


为了更深入地了解森林生物多样性的分布格局、形成与维持的机制,更好地进行森林生物多样性监测,1975年一些著名生态学家提出建立大样地的构思,并在BCI(Barro Colorado Island, Panama)进行了尝试(Condit,1995)。至今,美国Smithsonian研究院CTFS(the Center for Tropical Forest Science)已在全球热带地区的14个国家建立了18个森林生物多样性监测大样地(4~52 hm2),形成了热带森林生物多样性监测网络(
大样地的建设有效监测了物种的时空分布模式,为研究物种多样性的维持机制、物种空间分布格局、群落动态等提供了重要的研究平台,并取得了热带森林生物多样性形成与维持研究的大量成果(Condit,1995; Hubbell et al., 1999; Condit et al., 2002; Hooper et al., 2004; Condit et al.,2006; Hubbell,2006)。这些研究成果对物种共存的经典生态位理论提出质疑,创立了群落生态学的中性理论(Neutral theory)(Hubbell,2001),对热带森林物种多样性形成与维持作出了很好的解释。中性理论以物种间在个体水平上的对等性为前提,强调随机过程的重要性,第一次从基本生态学过程(出生、死亡、迁移、物种分化)出发,为在个体水平上探讨群落结构形成搭建了理论框架(Chave,2004),在生态学界引起了极大的反响(McGill, 2003; Volkov et al., 2003; Chase,2005;Condit et al.,2006;McGill et al.,2006)。
中国地跨热带到寒温带,是世界上生物多样性最丰富的国家之一,有必要借鉴CTFS大样地建设的思路和方法,进行森林生物多样性形成维持的研究。为此,中国科学院生物多样性委员会组织建立了中国森林生物多样性监测网络,分别在温带地区的长白山(Changbaishan,CBS)、中亚热带地区的古田山(Gutianshan,GTS)、南亚热带的鼎湖山(Dinghushan,DHS)和热带地区的西双版纳(Xishuangbanna,XBN)建立了20~25 hm2的固定监测大样地。本文报道鼎湖山南亚热带常绿阔叶林大样地(简称鼎湖山大样地,20 hm2 lower subtropical evergreen broadleaved forest plot in Dinghushan,DHS plot)的区系特征、物种组成、垂直结构、径级结构与空间分布等群落特征。
1 研究区、样地概况与研究方法
1.1 研究区概况
鼎湖山国家级自然保护区位于广东省肇庆市鼎湖区(23°09'21″~23°11'30″ N,112°30'39″~112°33'41″ E),面积为1 155 hm2,属于低山丘陵,最高峰鸡笼山海拔1 000.3 m,山体陡峭,坡度多在30°~45°之间。
该地区属南亚热带湿润季风气候,冬夏气候交替明显,年平均气温为20.9 ℃,最热月(7月)平均气温为28.0 ℃,最冷月(1月)平均气温为12.6 ℃,极端最高气温为 38.0 ℃,极端最低气温为-0.2 ℃。平均每年有 4 次霜冻,每次持续 1~2 d。1975~1995年年均降雨量为1 985 mm。4~9 月为主要降雨季节,月降雨量均超过200 mm。11月至翌年1月为旱季,月降雨量不足100 mm。年平均蒸发量为 1 115 mm。年平均相对湿度为80.3%(黄忠良等,1998)。每年平均遭遇3 次寒潮,寒潮常出现在 11月至翌年3 月。寒潮时气温骤降,偶有霜冻。7~9 月为台风季节,平均每年有 4 次台风登陆(黄伟峰和沈雪苹,1982)。
研究区内地带性土壤为发育于砂岩和砂页岩的赤红壤,山地垂直分布有黄壤和山地灌丛草甸土。赤红壤分布于海拔 300 m以下的丘陵地带,土层厚40~80 cm,pH 值为4.6,表土层有机质含量为4.3%;黄壤分布于海拔300~900 m,土层厚 40~90 cm,pH值范围为4.7~5.9,表土层有机质含量平均为 3.4%;900 m以上为山地灌丛草甸土,土层厚20~30 cm,pH 值为5,表土层有机质含量为9%以上。此外,尚有局部分布的耕型赤红壤(何金海等,1982)。
鼎湖山国家级自然保护区有野生被子植物 181科713 属1 566 种,裸子植物 4 科5 属 23 种,蕨类植物37科74 属131 种,苔藓植物 45 科86 属141 种。此外,约有 390 种栽培植物(
1.2 样地概况
图1
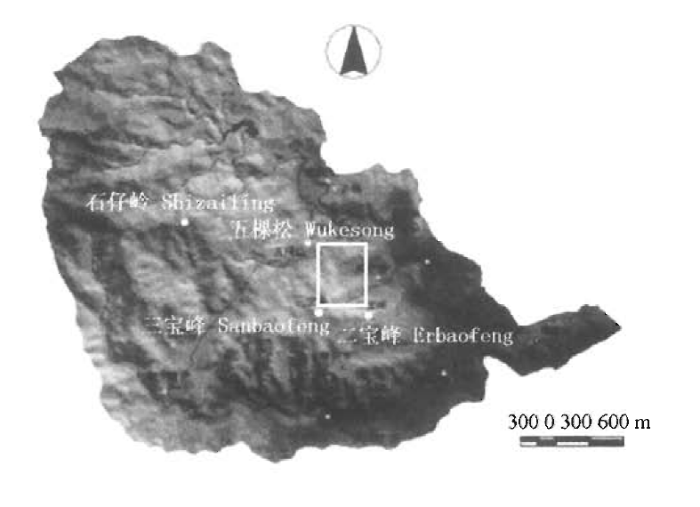
图1
中国鼎湖山南亚热带常绿阔叶林动态监测样地位置示意图
Fig.1
The location of 20 hm2 lower subtropical evergreen broadleaved forest plot in Dinghushan, China (DHS plot)
图2
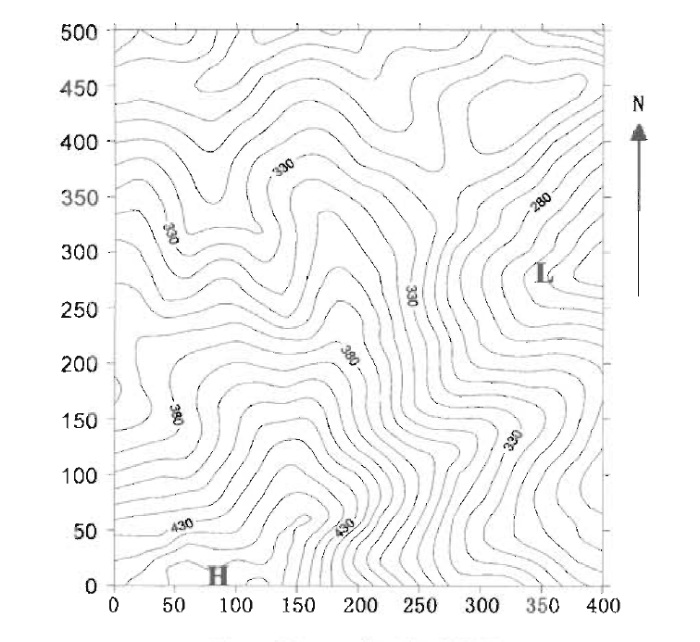
图2
鼎湖山大样地等高线图
H: 最高点为470 m The highest=470 m L: 最低点为230 m The lowest=230 m
Fig.2
The topography map of Dinghushan (DHS) plot
群落外貌表现为终年常绿,落叶成分很少,群落高达30 m,垂直结构复杂,成层现象较明显,一般可分5层,乔木上、中、下层,灌木层,草本层。此外还有藤本植物和附生植物组成的层间植物,交织攀附于乔木和灌木上。
1.3 样地的设置与调查
依据BCI大样地建设的技术规范(Condit,1998),中国森林生物多样性监测网络统一了调查研究方法。鼎湖山样地采用全站仪将20 hm2大样地划分为500个20 m×20 m的样方,样方的4个角用水泥桩作永久标记。在每个20 m×20 m的样方内用插值法细分为4个10 m×10 m和16个5 m×5 m的小样方。
野外调查以20 m×20 m样方为树种编号单元,以大样地的西南角为坐标原点,顺序排列20 m×20 m样方的行、列号。树种的个体编号共7位数,前4位数是20 m×20 m样方的行号和列号,第5位至第7位数为植株编号,对于分枝采用同一树号加分枝编号。以5 m×5 m小样方为基本测树单元,按顺时针挂牌标记每个胸径DBH(Diameter at breast height)≥1 cm的个体,记录树种名称、胸径、坐标和生长状况等信息,并建立数据库。
1.4 数据分析方法1
采用国际通用软件R 2.60 (
2 结果与分析
2.1 区系特征
表1 鼎湖山大样地木本植物统计
Table 1
| 类群 Group | 组成 Composition | 生长型 Growth type | |||||
|---|---|---|---|---|---|---|---|
| 科Family | 属Genus | 种Species | 乔木Arbor | 小乔木Subarbor | 灌木Shrub | ||
| 裸子植物Gymnosperm | 2 | 2 | 2 | 2 | - | - | |
| 被子植物Angiosperm | |||||||
| 双子叶植物Dicotyledon | 52 | 115 | 206 | 44 | 93 | 69 | |
| 单子叶植物Monocotyledon | 2 | 2 | 2 | 1 | - | 1 | |
| 合计 Total | 56 | 119 | 210 | 47 | 93 | 70 | |
表2 乔木树种物种数和个体数最大的前10个科
Table 2
| 科名 Family | 物种数与比率 No. of species (Proportion %) | 科名 Family | 个体数与比率 No. of individuals (Proportion %) |
|---|---|---|---|
| 樟科Lauraceae | 21 (10.00) | 樟科Lauraceae | 14 065 (19.64) |
| 大戟科Euphorbiaceae | 20 (9.52) | 桃金娘科Myrtaceae | 8 247 (11.52) |
| 茜草科Rubiaceae | 14 (6.67) | 茜草科Rubiaceae | 8 024 (11.20) |
| 桑科Moraceae | 11 (5.23) | 野牡丹科Melastomataceae | 5 398 (7.54) |
| 山茶科Theaceae | 9 (4.29) | 大戟科Euphorbiaceae | 5 114 (7.14) |
| 桃金娘科Myrtaceae | 9 (4.29) | 杜鹃花科Ericaceae | 4 860 (6.79) |
| 冬青科Aquifoliaceae | 8 (3.81) | 紫金牛科Myrsinaceae | 4 457 (6.22) |
| 杜鹃花科Ericaceae | 8 (3.81) | 山茶科Theaceae | 2 881 (4.02) |
| 杜英科Elaeocarpaceae | 6 (2.86) | 蝶形花科Papilionaceae | 2 791 (3.90) |
| 蝶形花科Papilionaceae | 5 (2.38) | 壳斗科Fagaceae | 2 604 (3.64) |
| 总计 Total | 111 (52.86) | 总计 Total | 58 441 (81.60) |
表3 鼎湖山大样地木本植物群落种子植物属的分布区类型统计
Table 3
| 分布区类型 Distribution area types | 属数No. of genera | 比率Rate (%) |
|---|---|---|
| 1 世界分布Cosmopolitan | - | - |
| 2 泛热带分布 Pantropic | 37 | 31.09 |
| 3 热带亚洲和热带美洲间断分布 Tropical Asia and Tropical America disjuncted | 5 | 4.20 |
| 4 旧世界热带分布 Old World Tropic | 18 | 15.12 |
| 5 热带亚洲至热带大洋洲分布 Tropical Asia and Tropical Australasia | 10 | 8.40 |
| 6 热带亚洲至热带非洲分布 Tropical Asia to Tropical Africa | 8 | 6.72 |
| 7 热带亚洲分布 Tropical Asia | 29 | 24.37 |
| 热带成分(分布型2~7) Tropical elements | 107 | 89.92 |
| 8 北温带分布 North Temperate | 3 | 2.52 |
| 9 东亚和北美洲间断分布 East Asia and North America disjuncted | 5 | 4.20 |
| 温带成分(分布型8~9) Temperate elements | 8 | 6.72 |
| 10 旧世界温带分布 Old World Temperate | - | - |
| 11 温带亚洲分布 Temperate Asia | - | - |
| 12 地中海区、西亚至中亚分布 Mediterranean, West Asia to Central Asia | - | - |
| 13 中亚分布 Central Asia | - | - |
| 14 东亚分布 East Asia | 4 | 3.36 |
| 15 中国特有分布 Endemic to China | - | - |
| 合计 Total | 119 | 100.00 |
2.2 物种组成
2.2.1 种-多度格局
样地具有较高的物种多样性,在20 hm2的样地中有210个物种,其中个体数量较多的前38个物种占总个体数的90%,个体数量最多的前9个物种占总个体数的50%,而个体数较少的110个种个体数之和还不到样地总个体数的1%(图3)。
图3
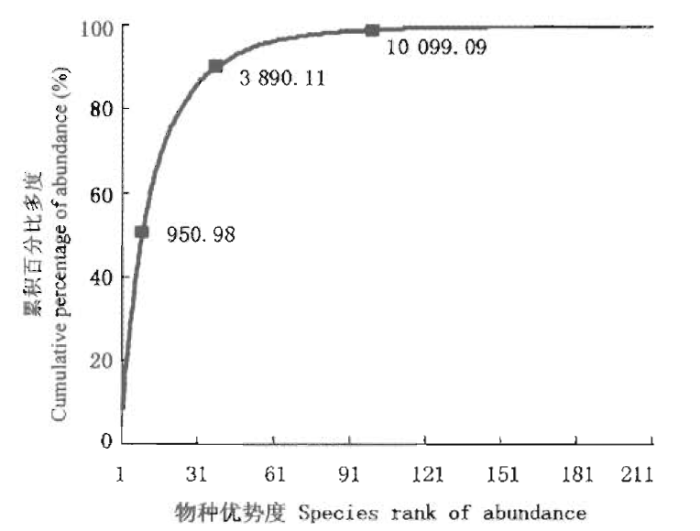
图3
鼎湖山大样地物种多度的累计分布图
Fig.3
Cumulative distribution map of species abundance in Dinghushan (DHS) plot
2.2.2 优势度
按树种重要值(Important value,简称IV)((相对频度+相对多度+相对胸高断面积)/3) )排序,样地中 IV≥1的树种有29种(表4),这些物种的多度与胸高断面积分别占样地总多度与总胸高断面积的83.69%与90.52%。IV最大的前3个物种为锥(Castanopsis chinensis)、木荷(Schima superba)和黄杞(Engelhardtia roxburghiana),均是乔木上层的优势种,同时也是胸高断面积最大的物种,三者之和为16.91 m2·hm-2,占样地总胸高断面积的56.06%。重要值较大的其它物种多为个体数量较多的物种(表5),其中红枝蒲桃(Syzygium rehderianum)、广东金叶子(Craibiodendron scleranthum var. kwangtungense)、香楠(Aidia canthioides)、厚壳桂(Cryptocarya chinensis)和黄果厚壳桂(C. concinna)等是乔木中下层的优势种。
表4 鼎湖山大样地的物种组成
Table 4
| 物种名 Species | 多度 Abundance | 分枝数 Ramification | 胸高断面积 Basal area (m2·hm-2) | 重要值 Important value |
|---|---|---|---|---|
| 锥Castanopsis chinensis | 2 311 | 546 | 9.304 | 12.3 |
| 木荷Schima superba | 2 296 | 260 | 4.132 | 6.6 |
| 黄杞Engelhardtia roxburghiana | 737 | 149 | 3.475 | 4.8 |
| 红枝蒲桃Syzygium rehderianum | 5 990 | 347 | 0.879 | 4.7 |
| 广东金叶子Craibiodendron scleranthum var. kwangtungense | 3 325 | 111 | 1.664 | 4.4 |
| 香楠Aidia canthioides | 5 996 | 422 | 0.213 | 4.1 |
| 厚壳桂Cryptocarya chinensis | 2 557 | 1 028 | 1.208 | 3.5 |
| 黄果厚壳桂Cryptocarya concinna | 4 478 | 106 | 0.177 | 3.3 |
| 云南银柴Aporusa yunnanensis | 3 747 | 378 | 0.436 | 3.0 |
| 罗伞树Ardisia quinquegona | 3 702 | 503 | 0.076 | 2.8 |
| 柏拉木Blastus cochinchinensis | 4 011 | 718 | 0.058 | 2.7 |
| 肖蒲桃Acmena acuminatissima | 1 484 | 441 | 1.096 | 2.6 |
| 光叶红豆Ormosia glaberrima | 2 702 | 116 | 0.288 | 2.2 |
| 黄叶树Xanthophyllum hainanense | 1 873 | 160 | 0.359 | 2.0 |
| 滇粤山胡椒Lindera metcalfiana | 2 118 | 369 | 0.190 | 1.9 |
| 肉实树Sarcosperma laurinum | 1 576 | 146 | 0.330 | 1.9 |
| 华润楠Machilus chinensis | 532 | 65 | 0.845 | 1.7 |
| 鼎湖钓樟Lindera chunii | 1 302 | 1 016 | 0.151 | 1.5 |
| 谷木Memecylon ligustrifolium | 1 263 | 74 | 0.092 | 1.4 |
| 小新木姜Neolitsea umbrosa | 1 352 | 154 | 0.169 | 1.3 |
| 短序润楠Machilus breviflora | 800 | 327 | 0.344 | 1.3 |
| 褐叶柄果木Mischocarpus pentapetalus | 1 252 | 95 | 0.044 | 1.3 |
| 鱼骨木Canthium dicoccum | 603 | 19 | 0.158 | 1.2 |
| 密花树Rapanea neriifoli | 750 | 56 | 0.289 | 1.2 |
| 九节Psychotria rubra | 908 | 54 | 0.016 | 1.2 |
| 毛缘杜鹃Rhododendron henryi var. concavum | 810 | 208 | 0.278 | 1.1 |
| 马尾松Pinus massonian | 169 | 6 | 0.700 | 1.0 |
| 岭南山竹子Garcinia oblongifolia | 652 | 48 | 0.104 | 1.0 |
| 三花冬青Ilex triflora | 642 | 28 | 0.234 | 1.0 |
| 其它Others | 11 679 | 1 042 | 2.860 | 21.1 |
| 合计Total | 71 617 | 8 992 | 30.167 | 100.0 |
DBH:胸径Diameter at breast height
表5 个体数最多的10个物种
Table 5
| 种名 Species | 个体数与比率 No. of individuals (Proportion %) |
|---|---|
| 香楠Aidia canthioides | 5 996 (8.37) |
| 红枝蒲桃Syzygium rehderianum | 5 990 (8.36) |
| 黄果厚壳桂Cryptocarya concinna | 4 478 (6.25) |
| 柏拉木Blastus cochinchinensis | 4 011 (5.60) |
| 云南银柴Aporusa yunnanensis | 3 747 (5.23) |
| 罗伞树Ardisia quinquegona | 3 702 (5.17) |
| 广东金叶子Craibiodendron scleranthum var. kwangtungense | 3 325 (4.64) |
| 光叶红豆Ormosia glaberrima | 2 702 (3.77) |
| 厚壳桂Cryptocarya chinensis | 2 556 (3.57) |
| 锥Castanopsis chinensis | 2 312 (3.23) |
| 合计 Total | 38 819 (54.20) |
2.2.3 种-面积格局
根据样地的种-面积曲线可知,巢式取样与随机取样两种方法均显示,鼎湖山大样地在取样面积为2 500 m2以下时,物种随面积的增加而急剧增加;面积在2 500 m2以上时,物种数增加比较缓慢,面积在接近20 hm2 时,物种数增加才趋于平稳,由此可见,样地的物种十分丰富。种-面积曲线用幂函数模型(S=CAz)进行拟合,随机取样公式为:S=6.69A0.284(R2=0.992,p<0.001);巢式取样为:S=6.59A0.285(R2=0.997, p<0.001),其中S为物种数,A为样方面积,所得的Z值接近于经典观测值1/4(Rosenzweig, 1995)。以此值推测出本地区50 hm2样地的物种数为277种,与CTFS 各热带地区的50 hm2样地相比,鼎湖山大样地的物种数高于一年具有6个月旱季的泰国HKK(Huai Kha Khaeng Wildlife Sanctuary, in Thailand)(248种)和印度Mudumalai(72种),接近于一年具有4个月旱季的巴拿马BCI(299种),而低于其它无旱季热带地区的样地,如马来西亚Pasoh(818种)和马来西亚Lambir(1 174种,52 hm2)(Condit et al., 2000)。
2.2.4 稀有种
按照Hubbell和Foster(1986)的定义,每公顷个体数等于或少于1的物种被认为是本样地的稀有种(Rare species)。样地的稀有种比例很高,共有110个,占总物种数的52.38%,其中只有1个个体的物种数为27,占总物种数的12.86%,而稀有种的个体总数为649株,仅占样地个体总数的0.91%。在全球已建立的多个大样地中仅次于HKK(60%以上),而其它大样地都不超过40%。
通常群落中稀有种较多的原因,首先取决于物种本身的种群特征与分布特性,有些物种在自然界中种群数量稀少或分布零散;其次是由于区系的交汇分布造成的,相邻气候带的物种在分布上相互渗透,一旦超出主要分布区域,其个体数量将明显下降;再之,人为或自然的干扰也会导致种群数量的减少,如过度采挖、偶然地人为带入、人为或自然的局部生境改变等。在鼎湖山大样地中有45%的稀有种源于物种本身的特性,有20%的稀有种源于区系交汇,人为干扰活动造成的稀有种占30%以上(表6)。
表6 鼎湖山大样地稀有种形成的原因
Table 6
| 形成原因 Forming factors | 物种数 Species number | 比率 Proportion (%) |
|---|---|---|
| 1. 物种特性 Species characteristics | 49 | 44.54 |
| 区域稀有种 Rare in Lower Subtropical Area | 32 | 29.09 |
| 鼎湖山分布较少 Rare in DHS Reserve | 17 | 15.45 |
| 2. 区系交汇Floristic transitional zone | 24 | 21.82 |
| 典型亚热带种类 Typical subtropical species | 18 | 16.36 |
| 沟谷分布种 Ravine species | 6 | 5.46 |
| 3. 干扰Disturbance | 37 | 31.82 |
| 阳生性树种 Heliophytes species | 19 | 17.27 |
| 中生性树种 Mesophyte species | 16 | 14.55 |
| 果树 Fruit tree | 2 | 1.82 |
| 合计 Total | 110 | 100.00 |
在CTFS的所有大样地中,HKK具有很高的稀有种比例,Bunyavejchewi 等(2004)将其原因主要归结为生境异质性和森林类型相嵌,以及干扰造成的相嵌结构的变化,而忽略了物种本身的特性。此外,由于鼎湖山保护区处于热带向中亚热带的气候过渡带,在低海拔地段分布着具有热带性质的沟谷雨林,在中山地段(海拔500 m以上)分布着具有中亚热带性质的山地常绿阔叶林。样地位于海拔230~470 m之间,高差达240 m。其下限接近沟谷雨林群落,上限接近山地常绿阔叶林,因此邻近区系的物种渗透也占了较大的比例。
图4
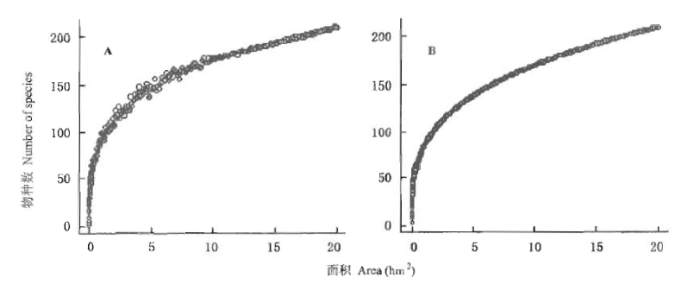
图4
鼎湖山大样地种-面积曲线
A: 随机取样Random sampling B: 巢式取样Nest sampling
Fig.4
Species-area curve in Dinghushan (DHS) plot
2.3 垂直结构
乔木上层的优势种比较显著,高20 m以上,主要由锥、木荷、黄杞等高大的阳性乔木构成,种类少,郁闭度仅为0.3。乔木中层是群落的主要层,高10~17 m,常见的种类有厚壳桂、黄叶树(Xanthophyllum hainanense)、华润楠(Machilus chinensis)、广东金叶子、肖蒲桃(Acmena acuminatissima)等中生、耐阴树种,郁闭度近0.6;乔木下层高3~9 m,由于一些乔木幼树和大灌木的进入,致使该层成分较复杂,物种多样性高,不同地段物种组成差异较大,主要有云南银柴(Aporusa yunnanensis)、厚壳桂、红枝蒲桃、肉实树(Sarcosperma laurinum)等,郁闭度仅为0.2左右。
灌木层高1.2~1.5 m,平均盖度为40%,种类组成丰富,多属于耐阴种类,以柏拉木(Blastus cochinchinensis)、罗伞树(Ardisia quinquegona)、九节(Psychotria rubra)为主,此外还有一些乔木的幼树。草本层的组成种类主要是几种阴生或耐阴的蕨类,如沙皮蕨(Hemigramma decurrins)、刺头复叶耳蕨(Arachniodes exilis)、金毛狗(Cibotium barometz)等。层间植物有丁公藤(Erycibe obtusifolia)、白叶瓜馥木(Fissistigma glaucescens)、白花油麻藤(Mucuna birdwoodiana)、扁担藤(Tetrastigma planicaule)等藤本植物,以及眼树莲(Dischidia chinensis)、石柑子(Pothos chinensis)、百足藤(Pothos repens)、小叶爬崖香(Piper arboricola)等附生植物。
2.4 径级结构
径级结构是植物群落稳定性和生长发育状况的重要指标。样地内DBH≥1 cm的树木的总胸高断面积为30.17 m2·hm-2。样方内乔木最大胸径为175 cm(锥)。样地中所有个体的径级分布(以1 cm等级排列)明显呈现倒“J"形(图5),表现出群落稳定与正常生长状态。从径级结构可以看出,DBH≤10 cm的个体数量居多,占83.58%,其中DBH≤4 cm的个体数占样地总个体数的60.86%;DBH>40 cm的个体数为504株,仅占0.70%。
图5
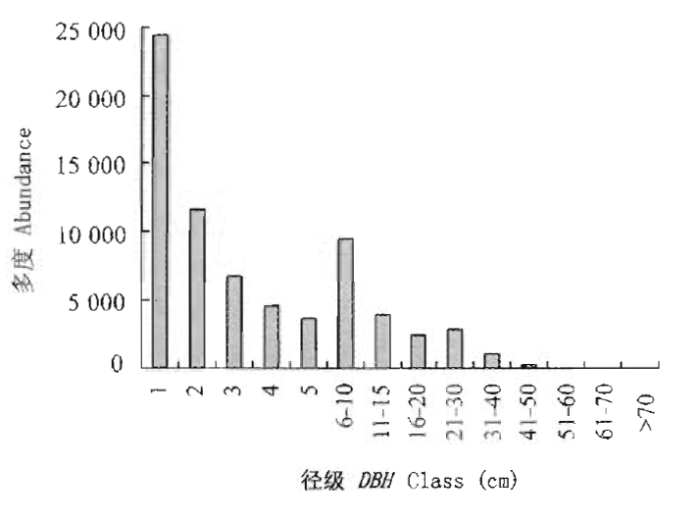
图5
鼎湖山大样地木本植物径级分布图
Fig.5
Distribution patterns of size class of diameter at breast height (DBH) of tree species in Dinghushan (DHS) plot
样地中DBH>10 cm的个体比例为16.42%,高于CTFS中热带雨林区的大样地,如BCI、Pasoh、Lambir、Sinharaja(均小于10%),而低于CTFS中热带季雨林区的大样地,如HKK(27.10%),Mudumalai (58.97%)(Condit et al., 2000),体现了鼎湖山的南亚热带季风气候特征。
根据对优势种径级结构的分布分析(图6),各树种的径级结构可归纳为4种类型:1)峰型(中径级个体储备型),此类物种为乔木上层优势种,其径级结构类似于正态分布,个体主要集中在中径级,小径级与大径级则相对较少,代表种为锥、木荷和黄杞;2)倒“J”型 (正常型),此类物种为乔木中层的优势种,径级结构的形状类似于倒置的“J”字,以DBH=1 cm的个体数最大,并随着DBH的增加个体数量逐步下降,代表种为厚壳桂、黄叶树和华润楠;3)类倒“J”型(偏正常型),此类物种占据乔木中、下层,径级结构近似于倒“J”型,但径级个体数最大值出现在DBH=2~4 cm,代表物种有肖蒲桃、广东金叶子和云南银柴等;4)“L”型(灌木型),此类物种分布于乔木下层和灌木层,个体集中在小径级处,大径级个体极少,代表种有柏拉木、香楠和黄果厚壳桂等。
图6
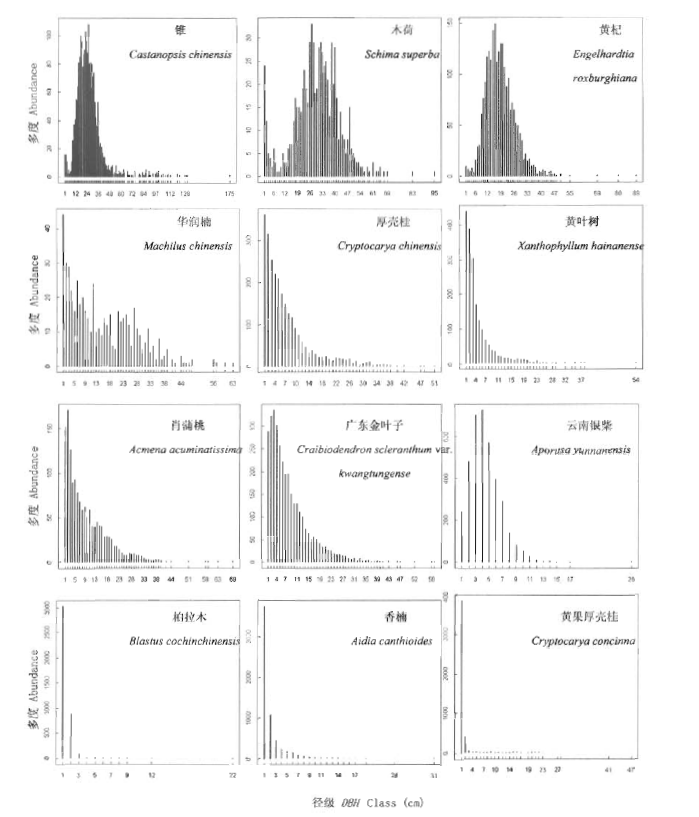
图6
鼎湖山大样地优势树种的径级结构图
Fig.6
Size-class distribution of dominant species in Dinghushan (DHS) plot
2.5 空间分布格局
对部分优势种按不同径级个体的点分布格局分析表明,优势种在各个层上的绝大部分尺度都是聚集分布的(图7),而不同径级的空间分布随物种的不同而呈现明显的差异,点格局显示了优势种在相同径级的分布(特别是在DBH=10~40 cm时)具有一定的空间互补性。
图7
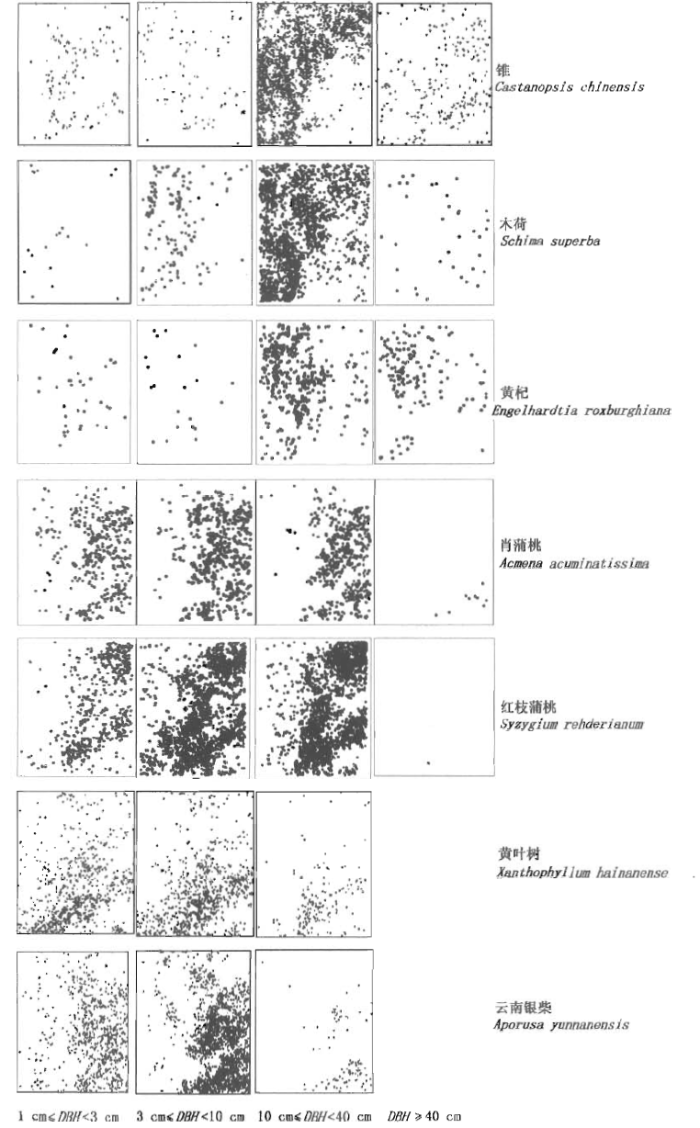
图7
鼎湖山大样地优势树种在4个不同径级的分布图
DBH: 胸径 Diameter at breast height
Fig.7
Distribution map of dominant species in Dinghushan (DHS) plot at four DBH classes
同一物种不同径级个体的空间分布规律基本一致,个别物种,如锥在中径级(10~40 cm)和大径级(40 cm以上)个体分布上存在着一定的互补关系,中径级个体主要分布于样地的西北部,大径级个体主要分布于东南部。同一层次、同一径级的不同物种之间也存在分布的互补性特征,如锥和黄杞都是乔木上层的优势种,其大径级个体的分布存在明显的互补性,锥的大径级个体主要分布于东南部,黄杞的大径级个体主要分布在西北角。在不同层次的优势种在空间分布格局上也存在互补关系,如在中径级个体分布上,乔木上层种锥和木荷主要分布在西北部,黄杞主要分布在西部;而乔木中、下层优势种肖蒲桃和广东金叶子主要分布在东部,黄叶树和云南银柴主要分布在东南角,与乔木上层树种形成互补。
对大径级(DBH>40 cm)的504个个体进行空间分布格局分析(图8),结果表明,所有这些个体随机分布于整个样地,说明样地的群落构成比较自然,没有受到大范围的干扰。但不同的物种在空间分布上存在一定的规律,锥是乔木上层的主要树种,其大径级个体聚集分布在山脊上,黄杞则聚集在山谷,华润楠主要出现在中海拔区域。
图8
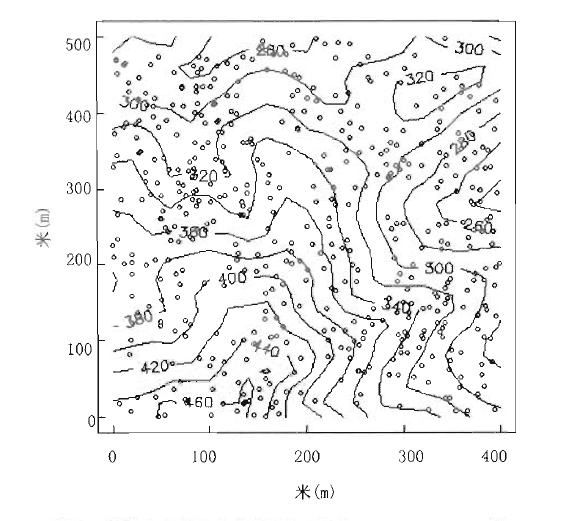
图8
鼎湖山大样地大径级乔木(DBH > 40 cm)的分布图
Fig.8
Distribution map of individuals with DBH > 40 cm in Dinghushan (DHS) plot
3 结论与讨论
通过对鼎湖山常绿阔叶林20 hm2样地物种组成与群落结构的分析发现,样地物种组成十分丰富,包括56科119属210种,种-面积曲线拟合显示其物种数量接近于BCI。区系特征明显,南亚热带的区系成分占绝对优势,并呈现由亚热带向热带过渡的特色。群落垂直结构复杂,地上成层现象较明显,优势树种明显,乔木上层的优势种锥、木荷和黄杞在群落中占有绝对优势地位,它们均为寿命长的阳生性树种,个体主要分布在中径级上,并以中径级作为主更新库来保持其在群落中长期的优势种地位。林下更新良好,红枝蒲桃、广东金叶子、香楠、厚壳桂、黄果厚壳桂等是乔木中下层的优势种,在样地中占有一定的比例。稀有种比例极高,有110种,占总物种数的52.38%,其中有45%的稀有种源于物种本身的特性,有20%源于区系交汇,人为或自然干扰造成的稀有种占30%以上。所有个体的径级分布(以1 cm等级排列)明显呈倒“J”形,表示群落稳定与正常生长状态。根据对优势种径级结构的分布分析,各树种的径级结构可归纳为4种类型:1)峰型(中径级个体储备型),此类物种为乔木上层优势种;2)倒“J”型 (正常型),此类物种为乔木中层的优势种;3)类倒“J”型(偏正常型),此类物种占据乔木的中、下层;4)“L”型(灌木型),此类物种分布于乔木下层和灌木层。从物种的空间分布格局来看,点格局分析得出优势种在各个层上的绝大部分尺度都是聚集分布的,而不同径级的空间分布随物种的不同而呈现出明显的差异性,点格局显示了优势种在相同径级的分布(特别是DBH=10~40 cm)具有一定的空间互补性,对大径级(DBH>40 cm)的504个个体进行空间分布格局分析,结果表明,所有个体随机分布于整个样地,但不同的物种在空间分布上存在一定的规律。
鼎湖山常绿阔叶林20 hm2样地与以往相同群落类型的1 hm2永久样地(孔国辉等,1998)相比,在物种组成与物种多度格局等方面都存在较大的差异。从物种组成看,1 hm2永久样地DBH≥1 cm的木本植物仅为123种,远小于20 hm2大样地的210个物种。从种-面积曲线的结果可知,在取样面积接近20 hm2时,曲线才趋于平缓,说明以20 hm2作为样地的尺度对研究鼎湖山常绿阔叶林更具完整性。从上层优势种锥、木荷和黄杞的多度格局来看,1 hm2样地由于取样面积的限制,这些树种的小径级个体很少,因而认为锥和黄杞为衰退种,木荷为近期进展远期衰退种(黄忠良等,1998),但20 hm2样地的结果显示,这3个优势种的多度是以中径级个体为主储备型结构的类型。由此可见,鼎湖山大样地能更为客观地反映不同尺度上鼎湖山常绿阔叶林群落水平的总体特征,能更好地为日后开展群落生态学的研究提供全面的数据。
长期以来,国内外学者对南亚热带常绿阔叶林植被进行了大量的研究(彭少麟,1996; 彭少麟和方炜,1994;彭少麟等,1998)。对季风常绿阔叶林植被分布、演替规律、群落结构和物种多样性等有了较为全面的了解。但是迄今为止,对南亚热带常绿阔叶林物种多样性维持机制的研究还不够充分,研究结果较少(叶万辉,2000;彭闪江等,2003;彭闪江和徐国良,2005;李林等,2006)。群落物种多样性的维持机制或组织方式的研究,一直是群落生态学研究的核心内容。目前国际上这方面的研究基本上是在热带雨林中开展的,基于热带雨林的研究提出了众多的理论假说。作为最具代表性的常绿阔叶林,鼎湖山大样地有其独特的复杂地形与过渡型气候带。那么,基于热带雨林推出的理论假说是否适用于南亚热带常阔叶林?该地带森林的群落组织方式是否有其特点?南亚热带森林群落中有着怎样的生物多样性格局与规律?生态位分化理论和中性理论等在南亚热带森林群落生物多样性格局与维持机制中有着怎样的作用和地位?这些问题的解决既可能为生物多样性的组织机制理论带来突破,又对发展完善当前的群落生态学理论具有十分重要的意义。同时,开展亚热带森林物种多样性维持机制的研究,对于加强亚热带森林的管理和保护也具有十分重要的现实意义。因此,对鼎湖山南亚热带常绿阔叶林群落特征的研究,将促进我们对这些问题的解决,进一步对群落多样性的维持机制做深入研究。
参考文献
Vegetation map of Dinghu Mountain Nature Reserve
Towards a really unified theory for metacommunities
Research in large, long-term tropical forest plot
Tropical Forest Census Plots: Methods and Results from Barro Colorado Island, Panama and a Comparison with Other Plots
The importance of demographic niches to tree diversity
Spatial patterns in the distribution of tropical tree species
Beta-diversity in tropical forest trees
The soils of Ding Hu Shan biosphere reserve
Factors affecting community composition of forest regeneration in deforested, abandoned land in Panama
The climatic ecological environments in the region of Ding Hu Shan
Climate of Dinghushan Biosphere Reserve
Neutral theory and the evolution of ecological equivalence
Light gap disturbances, recruitment limitation, and tree diversity in a neotropical forest
Floristic composition of Pasoh forest reserve, a lowland rain forest in peninsular Malaysia
Long-term monitoring of the low subtropical evergreen broad-leaved forest in Dinghushan Biosphere Reserve. Ⅰ. Species composition of Castanopsis chinensis, Cryptocarya concinna community and its contribution
Long-term ecological research of tropical rain forest in Sarawak
Study on the relationship between α diversity of plant community and environment on Dinghushan
Stand structure of Pasoh forest reserve, a lowland rain forest in peninsular Malaysia
A test of the unified neutral theory of biodiversity
Empirical evaluation of the neutral theory
Effects of habitat heterogeneity on forest community diversity in Dinghushan Biosphere Reserve
Seed traits of Castanopsis chinensis and its effects on seed predation patterns in Dinghushan Biosphere Reserve
Studies on the dynamics of dominant populations of Dinghushan forest during successions. Ⅲ. Cryptocarya chinensis and Cryptocarya concinna populations
The dynamics on organization in the successional process of Dinghushan Cryptocarya community
Neutral theory and relative species abundance in ecology
The vegetation of Ding Hu Shan Biosphere Reserve
Physico-geographical features of Ding Hu Shan and their dynamic analyses
The areal-types of Chinese genera of seed plants
The maintenance mechanism of plant community and its species diversity
Assessment of function area design in Dinghushan Biosphere Reserve using landscape ecology principles

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}