青藏高原及周边高山地区的植物繁殖生态学研究进展
张婵, 安宇梦, Yun JÄSCHKE, 王林林, 周知里, 王力平, 杨永平, 段元文
植物生态学报
2020, 44 ( 1):
1-21.
DOI: 10.17521/cjpe.2019.0296
青藏高原及周边高山地区孕育了极为丰富的植物多样性资源, 研究该地区植物如何顺利完成繁殖过程有助于我们理解植物对典型高山环境的进化和适应机制。该文综述了青藏高原地区高山植物在资源分配、繁殖方式、花部特征演化等方面的研究进展, 包括全球气候变化对植物繁殖特征的影响, 以及一些新技术和新方法在本研究领域的应用。在高山地区限制性环境中, 随海拔升高, 繁殖分配通常表现出增大的趋势, 其中投入到雄性资源的比例上升, 但具体的资源分配模式还要取决于植株的交配系统、个体大小、生活史特征、遗传特性以及环境中的资源有效性等。面对资源和传粉的双重限制, 植物在不同繁殖方式之间存在权衡, 当传粉者稀少时, 克隆繁殖和自交有利于繁殖保障; 而有性繁殖和异交能够提高种子的质量和后代的遗传多样性, 从而在复杂多变的气候条件下有利于种群的维持。因此, 不同繁殖方式的结合以及泛化的传粉互作网络可能是应对高山限制性环境的最优选择。花部特征的演化主要受到当地传粉者的选择压力, 但是外来传粉者、植食者、盗蜜者以及非生物环境(如温度、雨水和紫外辐射等)对花部性状的影响越来越受到重视。近年来, 青藏高原因其脆弱性和对气候变化的高度敏感性而在全球气候变化研究中备受关注, 以全球变暖和氮沉降增加为显著特征的全球气候变化正在直接或间接地影响着该地区高山植物的繁殖特征。气候变化影响植物和传粉者的物候并引起物种的迁移, 最终将导致植物与传粉者的时空不匹配。植物通过改变花部特征(花展示、花冠结构、花报酬的数量和质量)来响应气候变化, 这可能会改变其传粉者的类型、数量和访花行为, 从而最终影响植物的繁殖成功。3D打印和高通量测序等新技术和新方法的应用有助于促进植物繁殖生态学研究的进一步发展。3D打印的花能够精确控制其形态构造, 可以用于研究精细的花部特征变化对于传粉者行为的影响, 在此基础上与人工饲养的传粉者结合使用, 有助于进一步研究传粉者介导的花部特征演化。随着高通量测序技术的发展, 植物繁殖生态学领域, 尤其是花部特征演化的许多重要问题的潜在机制得以深入研究。该文最后提出了目前研究中需要注意的问题以及值得深入研究的发展方向。
View image in article
图2
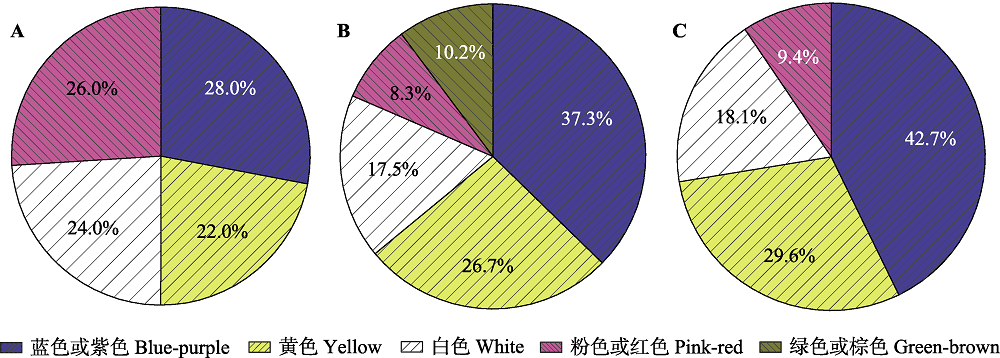
不同地区植物群落的花色比例。A, 尼泊尔境内喜马拉雅山区亚热带地区(海拔900-2 000 m)和亚高山地区(海拔3 000-4 100 m) 107种植物(Shrestha et al., 2014)。B, 中国西南横断山区(平均海拔> 4 300 m) 932种植物(Peng et al., 2014)。C, 中国西南横断山区(平均海拔> 4 300 m) 823种植物(Peng et al., 2012)。
正文中引用本图/表的段落
花部特征演化的最有效传粉者原则(most effective pollinator principle)认为在花部性状的进化过程中, 传粉者介导的选择扮演了重要的作用(黄双全, 2014)。花性状的组合(即形态、颜色、气味、报酬等)和特定的传粉者类群相关, 被称为传粉综合征(pollination syndromes)。在高山极端环境下, 传粉者在花部形态特征演化过程中发挥着极大的驱动作用。在群落水平上对横断山区27种开花植物的传粉者泛化水平(pollinator generalization level)的定量分析结果表明, 与辐射对称的花相比, 两侧对称的花增强了与特异性传粉者的相互作用, 从而增加了花粉落置的精确性(Gong & Huang, 2009)。对青藏高原东部高山草甸50种开花植物的花和叶特征的分析结果显示: 花大小的变异性显著低于叶片大小的变异性, 这表明传粉者对花部特征施加的稳定选择有利于花的稳定性(路宁娜和赵志刚, 2014)。六叶龙胆在高海拔地区雄蕊和雌蕊缩短, 花药与柱头间的距离(雌雄异位)缩小, 这是在传粉者稀少的情况下增加自花授粉能力的一种适应性策略(He at al., 2017)。毛茛科矮金莲花(Trollius farreri)在花粉限制压力更大的高海拔地区通过增大萼片等花展示的投入以吸引传粉者, 而在低海拔区域则增加了对退化花瓣这种产蜜结构的生物量分配, 这表明在高海拔地区花的展示结构对雌性繁殖成功的贡献要大于传粉报酬的贡献(孟金柳, 2010)。花色是一种吸引传粉者的显著视觉信号, 指示传粉者取食和传粉。高山植物群落中花的主导色彩会随着地域和季节的变化而不同, 这可能与传粉者的种类变化及其偏好有关(彭德力等, 2012)。针对尼泊尔境内喜马拉雅山区亚热带地区(海拔900-2 000 m)和亚高山地区(海拔3 000- 4 100 m) 107种植物的花色研究(图2A)发现, 其蓝紫色花的比例显著小于平均海拔4 300 m以上的横断山区(图2B、2C), 这可能与熊蜂作为横断山区主要传粉者有密切关系(Gong & Huang, 2011)。值得注意的是, 除了传粉者的选择压力, 植物的花色还可能受到其他生物因素(如植食者)和非生物因素(如紫外辐射)的影响, 青藏高原及周边高山地区丰富的植物种类和复杂多样的生境类型为检验花色的影响机制提供了有利条件, 但目前此类研究还十分欠缺, 有待今后开展深入广泛的研究。除了丰富的花色, 一些植物还演化出对传粉者具有特殊吸引和指示作用的花图案或花结构, 即“蜜导” (nectar guide), 借此控制访花者的落置方向和位置并进一步提高其访花效率(Leonard & Papaj, 2011; Hansen et al., 2012)。报酬(主要为花蜜)也是植物吸引传粉者的重要因素, 不同传粉者对花蜜的浓度、成分、颜色和分泌动态都表现出不同的偏好(Baker & Baker, 1983)。例如, 喜马拉雅山区分布的唇形科米团花(Leucosceptrum canum)通过花蜜的色彩和动态变化来吸引鸟类传粉者(Zhang et al., 2012); 我国西南山区分布的唇形科野拔子(Elsholtzia rugulosa)花蜜中的酚类化合物起到吸引传粉者同时又排斥非传粉者的双重作用(Zhang et al., 2016)。不同植物的花寿命具有显著差异, 花寿命的可塑性也是植物对传粉环境变化的一种适应对策(Trunschke & Stöcklin, 2017)。对青藏高原高寒草甸不同海拔11种开花植物的花寿命进行对比研究发现, 高海拔群落中植物的花寿命显著延长, 且可塑性更大, 说明植物通过提高花寿命及其可塑性以适应传粉者稀少的高寒环境(王玉贤等, 2018)。
值得注意的是, 蜜蜂养殖业的发展向自然生态系统中释放了大量的人工饲养蜜蜂, 进而对本土植物的花部特征演化产生了一定的选择压力.对青藏高原地区菊科钝苞雪莲(Saussurea nigrescens)的研究发现, 养蜂场附近的植株花蜜分泌量减少了60%以上, 并将结余的资源用于生产更多的花和种子(Mu et al., 2014).外来传粉者可能通过影响植物花部特征的进化而进一步作用于本地传粉者的访花行为和种群动态, 这对当地传粉网络稳定性的影响亟待深入研究.但当本地传粉昆虫缺乏时, 人工饲养的蜜蜂可协助本地植物完成传粉过程(Duan & Liu, 2007). ... Seasonal dynamics of terristrial net primary production in response to climate change in China 1 2003 ... 素有“世界屋脊”和地球“第三极”之称的青藏高原平均海拔超过4 000 m, 面积约2.5 × 106 km2, 是世界上面积最大、海拔最高的高原.本文所指的青藏高原及周边高山地区主要包括青海省和西藏自治区的全部以及四川省、云南省和甘肃省的部分地区, 邻国喜马拉雅高山地区的部分研究也有所涉及.青藏高原气候条件极具特色, 在植物生长季节, 主要受西南季风的控制, 降水及气温由东南向西北逐渐降低(An et al., 2001; Piao et al., 2003; 卢鹤立等, 2007).受地形和水热条件的影响, 青藏高原植被类型呈现出明显的水平和垂直地带性变化特征.整体来看, 青藏高原东南部以森林群落为主, 逐渐向西北过渡为高寒草甸、高寒草原及荒漠.青藏高原部分地区的垂直地带性变化同样非常明显, 随海拔升高由森林植被逐渐过渡为灌丛、草甸、流石滩, 而这一特征主要体现在藏东南海拔跨度较大的高山地区(郑度和陈伟烈, 1981). ... Disentangling visual and olfactory signals in mushroom-mimicking Dracula orchids using realistic three-dimensional printed flowers 1 2016 ... 3D打印技术的出现不仅突破性地解决了上述问题, 而且可以预见该技术在植物繁殖生态学研究领域具有广阔的应用前景.3D打印是以数字化模型为基础, 运用粉末状金属或塑料等可黏合材料, 通过逐层打印的方式直接建立与相应数学模型完全一致的三维物理实体模型(Gebler et al., 2014).近年来, 国外学者已经尝试将其应用于植物繁殖生态学中花部特征的进化研究, 通过打印出各式各样的花部结构来代替植物原结构进行模拟实验(Campos et al., 2015; Policha et al., 2016), 但是国内此类研究还未见报道.3D打印的花不仅仿真程度高, 而且可以根据研究目的的需要控制其形态构造.例如研究视觉信号时, 可以选择具有颜色的无味的原材料, 从而避免了颜料气味对传粉者的影响, 同时还可以构造出各种花部图案, 这在植物“蜜导”作用的研究中将发挥重要作用.研究嗅觉信号时, 可以首先分析花部挥发物的化合物组成, 然后用无味的原材料打印花部构造, 再将单一化合物或者组合化合物添加到3D打印的花上, 相同的方法还可以应用于花报酬(如花蜜量和花蜜成分)的研究.除了在物种水平上研究植物花部性状的选择压力, 3D打印技术还可以应用于群落水平上的繁殖生态学研究, 我们不仅可以模拟自然群落的组成, 还可以根据研究目的人为设置群落内斑块的大小和构成, 以及斑块间的距离等因素, 从群落水平上研究传粉者对花部特征的选择作用(王力平等, 2019). ... Effects of floral symmetry on pollination in Bidens aristosa 1 2015 ... 在以往的繁殖生态学研究中, 为了分析花部特征的演化, 特别是不同花部性状对传粉者的影响, 研究者常采用在野外实验中去除某些花部结构(Totland, 2004; Potts, 2015), 或者进行人工涂色的方法(Zhang et al., 2017).但是这些处理方式都存在一定缺陷, 首先破损后的花部位和颜料可能会释放某些化学物质, 从而影响传粉者的行为; 其次, 受限于这些简单的去除或者涂色方法, 无法满足对一些复杂的花部构造的分析.虽然后来一些研究中应用了纸花和树脂玻璃等人造花, 但是仍然难以模拟自然界复杂的花部特征, 同时可控性也较低. ... 外来引种植物印楝和辣木对红河干热河谷传粉网络的影响 1 2018 ... 青藏高原地区具有极为丰富的动植物多样性资源, 植物与传粉者间经过长期适应进化而形成了复杂的传粉网络, 传粉互作网络在维持生物多样性和生态系统服务功能中扮演着重要角色.目前, 我国在群落水平的传粉研究主要关注传粉网络的时空动态(Fang & Huang, 2012, 2016a; 赵延会, 2016), 外来植物(起国海, 2018; 土艳丽等, 2019)和动植物的系统发育(赵延会, 2016)对传粉网络的影响, 群落水平上的异种花粉转移(Fang & Huang, 2013, 2016b; 杨洁羽, 2018)等方面.连续3年对香格里拉高山草甸29种植物的花部特征与传粉功能群的关系进行研究, 结果表明, 虽然传粉网络结构是嵌套的、不对称的, 传粉者的访问偏好仍支持了传粉者在花部特征中的选择作用(Gong & Huang, 2011).此外, 该高山草甸中的开花植物与传粉者的种类在年际间变动较大, 但植物与其传粉者的连接及其强度基本都是稳定的, 而且熊蜂位于传粉网络的核心位置并对传粉功能起关键作用(Fang & Huang, 2012, 2016a).同地区内异种花粉转移的研究结果表明, 昆虫种间访问比例仅占所有访问次数的4%, 但是柱头上的异种花粉比例高达22%, 泛化植物接受多种其他植物的花粉, 但是不会输出更多的花粉给其他植物, 接受较多种间访问的植物具有较低的异种花粉落置数量, 这是共同开花植物降低异种花粉转移负面影响的一种适应策略(Fang & Huang, 2013, 2016b).在喜马拉雅-横断山地区玉龙雪山上海拔2 700-3 910 m范围内4个群落的传粉网络研究发现, 随着海拔升高, 传粉网络的特化水平显著升高而嵌套结构显著降低, 并且昆虫访花网络与传粉网络的拓扑结构存在显著变化(Zhao et al., 2019).另外, 植物及传粉者最近种间亲缘关系指数随海拔升高显著增加, 表明系统发育结构由离散逐渐转变为聚集, 然而植物及传粉者系统发育信号强度表现出不对称性, 传粉者系统发育关系的影响显著高于植物(赵延会, 2016).过度放牧、生物入侵等都会显著改变动植物的物种丰富度和多样性, 从而改变当地的传粉网络结构, 对生态系统功能产生负面影响(Soares et al., 2017).青藏高原东部高寒草甸的研究结果表明, 放牧降低了群落内接收多种花粉的物种所占的比例, 增加了两侧对称花的异种花粉数量(杨洁羽, 2018).而利用西藏入侵植物菊科印加孔雀草(Tagetes minuta)传粉者携带的花粉构建的植物花粉和传粉者网络结果表明, 印加孔雀草为泛化传粉系统, 并已经成功利用当地多种泛化传粉者为其授粉, 顺利整合进当地传粉网络(土艳丽等, 2019).此外, 全球气候变暖通过改变群落物种多样性, 或者影响植物与传粉者之间在物候、形态和分布区等方面的匹配性, 从而最终改变植物-传粉者网络的结构(肖宜安等, 2015). ... 外来引种植物印楝和辣木对红河干热河谷传粉网络的影响 1 2018 ... 青藏高原地区具有极为丰富的动植物多样性资源, 植物与传粉者间经过长期适应进化而形成了复杂的传粉网络, 传粉互作网络在维持生物多样性和生态系统服务功能中扮演着重要角色.目前, 我国在群落水平的传粉研究主要关注传粉网络的时空动态(Fang & Huang, 2012, 2016a; 赵延会, 2016), 外来植物(起国海, 2018; 土艳丽等, 2019)和动植物的系统发育(赵延会, 2016)对传粉网络的影响, 群落水平上的异种花粉转移(Fang & Huang, 2013, 2016b; 杨洁羽, 2018)等方面.连续3年对香格里拉高山草甸29种植物的花部特征与传粉功能群的关系进行研究, 结果表明, 虽然传粉网络结构是嵌套的、不对称的, 传粉者的访问偏好仍支持了传粉者在花部特征中的选择作用(Gong & Huang, 2011).此外, 该高山草甸中的开花植物与传粉者的种类在年际间变动较大, 但植物与其传粉者的连接及其强度基本都是稳定的, 而且熊蜂位于传粉网络的核心位置并对传粉功能起关键作用(Fang & Huang, 2012, 2016a).同地区内异种花粉转移的研究结果表明, 昆虫种间访问比例仅占所有访问次数的4%, 但是柱头上的异种花粉比例高达22%, 泛化植物接受多种其他植物的花粉, 但是不会输出更多的花粉给其他植物, 接受较多种间访问的植物具有较低的异种花粉落置数量, 这是共同开花植物降低异种花粉转移负面影响的一种适应策略(Fang & Huang, 2013, 2016b).在喜马拉雅-横断山地区玉龙雪山上海拔2 700-3 910 m范围内4个群落的传粉网络研究发现, 随着海拔升高, 传粉网络的特化水平显著升高而嵌套结构显著降低, 并且昆虫访花网络与传粉网络的拓扑结构存在显著变化(Zhao et al., 2019).另外, 植物及传粉者最近种间亲缘关系指数随海拔升高显著增加, 表明系统发育结构由离散逐渐转变为聚集, 然而植物及传粉者系统发育信号强度表现出不对称性, 传粉者系统发育关系的影响显著高于植物(赵延会, 2016).过度放牧、生物入侵等都会显著改变动植物的物种丰富度和多样性, 从而改变当地的传粉网络结构, 对生态系统功能产生负面影响(Soares et al., 2017).青藏高原东部高寒草甸的研究结果表明, 放牧降低了群落内接收多种花粉的物种所占的比例, 增加了两侧对称花的异种花粉数量(杨洁羽, 2018).而利用西藏入侵植物菊科印加孔雀草(Tagetes minuta)传粉者携带的花粉构建的植物花粉和传粉者网络结果表明, 印加孔雀草为泛化传粉系统, 并已经成功利用当地多种泛化传粉者为其授粉, 顺利整合进当地传粉网络(土艳丽等, 2019).此外, 全球气候变暖通过改变群落物种多样性, 或者影响植物与传粉者之间在物候、形态和分布区等方面的匹配性, 从而最终改变植物-传粉者网络的结构(肖宜安等, 2015). ... Plant molecular phylogeography in China and adjacent regions: Tracing the genetic imprints of Quaternary climate and environmental change in the world’s most diverse temperate flora 1 2011 ... 青藏高原独特的地形和气候条件孕育了极为丰富的植物多样性资源, 其中高原东南部的东喜马拉雅-横断山脉地区不仅是我国特有植物种类最为丰富的地区, 而且是现代北温带和高山植物区系的重要分化和起源中心(Huang et al., 2016).青藏高原隆升引起地理格局和自然环境的剧烈变化, 高山、峡谷等地理屏障阻碍了物种间基因流动, 提供了异质性生境和新的生态位, 从而驱动了植物的辐射演化和种类多样化(Yu et al., 2019).青藏高原地区维管植物种数约占中国的1/2, 其中仅分布于该区域的特有种类占35%以上(孙航, 2013), 特有种子植物共计113科519属3 764种.分子钟的证据表明, 青藏高原一些大属类群都是伴随着高原隆升分化形成的(于海彬等, 2018).进入第四纪更新世后(2.6-0.1 Ma), 冰期-间冰期的周期性气候波动进一步造就了青藏高原现今的植物多样性分布格局(Yu et al., 2019).谱系地理学研究指出, 该地区高山植物群体演化历史主要分为两种模式: 一、冰期退却到高原边缘的避难所, 冰后期回迁到高原面, 其中横断山脉地区常作为主要的避难所, 这也是该地区植物多样性丰富的原因之一; 二、地理隔离使得高原内部也存在很多避难所(含微型避难所), 如三江源地区, 植物在冰后期发生局域性扩张, 使得这些地区的物种多样性也比较丰富(Qiu et al., 2011). ... 甘青铁线莲花部特征的海拔变异及其繁殖适应 1 2017 ... 低温胁迫、暴雨频发以及强烈的紫外辐射是高山环境的典型特征.为应对这一系列极端环境, 高山植物往往会增加对支持及保护结构的投资, 如毛茛科甘青铁线莲(Clematis tangutica)在高海拔地区萼片十分发达, 以此来抵御外界不利因素对雌雄配子体的伤害(屈博, 2017).温度是植物生长与繁殖过程中重要的环境因子, 花发育、花蜜分泌等过程均受温度影响, 特别是在受到低温胁迫的高山植物带, 温度在花的形态、颜色和报酬等特征的进化过程中发挥着重要作用(Scaven & Rafferty, 2013).例如, “温室植物” (green house plant)菊科毡毛雪莲(Saussurea velutina)的苞片对花序具有保温作用, 有助于其发育(Yang & Sun, 2009); “棉毛植物” (cotton plant)菊科水母雪兔子(Saussurea medusa)的绒毛苞片除了具有保温效应, 还具有防水以及反射瞬时极端辐射的作用(Yang et al., 2008).此外, 在存在热量限制的高山地区, 向日运动(Zhang et al., 2010)及花闭合现象(He et al., 2006)也都是对低温环境较为常见的适应机制.大量研究发现, 花数量、花大小和花的对称性等特征均与温度紧密相关(Hoover et al., 2012; Yu et al., 2016).对青藏高原嵩草草甸64种开花植物花大小与花温度的测量结果表明, 花温度积累与花大小呈正相关关系, 且两侧对称花的温度积累比辐射对称花更为迅速(张国鹏, 2017).此外, 温度还可能通过影响植株的花蜜量、花蜜浓度和花蜜成分, 改变对传粉者的吸引力, 甚至影响传粉者的寿命(Hoover et al., 2012; Mu et al., 2015).高山带降水事件频发, 雨水会冲刷掉部分花粉、降低花粉的活性并稀释花蜜浓度(Huang et al., 2002; He et al., 2006), 因此一些植物进化出特殊的花部结构(比如下垂的花冠或者起遮盖作用的苞片等)来防止花粉受到雨水的影响(Huang et al., 2002; Sun et al., 2008).紫外辐射也会严重降低花粉的活性(Torabinejad et al., 1998).对横断山区的42种高山植物两种不同花部结构特征(即花粉暴露型与花粉遮盖型)研究表明, 花部具有的保护结构能有效地阻挡紫外辐射对花粉活力的影响(Zhang et al., 2014a).另外, 一些植物还演化出能够吸收紫外光的花部区域, 从而保护花粉等繁殖器官免受紫外辐射的伤害(Koski & Ashman, 2015).以上结果均表明紫外辐射在高山植物花部特征的进化过程中起到了非常重要的作用. ... 甘青铁线莲花部特征的海拔变异及其繁殖适应 1 2017 ... 低温胁迫、暴雨频发以及强烈的紫外辐射是高山环境的典型特征.为应对这一系列极端环境, 高山植物往往会增加对支持及保护结构的投资, 如毛茛科甘青铁线莲(Clematis tangutica)在高海拔地区萼片十分发达, 以此来抵御外界不利因素对雌雄配子体的伤害(屈博, 2017).温度是植物生长与繁殖过程中重要的环境因子, 花发育、花蜜分泌等过程均受温度影响, 特别是在受到低温胁迫的高山植物带, 温度在花的形态、颜色和报酬等特征的进化过程中发挥着重要作用(Scaven & Rafferty, 2013).例如, “温室植物” (green house plant)菊科毡毛雪莲(Saussurea velutina)的苞片对花序具有保温作用, 有助于其发育(Yang & Sun, 2009); “棉毛植物” (cotton plant)菊科水母雪兔子(Saussurea medusa)的绒毛苞片除了具有保温效应, 还具有防水以及反射瞬时极端辐射的作用(Yang et al., 2008).此外, 在存在热量限制的高山地区, 向日运动(Zhang et al., 2010)及花闭合现象(He et al., 2006)也都是对低温环境较为常见的适应机制.大量研究发现, 花数量、花大小和花的对称性等特征均与温度紧密相关(Hoover et al., 2012; Yu et al., 2016).对青藏高原嵩草草甸64种开花植物花大小与花温度的测量结果表明, 花温度积累与花大小呈正相关关系, 且两侧对称花的温度积累比辐射对称花更为迅速(张国鹏, 2017).此外, 温度还可能通过影响植株的花蜜量、花蜜浓度和花蜜成分, 改变对传粉者的吸引力, 甚至影响传粉者的寿命(Hoover et al., 2012; Mu et al., 2015).高山带降水事件频发, 雨水会冲刷掉部分花粉、降低花粉的活性并稀释花蜜浓度(Huang et al., 2002; He et al., 2006), 因此一些植物进化出特殊的花部结构(比如下垂的花冠或者起遮盖作用的苞片等)来防止花粉受到雨水的影响(Huang et al., 2002; Sun et al., 2008).紫外辐射也会严重降低花粉的活性(Torabinejad et al., 1998).对横断山区的42种高山植物两种不同花部结构特征(即花粉暴露型与花粉遮盖型)研究表明, 花部具有的保护结构能有效地阻挡紫外辐射对花粉活力的影响(Zhang et al., 2014a).另外, 一些植物还演化出能够吸收紫外光的花部区域, 从而保护花粉等繁殖器官免受紫外辐射的伤害(Koski & Ashman, 2015).以上结果均表明紫外辐射在高山植物花部特征的进化过程中起到了非常重要的作用. ... Physiological effects of climate warming on flowering plants and insect pollinators and potential consequences for their interactions 1 2013 ... 低温胁迫、暴雨频发以及强烈的紫外辐射是高山环境的典型特征.为应对这一系列极端环境, 高山植物往往会增加对支持及保护结构的投资, 如毛茛科甘青铁线莲(Clematis tangutica)在高海拔地区萼片十分发达, 以此来抵御外界不利因素对雌雄配子体的伤害(屈博, 2017).温度是植物生长与繁殖过程中重要的环境因子, 花发育、花蜜分泌等过程均受温度影响, 特别是在受到低温胁迫的高山植物带, 温度在花的形态、颜色和报酬等特征的进化过程中发挥着重要作用(Scaven & Rafferty, 2013).例如, “温室植物” (green house plant)菊科毡毛雪莲(Saussurea velutina)的苞片对花序具有保温作用, 有助于其发育(Yang & Sun, 2009); “棉毛植物” (cotton plant)菊科水母雪兔子(Saussurea medusa)的绒毛苞片除了具有保温效应, 还具有防水以及反射瞬时极端辐射的作用(Yang et al., 2008).此外, 在存在热量限制的高山地区, 向日运动(Zhang et al., 2010)及花闭合现象(He et al., 2006)也都是对低温环境较为常见的适应机制.大量研究发现, 花数量、花大小和花的对称性等特征均与温度紧密相关(Hoover et al., 2012; Yu et al., 2016).对青藏高原嵩草草甸64种开花植物花大小与花温度的测量结果表明, 花温度积累与花大小呈正相关关系, 且两侧对称花的温度积累比辐射对称花更为迅速(张国鹏, 2017).此外, 温度还可能通过影响植株的花蜜量、花蜜浓度和花蜜成分, 改变对传粉者的吸引力, 甚至影响传粉者的寿命(Hoover et al., 2012; Mu et al., 2015).高山带降水事件频发, 雨水会冲刷掉部分花粉、降低花粉的活性并稀释花蜜浓度(Huang et al., 2002; He et al., 2006), 因此一些植物进化出特殊的花部结构(比如下垂的花冠或者起遮盖作用的苞片等)来防止花粉受到雨水的影响(Huang et al., 2002; Sun et al., 2008).紫外辐射也会严重降低花粉的活性(Torabinejad et al., 1998).对横断山区的42种高山植物两种不同花部结构特征(即花粉暴露型与花粉遮盖型)研究表明, 花部具有的保护结构能有效地阻挡紫外辐射对花粉活力的影响(Zhang et al., 2014a).另外, 一些植物还演化出能够吸收紫外光的花部区域, 从而保护花粉等繁殖器官免受紫外辐射的伤害(Koski & Ashman, 2015).以上结果均表明紫外辐射在高山植物花部特征的进化过程中起到了非常重要的作用. ... Climate change impacts on pollination 1 2016 ... 增温影响植物的物候并引起物种的迁移, 最终将导致植物与传粉者的时空不匹配, 从而扰乱两者的协同进化关系, 植物因缺乏合适的传粉者而导致传粉失败, 其通过有性繁殖产生的个体数目会显著降低, 并可能最终导致植物群落衰退(Settele et al., 2016).增温对不同开花物候参数的影响程度和模式不同, 其中研究最多的是初花时间.增温对初花时间影响的一般规律是: 随着气温升高, 多数物种开花时间提前, 少数物种开花时间延迟, 也有的植物没有明显变化(肖宜安等, 2015).例如, 在藏北高寒草甸的模拟增温实验中, 增温处理导致不同类型植物的繁殖时间发生明显偏移, 这可能会影响群落中相邻开花种类的繁殖重叠时间, 进而改变物种间的竞争关系(朱军涛, 2016).对20世纪20年代以来横断山地区蔷薇属(Rosa)和栒子属(Cotoneaster) 12种植物腊叶标本的研究发现, 气候变化背景下蔷薇属植物开花物候显著延迟, 而栒子属植物的开花物候没有明显变化, 这反映出即使在亲缘关系很近的物种间, 开花物候对气候变化的响应也同样存在显著的种间差异(Yu et al., 2016).以往研究指出, 不同物种之间物候期的响应差异主要是因为植物生活型、开花时间、传粉类型等特性的差别(Fitter & Fitter, 2002; Dorji et al., 2013).与多年生、晚期开花和风媒传粉植物相比, 一年生、早期开花和虫媒传粉植物对气候变暖的影响更敏感(Hegland et al., 2009; Munson & Long, 2017).另外, 不同根系深度的植物对增温引起的表层土壤水分胁迫的响应和敏感性也不同, 与浅根植物相比, 深根植物的物候过程较少受到土壤水分的限制(朱军涛, 2016). ... Flower colour and phylogeny along an altitudinal gradient in the Himalayas of Nepal 2 2014 ... 花部特征演化的最有效传粉者原则(most effective pollinator principle)认为在花部性状的进化过程中, 传粉者介导的选择扮演了重要的作用(黄双全, 2014).花性状的组合(即形态、颜色、气味、报酬等)和特定的传粉者类群相关, 被称为传粉综合征(pollination syndromes).在高山极端环境下, 传粉者在花部形态特征演化过程中发挥着极大的驱动作用.在群落水平上对横断山区27种开花植物的传粉者泛化水平(pollinator generalization level)的定量分析结果表明, 与辐射对称的花相比, 两侧对称的花增强了与特异性传粉者的相互作用, 从而增加了花粉落置的精确性(Gong & Huang, 2009).对青藏高原东部高山草甸50种开花植物的花和叶特征的分析结果显示: 花大小的变异性显著低于叶片大小的变异性, 这表明传粉者对花部特征施加的稳定选择有利于花的稳定性(路宁娜和赵志刚, 2014).六叶龙胆在高海拔地区雄蕊和雌蕊缩短, 花药与柱头间的距离(雌雄异位)缩小, 这是在传粉者稀少的情况下增加自花授粉能力的一种适应性策略(He at al., 2017).毛茛科矮金莲花(Trollius farreri)在花粉限制压力更大的高海拔地区通过增大萼片等花展示的投入以吸引传粉者, 而在低海拔区域则增加了对退化花瓣这种产蜜结构的生物量分配, 这表明在高海拔地区花的展示结构对雌性繁殖成功的贡献要大于传粉报酬的贡献(孟金柳, 2010).花色是一种吸引传粉者的显著视觉信号, 指示传粉者取食和传粉.高山植物群落中花的主导色彩会随着地域和季节的变化而不同, 这可能与传粉者的种类变化及其偏好有关(彭德力等, 2012).针对尼泊尔境内喜马拉雅山区亚热带地区(海拔900-2 000 m)和亚高山地区(海拔3 000- 4 100 m) 107种植物的花色研究(图2A)发现, 其蓝紫色花的比例显著小于平均海拔4 300 m以上的横断山区(图2B、2C), 这可能与熊蜂作为横断山区主要传粉者有密切关系(Gong & Huang, 2011).值得注意的是, 除了传粉者的选择压力, 植物的花色还可能受到其他生物因素(如植食者)和非生物因素(如紫外辐射)的影响, 青藏高原及周边高山地区丰富的植物种类和复杂多样的生境类型为检验花色的影响机制提供了有利条件, 但目前此类研究还十分欠缺, 有待今后开展深入广泛的研究.除了丰富的花色, 一些植物还演化出对传粉者具有特殊吸引和指示作用的花图案或花结构, 即“蜜导” (nectar guide), 借此控制访花者的落置方向和位置并进一步提高其访花效率(Leonard & Papaj, 2011; Hansen et al., 2012).报酬(主要为花蜜)也是植物吸引传粉者的重要因素, 不同传粉者对花蜜的浓度、成分、颜色和分泌动态都表现出不同的偏好(Baker & Baker, 1983).例如, 喜马拉雅山区分布的唇形科米团花(Leucosceptrum canum)通过花蜜的色彩和动态变化来吸引鸟类传粉者(Zhang et al., 2012); 我国西南山区分布的唇形科野拔子(Elsholtzia rugulosa)花蜜中的酚类化合物起到吸引传粉者同时又排斥非传粉者的双重作用(Zhang et al., 2016).不同植物的花寿命具有显著差异, 花寿命的可塑性也是植物对传粉环境变化的一种适应对策(Trunschke & Stöcklin, 2017).对青藏高原高寒草甸不同海拔11种开花植物的花寿命进行对比研究发现, 高海拔群落中植物的花寿命显著延长, 且可塑性更大, 说明植物通过提高花寿命及其可塑性以适应传粉者稀少的高寒环境(王玉贤等, 2018).
本文的其它图/表
|

{kind=link}