


20世纪以来,生物科学已经逐渐成熟,可预测性逐渐增强,随着这种成熟的逐渐专业化,生物学研究方向的窄化现象不仅发生在生物组织的不同水平上(如,分子、个体、种群和生态系统),而且发生在特定的模式生物范围内(如,线虫(Anguina)、水蚤(Daphnia)、果蝇(Drosophila)及拟南芥(Arabidopsis)),或者特定的生境中(如,湖泊、森林、海洋和草地)(Elser et al., 2000b)。这种生物学知识的逐渐分化,使得不同领域的联系变得更加困难,从而阻碍了把整个生物界作为一个功能整体来进行研究(Allen & Hoekstra, 1992; Vogel, 1998)。目前,生物学家面临着发展综合性理论去研究生物系统的挑战,这种理论必须把我们已深刻了解的个体水平生物有机体联系起来,而且必须强调各种生物群体和生态系统的一致性特征(Michaels, 2003)。
地球上的所有生物是否具有统一的特征呢?是否可能在现代生物科学框架下提出一种理论去连接各种不同的组织水平(从基因的分子结构到生态系统)?要整合各种组织水平、有机体类型和各种生境的生物学知识确实是一个很大挑战。一个结合热力学第一定律、生物进化的自然选择原理和分子生物学中心法则的理论应运而生(Michaels, 2003),这一理论就是“生态化学计量学”(Ecological stoichiometry)或称为“化学计量生态学”(Stoichiometric ecology)。
生态化学计量学理论对于生物学研究具有重要意义,2004年第85卷第5期Ecology杂志刊登了“Ecological stoichiometry”特别专栏,介绍了欧美多个国家有关研究小组的最新研究进展。由此可见该杂志编辑部希望生态化学计量学这一新兴学科能够引起科学家们更广泛的关注。相对国际上对生态化学计量学研究十分活跃而言,生态化学计量学在中国目前尚没有受到广泛关注(张明霞,2003),鉴于此,本文介绍了生态化学计量学的概念、研究历史、理论及应用情况,期望对促进国内相关研究工作的开展起到一个抛砖引玉的作用。
1 生态化学计量学的概念
Stoichiometry是一个化学术语,它起源于希腊语的两个词汇“stoicheion”和“metron”,中文含义分别为“元素”和“测量”,因此,stoichiometry意为“测量元素的科学”,在化学中称为化学计量学。化学计量学是指研究参加化学反应的物质量或者是化学反应产物质量的科学,它最早由Richter在1792年提出(Sherman & Kuselman,1999),主要处理物质转换和化学反应中物质的质量间关系问题,重点强调物质的化学成分与结构关系,如化学反应的质量、原子量、分子量和百分比计算。作为化学反应方程中的系数,摩尔比(Molar ratio)在解决化学计量问题中占有中心位置。化学计量学主要原理包括:1)定比定律(Law of constant composition,也称作Law of definite proportions),其内涵为,每种化合物都有完全确定的组成;2)倍比定理(Law of multiple proportions),其内涵为,不同的元素化合时,这些元素的原子按简单整数比结合成化合物;3)质量守恒原理(Mass conservation principle),其内涵为,参加化学反应物质的原子种类和数量与化学反应产物的原子种类和数量保持不变。
生物世界都是由元素组成的,因此,生态学中也有化学计量规律,就像实验室的化学反应一样。生态交互作用无疑要涉及化学元素的重组,因此,有机体交互作用过程中化学元素的组合也要遵循质量守恒定律。从化学计量学角度来看,整个有机体就如一个单一的复杂分子(Sterner & Elser, 2002),分子内部和分子之间不断进行着化学重组。因此,从这一角度来说,当研究者想要了解某种生物的生物学特性时,无疑应首先确定该生物的化学组成特征。在这种意义下,每个有机体或生态系统能够被还原为一个简单的化学计量组成。尽管这种简化并没有实际作用,但可以引发我们产生一些思考:各种有机体是否存在一个固定的化学计量比值?不同有机体的化学元素组成是否相互影响?如果有机体所需要的其中一个组成元素不足,将会出现什么情况?而这种有机体会对生态系统产生什么影响(Michaels, 2003)?
Reiners是最早把化学计量学作为生态系统功能的一个补充模型的研究者之一,他认为化学计量学补充并延伸了生态能量学(Ecological energetics)(Reiners, 1986)。生态能量学自从Lindeman(1942)提出后,就一直是生态学中生物物理分析的主要模式。化学计量学思想在生态学中应用的关键就是实现生物实体(如,分子、细胞器、细胞和有机体等)能够根据它们的元素组成而加以明显区分,因为这些元素的组成差异是与重要生态功能相联系的(Elser et al., 1996),因此,生物体与生物体之间、生物体与非生物环境之间的相互作用方式不仅会受到相关生物体对元素需求的强烈影响,也会受到周围环境化学元素平衡状况的影响(Elser & Urabe, 1999)。
生态化学计量学理论认为有机体都是由元素构成的,这些元素的比值不仅决定了有机体的关键特征,也决定了有机体对资源数量和种类的需求(Schimel, 2003)。实际上,有机体的这种特征包含了3层涵义:第一,因为元素是有机体的最基本组成,所以生物进化将明显影响有机体的元素比值,因此,生态化学计量学理论可以直接与生物进化相联系;第二,环境对有机体元素比值的影响很大,不同的地质、气候和生物等因素都会影响它;第三,有机体通过消耗和释放不同于环境元素比值的元素,从而对其周围环境元素的比值产生影响。因此,环境的元素化学计量比值和有机体的化学计量比值之间就形成了复杂的反馈关系,一旦两者的化学计量比值不相匹配,就会引发有机体种群行为和进化的改变(Schimel, 2003),也会影响生物的生长发育过程和形态的改变(Méndez & Karlsson,2005)。可以认为,有机体的生态化学计量学理论连接了生物系统的3个尺度:分子水平(进化机理)、有机体水平和大尺度的生态系统过程。
生态化学计量学在研究元素动态和元素间关系的基础上建立一个共识,即有机体是由元素组成,而这些元素进一步组成了各种各样的有机化合物(还有一些无机骨架和金属螯合物),每种主要的化合物都具有自己独特的化学计量值。比如,碳水化合物含C和N丰富而含P少,核苷含N和P很高。在有机体水平,生物的化学组成可以反映进化的结果,这些进化的结果包括影响生物生长速率、健康状况、新陈代谢、结构和生态演替的某些方面(Michaels, 2003)。
活有机体的元素组成相对比较稳定,这种稳定性是通过它们积极调控细胞组成成分或体内环境来保持的。植物对其生物化学环境的选择具有更大弹性,因此,它们的元素组成也比动物和微生物变化更大。但是,所有有机体的化学计量值的差异比我们通常想象的要大,而且物种之间的变化可能与分类群间的变化一样大(Michaels, 2003)。总体上,植物无机养分的比值变化范围很大,而以植物或其它动物为食的动物养分元素组成变化范围较小。
为什么要在生态学中聚焦于化学元素呢?首先,元素容易在不同生物组织水平间移动,因此,能够计算元素的组成和估测进入或流出各种生物实体化学元素的通量;其次,元素是不变的,因此可以利用质量守恒原理去研究元素在各组织水平间的流动量;第三,有机体都是化学实体,都是由化学反应产生、维持和增殖的,尽管这些反应都是以很复杂和耦合的反应网形式进行的(Sterner & Elser, 2002)。
2 生态化学计量学研究历史
化学计量学理论在生态学上的应用可以追溯到1862年李比希提出的最小量定律(Liebig's law of the minimum),该理论认为在组成生物体的资源中,相对需求而言供应量最少的资源将成为限制该生物生长的主要因素,即有机体受到与需求量相比供应量最小资源的限制。该理论阐明的一个重要思想就是生物体的元素组成平衡对生物体生长是非常重要的。李比希认为化学在动物和植物生理学研究中具有重要地位,他的理论已经促成了一系列对有机体适应贫瘠环境的生态学和生理学研究,其中许多研究表明,限制性元素含量不同,有机体元素组成会产生相应变化。此后,Lotka(1925)在他的《物理生物学的基础》(Elements of Physical Biology)一书中,提出了捕食者-猎物相互作用模型,定量阐述了生物之间的相互作用关系。Lotka是第一个考虑物理-化学系统热力学定律与生物世界相联系的人,他的思想在生态学得到广泛应用并发展出了许多生态学基础理论:最佳取食理论(Optimal foraging)(Belovsky, 1978)、资源比理论(Resource-ratio theory)(Tilman, 1982)、海洋生物地球化学循环中的Redfield比值(Redfield, 1958)和养分利用效率(Vitousek, 1982)等。因此,也有人认为Lotka的理论是生态化学计量学理论的最早起源(Elser et al., 2000b)。尽管Redfield认识到有机体的化学计量组成总是不断变化的,但是,他对海洋中的无机养分比值和有机体的元素组成之间存在显著的一致性感到很惊奇,通过试验他确定了当养分不受到限制时浮游植物的元素比值关系为C:N:P = 106:16:1,这一比值关系被称为Redfield比值(Redfield ratio)(Redfield,1958; Redfield et al., 1963)。尽管后来证明C:N:P比值是不断变化的,但是,海洋浮游植物中C:N:P比值明显恒定性假设的提出,促成了海洋生物地球化学研究的极大发展。Tilman (1982)在Lotka-Volterra理论的基础上提出了资源比理论,这个理论认为单个物种的生长速率是限制性资源可利用量的函数,而资源的可利用量又是环境的资源供应速率和物种消耗速率的函数;资源在物种生长速率和物种间的竞争中起着关键的作用,因此,可以利用资源的流动和含量来预测物种的多度格局、物种共存和生物多样性。Vitousek(1982)提出了养分利用效率的概念,他把养分利用效率定义为,“植物损失单位养分造成的有机质从植物体损失的量或永久性储存单位养分而获得永久性储存在植物体内的有机质数量。”该概念可以简单地理解为单位养分的生产力,实际上它表述了碳(生物量)与养分的比值关系。养分利用效率提出后逐渐成为生态系统生态学的一个中心概念。
Reiners (1986)在其文章“Complementary models for ecosystems”中首次明确地把化学计量学理论作为生态系统研究的一个补充理论,从而使生态学与化学计量学有机结合起来。在该文中,Reiners阐明了化学计量学理论在生态学中的应用,并且提出了结合化学计量学理论的生态学研究理论模型。尽管作者并未直接使用生态化学计量学一词表示这一理论,但实际上该理论模型已经很清晰地阐明了生态化学计量学的基本理论框架。Reiners的生态化学计量学思想提出后,许多研究者,特别是海洋生物学家开始注意这一新的理论,大量的研究开始于20世纪90年代,这些研究验证并补充了生态化学计量学理论。以Sterner和Elser(2002)的著作《生态化学计量学:从分子到生物圈的元素生物学》一书的出版为标志,生态化学计量学理论基本得到完善,这一理论也逐渐被许多生物学家认同(Harris, 2003; Schindler, 2003)。著名生态学家Vitousek在该书前言中坦言,该书的出版是生态学研究史上的重要里程碑。Michaels(2003)认为生态化学计量学理论将不同尺度、不同生物群系和不同研究领域的生物学研究有机联系起来了。
3 生态化学计量学的基本理论
作为一门新学科,生态化学计量学的正式提出只有不到20年的历史。但是,由于化学计量学在生态学上的应用已有很长的历史,生态化学计量学理论的发展、完善及应用过程十分迅速。
图1
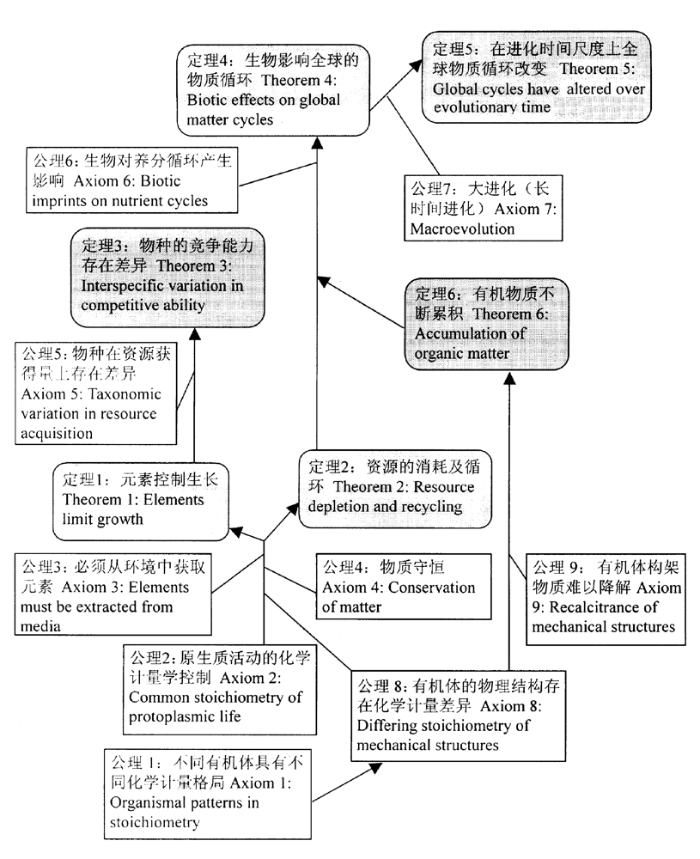
此外,生态化学计量学还发展了两个重要的具体理论,即动态平衡原理和生长速率理论。有机体化学元素组成的动态平衡原理是生态化学计量学理论成立的理论基础。正常的生物体的养分元素组成基本比较稳定,即使外界环境不断变化下,其组成也不会发生很大变化;但是,如果受到某种极端环境因子的影响,造成元素组成产生巨大变化,超出了其能够忍受的极限,这种有机体就可能无法生存,这就是生态学上的“Shelford耐受定律”在有机体元素组成上的一个反映。因此,有机体要生存必需具备一个负反馈机制,即,有机体内部环境(比如,消费者的元素计量比值)和外部环境(比如,食物的元素计量比)保持一种相对稳定的格局。这种有机体与其环境保持一种相对稳定的平衡状态的现象就被称为“动态平衡”(Homeostasis)。Kooijman(1995)以化学计量学的观点定义动态平衡为“使大多数有机体保持身体化学元素组成相对稳定的能力,尽管环境(包括有机体的食物)的化学元素组成发生变化”。生物学中的动态平衡就要使有机体内部的环境(如消费者的元素计量比等)不随外部环境(如资源的元素计量比等)的变化而剧烈变化,从而使整个有机体基本保持稳定,因此,动态平衡被认为是生命的本质特征。有机体能够控制它们自身的许多特性,包括养分平衡、pH值稳定等,从而在一定范围内维持其稳定性。在生态化学计量学中,有机体元素的动态平衡是指有机体中元素组成与它们周围环境(包括它们利用的资源)养分元素供应保持相对稳定的一种状态(Sterner & Elser,2002)。动态平衡使生物的化学物质组成产生不同程度的变化以及保持一定程度的稳定,因此,它是生态化学计量学存在的理论基础。
“生长速率理论”(Growth rate hypothesis, GRH)是有机体生态化学计量控制的基本途径,这个理论提供了生态化学计量控制生命进化、细胞生物学特性、种群动态和生态系统功能机制的基本框架(Reiners,1986; Elser et al., 2000b; Vanni et al., 2002)。这种与C:N:P化学计量比、RNA分配和有机体生活史相关的机制就被称为生长速率理论(Elseret al., 1996, 2000a)。GRH认为生物体必须改变它们的C:N:P比值以适应生长速率的改变。对许多异养生物来说,高的生长速率不仅对应高的N:C和P:C比,而且对应低的N:P比值(Elseret al., 2003; Makino et al., 2003)。GRH最初起源于对最适条件下,不同物种生物量C:P、N:P和最大生长速率的比较研究(Elseret al., 1996; Main et al., 1997),这种理论也适用于同种生物的生物量C:N和N:P比变化的研究(Hessen,1990),因为生长速率和RNA含量会随年龄增加而降低(Gorokhova & Kyle, 2002)。
生长速率理论把生物组织的不同层次(分子、个体、种群和生态系统)有机联系起来了。生长速率理论认为,单个有机体P的生物地球化学过程与生长和繁殖紧密相关(它是该种群动态的一个主要决定者)(Elser et al., 1996)。具体来说,有机体C:N:P比的变化主要是由生物体P含量的变化决定(Vanniet al., 2002)。生物体P的变化至少部分受不同生长或发育速率的有机体rRNA中P含量变化的影响(Elser et al., 1996)。有机体必须调整它们的C:N:P化学计量比来适应生长速率的改变,因此,生长快速的有机体通常具有较低的C:P和N:P比值(Elseret al., 2003; Makino et al., 2003)。这种较低的C:P和N:P比值反映了分配到rRNA中P的增加,因为核糖体需要快速地合成蛋白质以支持快速生长。这种直接或间接地造成生活史特性产生变化的演化过程会对有机体的元素组成产生影响(Elseret al., 2000b),因此,生长速率理论将细胞的物质分配与生物体的功能及其动态特征联系起来了。GRH假设已经在许多物种上得到验证,比如,异养细菌大肠杆菌(Escherichia coli)(Makino et al., 2003),以及浮游动物和陆地上的昆虫等(Elser et al., 2003)。但是,这些验证都是基于单个物种研究基础上获得的。Makino和Cotner(2004)通过多种细菌混合的实验表明,高C:P比(≥373)供应下的细菌群落,其RNA含量增加,生物量中P含量的增加与细菌生长速率成正比,正如GRH预测,高的生长速率造成低的生物量C:P和N:P比值;而当以低的C:P比(93:1)供应时,虽然细菌RNA含量仍然增加,但是,生物量C:P和N:P也随着生长速率的增加而增加,这种情况却不符合GRH理论的预测结果。因此,他认为只有在P限制的情况下(C:P比很高),由GRH预测的生态化学计量比值与生长速率的关系在生物群落和个体水平上都成立;而在不受P限制的情况下,细菌群落生长对养分供应的反应不符合GRH预测的结果,这可能是由于细菌群落的生长对养分计量比值的反应比单个有机体更加复杂的原因。
4 生态化学计量学的应用
自Reiners(1986)提出生态化学计量学思想以来,许多科学家对此开展了研究,生态化学计量学的应用领域不断拓展,目前生态化学计量学理论已经应用于包括营养动态(Sterner & Hessen, 1994; Andersen, 1997)、微生物营养(Tezuka 1989; Chrzanowski et al., 1996)、寄主-病原关系(Smith, 1993a)、共生(Smith, 1993b)、比较生态系统分析(Comparative ecosystem analysis)(Downing, 1997; Jaenike & Markow, 2003)、消费者驱动的养分循环(Sterner, 1990; Urabe et al., 1995; Elser & Urabe, 1999)、生物的养分限制(Koerselman & Meuleman, 1996; Aerts & Chapin, 2000; Tessier & Raynal, 2003)、碳循环(Lerman et al., 2000; Hessen et al., 2004)、种群动态(Urabe et al., 2002; Andersen et al., 2004)、森林演替与衰退(Wardle et al., 2004)和生态系统养分供应与需求的平衡(Vitousek, 2003; Schade et al., 2005)等研究,生态化学计量学理论不断得到丰富和验证。本文只重点介绍生态化学计量学理论在消费者驱动的养分循环、限制性养分元素判别以及全球C、N、P循环等方面的应用进展。
4.1 消费者驱动的养分循环
通过把生态化学计量学应用到消费者驱动的养分循环过程,生态学家逐渐认识到消费在控制诸如养分循环等生态系统过程中的作用。最近20年来,在海洋和陆地生态系统的养分循环和营养关系的理解上取得了很大进步。
消费者和它们食物的化学组成的差异能够影响消费者的取食行为(White, 1993)、消费者种群稳定性(White, 1993; Sterner & Hessen, 1994; Andersen, 1997)和群落组织结构(White, 1993; Andersen, 1997)。自养生物的养分含量变化很大,因此,其C:N:P比值变化也很大。相比之下,消费者的养分含量变化范围更小,通常维持一个比较严格的C:N:P比值。因此,当消费者和它们的食物之间的化学计量比值出现不平衡时,消费者的生长和活动将会受到影响(Loganet al., 2004; Hessen et al., 2004)。化学计量平衡也会对营养动态(Urabe & Sterner 1996; Sterner et al., 1998)和食物网中的生物地球化学循环(Andersen, 1997; Sterner et al., 1992; Urabe et al., 1995; Elser & Urabe, 1999)产生影响。许多研究表明(Stelzer & Lamberti, 2002; Ward & Cummins 1979; Elser et al., 2003),低的食物质量(高C:N或C:P比)将会显著降低消费者的生长速率。实验室模拟(Urabeet al., 1997)和原位试验(Elser et al., 2003)研究都表明,当喂以高C:P比的藻类时,水蚤的生长受到P限制。Frost和Elser(2002)通过控制蜉蝣(Mayfly)的食物及其C:P比值试验发现,供应食物量不足会造成蜉蝣的生长速率下降;而在食物供应充足的情况下,喂以低C:P比值的食物时蜉蝣生长更快,并且蜉蝣的P含量与其食物的P含量正相关。Sterner和George(2000)研究了海洋中鲤科中的几种鱼的C:N:P比值及其与食物(海藻)的C:N:P比值之间的关系,探讨了通过鱼驱动的食物链的养分循环状况,结果发现,鲤鱼的N:P比值与它们的食物的N:P比值相似;不同湖泊的鲤鱼养分含量变化很小,尽管食物的养分含量变化很大;C:N:P比值稳定性的维持,是由鲤鱼对养分的平衡吸收而不是由鲤鱼对食物的消化速率决定的。
生态化学计量学理论在陆地生态系统消费者驱动的养分循环研究中的应用相对较少。GRH理论认为以含P量丰富的物质为食物的有机体具有更高的P和RNA含量,以及更快的生长速率。这种机制暗示一种潜在的因果关系链,即土壤有效P含量影响植物生长和组织P含量,然后进一步影响取食者的P和核酸含量,生长和繁殖以及种群动态。Schade等(2003)研究了一个沙地生态系统土壤中有效P的时空分布对一种豆科灌木(Prosopis velutina)和它的一种主要昆虫类取食者(Sabinia setosa)的化学计量组成的影响。结果表明,这种灌木叶片的C:P比值受到了土壤有效P时空分布变化的影响,两者呈负相关;正如GRH理论预测,取食者P和RNA含量能够反映土壤有效P和这种灌木叶片的C:P比值,而且这种取食者的种群数量也与植物叶片的C:P比成反比。Elser等(2000a)通过比较陆地植物叶片的元素比值与昆虫取食的关系,表明这些陆地植物的平均C:P和N:P比都高于这些植物取食者的相应比值,这有可能是因为植物通过低的P含量来限制取食者的种群发展。
4.2 限制性元素的判断
养分的可利用性/供应量能够影响许多陆地、水生和海洋生态系统的生产力、生物组成、生物多样性、物种动态,以及植物、动物和微生物种群间的交互作用(Vitousek, 2004),养分的供应量是否充足是影响有机体生长、种群结构、物种相互作用和生态系统稳定性的重要因素,因此,对这种供应量不足的养分类型的判断就成为维护这个系统稳定的前提。生态化学计量学涉及到消费者和它们的食物的元素比率(C:N、C:P和N:P比),并且认为相对需求量来说供应量最少的元素就是“限制性”元素。同时,生态化学计量学理论认为有机体的元素组成比值是动态稳定的,有机体存在一个相对稳定的C、N、P元素比值关系,而其中任何一种元素的巨大变化都将使这一比值发生变化。因此,通过C:N:P比值的变化,该理论可以用来判断限制有机体生长、发育或繁殖的元素类型。许多研究也表明生态计量比值能够有效反映出限制性元素的种类(Andersen & Hessen,1991; Koerselman & Meuleman, 1996; Güsewell,2004; Ågren, 2004)。有机物、食物网和生态系统中储存的C在一定程度上是由质量守恒原理和其它关键养分元素(如N、P等)的可获得量控制的。养分限制意味着C的相对过量(如,C:N、C:P上升),同时,也意味着自养和异养生物的C汇可能不同。总体上,C与养分比值通常较高且变化灵活的自养生物和分解者,与具有较低C与养分比值和生态化学控制较严(变化幅度小)的消费者的C:N:P比值差异较大。因此,食物网中的C利用效率可能主要由自养生物的养分元素控制,当食物(自养生物)中的C与养分比值达到那些消费者的C与养分比值时,C利用效率将会增加(Hessenet al., 2004)。
化学计量比值的变化能够改变植食动物之间的养分竞争,从而改变自养生物养分限制的种类。在研究海洋和湖泊等水域生态系统过程中,对于海洋水域中限制性元素种类的判断经历了很长的争论。早期,海洋生态学家(如,Boynton et al., 1982; Smith, 1984)认为海洋中N是主要的限制性元素,因此,他们主要是定量研究浮游动物的N循环过程;而另一些生态学家认为,由于海洋可以通过对大气N2的固定来保证N的供应,因此,P是海洋中最短缺的元素(Meybeck, 1982)。这一时期,海洋生态学家除了考虑浮游动物对海洋初级生产力的影响,几乎没有考虑过浮游动物造成的N和P养分的再生以及这种再生的重要意义(Downing, 1997)。
但是,20世纪80年代,Lehman(1984)的研究指出,浮游动物并不是以同样的效率循环利用N和P,这可能造成浮游动物通过改变海洋生态系统的N:P比值供应而影响浮游植物群落。Elser等(1988)研究发现,由于鱼类取食产生变化,浮游动物群落的组成从水蚤(Daphnia)占优势转变为桡足类动物(Calanoid copepod)占优势,这个过程同时伴随着限制浮游植物生长的养分元素类型的变化。在水蚤占优势的情况下,浮游植物的生长受到P的限制,而在桡足类动物占优势的情况下浮游植物的生长就会受到N的限制。Andersen和Hessen(1991)的研究表明,与水蚤相比,桡足类动物能够稍微提高N含量,但却能够大量降低P含量。因此,水蚤具有较低的N:P比值(约为14:1),而大多数桡足类动物的N:P比值较高(约为30~50:1)。Sterner等(1992)总结Elser等(1988)的报道认为,这是消费者驱动的化学计量改变的结果。水蚤占优势的浮游动物的聚集不同程度地造成动物生物量中积累了大量的P,而N却以一种相对较高的速率进行循环,因此,使浮游植物的生长朝着P限制的方向发展。当由桡脚类动物占优势造成的浮游动物聚集时,浮游动物群落积累大量N,而P的循环利用速率很快,从而导致浮游植物的生长受到N的限制。
在陆地生态系统中,土壤细菌的C:N比值一般为6,而植物组织和凋落物的C:N比值通常大于100。因此,Vitousek等(2002)认为,在陆地生态系统中,相对于它们的需求量,动物和微生物都是生存于一个C含量丰富而N不足的世界中。动物的营养和生长经常受它们的食物N含量的限制,造成动物体内蛋白质含量不足(White, 1993)。微生物在分解过程中也会受到凋落物中N含量的影响,为了自身的繁殖和生长,微生物会保留大部分从分解基质中获得的N,并且也直接从土壤中吸收无机N加以补充,N从有机物转化到生物可利用状态的过程落后于植物凋落物的分解过程,因此,植物往往容易受到N不足的限制。生物固N是一个非常耗能的过程,需要消耗大量的ATP(含P)。许多研究也表明,在陆地和海洋生态系统中,藻青菌类(Cyanobacterial systems)微生物受到P的供应不足的限制,从而抑制N的固定(Smith, 1993a; Crews, 1993a, 1993b)。最近一项研究也发现,在长期缺乏灾难性干扰的情况下,森林生态系统原始演替或次生演替的后期阶段,常常伴随着生产力的下降以及腐殖质和新鲜凋落物中N:P比的增加,表明随着时间推移森林生态系统越来越受到P的限制(Wardleet al., 2004)。
植物叶片的N:P临界比值被认为可以作为判断环境对植物生长的养分供应状况的指标(Wassenet al., 1995; Aerts & Chapin, 2000; Güsewell,2004)。Sakamoto(1966)发现当海藻N:P比值在10~17范围内时,N和P对叶绿素的生产量的作用几乎相同(N、P同时限制或都不限制生产量),当N:P比小于10时叶绿素的生产量仅受N的限制,而当N:P比值大于17时,叶绿素的产生量只受P的限制。Wassen等(1995)、Koerselman和Meuleman(1996)通过对湿地植被施肥试验表明,N:P比大于16时,可以认为这个系统是受P限制的;而当N:P比小于14时,这一生态系统被认为是受N限制的。在N:P比为14~16范围内,被认为是受到2种元素共同限制或2种元素都不缺少。Braakhekke和Hooftman(1999)通过对土壤养分贫瘠的草地进行施肥试验,认为植物叶片N或P不足的N:P比临界值分别为10和14。结合叶片中N和P含量大小,Braakhekke和Hooftman(1999)认为N:P比大于14而植物叶片P含量低于1.0 mg·g-1时,这个系统是受P限制的;而当N:P比小于10和植物叶片N含量低于20.0 mg·g-1时,这一生态系统被认为是受N限制的。N:P比在10~14范围内的被认为是受到2种元素共同限制(当P < 1.0 mg·g-1和N < 20.0 mg·g -1时)或两种元素都不缺少(当P > 1.0 mg·g -1和N > 20.0 mg·g -1时)。当然,由于研究区域、植物器官及植物种类(种间和种内)的差异,N:P临界指标也会发生很大变化(Güsewell,2004)。
4.3 全球C、N、P生物地球化学循环
元素在全球生物地球化学循环中并不是单个进行的,元素之间存在相互作用。在所有元素中,C代表了地球上所有生命形式的通用媒介,许多元素之间的交互作用都是通过C与其它养分的比值关系来进行调节的(Melillo et al., 2003)。因此,生物的C:N:P化学计量比对全球C和养分循环具有重要影响,生物量中C与养分比值的差异能够调控生态系统中有机物质的去处,影响生物圈中C的消耗或固定过程。目前,人类活动对生物地球化学循环的影响逐渐加剧,而人类活动对全球C循环的影响与自然界本身的C循环过程完全不同,因此,必须了解生物群落中C、P、N的耦合关系,以更好地预测人类干扰对区域生态系统和生物圈的影响(Sterner & Elser,2002)。
目前,全球环境和气候变化对N和P在生物地球化学C循环中的控制性作用的关系存在两个问题:1)陆地上有多少新的初级生产量可以归功于土壤有机物质的分解和化肥的施用?2)海洋中有多少新的初级生产量是由陆地运输到海洋的养分和由深海运输到海洋表层而产生的?通过C:N:P化学计量关系的确立,生态化学计量学为这些问题的回答提供了契机(Lermanet al., 2000)。
海洋的生产量占整个生物圈生产量的50%左右,它是大量有机物质长期储存的地方,因此,在很大程度上海洋能够影响全球气候。“生物泵”(Biological pump)指的是海洋中C从表层向深层净移动的过程,这个过程能够导致表层水域生产者对CO2的同化和表层生产者固定的C往温跃层以下运输(Lalli & Parsons, 1997),结果促进了海洋对CO2的吸收,使C深埋于海洋底层沉积物中。元素化学计量比从两个方面影响生物泵功能的发挥(Falkowski et al., 2000),一个是上层海水中溶解养分(N和P)的利用效率;另外一个就是初级生产中形成的有机物质的C:N:P化学计量值,只有在表层生产的物质C与养分比值大于从中层进入表层的物质C与养分比值情况下,生物泵才能使C净储存于深海(Karl,1999)。整体上,海洋中C:N:P比值是比较平衡的(约等于Redfield比值)。但是,由于世界海洋面积约占整个地球的70%,如果海洋中的C:N:P比值发生哪怕一个微小的变化都可能会影响全球的碳循环。海洋中的可溶性有机质(Dissolved organic matter)是生物圈中最大的碳库之一,Hopkinson和Vallino(2005)通过总结近10年来的世界几大海洋的研究,发现海洋中的可溶性有机质是以一种C:N:P=199:20:1的比值进行矿化的,这个比值显著高于Redfield比值(106:16:1),整个海洋的C:N和C:P比值表现为一种C丰富的状态,即C分别比Redfield比值高出62%和87%。因此,如果进入到了海洋表层水体中的N和P增加,那么就将有更多的可溶性有机碳进入海洋中。对厄尔尼诺引起的南方涛动(ENSO)研究表明,在ENSO期间,海水增暖造成海洋表层混合层变窄和垂直的养分供应量减少,海洋生物量的C与养分比值增加,可能原因是混合层的窄化使得表层水域光/养分比值增加,从而减缓了表层的光限制,加剧了养分的限制(Karl,1999)。
人类活动正在史无前例地改变全球生物地球化学循环(Falkowski et al., 2000),这些改变大部分是与人类的食物生产、城市化、工业化和对水分过度利用等活动有关。陆地与水域生物地球化学循环的改变并不是单个元素循环的变化,而是多种元素共同变化的,自然界元素间的相互关系也受到人类活动的改变(Melillo et al., 2003)。而自然界本身的C、N、P生物地球化学循环的耦合作用已经很复杂,因此,如何定量评价人类的影响?人类改变后的全球C:N:P计量比值是多少?应用生态化学计量学理论能够部分回答这些问题。人类对全球C、N、P的影响主要通过生物量生产(主要通过农业)和能量消耗两个方面,人类活动,诸如化石燃料燃烧、土地利用变化和工业化等造成了大量CO2释放(Hoffert et al., 1998; Austin et al., 2003);化肥的生产、化石燃料的燃烧和固氮作物的推广所造成的N输入远大于自然系统对N的固定量(Galloway & Cowling, 2002);而矿石的开采、化肥的施用以及工业上P的生产所造成的P输入也远超过自然界中由岩石风化产生的P(Bennett et al., 2001)。Falkowski等(2000)研究发现,人类活动使C、N、P通量分别增加了13%、108%和400%,自然过程的全球C:N:P计量比约为20 333:43:1,人类活动造成的C:N:P计量比约为667:12:1(表1)。从C:N:P计量比的对比可以看出,人类对N和P循环的影响强于对C循环的影响,与C相比人类对全球生态系统的影响主要是增加了N和P。由于人类的这种大规模的“富营养化”活动,整体上,地球上的总体生物群的养分利用效率(单位养分的生物量生产能力)可能出现了下降。
表1 人类活动对C、N、P全球生物地球化学循环的影响(Falkowski et al., 2000)
Table 1
| 元素 Element | 产生来源 Sources of flux | 产生大小 Magnitude of flux (109 kg·a-1) | 人类的影响 The change due to human activities (%) | |
|---|---|---|---|---|
| 自然 Natural | 人类 Anthropogenic | |||
| C | 陆地生物呼吸和分解Terrestrial respiration and decay CO2 | 61 000 | 8 000 | +13 |
| 化石燃料燃烧和土地利用改变Fossil fuel and land use CO2 | ||||
| N | 自然界生物固氮Natural biological fixation | 130 | 140 | +108 |
| 农作物、化石燃料燃烧和施肥Fixation owing to rice cultivation, combustion of fossil fuels, and production of fertilizer | ||||
| P | 风化Chemical weathering | 3 | 12 | +400 |
| 采矿Mining | ||||
在陆地生态系统C、N、P循环研究中,McGroddy等(2004)通过总结世界范围内森林生态系统的研究结果发现,不同的生物群(温带阔叶林、温带针叶林和热带森林)具有不同的C:N:P比值,而对于整个全球森林生态系统来说,树木叶片的C、N、P原子比率为1 212:28:1,森林凋落物的C、N、P原子比率为3 007:45:1。树木叶片和凋落物的C:N:P比值表明,陆地生态系统植被的C含量相对于N和P要高于淡水自养生物(淡水生态系统自养生物的C:N:P比值约为306:30:1 (Elseret al., 2000a)),但两类生态系统的N:P比值相似。Ågren (2004)对一种水生植物和一种陆生植物的N:P比值进行比较后,也发现水生植物和陆生植物的N:P比值基本相似,而两者之间的微小差异可能主要来源于水生植物比陆生植物具有更大的最大相对生长速率,而且前者对养分的奢侈吸收能力更强。这些研究表明,两类生态系统中自养生物光合器官的N:P比值可能都受到相似生理过程的控制。Jackson等(1997)的研究也发现,全球植被细根的C:N:P比值为1 158:24:1,与树木叶片的C:N:P比值相近。凋落叶片的C与养分比值大于叶片的C与养分比值,表明全球尺度上养分再吸收(特别是对P的再吸收)是陆地森林植被的一个重要机制。Vitousek(2003)通过对夏威夷不同土壤发育阶段森林生态系统主要组成树种的不同组织C:N、N:P比值的研究发现,桃金娘(Metrosideros polymorpha)花、叶片、叶片凋落物、根系和木质部分的C:N比值在不同生态系统中差异很大,而N:P比值基本相似;各发育阶段的不同树种对应组织的C:N比值差异也很大,而N:P比值也基本相似,表现出N:P比值比较恒定,而C:N比值变化较大。Vitousek(2003)认为化学计量的(Stoichiometric)和弹性的(Flexible)生态交互作用共同控制着生物对元素的需求和供应,而这两种作用的共存也是导致陆地生态系统复杂性的因素之一。McGroddy等(2004)和Vitousek(2003)等的工作已经为陆地生态系统的研究提供了一定的基础,但是,陆地生态系统的复杂性决定了目前的研究还远远不够,有待于进一步加强。
5 结论
化学元素是生物体最本质的组成成分,它能够对有机体的许多行为进行有序调控,生态化学计量学理论被认为在所有生物系统和全球生物地球化学循环研究中具有重要价值(Michaels, 2003),但是,生态化学计量学并不是万能的,它至少在目前还不能回答太多生物学问题。由于受到研究区域集中(海洋生物、微生物相对较多)、研究物种数少(检验理论的物种数目有限)和理论提出时间短等限制,以及部分研究者对一些物种的研究未能验证生态化学计量学理论,关于生态化学计量学的研究还远远不够,生态化学计量学理论是否适用于全部生物还有待于进一步检验。目前,生态化学计量学是生态学研究的一个热点,发展前景十分诱人。今后这方面的研究将可能主要集中于以下几个方面:
1)陆地生态系统生态化学计量
生态化学计量学理论的提出、理论的验证和丰富大都是基于对海洋生物的研究获得的,而对于陆地生物的研究相对很少,这可能是因为陆地生态系统生物地球化学循环的周转速率很慢,而且,陆地生态系统中富含各种难以分解的含C化学物质,以及影响C:N:P计量比值的因素更多,这就使陆地生态系统比海洋生态系统更为复杂。因此,这一理论还需要在对陆地生物进行广泛研究的基础上得以最终验证和确立。
2)养分胁迫下生物生长发育的适应机制
生物对养分胁迫的响应一直是生态学研究的热点问题之一,胁迫响应机制的探讨从分子、细胞、生理生态再到种群动态层次都有,但问题似乎仍未解决,生态化学计量学理论的应用有望在这一问题上获得更多确定的答案。
3)利用生态化学计量学理论解释生命现象
生态化学计量学理论认为,生物的生命形式可以简化成简单的元素比值,因此,这一简化有可能促进对生物起源、进化、生命维持以及生态系统水平养分供应与需求平衡机制的探索。
4)全球C、N、P循环
CO2是重要的温室气体,C循环是生态学领域的一个热点问题,全球C“失汇”及全球C循环的稳定性如何维持仍然困扰着生态学家,而生态化学计量学的提出可以阐明海洋和陆地植被对C的吸收及释放机制。这种释放与吸收机制一旦研究清楚,将有利于人类了解甚至是控制全球C循环。目前已经有少部分研究表明了生态化学计量学理论在全球C循环研究中的重要作用,因此,这方面的研究有待于进一步拓展。同样,与C循环关系密切的N、P循环也将是今后研究的重点。在全球生物地球化学研究中有许多急需解决的问题需要利用生态化学计量学理论去解决。可以预期,生态化学计量学将继续得到更蓬勃的发展。
参考文献
The mineral nutrition of wild plants revisited: a re-evaluation of processes and patterns
The C:N:P stoichiometry of autotrophs-theory and observations
Stoichiometry and population dynamics
Carbon, nitrogen, and phosphorus content of freshwater zooplankton
Human disruption of element interactions: drivers, consequences, and trends for the twenty-first century
In: Melillo JM, Field CB, Moldan B eds.
Diet optimization in a generalist herbivore: the moose
Human impact on erodable phosphorus and eutrophication: a global perspective
A comparative analysis of nutrients and other factors influencing estuarine phytoplankton production
In: Kennedy VS ed.
The resource balance hypothesis of plant species diversity in grassland
Carbon, nitrogen and phosphorus budgets for the mixotrophic phytoflagellate Poterioochromonas malhamensis (Chrysophyceae) during bacterial ingestion
Foliar and surface soil nutrient concentrations of Pinus spp. plantations in relation to species and stand age in Zhanggutai sandy land, northeast China
Element ratios and growth dynamics of bacteria in an oligotrophic Canadian shield lake
Nutrient limitation to nitrogen fixation in young volcanic sites
Phosphorus regulation of nitrogen fixation in a traditional Mexican agroecosystem
Marine nitrogen: phosphorus stoichiometry and the global N:P cycle
Growth rate—stoichiometry couplings in diverse biota
Organism size, life history, and N:P stoichiometry: towards a unified view of cellular and ecosystem processes
Zooplankton-mediated transitions between N and P limited algal growth
Nutritional constraints in terrestrial and freshwater food webs

Biological and environmental contrasts between aquatic and terrestrial systems have hindered analyses of community and ecosystem structure across Earth's diverse habitats. Ecological stoichiometry provides an integrative approach for such analyses, as all organisms are composed of the same major elements (C, N, P) whose balance affects production, nutrient cycling, and food-web dynamics. Here we show both similarities and differences in the C:N:P ratios of primary producers (autotrophs) and invertebrate primary consumers (herbivores) across habitats. Terrestrial food webs are built on an extremely nutrient-poor autotroph base with C:P and C:N ratios higher than in lake particulate matter, although the N:P ratios are nearly identical. Terrestrial herbivores (insects) and their freshwater counterparts (zooplankton) are nutrient-rich and indistinguishable in C:N:P stoichiometry. In both lakes and terrestrial systems, herbivores should have low growth efficiencies (10-30%) when consuming autotrophs with typical carbon-to-nutrient ratios. These stoichiometric constraints on herbivore growth appear to be qualitatively similar and widespread in both environments.
Biological stoichiometry from genes to ecosystems
The stoichiometry of consumer-driven nutrient recycling: theory, observations, and consequences
The global carbon cycle: a test of our knowledge of Earth as a system
Growth responses of littoral mayflies to the phosphorus content of their food
Reactive nitrogen and the world: 200 years of change
Analysis of nucleic acids in Daphnia: development of methods and ontogenetic variations in RNA-DNA content
N:P ratios in terrestrial plants: variation and functional significance
Book review: ecological stoichiometry: biology of elements from molecules to the biosphere
Carbon, nitrogen and phosphorus status in Daphnia at varying food conditions
Carbon sequestration in ecosystems: the role of stoichiometry
Energy implications of future stabilization of atmospheric CO 2 content
Efficient export of carbon to the deep ocean through dissolved organic matter
Oceanic dissolved organic carbon (DOC) constitutes one of the largest pools of reduced carbon in the biosphere. Estimated DOC export from the surface ocean represents 20% of total organic carbon flux to the deep ocean, which constitutes a primary control on atmospheric carbon dioxide levels. DOC is the carbon component of dissolved organic matter (DOM) and an accurate quantification of DOM pools, fluxes and their controls is therefore critical to understanding oceanic carbon cycling. DOC export is directly coupled with dissolved organic nitrogen and phosphorus export. However, the C:N:P stoichiometry (by atoms) of DOM dynamics is poorly understood. Here we study the stoichiometry of the DOM pool and of DOM decomposition in continental shelf, continental slope and central ocean gyre environments. We find that DOM is remineralized and produced with a C:N:P stoichiometry of 199:20:1 that is substantially lower than for bulk pools (typically >775:54:1), but greater than for particulate organic matter (106:16:1--the Redfield ratio). Thus for a given mass of new N and P introduced into surface water, more DOC can be exported than would occur at the Redfield ratio. This may contribute to the excess respiration estimated to occur in the interior ocean. Our results place an explicit constraint on global carbon export and elemental balance via advective pathways.
A global budget for fine root biomass, surface area, and nutrient contents
A sea of change: biogeochemical variability in the North Pacific Subtropical Gyre
The vegetation N:P ratio: a new tool to detect the nature of nutrient limitation
The stoichiometry of animal energetics
and their impacts on the structure of phytoplankton communities
In: Meyers DG, Strickler JR eds. Trophic Dynamics Within Aquatic Ecosystems.
Nitrogen and phosphorus controls of the carbon cycle
Control of CNP homeostasis in herbivore consumers through differential assimilation
Stoichiometric analysis recognizes that a herbivore is a mixture of multiple chemical elements, especially C, N, and P, that are fixed in various proportions. In the face of a variable quality food supply, herbivores must regulate ingested nutrients to maintain a homeostatic state. We develop a dynamic mathematical model, based on differential assimilation, that controls the C:N and C:P ratios in a herbivore within given tolerance ranges; the actual mathematical mechanism is to define the absorption coefficients to be dependent on these elemental ratios. The model inputs variable, time-dependent food and grazing rates and calculates the net dynamic consumer production (growth) of C, N, and P. It shows that total growth is equally sensitive to variations in both food quality and in assimilation rates, and it shows quantitatively how C-, N-, or P-limited growth could occur at different periods during the development period. The analysis generalizes earlier work limited to just two elements.
Differential mechanical force distributions are increasingly recognized to provide important feedback into the control of an organ's final size and shape. As a second messenger that integrates and relays mechanical information to the cell, calcium ions (Ca(2+)) are a prime candidate for providing important information on both the overall mechanical state of the tissue and resulting behavior at the individual-cell level during development. Still, how the spatiotemporal properties of Ca(2+) transients reflect the underlying mechanical characteristics of tissues is still poorly understood. Here we use an established model system of an epithelial tissue, the Drosophila wing imaginal disc, to investigate how tissue properties impact the propagation of Ca(2+) transients induced by laser ablation. The resulting intercellular Ca(2+) flash is found to be mediated by inositol 1,4,5-trisphosphate and depends on gap junction communication. Further, we find that intercellular Ca(2+) transients show spatially non-uniform characteristics across the proximal-distal axis of the larval wing imaginal disc, which exhibit a gradient in cell size and anisotropy. A computational model of Ca(2+) transients is employed to identify the principle factors explaining the spatiotemporal patterning dynamics of intercellular Ca(2+) flashes. The relative Ca(2+) flash anisotropy is principally explained by local cell shape anisotropy. Further, Ca(2+) velocities are relatively uniform throughout the wing disc, irrespective of cell size or anisotropy. This can be explained by the opposing effects of cell diameter and cell elongation on intercellular Ca(2+) propagation. Thus, intercellular Ca(2+) transients follow lines of mechanical tension at velocities that are largely independent of tissue heterogeneity and reflect the mechanical state of the underlying tissue.
N:P stoichiometry and ontogeny of crustacean zooplankton: a test of the growth rate hypothesis
Elemental stoichiometry of a heterotrophic bacterial community in a freshwater lake: implications for growth- and resource-dependent variations
Are bacteria more like animals than plants? Growth rate and resource dependence of bacterial C:N:P stoichiometry
Elemental stoichiometry of Drosophila and their hosts
Scaling of C:N:P stoichiometry in forests worldwide: implications of terrestrial Redfield-type ratios
Nutrient stoichiometry in Pinguicula vulgaris: nutrient availability, plant size, and reproductive status
Carbon, nitrogen, and phosphorus transport by world rivers
return of nitrogen to the soil
.In: Paul EA, Clark FE eds.
The biological control of chemical factors in the environment
The influence of organisms on the composition of seawater
In: Hill MN ed. The Sea Vol. 2.
Primary production by phytoplankton community in some Japanese lakes and its dependence on lake depth
A conceptual framework for ecosystem stoichiometry: balancing resource supply and demand
Stoichiometric tracking of soil nutrients by a desert insect herbivore
Stoichiometry and chemical metrology: Karl Fisher reaction
Phosphorus versus nitrogen limitation in the marine environment
Applicability of resource-ratio theory to microbial ecology
Implications of resource-ratio theory for microbial ecology
Ecological stoichiometry in running waters: periphyton chemical composition and snail growth
The ratio of nitrogen to phosphorus resupplied by herbivores: zooplankton and the algal competitive arena
Carbon: phosphorus stoichiometry and food chain production
Stoichiometric relationships among producers and consumers in food webs
Carbon, nitrogen, and phosphorus stoichiometry of cyprinid fishes
Algal nutrient limitation and the nutrition of aquatic herbivores
Use of nitrogen to phosphorus ratios in plant tissue as an indicator of nutrient limitation and nitrogen saturation
The C:N:P ratios of phytoplankton determine the relative amounts of dissolved inorganic nitrogen and phosphorus released during aerobic decomposition
Reduced light increases herbivore production due to stoichiometric effects of light: nutrient balance
Contribution of metazoan plankton to the cycling of N and P in Lake Biwa
Regulation of herbivore growth by the balance of light and nutrients
Phosphorus-limitation of Daphnia growth: is it real?
Stoichiometry of nutrient recycling by vertebrates in a tropical stream: linking biodiversity and ecosystem function
Nutrient cycling and nutrient use efficiency
Stoichiometry and flexibility in the Hawaiian model system
In: Melillo JM, Field CB, Moldan B eds.
Nitrogen and nature
Anthropogenic changes to the global N cycle are important in part because added N alters the composition, productivity, and other properties of many natural ecosystems substantially. Why does added N have such a large impact? Why is N in short supply in so many natural ecosystems? Processes that slow the cycling of N relative to other elements and processes that control ecosystem-level inputs and outputs of N could cause N supply to limit the dynamics of ecosystems. We discuss stoichiometric differences between terrestrial plants and other organisms, the abundance of protein-precipitating plant defenses, and the nature of the C-N bond in soil organic matter as factors that can slow N cycling. For inputs, the energetic costs of N fixation and their consequences, the supply of nutrients other than N, and preferential grazing on N-fixers all could constrain the abundance and/or activity of biological N-fixers. Together these processes drive and sustain N limitation in many natural terrestrial ecosystems.
Effects of food quality on growth of a stream detritivore,Paraendipes albimanus (Meigen)(Diptera: Chironomidae)
Ecosystem properties and forest decline in contrasting long-term chronosequences
During succession, ecosystem development occurs; but in the long-term absence of catastrophic disturbance, a decline phase eventually follows. We studied six long-term chronosequences, in Australia, Sweden, Alaska, Hawaii, and New Zealand; for each, the decline phase was associated with a reduction in tree basal area and an increase in the substrate nitrogen-to-phosphorus ratio, indicating increasing phosphorus limitation over time. These changes were often associated with reductions in litter decomposition rates, phosphorus release from litter, and biomass and activity of decomposer microbes. Our findings suggest that the maximal biomass phase reached during succession cannot be maintained in the long-term absence of major disturbance, and that similar patterns of decline occur in forested ecosystems spanning the tropical, temperate, and boreal zones.
Nutrient concentrations in mire vegetation as a measure of nutrient limitation in mire ecosystems
Application of N:P stoichiometry to ecology studies
Differential responses of N:P stoichiometry ofLeymus chinensis and Carex korshinskyi to N additions in a steppe ecosystem in Nei Mongol

{kind=link}
{kind=link}