


热带云雾林指潮湿热带地区经常被云雾掩盖的森林(Stadtmüller, 1987), 主要分布在热带美洲、热带非洲和热带亚洲地区(Bubb et al., 2004)。森林群落高度、外貌、生物多样性和群落结构等都与低海拔热带森林有明显的区别: 树木高度明显减小, 以小径级乔木为主, 植株密度较大, 树干常弯曲, 小叶革质单叶占有最大比重, 类似于旱生生境中的旱生形态(Tanner, 1977; Williams-Linera, 2002; Nomura & Kikuzawa, 2003); 群落物种多样性较低海拔森林偏低(Terborgh, 1977; Hietz & Briones, 1998; Vazquez & Givnish, 1998), 附生苔藓、地衣、蕨类和兰科植物非常丰富。热带云雾林独特的群落结构和多样性与其特殊环境条件密切相关。热带云雾林常呈岛屿状分布于海拔500-3 900 m的山顶或山脊, 因山风强烈、气温低、云雾多, 土壤潮湿而常处于饱和状态(Aldrich et al., 1997)。热带云雾林具有保持水土、调节水源等重要作用, 特别是在相对干旱的季节, 仍能提供比一般森林更多的水源, 对维持生态系统功能有重要意义(Bruijnzeel & Hamilton, 1998); 其林内雾水沉积是海边和较高山地森林生态系统水分和养分循环的重要构成要素(Nadkarni & Solano, 2002; Holder, 2004)。在全球气候变化背景下, 热带云雾林被认为是对气候变化敏感的典型生态系统(Walther et al., 2002), 是世界上受威胁最严重而研究最少的森林(Aldrich et al., 1997)。
热带山地常绿林和热带山顶矮林是我国热带云雾林的主要类型(Bubb et al., 2004), 在我国植被中属于阔叶林植被型亚纲山地常绿苔藓林植被型组(吴征镒, 1995), 主要分布在云南和海南岛(Bubb et al., 2004)。云南热带云雾林群落的物种组成和区系成分等已有研究(施济普, 2007; Shi & Zhu, 2009)。海南岛热带云雾林主要分布于霸王岭、五指山、尖峰岭、吊罗山和鹦哥岭等林区海拔1 200 m以上的山顶地段(胡玉佳和李玉杏, 1992)。蒋有绪和卢俊培(1991)、杨小波等(1994)和余世孝等(2001)分别对尖峰岭、五指山和霸王岭热带云雾林的群落结构特征进行过研究。森林环境条件是影响群落结构、生物多样性和生态系统功能的重要要素(Whitmore, 1990; 吴征镒, 1995), 但目前还没有针对热带云雾林环境研究方面的报道。本文以霸王岭热带云雾林为例, 系统地分析比较了热带云雾林的光照、温度、水分、土壤因子和地形因子等环境特征, 为进一步深入分析热带云雾林的生物多样性、生态系统功能及其对气候变化的响应等生态学过程奠定基础, 并为森林生态系统管理提供基础性资料。
1 研究地点和研究方法
1.1 研究区概况、样地概况
研究样地位于海南省昌江县和白沙县境内的霸王岭国家级自然保护区。霸王岭自然保护区总面积约50 000 hm2, 地理坐标为18º50′-19º05′ N, 109º05′-109º25′ E。以山地为主, 海拔100-1 654 m。该地区属热带季风气候, 干湿季明显, 5-10月为雨季, 11月至次年4月为旱季。海拔100 m位置年平均气温23.6 ℃, 年平均降水量1 677.1 mm (Zang et al., 2005)。随着海拔增加, 植被依次为热带低地雨林、热带山地雨林、热带山地常绿林和热带山顶矮林。土壤以砖红壤为代表类型, 随着海拔增加逐渐过渡为山地红壤、山地黄壤和山地草甸土(胡玉佳和李玉杏, 1992)。样地分别设置在热带山地常绿林(tropical montane evergreen forest, TMEF)和热带山顶矮林(tropical montane dwarf forest, TMDF)。热带山地常绿林设置两个样地(地理坐标: TMEF1, 19º05′24.5″ N, 109º12′56.2″ E; TMEF2, 19º05′33.2″ N, 109º12′ 53.2″ E), 每个样地随机设置4个2 500 m2的样方。热带山顶矮林设置3个样地(地理坐标: TMDF1, 19º05′04.8″ N, 109º12′43.5″ E; TMDF2, 19º05′12.4″ N, 109º12′36.4″ E; TMDF3, 19º05′57″ N, 109º12′54.8″ E), 3个样地分别随机设置4个、5个和2个2 500 m2的样方。不同样地间水平距离在100 m以上, 所有样地的坡向为东坡。热带山地常绿林和热带山顶矮林都为原始林, 两者样地概况见表1。TMEF的优势树种有陆均松(Dacrydium pierrei)、岭南青冈(Cyclobalanopsis championii)、大果马蹄荷(Exbucklandia tonkinensis)、碟斗青冈(C. disciformis)、厚皮香(Ternstroemia gymnanthera)和木荷(Schima superba); TMDF的优势树种有蚊母树(Distylium racemosum)、丛花山矾(Symplocospoilanei)、海南五针松(Pinus fenzeliana)、碎叶蒲桃(Syzygium buxifolium)、乌心樟(Cinnamomum tsoi)、黄杞(Engelhardtia roxburghiana)和毛棉杜鹃花(Rhododendron moulmainense)等。
表1 热带山地常绿林和热带山顶矮林研究样地比较(平均值±标准偏差)
Table 1
| 研究样地 Study site | 海拔 Altitude (m) | 郁闭度 Canopy density | 平均树高 Mean tree height (m) | 植株密度 Stem density (stems·100 m-2) | 基面积 Basal area (m2·100 m-2) | 物种丰富度 Species richness |
|---|---|---|---|---|---|---|
| 热带山地常绿林 TMEF | 1 200 ± 4.34 | >0.7 | 5.55 ± 0.59 | 52.2 ± 6.4 | 0.43 ± 0.16 | 18.6 ± 2.3 |
| 热带山顶矮林 TMDF | 1 300 ± 8.52 | 0.5-0.7 | 4.01 ± 0.58 | 115.3 ± 24.5 | 0.25 ± 0.09 | 29.7 ± 1.5 |
所调查的植株胸径均在1 cm以上。
Data were obtained based on trees with diameter at breast height ≥ 1 cm.
1.2 研究方法
1.2.1 光合有效辐射数据采集
光合有效辐射(photosynthetically active radiation, PAR)数据在2009年6月5日无云的全日晴天采集。用2个Field Scount光量子计(3415FSE, Spectrum Technologies Inc., New Jersey, USA), 从8:00到16:00每隔2 h采一次样。采样时, 沿每个样方对角线均匀设定5个点, 手持Field Scount光量子计在每个点距离地面1.3 m的地方测定PAR。
1.2.2 空气温度、相对湿度测定
温度、湿度数据采集时间从2009年5月至10月。将5个HOBO Pro温湿度自动记录仪(HOBO U23- 001, Onset, MA, USA)分别放置在上述样地中间, 温度、湿度仪绑定在离地面1.3 m处, 每隔1 h自动测定, 并同时记录空气温度和相对湿度。
1.2.3 土壤数据的采集
土壤数据于2009年雨季采集。每个样方中土壤采集样点与PAR测定样点相同, 每个样方采集5个土样。去除土壤表层枯枝落叶, 挖深0.2 m剖面, 先根据土壤颜色、颗粒大小、黏性等物理性质判断腐殖质层, 并测定其厚度; 然后自上而下取20 cm混合土样; 最后在取土位置用自制的1.5 m钢钎打入土壤, 直至到达土壤母岩, 土壤厚度即为没入土壤的钢钎长度。所有土样自然风干后测定其成分含量: 有机质用高温外热重铬酸钾氧化-容量法测定; 全磷的测定方法是用HClO4-H2SO4消化法分解样品, 然后用钼锑抗比色法测定。全氮、全钾含量和pH值分别用凯氏定氮法、NaOH熔融-火焰光度法和电位法测定。水解性氮、速效磷和速效钾分别用碱解扩散法、盐酸-氟化铵法和乙酸铵提取火焰光度法测定(中国土壤学会农业化学专业委员会, 1983)。
1.2.4 地形数据的采集
地形数据包括坡度、岩石裸露比例和海拔高度。坡度用坡度坡向仪测定, 估算裸露出地表的石头面积占样方面积的比例即为岩石裸露比例, 海拔高度用海拔仪测定。
1.2.5 数据处理和分析
计算热带山地常绿林和热带山顶矮林一天中不同时段的平均PAR、不同月份日平均空气温度和相对湿度、各土壤成分的平均值、各地形因子的平均值(±标准偏差)。岩石裸露比例和相对湿度数据用反正弦变换。同一天不同时段的PAR差异及不同月份的日平均温度湿度差异用One-way ANOVAs检验, 并用Tukey-Kramer HSD检验进行多重比较。两种森林类型间土壤因子和地形因子的差异用Wilcoxcon检验。不同环境因子间的作用大小用主成分分析(PCA)后, 根据每个变量的负荷值进行比较。首先建立样地×环境变量矩阵, 然后用R2.9.2中Vegan包导入矩阵分析, 当累积解释方差比例达到80%时, 取前面所有排序轴分析, 并根据各环境因子在PCA轴上负荷的大小判断其作用大小。环境因子间的相关性用Pearson相关性分析, 相关性大小用Student’s t检验。所有数据统计分析在R2.9.2中ad4、ape和vegan包进行(R Development Core Team, 2009)。
2 研究结果
2.1 PAR和空气温度湿度特征
热带山地常绿林和热带山顶矮林的PAR在一天中的不同时段有显著差异(山地常绿林: F(3,28) = 14.48, p < 0.001; 热带山顶矮林: F(3,40) = 4.61, p = 0.007), 且呈单峰曲线分布(图1A), 10:00-12:00, PAR最强。多重比较的结果表明: 热带山地常绿林8:00-10:00与10:00-12:00、10:00-12:00与14:00- 16:00、12:00-14:00与14:00-16:00时段PAR有显著差异(p = 0.001; p < 0.001; p = 0.004), 而其他时段间无显著差异(p = 0.15; p = 0.37; p = 0.07); 热带山地矮林8:00-10:00与10:00-12:00、10:00-12:00与14:00- 16:00时段PAR有显著差异(p = 0.012; p = 0.016), 其他时段间无显著差异(p = 0.99; p = 0.88; p = 0.92; p = 0.07)。热带山地常绿林一天中各个时段的PAR都比热带山顶矮林显著低(图1A)。
图1
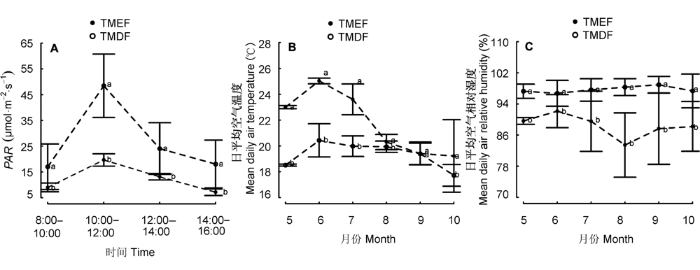
图1
热带山地常绿林(TMEF)和热带山顶矮林(TMDF)间光合有效辐射(PAR)和日平均空气温度湿度的比较(平均值±标准误差)。A, 光合有效辐射。B, 日平均空气温度。C, 日平均空气相对湿度。不同字母(a, b)表示两者有显著差异(p < 0.05)。
Fig. 1
Comparisons in photosynthetically active radiation (PAR), mean daily air temperature and air relative humidity between tropical montane evergreen forest (TMEF) and tropical montane dwarf forest (TMDF) (mean ± SE). A, Photosynthetically active radiation. B, Mean daily air temperature. C, Mean daily relative humidity. Different letters (a, b) indicate significant difference at p < 0.05.
热带山地常绿林和热带山顶矮林在不同月份的日平均空气温度有极显著差异(热带山地常绿林: F(5,354) = 188.7, p < 0.001; 热带山顶矮林: F(5,534) = 147.6, p < 0.001), 且呈单峰曲线分布(图1B), 6月份日平均空气温度最高。多重比较结果显示, 热带山地常绿林6月与8月、6月与9月及6月与10月日平均空气温度有极显著差异(p < 0.001); 而8-10月的日平均空气温度无显著差异(p = 0.20; p = 0.09; p = 0.98); 热带山顶矮林除了7月与8月外(p = 0.98), 其他月份间都有极显著差异(p < 0.001)。5-10月热带山地常绿林和热带山顶矮林的平均空气温度分别为(21.76 ± 2.44) ℃和(19.33 ± 1.03) ℃, 除9月份, 热带山地常绿林各月的平均空气温度均比热带山顶矮林显著高。
比较不同月份日平均空气相对湿度, 热带山地常绿林和热带山顶矮林均有显著差异(热带山地常绿林: F(5,354) = 5.70, p = 0.001; 热带山顶矮林: F(5,534) = 4.74, p < 0.001), 且呈倒“S”型分布(图1C)。多重比较结果表明, 热带山地常绿林日平均空气相对湿度在7月与8月及8月与10月有显著差异(p = 0.01; p = 0.001), 其他月份间无显著差异(p > 0.05); 热带山顶矮林日平均空气相对湿度在7月与9月及9月与10月有显著差异(p < 0.001; p = 0.04), 而其他月份间无显著差异(p > 0.05)。两种森林类型比较, 5-10月热带山地常绿林和热带山顶矮林平均空气相对湿度分别为(88.44 ± 2.90)%和(97.71 ± 0.80)%, 热带山地常绿林各月日平均空气相对湿度均比热带山顶矮林显著小。
2.2 土壤和地形特征
表2 热带山地常绿林和热带山顶矮林土壤因子比较(平均值±标准偏差)
Table 2
| 土壤因子 Soil factor | 植被类型 Forest type | 土壤因子 Soil factor | 植被类型 Forest type | |||
|---|---|---|---|---|---|---|
| 热带山地常绿林 TMEF | 热带山顶矮林 TMDF | 热带山地常绿林 TMEF | 热带山顶矮林 TMDF | |||
| 全氮 Total nitrogen (g·kg-1) | 2.25 ± 0.59a | 1.15 ± 0.38b | 有效钾 Available potassium (mg·kg-1) | 21.97 ± 1.39a | 31.97 ± 13.79a | |
| 全磷 Total phosphorous (g·kg-1) | 0.79 ± 0.14a | 0.49 ± 0.32b | 有机质 Organic matter (g·kg-1) | 64.87 ± 22.23a | 36.71 ± 12.99b | |
| 全钾 Total potassium (g·kg-1) | 12.68 ± 1.67b | 65.31 ± 12.50a | pH | 4.44 ± 0.29a | 3.96 ± 0.19b | |
| 速效氮 Available nitrogen (mg·kg-1) | 153.02 ± 28.10a | 84.50 ± 15.44b | 腐殖质厚度 Humus thickness (cm) | 8.75 ± 0.67a | 9.15 ± 1.22a | |
| 有效磷 Available phosphorous (mg·kg-1) | 11.00 ± 1.65b | 19.46 ± 4.25a | 土壤厚度 Soil thickness (cm) | 65.35 ± 8.63a | 54.90 ± 8.55b | |
不同字母表示有显著差异(p < 0.05)。
Different letters at the top right corner of mean values indicate significant difference at p < 0.05.
图2
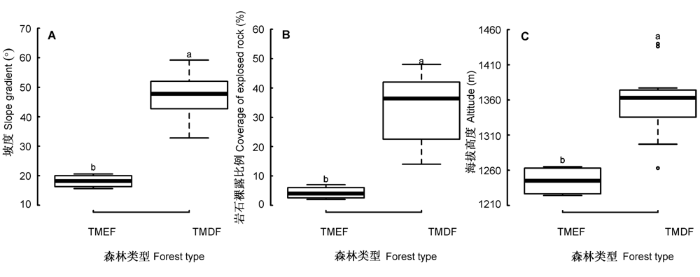
图2
热带山地常绿林(TMEF)和热带山顶矮林(TMDF)地形因子的比较。A, 坡度。B, 岩石裸露比例。C, 海拔高度。箱线图上不同字母(a, b)表示两者有显著差异(p < 0.05)。
Fig. 2
Comparison in topographic conditions between tropical montane evergreen forest (TMEF) and tropical montane dwarf forest (TMDF).A, Slope. B, Coverage of exposed rock. C, Altitude. Different letters (a, b) above the boxplot indicate significant difference at p < 0.05.
2.3 环境因子主成分分析(PCA)
图3
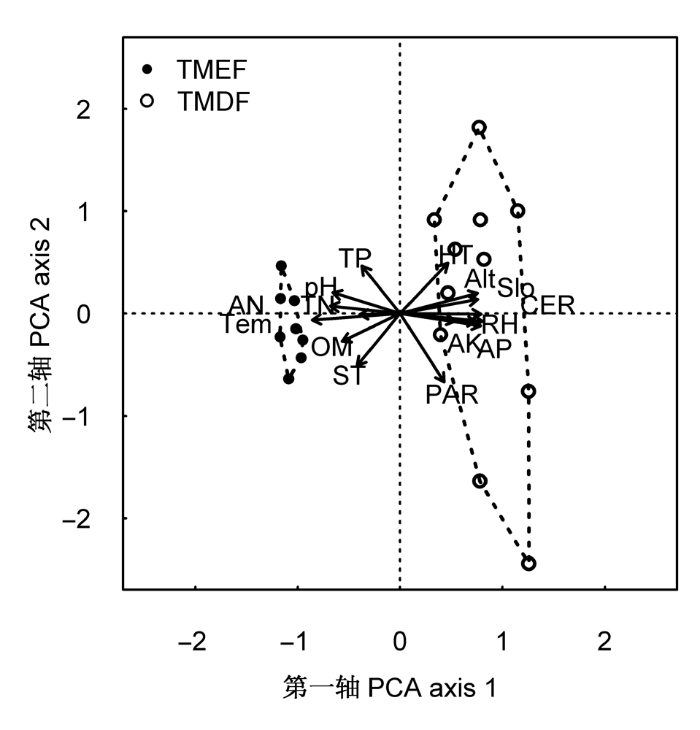
图3
热带山地常绿林(TMEF)和热带山顶矮林(TMDF)样方随环境变量变化的PCA排序图。AK, 有效钾; Alt, 海拔高度; AN, 速效氮; AP, 有效磷; CER, 岩石裸露比例; HT, 腐殖质厚度; OM, 有机质; PAR, 光合有效辐射; RH, 日平均空气相对湿度; Slo, 坡度; ST, 土壤厚度; Tem, 日平均空气温度; TK, 全钾; TN, 全氮; TP, 全磷。
Fig. 3
Biplot of principal component analysis (PCA) showing changes in environmental variables across tropical montane evergreen forest (TMEF) and tropical montane dwarf forest (TMDF).AK, available potassium; Alt, altitude; AN, available nitrogen; AP, available phosphorus; CER, coverage of exposed rock; HT, humus thickness; OM, organic matter; PAR, photosynthetically active radiation; RH, mean daily air relative humidity; Slo, slope gradient; ST, soil thickness; Tem, mean daily air temperature; TK, total potassium; TN, total nitrogen; TP, total phosphorus.
表3 主成分分析(PCA)中各环境变量在前两个排序轴的负荷值及解释方差
Table 3
| 环境变量 Environmental variables | PCA1 | PCA2 | PCA3 | PCA4 |
|---|---|---|---|---|
| 海拔高度 Altitude (m) | 0.86 | 0.23 | -0.32 | 0.07 |
| 日平均空气相对湿度 Mean daily air relative humidity (%) | 0.85 | -0.10 | -0.45 | -0.01 |
| 日平均空气温度 Mean daily air temperature (℃) | -0.98 | -0.07 | -0.12 | -0.004 |
| 坡度 Slope gradient (º) | 0.90 | -0.01 | 0.34 | 0.11 |
| 岩石裸露比例 Coverage of exposed rock (%) | 0.92 | -0.09 | -0.28 | 0.06 |
| 土壤厚度 Soil thickness (cm) | -0.48 | -0.58 | 0.34 | 0.23 |
| 腐殖质厚度 Humus thickness (cm) | 0.53 | 0.56 | 0.46 | 0.17 |
| 光合有效辐射 Photosynthetically active radiation, PAR (µmol∙m-2∙s-1) | 0.49 | -0.76 | -0.15 | 0.04 |
| 全氮 Total nitrogen (g∙kg-1) | -0.78 | 0.08 | -0.27 | 0.25 |
| 全磷 Total phosphorus (g∙kg-1) | -0.42 | 0.53 | -0.53 | 0.30 |
| 全钾 Total potassium (g∙kg-1) | 0.87 | 0.16 | 0.34 | 0.001 |
| 速效氮 Available nitrogen (mg∙kg-1) | -0.45 | -0.01 | -0.15 | -0.51 |
| 有效磷 Available phosphorus (mg∙kg-1) | 0.90 | -0.14 | -0.33 | -0.002 |
| 有效钾 Available potassium (mg∙kg-1) | 0.61 | -0.08 | -0.16 | 0.11 |
| 有机质 Organic matter (g∙kg-1) | -0.65 | -0.31 | -0.07 | 0.43 |
| pH | -0.75 | 0.24 | -0.05 | 0.04 |
| 特征值 Eigenvalue | 8.76 | 1.80 | 1.49 | 1.11 |
| 方差比例 Proportion of variance | 0.547 | 0.113 | 0.090 | 0.070 |
| 累积方差比例 Proportion of cumulative variance | 0.547 | 0.660 | 0.753 | 0.822 |
加粗数字表示该变量的负荷绝对值大于0.75。
Bold data indicate that the absolute loadings of environmental variables are over 0.75.
2.4 环境因子相关性
图4
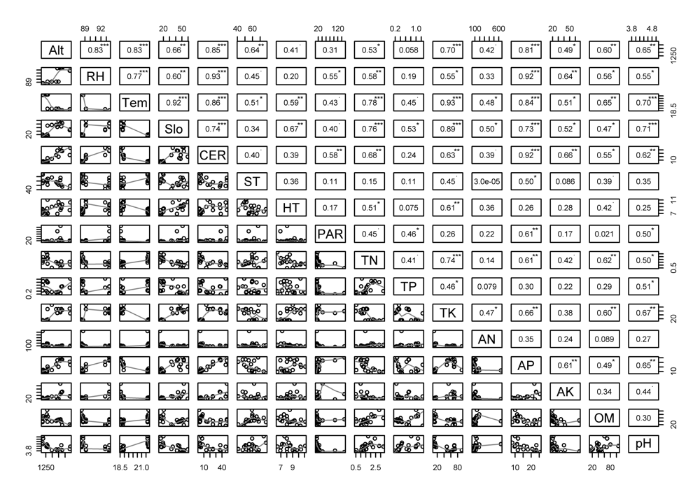
图4
热带山地常绿林(TMEF)和热带山顶矮林(TMDF)环境因子间的相关性分析: 相关图解为环境因子间的相关系数及其显著性。该图是将热带山地常绿林和热带山顶矮林的环境因子数据合并后的相关性分析结果。AK, 有效钾(mg·kg-1); Alt, 海拔高度(m); AN, 速效氮(mg·kg-1); AP, 有效磷(mg·kg-1); CER, 岩石裸露比例(%); HT, 腐殖质厚度(cm); OM, 有机质(g·kg-1); PAR, 光合有效辐射(µmol∙m-2∙s-1); RH, 日平均空气相对湿度(%); Slo, 坡度(º); ST, 土壤厚度(cm); Tem, 日平均空气温度(℃); TK, 全钾(g·kg-1); TN, 全氮(g·kg-1); TP, 全磷(g·kg-1)。*, p < 0.05; **, p < 0.01; ***, p < 0.001; 无*的数字表示 p > 0.05。
Fig. 4
Correlation among environmental conditions in both tropical montane evergreen forest (TMEF) and tropical montane dwarf forest (TMDF), indicating correlation coefficients of environmental factors and their significance. The figure was obtained with correlation analysis after combining environmental data of TMEF and TMDF.AK, available potassium (mg·kg-1); Alt, altitude (m); AN, available nitrogen (mg·kg-1); AP, available phosphorous (mg·kg-1); CER, coverage of exposed rock (%); HT, humus thickness (cm); OM, organic matter (g·kg-1); PAR, photosynthetically active radiation (µmol ∙ m-2 ∙ s-1); RH, mean daily air relative humidity (%); Slo, slope gradient (º); ST, soil thickness (cm); Tem, mean daily air temperature (℃); TK, total potassium (g·kg-1); TN, total nitrogen (g·kg-1); TP, total phosphorus (g·kg-1). *, p < 0.05; **, p < 0.01; ***, p < 0.001; figures without * indicate p > 0.05.
3 讨论
3.1 热带云雾林的划分
能否把热带山地常绿林和热带山顶矮林划为热带云雾林, 对此尚存在争议。有学者认为: 热带山地常绿林分布带很窄, 在植物区系的组成上是由热带山地雨林向热带山顶苔藓矮林过渡的类型, 很多种类属于热带山地雨林的共有种类, 因此在植被垂直分布梯度上应归入热带山地雨林类型(陈树培, 1982; 黄全等, 1986)。本文根据国际惯用的方法, 从森林环境角度划分热带云雾林, 即云雾出现频度高、空气湿度大的森林为热带云雾林(Stadtmüller, 1987; Bubb et al., 2004)。根据本研究结果, 热带山地常绿林和热带山顶矮林云雾出现频率都较高, 5-10月热带山地常绿林和热带山顶矮林日平均空气相对湿度在88%以上, 且在98天以上的时间内空气湿度达到100%。一些学者也把热带山地常绿林从热带山地雨林中单列出来, 称为热带山地苔藓林或热带山地常绿林, 并认为它们比热带山地雨林云雾多、湿度大(陆阳等, 1986; 余世孝等, 1993)。在另一些研究中, 热带山地常绿林也被称为热带云雾林(余世孝等, 2001)。
3.2 热带云雾林的光照特征
调查发现: 霸王岭热带云雾林地处海拔较高的山脊地带, 且主要分布在东坡, 林冠光照充足, 一天不同时段的光合有效辐射呈单峰曲线。热带山顶矮林下的光照显著高于热带山地常绿林(图1A), 主要原因有以下3个方面: 首先, 与热带山地常绿林树木平均高度(5.55 ± 0.59) m大于热带山顶矮林树木平均高度(4.01 ± 0.58) m)有关。研究表明, 透过树叶的光照强度往往从林冠到林下呈指数级数递减(Chazdon & Fetcher, 1984; Kitajima et al., 2005), 随着树高增加, 林下可利用光呈垂直递减趋势(Monsi & Saeki, 2005)。其次, 与热带山地常绿林冠层郁闭度(> 70%)大于热带山顶矮林冠层郁闭度(50%-70%)有关。调查发现, 热带云雾林群落的郁闭度与林下光合有效辐射显著负相关(r = -0.56, p = 0.01), 郁闭热带山地常绿林能遮挡大部分冠层光照, 使穿透到林下的光照较弱。最后, 与地形有关, 光合有效辐射与坡度显著正相关(图4)。由于热带山地常绿林坡度显著小于热带山顶矮林坡度(图2A), 热带山地常绿林群落的乔木冠层容易互相重叠遮挡光照, 而热带山顶矮林冠层重叠少, 光照易穿透冠层到达地表, 因而林下光照强。主成分分析表明, 光合有效辐射是影响热带云雾林植被分布的重要因素(图3), 光照对于云雾林植物的生长和更新、物种分布、植物功能性状变异等有重要意义(Emborg, 1998; Ackerly et al., 2002; Ostertag et al., 2008)。
3.3 热带云雾林的空气温度和湿度特征
由于热带云雾林分布海拔高, 不同月份的日平均空气温度差异大(图1B)。研究发现, 除9月份外, 热带山地常绿林各月份日平均空气温度都比热带山顶矮林高。这与前者分布的海拔比后者低有关(图2C), 空气温度与海拔高度显著负相关(图4)。热带云雾林空气温度还可能与空气湿度有关, 本研究中两者呈显著负相关关系。湿润环境往往对温度有较强的调节能力, 相对而言, 干燥的热带山地常绿林对空气温度的调节能力较差, 因而温度较高。与低海拔的东二林场(109°10′31.865″ E, 19°6′39.442″ N, 海拔905 m)和南岔河(109°11′57.251″ E, 19°6′38.983″ N, 海拔 600 m) (中国林业科学研究院热带林业研究所的观测数据, 未发表)比较, 东二林场和南岔河5-10月的日平均温度分别为(22.24 ± 1.15) ℃和(24.40 ± 1.38) ℃, 比热带山地常绿林和热带山顶矮林日平均空气温度显著高(One-way ANOVA检验, F(3,20) = 10.14, p < 0.001)。相关文献也证实, 热带云雾林的空气温度比低海拔植被低(胡玉佳和李玉杏, 1992; 杨小波等, 1994; 黄世能等, 2000)。主成分分析和相关性分析表明, 空气温度负荷最大, 且与其他环境因子显著相关(表3; 图4)。这证实了温度是高海拔森林群落环境因子影响植物生长、储存水分和养分能力的最重要因素(Körner, 1989; Roche et al., 2004)。与云南地区热带云雾林相比, 海南岛热带云雾林由于所处海拔较低, 所以空气温度较高(Shi & Zhu, 2009)。
5-10月热带山地常绿林和热带山顶矮林日平均空气相对湿度在88%以上, 显示其群落环境湿润特点(Aldrich et al., 1997)。较高空气相对湿度与高频率云雾弥漫有关。经调查发现, 在5-10月186天中, 热带山地常绿林和热带山顶矮林各自平均有98天和152天空气湿度达到100%。饱和的空气形成水滴或水平降水, 一方面为森林动植物特别是附生的苔藓、地衣、蕨类和兰科植物提供充足的养分和水分(Holder, 2004), 另一方面为低海拔森林提供充足的水源(Bruijnzeel & Hamilton, 1998), 也促进海边和较高山地森林生态系统水分和养分的循环(Nadkarni & Solano, 2002; Holder, 2004)。热带山地常绿林的空气相对湿度比热带山顶矮林小, 这可能与海拔较低有关(图4)。与低海拔的东二林场和南岔河山地雨林比较(中国林业科学研究院热带林业研究所观测数据, 未发表), 东二林场和南岔河5-10月的日平均相对湿度分别为(85.72 ± 3.04)%和(81.94 ± 5.14)%, 显著低于热带山地常绿林和热带山顶矮林的日平均空气相对湿度(One-way ANOVA检验, F(3,20) = 43.97, p < 0.001)。
3.4 热带云雾林的土壤特征
热带山地常绿林的土壤肥力比热带山顶矮林高(表2)。热带山地常绿林的有机质、全氮、全磷和速效氮含量都比热带山顶矮林高, 这可能与热带山地常绿林群落的物种丰富、结构复杂、地面凋落物多有关。而且, 热带山地常绿林的空气温度相对较高, 可能土壤内微生物活动较强, 分解枯枝落叶的能力较强, 从而使得土壤有机质含量相对较大。热带山地常绿林和热带山顶矮林的土壤肥力差异, 也将为植物提供不同水平的营养来源, 物种间因资源水平差异形成不同的竞争格局, 从而影响植物的多样性(Huston, 1980; Gentry, 1988)。另外, 主成分和相关性分析表明, 磷有较高负荷且与其他环境因子密切相关(图3, 图4), 这说明土壤磷元素可能是影响热带云雾林植被分布的重要因子。磷在热带云雾林群落成为限制性因子, 可能是由于土壤呈酸性(表2), 土壤中H2PO4-2易与Al3+和Fe3+形成难溶复合物而不易被植物吸收(Bohn et al., 2001)。与霸王岭热带山地雨林的土壤成分比较(邓福英, 2007), 热带山地雨林的土壤速效氮、有效钾含量和pH显著高于热带云雾林(W = 2, p < 0.001; W = 0, p < 0.001; W = 2, p = 0.001), 而全磷和有效磷含量显著低于热带云雾林(Wilcoxcon检验, W = 75, p = 0.02; W = 72, p = 0.04); 其他土壤因子差异不显著。与云南苏典、黄连山和分水岭的热带云雾林(施济普, 2007)比较, 云南地区热带云雾林的土壤全氮、速效氮和有机质含量都比霸王岭热带云雾林高(Wilcoxcon检验, 全氮: W = 0, p = 0.002; 速效氮: W = 0, p = 0.002; 有机质: W = 0, p < 0.001), 而两者间的全磷、有效磷及pH无显著差异, 整体上云南地区热带云雾林的土壤肥力较高。
总之, 热带云雾林雨季的环境独特: 一天中光合有效辐射呈单峰曲线变化; 日平均空气温度在22 ℃以下, 呈单峰曲线变化; 空气相对湿度在88%以上, 呈倒“S”型曲线变化。与热带山地雨林比较, 热带云雾林坡度和海拔高度较大, 土壤肥力较低。其中, 空气温度、有效磷、全钾、全氮及地形因子对热带云雾林植被分布有重要影响。本文主要研究热带云雾林雨季环境特征, 后续研究可对旱季及多年的环境参数进行监测, 为研究热带云雾林生物多样性、生态系统功能及其对气候变化的响应等生态学过程奠定基础。
致谢
国家自然科学基金(30430570)、中国林业科学研究院森林生态与环境保护研究所基金(RIFEEP200901)和香港嘉道理农场与植物园奖学金资助。感谢霸王岭自然保护区杨秀森高工和王进强、海南大学杨小波教授、李东海副教授和杨福孙博士在野外调查中提供的帮助。感谢中国林业科学研究院热带林业研究所提供相关数据。
参考文献
Leaf size, specific leaf area and microhabitat distribution of chaparral woody plants: constrasting patterns in species level and community level analyses

We examined variation in leaf size and specific leaf area (SLA) in relation to the distribution of 22 chaparral shrub species on small-scale gradients of aspect and elevation. Potential incident solar radiation (insolation) was estimated from a geographic information system to quantify microclimate affinities of these species across north- and south-facing slopes. At the community level, leaf size and SLA both declined with increasing insolation, based on average trait values for the species found in plots along the gradient. However, leaf size and SLA were not significantly correlated across species, suggesting that these two traits are decoupled and associated with different aspects of performance along this environmental gradient. For individual species, SLA was negatively correlated with species distributions along the insolation gradient, and was significantly lower in evergreen versus deciduous species. Leaf size exhibited a negative but non-significant trend in relation to insolation distribution of individual species. At the community level, variance in leaf size increased with increasing insolation. For individual species, there was a greater range of leaf size on south-facing slopes, while there was an absence of small-leaved species on north-facing slopes. These results demonstrate that analyses of plant functional traits along environmental gradients based on community level averages may obscure important aspects of trait variation and distribution among the constituent species.
Tropical Montane Cloud Forests: an Urgent Priority for Conservation
Climatic conditions and tropical montane forest productivity: the fog has not lifted yet the structure and functioning of montane tropical forest: control by climate, soils and disturbance
Photosynthetic light environments in a lowland tropical rainforest in Costa-Rica
The vegetation and vegetational regionalization of Ledong Country, Hainan Island
Classification of Plant Functional Groups and Identification of the Ecological Keystone Species in the Tropical Montane Rain Forest of Hainan Island, China
(海南岛热带山地雨林植物功能群划分及生态关键种的确定)
Understory light conditions and regeneration with respect to the structural dynamics of a near-natural temperate deciduous forest in Denmark
Changes in plant community diversity and floristic composition on environmental and geographical gradients
Correlation between water relations and within-canopy distribution of epiphytic ferns in a Mexican cloud forest
Rainfall interception and fog precipitation in a tropical montane cloud forest of Guatemala
Study of tropical vegetation series in Jianfengling Region, Hainan Island
Composition and geographical elements of the spermatophytic flora from Jianfengling area of Hainan Island
Soil nutrients and tree species richness in Costa Rican forests
Variation in crown light utilization characteristics among tropical canopy trees
The nutritional status of plants from high altitudes. A worldwide comparison
Are plants at high altitudes short in nutrients? In order to answer this question the mineral nutrient content of leaves from over 150 plant species from 9 different mountain areas of all major climatic zones were analyzed (Kjeldahl nitrogen in all, phosphate in half of the samples, K, Mg, Mn, Ca in the Alps only). The majority of data are from herbaceous perennials, but shrubs and trees were studied as well. N-partitioning was studied in 45 herbaceous species from contrasting altitudes in the Alps. The survey falls into three categories: (1) comparisons of whole communities of species from contrasting altitudes, (2) analysis of altitudinal gradients, and (3) additional collections from high altitude sites alone. Unlike the other mineral nutrients, nitrogen content follows consistent altitudinal and latitudinal trends. The higher altitude sample always had higher N content per unit leaf area, irrespective of life form, wherever comparable plants (the same or related species) were investigated at contrasting altitudes. N content per unit dry weight (%) increased with altitude in herbaceous plants (in some species >4%), but was remarkably stable in evergreen woody plants (around 1%). The mean fraction of total plant N allocated to leaves of herbaceous plants in the Alps was the same at low and high altitude (1/3 of total). Leaf N (%) from the regional upper limits of higher plant life reveals a latitudinal decrease from subarctic to equatorial mountains, which may be related to the duration of annual leaf activity. Since mean N content per leaf area hardly differs between the uppermost sites, life span expectation (sink-duration) seems to control carbon investments rather than N input per leaf area. The growth of leaves at high altitude seems to be controlled in a way that leads to comparatively high nutrient contents, which in turn support high metabolic activity. Inherent developmental growth constraints inhibit nutrient dilution in the plant body and thus defy the application of classical concepts of plant-nutrient versus soil-nutrient relations developed for lowlands and in particular for cultivated plants. The results re-emphasize the global significance of links between nitrogen content, leaf sclerophylly, leaf longevity and photosynthetic capacity.
Vegetation of Bawangling Gibbon Natural Reserve in Hainan Island
On the factor light in plant communities and its importance for matter production
Potential effects of climate change on canopy communities in a tropical cloud forest: an experimental approach
Global climate change models predict reduced cloud water in tropical montane forests. To test the effects of reduced cloud water on epiphytes, plants that are tightly coupled to atmospheric inputs, we transplanted epiphytes and their arboreal soil from upper cloud forest trees to trees at slightly lower elevations that are naturally exposed to less cloud water. Control plants moved between trees within the upper site showed no transplantation effects, but experimental plants at lower sites had significantly higher leaf mortality, lower leaf production, and reduced longevity. After the epiphytes died, seedlings of terrestrial gap-colonizing tree species grew from the seed banks within the residual mats of arboreal soil. Greenhouse experiments confirmed that the death of epiphytes can result in radical compositional changes of canopy communities. Thus, tropical montane epiphyte communities constitute both a potentially powerful tool for detecting climate changes and a rich arena to study plant/soil/seed interactions under natural and manipulated conditions. This study also provides experimental evidence that the potential effects of global climate change on canopy and terrestrial communities can be significant for cloud forest biota. Results suggest there will be negative effects on the productivity and longevity of particular epiphytes and a subsequent emergence of an emerging terrestrial component into the canopy community from a previously suppressed seed bank.
Productive phenology of tropical montane forest: fertilization experiment along a moisture gradient
Understory colonization of Eucalyptus plantations in Hawaii in relation to light and nutrient levels
R: A Language and Environment for Statistical Computing
Congruency analysis of species ranking based on leaf traits: Which traits are the more reliable?
Community Ecology and Biogeography of the Mossy Dwarf Forest in Yunnan
(云南山顶苔藓矮林群落生态学与生物地理学研究)
Tree species composition and diversity of tropical mountain cloud forest in the Yunnan, southwestern China
Cloud Forest in the Humid Tropics: A Bibliographic Review
Four montane rain forests of Jamaica: a quantitative characterization of the floristics, the soils and the foloar mineral levels and a discussion of the interrelation
Bird species diversity on an Andean elevational gradient
Altitudinal gradients in tropical forest composition, structure, and diversity in the Sierra de Manantlan
Ecological responses to recent climate change
There is now ample evidence of the ecological impacts of recent climate change, from polar terrestrial to tropical marine environments. The responses of both flora and fauna span an array of ecosystems and organizational hierarchies, from the species to the community levels. Despite continued uncertainty as to community and ecosystem trajectories under global change, our review exposes a coherent pattern of ecological change across systems. Although we are only at an early stage in the projected trends of global warming, ecological responses to recent climate change are already clearly visible.
Tree species richness complementarily, disturbance and fragmentation in a Mexican tropical montane cloud forest
The forest vegetation of Wuzhishan Mountain in Hainan Island. I. The forest vegetation types of Wuzhishan Mountain
Spatial analysis of species diversity in the tropical vegetations along the vertical belt at Bawangling Natural Reserve, Hainan Island
The tropical montane rain forest of Bawangling Natural Reserve, Hainan Island. I. The permanent plots and the community types
Within community patch dynamics in a tropical montane rain forest of Hainan Island, South China

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}