
长期氮添加对典型草原植物多样性与初级生产力的影响及途径
收稿日期: 2019-10-01
修回日期: 2020-01-09
网络出版日期: 2020-02-24
基金资助
国家重点研发计划(2016YFC0500801);国家自然科学基金(41671046);国家自然科学基金(31400393);国家自然科学基金(31600365)
Effects and pathways of long-term nitrogen addition on plant diversity and primary productivity in a typical steppe
Received date: 2019-10-01
Revised date: 2020-01-09
Online published: 2020-02-24
Supported by
National Key R&D Program of China(2016YFC0500801);National Natural Science Foundation of China(41671046);National Natural Science Foundation of China(31400393);National Natural Science Foundation of China(31600365)
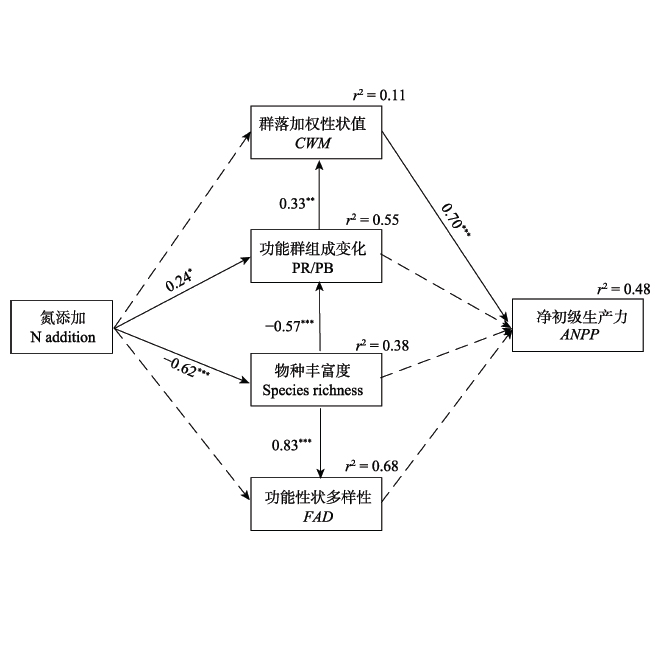
氮(N)沉降对陆地生态系统的结构和功能已产生了重要的影响, N也是中国北方草原植物生长和初级生产力的主要限制性元素。物种多样性和功能多样性是揭示生物多样性对生态系统功能维持机制的关键指标, 然而, 关于长期N添加下草原物种多样性与功能多样性的关系, 及其对初级生产力的影响途径及机制, 尚不十分清楚。为此, 该研究依托在内蒙古典型草原建立的长期N添加实验平台, 实验处理包括1个完全对照(不添加任何肥料)和6个N添加水平(0、1.75、5.25、10.50、17.50和28.00 g·m-2·a-1), 研究了长期N添加对典型草原物种多样性、功能多样性和初级生产力的影响大小及途径。结果表明: 1) N添加显著降低了典型草原的物种丰富度和Shannon-Wiener指数, 但对功能多样性(包括功能性状多样性指数和群落加权性状值)无显著的影响。2)结构方程模型分析表明, 功能多样性主要受物种丰富度的影响, 但是物种多样性减少并没有导致功能多样性降低, 其原因主要是功能群组成发生了改变, 即群落内多年生根茎禾草所占比例显著增加, 以致群落加权性状值变化不大。3) N通过影响物种丰富度和功能群组成, 间接影响群落加权性状值, 进而影响群落净初级生产力。其中, 群落加权性状值是最重要的影响因子, 可解释48%的初级生产力变化, 表明初级生产力主要是由群落内优势物种的生物量及功能性状所决定, 因此该研究的结果很好地支持了质量比假说。
王玉冰,孙毅寒,丁威,张恩涛,李文怀,迟永刚,郑淑霞 . 长期氮添加对典型草原植物多样性与初级生产力的影响及途径[J]. 植物生态学报, 2020 , 44(1) : 22 -32 . DOI: 10.17521/cjpe.2019.0260
Aims Nitrogen (N) deposition has made great impacts on the structure and function of terrestrial ecosystems in recent decades, and N is known as the main limiting element of plant growth and primary productivity in the northern grassland of China. Species diversity and functional diversity have been widely recognized as key indicators of the mechanisms of biodiversity maintenance for ecosystem functions. However, the effect of long-term N addition on the relationship between species diversity and functional diversity, and the corresponding impacts on primary productivity have rarely been studied; particularly the underpinning mechanisms remain unclear. Our objective is to examine the effects and pathways of long-term N addition on species diversity, functional diversity and primary productivity in a typical steppe.
Methods A manipulative N addition experiment located in a typical steppe of the Nei Mongol grassland has been conducted for 18 years. The experimental design included seven levels of N addition rate (i.e., control, 0, 1.75, 5.25, 10.50, 17.50, 28.00 g·m-2·a-1) with nine replicates for each treatment. The plant functional traits of dominant species, species richness and composition, and aboveground net primary productivity (ANPP) were determined. In addition, species diversity, functional attribute diversity and community-weighted mean traits were calculated.
Important findings 1) N addition significantly reduced species richness and Shannon-Wiener index, but had little effect on functional diversity, including functional attribute diversity and community-weighted mean traits. 2) Structural equation model analyses showed that functional diversity was mainly affected by species richness, whereas the decrease in species diversity didn’t lead to the decrease in functional diversity. The community- weighted mean traits did not change under N addition, which was mainly due to the shift in functional group composition, that is, the proportion of perennial rhizome grass in plant community increased greatly under N addition. 3) The effect of N addition on ANPP was through direct pathways of species richness and shift in functional group composition, and further via an indirect pathway of community-weighted mean traits. Particularly, the community-weighted mean traits were the most important factors and explained 48% of total variation in ANPP, implying that the primary productivity is mainly determined by the biomass and functional traits of dominant species in the plant community. Our results well supported the mass-ratio hypothesis.
| [1] | Bai YF, Wu JG, Clark CM, Naeem S, Pan QM, Huang JH, Zhang LX, Han XG (2010). Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: Evidence from Inner Mongolia grasslands. Global Change Biology, 16, 358-372. |
| [2] | Bennett JA, Riibak K, Tamme R, Lewis RJ, P?rtel M (2016). The reciprocal relationship between competition and intraspecific trait variation. Journal of Ecology, 104, 1410-1420. |
| [3] | Bernhardt-R?mermann M, R?mermann C, Pillar VD, Kudernatsch T, Fischer A (2010). High functional diversity is related to high nitrogen availability in a deciduous forest?—?Evidence from a functional trait approach. Folia Geobotanica, 45, 111-124. |
| [4] | Brym ZT, Lake JK, Allen D, Ostling A (2011). Plant functional traits suggest novel ecological strategy for an invasive shrub in an understorey woody plant community. Journal of Applied Ecology, 48, 1098-1106. |
| [5] | Cadotte MW (2017). Functional traits explain ecosystem function through opposing mechanisms. Ecology Letters, 20, 989-996. |
| [6] | Cardinale BJ, Wright JP, Cadotte MW, Carroll IT, Hector A, Srivastava DS, Loreau M, Weis JJ (2007). Impacts of plant diversity on biomass production increase through time because of species complementarity. Proceedings of the National Academy of Sciences of the United States of America, 104, 18123-18128. |
| [7] | Chen DM, Xing W, Lan ZC, Saleem M, Wu Y, Hu SJ, Bai YF (2019). Direct and indirect effects of nitrogen enrichment on soil organisms and carbon and nitrogen mineralization in a semi-arid grassland. Functional Ecology, 33, 175-187. |
| [8] | Clark CM, Tilman D (2008). Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands. Nature, 451, 712-715. |
| [9] | de Bello F, Lep? J, Sebastià MT (2006). Variations in species and functional plant diversity along climatic and grazing gradients. Ecography, 29, 801-810. |
| [10] | Duprè C, Stevens CJ, Ranke T, Bleeker A, Peppler-Lisbach C, Gowing DJG, Dise NB, Dorland E, Bobbink R, Diekmann M (2010). Changes in species richness and composition in European acidic grasslands over the past 70 years: The contribution of cumulative atmospheric nitrogen deposition. Global Change Biology, 16, 344-357. |
| [11] | Finegan B, Pe?a-Claros M, de Oliveira A, Ascarrunz N, Bret-Harte MS, Carre?o-Rocabado G, Casanoves F, Díaz S, Eguiguren Velepucha P, Fernandez F, Licona JC, Lorenzo L, Salgado Negret B, Vaz M, Poorter L (2015). Does functional trait diversity predict above-ground biomass and productivity of tropical forests? Testing three alternative hypotheses. Journal of Ecology, 103, 191-201. |
| [12] | Gagic V, Bartomeus I, Jonsson T, Taylor A, Winqvist C, Fischer C, Slade EM, Steffan-Dewenter I, Emmerson M, Potts SG, Tscharntke T, Weisser W, Bommarco R (2015). Functional identity and diversity of animals predict ecosystem functioning better than species-based indices. Proceedings of the Royal Society B, 282, 20142620. DOI: 10.1098/rspb.2014.2620. |
| [13] | Grime JP (1998). Benefits of plant diversity to ecosystems: Immediate, filter and founder effects. Journal of Ecology, 86, 902-910. |
| [14] | Gross N, Kunstler G, Liancourt P, de Bello F, Suding KN, Lavorel S (2009). Linking individual response to biotic interactions with community structure: A trait-based framework. Functional Ecology, 23, 1167-1178. |
| [15] | Hooper DU, Chapin III FS, Ewel JJ, Hector A, Inchausti P, Lavorel S, Lawton JH, Lodge DM, Loreau M, Naeem S, Schmid B, Set?l? H, Symstad AJ, Vandermeer J, Wardle DA (2005). Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecological Monographs, 75, 3-35. |
| [16] | Hooper DU, Vitousek PM (1998). Effects of plant composition and diversity on nutrient cycling. Ecological Monographs, 68, 121-149. |
| [17] | Kardol P, Wardle DA (2010). How understanding aboveground-?belowground linkages can assist restoration ecology. Trends in Ecology & Evolution, 25, 670-679. |
| [18] | Knops JM, Reinhart K (2000). Specific leaf area along a nitrogen fertilization gradient. The American Midland Naturalist, 144, 265-272. |
| [19] | Kraft NJB, Godoy O, Levine JM (2015). Plant functional traits and the multidimensional nature of species coexistence. Proceedings of the National Academy of Sciences of the United States of America, 112, 797-802. |
| [20] | Kunstler G, Falster D, Coomes DA, Hui F, Kooyman RM, Laughlin DC, Poorter L, Vanderwel M, Vieilledent G, Wright SJ, Aiba M, Baraloto C, Caspersen J, Cornelissen JHC, Gourlet-Fleury S, Hanewinkel M, Herault B, Kattge J, Kurokawa H, Onoda Y, Pe?uelas J, Poorter H, Uriarte M, Richardson S, Ruiz-Benito P, Sun IF, St?hl G, Swenson NG, Thompson J, Westerlund B, Wirth C, Zavala MA, Zeng HC, Zimmerman JK, Zimmermann NE, Westoby M (2016). Plant functional traits have globally consistent effects on competition. Nature, 529, 204-207. |
| [21] | Lan ZC, Bai YF (2012). Testing mechanisms of N-enrichment-?induced species loss in a semiarid Inner Mongolia grassland: Critical thresholds and implications for long-term ecosystem responses. Philosophical Transactions of the Royal Society B-Biological Sciences, 367, 3125-3134. |
| [22] | Lan ZC, Jenerette GD, Zhan SX, Li WH, Zheng SX, Bai YF (2015). Testing the scaling effects and mechanisms of N-induced biodiversity loss: Evidence from a decade-long grassland experiment. Journal of Ecology, 103, 750-760. |
| [23] | Laughlin DC (2014). Applying trait-based models to achieve functional targets for theory-driven ecological restoration. Ecology Letters, 17, 771-784. |
| [24] | Lavorel S, Grigulis K, McIntyre S, Williams NSG, Garden D, Dorrough J, Berman S, Quétier F, Thébault A, Bonis A (2008). Assessing functional diversity in the field—?Methodology matters! Functional Ecology, 22, 134-147. |
| [25] | Li W, Cheng JM, Yu KL, Epstein HE, Guo L, Jing GH, Zhao J, Du GZ (2015). Plant functional diversity can be independent of species diversity: Observations based on the impact of 4-yrs of nitrogen and phosphorus additions in an alpine meadow. PLOS ONE, 10, e0136040. DOI: 10.1371/?journal.pone.0136040. |
| [26] | Li XG, Zhu ZH, Zhou XS, Yuan FR, Fan RJ, Xu ML (2011). Effects of clipping, fertilizing and watering on the relationship between species diversity, functional diversity and primary productivity in alpine meadow of China. Chinese Journal of Plant Ecology, 35, 1136-1147. |
| [26] | [ 李晓刚, 朱志红, 周晓松, 袁芙蓉, 樊瑞俭, 许曼丽 (2011). 刈割、施肥和浇水对高寒草甸物种多样性、功能多样性与初级生产力关系的影响. 植物生态学报, 35, 1136-1147.] |
| [27] | Maire V, Gross N, B?rger L, Proulx R, Wirth C, da Silveira Pontes L, Soussana JF, Louault F (2012). Habitat filtering and niche differentiation jointly explain species relative abundance within grassland communities along fertility and disturbance gradients. New Phytologist, 196, 497-509. |
| [28] | Mokany K, Ash J, Roxburgh S (2008). Functional identity is more important than diversity in influencing ecosystem processes in a temperate native grassland. Journal of Ecology, 96, 884-893. |
| [29] | Mouchet MA, Villéger S, Mason NWH, Mouillot D (2010). Functional diversity measures: An overview of their redundancy and their ability to discriminate community assembly rules. Functional Ecology, 24, 867-876. |
| [30] | Naeem S, Wright JP (2003). Disentangling biodiversity effects on ecosystem functioning: Deriving solutions to a seemingly insurmountable problem. Ecology Letters, 6, 567-579. |
| [31] | Orwin KH, Buckland SM, Johnson D, Turner BL, Smart S, Oakley S, Bardgett RD (2010). Linkages of plant traits to soil properties and the functioning of temperate grassland. Journal of Ecology, 98, 1074-1083. |
| [32] | Pan QM, Bai YF, Wu JG, Han XG (2011). Hierarchical plant responses and diversity loss after nitrogen addition: Testing three functionally-based hypotheses in the Inner Mongolia grassland. PLOS ONE, 6, e20078. DOI: 10.1371/?journal.pone.0020078. |
| [33] | Pardo LH, Fenn ME, Goodale CL, Geiser LH, Driscoll CT, Allen EB, Baron JS, Bobbink R, Bowman WD, Clark CM, Emmett B, Gilliam FS, Greaver TL, Hall SJ, Lilleskov EA, Liu LL, Lynch JA, Nadelhoffer KJ, Perakis SS, Robin-Abbott MJ, Stoddard JL, Weathers KC, Dennis RL (2011). Effects of nitrogen deposition and empirical nitrogen critical loads for ecoregions of the United States. Ecological Applications, 21, 3049-3082. |
| [34] | Payne RJ, Dise NB, Field CD, Dore AJ, Caporn SJ, Stevens CJ (2017). Nitrogen deposition and plant biodiversity: Past, present, and future. Frontiers in Ecology and the Environment, 15, 431-436. |
| [35] | Pérez-Ramos IM, Díaz-Delgado R, de la Riva EG, Villar R, Lloret F, Mara?ón T (2017). Climate variability and community stability in Mediterranean shrublands: The role of functional diversity and soil environment. Journal of Ecology, 105, 1335-1346. |
| [36] | Petchey OL, Gaston KJ (2002). Functional diversity (FD), species richness and community composition. Ecology Letters, 5, 402-411. |
| [37] | Prado-Junior JA, Schiavini I, Vale VS, Arantes CS, van der Sande MT, Lohbeck M, Poorter L (2016). Conservative species drive biomass productivity in tropical dry forests. Journal of Ecology, 104, 817-827. |
| [38] | Spasojevic MJ, Suding KN (2012). Inferring community assembly mechanisms from functional diversity patterns: The importance of multiple assembly processes. Journal of Ecology, 100, 652-661. |
| [39] | Suding KN, Collins SL, Gough L, Clark C, Cleland EE, Gross KL, Milchunas DG, Pennings S (2005). Functional- and abundance-based mechanisms explain diversity loss due to N fertilization. Proceedings of the National Academy of Sciences of the United States of America, 102, 4387-4392. |
| [40] | Tylianakis JM, Didham RK, Bascompte J, Wardle DA (2008). Global change and species interactions in terrestrial ecosystems. Ecology Letters, 11, 1351-1363. |
| [41] | Valencia E, Maestre FT, Le Bagousse-Pinguet Y, Quero JL, Tamme R, B?rger L, García-Gómez M, Gross N (2015). Functional diversity enhances the resistance of ecosystem multifunctionality to aridity in Mediterranean drylands. New Phytologist, 206, 660-671. |
| [42] | Violle C, Navas ML, Vile D, Kazakou E, Fortunel C, Hummel I, Garnier E (2007). Let the concept of trait be functional! Oikos, 116, 882-892. |
| [43] | Walker B, Kinzig A, Langridge J (1999). Original articles: Plant attribute diversity, resilience, and ecosystem function: The nature and significance of dominant and minor species. Ecosystems, 2, 95-113. |
| [44] | Wan HW, Yang Y, Bai SQ, Xu YH, Bai YF (2008). Variations in leaf functional traits of six species along a nitrogen addition gradient in Leymus chinensis steppe in Inner Mongolia. Journal of Plant Ecology (Chinese version), 32, 611-621. |
| [44] | [ 万宏伟, 杨阳, 白世勤, 徐云虎, 白永飞 (2008). 羊草草原群落6种植物叶片功能特性对氮素添加的响应. 植物生态学报, 32, 611-621.] |
| [45] | Xu ZW, Li MH, Zimmermann NE, Li SP, Li H, Ren HY, Sun H, Han XG, Jiang Y, Jiang L (2018). Plant functional diversity modulates global environmental change effects on grassland productivity. Journal of Ecology, 106, 1941-1951. |
| [46] | Yang HJ, Li Y, Wu MY, Zhang Z, Li LH, Wan SQ (2011). Plant community responses to nitrogen addition and increased precipitation: The importance of water availability and species traits. Global Change Biology, 17, 2936-2944. |
/
| 〈 |
|
〉 |
