


种群空间格局分析是研究种群特征、种群间相互作用以及种群与环境关系的重要手段, 一直是生态学的热点之一(Wiegand & Moloney, 2004)。探索森林立木的空间分布格局可以为未充分认识的生态问题提供更好的理论解释(Franklin et al., 2002; Salas et al., 2006, Flügge et al., 2012), 对理解森林生态系统的种群动态变化至关重要(Veblen et al., 1979)。许多物种共存和群落结构的现象和理论已经被提出, 例如, 热带雨林同种相邻成体的距离较远(Janzen, 1970), 成年个体分布相对均匀(Hubbell, 1979), 具有丰富的生物物种和相对低的立木密度(Condit et al., 2000)等, 这些都涉及空间格局分析(Li et al., 2009)。物种的空间格局分析在热带森林的研究中一直是一个相当重要的研究方向(Wiegand & Moloney, 2004)。
自然界中大多数物种空间分布的聚集程度和尺度体现着物种如何利用资源, 如何被捕食或寄生, 以及物种的繁殖特性。影响物种分布格局的可能机理主要有扩散限制、生境异质性、Janzen-Connell假说和邻体竞争。(1)扩散限制: 扩散限制可以在空间上使物种分离而减弱种间竞争程度, 热带雨林不同物种的空间格局与扩散限制有密切关系, 扩散距离和植株密度是影响空间格局的主要因素(Seidler & Plotkin, 2006; Lin et al., 2011)。(2)生境异质性: 环境的筛选作用使物种聚集在适宜的环境中, 生态位分化促使物种共存(Lai et al., 2009; Baldeck et al., 2013)。(3) Janzen-Connell假说: 由于捕食者和病原体对种子和幼苗的捕食和寄生, 幼苗、幼树的分布远离母树(Connell, 1971; Liu et al., 2012), 在母树周围的一定范围内为其他物种提供了定居空间, 此假说是物种共存的主要机制之一。(4)邻体竞争: 幼苗定居、生长、存活的各个阶段, 都存在由于邻体竞争而发生的密度制约的死亡作用, 这减小了种群的聚集程度, 导致较高的生物多样性(Wills et al., 1997; He & Duncan, 2000; Johnson et al., 2012)。众多的解释机制说明物种空间格局形成的复杂性和探索其不同形成原因的必要性。
尖峰岭是我国目前为数不多的保存较完好的具有热带雨林性质的森林区域(方精云等, 2004)。该地森林类型丰富, 从低海拔到高海拔依次形成滨海有刺灌丛、稀树草原、热带半落叶季雨林、热带常绿季雨林、热带沟谷雨林、热带山地雨林以及山顶苔藓矮林等植被类型, 热带山地雨林则为该地区发育最为完善、结构最为复杂的类型(李意德等, 2002)。白颜树(Gironniera subaequalis)为榆科白颜树属的常绿乔木, 属于尖峰岭热带原始林中第二层优势乔木。
本研究采用成对相关函数(pair correlation function, PCF)分析白颜树种群各个径级的空间分布格局特征, 采用双变量成对相关函数和标签关联函数(mark connection functions, MCF)分析不同生活史阶段的空间关联性; 采用Berman检验方法检验地形因子对个体分布的影响, 试图揭示白颜树种群发育过程中的空间动态, 并讨论上述几种机制在格局形成过程中所起的作用。
1 材料和方法
1.1 自然概况
尖峰岭地处海南岛西南部, 北与东方县接壤, 东、南均与乐东县相邻, 研究区域位于18° N, 108°- 109° E之间。属低纬度热带岛屿季风气候, 年降水量2449.0 mm (周璋等, 2009), 年平均气温24.5 ℃, 最冷月平均气温19.4 ℃, 最热月平均气温27.3 ℃, 干湿两季明显。尖峰岭核心区还保留有未经砍伐、保持原生状态的热带山地雨林。地带性土壤为燥红土、褐色砖红壤、砖红壤性黄壤和山地黄壤等类型(李意德等, 2002)。
1.2 研究方法
1.2.1 样地调查
海南尖峰岭60 hm2大样地建设参照1980年美国史密森热带研究所热带森林研究中心(Center for Tropical Forest Science, CTFS)在巴拿马Barro Colorado Island建立50 hm2样地制定的技术规范进行。整个样地呈长方形, 采用全站仪(南方测绘NTS-310B/R, 广州)划分为1500个20 m × 20 m的样方。400 m2的大样方用水泥角桩划分, 每个大样方又分为16个5 m × 5 m的小样方, 在每个5 m × 5 m的单元内, 对植物群落进行全面调查。记录每个样方内胸径(1.3 m处)≥1 cm乔木树种的物种学名、胸径及生长情况, 并测量个体空间位置坐标, 坐标值直接用距离(m)表示。同时利用测量得到的地形数据计算每个20 m × 20 m样方的海拔、坡度、坡向、坡位参数。
1.2.2 点格局分析方法
本研究采用现代点格局函数分析各径级的空间分布特征和不同生活史阶段之间的空间关联。
Ripley’s K函数公式如下(Ripley, 1976):
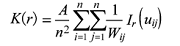
uij表示个体i和个体j之间的距离, 当uij ≥ r时, Ir= 0, 当uij ≤ r时, Ir= 1; Wij是以i为圆心, uij为半径的圆位于样地内的周长与圆周长之比, 为一个修正值; A为样地面积, n为样地中个体总数; r表示空间尺度(张金屯, 1998)。
双变量的K函数:
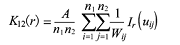
式中, n1与n2表示不同生活史阶段的个体数量, 其余参数与式(1)中相同。
成对相关函数(下文简称g函数)消除了K函数在大尺度上的累积效应, 其表达式为:
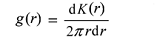
g函数也有相应的单变量和双变量形式, 对于完全随机分布, g(r) = 1。
标签关联函数(下文简称P函数)是一组条件概率表达式, 两个阶段间的P函数包括3个表达式: P11, P12, P22, 表示取距离为r的任意两个个体, 属性分别为1,1 (或1,2; 2,2)的可能性, 假设1个体和2个体出现的概率为p1和p2, 那么理论值P11 = p12; P12 = 2p1p2; P22 = p22。因为分母相当于随机标签零假设的g函数, 所以P12函数(式4, 其中λ1与λ2表示相应的密度, g(r) = [λ1λ1g11(r) + λ1λ2g12(r) +λ2λ1g21(r) + λ2λ2g22(r)]/(λ1 + λ2)2)排除了可能存在的大尺度环境异质性对两径级个体分布的影响, 反映了两种个体间绝对的相互作用(Getzin et al., 2008; Illian et al., 2008)。
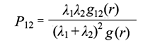
选择具有明确生态学意义、能准确描述数据偏离理论程度的零假设模型(null model)对空间点格局分析非常关键。不同的研究目的和问题假设要选择不同的零模型(王鑫厅等, 2012)。本文用到的零模型有: (1)完全空间随机模型(complete spatial randomness, CSR): 假设物种的空间分布不受任何生物或非生物过程影响, 在研究区域内各点出现的概率相同; (2)异质泊松模型(heterogeneous Poisson, HP): 依据密度函数λ(s) (s表示样地内任意一点的位置, 可用坐标(x, y)表示)决定个体的分布, 可以排除大尺度环境异质性的影响, 本研究采用高斯核函数(Diggle, 2003)进行密度估计, 选取标准差sigma = 15 m消除大尺度上的环境异质性对空间格局的影响; (3)先决条件模型(antecedent condition, AC): 在生成双变量g函数的包迹线时, 假设2个体的位置不变(假设2先存在, 2表示较大龄级个体), 1个体的分布在本研究中采用sigma = 15 m的异质泊松模型模拟; (4)随机标签模型(random labeling, RL): 个体的位置不变, 属性随机赋予每个个体。对于单变量g函数采用完全随机模型和异质泊松模型; 双变量g函数采用完全随机模型和先决条件模型; 而P函数结果表示条件概率, 相当于采用随机标签模型。通过99次Monte Carlo随机模拟, 分别利用模拟的最大值和最小值生成上下两条包迹线, 计算99%的置信区间。
1.2.3 地形因子检验方法
用Berman检验方法(Berman, 1986; Zhu et al., 2013)检验地形因子对各生活史阶段个体分布的影响。计算所有个体所在位置xi的环境变量值v(xi)的平均值Sobs, 以20 m × 20 m样方为单位每次沿x轴或y轴移动一个单位生成零假设模型, 计算每个零模型相应的Ssim, 显著度由Sobs在Ssim中的排序位置决定。
本文所有分析计算使用R 3.0.1 (http://www. rproject.org)软件完成。
2 结果
尖峰岭热带山地雨林的树种重要值相对较小, 没有占绝对优势的树种, 其中白颜树的重要值在整个样地中排名第二, 仅次于棕榈科的大叶蒲葵(Livistona saribus)。白颜树的相对胸高断面积、相对密度和相对频度都较大, 其中相对胸高断面积也仅次于大叶蒲葵(表1)。
表1 白颜树在群落中的数量指标
Table 1
| 重要值 Importance value | 相对胸高断面积 Relative cross-section area at the breast height | 相对密度 Relative density | 相对频度 Relative frequency | |
|---|---|---|---|---|
| 数值 Value | 3.01 | 5.50 | 2.28 | 1.24 |
| 在样地内的排名 Relative position of all species | 2 | 2 | 9 | 4 |
60 hm2样地中的白颜树共有10022株, 按胸径等距分为6个径级, 再把I、II划为幼树, III、IV划为中龄树, V、VI划为成年树(表2; 图1)。白颜树的小径级个体数量较多而大径级个体数量较少, 径级分布呈现倒“J”型, 群落处于增长或稳定状态。
表2 生活史的划分
Table 2
| 幼树 Young trees (44%) | 中龄树 Middle-aged trees (34.8%) | 成年树 Adult trees (21.2%) | ||||||
|---|---|---|---|---|---|---|---|---|
| 径级I Class I | 径级II Class II | 径级III Class III | 径级IV Class IV | 径级V Class V | 径级VI Class VI | |||
| 胸径 DBH | 1 cm ≤ DBH < 5 cm | 5 cm ≤ DBH < 10 cm | 10 cm ≤ DBH < 15 cm | 15 cm ≤ DBH < 20 cm | 20 cm ≤ DBH < 25 cm | DBH ≥ 25 cm | ||
| 株数 Number of stems | 2 503 | 1 948 | 1 830 | 1 655 | 1 194 | 932 | ||
DBH, diameter at the breast height.
图1
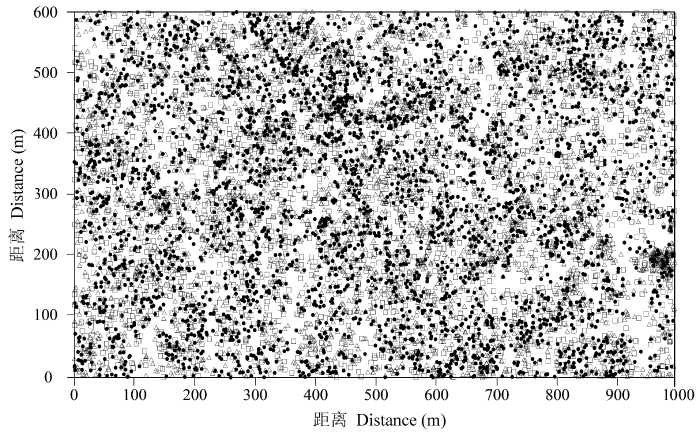
图1
白颜树所有个体的空间分布。
三角形代表幼树; 黑色圆点代表中龄树; 正方形代表成年树。
Fig. 1
Spatial distribution of all stems of Gironniera subaequalis.
Young trees are marked by the triangles; middle-aged trees are marked by the black dots; and adult trees are marked by the squares.
2.1 各径级空间点格局分析
基于完全随机分布(CSR)和异质泊松分布(HR) 零模型的分析都表明, 随着径级增加, 白颜树的空间聚集程度和尺度逐渐减小, 分布特征由聚集型过渡为随机型、均匀型(图2): CSR零模型分析显示I、II、III径级的聚集尺度分别是0-43 m、0-43 m、0-31 m; IV径级在0-6 m内随机分布, 在6-20 m之间有微弱的聚集分布特征; V径级除了在0-2 m表现出很微弱的均匀分布外, 所有尺度上都是随机分布; VI径级在0-5 m内表现为均匀分布, 而在较大的尺度上有微弱的聚集分布特征。HR零模型分析排除了15 m以上的大尺度环境异质性的影响, 结果显示I、II、III径级的聚集尺度分别是0-8 m, 0-7 m, 0-5 m; IV径级在0-4 m内均匀分布, 其他范围内随机分布, V径级在0-3 m内均匀分布, VI径级在0-10 m内均匀分布(图2)。
图2
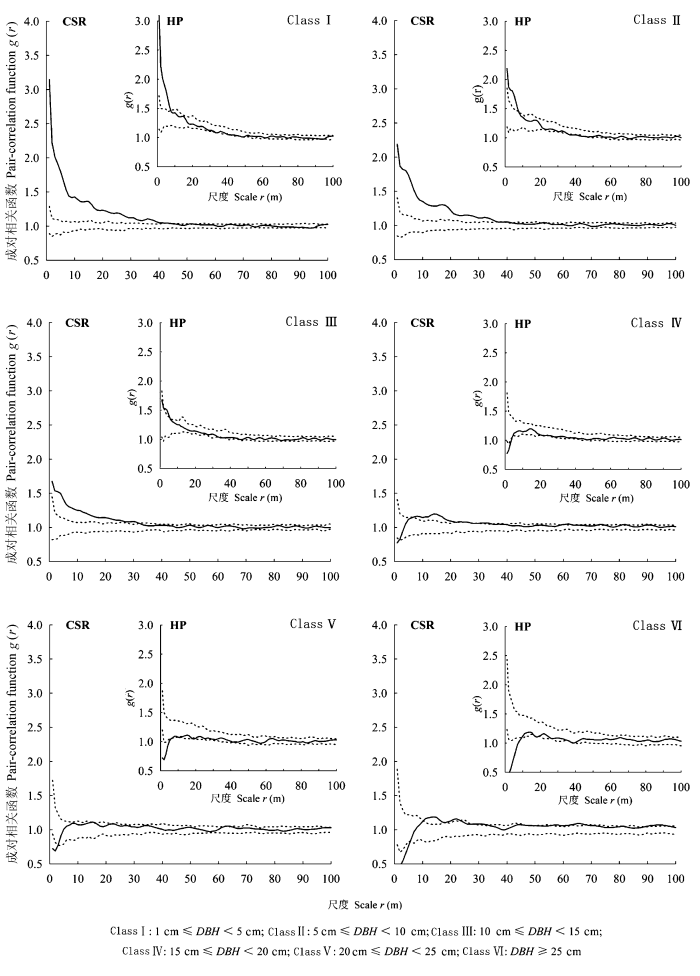
图2
白颜树各径级的成对相关函数。
实线表示成对相关函数, 虚线表示99%包迹线。零模型为完全空间随机模型(CSR)和sigma = 15 m的异质泊松模型(HP)。
Fig. 2
Univariate pair-correlation functions of different age classes of Gironniera subaequalis.
Solid lines are pair-correlation functions, and dotted lines are 99% confidence envelopes. Null models: complete spatial randomness (CSR) and Heterogeneous Poisson (HP) with sigma = 15 m.
2.2 不同生活史阶段空间相关关系
基于完全空间随机零模型的双变量g函数分析结果表明(图3): 幼树和中龄树两个阶段个体在0-48 m的尺度上呈现空间正关联, 随后还有几个小区间函数值位于包迹线以上, 呈现出微弱的空间正关联, 这可能是随机波动引起的; 幼树和成年树在0-5 m内负关联, 大于5 m后无关联; 中龄树与成年树在0-4 m内负关联, 大于4 m后表现出微弱的正关联关系。排除环境异质性的分析结果表明(图3): 幼树和中龄树两个阶段个体在0-10 m; 14-15 m的尺度上呈现空间正关联, 其他尺度上无关联; 幼树和成年树在0-5 m内负关联, 大于5 m后无关联; 中龄树与成年树在0-4 m内负关联, 大于4 m后几乎无空间关联。虽然成年树与其余两个阶段之间都呈现负关联, 但成年树与中龄树之间负关联的尺度要比与幼树之间的小, 而函数曲线与理论值的偏离程度大。
图3
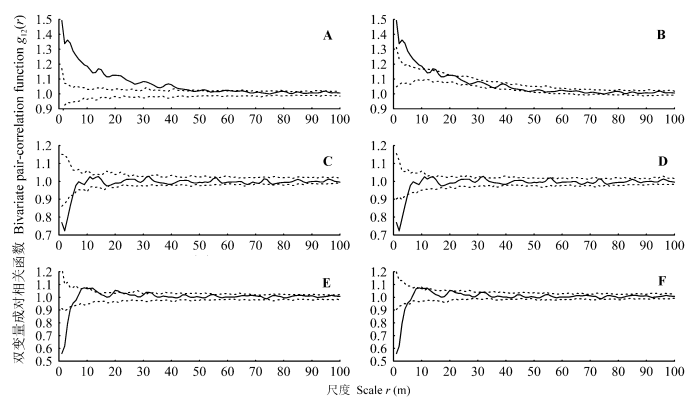
图3
白颜树各个生活史阶段之间的双变量成对相关函数。实线表示双变量成对相关函数; 虚线表示99%包迹线。
A, 包迹线采用完全空间随机零模型的幼树与中龄树的关系。B, 包迹线采用先决条件零模型的幼树与中龄树的关系。C, 包迹线采用完全空间随机零模型的幼树与成年树的关系。D, 包迹线采用先决条件零模型的幼树与成年树的关系。E, 包迹线采用完全空间随机零模型的中龄树与成年树的关系。F, 包迹线采用先决条件零模型的中龄树与成年树的关系。
Fig. 3
Bivariate pair-correlation functions between pairs of life-history stages of Gironniera subaequalis. Solid lines are bivariate pair-correlation functions and dotted lines are 99% confidence envelopes.
A, Relationship between young trees and middle-aged trees with envelopes calculated by complete spatial randomness null model. B, Relationship between young trees and middle-aged trees with envelops calculated by antecedent condition null model. C, Relationship between young trees and adult trees with envelopes calculated by complete spatial randomness null model. D, Relationship between young trees and adult trees with envelops calculated by antecedent condition null model. E, Relationship between middle-aged trees and adult trees with envelopes calculated by complete spatial randomness null model. F, Relationship between middle-aged trees and adult trees with envelops calculated by antecedent condition null model.
P函数排除了大尺度上环境异质性的影响, 分析结果与g函数有所差异(图4): 中龄树和幼树在1-29 m内相分离, 其中幼树在28 m内聚集; 中龄树个体之间在6 m内被幼树分离。幼树和成年树在42 m内相分离, 其中幼树在0-28 m内聚集; 成年树在9 m内分离。中龄树和成年树在19 m内相分离, 其中中龄树在9 m的范围内聚集; 成年树在6 m内分离。而在相对大的尺度上各函数值都在包迹线之内, 表示两个生活史阶段的个体随机混和。
图4
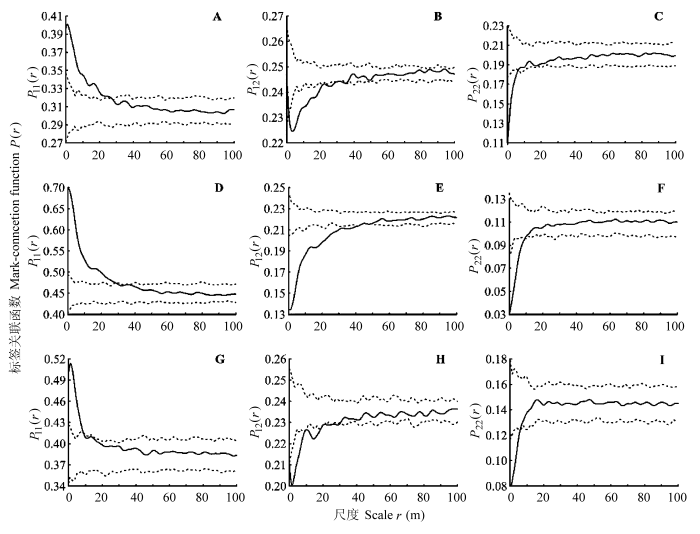
图4
标签关联函数P(r)。
A, 从所有幼树和中龄树中任选两个距离为r的个体, 两者均为幼树的概率。B, 从所有幼树和中龄树中任选两个距离为r的个体, 其中一个为幼树, 另一个为中龄树的概率。C, 从所有幼树和中龄树中任选两个距离为r的个体, 两者均为中龄树的概率。D, 从所有幼树和成年树中任选两个距离为r的个体, 两者均为幼树的概率。E, 从所有幼树和成年树中任选两个距离为r的个体, 其中一个为幼树, 另一个为成年树的概率。F, 从所有幼树和成年树中任选两个距离为r的个体, 两者均为成年树的概率。G, 从所有中龄树和成年树中任选两个距离为r的个体, 两者均为中龄树的概率。H, 从所有中龄树和成年树中任选两个距离为r的个体, 其中一个为中龄树, 另一个为成年树的概率。I, 从所有中龄树和成年树中任选两个距离为r的个体, 两者均为成年树的概率。实线表示标签关联函数; 虚线表示99%包迹线。
Fig. 4
The mark-connection functions P(r).
A, Probabilities by randomly choosing two individuals (separated by distance r) from all young trees and middle-aged trees, when both are young trees. B, Probabilities by randomly choosing two individuals (separated by distance r) from all young trees and middle-aged trees, when one is a young tree and the other is a middle-aged tree. C, Probabilities by randomly choosing two individuals (separated by distance r) from all young trees and middle-aged trees, when both are middle-aged trees. D, Probabilities by randomly choosing two individuals (separated by distance r) from all young trees and adult trees, when both are young trees. E, Probabilities by randomly choosing two individuals (separated by distance r) from all young trees and adult trees, when one is a young tree and the other is an adult tree. F, Probabilities by randomly choosing two individuals (separated by distance r) from all young trees and adult trees, when both are adult trees. G, Probabilities by randomly choosing two individuals (separated by distance r) from all middle-aged trees and adult trees, when both are middle-aged trees. H, Probabilities by randomly choosing two individuals (separated by distance r) from all middle-aged trees and adult trees, when one is a young tree and the other is an adult tree. I, Probabilities by randomly choosing two individuals (separated by distance r) from all middle-aged trees and adult trees, when both are adult trees. Solid lines are mark-connection functions, and dotted lines are 99% confidence envelopes.
2.3 地形因子对各个阶段个体分布的影响
坡度对每个生活史阶段的个体分布都有显著影响: 海拔仅对幼树影响显著, 对中龄树和成年树无显著影响; 凹凸度对幼树和中龄树影响显著, 对成年树无显著影响(表3)。
表3 地形因子对白颜树各生活史阶段个体分布的影响
Table 3
| 幼树 Young tree | 中龄树 Middle-aged tree | 成年树 Adult tree | |
|---|---|---|---|
| 坡度 Slope | + | + | + |
| 海拔 Elevation | + | - | - |
| 凹凸度 Convex | + | + | - |
+, significant positive correlation; -, uncorrelated; p ≤ 0.025.
+, 显著正相关; -, 无相关性; p ≤ 0.025。
3 讨论
本研究发现, 白颜树的I、II、III、IV径级聚集分布, 且聚集程度和尺度随径级增大而减小, 属于成年树的VI、VII径级在小尺度上呈现均匀分布(图2)。以往的众多研究发现自然界中大多数物种在一定尺度上都会表现出聚集分布特征, 这样的格局有利于发挥群体效应, 形成适于自身生长的环境, 增加种间竞争力, 抵御外来种入侵, 维持种群的正常发展(韩路等, 2007)。Condit等(2000)对6个不同地点的热带森林研究发现, 有大于97.8%的物种聚集分布, 在中国的鼎湖山亚热带森林中有96.1%的物种聚集分布(Li et al., 2009), 在海南岛霸王岭热带低地雨林中, 呈现聚集分布的树种约70%, 随着尺度的增大, 聚集强度逐渐减小(黄运峰等, 2012), 西双版纳20 hm2热带森林样地内的4个上层优势树种的幼树和中龄树以聚集分布为主, 而成年树则以随机分布为主(兰国玉等, 2008), 本研究与上述研究结果一致。物种本身的生物学特性、群落中其他的生物因素和环境因素都影响着种群的空间格局。
3.1 扩散限制
许多研究都说明扩散对格局形成起到至关重要的作用(Pinto & MacDougall, 2010; Shen et al., 2013)。对热带森林561个物种的研究表明: 空间聚集的强度、尺度与种子扩散方式有关(Seidler & Plotkin, 2006)。对鼎湖山124个个体数大于10的物种的研究发现, 只依靠风或动物扩散的物种比同时依靠两种方式扩散的物种聚集程度大(Li et al., 2009)。扩散限制是导致热带地区树种聚集分布的重要机制之一(Wiegand et al., 2007)。扩散对格局形成的作用直观的表现是在一定的尺度上幼树与成年树正关联。本研究中双变量g函数表明成年树和幼树在小尺度上负关联(图3C, 3D), 标签关联函数表明幼树本身聚集分布, 但与母树分离(图4D, 4E), 因此母树作为种源使幼苗、幼树在其周围聚集分布的作用在本研究中没有明确表现出来。以异质泊松过程为零模型的g函数消除了大尺度上的环境异质性后, I、II、III径级仍然聚集分布, 这很可能是扩散的结果(图2); 用先决条件零模型排除环境异质性后, 成对相关函数显示幼树和中龄树在10 m内空间正关联(图3B), 而幼树和中龄树的标签关联函数中, P12(r)函数值小于理论值说明幼树和中龄树个体间是相分离的(图4B), 所以小尺度上幼树和中龄树的空间正关联可能是由于扩散导致的斑块重合。森林植物群落形成过程中的一个重要特征是扩散限制先设定好一个模板或一个格局(Levine & Murrell, 2003), 之后由于个体受生境的影响而改变原有的格局分布(黄运峰等, 2012)。由于样地中白颜树成年树密度大, 而且分布较均匀, 其种子较小而数量多, 可能会导致不同母树的种子空间上重叠在一起; 其种子颜色鲜艳, 能依靠鸟类传播, 扩散距离较远, 扩散过程中, 一部分种子会被鸟类取食, 这些在很大程度上使胸径1 cm以上的个体分布异于理论的扩散格局。此外, 白颜树雌雄异株, 本研究缺少成年树性别的信息, 故无法明确体现出种子的扩散限制所起的作用。
3.2 环境异质性
环境影响着植物的整个生活史, 从而影响种群的空间格局。在小尺度上, 环境异质性可能仅对少量物种的分布有显著影响(Harms et al., 2001), 而在大尺度上环境异质性被认为是产生植被空间格局的主要原因(Clark et al., 1998; Takyu et al., 2002), 在大于10 m的尺度上, 如果呈现聚集分布, 就可以解释为是受到生境异质性的影响(Getzin et al., 2008), CSR零模型(存在环境异质性)分析显示I、II、III三个阶段聚集分布的最大尺度均在30 m以上, IV径级虽然在较小尺度内变为随机分布, 但在6-20 m又表现为聚集分布, 可以推测环境异质性在其中起了作用(图2); 以CSR为零模型的幼树和中龄树的双变量g函数也显示出大尺度上的正关联(图3), 而幼树和中龄树的标签关联函数排除了大尺度上环境的影响, 说明幼树和中龄树个体间是相分离的(图4B), 其中P12(r)在包迹线以下的最大尺度可能是中龄树和幼树斑块中心相距的平均距离, 由于幼树的聚集程度大, 镶嵌在中龄树的斑块中, 使中龄树个体在6 m内被分离, 这充分说明幼树和中龄树的空间正关联不是个体间的直接作用导致的, 而是由于环境的影响各自以斑块的形式分布在适宜的生境中。
有些物种的大树会表现出比幼树更强的聚集分布, 这可能是由于环境的斑块性所致, 这些物种的幼年个体分布广泛, 随着时间的推移, 处于劣势环境中的个体死亡, 导致大多数成年树生活在种群更适应的环境中(Condit et al., 2000), 但白颜树的聚集程度随着径级增大而减小, 所以环境对胸径1 cm以上的白颜树个体的影响, 不足以造成I-VI阶段的格局特征变化。结合坡度、海拔、凹凸度3个地形因子都对幼树影响显著, 坡度和凹凸度对中龄树影响显著, 仅坡度对成年树影响显著(表3), 说明白颜树的小径级个体比大径级个体对环境更为敏感, 我们推测环境的选择性在种子扩散、萌发过程中或是幼苗胸径还未长到1 cm之前更为明显; Baldeck等(2013)对1 cm以上的立木研究发现同径级的亚群落结构和组成相似, 可以推断对于大多数物种, 环境的筛选作用发生在更早的时期, 与本研究结果一致。
3.3 密度依赖死亡
在大径级的个体中, 个体死亡概率依赖于其本身生长发育情况和竞争环境(Batista & Maguire, 1998)。如果个体是随机死亡的, 死亡后的活立木空间格局应该保持不变, 而密度依赖死亡会减弱聚集程度(Condit et al., 2000), 甚至使分布格局变为随机或均匀分布。密度依赖死亡在热带森林中普遍存在, 是物种共存的主要机制之一(Bagchi et al., 2011)。密度依赖死亡作用是由个体间竞争引起的(Murrell, 2009), 由于竞争死亡, 个体的空间分布越来越均匀。本研究中I、II、III、IV四个径级聚集程度不断减弱, V、VI在小距离上是均匀分布的, 且VI比V均匀分布的强度和尺度都大(图2), 虽然没有死亡前后的对比数据, 但是可以推断密度依赖死亡作用的存在。在较小的尺度上白颜树的成年树不仅与幼树、中龄树空间负关联, 成年树本身也表现出均匀分布, 可能是由于成年树对空间、资源等占有较多, 使同种个体在其周围一定的范围内很难生存, 说明白颜树个体发育中经历了强烈的种内竞争, 成年树阶段最终呈现出均匀分布格局。关于密度依赖死亡对格局形成的作用, 本研究只分析了种内竞争的影响, 而来自其他物种尤其是来自优势种的种间竞争对空间格局的形成也非常关键(Hu et al., 2012), 热带森林具有极高的物种多样性, 相邻植株往往不是同种个体, 因此异种个体的影响也是空间格局形成的一种主要机制。
参考文献
Spatial patterns reveal negative density dependence and habitat associations in tropical trees

Understanding how plant species coexist in tropical rainforests is one of the biggest challenges in community ecology. One prominent hypothesis suggests that rare species are at an advantage because trees have lower survival in areas of high conspecific density due to increased attack by natural enemies, a process known as negative density dependence (NDD). A consensus is emerging that NDD is important for plant-species coexistence in tropical forests. Most evidence comes from short-term studies, but testing the prediction that NDD decreases the spatial aggregation of tree populations provides a long-term perspective. While spatial distributions have provided only weak evidence for NDD so far, the opposing effects of environmental heterogeneity might have confounded previous analyses. Here we use a novel statistical technique to control for environmental heterogeneity while testing whether spatial aggregation decreases with tree size in four tropical forests. We provide evidence for NDD in 22% of the 139 tree species analyzed and show that environmental heterogeneity can obscure the spatial signal of NDD. Environmental heterogeneity contributed to aggregation in 84% of species. We conclude that both biotic interactions and environmental heterogeneity play crucial roles in shaping tree dynamics in tropical forests.
Habitat filtering across tree life stages in tropical forest communities
Modeling the spatial structure of topical forests
Testing for spatial association between a point process and another stochastic process
Edaphic variation and the mesoscale distribution of tree species in a neotropical rain forest
Spatial patterns in the distribution of tropical tree species
Fully mapped tree census plots of large area, 25 to 52 hectares, have now been completed at six different sites in tropical forests, including dry deciduous to wet evergreen forest on two continents. One of the main goals of these plots has been to evaluate spatial patterns in tropical tree populations. Here the degree of aggregation in the distribution of 1768 tree species is examined based on the average density of conspecific trees in circular neighborhoods around each tree. When all individuals larger than 1 centimeter in stem diameter were included, nearly every species was more aggregated than a random distribution. Considering only larger trees (>/= 10 centimeters in diameter), the pattern persisted, with most species being more aggregated than random. Rare species were more aggregated than common species. All six forests were very similar in all the particulars of these results.
On the role of natural enemies in preventing competitive exclusion in some marine animals and in rain forest trees
Statistical Analysis of Spatial Point Patterns
Community structures and species richness in the montane rain forest of Jianfengling, Hainan Island, China
海南岛尖峰岭山地雨林的群落结构、物种多样性以及在世界雨林中的地位
The memory of spatial patterns: changes in local abundance and aggregation in a tropical forest
The current spatial pattern of a population is the result of previous individual birth, death, and dispersal events. We present a simple model followed by a comparative analysis for a species-rich plant community to show how the current spatial aggregation of a population may hold information about recent population dynamics. Previous research has shown how locally restricted seed dispersal often leads to stronger aggregation in less abundant populations than it does in more abundant populations. In contrast, little is known about how changes in the local abundance of a species may affect the spatial distribution of individuals. If the level of aggregation within a species depends to some extent on the abundance of the species, then changes in abundance should lead to subsequent changes in aggregation. However, an overall change of spatial pattern relies on many individual birth and death events, and a surplus of deaths or births may have short-term effects on aggregation that are opposite to the long-term change predicted by the change in abundance. The change in aggregation may therefore lag behind the change in abundance, and consequently, the current aggregation may hold information about recent population dynamics. Using an individual-based simulation model with local dispersal and density-dependent competition, we show that, on average, recently growing populations should be more aggregated than shrinking populations of the same current local abundance. We tested this hypothesis using spatial data on individuals from a long-term tropical rain forest plot, and find support for this relationship in canopy trees, but not in understory and shrub species. On this basis we argue that current spatial aggregation is an important characteristic that contains information on recent changes in local abundance, and may be applied to taxonomic groups where dispersal is limited and within-species aggregation is observed.
Disturbances and structural development of natural forest ecosystems with silvicultural implications, using Douglas-fir forests as an example
Heterogeneity influences spatial patterns and demographics in forest stands
Spatial distribution patterns and dynamics of major population in Populus euphratica forest in upper reaches of Tarim River
塔里木河上游天然胡杨林种群空间分布格局与动态研究
Habitat associations of trees and shrubs in a 50-ha neotropical forest plot
Density-dependent effects on tree survival in an old-growth Douglas fir forest
Dominant species and dispersal limitation regulate tree species distributions in a 20-ha plot in Xishuangbanna, southwest China
Spatial pattern of trees in tropical lowland rain forest in Bawangling of Hainan Island, China
海南岛霸王岭热带低地雨林树木的空间格局. 植物生态学报,
Tree dispersion, abundance, and diversity in a tropical dry forest
Patterns of tree abundance and dispersion in a tropical deciduous (dry) forest are summarized. The generalization that tropical trees have spaced adults did not hold. All species were either clumped or randomly dispersed, with rare species more clumped than common species. Breeding system was unrelated to species abundance or dispersion, but clumping was related to mode of seed dispersal. Juvenile densities decreased approximately exponentially away from adults. Rare species gave evidence of poor reproductive performance compared with their performance when common in nearby forests. Patterns of relative species abundance in the dry forest are compared with patterns in other forests, and are explained by a simple stochastic model based on random-walk immigration and extinction set in motion by periodic community disturbance.
Statistical Analysis and Modelling of Spatial Point Patterns. John Wiley & Sons, London
Herbivores and the number of tree species in tropical forests
Consp- ecific negative density dependence and forest diversity
Conspecific negative density-dependent establishment, in which local abundance negatively affects establishment of conspecific seedlings through host-specific enemies, can influence species diversity of plant communities, but the generality of this process is not well understood. We tested the strength of density dependence using the United States Forest Service's Forest Inventory and Analysis database containing 151 species from more than 200,000 forest plots spanning 4,000,000 square kilometers. We found that most species experienced conspecific negative density dependence (CNDD), but there was little effect of heterospecific density. Additionally, abundant species exhibited weaker CNDD than rarer species, and species-rich regions exhibited stronger CNDD than species-poor regions. Collectively, our results provide evidence that CNDD is a pervasive mechanism driving diversity across a gradient from boreal to subtropical forests.
Species-habitat associations change in a subtropical forest of China
Establishment of Xishuangbanna tropical forest dynamics plot: species compositions and spatial distribution patterns
西双版纳热带森林动态监测样地——树种组成与空间分布格局
The community-level consequences of seed dispersal patterns
Spatial distributions of tree species in a subtropical forest of China
Point patterns of tree distribution determined by habitat heterogeneity and dispersal limitation
Experimental evidence for a phylogenetic Janzen- Connell effect in a subtropical forest
Observational evidence increasingly suggests that the Janzen-Connell effect extends beyond the species boundary. However, this has not been confirmed experimentally. Herein, we present both observational and experimental evidence for a phylogenetic Janzen-Connell effect. In a subtropical forest in Guangdong province, China, we observed that co-occurring tree species are less phylogenetically related than expected. The inhibition effects of neighbouring trees on seedling survival decreased with increasing phylogenetic distance between them. In a shade-house experiment, we studied seedling survival of eight species on soil collected close to Castanopsis fissa relative to their survival on soil close to their own adult trees, and found that this relative survival rate increased with phylogenetic distance from C. fissa. This phylogenetic signal disappeared when seedlings were planted in fungicide-treated soil. Our results clearly support negative effects of phylogenetically similar neighbouring trees on seedling survival and suggest that these effects are caused by associated host-specific fungal pathogens.
On the emergent spatial structure of size-structured populations: When does self-thinning lead to a reduction in clustering?
Dispersal limitation and environmental structure interact to restrict the occupation of optimal habitat
The second-order analysis of stationary point processes
Spatial patterns in an old-growth Nothofagus obliqua forest in south-central Chile
Seed dispersal and spatial pattern in tropical trees
Theories of tropical tree diversity emphasize dispersal limitation as a potential mechanism for separating species in space and reducing competitive exclusion. We compared the dispersal morphologies, fruit sizes, and spatial distributions of 561 tree species within a fully mapped, 50-hectare plot of primary tropical forest in peninsular Malaysia. We demonstrate here that the extent and scale of conspecific spatial aggregation is correlated with the mode of seed dispersal. This relationship holds for saplings as well as for mature trees. Phylogenetically independent contrasts confirm that the relationship between dispersal and spatial pattern is significant even after controlling for common ancestry among species. We found the same qualitative results for a 50-hectare tropical forest plot in Panama. Our results provide broad empirical evidence for the importance of dispersal mode in establishing the long-term community structure of tropical forests.
Quantifying effects of habitat heterogeneity and other clustering processes on spatial distributions of tree species
Spatially explicit consideration of species distribution can significantly add to our understanding of species coexistence. In this paper, we evaluated the relative importance of habitat heterogeneity and other clustering processes (e.g., dispersal limitation, collectively called the non-habitat clustering process) in explaining the spatial distribution patterns of 341 tree species in three stem-mapped 25-50 ha plots of tropical, subtropical, and temperate forests. Their relative importance was estimated by a method that can take one mechanism into account when estimating the effects of the other mechanism and vice versa. Our results demonstrated that habitat heterogeneity was less important in explaining the observed species patterns than other clustering processes in plots with flat topography but was more important in one of the three plots that had a complex topography. Meanwhile, both types of clustering mechanisms (habitat or non-habitat) were pervasive among species at the 50-ha scale across the studied plots. Our analyses also revealed considerable variation among species in the relative importance of the two types of mechanism within each plot and showed that this species-level variation can be partially explained by differences in dispersal mode and growth form of species in a highly heterogeneous environment. Our findings provide new perspectives on the formation of species clustering. One important finding is that a significant species-habitat association does not necessarily mean that the habitat heterogeneity has a decisive influence on species distribution. The second insight is that the large species-level variation in the relative importance of the two types of clustering mechanisms should not be ignored. Non-habitat clustering processes can play an important role on species distribution.
Effects of topography on tropical lower montane forests under different geological conditions on Mount Kinabalu, Borneo
Tree regeneration strategies in a lowland Nothofagus-dominated forest in south-central Chile
Point pattern analysis based on different null models for detecting spatial patterns
基于不同零模型的点格局分析
Rings, circles, and null- models for point pattern analysis in ecology
Analyzing the spatial structure of a Sri Lankan tree species with multiple scales of clustering
Clustering at multiple critical scales may be common for plants since many different factors and processes may cause clustering. This is especially true for tropical rain forests for which theories explaining species coexistence and community structure rest heavily on spatial patterns. We used point pattern analysis to analyze the spatial structure of Shorea congestiflora, a dominant species in a 25-ha forest dynamics plot in a rain forest at Sinharaja World Heritage Site (Sri Lanka), which apparently shows clustering at several scales. We developed cluster processes incorporating two critical scales of clustering for exploring the spatial structure of S. congestiflora and interpret it in relation to factors such as competition, dispersal limitation, recruitment limitation, and Janzen-Connell effects. All size classes showed consistent large-scale clustering with a cluster radius of approximately 25 m. Inside the larger clusters, small-scale clusters with a radius of 8 m were evident for recruits and saplings, weak for intermediates, and disappeared for adults. The pattern of all trees could be divided into two independent patterns: a random pattern (nearest neighbor distance > 8 m) comprising approximately 12% of the trees and a nested double-cluster pattern. This finding suggests two independent recruitment and/or seed dispersal mechanisms. Saplings were several times as abundant as recruits and may accumulate several recruit generations. Recruits were only weakly associated with adults and occupied about half of the large-scale clusters, but saplings almost all. This is consistent with recruitment limitation. For approximately 70% (95%) of all juveniles the nearest adult was less than 26 m away (53 m), suggesting a dispersal limitation that may also be related to the critical large-scale clustering. Our example illustrates the manner in which the use of a specific and complex null hypothesis of spatial structure in point pattern analysis can help us better understand the biology of a species and generate specific hypotheses to be further investigated in the field.
Strong density- and diversity-related effects help to maintain tree species diversity in a neotropical forest
Analysis of spatial point pattern for plant species
植物种群空间分布的点格局分析
Climate changes characteristics over tropical mountain rainforest in Jianfengling during the recent 26 years: radiation, moisture, and wind factors
海南岛尖峰岭热带山地雨林区26年的气候变化特征——光、水和风因子
The relative importance of Janzen-Connell effects in influence- ing the spatial patterns at the gutianshan subtropical forest
The Janzen-Connell hypothesis is among the most important theories put forward to explain species coexistence in species-rich communities. However, the relative importance of Janzen-Connell effects with respect to other prominent mechanisms of community assembly, such as dispersal limitation, self-thinning due to competition, or habitat association, is largely unresolved. Here we use data from a 24-ha Gutianshan subtropical forest to address it. First we tested for significant associations of adults, juveniles, and saplings with environmental variables. Second we evaluated if aggregation decreased with life stage. In a third analysis we approximately factored out the effect of habitat association and comprehensively analyzed the spatial associations of intraspecific adults and offspring (saplings, juveniles) of 46 common species at continuous neighborhood distances. We found i) that, except for one, all species were associated with at least one environmental variable during at least one of their life stages, but the frequency of significant habitat associations declined with increasing life stage; ii) a decline in aggregation with increasing life stage that was strongest from juveniles to adults; and iii) intraspecific adult-offspring associations were dominated by positive relationships at neighborhood distances up to 10 m. Our results suggest that Janzen-Connell effects were not the dominant mechanisms in structuring the spatial patterns of established trees in the subtropical Gutianshan forest. The spatial patterns may rather reflect the joint effects of size-dependent self-thinning, dispersal limitation and habitat associations. Our findings contribute to a more comprehensive understanding of the relative importance of Janzen-Connell effects in influencing plant community structure under strong topographic heterogeneity.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}