|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
陆地生境岛屿藓类植物小岛屿效应驱动因素分析——以太行山脉中段山顶为例
植物生态学报
2023, 47 (1):
65-76.
DOI: 10.17521/cjpe.2022.0272
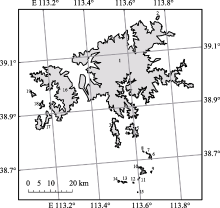
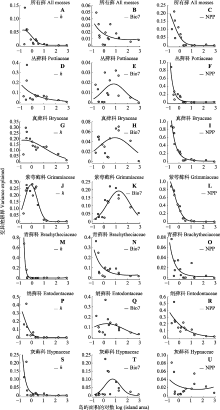
小岛屿效应描述了种-面积关系的一种特殊现象, 是当前生物地理学和生物多样性研究理论框架的重要组成部分。随着气候变暖, 山顶物种的生存受到威胁, 然而以山顶生境岛屿为载体对小岛屿效应的研究还十分缺乏。该研究以太行山脉中段19个面积0.06-801.58 km2的山顶生境岛屿为研究区, 在2019-2021年的夏秋季对藓类进行调查。共记录到藓类131种, 隶属于23科68属。采用6种种-面积关系回归模型, 分别检测了所有藓和6个常见藓科是否存在小岛屿效应。根据小岛屿效应形成机制的生境多样性假说、灭亡假说和营养补给假说, 选择了岛屿高度、温度年变化范围和单位面积净初级生产力作为变量, 对小岛屿效应的驱动因素进行分析。在各类群组中, 使用多元线性回归和变差分解分别评估上述3个变量对物种丰富度变化的线性影响。首先使用5个面积最小的岛屿进行分析, 计算出3个变量对物种丰富度变化的贡献, 然后以迭代的方式逐次加入面积更大的1个岛屿, 并再次进行变差分解分析。最后使用广义线性回归分析了3个变量对物种丰富度变化的贡献在迭代过程中的变化趋势。结果显示, 所有藓和6个常见藓科均存在小岛屿效应, 其面积阈值分布在0.36-106.91 km2间。各组藓类小岛屿效应的驱动因素具有差异性, 其中, 除了紫萼藓科之外, 其他各组均支持生境假说; 除了丛藓科、真藓科、紫萼藓科和灰藓科之外, 其他各组均支持灭亡假说; 而各组在不同程度上普遍支持营养补给假说。整体而言, 面积约为10 km2以上的岛屿维持了大量的藓类物种多样性, 需要重点保护。对于生境要求较为单一的紫萼藓科而言, 保护石生环境是保护其物种多样性的关键; 而对于其他科藓类而言, 生境类型多样性的保护是维持物种多样性的重要保障。营养补给假说的普遍适用性揭示了山顶下方森林生态系统的资源补给作用减缓了由于面积的减小而造成的藓类物种数量下降, 因此保护山顶下方林地内物种多样性和群落稳定性对维持山顶藓类物种多样性具有重要意义。
表3
太行山脉中段山顶生境岛屿藓类植物的种-面积关系拟合结果
正文中引用本图/表的段落
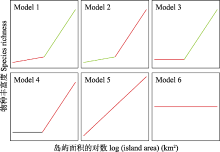
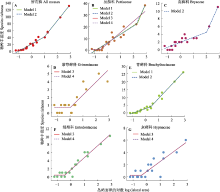
根据AICc值, 所有藓和6个常见科藓类均存在小岛屿效应(图3; 表3)。先缓后升的两段式模型(模型1和2; ?AICc = 0)是所有藓的最优种-面积关系。次优模型为先平后升的两段式模型(模型3和4)。最优模型与次优模型的?AICc > 2, 表明先缓后升的两段式模型(模型1和2)是拟合所有藓种-面积关系的最佳模型。根据最佳模型提供的T, 所有藓的面积阈值为100.993 = 9.84 km2。对于丛藓科而言, 先缓后升的两段式模型(模型1和2)是最优的种-面积关系; 次优模型为先平后升的两段式模型(模型3和4)。最优模型与次优模型的?AICc < 2, 表明次优模型与最优模型具有同等解释效力。因此, 其面积阈值是4个模型所对应的面积阈值的平均值, 即(102.338 + 102.228 + 10-0.565 + 10-0.565)/4 = 96.83 km2。同理, 真藓科的最佳种-面积关系为模型2, 其面积阈值为106.91 km2。紫萼藓科的最佳种-面积关系为模型3和4, 其面积阈值为1.03 km2。青藓科的最佳种-面积关系为模型1和2, 其面积阈值为2.42 km2。绢藓科的最佳种-面积关系为模型3和4, 其面积阈值为0.36 km2。灰藓科的最佳种-面积关系为模型3和4, 其面积阈值为0.43 km2。
一般而言, 对于资源需求相对较高、扩散能力较低、生存能力较弱的生物类群, 其面积阈值往往较高(Lomolino & Weiser, 2001; Gao & Perry, 2016a)。Yu等(2020)对舟山群岛苔藓植物的小岛屿效应研究中发现侧蒴藓类的面积阈值高于顶蒴藓类, 并将其归因于顶蒴藓类的抗旱性强于侧蒴藓类。然而, 本研究并未发现侧蒴藓类的面积阈值普遍地高于顶蒴藓类, 这可能是由于3方面原因导致的。第一, 虽然总体而言侧蒴藓类的耐旱性弱于顶蒴藓类, 但是同为侧蒴藓类的绢藓科、青藓科与灰藓科之间本身在耐旱性上也有着很大的差异性, 绢藓科、青藓科更倾向于阴生, 灰藓科则倾向于抗干旱型, 笼统地对侧蒴藓类和顶蒴藓类进行比较, 可能得不出精准的科学结论。第二, 在其他方面, 侧蒴藓类可能具有优越的适应能力。首先, 根据观察, 一些侧蒴藓类物种如灰青藓(Brachythecium glauculum)和深绿绢藓(Entodon luridus)在山顶广泛分布; 而一些顶蒴藓类物种如刺叶真藓(Bryum lonchocaulon)和圆蒴连轴藓(Schistidium apocarpum)只分布在最大的山顶。这表明一些侧蒴藓类物种具有更强的扩散能力。其次, 从孢子体着生位置上看, 顶蒴藓类的蒴柄着生于茎顶, 通常不分枝, 如小树一般直立生长, 多表现为丛集型; 而侧蒴藓类的蒴柄从叶腋中发生(Vanderpoorten & Goffinet, 2009), 通常以平铺型、交织型出现(Kürschner, 2004)。在风速较高的山顶处, 更加贴近地面、相互交织生长的侧蒴藓类比顶蒴藓类可能表现出更强的固着性。另外, 除了上述扩散和风速条件, pH、CaCO3含量、植被盖度、有机质含量以及土壤质地等也都是影响藓类植物分布的重要环境因素(张元明等, 2002), 因此仅仅从抗旱性一个角度并不能准确地比较不同类群对于环境的综合适应能力。第三, 本研究仅使用了两片段的分段回归模型对小岛屿效应进行检测, 并未尝试三片段的分段回归模型。由于三片段的分段回归模型拥有更多的模型参数, 一般要求岛屿数量在40座以上时方可应用(Gao et al., 2019; Gao & Wang, 2022)。未来的研究工作可以继续调查周围更多的山顶, 使更复杂的种-面积关系模型得以应用, 以期对藓类植物小岛屿效应的面积阈值进行更精确的估计。
本文的其它图/表
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||