


养分是影响生态系统结构和功能的重要因素, 其供应量是否充足是影响有机体生长、种群结构和生态系统稳定性的重要因素(曾德慧和陈广生, 2005)。氮素和磷素是生物体的重要组成成分, 是生命活动过程所必需的大量营养元素, 也是土壤生产力的重要限制性因素。在大多数的陆地生态系统中, 氮和磷是限制生物生长的重要限制因子(Güsewell, 2004)。已有研究认为植物叶片的氮磷比(N:P)可以作为判断环境因子, 特别是土壤对植物生长的养分供应状况的指标(Aerts & Chapin, 2000; Güsewell, 2004; Wassen et al., 1995)。
近年来, 大气中含氮物质浓度不断增加, 其来源和分布迅速扩展到全球范围(Galloway & Coeling, 2002; Galloway et al., 2003), 氮沉降导致的土壤氮素增加将对全球陆地生产力产生深远的影响(Reich et al., 2006; Magnani et al., 2007; Mo et al., 2008)。相对于氮素, 陆地生态系统土壤中磷的来源相当稳定, 因此, 土壤中的氮磷比失调越来越严重的趋势不可避免。也有研究认为, 热带、亚热带丰富的氮可以促进植物对磷的吸收(Houlton et al., 2008), 但是即便如此, 土壤中的磷素有效性还是不足以平衡氮素有效性(Asner et al., 2001; He et al., 2002), 因此磷素成为许多陆地生态系统生物生长和重要生态过程的限制因子(He et al., 2002; Kellogg & Bridghain, 2003), 尤其是分布在低纬度地区的热带、亚热带生态系统(Ae et al., 1990; Kellogg & Bridghain, 2003)。Wardle等(2004)研究发现, 在缺乏灾难性干扰的情况下, 森林生态系统演替到后期, 常常会出现生产力下降, 并且新鲜凋落物和腐殖质中N:P增加, 说明随着演替进行, 森林生态系统越来越受到P的限制。我国南亚热带森林分布在低纬度地区, 是有效磷缺乏的区域, 而该地区工业化、城市化迅速发展, 存在高氮沉降的现象, 植被和土壤中氮、磷状况及其比值特征需要深入研究。
本文基于多年的数据资料, 研究鼎湖山森林生态系统不同演替阶段上土壤和植被的N、P的化学计量特征, 特别深入分析了土壤与植物化学计量特征之间的关系, 试图探讨演替进展过程对土-植系统中营养元素在化学计量上的反映。
1 研究地概况
鼎湖山位于广东省肇庆地区境内(23°10′ N, 112°34′ E)。该地区多为丘陵和低山, 海拔为100- 700 m, 最高峰鸡笼山海拔1 000.3 m。气候属南亚热带季风湿润型气候, 冬夏气候交替明显。年平均气温20.9 ℃, 最热月7月, 最冷月1月, 极端最高气温38.0 ℃, 极端最低气温-0.2 ℃, 年降雨量1 860 mm, 4-9月为主要降雨季节, 11月至翌年1月为少雨季节, 年平均蒸发量1 115 mm, 年平均相对湿度82%。灾害性天气为寒潮和台风。土壤由不同颜色、硬度与质地的砂岩、砂页岩、页岩和石英砂岩发育而成, 主要土壤类型为赤红壤和红壤。
本研究的样地分布在赤红壤上同一演替系列的3种植物群落, 即演替初期的马尾松(Pinus massoniana)林、演替中期的针叶阔叶混交林和演替后期的季风常绿阔叶林(简称季风林)(彭少麟和王伯荪, 1993)。马尾松林样地海拔150-250 m, 马尾松是现有的唯一乔木层, 林下灌木丰富, 主要为桃金娘(Rhodomyrtus tomentosa)和三叉苦(Evodia lepta)等, 荷木(Schima superba)等阔叶树种幼苗丰富。混交林样地海拔150-250 m, 针叶树是马尾松, 阔叶树种主要有荷木、锥栗(Castanopsis chinensis)和黧蒴锥(C. fissa)等, 针阔比例约3:7。季风林样地海拔200-300 m, 群落外貌终年常绿, 垂直结构复杂。优势树种主要有锥栗、荷木、厚壳桂(Cryptocarya chinensis)、黄果厚壳桂(C. concinna)和华润楠(Machilus chinensis)等。
2 研究方法
2.1 采样处理方法
2.1.1 土壤
在以上3个林型的样地内, 按照0-10、10-20、20-40和40-60 cm的层次, 用直径2 cm的土钻采集10个土壤样品, 每个样品为每个位置点直径1 m范围内的4钻土混合而成, 装入土袋, 带回实验室。土样制备: 将土样放至通风干燥处, 剔除石粒和草根等杂物, 自然风干, 再用滚轴磨成2 mm的粉末, 装瓶待用。
2.1.2 植被
分别采集针叶林的马尾松, 混交林的马尾松、荷木和锥栗, 季风林的锥栗、荷木、厚壳桂、黄果厚壳桂和华润楠等优势种的根、树皮、树芯、叶和枝, 带回实验室烘干、磨碎、过60目筛, 装瓶供测试。
2.2 分析方法
分析项目为土壤全氮、全磷, 植物各器官的全氮、全磷。植物全氮和土壤全氮用凯氏法测定, 植物全磷用硝酸-高氯酸消煮-钼锑抗分光光度法(GB 7887-87, 分光光度计, 上海)测定, 土壤全磷用硫酸- 高氯酸消煮-钼锑抗比色法(GB 7852-87)测定。植物样品分析: 同一物种之间的样品得出算术平均数, 不同演替阶段林型的养分含量用所选优势种的养分含量根据物种的重要值计算加权平均值来表示, 得出不同林型植物体各器官的氮磷含量值。
采用SPSS 11.5统计软件进行ANOVA分析。不同演替阶段森林土壤的氮磷含量、植物的氮磷含量、土壤氮磷比和植物氮磷比通过单因素方差分析(One-Way ANOVA)的LSD法进行显著性检验。不同演替阶段的共有种的氮磷含量、氮磷比采用t检验。所有结果p < 0.05, 即差异显著。
3 结果
3.1 土壤中的N、P含量
鼎湖山不同演替阶段的森林土壤中N的含量随演替进行而增加(图1), 0-10 cm土层增加得更为显著, 该土层中演替初期(马尾松林) N含量为0.440 g·kg-1、演替中后期(混交林和季风林) N含量分别达到0.843和1.023 g·kg-1, 为演替初期的2-2.5倍。对同一演替阶段、不同土层的N含量作差异显著性检验, 各林型中表层(0-10 cm)土壤N含量与底层土壤的分布差异非常明显(p < 0.05), 表层富集现象尤为突出; 而对同一土层、不同演替阶段的N含量作差异显著性检验, 演替初期与演替中后期的各层土壤均存在显著差异(p < 0.05), 但演替中后期之间各层土壤N含量差异不显著(p > 0.05), 说明演替中后期, 土壤中N的输入与输出趋于平衡, 增加趋势变缓(图1)。
图1
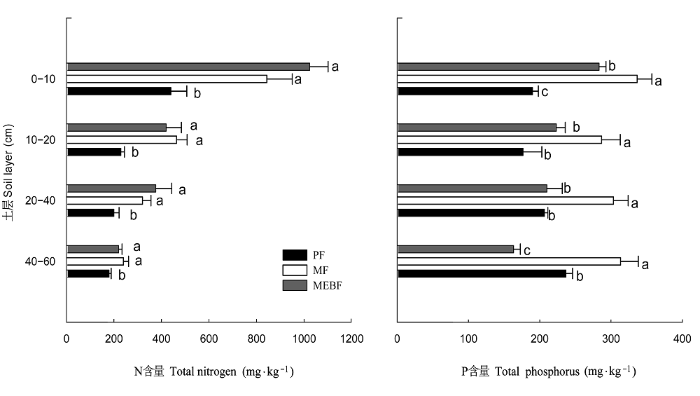
图1
鼎湖山3种不同演替阶段森林土壤的氮磷含量。误差线代表标准误(SE); 同一土层具有不同字母的表示差异显著 (p < 0.05)。图中氮、磷值为2005年数据。MEBF, 季风常绿阔叶林; MF, 混交林; PF, 马尾松林。
Fig. 1
Soil total nitrogen and phosphorus concentrations among three forest types at different succession stages in Dinghushan. Different letters indicate significant differences at the confidence level of p < 0.05 in the same soil layer among the three forests. Data source: 2005. MEBF, monsoon evergreen broad-leaved forest; MF, mixed forest; PF, pine forest.
土壤中P含量以演替中期混交林最为丰富, 其0-10 cm土层中P的含量为0.337 g·kg-1, 而演替前期和后期土壤中P的含量相对较低, 马尾松林和季风林土壤中P含量分别为0.190和0.283 g·kg-1。可见, 不同演替阶段土壤中P的含量表现出较大的差异 (p < 0.05)。但P的含量随土层的变化与N在土层中的分布大不相同, 没有表现出显著差异, 在不同土壤层次上变化幅度不大, 垂直分布相当均匀。因此, 可以看出该研究区域森林土壤中N的含量在土壤剖面上呈现出“倒金字塔”的分布, 而P含量在土壤剖面上表现的是“圆柱体”分布模式(图1)。
3.2 植被中的N、P含量
在所有的植物器官中, 叶片的N、P含量最高(图2), 3个演替阶段森林植被叶片N的含量超过了2%, P的含量在0.08%-0.10%范围内; 随着演替的进行, 叶片中的N、P含量都呈减少的趋势。对不同演替阶段叶片N、P含量进行方差分析发现, 马尾松林与演替中后期的混交林和季风林均存在显著差异(p < 0.05), 但混交林与季风林差异不显著(p > 0.05)。根、枝和皮的N、P含量相差不大, 其中根系中N、P的含量都以马尾松林为最多(0.93%和0.05%), 混交林和季风林根系中N、P的含量彼此相当。所有演替阶段树干的N、P含量最低。
图2
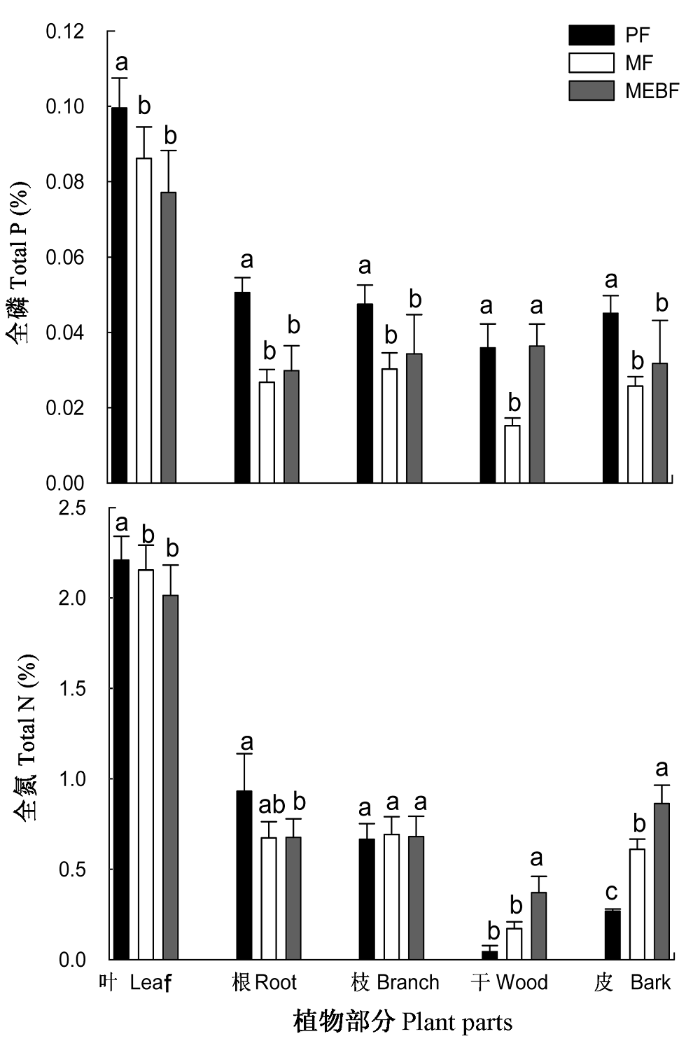
图2
鼎湖山3种不同演替阶段植被的N、P含量。季风林数据时间范围是1992-2005年; 混交林为2001-2006年; 马尾松林为1992-2006年。同一部分具有不同字母的表示差异显著(p < 0.05)。MEBF、MF、PF, 同
Fig. 2
Plant total nitrogen and phosphorus concentrations among three types of forests at different succession stages in Dinghushan. Data source: MEBF, from 1992 to 2005; MF, from 2001 to 2006; PF, from 1992 to 2006. Different letters indicate significant differences at the confidence level of p<0.05 in the same plant parts among the three forests; MEBF, MF, PF, see
同一树种N、P的含量在不同演替阶段也存在着差异(表1)。对不同演替阶段的相同物种的叶片和根系的N、P含量进行t检验, 结果表明, 马尾松林的马尾松叶片和混交林的马尾松叶片的P含量、混交林荷木叶片和季风林荷木叶片的N含量差异极显著 (p < 0.01)。另外, 不同演替阶段的相同树种马尾松、荷木和锥栗的根系的P含量均存在显著性差异(p < 0.05), 而根系N的含量只在锥栗中存在显著性差异(p < 0.05)。
表1 不同演替阶段相同物种的N、P含量差异分析
Table 1
| 共有种 Common species1) | 叶 Leaf | 根 Root | |||||
|---|---|---|---|---|---|---|---|
| N | P | N:P | N | P | N:P | ||
| 马尾松 Pinus massoniana | 0.179 | 0.001** | 0** | 0.793 | 0** | 0** | |
| 荷木 Schima superba | 0** | 0.064 | 0** | 0.092 | 0.043* | 0.101 | |
| 锥栗 Castanopsis chinensis | 0.621 | 0.230 | 0.27 | 0.020* | 0** | 0.001** | |
表中数值为p值。1), 马尾松为马尾松林和混交林的共有种。荷木、锥栗为混交林和季风林的共有种。*, p < 0.05; **, p < 0.01。
The data in the table is p value. 1) the common species between the pine forest and the mixed forest is Pinus massoniana; Schima superba and Castanopsis chinensis are common species between the mixed forest and the monsoon evergreen broad-leaved forest. *, p < 0.05; **, p < 0.01.
3.3 土壤和植物中的N、P化学计量特征
图3
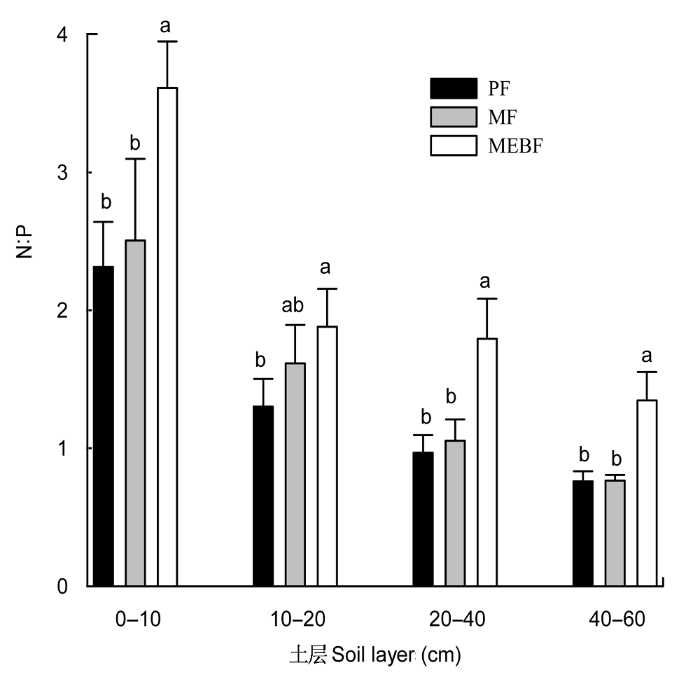
图3
鼎湖山3种林型土壤的N:P。MEBF、MF、PF, 同
Fig. 3
N:P ratio in the soil of three types of forests in Dinghushan. MEBF, MF, PF, see
图4
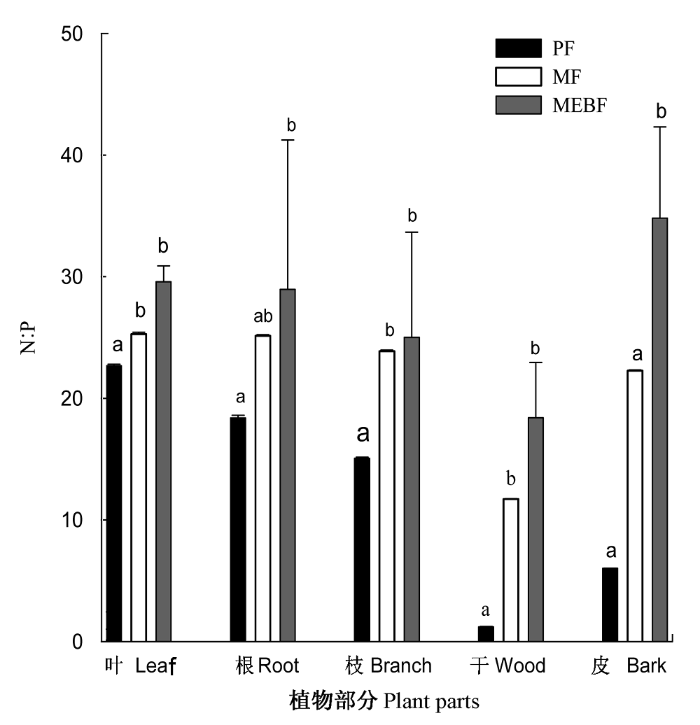
图4
鼎湖山3种林型植物的N:P。同一部分具有不同字母的表示差异显著(p < 0.05)。MEBF、MF、PF, 同
Fig. 4
N:P ratio in the plant of three types of forests in Dinghushan. Different letters indicate significant differences at the confidence level of p < 0.05 in the same plant parts among the three forests. MEBF, MF, PF, see
4 讨论
4.1 南亚热带森林生态系统植物与土壤中N:P特征
土壤作为植物体营养元素的主要来源, 其养分的变化和分布情况会对植物体的生长发育产生很大的影响。鼎湖山森林土壤中N的含量在土壤剖面上呈现出“倒金字塔”的分布, P含量在土壤剖面上表现的是“圆柱体”分布模式(图1)。这主要是由于N、P的来源不同造成的。土壤N素主要来源是凋落物的归还和大气氮沉降(鼎湖山降水氮沉降量为38.4 kg N·hm-2·a-1 (周国逸和闫俊华, 2001)), 导致氮素首先在土壤表层密集, 然后再随水或者其他介质向下层迁移扩散, 形成土壤氮素的浓度从表层到底层越来越低的一个渐进的分布格局。土壤P素的来源相对固定, 主要是通过岩石的风化。由于岩石风化是一个漫长的过程, 风化的程度在0-60 cm的土壤层中差异不大, 这就使得P素在土壤中的垂直分布呈现上下差异不大的“圆柱体”形状。
植物与土壤是生态系统生物因子与环境因子的代表, 它们的N:P存在着显著的差异, 这可能是土壤与植物各自执行不同的功能决定的。鼎湖山针叶林、混交林和季风林的植物叶片的N:P分别为对应林型0-10 cm土壤N:P的9.7、10.1和8.2倍。但是随着演替的进行, 植物和土壤的N:P增加, 说明了演替的过程中, 植物与土壤的变化是同步的。
植物与土壤作为营养元素生物地球化学循环的不同环节, 二者之间存在着必然的联系。根系是植物从土壤中直接吸收营养元素的最主要的器官(陆时万等, 1991), 是土-植系统联系的纽带。根系对土壤变化的反应最敏感, 因为植物体其他器官会受到根系作用的缓冲。植物体需要的N:P与土壤差别很大, 这就需要根系有选择地吸收土壤中的氮、磷元素。相对来说, 植物体的N:P更大, 所以根系就要吸收相对更多的氮量来提高N:P, 以满足植物体的需要。叶片是植物体营养元素利用的最主要的场所。植物体营养元素的流失主要是通过叶片的凋落(官丽莉等, 2004), 所以叶片也是植物体营养元素的输出端。根系和叶片作为植物体这个系统营养元素的输入和输出端, 为了保持这个系统的动态平衡, 二者的营养元素的比例应该一致。本研究的结果恰恰证明了这一点。研究发现鼎湖山演替阶段森林的植物叶片和根系的N:P比较接近。某个系统的营养输入和输出的比例一致, 说明该系统稳定。因此, 本文认为根系和叶片N:P的比较可以作为植物生长状况的一个评价指标。
鼎湖山森林演替序列上各土层中N:P呈现明显增加趋势。通过数据(图1, 图3)可以看出, N的增加是演替序列土壤N:P增加的主要原因。土壤中N的含量从针叶林、混交林到季风林持续增加, 主要原因有以下几点。1)凋落物的归还。自然森林生态系统的土壤中, N的最主要来源是凋落物的归还。凋落物的量和分解速率是决定土壤N的重要因素。一般来说, 阔叶林凋落物输入到土壤中的N多于针叶林, 混交林多于纯林(彭少麟和刘强, 2002)。在鼎湖山的3种林型中, 季风林的凋落物分解最快, 混交林次之, 针叶林最慢(莫江明等, 2004)。另外针叶林、混交林和季风林的年凋落物量分别为3.31 × 103、8.50 × 103和8.28 × 103 kg·hm-2 (张德强等, 2000)。2)氮沉降。除了植被凋落物的归还以外, 大气氮沉降是森林土壤氮的另一个重要来源, 尤其是在高氮沉降的珠江三角洲地区, 据研究显示, 鼎湖山降水氮沉降量为38.4 kg N·hm-2·a-1 (周国逸和闫俊华, 2001), 远远超出森林植被在生长季对氮的需求量(5-8 kg N·hm-2·a-1) (即生态系统达到氮饱和状态)。大气氮沉降在森林受到林冠的截留作用, 使得不同林型的土壤最后实际接收的氮量有所差别。方运霆等(2005)发现鼎湖山3种林型的穿透雨中铵态氮的含量分别为1.85、2.36和1.92 mg·L-1, 硝态氮的含量分别为3.34、2.34和5.65 mg·L-1, 阔叶林穿透雨中无机氮含量分别比混交林和针叶林高出61.1%和45.9%。
在土壤N含量增加成为既定事实的前提下, 植物的主要器官叶片和根系中的N并没有相应地表现出增加的趋势, 这和Schleppi等(1999)在瑞士中部的云杉(Picea abies)林的实验结果比较吻合, 他们通过N增加实验发现, 树木和地面植被的含氮量并没有明显增加, 只有一小部分的标记氮进入地上植被, 绝大部分进入土壤。所以鼎湖山演替序列上植被N:P的升高主要是P造成的, 即演替序列上植被各器官中P的含量是下降的。由于P本身的生物地球化学循环特征, 在土壤发育的过程中, 由于风化侵蚀, 土壤中原生矿物逐渐消失。土壤中的磷酸盐从非闭蓄态的有机矿物形式转变成为闭蓄态和有机结合的形式, 难以利用(Crews et al., 1995)。所以森林演替的过程中, 可被植物吸收利用的有效磷越来越有限, 就导致植物体各器官的含磷量逐渐降低。
4.2 P对南亚热带森林生态系统的限制作用
低纬度地区土壤中P缺乏已经基本成为公认的事实(Ae et al., 1990; Asner et al., 2001; He et al., 2002; Houlton et al., 2008; Kellogg & Bridghain, 2003), 在长期缺乏大的干扰的演替过程中, 到了后期土壤P也有下降的趋势(Wardle et al., 2004), 本研究也可以证明这一点。
首先, 相对于叶片来说, 根系更直接地受到土壤的影响。从鼎湖山不同演替阶段的共有物种叶片和根系P含量差异分析(表1)可以看出, 叶片P的显著性差异只存在于演替的前期和中期, 而根系P含量在演替的前、中、后期3个阶段都存在显著性差异, 这和鼎湖山表层土壤P含量的特征相同。这个结果也在一定程度上说明土壤中P含量的变化首先影响到根系, 也就是说土壤中缺P的这种限制性作用首先体现在了植物根系上。
其次, 植物叶片的N:P在许多研究中都被作为判断植物生长限制因子的指标(Aerts & Chapin, 2000; Güsewell, 2004; Wassen et al., 1995)。Koer- selman和Meuleman (1996)通过总结一些沼泽、欧石楠荒原、湿草地和沙丘等生态系统的施肥实验的结果得出结论, 即N:P < 14, 表明是氮限制, N:P > 16表明是磷限制, N:P处于二者之间为氮磷共同限制或者二者都不限制, 这个结论被广泛地应用于生态系统限制因子的判断(阎恩荣等, 2008)。也有研究认为, 生态系统本身的复杂性决定了不能够采用单一的指标来判断系统的限制因子, 不同研究区域、生态系统类型以及植被种类的不同, N:P临界指标会有很大的变化(Güsewell, 2004)。但是一般可以接受的观点是: 较低的N:P一般反映植物受到氮限制, 较高的N:P反映植物受到磷限制。问题是当N:P升高到什么程度才是磷限制, 仍然是难以确定的一个临界值。鼎湖山演替系列上的3种森林植被叶片的N:P远远超过16, 分别为22.67、25.28和29.58, 这远高于其他研究结果, 所以可以相对地反映出鼎湖山森林生态系统受磷的限制比较严重。
最后, 从绝对的量上比较分析, 我国土壤全磷量平均约为0.56 mg·g-1 (Han et al., 2005), 而位于南亚热带的鼎湖山森林土壤含磷量为0.15-0.3 mg·g-1, 这也证明了土壤P库随纬度降低而降低的规律(Hedin, 2004; Zhang et al., 2005)。而与同纬度带上植物体P含量相比, 莫江明等(2000)的研究表明鼎湖山植物叶片P含量的平均水平只相当于该气候带总平均水平的43%。另外, 我国南亚热带氮沉降量不断增加, 土壤中的氮磷比失调, 使得P成为生态系统生物生长和重要生态过程的限制因子(He et al., 2002)。
致谢
国家自然科学基金(30872012和40730102)资助项目。
参考文献
Phosphorus uptake by pigeon pea and its role in cropping systems of the Indian subcontinent
The mineral nutrition of wild plants revisited: a re-evaluation of processes and patterns
Physical and biogeochemical controls over terrestrial ecosystem responses to nitrogen deposition
Changes in soil phosphorus fractions and ecosystem dynamics across a long chronosequence in Hawaii
Acidity and inorganic nitrogen concentrations in soil solution in short-term response to N addition in subtropical forests
The nitrogen cascade
Reactive nitrogen and the world: 200 years of change
Twenty years of litter fall dynamics in subtropical evergreen broad-leaved forests at the Dinghushan Forest Ecosystem Research Station
N:P ratios in terrestrial plants: variation and functional significance
Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China
Interactions between soil moisture content and phosphorus supply in spring wheat plants grown in pot culture
Global organization of terrestrial plant nutrient interactions
A unifying framework for dinitrogen fixation in the terrestrial biosphere

Dinitrogen (N(2)) fixation is widely recognized as an important process in controlling ecosystem responses to global environmental change, both today and in the past; however, significant discrepancies exist between theory and observations of patterns of N(2) fixation across major sectors of the land biosphere. A question remains as to why symbiotic N(2)-fixing plants are more abundant in vast areas of the tropics than in many of the mature forests that seem to be nitrogen-limited in the temperate and boreal zones. Here we present a unifying framework for terrestrial N(2) fixation that can explain the geographic occurrence of N(2) fixers across diverse biomes and at the global scale. By examining trade-offs inherent in plant carbon, nitrogen and phosphorus capture, we find a clear advantage to symbiotic N(2) fixers in phosphorus-limited tropical savannas and lowland tropical forests. The ability of N(2) fixers to invest nitrogen into phosphorus acquisition seems vital to sustained N(2) fixation in phosphorus-limited tropical ecosystems. In contrast, modern-day temperatures seem to constrain N(2) fixation rates and N(2)-fixing species from mature forests in the high latitudes. We propose that an analysis that couples biogeochemical cycling and biophysical mechanisms is sufficient to explain the principal geographical patterns of symbiotic N(2) fixation on land, thus providing a basis for predicting the response of nutrient-limited ecosystems to climate change and increasing atmospheric CO(2).
Phosphorus retention and movement across an ombrotrophic-minerotrophic peatland gradient
The vegetation N:P ratio: a new tool to detect the nature of nutrient limitation
The human footprint in the carbon cycle of temperate and boreal forests
Litter decomposition and its responses to simulated N deposition for the major plants of Dinghushan forests in subtropical China
Distribution pattern of nutrient elements in plants of Dinghushan lower subtropical evergreen broad-leaved forest
Nitrogen addition reduces soil respiration in a mature tropical forest in southern China
The dynamics of forest litter and its responses to global warming
Forest succession at Dinghushan, Guangdong, China
Nitrogen limitation constrains sustainability of ecosystem response to CO2
Enhanced plant biomass accumulation in response to elevated atmospheric CO2 concentration could dampen the future rate of increase in CO2 levels and associated climate warming. However, it is unknown whether CO2-induced stimulation of plant growth and biomass accumulation will be sustained or whether limited nitrogen (N) availability constrains greater plant growth in a CO2-enriched world. Here we show, after a six-year field study of perennial grassland species grown under ambient and elevated levels of CO2 and N, that low availability of N progressively suppresses the positive response of plant biomass to elevated CO2. Initially, the stimulation of total plant biomass by elevated CO2 was no greater at enriched than at ambient N supply. After four to six years, however, elevated CO2 stimulated plant biomass much less under ambient than enriched N supply. This response was consistent with the temporally divergent effects of elevated CO2 on soil and plant N dynamics at differing levels of N supply. Our results indicate that variability in availability of soil N and deposition of atmospheric N are both likely to influence the response of plant biomass accumulation to elevated atmospheric CO2. Given that limitations to productivity resulting from the insufficient availability of N are widespread in both unmanaged and managed vegetation, soil N supply is probably an important constraint on global terrestrial responses to elevated CO2.
Three years of increased nitrogen deposition do not affect the vegetation of a montane forest ecosystem
Ecosystem properties and forest decline in contrasting long-term chronosequences
Nutrient concentrations in mire vegetation as a measure of nutrient limitation in mire ecosystems
N:P stoichiometry in secondary succession in evergreen broad-leaved forest, Tiantong, East China
Ecological stoichiometry: a science to explore the complexity of living systems
Pools and distributions of soil phosphorus in China
The litter fall of respresentative forests of successional series in Dinghushan
The influence of regional atmospheric precipitation characteristics and its element inputs on the existence and development of Dinghushan forest ecosystems

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}