


通常人们认为施肥导致物种丰富度下降是由竞争引起的(Newman, 1973; Tilman, 1982; Grime, 2002; Lan & Bai, 2012)。施肥导致资源供应的不对称性, 因此改变了地上、地下或者是总竞争强度(Newman, 1973; Tilman, 1982), 进而引起物种丰富度的下降。代表性的研究如下: Grime (2002)最早提出多样性下降可能是由于施肥导致地上和地下竞争力增强而引起的; Newman (1973)和Hautier等 (2009)认为施肥导致光竞争增强而引起了物种多样性的降低; 而Dickson和Bryan (2011)认为, 即使光不是限制因子, 施肥也会降低多样性, 主张应结合多个因子解释多样性下降; Rajaniemi (2002)则认为物种多样性下降是由施肥后地下竞争增强引起的; Niu等(2008)研究表明: 施肥明显改变了植物的繁殖分配策略, 最终导致地上和地下竞争力共同作用引起物种丰富度的变化, 有力地支持了总竞争力假说。但也有研究表明施肥后密度发生变化, 进而引起物种多样性的下降。Luo等(2006)的研究表明, 多样性下降主要是由于施肥降低了群落的密度, 密度下降引起了物种丰富度的下降; 而Stevens和Carson (1999b)认为, 施肥后, 小个体植物在光竞争中处于不利地位, 而引起的随机死亡导致了多样性的下降, 有力地支持了群落水平的自疏假说(即密度下降引起了物种丰富度的损失)。但也有人认为是其他方面的原因导致施肥后多样性下降, 例如: Stevens等(2004)的研究表明施肥会引起物种补偿能力的降低(特别是小个体物种), 进而降低了物种丰富度; Gough等(2012)的研究表明, 由于生活型的差异, 使得不同的功能群对施肥的响应也不同; Hillebrand等(2007)的研究表明, 施肥对多样性的影响依赖于群落的均匀度、生态系统的生产力、植物生物量对施肥的响应等众多因素。尽管这些研究都解释了施肥后物种丰富度为什么会下降, 但是还没有形成一个统一的认识。因此要彻底了解物种丰富度对施肥的响应机制, 还面临着巨大的挑战(Abrams, 1995; Waide et al., 1999; Mittelbach et al., 2001)。
CV = σ / μ
CV、σ和μ分别代表植物个体大小的变异系数、标准偏差和平均值。
关于地下竞争的研究报道虽然很多, 但很少有实验直接定量地下竞争。Tilman (1988)用R*定性地预测地下竞争的强弱: 通常R*越低, 其地下竞争强度越强。R*可以用单位植物氮含量表示, 一般认为单位植物氮含量越低, 氮的利用效率就越高, 对土壤氮的竞争力也就越强(van Ruijven & Berendse, 2005)。青藏高原东部高寒草甸主要是受氮限制的(Niu et al., 2008), 为了研究物种对氮竞争力的强弱是否影响多样性, 本实验用单位植物氮含量(mg·g-1)表示该物种的地下氮的竞争力, 通常认为单位植物氮含量越低, 其对土壤氮的竞争力也就越强。本实验拟通过测量物种间单位植物氮含量存在显著差异的物种对数目的变化来检验物种对氮竞争力的强弱如何影响多样性。以上方法使我们能够定性地探索施肥后青藏高原东部高寒草甸物种多样性下降的潜在机制。
在青藏高原东部高寒草甸植物群落, 施肥降低了多样性, 同时增加了生产力, 但是目前对多样性下降的潜在机制并不清楚。本研究试图通过施肥实验来检验经典的竞争理论的重要性, 并对群落多样性的维持和丧失机制做出解释。本研究主要探讨以下两个方面的问题: 施肥后物种多样性下降的原因是什么?放牧是否改变了施肥对物种多样性的影响?
1 材料和方法
1.1 研究地概况
本研究地点在青藏高原东部的兰州大学高寒草甸与湿地生态野外定位研究站的玛曲哇啦卡试验场。该野外定位研究站位于甘肃省玛曲县境内(35.58° N, 101.53° E), 海拔3500 m。年平均气温为1.2 ℃, 最冷月为1月(平均气温-10 ℃), 最热月为 7月(平均气温11.7 ℃)。近35年的平均年降水量是620 mm, 主要分布在短的、湿冷的夏季。本地区年光照时间超过2580 h, 而霜期超过270天, 没有绝对的无霜期。
该地区比较典型的优势物种有: 四川嵩草(Kobresia setchwanensis)、冷地早熟禾(Poa crymophila)、垂穗披碱草(Elymus nutans)和草玉梅(Anemone rivularis)等。平均地上生物量是280-400 g·m-2(干物质生物量), 每0.25 m2有20-35个物种。在1999年10月份, 一块450 m × 220 m的样地在春秋季节被围栏封育。围栏外草场载畜量是0.18头牛·hm-2和3.68只羊·hm-2, 每年的7月中旬到9月中旬, 这些牲畜被转移到夏季牧场去放牧。
1.2 试验设计
本试验利用随机的小区(随机区组)试验设计, 包括两个放牧处理(放牧和封育)和两个施肥处理(0和30 g·m-2)总共4个组合, 每个组合8个重复, 共计32个样方。在2007年5月底, 在放牧地和封育地选定32个小区, 每个小区的大小是8 m × 4 m, 小区间有2 m的缓冲区, 缓冲区不施肥。用(NH4)2HPO4 (天津贸易公司, 天津)确立了两个施肥梯度, 其氮和磷的施用量分别为0、6.3 g N·m-2和0、7.0 g P·m-2。每年的5月底在雨前进行施肥。在2008和2009年, 把每个小区分成两部分, 即: 4 m × 4 m的植被监测区和4 m × 4 m的个体采样区。
1.3 个体取样
本研究选取26个常见物种进行抽样和测量, 这些物种占据了青藏高原东部高寒草甸大于90%的地上生物量。在每个处理的采样区, 于2008年和2009年的7月20号至9月5号(物种花期)采样, 每个物种采20个个体, 每年总共采集2600个(26个物种× 20个个体× 5个处理)个体。由于不可能抽取完整的根部, 因此抽取每个个体的地上部分生物量用以表示该个体的总生物量。对于克隆植物而言, 把每个无性系分株看作一个个体(Luoet al., 2006)。每个个体在实验室内被分为茎﹑叶和果实, 然后在80 ℃下烘干至恒重。最后用天平称出每部分的质量, 精确到10-4g。为了计算物种的叶面积, 在每个处理上, 每个物种抽取了20枚完整的叶, 新鲜的叶在neat image软件下被扫描, 然后在实验室内80 °C下烘干至恒重。最后用天平称出每部分的质量, 精确到10-4g。最后, 用流动注射分析仪(美国路易企业有限公司, 香港)测定地上部分单位植物氮含量(浓度单位: mg·g-1), 每个物种在每个处理上测了3个重复。
1.4 植被监测
在每个小区选取一个0.5 m × 0.5 m的样方。为了防止边缘效应, 限制该样方离边界至少50 cm。在每个小区, 测量物种丰富度和多度, 只有根生长在样方内的个体才被计算。在每年的8月底, 样方内的所有个体被齐地剪取, 并分成4个功能群(禾本科植物、豆科植物、莎草和杂草), 剪取的所有草样在80 ℃下烘干至恒重。最后用天平称出各部分的质量, 精确到10-4g。
1.5 数据分析
用双因素方差分析检验施肥和放牧对物种个体大小(指物种个体平均生物量, 以下同)和物种丰富度的影响。施肥和放牧被看作是固定因素, 个体大小和物种丰富度被看作是因变量。用多重比较方法检验了个体大小和物种丰富度在各处理间的差异。物种高度、单位植物氮含量和物种多度被对数转换以满足方差齐性。利用配对t检验, 检测了在放牧地和封育地施肥对单位植物氮含量和物种高度的影响。利用χ2统计分析检验了放牧地和封育地施肥后单位植物氮含量在物种间的差异。为了说明地下竞争力在物种间的差异, 利用多重比较的方法检验了单位植物氮含量在26个物种间的差异。用Shapiro-Wilk test和 Levene’s test检验方差齐性和正态性。所有的分析都在SPSS 15.0上进行。
2 结果
2.1 施肥对物种丰富度的影响
图1
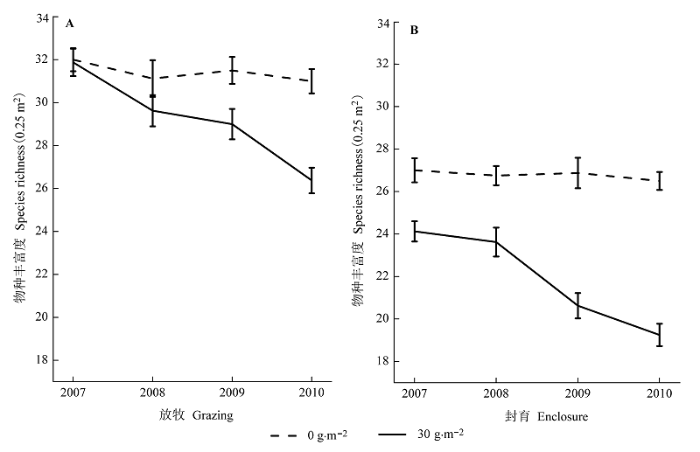
图1
施肥(30 g·m-2)对放牧地(A)和封育地(B)物种丰富度的影响(平均值±标准误差)。
Fig. 1
Effects of fertilization (30 g·m-2) on species richness in both grazed plots (A) and enclosure (B) (mean ± SE).
2.2 施肥对个体大小不整齐性的影响
在封育地, 施肥导致个体大小的不整齐性增加了15% (F1, 30 = 8.019, p < 0.001), 而在放牧地, 施肥并没有改变物种间的个体大小不整齐性( F1,30 = 0.001, p > 0.05) ( 图2)。
图2
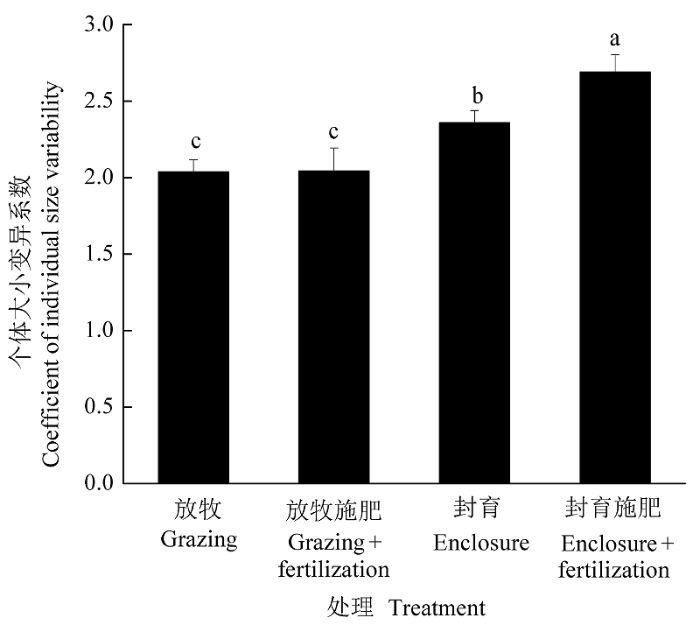
图2
施肥(30 g·m-2)对放牧地和封育地个体大小变异系数的影响(平均值±标准误差)。
不同小写字母表示差异显著(p < 0.05)。
Fig. 2
Effects of fertilization (30 g·m-2) on coefficient of individual size variability both in grazed plots and enclosure (mean ± SE).
Different lowercase letters indicate significant differences among treatments (p < 0.05).
2.3 施肥对物种平均高度的影响
表1 施肥对放牧地和封育地物种高度(2007、2008和2009年的平均值)的影响
Table 1
| 物种 Species | 植物高度 Plant height (cm) | |||||
|---|---|---|---|---|---|---|
| 封育 Enclosure | 封育施肥 Enclosure + fertilization | 放牧 Grazing | 放牧施肥 Grazing + fertilization | |||
| 青藏大戟 Euphorbia altotibetica | 10.5 | 19.9 | 6.9 | 7.8 | ||
| 钝裂银莲花 Anemone obtusiloba | 11.4 | 18.4 | 5.4 | 9.2 | ||
| 垂穗披碱草 Elymus nutans | 40.5 | 66.7 | 26.5 | 24.2 | ||
| 莓叶委陵菜Potentilla fragarioids | 11.0 | 12.0 | 6.6 | 6.1 | ||
| 阿尔泰蒲公英Taraxacum altaicum | 6.5 | 15.9 | 7.1 | 7.5 | ||
| 米口袋Gueldenstaedtia verna | 8.7 | 15.1 | 5.5 | 5.8 | ||
| 蓬子菜Galium verum | 14.1 | 25.3 | 6.6 | 9.0 | ||
| 四川嵩草Kobresia setchwanensis | 34.1 | 42.2 | 21.3 | 25.2 | ||
| 马尔康柴胡 Bupleurum malconense | 20.5 | 20.0 | 7.9 | 6.9 | ||
| 冷地早熟禾Poa crymophila | 33.1 | 56.3 | 16.4 | 18.1 | ||
| 钝苞雪莲Saussurea nigrescens | 13.6 | 14.2 | 6.4 | 7.0 | ||
| 高山韭Allium sikkimense | 17.5 | 27.6 | 8.9 | 14.3 | ||
| 菭草Koeleria cristata | 22.7 | 46.3 | 22.5 | 17.0 | ||
| 细叶亚菊Ajania tenuifolia | 12.7 | 20.6 | 8.2 | 5.5 | ||
| 草玉梅Anemone rivularis | 20.0 | 29.2 | 11.9 | 9.9 | ||
| 花苜蓿Medicago ruthenica | 10.1 | 19.0 | 5.5 | 6.7 | ||
| 椭圆叶花锚Halenia elliptica | 20.1 | 19.4 | 9.9 | 8.9 | ||
| 展毛翠雀花 Delphinium kamaonense | 16.0 | 25.1 | 8.0 | 8.2 | ||
| 丝叶毛茛 Ranunculus tanguticusvar.capillaceus | 13.2 | 23.3 | 7.4 | 8.2 | ||
| 乳白香青Anaphalis lactea | 8.1 | 8.6 | 3.6 | 4.8 | ||
| 甘肃棘豆Oxytropis kansuensis | 12.1 | 16.0 | 7.1 | 6.5 | ||
| 大籽蒿Artemisia sieversiana | 17.1 | 34.4 | 6.2 | 10.1 | ||
| 甘肃马先蒿 Pedicularis kansuensis | 15.4 | 21.8 | 7.6 | 5.9 | ||
| 高原毛茛Ranunculus tanguticus | 10.2 | 14.7 | 6.9 | 7.3 | ||
| 芒剪股颖 Agrostis trinii | 21.9 | 39.2 | 15.0 | 18.6 | ||
| 扁蕾Gentianopsis barbata | 19.6 | 25.4 | 15.5 | 16.9 | ||
表2 施肥对封育地和放牧地组成物种单位植物氮含量(2008和2009年的平均值)的影响
Table 2
| 物种 Species | 平均植物氮含量 Mean plant nitrogen content (mg·g-1) | |||
|---|---|---|---|---|
| 封育 Enclosure | 封育施肥 Enclosure + fertilization | 放牧 Grazing | 放牧施肥 Grazing + fertilization | |
| 青藏大戟 Euphorbia altotibetica | 14.7 | 15.9 | 11.7 | 14.5 |
| 钝裂银莲花 Anemone obtusiloba | 14.5 | 15.8 | 16.5 | 16.2 |
| 垂穗披碱草 Elymus nutans | 7.6 | 7.5 | 11.5 | 10.0 |
| 莓叶委陵菜Potentilla fragarioids | 13.3 | 13.8 | 13.0 | 15.3 |
| 阿尔泰蒲公英Taraxacum altaicum | 15.5 | 17.3 | 18.1 | 17.0 |
| 米口袋Gueldenstaedtia verna | 21.6 | 24.9 | 24.8 | 25.7 |
| 蓬子菜Galium verum | 14.2 | 14.8 | 15.2 | 20.0 |
| 四川嵩草Kobresia setchwanensis | 10.3 | 11.9 | 9.6 | 10.7 |
| 马尔康柴胡 Bupleurum smithii | 13.0 | 14.8 | 18.5 | 16.2 |
| 冷地早熟禾Poa crymophila | 7.7 | 7.1 | 6.5 | 8.2 |
| 钝苞雪莲 Saussurea nigrescens | 10.4 | 9.8 | 11.2 | 12.2 |
| 高山韭Allium sikkimense | 14.5 | 16.4 | 14.1 | 13.7 |
| 菭草Koeleria cristata | 9.2 | 10.6 | 10.3 | 11.6 |
| 细叶亚菊Ajania tenuifolia | 13.3 | 10.2 | 14.5 | 16.7 |
| 草玉梅Anemone rivularis | 14.3 | 17.3 | 16.9 | 15.4 |
| 花苜蓿Medicago ruthenica | 22.9 | 25.0 | 25.9 | 27.0 |
| 椭圆叶花锚Halenia elliptica | 13.1 | 13.3 | 14.7 | 14.7 |
| 展毛翠雀花 Delphinium kamaonense | 10.7 | 11.6 | 13.4 | 13.7 |
| 丝叶毛茛 Ranunculus tanguticusvar.capillaceus | 13.5 | 15.5 | 11.3 | 13.7 |
| 乳白香青Anaphalis lactea | 9.9 | 10.1 | 10.5 | 10.9 |
| 甘肃棘豆Oxytropis kansuensis | 27.1 | 28.3 | 28.8 | 29.7 |
| 大籽蒿Artemisia sieversiana | 12.1 | 12.4 | 12.8 | 18.7 |
| 甘肃马先蒿 Pedicularis kansuensis | 19.5 | 21.8 | 23.3 | 27.3 |
| 高原毛茛Ranunculus tanguticus | 12.8 | 13.5 | 11.9 | 12.9 |
| 芒剪股颖 Agrostis trinii | 9.5 | 10.4 | 12.4 | 13.7 |
| 扁蕾Gentianopsis barbata | 10.5 | 14.4 | 15.2 | 16.3 |
2.4 施肥对单位植物氮含量的影响
在封育地的325个物种对间, 有308个物种对的单位植物氮含量存在显著差异, 施肥后, 只有107个物种对间有显著差异。在放牧地的325个物种对间, 有100个存在显著差异, 施肥后, 则有114个物种对间有显著差异(图3)。根据χ2检验发现, Pearson χ2= 374.5, p< 0.001, 证明结果不需要校正; 似然比卡方χ2= 433.2, p< 0.001, 表明施肥后放牧地和封育地物种间的单位植物氮含量存在显著差异。
图3
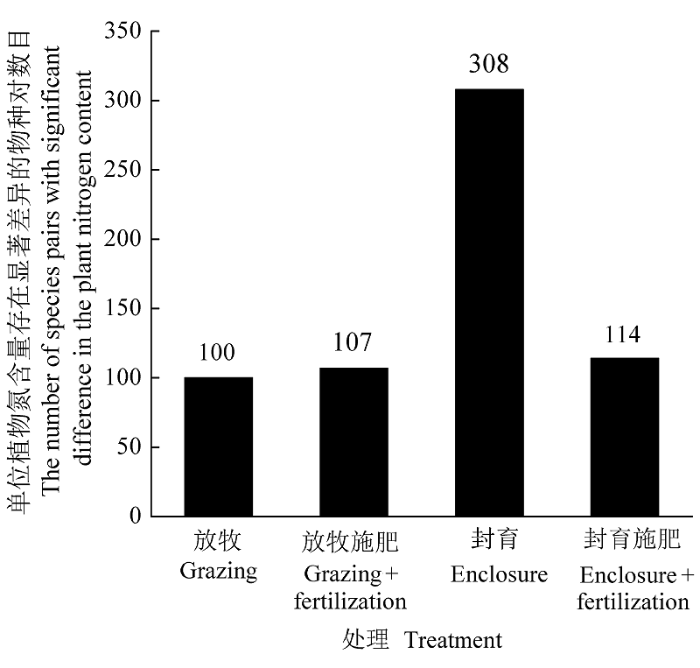
图3
施肥对封育地和放牧地单位植物氮含量存在显著差异的物种对数目的影响。
Fig. 3
Effects of fertilization on species pairs with a significant difference in plant nitrogen content both in enclosure and grazed plots.
3 讨论
3.1 在放牧地和封育地光竞争强度的变化对多样性的影响
封育地施肥后, 物种丰富度减小了, 个体大小不对称性增加了。个体较大的植物能获得更多的光资源, 快速地成为优势物种而妨碍其他植物的生长, 进而加速了小个体物种的消失(Weiner, 1985), 使得小个体物种在数量上更稀少。即使这些小个体的物种没有死亡(生理), 但由于遗传漂变的影响, 它们种群数量之小已经不能维持自身的存在, 它们在遗传上已经死亡(Weiner, 1985; Moles et al., 2009)。因此这种依赖个体大小的空间分布模式为解释物种多样性的下降机制提供了新的视点。现有的结果表明: 在光受限的环境里, 由于物种间高度的差异, 光竞争的不对称性对施肥后物种丰富度下降起到了重要作用。当然, 除高度之外, 其他物种特征的不同可能也会影响物种对施肥的响应, 例如, Gough等(2012)的研究表明, 虽然丛生禾草和根茎禾草都有较高的高度, 但是施肥后长寿命的丛生禾草多度下降了, 周转率快的根茎禾草多度增加了。这个研究表明了生活型及周转率对物种多样性下降的影响。因此, 未来的研究更应该在全球尺度上结合实验方法、植被类型和环境因子, 从物种特征这一方面探讨施肥后多样性下降的内在机制。
相对于封育地, 放牧地使得施肥对多样性的影响更加复杂。Hillebrand等(2007)的研究表明: 放牧和施肥对多样性的影响依赖于群落的均匀度、生态系统的生产力、植物生物量对施肥的响应, 以及食草动物对植物偏好等众多因素。而在本研究中, 尽管施肥降低了物种丰富度, 但是个体大小不等在各个施肥梯度上并没有显著差异。这可能与放牧地整体上的光环境有关。放牧后, 光成为一个中性特征, 植物在截取光的能力上并没有明显的差异, 光竞争减弱。特别是小个体植物, 尽管高度低, 但由于放牧移除了大个体的生物量, 使它们从光竞争中释放出来, 从而在这个过程中获利最多。另外, 放牧也可以通过减小茎密度减小光竞争(Aarssen et al., 2006), 进而会导致地上竞争减小。这可能是施肥后放牧地物种丰富度的变化比封育地缓慢的原因。需要注意的是, 尽管放牧地施肥后物种丰富度的变化与封育地施肥相比下降缓慢, 但是这在统计上还是显著的, 说明除了光外, 还存在着其他因素影响着物种丰富度的变化。
3.2 在放牧地和封育地地下竞争强度的变化对多样性的影响
封育地施肥后, 地下氮竞争的强度减弱了, 而光竞争增强了。这与Newman (1973)和Tilman (1988)的研究结果一致: 随着生产力的增加, 植物间从地下竞争逐渐转变为地上竞争。本研究还发现氮的利用效率在施肥后下降了, 这说明施肥后植物吸收的氮增加了, 植物通过增加氮吸收增加了光合效率, 进而提高了生物量, 增加了对光的竞争强度。在放牧地施肥后, 对氮资源不对称竞争的物种对数目有相对微弱的增加(地下不对称竞争的物种对数从100增加到114), 这可能部分解释了物种丰富度的变化。由于施肥增加了禾本科植物的比例, 禾本科植物通常有着较高密度的根系, 广泛分布的须根系能高效地吸收地下空间和资源, 这将减少其他植物幼苗生长的可利用空间和资源, 并进一步影响它们的成功建植。因此, 物种在根体积上的差别将在这种高密度根系的环境下对物种的共存起重要作用。另外, 由于氮的利用效率在物种间存在差异, 地下竞争是不对称的, 这与Weiher等(1998)与Newbery和Newman (1978)的实验结果不一致, 他们认为地下竞争是对称的。这可能有以下两个方面的原因: (1)他们的实验一般持续时间较短, 随着实验的进行, 资源越来越受到限制, 这种竞争有可能慢慢地转变为不对称性的。(2)在他们的实验中, 有限的个体密度和丰富的可利用性资源意味着地下竞争将会很弱, 不利于检测地下竞争强度在物种间的差异。本研究主要考虑施肥后竞争力在不同土地利用方式下的变化, 而忽视了密度及土壤种子库变化对多样性的影响。Luo等(2006)的研究表明, 密度的变化最多可引起多样性42.9%的下降; Stevens等(2004)的研究也表明, 施肥会引起物种补偿能力的降低而影响多样性。因此群落水平自疏假说以及物种补偿能力的变化对多样性下降起着不可忽视的作用。
3.3 展望
未来关于施肥后多样性下降的研究应加强以下三个方面: 1)在全球尺度上, 物种特征的变化对多样性下降的影响; 2)密度的变化(群落水平自疏假说)对多样性下降的影响; 3)长期施肥后种子限制以及幼苗定居限制对多样性下降的影响。
参考文献
Why are there so many small plants? Implications for species coexistence
Monotonic or unimodal diversity- productivity gradients: What does competition theory predict?
Biodiversity loss and its impact on humanity

The most unique feature of Earth is the existence of life, and the most extraordinary feature of life is its diversity. Approximately 9 million types of plants, animals, protists and fungi inhabit the Earth. So, too, do 7 billion people. Two decades ago, at the first Earth Summit, the vast majority of the world's nations declared that human actions were dismantling the Earth's ecosystems, eliminating genes, species and biological traits at an alarming rate. This observation led to the question of how such loss of biological diversity will alter the functioning of ecosystems and their ability to provide society with the goods and services needed to prosper.
Fertilization decreases plant biodiversity even when light is not limiting
P>Many researchers hypothesize that plant richness declines at high soil fertility (and high productivity) due to light limitation. We tested this hypothesis in an old-field by independently manipulating fertilization and light levels via shade cloth (decreased light), vegetation tie-backs (increased light) and vegetation clipping (increased light). Droughts occurred during two of the four years of the study, and we found that higher light levels were generally associated with decreased plant richness in drought years but increased plant richness in wet years. Most importantly, fertilization decreased richness whether light availability limited richness (wet years) or did not limit richness (drought years), and the effects of fertilization and light manipulation treatments were additive. These results suggest that effects of fertilization on plant richness are at least partly independent of light levels and that competition for resources other than light plays a substantial role in the decline of plant richness after fertilization.
Incorporating clonal growth form clarifies the role of plant height in response to nitrogen addition
Nutrient addition to grasslands consistently causes species richness declines and productivity increases. Competition, particularly for light, is often assumed to produce this result. Using a long-term dataset from North American herbaceous plant communities, we tested whether height and clonal growth form together predict responses to fertilization because neither trait alone predicted species loss in a previous analysis. Species with a tall-runner growth form commonly increased in relative abundance in response to added nitrogen, while short species and those with a tall-clumped clonal growth form often decreased. The ability to increase in size via vegetative spread across space, while simultaneously occupying the canopy, conferred competitive advantage, although typically only the abundance of a single species within each height-clonal growth form significantly responded to fertilization in each experiment. Classifying species on the basis of two traits (height and clonal growth form) increases our ability to predict species responses to fertilization compared to either trait alone in predominantly herbaceous plant communities.
Competition for light causes plant biodiversity loss after eutrophication
Human activities have increased the availability of nutrients in terrestrial and aquatic ecosystems. In grasslands, this eutrophication causes loss of plant species diversity, but the mechanism of this loss has been difficult to determine. Using experimental grassland plant communities, we found that addition of light to the grassland understory prevented the loss of biodiversity caused by eutrophication. There was no detectable role for competition for soil resources in diversity loss. Thus, competition for light is a major mechanism of plant diversity loss after eutrophication and explains the particular threat of eutrophication to plant diversity. Our conclusions have implications for grassland management and conservation policy and underscore the need to control nutrient enrichment if plant diversity is to be preserved.
Consumer versus resource control of producer diversity depends on ecosystem type and producer community structure
Testing mechanisms of N-enrichment induced species loss in a semiarid Inner Mongolia grassland: critical thresholds and implications for long-term ecosystem responses
Importance of assemblage- level thinning: a field experiment in an alpine meadow on the Tibet Plateau
What is the observed relationship between species richness and productivity?
Global patterns in plant height
Competition between grassland plants of different initial sizes
Four species of grassland plant, Plantago lanceolata, Holcus lanatus, Lolium perenne and Rumex acetosa, were grown as monocultures and mixtures in pots of nutrient poor soil in a glasshouse for 8 months. There were four plants per pot and these were arranged in two competition modes: either root and shoot interactions were permitted, or only roots allowed to interact by using above-ground partitions. Time of introduction of seedlings was varied to give a range of plant size ratios at the start of the experiment. The factorial design catered for all combinations of species, competition modes and planting times, replicated in four blocks. The shoots were clipped at a fixed height at each of five harvests. Rumex grew badly and was mostly omitted from analysis of the data.By (i) following the change in the relationship of clip dry weights against planting time with successive harvests, (ii) plotting the change in the logarithm of the ratio of cumulative clip dry weights with time and (iii) the use of de Wit logarithmic ratio plots it was demonstrated that each monoculture and mixture combination's ratios of plant weights converged towards stable equilibrium values. Three hypotheses are put forward to explain why in monocultures a smaller plant was at a competitive advantage relative to a larger neighbour and was not suppressed in its growth by the latter. In mixtures this plant size effect was superimposed to different extents on the relative aggressiveness of the species considered. It was concluded that in a nutrient poor soil, when competition for light was low, root interactions can promote the co-existence of neighbouring plant species.
Competition and diversity in herbaceous vegetation
The role of biomass allocation strategy in diversity loss due to fertilization
Why does fertilization reduce plant species diversity? Testing three competition-based hypotheses
Impact of nitrogen deposition on the species richness of grasslands
A transect of 68 acid grasslands across Great Britain, covering the lower range of ambient annual nitrogen deposition in the industrialized world (5 to 35 kg Nha-1 year-1), indicates that long-term, chronic nitrogen deposition has significantly reduced plant species richness. Species richness declines as a linear function of the rate of inorganic nitrogen deposition, with a reduction of one species per 4-m2 quadrat for every 2.5 kg Nha-1 year-1 of chronic nitrogen deposition. Species adapted to infertile conditions are systematically reduced at high nitrogen deposition. At the mean chronic nitrogen deposition rate of central Europe (17 kg Nha-1 year-1), there is a 23% species reduction compared with grasslands receiving the lowest levels of nitrogen deposition.
The significance of assemblage-level thinning for species richness
Plant density determines species richness along an experimental fertility gradient
Functional and abundance-based mechanisms explain diversity loss due to N fertilization
Diversity-productivity relationships: initial effects, long-term patterns, and underlying mechanisms
The relationship between productivity and species richness
Community assembly rules, morphological dispersion, and the coexistence of plant species
Size hierarchies in experimental populations of annual plants
Stochastic and deterministic processes together determine alpine meadow plant community composition on the Tibetan Plateau
To predict the consequences of environmental change on the structure and composition of communities, it is necessary to also understand the regional drivers underlying the structuring of these communities. Here, we have taken a hypothesis-based approach to test the relative importance of niche versus neutral processes using niche overlap, species traits and population asynchrony in two crossed treatments of fertilization and grazing in an alpine meadow community. Our results suggested that the observed species biomass overlap was not significantly different between treatments of grazing, grazing x fertilization and grazer exclusion. In contrast, the species biomass overlap was higher than expected in fertilization treatments when grazers were excluded. On the one hand, we found no relationship between species traits and relative abundance in grazing, grazing x fertilization and grazer-exclusion treatments; on the other hand, mechanistic trait-based theory could be used to predict species relative abundance patterns in fertilization treatments when grazers were excluded. From grazing to fertilization, when grazers were excluded, there was a slight increase in species synchrony, which indicated that the complementary dynamic of species gradually changed from complete independence into synchronously fluctuating with increasing fertilization. Based on the above results, we concluded that stochastic and deterministic processes formed ends of a continuum from grazing to fertilization when grazers were excluded in an alpine meadow plant community, and the importance of niche differences between species in structuring grassland communities increased with increasing fertilization and decreased with grazing.
The effects of long-term fertilization on the temporal stability of alpine meadow communities
Recent theoretical and experimental work suggests that species diversity enhances the temporal stability of communities. However, empirical support largely comes from experimental communities. The relationship between diversity and stability in natural communities, and the ones facing environmental changes in particular, has received less attention. We created a gradient of fertility in a natural alpine meadow community to test the effects of diversity and fertilization on the temporal variability of community cover and cover of component species and to determine the importance of asynchrony, portfolio effects, cover and dominance for diversity-stability relationships. Although fertilization strongly reduced species richness, the temporal stability in community cover increased with fertilization. Most species showed a decline of temporal stability in mean population cover with fertilization, but two grass species, which dominated fertilized communities after 10 years, showed an increase of stability. Detailed analysis revealed that the increased dominance of these two highly stable grass species was associated with increased community stability at high levels of fertilization. In contrast, we found little support for other mechanisms that have been proposed to contribute to community stability, such as changes in asynchrony and portfolio effects. We conclude that the presence of highly productive species that have stabilizing properties dominate fertilized assemblages and enhance ecosystem stability.
The effect of environmental and phylogenetic drivers on community assembly in an alpine meadow community
To predict the consequences of environmental change on plant communities at local scales, one needs to understand the regional drivers structuring these communities. Here, we used a formal analytical framework incorporating functional traits and evolutionary histories to understand the importance of environmental filtering and species interactions in the assembly of alpine plant communities. The study was conducted in the Tibetan Plateau using field plots experiencing changes in land use (fertilization and grazing). We observed evidence for both trait-based convergence (associated with plant height and tissue nitrogen) and divergence (associated with specific leaf area) within alpine plant communities, suggesting that environmental filtering and limiting similarity are acting simultaneously during assembly processes. Although we did not observe evidence of phylogenetic niche conservatism in relation to intensified land use, we did observe support for the phylogenetic structure of plant communities influencing community-weighted mean trait values, suggesting that evolutionary constraints represent a significant driver of community assembly in this system. Therefore, evolutionary and ecological processes may have independent effects on alpine plant communities facing land use intensification.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}