|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
光响应曲线的指数改进模型与常用模型比较
植物生态学报
2012, 36 (12):
1277-1285.
DOI: 10.3724/SP.J.1258.2012.01277
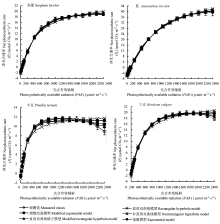
光响应曲线的参数是研究植物生理状态的重要指标, 常用的光响应曲线模型无法准确地计算出光饱和点和最大净光合速率。该文利用光响应曲线新模型——指数改进模型、直角双曲线模型、直角双曲线修正模型、非直角双曲线模型和指数模型, 拟合高粱(Sorghum bicolor)、苋(Amaranthus tricolor)、大麦(Hordeum vulgare)和半夏(Pinellia ternata)的光响应曲线, 并随机选取部分数据进行检验, 得到了各模型计算出的主要生理参数, 并对这些数据进行了比较分析, 讨论了各模型之间的优缺点和准确性, 描述了C3、C4植物光响应的适宜性。结果表明, 基于C3植物得到的指数改进模型和直角双曲线修正模型能较准确地计算出C3、C4植物饱和光强和最大净光合速率, 并在描述光响应曲线时比另外3个模型具有更高的精确性和适宜性。实验结果可为光响应曲线模型在C3和C4光合途径植物中的应用提供参考。
表2
不同模型计算出的4种材料的生理参数和观测值的比较
正文中引用本图/表的段落
从表2可见, 所有模型得出的LCP均在观测值范围内。Pmax除指数改进模型和直角双曲线修正模型的计算值比较接近观测值外, 非直角双曲线模型、直角双曲线模型和指数模型计算出的Pmax均不同程度地大于观测值。各模型计算的Rdark除直角双曲线模型略大外, 都比较接近观测值。另外, 只有指数改进模型和直角双曲线修正模型才能通过模型直接计算LSP。‘藏青320’ 2个模型的LSP计算值均比较接近观测值。但是由这2个模型计算的高粱的LSP分别为1952和2071 μmol·m-2·s-1。苋直角双曲线修正模型的LSP为2574 μmol·m-2·s-1, 稍大于观测值2200 μmol·m-2·s-1; 半夏指数改进模型的LSP为967 μmol·m-2·s-1, 略小于观测值1200 μmol·m-2·s-1。
光响应曲线的数学模型可以快速地估算许多有生理意义的参数, 例如: Pmax、Rdark和LCP。在农业生产中, 当PAR超过LSP时, 为了避免作物产生光抑制, 增加产量, 常采取遮阴或浇水的办法来保证作物正常的光合作用(许大全, 1997)。因此LSP是作物的一个重要生理指标。苋直角双曲线修正模型的LSP为2574 μmol·m-2·s-1, 略大于观测值2200 μmol·m-2·s-1。苋的指数改进模型的MSE和MAE比直角双曲线修正模型小, 这说明苋的指数改进模型描述的光响应曲线比直角双曲线修正模型的要准确。于是苋的LSP为指数改进模型的计算值2295 μmol·m-2·s-1。根据以上分析方法, 高粱的LSP为指数改进模型的计算值1952 μmol·m-2·s-1; 半夏的LSP为直角双曲线修正模型的计算值1122 μmol·m-2·s-1; ‘藏青320’的LSP为直角双曲线修正模型的计算值1580 μmol·m-2·s-1。这表明指数改进模型和直角双曲线修正模型在不同植物具体的LSP应用中存在一定差异, 但是计算出的LSP值和观测值相比, 差异都不大。在C3、C4植物的光响应曲线应用中, 直角双曲线模型、非直角双曲线模型和指数模型计算出的Pmax都不同程度地大于观测值, 而指数改进模型和直角双曲线修正模型的Pmax和观测值均比较接近(表2) (Ye, 2007; Chen et al., 2011)。各模型的LCP和φ0计算值的差异不大。Rdark除直角双曲线模型的大于观测值外, 其他4个模型的Rdark都比较接近观测值。由此可见, 不同光合途径的植物由指数改进模型和直角双曲线修正模型计算的光响应曲线生理参数均比较接近观测值。
本文的其它图/表
|