectar robbers of some plants with nectar spurs. A, Bombus secures visiting Delphinium yuanum. B, Solitary bee visiting Epimedium pubescens. C, Anthophoridae bee visiting Corydalis incisa. D, Bombus sp. visiting Halenia elliptica. E, Bombus sp. visiting Impatiens compta. F, Anthophoridae bee visiting Linaria maroccana. G, Anthophoridae bee robbing nectar of Corydalis sheareri. H, Xylocopa appendiculata robbing nectar of Impatiens balsamina. Photoed by JIN Xiao-Fang.
| [1] |
Almouslem AB, Tilney-Bassett RAE (1989). The inheritance of flower doubleness and nectary spur in Pelargonium × hortorum Bailey. Euphytica, 41, 23-29.
DOI URL |
| [2] |
Antoń S, Kamińska M (2015). Comparative floral spur anatomy and nectar secretion in four representatives of Ranunculaceae. Protoplasma, 252, 1587-1601.
DOI PMID |
| [3] |
Ayers TJ (1987). Four species from western Mexico new to Lobelia (Campanulaceae: Lobelioideae). Brittonia, 39, 417-422.
DOI URL |
| [4] |
Ayers TJ (1990). Systematics of Heterotoma (Campanulaceae) and the evolution of nectar spurs in the New World Lobelioideae. Systematic Botany, 15, 296-327.
DOI URL |
| [5] |
Bateman RM, Rudall PJ, James KE (2006). Phylogenetic context, generic affinities and evolutionary origin of the enigmatic Balkan orchid Gymnadenia frivaldii Hampe ex Griseb. Taxon, 55, 107-118.
DOI URL |
| [6] | Bateman RM, Sexton R (2008). Is spur length of Platanthera species in the British Isles adaptively optimized or an evolutionary red herring? Watsonia, 27, 1-22. |
| [7] |
Bell AK, Roberts DL, Hawkins JA, Rudall PJ, Box MS, Bateman RM (2009). Comparative micromorphology of nectariferous and nectarless labellar spurs in selected clades of subtribe Orchidinae (Orchidaceae). Botanical Journal of the Linnean Society, 160, 369-387.
DOI URL |
| [8] |
Blaxland K (2004). A new species of Viola (Violaceae) from south-west Turkey. Botanical Journal of the Linnean Society, 145, 505-509.
DOI URL |
| [9] |
Boberg E, Alexandersson R, Jonsson M, Maad J, Ågren J, Nilsson LA (2014). Pollinator shifts and the evolution of spur length in the moth-pollinated orchid Platanthera bifolia. Annals of Botany, 113, 267-275.
DOI PMID |
| [10] |
Boehm MMA, Jankowski JE, Cronk QCB (2022). Plant- pollinator specialization: origin and measurement of curvature. The American Naturalist, 199, 206-222.
DOI URL |
| [11] | Box MS (2010). Role of KNOX Genes in the Evolution and Development of Floral Nectar Spurs. PhD dissertation, University of Cambridge, Cambridge, UK. |
| [12] |
Box MS, Bateman RM, Glover BJ, Rudall PJ (2008). Floral ontogenetic evidence of repeated speciation via paedomorphosis in subtribe Orchidinae (Orchidaceae). Botanical Journal of the Linnean Society, 157, 429-454.
DOI URL |
| [13] |
Box MS, Dodsworth S, Rudall PJ, Bateman RM, Glover BJ (2011). Characterization of Linaria KNOX genes suggests a role in petal-spur development. The Plant Journal, 68, 703-714.
DOI URL |
| [14] |
Bruce EA (1953). Notes on African Pedaliaceae. Kew Bulletin, 8, 417-429.
DOI URL |
| [15] |
Brunet J, van Etten ML (2019). The response of floral traits associated with pollinator attraction to environmental changes expected under anthropogenic climate change in high-altitude habitats. International Journal of Plant Sciences, 180, 954-964.
DOI URL |
| [16] |
Carmo-Oliveira R, Oliveira PE, Lange de Morretes B (2017). Appendicular origin and structure of the spur of Vochysiaceae flowers. Acta Botanica Brasilica, 31, 433-444.
DOI URL |
| [17] |
Chapurlat E, Ågren J, Sletvold N (2015). Spatial variation in pollinator-mediated selection on phenology, floral display and spur length in the orchid Gymnadenia conopsea. New Phytologist, 208, 1264-1275.
DOI PMID |
| [18] |
Chowlu K, Angela N, Rao AN, Vij SP (2012). Two new additions to the orchid flora of Manipur, India. Kew Bull, 16, 1.
DOI URL |
| [19] | Cole TCH, Hilger HH, Stevens P (2019). Angiosperm phylogeny poster (APP)—Flowering plant systematics, 2019. PeerJ Preprints. DOI: 10.7287/peerj.preprints.2320v6. |
| [20] | Cronk QCB, Bateman RM, Hawkins JA (2002). Developmental Genetics and Plant Evolution. CRC Press, Boca Raton. |
| [21] | Cullen E (2020). The Evolution and Development of Nectar Spurs. PhD dissertation, University of Cambridge, Cambridge, UK. |
| [22] |
Cullen E, Fernández-Mazuecos M, Glover BJ (2018). Evolution of nectar spur length in a clade of Linaria reflects changes in cell division rather than in cell expansion. Annals of Botany, 122, 801-809.
DOI PMID |
| [23] |
Dodsworth S (2017). Petal, sepal, or tepal? B-genes and monocot flowers. Trends in Plant Science, 22, 8-10.
DOI PMID |
| [24] |
Don D (1841). A monograph of the genus Disporum. Transactions of the Linnean Society of London, 18, 513-524.
DOI URL |
| [25] |
Dransfield J, Comber JB, Smith G (1986). A synopsis of Corybas (Orchidaceae) in west Malesia and Asia. Kew Bulletin, 41, 575-613.
DOI URL |
| [26] |
Edwards MB, Ballerini ES, Kramer EM (2022). Complex developmental and transcriptional dynamics underlie pollinator-driven evolutionary transitions in nectar spur morphology in Aquilegia (columbine). American Journal of Botany, 109, 1360-1381.
DOI PMID |
| [27] |
Ellis AG, Johnson SD (1999). Do pollinators determine hybridization patterns in sympatric Satyrium (Orchidaceae) species? Plant Systematics and Evolution, 219, 137-150.
DOI URL |
| [28] | Endress PK, Matthews ML (2006). Elaborate petals and staminodes in eudicots: diversity, function, and evolution. Organisms Diversity & Evolution, 6, 257-293. |
| [29] |
Fernández-Mazuecos M, Blanco-Pastor JL, Juan AN, Carnicero P, Forrest A, Alarcón M, Vargas P, Glover BJ (2019). Macroevolutionary dynamics of nectar spurs, a key evolutionary innovation. New Phytologist, 222, 1123-1138.
DOI PMID |
| [30] | Fleischmann A (2018). Systematics and Evolution of Lentibulariaceae: II. Genlisea. Oxford University Press, Oxford. |
| [31] | Fulton M, Hodges SA (1999). Floral isolation between Aquilegia formosa and Aquilegia pubescens. Proceedings of the Royal Society of London. Series B: Biological Sciences, 266, 2247-2252. |
| [32] |
Graham SA (1990). New species of Cuphea section Melvilla (Lythraceae) and an annotated key to the section. Brittonia, 42, 12-32.
DOI URL |
| [33] | Grant V, Grant KA (1965). Flower Pollination in the Phlox Family. Columbia University Press, New York. |
| [34] | Grant V, Temeles EJ (1992). Foraging ability of rufous hummingbirds on hummingbird flowers and hawkmoth flowers. Proceedings of the National Academy of Sciences of the United States of America, 89, 9400-9404. |
| [35] |
Guo MY, Pang XH, Xu YQ, Jiang WJ, Liao BS, Yu JS, Xu J, Song JY, Chen SL (2022). Plastid genome data provide new insights into the phylogeny and evolution of the genus Epimedium. Journal of Advanced Research, 36, 175-185.
DOI URL |
| [36] |
Hagen KB, Kadereit JW (2003). The diversification of Halenia (Gentianaceae): ecological opportunity versus key innovation. Evolution, 57, 2507-2518.
PMID |
| [37] | Hilton LD, Harder SC (2006). Ecology and Evolution of Flowers. Oxford University Press, Oxford. |
| [38] |
Hobbhahn N, Johnson SD, Bytebier B, Yeung LD, Harder LD (2013). The evolution of floral nectaries in Disa (Orchidaceae: Disinae): recapitulation or diversifying innovation? Annals of Botany, 112, 1303-1319.
DOI PMID |
| [39] |
Hodges SA (1995). The influence of nectar production on hawkmoth behavior, self pollination, and seed production in Mirabilis multiflora (Nyctaginaceae). American Journal of Botany, 82, 197-204.
DOI URL |
| [40] |
Hodges SA (1997). Floral nectar spurs and diversification. International Journal of Plant Sciences, 158, S81-S88.
DOI URL |
| [41] | Hodges SA, Arnold ML (1995). Spurring plant diversification: Are floral nectar spurs a key innovation? Proceedings of the Royal Society of London. Series B: Biological Sciences, 262, 343-348. |
| [42] |
Hodges SA, Fulton M, Yang JY, Whittall JB (2004). Verne Grant and evolutionary studies of Aquilegia. New Phytologist, 161, 113-120.
DOI URL |
| [43] |
Holmgren NH (1992). Two new species of Viola (Violaceae) from the Intermountain West, USA. Brittonia, 44, 300-305.
DOI URL |
| [44] |
Hořák D, Janeček Š (2021). A geographical perspective on the relationship between Impatiens spur lengths and bill lengths of sunbirds in Afrotropical Mountains. Ecology and Evolution, 11, 3120-3129.
DOI URL |
| [45] |
Inouye DW (1980). The terminology of floral larceny. Ecology, 61, 1251-1253.
DOI URL |
| [46] |
Irwin RE, Brody AK (1998). Nectar robbing in Ipomopsis aggregata: effects on pollinator behavior and plant fitness. Oecologia, 116, 519-527.
DOI URL |
| [47] |
Irwin RE, Bronstein JL, Manson JS, Richardson L (2010). Nectar robbing: ecological and evolutionary perspectives. Annual Review of Ecology, Evolution, and Systematics, 41, 271-292.
DOI URL |
| [48] |
Irwin RE, Maloof JE (2002). Variation in nectar robbing over time, space, and species. Oecologia, 133, 525-533.
DOI PMID |
| [49] |
Jabbour F, Renner SS (2012). Spurs in a spur: perianth evolution in the Delphinieae (Ranunculaceae). International Journal of Plant Sciences, 173, 1036-1054.
DOI URL |
| [50] |
Janeček Š, Bartoš M, Njabo KY (2015). Convergent evolution of sunbird pollination systems of Impatiens species in tropical Africa and hummingbird systems of the New World. Biological Journal of the Linnean Society, 115, 127-133.
DOI URL |
| [51] |
Janssens SB, Smets EF, Vrijdaghs A (2012). Floral development of Hydrocera and Impatiens reveals evolutionary trends in the most early diverged lineages of the Balsaminaceae. Annals of Botany, 109, 1285-1296.
DOI PMID |
| [52] |
Johnson SD (1996). Bird pollination in South African species of Satyrium (Orchidaceae). Plant Systematics and Evolution, 203, 91-98.
DOI URL |
| [53] | Johnson SD, Anderson B (2010). Coevolution between food-rewarding flowers and their pollinators. Evolution, 3, 32-39. |
| [54] |
Johnson SD, Ellis A, Dötterl S (2007). Specialization for pollination by beetles and wasps: the role of lollipop hairs and fragrance in Satyrium microrrhynchum (Orchidaceae). American Journal of Botany, 94, 47-55.
DOI PMID |
| [55] |
Johnson SD, Peter CI, Ellis AG, Boberg E, Botes C, van der Niet T (2011). Diverse pollination systems of the twin-spurred orchid genus Satyrium in African grasslands. Plant Systematics and Evolution, 292, 95-103.
DOI URL |
| [56] |
Johnson SD, Steiner KE (1997). Long-tongued fly pollination and evolution of floral spur length in the Disa draconis complex (Orchidaceae). Evolution, 51, 45-53.
DOI PMID |
| [57] | Kawasaki ML (1992). Systematics of Erisma (Vochysiaceae). Wiley Press, Hoboken, USA. |
| [58] |
Kil JH, Park SH, Kim YH, Lee DB (2009). Unrecorded and introduced taxon in Korea: Cymbalaria muralis P. Gaetrn. (Scrophulariaceae). Korean Journal of Plant Taxonomy, 39, 120-123.
DOI URL |
| [59] |
Koehler S, Williams NH, Whitten WM, do Carmo E do Amaral M (2002). Phylogeny of the Bifrenaria (Orchidaceae) complex based on morphology and sequence data from nuclear rDNA internal transcribed spacers (ITS) and chloroplast trnL-trnF region. International Journal of Plant Sciences, 163, 1055-1066.
DOI URL |
| [60] |
Koopman MM, Ayers TJ (2005). Nectar spur evolution in the Mexican lobelias (Campanulaceae: Lobelioideae). American Journal of Botany, 92, 558-562.
DOI URL |
| [61] | Kowalkowska A, Margońska H, Kozieradzka-Kiszkurno M (2010). Comparative anatomy of the lip spur and additional lateral sepal spurs in a three-spurred form (f. fumeauxiana) of Anacamptis pyramidalis. Acta Biologica Cracoviensia Series Botanica, 52, 13-18. |
| [62] |
Kowalkowska AK, Margońska HB, Kozieradzka-Kiszkurno M, Bohdanowicz J (2012). Studies on the ultrastructure of a three-spurred fumeauxiana form of Anacamptis pyramidalis. Plant Systematics and Evolution, 298, 1025-1035.
DOI URL |
| [63] | Krishna S, Keasar T (2018). Morphological complexity as a floral signal: from perception by insect pollinators to co-evolutionary implications. International Journal of Molecular Sciences, 19, 1681. DOI: 10.3390/ijms19061681. |
| [64] | Kurzweil H, Manning JC (2005). A synopsis of the genus Disperis Sw. (Orchidaceae). Adansonia, 27, 155-207. |
| [65] | Li DF, Yan XC, Lin Y, Wang L, Wang Q (2021). Do flowers removed of either nectar or pollen attract fewer bumblebee pollinators? An experimental test in Impatiens oxyanthera. AoB Plants, 13, plab029. DOI: 10.1093/aobpla/plab029. |
| [66] |
Li MH, Zhang GQ, Lan SR, Liu ZJ, Consortium CP (2016). A molecular phylogeny of Chinese orchids. Journal of Systematics and Evolution, 54, 349-362.
DOI URL |
| [67] |
Li YX, Quan QM, Sun GL (2009). Effect of floral morphology on fruit set in Epimedium sagittatum (Berberidaceae). Plant Systematics and Evolution, 279, 51-58.
DOI URL |
| [68] |
Louis P, Decraene R, Smets EF (2001). Floral developmental evidence for the systematic relationships of Tropaeolum (Tropaeolaceae). Annals of Botany, 88, 879-892.
DOI URL |
| [69] |
Lustofin K, Świątek P, Miranda VFO, Płachno BJ (2020). Flower nectar trichome structure of carnivorous plants from the genus butterworts Pinguicula L. (Lentibulariaceae). Protoplasma, 257, 245-259.
DOI |
| [70] | Ma WL (2018). Fine Structure and Morphology of Plants in China. Higher Education Press, Beijing. |
| [马炜梁 (2018). 中国植物精细解剖. 高等教育出版社, 北京.] | |
| [71] |
Macior LW (1970). The pollination ecology of Dicentra cucullaria. American Journal of Botany, 57, 6-11.
DOI URL |
| [72] |
Macior LW (1975). The pollination ecology of Delphinium tricorne (Ranunculaceae). American Journal of Botany, 62, 1009-1016.
DOI URL |
| [73] |
Macior LW (1978). Pollination interactions in sympatric Dicentra species. American Journal of Botany, 65, 57-62.
DOI URL |
| [74] |
Mack JLK, Davis AR (2015). The relationship between cell division and elongation during development of the nectar- yielding petal spur in Centranthus ruber (Valerianaceae). Annals of Botany, 115, 641-649.
DOI URL |
| [75] |
Manning JC, Goldblatt P, Forest F (2009). A revision of Fumariaceae (Fumarioideae) in southern Africa, including naturalized taxa. Bothalia, 39, 45-65.
DOI URL |
| [76] |
Miller RB (1978). The pollination ecology of Aquilegia elegantula and A. caerulea (Ranunculaceae) in Colorado. American Journal of Botany, 65, 406-414.
DOI URL |
| [77] |
Miller RB (1981). Hawkmoths and the geographic patterns of floral variation in Aquilegia caerulea. Evolution, 35, 763-774.
DOI URL |
| [78] |
Miller RB, Willard CL (1983). The pollination ecology of Aquilegia micrantha (Ranunculaceae) in Colorado. The Southwestern Naturalist, 28, 157-164.
DOI URL |
| [79] |
Minelli A (2016). Species diversity vs. morphological disparity in the light of evolutionary developmental biology. Annals of Botany, 117, 781-794.
DOI PMID |
| [80] |
Newman DA, Thomson JD (2005). Effects of nectar robbing on nectar dynamics and bumblebee foraging strategies in Linaria vulgaris (Scrophulariaceae). Oikos, 110, 309-320.
DOI URL |
| [81] | Nguyen VC, Averyanov LV, Maisak TV, Nguyen TLT, Nguyen VK, Truong BV (2022). Gastrochilus pankajkumarii, (Aeridinae, Epidendroideae, Orchidaceae), a new lithophytic orchid from southern Vietnam. Taiwania, 67, 35-39. |
| [82] | Orellana MR, Rovira AM, Blanché C, Bosch M (2008). Effects of local abundance on pollination and reproduction in the narrow endemic endangered species Delphinium bolosii (Ranunculaceae). Orsis, 23, 027-046. |
| [83] | Pansarin ER, Pansarin LM, Alves-dos-Santos I (2015). Floral features, pollination biology, and breeding system of Comparettia coccinea (Orchidaceae: Oncidiinae). Flora- Morphology, Distribution, Functional Ecology of Plants, 217, 57-63. |
| [84] |
Paton A (1997). Classification and species of Platostoma and its relationship with Haumaniastrum (Labiatae). Kew Bulletin, 52, 257-292.
DOI URL |
| [85] |
Peter CI, Johnson SD (2009). Pollination by flower chafer beetles in Eulophia ensata and Eulophia welwitschii (Orchidaceae). South African Journal of Botany, 75, 762-770.
DOI URL |
| [86] |
Płachno BJ, Stpiczyńska M, Adamec L, Miranda VFO, Świątek P (2018). Nectar trichome structure of aquatic bladderworts from the section Utricularia (Lentibulariaceae) with observation of flower visitors and pollinators. Protoplasma, 255, 1053-1064.
DOI |
| [87] |
Puzey JR, Gerbode SJ, Hodges SA, Kramer EM, Mahadevan L (2012). Evolution of spur-length diversity in Aquilegia petals is achieved solely through cell-shape anisotropy. Proceedings of the Royal Society B: Biological Sciences, 279, 1640-1645.
DOI URL |
| [88] | Rebocho AB, Southam P, Kennaway JR, Bangham JA, Coen E (2017). Generation of shape complexity through tissue conflict resolution. Elife, 6, e20156. DOI: 10.7554/eLife.20156. |
| [89] |
Robertson JL, Wyatt R (1990). Evidence for pollination ecotypes in the yellow-fringed orchid, Platanthera ciliaris. Evolution, 44, 121-133.
DOI PMID |
| [90] |
Rudall PJ, Manning JC, Goldblatt P (2003). Evolution of floral nectaries in Iridaceae. Annals of the Missouri Botanical Garden, 90, 613-631.
DOI URL |
| [91] | Rust RW (1979). Pollination of Impatiens capensis: pollinators and nectar robbers. Journal of the Kansas Entomological Society, 52, 297-308. |
| [92] | Shivanna KR (2014). Biotic pollination: How plants achieve conflicting demands of attraction and restriction of potential pollinators//Ramawat KG, Merillon JM, Shivanna KR. Reproductive Biology of Plants. CRC Press, Baca Raton. |
| [93] |
Stebbins GL (1970). Adaptive radiation of reproductive characteristics in angiosperms, I: Pollination mechanisms. Annual Review of Ecology and Systematics, 1, 307-326.
DOI URL |
| [94] |
Steiner KE (1996). Chromosome numbers and relationships in tribe Hemimerideae (Scrophulariaceae). Systematic Botany, 21, 63-76.
DOI URL |
| [95] | Steiner KE, Whitehead VB (1988). The association between oil-producing flowers and oil-collecting bees in the Drakensberg of southern Africa. Monogr. Monographs in Systematic Botany from the Missouri Botanical Garden, 25, 259-277. |
| [96] |
Steiner KE, Whitehead VB (1990). Pollinator adaptation to oil-secreting flowers—Rediviva and Diascia. Evolution, 44, 1701-1707.
DOI PMID |
| [97] |
Stpiczyńska M, Matusiewicz J (2001). Anatomy and ultrastructure of spur nectary of Gymnadenia conopsea (L.) Orchidaceae. Acta Societatis Botanicorum Poloniae, 70, 267-272.
DOI URL |
| [98] | Suddee S, Paton A, Parnell J, Puudjaa P, Keiwbang W, Rueangruea S (2019). Five new species of Platostoma (Lamiaceae) from North-Eastern Thailand. Thai Forest Bulletin (Botany), 47, 226-240. |
| [99] |
Suetsugu K, Fukushima S (2014). Bee pollination of the endangered orchid Calanthe discolor through a generalized food-deceptive system. Plant Systematics and Evolution, 300, 453-459.
DOI URL |
| [100] |
Suetsugu K, Naito RS, Fukushima S, Kawakita A, Kato M (2015). Pollination system and the effect of inflorescence size on fruit set in the deceptive orchid Cephalanthera falcata. Journal of Plant Research, 128, 585-594.
DOI PMID |
| [101] | Sutton DA (1988). A Revision of the Tribe Antirrhineae. Oxford University Press, Oxford. |
| [102] |
Svensson MG, Rydell J, Töve J (1998). Deep flowers for long tongues. Trends in Ecology & Evolution, 13, 259-260.
DOI URL |
| [103] |
Szlachetko DL (1995). Eltroplectris brachycentron Szlachetko (Orchidaceae), a new orchid species from Bolivia. Novon, 5, 375-378.
DOI URL |
| [104] | Teoh ES (2022). Orchid Species from Himalaya and Southeast Asia Vol. 3 (R-Z). Springer Press, Berlin. |
| [105] |
The Angiosperm Phylogeny Group (2016). An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Botanical Journal of the Linnean Society, 181, 1-20.
DOI URL |
| [106] |
Thompson JN (1999). Specific hypotheses on the geographic mosaic of coevolution. The American Naturalist, 153, S1-S14.
DOI URL |
| [107] |
Travers SE, Temeles EJ, Pan I (2003). The relationship between nectar spur curvature in jewelweed (Impatiens capensis) and pollen removal by hummingbird pollinators. Canadian Journal of Botany, 81, 164-170.
DOI URL |
| [108] |
van der Niet, Jürgens A, Johnson SD (2010). Pollinators, floral morphology and scent chemistry in the southern African orchid genus Schizochilus. South African Journal of Botany, 76, 726-738.
DOI URL |
| [109] |
Vandelook F, Janssens SB, Gijbels P, Fischer E, van den Ende W, Honnay O, Abrahamczyk S (2019). Nectar traits differ between pollination syndromes in Balsaminaceae. Annals of Botany, 124, 269-279.
DOI PMID |
| [110] |
Vargas P, Liberal I, Ornosa C, Gómez JM (2017). Flower specialisation: the occluded corolla of snapdragons (Antirrhinum) exhibits two pollinator niches of large long-tongued bees. Plant Biology, 19, 787-797.
DOI PMID |
| [111] | Venter H, Johannes T (1979). A Monograph of Monsonia L. (Geraniaceae). Springer Press, Berlin. |
| [112] |
Vogel S (1984). The Diascia flower and its bee—An oil-based symbiosis in southern Africa. Acta Botanica Neerlandica, 33, 509-518.
DOI URL |
| [113] |
Wang Q, Li YX, Pu XZ, Zhu LY, Tang Z, Liu Q (2013). Pollinators and nectar robbers cause directional selection for large spur circle in Impatiens oxyanthera (Balsaminaceae). Plant Systematics and Evolution, 299, 1263-1274.
DOI URL |
| [114] | Wang XY, Tang XX, Tong ZY, Huang SQ (2019). Geographic mosaic theory of coevolution in plant and pollinator interaction: process, evidence, prospect. Chinese Science Bulletin, 64, 1702-1710. |
| [王晓月, 汤晓辛, 童泽宇, 黄双全 (2019). 植物与传粉者地理镶嵌的协同演化: 过程、证据与展望. 科学通报, 64, 1702-1710.] | |
| [115] |
Waser NM (1988). Comparative pollen and dye transfer by pollinators of Delphinium nelsonii. Functional Ecology, 2, 41-48.
DOI URL |
| [116] |
Wasserthal LT (1997). The pollinators of the Malagasy star orchids Angraecum sesquipedale, A. sororium and A. compactum and the evolution of extremely long spurs by pollinator shift. Botanica Acta, 110, 343-359.
DOI URL |
| [117] |
Watthana S (2007). The genus Pomatocalpa (Orchidaceae) a taxonomic monograph. Harvard Papers in Botany, 11, 207-256.
DOI URL |
| [118] |
Weryszko-Chmielewska E, Bożek M (2008). Structure of trichomatous nectaries in flowers of Lonicera kamtschatica (Sevast.) Pojark. Acta Agrobotanica, 61, 13-26.
DOI URL |
| [119] |
Whittall JB, Hodges SA (2007). Pollinator shifts drive increasingly long nectar spurs in columbine flowers. Nature, 447, 706-709.
DOI |
| [120] | Xie SY, Hou XQ, Zhang XH (2022). Are the spurs more complex than other petal types in Epimedium? Evidence from development, micromorphology, and nectary structure. Flora, 293, 152101. DOI: 10.1016/j.flora.2022.152101. |
| [121] | Yang ML, Wang LL, Zhang GP, Meng LH, Yang YP, Duan YW (2018). Equipped for migrations across high latitude regions? Reduced spur length and outcrossing rate in a biennial Halenia elliptica (Gentianaceae) with mixed mating system along a latitude gradient. Frontiers in Genetics, 9, 223. DOI: 10.3389/fgene.2018.00223. |
| [122] |
Young HJ (2008). Selection on spur shape in Impatiens capensis. Oecologia, 156, 535-543.
DOI URL |
| [123] |
Yousefi N, Zarre S, Heubl G (2016). Molecular phylogeny of the mainly Mediterranean genera Chaenorhinum, Kickxia and Nanorrhinum (Plantaginaceae, tribe Antirrhineae), with focus on taxa in the Flora Iranica region. Nordic Journal of Botany, 34, 455-463.
DOI URL |
| [124] |
Zhang R, Min Y, Holappa LD, Walcher-Chevillet CL, Duan XS, Donaldson E, Kong HZ, Kramer EM (2020). A role for the Auxin Response Factors ARF6 and ARF8 homologs in petal spur elongation and nectary maturation in Aquilegia. New Phytologist, 227, 1392-1405.
DOI PMID |
| [125] |
Zhang YW, Wang Y, Guo YH (2006). The effects of nectar robbing on plant reproduction and evolution. Chinese Journal of Plant Ecology (Chinese Version), 30, 695-702.
DOI URL |
|
[张彦文, 王勇, 郭友好 (2006). 盗蜜行为在植物繁殖生态学中的意义. 植物生态学报, 30, 695-702.]
DOI |
|
| [126] |
Zhao L, Gong JZ, Zhang XH, Liu TQ, Ma X, Ren Y (2016). Floral organogenesis in Urophysa rockii, a rediscovered endangered and rare species of Ranunculaceae. Botany, 94, 215-224.
DOI URL |
| [127] |
Zou X, Fountain DW, Morgan ER (2001). Anatomical and morphological studies of seed development in Sandersonia aurantiaca (Hook.). South African Journal of Botany, 67, 183-192.
DOI URL |
| [1] | 张彦文, 王勇, 郭友好. 盗蜜行为在植物繁殖生态学中的意义[J]. 植物生态学报, 2006, 30(4): 695-702. |
| [2] | 邓晓保, 任盘宇, 李庆军. 云南草蔻花蜜分泌格局与访花动物行为及其对果实和种子产量的影响[J]. 植物生态学报, 2005, 29(2): 274-280. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2026 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19
![]()