昼夜不对称增温对长白山阔叶红松林碳汇能力的影响
李伟斌, 张红霞, 张玉书, 陈妮娜
植物生态学报
2023, 47 ( 9):
1225-1233.
DOI: 10.17521/cjpe.2022.0478
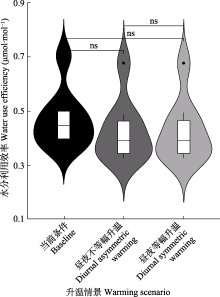
过去50年的气温数据表明全球陆地表面在夜间比白天变暖更快, 然而以往的研究大多关注全天对等增温的影响, 对昼夜不对称增温效应的认识不足。该研究利用光合增益和水力成本优化模型分析了两种增温情景(昼夜等幅升温和昼夜不等幅升温)对长白山阔叶红松林植被动态的影响。结果表明: 光合增益和水力成本优化模型可以很好地模拟长白山阔叶红树林的碳收支状态(R2 = 0.67, p < 0.001)。增温普遍促进了长白山阔叶红树林的碳汇(11.2%-13.8%), 但未显著改变其水分利用效率; 而不同增温情景对年固碳量的促进作用并无显著差异。与此同时, 增温增加了森林植被的水分压力, 从而增加了植物的导水率损失百分数(水力脆弱性, 1.1%)。由此可见, 相比于当前气候条件, 所有增温情景均会提高森林的碳汇能力, 但同时也会加大森林的死亡风险, 进而降低森林碳汇潜力。
View image in article
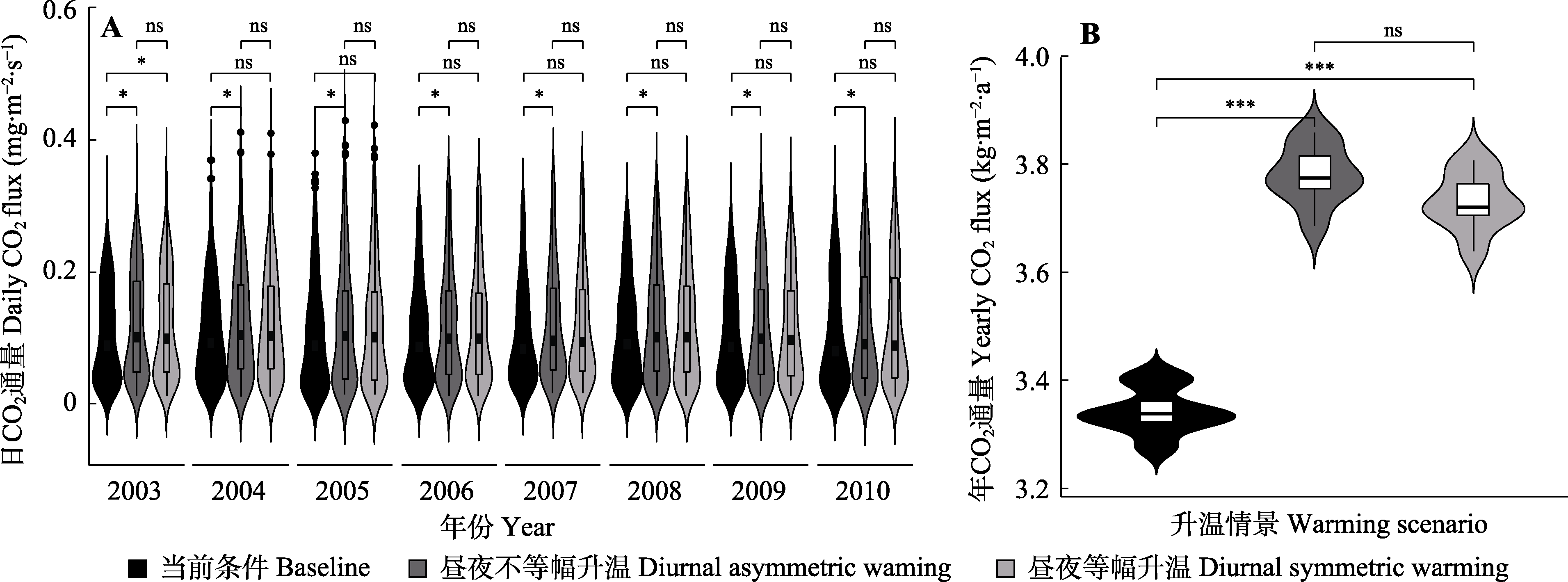
图2
2003-2010年不同增温情景下长白山阔叶红松林CO2通量的变化. A, 日平均动态。B, 年累计固碳量。*, p < 0.05; ***, p < 0.001; ns, p > 0.05。
正文中引用本图/表的段落
长白山阔叶红松林的固碳速率(CO2通量)具有明显的年际差异(图2A); 在2003-2010年自然条件下, 长白山阔叶红松林的平均年累计固碳量约为3.3 kg CO2·m-2·a-1 (图2B)。不同增温情景下长白山阔叶红松林固碳量的模拟结果表明, 增温会显著促进长白山阔叶红松林对大气CO2的吸收速率(固碳速率;图2)。总体而言, 虽然不同增温情景之间的结果并无显著差异, 但昼夜不等幅升温和昼夜等幅升温对年累计固碳量的增加效率分别为12.5%-13.8%和11.2%-12.5% (图2B)。
本研究利用Sperry模型模拟了长白山阔叶红松林2003-2010年的净光合速率, 结果表明模拟结果与通量净生产力具有显著的一致性, 表明该模型可准确地模拟我国长白山阔叶红松林的植被生长动态。基于此模型对昼夜等幅升温效应的模拟发现全球变暖将显著增加长白山阔叶红松林的碳汇能力, 但并未改变其水分利用效率, 这与之前基于全天对等增温研究的结果(Liang et al., 2013; Cao et al., 2018)一致。对昼夜不对称增温的模拟结果表明: 昼夜不等幅升温显著提高了该区森林的植被碳固定速率(提高了12.5%-13.8%; 图2), 这与Peng等(2013)利用遥感数据在北半球陆地生态系统中得到的结果相似, 即夜间增温对温带地区植被的生长具有正向影响。虽然在白天增温能够提高光合酶的活性, 促进光合效率(Liang et al., 2013); 然而白天增温也会通过提高植物的蒸腾引起较大的土壤水分损失, 植物只能通过降低气孔开放以减少水分损失, 这同时抑制了光合作用(Zhang et al., 2020a)。相反, 虽然夜间增温可以增加呼吸作用的碳消耗, 但是由于夜间蒸腾速率较小, 节约的土壤水分可供白天光合作用使用, 利于白天的光合作用。综合来看, 虽然昼夜等幅升温也有利于长白山阔叶红松林的生长(主要受益于增强的光合酶活性), 但是远不及昼夜不等幅升温带来的“光合的超补偿作用” (朱军涛和郑家禾, 2022), 补偿程度为12.5%-13.8%; 当然上述补偿程度还需通过模拟控制实验进行验证。
1)植物参数中两个值分别为红松和阔叶树种参数. ... The response of stomatal conductance to seasonal drought in tropical forests 1 2020 ... 气孔的开合是调节大气CO2进入植物叶片进行光合固碳和植物水分蒸腾散失的关键, 是模拟全球变暖下植物碳汇潜力的基础, 而现有的陆面过程模型难以反映气孔对复杂环境条件的响应(Wu et al., 2020).Sperry等(2017)开发的光合增益和水力成本优化模型假设植物可以通过调节气孔使其在最少蒸腾失水的情况下, 保证足够的CO2进入气孔以取得最大光合速率, 已被用于干旱、全球变暖以及大气CO2浓度升高等全球变化背景下森林生态系统生长碳汇以及死亡率的模拟研究(Sperry et al., 2019; Li et al., 2022).与遥感数据的分析结果不同, 利用模型可以评估、预测任何可能发生的气候情景下研究区植被碳汇潜力的大小, 并能有效地检验单一因子(如温度)变化对结果的影响, 是分析不同气候情景影响的有效手段(Peng et al., 2013). ... Terrestrial carbon cycle affected by non-uniform climate warming 1 2014 ... 已有的增温模拟实验和模型结果均表明全球变暖能够通过促进光合作用而提高生态系统的碳固定量(Bai et al., 2013; Liang et al., 2013; Zhang et al., 2015; Du et al., 2020; Li et al., 2022), 然而, 这些研究并未考虑昼夜增温速率差异的影响.作为影响陆地生态系统碳汇能力的关键因素, 越来越多的研究开始关注昼夜不对称增温的影响(Wan et al., 2009; Peng et al., 2013; Tan et al., 2015; 赵杰等, 2018).例如, Peng等(2013)结合遥感数据和大气反演模型系统地分析了1982-2009年期间的昼夜不对称增温对北半球陆地生态系统碳汇通量的影响, 结果表明白天增温能够促进寒、温带湿润地区植被的生长, 但不利于温带干旱和半干旱(水分条件较差)地区植被的生长, 而夜间增温对植被生长的影响正好相反.同样, 赵杰等(2018)利用遥感数据在中国温带地区的研究发现植被活动对白天增温的响应比夜间增温更大, 且多为正向反应, 然而这种响应具有较大的季节异质性(Tan et al., 2015).此类研究仅关注了历史时期的昼夜不同速增温对植被动态的影响, 并未涉及不同增温情景影响的差异, 以及这种增温效应对植被后期生长、生存的影响.未来气候变暖持续加快, 将进一步加大昼夜增温幅度的差异, 温度的升高将引起饱和水汽压差(VPD)指数式的上升, 增加植物的水分压力(Grossiord et al., 2020), 尤其是在气孔导度相对较高的白天, 引起植被生长的停滞, 甚至死亡(McDowell & Allen, 2015; Hartmann et al., 2022; Gong & Hao, 2023), 进而改变未来陆地植被的分布及碳汇能力.因此, 探讨不对称增温对植被碳汇能力及植被对新环境适应能力的影响, 可提高对未来气候变化情景下陆地生态系统碳汇预测的准确性(Xia et al., 2014; 朱军涛和郑家禾, 2022). ... 陆地生态系统过程对气候变暖的响应与适应 1 2020 ... 全球气候变暖已成为不争的事实, 从1850- 1900年到2006-2015年全球平均陆地表面气温上升了1.53 ℃ (IPCC, 2019).而且, 越来越多的研究结果表明全球陆地变暖速率存在较大的昼夜差异, 已有的数据表明过去50年来全球陆地系统夜间温度的上升速率是白天的1.4倍(Vose et al., 2005; Das et al., 2011; IPCC, 2019).由于植物生理过程的昼夜差异(如白天以光合碳吸收为主, 而呼吸碳消耗贯穿全天), 这种不对称增温将对植物的生存策略及碳源、汇产生重要影响(Liang et al., 2013; Peng et al., 2013; Li et al., 2018; McDowell et al., 2020; 夏建阳等, 2020; Hartmann et al., 2022), 从而对未来陆地生态系统的碳汇能力产生复杂影响.因此, 进一步探讨这种昼夜不对称增温对陆地生态系统植被动态的具体影响对于厘清全球变暖对陆地碳循环的影响具有重要意义(朱军涛和郑家禾, 2022). ... 陆地生态系统过程对气候变暖的响应与适应 1 2020 ... 全球气候变暖已成为不争的事实, 从1850- 1900年到2006-2015年全球平均陆地表面气温上升了1.53 ℃ (IPCC, 2019).而且, 越来越多的研究结果表明全球陆地变暖速率存在较大的昼夜差异, 已有的数据表明过去50年来全球陆地系统夜间温度的上升速率是白天的1.4倍(Vose et al., 2005; Das et al., 2011; IPCC, 2019).由于植物生理过程的昼夜差异(如白天以光合碳吸收为主, 而呼吸碳消耗贯穿全天), 这种不对称增温将对植物的生存策略及碳源、汇产生重要影响(Liang et al., 2013; Peng et al., 2013; Li et al., 2018; McDowell et al., 2020; 夏建阳等, 2020; Hartmann et al., 2022), 从而对未来陆地生态系统的碳汇能力产生复杂影响.因此, 进一步探讨这种昼夜不对称增温对陆地生态系统植被动态的具体影响对于厘清全球变暖对陆地碳循环的影响具有重要意义(朱军涛和郑家禾, 2022). ... Divergences in hydraulic conductance and anatomical traits of stems and leaves in three temperate tree species coping with drought, N addition and their interactions 1 2020a ... 本研究利用Sperry模型模拟了长白山阔叶红松林2003-2010年的净光合速率, 结果表明模拟结果与通量净生产力具有显著的一致性, 表明该模型可准确地模拟我国长白山阔叶红松林的植被生长动态.基于此模型对昼夜等幅升温效应的模拟发现全球变暖将显著增加长白山阔叶红松林的碳汇能力, 但并未改变其水分利用效率, 这与之前基于全天对等增温研究的结果(Liang et al., 2013; Cao et al., 2018)一致.对昼夜不对称增温的模拟结果表明: 昼夜不等幅升温显著提高了该区森林的植被碳固定速率(提高了12.5%-13.8%; 图2), 这与Peng等(2013)利用遥感数据在北半球陆地生态系统中得到的结果相似, 即夜间增温对温带地区植被的生长具有正向影响.虽然在白天增温能够提高光合酶的活性, 促进光合效率(Liang et al., 2013); 然而白天增温也会通过提高植物的蒸腾引起较大的土壤水分损失, 植物只能通过降低气孔开放以减少水分损失, 这同时抑制了光合作用(Zhang et al., 2020a).相反, 虽然夜间增温可以增加呼吸作用的碳消耗, 但是由于夜间蒸腾速率较小, 节约的土壤水分可供白天光合作用使用, 利于白天的光合作用.综合来看, 虽然昼夜等幅升温也有利于长白山阔叶红松林的生长(主要受益于增强的光合酶活性), 但是远不及昼夜不等幅升温带来的“光合的超补偿作用” (朱军涛和郑家禾, 2022), 补偿程度为12.5%-13.8%; 当然上述补偿程度还需通过模拟控制实验进行验证. ... Responses of functional traits to seven-year nitrogen addition in two tree species: coordination of hydraulics, gas exchange and carbon reserves 1 2020b ... 本研究选取的长白山原始阔叶红松林位于我国吉林省东南部的中朝交界处(41.38°-42.60° N, 126.92°-129.00° E), 海拔500-1 100 m, 属于典型的温带大陆性季风山地气候(李伟斌等, 2014), 夏季短且温暖湿润, 冬季寒冷且漫长(长达7个月).该研究区近34年(1982-2015)的年平均气温为4.0 ℃, 年降水量为711.3 mm, 主要集中在6-9月, 约占全年降水量的80%.该区域地势平坦(坡度1°-5°), 土壤为发育于火山灰沙砾土的暗棕色森林土, 主要建群针叶树种为红松, 阔叶树种包括蒙古栎(Quercus mongolica)、水曲柳(Fraxinus mandshurica)、紫椴(Tilia amurensis)和五角槭(Acer pictum subsp. mono)等, 林分结构复杂, 平均林龄180年左右, 立木约560株·hm-2, 平均株高26 m, 郁闭度0.8, 最大叶面积指数接近6.0 m2·m-2 (李伟斌等, 2014; Zhang et al., 2020b). ... 长白山自然保护区生态系统碳平衡研究 1 2003 ... 长白山阔叶红松(Pinus koraiensis)林是我国温带主要森林类型, 对我国森林生态系统固碳功能的发挥具有重要作用(张娜等, 2003; Li et al., 2017), 揭示其对昼夜不对称增温的生理响应, 对科学表达我国温带森林碳动态, 准确评估和预测全球变化背景下森林生态系统碳收支具有重要意义.本研究通过假定不同的增温情景(昼夜等幅升温和昼夜不等幅升温), 利用光合增益和水力成本优化模型评估了昼夜不对称增温对长白山阔叶红松林碳汇动态和死亡风险的影响.具体来说, 本研究使用2003-2010年的气候数据分别模拟了不同增温模式下长白山阔叶红松林的固碳速率, 并在树木死亡机制(McDowell et al., 2008)的基础上评估了不同增温情景对未来森林死亡风险的影响, 以期提高昼夜不对称增温对森林植被动态影响的认识, 为准确评估未来气候变化情景下森林生态系统碳汇能力奠定基础. ... 长白山自然保护区生态系统碳平衡研究 1 2003 ... 长白山阔叶红松(Pinus koraiensis)林是我国温带主要森林类型, 对我国森林生态系统固碳功能的发挥具有重要作用(张娜等, 2003; Li et al., 2017), 揭示其对昼夜不对称增温的生理响应, 对科学表达我国温带森林碳动态, 准确评估和预测全球变化背景下森林生态系统碳收支具有重要意义.本研究通过假定不同的增温情景(昼夜等幅升温和昼夜不等幅升温), 利用光合增益和水力成本优化模型评估了昼夜不对称增温对长白山阔叶红松林碳汇动态和死亡风险的影响.具体来说, 本研究使用2003-2010年的气候数据分别模拟了不同增温模式下长白山阔叶红松林的固碳速率, 并在树木死亡机制(McDowell et al., 2008)的基础上评估了不同增温情景对未来森林死亡风险的影响, 以期提高昼夜不对称增温对森林植被动态影响的认识, 为准确评估未来气候变化情景下森林生态系统碳汇能力奠定基础. ... 碳中和目标下中国森林固碳量跨期分配及成本 1 2022 ... 由于本研究是建立在假定的增温情景和模型之上的, 因此存在以下三方面的不确定性: 1)模型选择的不确定性, 本研究所用的Sperry模型相对比较简单, 未考虑植物本身对不对称增温的适应性(朱军涛和郑家禾, 2022), 如植物根系特征(Bai et al., 2012)、碳分配(Bai et al., 2012; Li et al., 2020)和养分吸收(Prieto & Querejeta, 2020)等的变化, 限制了对未来情景的精确预测; 2)情景假设的不确定性, 1.5 ℃的增温幅度相对较高, 尤其在我国为实现“碳中和”目标的努力下(张祥华等, 2022), 这可能会高估长白山阔叶红松林的固碳能力及夜间增温所带来的“光合超补偿作用”; 3)增温引起水分压力的不确定性, 增温将引起VPD的指数式上升(Grossiord et al., 2020), 但在本研究中并未因为增温而改变模型输入的VPD, 这同样将高估固碳能力, 而低估导水率损失百分比(低估水力学失败发生的可能性) (Li et al., 2022).此外, 增温对未来降水格局的变化以及未来高CO2环境对植物水分利用效率的影响均未考虑, 增加了结果的不确定性. ... 碳中和目标下中国森林固碳量跨期分配及成本 1 2022 ... 由于本研究是建立在假定的增温情景和模型之上的, 因此存在以下三方面的不确定性: 1)模型选择的不确定性, 本研究所用的Sperry模型相对比较简单, 未考虑植物本身对不对称增温的适应性(朱军涛和郑家禾, 2022), 如植物根系特征(Bai et al., 2012)、碳分配(Bai et al., 2012; Li et al., 2020)和养分吸收(Prieto & Querejeta, 2020)等的变化, 限制了对未来情景的精确预测; 2)情景假设的不确定性, 1.5 ℃的增温幅度相对较高, 尤其在我国为实现“碳中和”目标的努力下(张祥华等, 2022), 这可能会高估长白山阔叶红松林的固碳能力及夜间增温所带来的“光合超补偿作用”; 3)增温引起水分压力的不确定性, 增温将引起VPD的指数式上升(Grossiord et al., 2020), 但在本研究中并未因为增温而改变模型输入的VPD, 这同样将高估固碳能力, 而低估导水率损失百分比(低估水力学失败发生的可能性) (Li et al., 2022).此外, 增温对未来降水格局的变化以及未来高CO2环境对植物水分利用效率的影响均未考虑, 增加了结果的不确定性. ... A meta-analysis of the effects of experimental warming on soil carbon and nitrogen dynamics on the Tibetan Plateau 1 2015 ... 已有的增温模拟实验和模型结果均表明全球变暖能够通过促进光合作用而提高生态系统的碳固定量(Bai et al., 2013; Liang et al., 2013; Zhang et al., 2015; Du et al., 2020; Li et al., 2022), 然而, 这些研究并未考虑昼夜增温速率差异的影响.作为影响陆地生态系统碳汇能力的关键因素, 越来越多的研究开始关注昼夜不对称增温的影响(Wan et al., 2009; Peng et al., 2013; Tan et al., 2015; 赵杰等, 2018).例如, Peng等(2013)结合遥感数据和大气反演模型系统地分析了1982-2009年期间的昼夜不对称增温对北半球陆地生态系统碳汇通量的影响, 结果表明白天增温能够促进寒、温带湿润地区植被的生长, 但不利于温带干旱和半干旱(水分条件较差)地区植被的生长, 而夜间增温对植被生长的影响正好相反.同样, 赵杰等(2018)利用遥感数据在中国温带地区的研究发现植被活动对白天增温的响应比夜间增温更大, 且多为正向反应, 然而这种响应具有较大的季节异质性(Tan et al., 2015).此类研究仅关注了历史时期的昼夜不同速增温对植被动态的影响, 并未涉及不同增温情景影响的差异, 以及这种增温效应对植被后期生长、生存的影响.未来气候变暖持续加快, 将进一步加大昼夜增温幅度的差异, 温度的升高将引起饱和水汽压差(VPD)指数式的上升, 增加植物的水分压力(Grossiord et al., 2020), 尤其是在气孔导度相对较高的白天, 引起植被生长的停滞, 甚至死亡(McDowell & Allen, 2015; Hartmann et al., 2022; Gong & Hao, 2023), 进而改变未来陆地植被的分布及碳汇能力.因此, 探讨不对称增温对植被碳汇能力及植被对新环境适应能力的影响, 可提高对未来气候变化情景下陆地生态系统碳汇预测的准确性(Xia et al., 2014; 朱军涛和郑家禾, 2022). ... 中国温带昼夜增温的季节性变化及其对植被动态的影响 2 2018 ... 已有的增温模拟实验和模型结果均表明全球变暖能够通过促进光合作用而提高生态系统的碳固定量(Bai et al., 2013; Liang et al., 2013; Zhang et al., 2015; Du et al., 2020; Li et al., 2022), 然而, 这些研究并未考虑昼夜增温速率差异的影响.作为影响陆地生态系统碳汇能力的关键因素, 越来越多的研究开始关注昼夜不对称增温的影响(Wan et al., 2009; Peng et al., 2013; Tan et al., 2015; 赵杰等, 2018).例如, Peng等(2013)结合遥感数据和大气反演模型系统地分析了1982-2009年期间的昼夜不对称增温对北半球陆地生态系统碳汇通量的影响, 结果表明白天增温能够促进寒、温带湿润地区植被的生长, 但不利于温带干旱和半干旱(水分条件较差)地区植被的生长, 而夜间增温对植被生长的影响正好相反.同样, 赵杰等(2018)利用遥感数据在中国温带地区的研究发现植被活动对白天增温的响应比夜间增温更大, 且多为正向反应, 然而这种响应具有较大的季节异质性(Tan et al., 2015).此类研究仅关注了历史时期的昼夜不同速增温对植被动态的影响, 并未涉及不同增温情景影响的差异, 以及这种增温效应对植被后期生长、生存的影响.未来气候变暖持续加快, 将进一步加大昼夜增温幅度的差异, 温度的升高将引起饱和水汽压差(VPD)指数式的上升, 增加植物的水分压力(Grossiord et al., 2020), 尤其是在气孔导度相对较高的白天, 引起植被生长的停滞, 甚至死亡(McDowell & Allen, 2015; Hartmann et al., 2022; Gong & Hao, 2023), 进而改变未来陆地植被的分布及碳汇能力.因此, 探讨不对称增温对植被碳汇能力及植被对新环境适应能力的影响, 可提高对未来气候变化情景下陆地生态系统碳汇预测的准确性(Xia et al., 2014; 朱军涛和郑家禾, 2022). ...
本文的其它图/表
|


{kind=link}