


陆地生态系统的生物量是全球碳循环过程中的重要组成部分(Scurlock et al., 2002), 也是估算地球支持能力的重要指标(方精云等, 1996)。草地生态系统生物量的大小不仅很大程度上决定了其生态系统的碳积累, 而且决定其产草量, 因而影响其生态与生产服务功能(Jobbágy & Sala, 2000)。草地生态系统生物量大小受到土壤中可利用营养元素、土壤pH值、温度、降水等多种理化因子的影响(Chapin & Matson, 2011)。其中氮、磷均是植物生长所必需的大量营养元素, 在植物生长、发育和繁殖等过程中有着重要的作用, 自然状态下, 很多生态系统生产力都表现为氮限制或者磷限制或者氮磷共限制(Elser et al., 2007; Vitousek et al., 2010)。
氮是许多生态系统中限制植物生长的最关键营养元素, 因此氮输入的增加可提高土壤中可利用氮的含量, 消除或缓解氮的限制, 促进植物的生长, 所以氮添加一般会促进草地生态系统的地上生物量(Zavaleta et al., 2003; LeBauer & Treseder, 2008; Lee et al., 2010); 磷是草地生态系统中除氮以外限制植物生长的最重要营养元素(Vance et al., 2003), 因此磷的添加一般也会促进植物生长而增加地上生物量(Elser et al., 2007)。但是一些生态系统主要受氮的限制, 而另一些生态系统主要受磷的限制, 这是由于: (1)自然条件下, 氮进入陆地生态系统的主要途径是固氮生物对大气中氮气的固定, 而磷则主要通过原生矿物的风化作用进入生态系统(Odum & Barrett, 2005), 因此一般认为生态系统在演替早期主要表现为氮限制, 在演替后期则主要为磷限制(Verhoeven et al., 1996); (2)控制氮可利用性的反应主要为生物反应, 而控制磷可利用性的反应则主要是地球化学反应(Chapin & Matson, 2011); (3)工业革命以来, 人类活动对环境的影响日益深重, 有些生态系统接受了大量的外源性氮磷输入, 引起生态系统营养限制的变化, 所以冻原生态系统(Shaver, 1986)及受火烧干扰频繁的生态系统(Raison, 1979; Hungate et al., 2003)被认为主要受可利用氮限制, 而具古老土壤的热带地区生态系统等则被认为主要受可利用磷限制(Vitousek, 2004)。
青藏高原高寒草甸地处高海拔地区(Zhao & Zhou, 1999), 尽管土壤中储存着大量的氮素和磷素, 特别是有机氮和有机磷, 但是由于低温的限制, 有机氮、磷矿化作用比较低, 可利用氮、磷含量仅占土壤全氮、全磷含量的1%左右, 植物的生长极有可能受到可利用氮、磷的限制(周兴民, 2001)。在青藏高原海北高寒草甸和甘南高寒草甸的一系列氮添加实验表明, 氮添加可促进植物生长(沈振西等, 2002; Ren et al., 2010; Song et al., 2012), 证实高寒草甸植物生长受土壤中可利用氮的限制, 但高寒草甸植物生长主要受氮限制, 还是磷限制, 抑或氮磷共限制尚不明确。近年来青藏高原正经历着明显的增温过程(Liu & Chen, 2000; IPCC, 2007), 土壤增温促进了土壤有机质的分解(Luo et al., 2010; Rui et al., 2012), 另一方面, 青藏高原东缘地区近年氮沉降明显, 并具有逐年增加的趋势(Lü & Tian, 2007), 这均会使青藏高原高寒草甸土壤中的可利用氮、磷含量增加, 因此研究该区域生态系统植物生物量对氮、磷添加的响应, 不仅有助于深入了解全球变化背景下高寒草甸生态系统碳输入过程的变化, 对研究区域和全球碳平衡具有重要意义, 而且将为高寒草甸的健康发展和科学管理提供理论依据。
本研究选取位于青藏高原东北部的高寒草甸生态系统, 研究该生态系统群落生物量对氮、磷添加的响应, 拟解决两个问题: (1)氮、磷添加如何通过影响不同功能群植物生长而影响青藏高原高寒草甸生物量? (2)青藏高原高寒草甸植物生长主要受氮限制, 磷限制, 还是氮磷共限制?
1 材料和方法
1.1 研究区域的自然概况
本研究在青海海北高寒草地生态系统国家野外科学观测研究站(海北站)进行。海北站地处青藏高原东北隅的青海海北藏族自治州门源县境内, 地理范围为: 37°29′-37°45′ N, 101°12′-101°23′ E, 处于祁连山北支冷龙岭东段南麓坡地的大通河河谷西段, 海拔为3200-3600 m (气象观测站海拔高度为3220 m)。由于地处亚洲大陆腹地, 海北站站区属于典型的高原大陆性气候, 夏季为东南季风气候, 冬季受西伯利亚寒流的影响, 一年中无明显的四季之分, 只有冷暖季之别, 暖季短暂而凉爽湿润, 冷季漫长而寒冷干燥。海北站多年(1981-2000年)平均气温为-1.7 ℃, 极端最高气温23.7 ℃, 极端最低气温为-37.1 ℃; 年降水量为561 mm, 降水多集中在5-9月, 约占全年降水量的80%。海北站站区的土壤呈现土壤发育年轻、土层薄、表层具有较厚的草皮层, 草皮层以下具有较厚的腐殖质层, 有机质含量及全量养分丰富而有效养分贫乏等特征。本研究选择了长期适应于高寒生态环境的耐寒中生植物高寒矮生嵩草(Kobresia humilis)草甸为研究对象。矮生嵩草草甸植被具有植物生长低矮、群落总覆盖度高(90%以上)、种类组成较多、群落结构简单, 一般为单层结构, 初级生产力较高等特征, 其植物群落以矮生嵩草、异针茅(Stipa aliena)、垂穗披碱草(Elymus nutans)、早熟禾(Poa annua)、黄花棘豆(Oxytropis ochrocephala)、花苜蓿(Medicago ruthenica)、麻花艽(Gentiana straminea)等为优势种。
1.2 实验设计
2009年5月, 我们选择了位于海北站东北方向约2 km处的一片草场作为本研究的实验样地。实验前此处为当地牧民的冬春季牧场, 因为本实验为养分添加实验, 为排除实验前冬季放牧过程中动物粪便对实验结果的影响, 围栏后人工捡除样方内所有可见的动物粪便。
实验采取完全随机区组设计, 共4个处理, 分别为: 对照; 氮添加(N); 磷添加(P); 氮磷同时添加(NP)。N的施用量为10 g·m-2·a-1, 以尿素的形式添加; P的施用量为5 g·m-2·a-1, 以Ca(H2PO4)2·H2O的形式添加。每个处理6个重复, 共计24个样方, 样方面积为 3 m × 3 m。
将预先称好的颗粒状肥料一次性均匀撒在样方内进行养分添加。为保证施肥效果, 施肥时间选择在生长季中期的一个雨天或晴天的傍晚进行, 雨水或次日形成的露水使得肥料迅速溶解进入土壤。实验期间, 养分添加日期分别为2009年7月15日、2010年7月5日、2011年7月15日和2012年6月22日。
对照处理不采取任何施肥措施。
1.3 地上生物量、地下生物量的测定及地下和地上生物量比值
地上生物量和地下生物量均用收获法测定。连续4年养分添加后, 在2012年生长季盛期(8月下旬至9月上旬), 在每个样方设置0.25 m × 0.25 m的样方, 收获每个样方框内植物的地上部分, 分功能群之后于65 ℃烘干至恒重。称量后, 计算单位面积内群落尺度和功能群尺度上的地上生物量。实验分3个功能群: 禾草, 即禾本科植物; 莎草, 即莎草科植物; 杂类草, 包括除禾本科及莎草科以外的草本植物, 测定指标包括各功能群的绝对生物量, 及其在群落总生物量中所占的比例。
在收获了植物地上部分的样方框内, 用3.5 cm直径的根钻, 以0-10、10-20、20-30和30-40 cm的土层, 每层三钻, 取得土柱。将每个样方同一层次的3个土柱混合后装于网袋, 带回实验室用水将植物根系冲洗干净后, 于65 ℃烘干至恒重。称量后计算单位面积内的地下生物量。
以地下和地上生物量比值(R/S)表示生物量在地上、地下间的分配关系, 计算方法: R/S =地下生物量/地上生物量。
1.4 生物量碳库估算
本文中使用常数(0.45)将所测定的生物量统一转化为生物量碳含量, 单位为g C·m-2 (Piao et al., 2007)。
1.5 统计分析
用双因素方差分析(two-way ANOVA)确定N添加和P添加对地上生物量、地下生物量、总生物量、R/S、各功能群地上绝对生物量及其在群落中所占的比例的主效应及交互效应。实验数据的统计分析均在SPSS 16.0 (SPSS, Chicago, USA)中进行, 统计图形在SigmaPlot 10.0中绘制。
2 结果
2.1 不同功能群植物的地上生物量
N、P添加均极显著增加了禾草的地上绝对生物量及其在群落总生物量中所占的比例, 同时均显著降低了杂类草在群落总生物量中的比例, 此外P添加极显著降低了莎草地上绝对生物量及其在群落总生物量中所占的比例(表1)。对各功能群的地上绝对生物量, N添加使禾草生物量增加了50.5%; P添加使禾草生物量增加了89.8%, 使莎草生物量降低了82.9% (图1); 对各功能群在群落总生物量中所占的比例, N添加使禾草的比例增加了25.9%, 使杂类草的比例降低了35.6%; P添加使禾草的比例增加了31.5%, 使莎草和杂类草的比例分别降低了88.4%和33.7% (图2)。无论是对各功能群的绝对生物量或其在群落总生物量中所占的比例, N、P添加均无交互作用(表1)。
图1
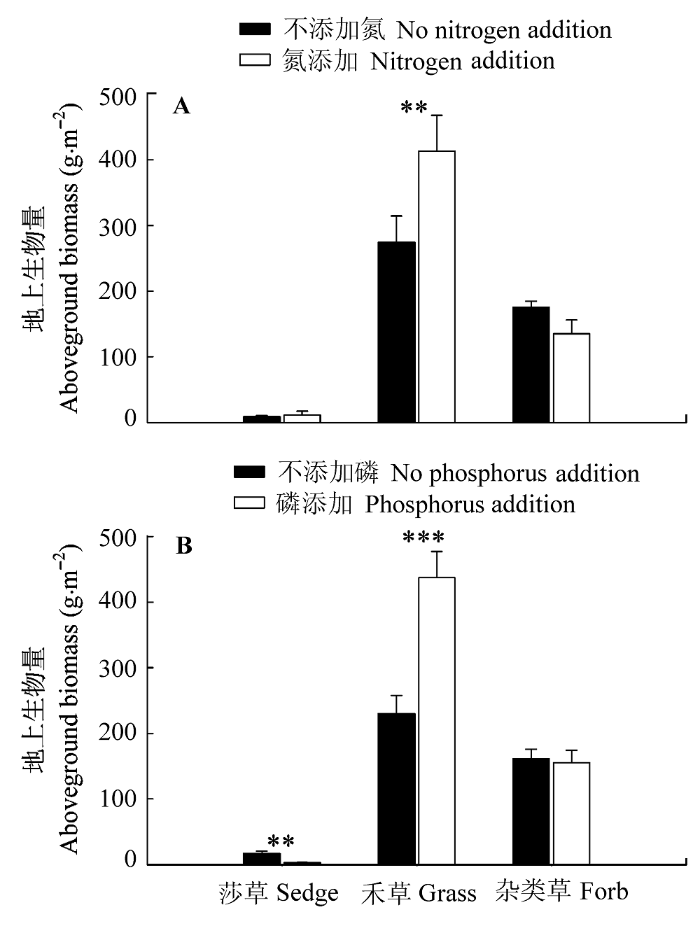
图1
氮(A)、磷(B)添加对3种不同功能群地上生物量的影响(平均值±标准误差)。
Fig. 1
Effects of nitrogen addition (A) and phosphorus addition (B) on aboveground biomass of three different functional groups (mean ± SE). **, p < 0.01; ***, p < 0.001.
图2
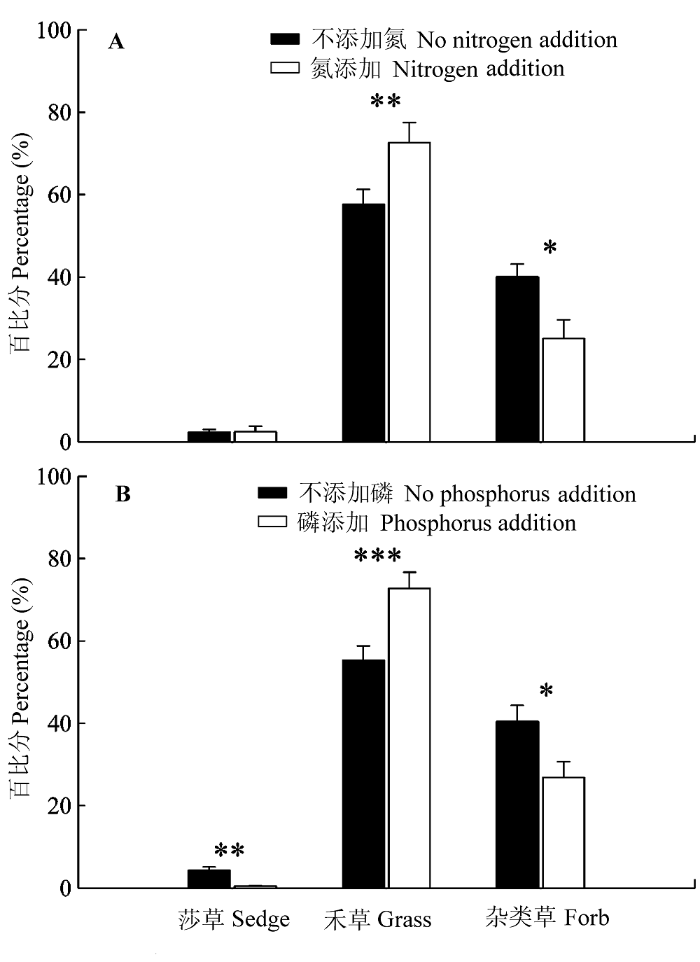
图2
氮(A)、磷(B)添加对3种不同功能群的地上生物量在群落总生物量中所占比例的影响(平均值±标准误差)。
Fig. 2
Effects of nitrogen addition (A) and phosphorus addition (B) on the proportions of the three different functional groups aboveground biomass of the community biomass (mean ± SE). *, p < 0.05; **, p < 0.01; ***, p < 0.001.
表1 氮、磷添加对不同功能群的地上生物量(g·m-2·a-1)和其在群落总生物量中所占比例(%)的影响的双因素方差分析表
Table 1
| 氮添加 N addition | 磷添加 P addition | 氮磷交互作用 N × P interaction | ||||||
|---|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | |||
| 莎草生物量 Sedge biomass | 0.28 | 0.611 | 15.60 | 0.006 | 2.75 | 0.141 | ||
| 莎草生物量 Sedge biomass (%) | 0.01 | 0.910 | 17.53 | 0.004 | 0.79 | 0.403 | ||
| 禾本生物量 Grass biomass | 15.24 | 0.006 | 39.95 | <0.001 | 0.80 | 0.400 | ||
| 禾本生物量 Grass biomass (%) | 12.78 | 0.009 | 19.64 | 0.003 | 0.19 | 0.676 | ||
| 杂类草生物量 Forb biomass | 1.91 | 0.210 | 0.13 | 0.730 | 0.97 | 0.358 | ||
| 杂类草生物量 Forb biomass (%) | 9.91 | 0.016 | 9.34 | 0.018 | 0.04 | 0.844 | ||
加粗数字表示效应显著。
Bold numbers indicate significant effects.
2.2 地上生物量、地下生物量及总生物量和R/S
对照处理地上生物量为326 g·m-2·a-1, N、P添加均显著促进了地上生物量的增加, 分别提高了24% (p = 0.008)和52% (p < 0.001); 对照处理地下生物量为1240 g·m-2·a-1, N添加对地下生物量没有显著影响(p = 0.216), 而P添加有增加地下生物量的趋势(27%, p = 0.060)。对照处理总生物量为1566g·m-2·a-1, N添加对总生物量无显著影响(p = 0.969), 而P添加显著促进了总生物量(30%, p = 0.008)。对于地上、地下及总植物生物量, N、P添加均无交互作用(表2; 图3)。对照处理R/S为3.96, N添加对R/S的降低作用接近显著(p = 0.086), 而P添加对其影响不显著(表2)。
表2 氮、磷添加对地上生物量(AGB)、地下生物量(BGB)、总生物量(TB)和地下地上生物量比(R/S)影响的双因素方差分析表
Table 2
| 氮添加 N addition | 磷添加 P addition | 氮磷交互作用 N × P interaction | ||||||
|---|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | |||
| AGB (g·m-2·a-1) | 13.61 | 0.008 | 66.38 | < 0.001 | 0.95 | 0.363 | ||
| BGB (g·m-2·a-1) | 1.80 | 0.216 | 4.82 | 0.060 | 0.02 | 0.880 | ||
| TB (g·m-2·a-1) | 0.002 | 0.969 | 15.22 | 0.008 | 1.54 | 0.260 | ||
| R/S | 3.50 | 0.086 | 1.24 | 0.309 | 0.07 | 0.806 | ||
加粗数字表示效应显著。
Bold numbers indicate significant effects.
图3
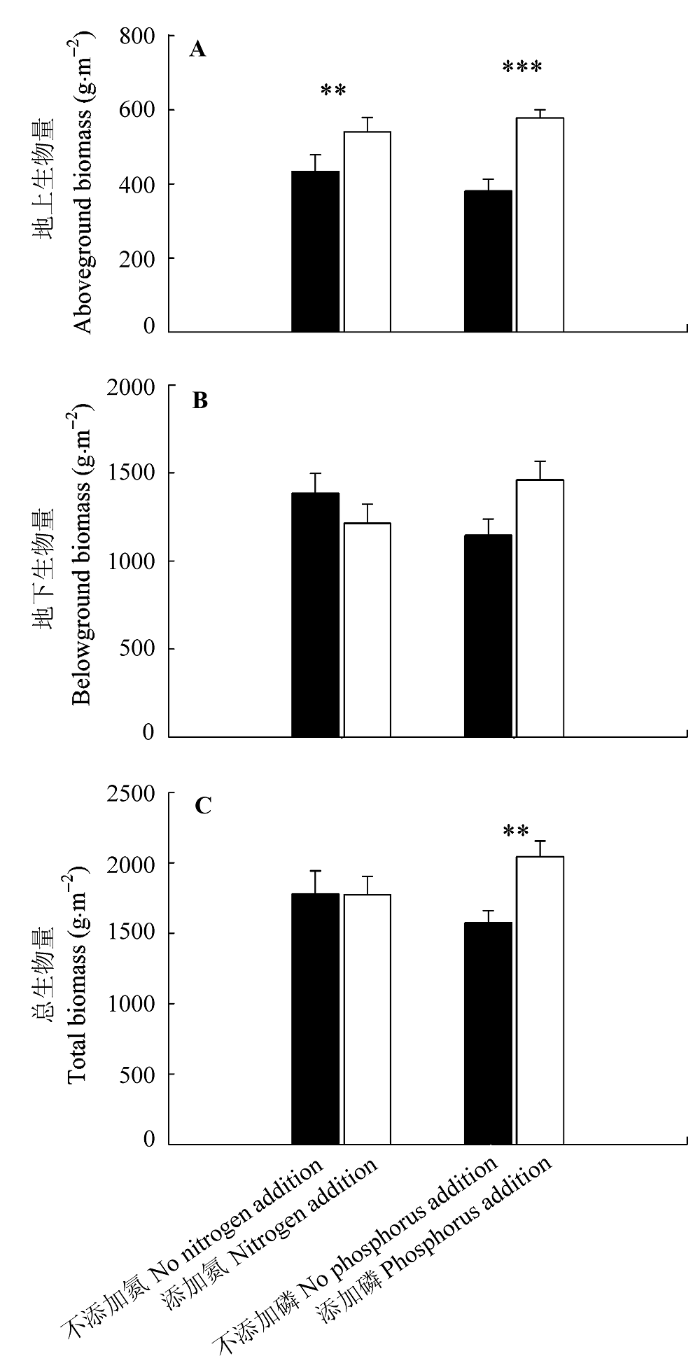
图3
氮、磷添加对地上生物量(A)、地下生物量(B)和总生物量(C)的影响(平均值±标准误差)。
Fig. 3
Effects of nitrogen and phosphorus additions on aboveground biomass (A), belowground biomass (B) and total biomass (C) (mean ± SE). **, p < 0.01; ***, p < 0.001.
2.3 生物量碳库
表3列出了青藏高原高寒草甸生态系统的生物量碳库。不同处理下地上生物量碳库为: 对照, 146.5 g C·m-2; N添加, 204.8 g C·m-2; P添加, 250.9 g C·m-2; NP添加, 281.5 g C·m-2。不同处理下地下生物量碳库为: 对照, 558.0 g C·m-2; N添加, 510.5 g C·m-2; P添加, 709.1 g C·m-2; NP添加, 598.8 g C·m-2。总生物量碳库为: 对照, 704.6 g C·m-2; N添加, 715.3 g·C m-2; P添加, 960.0 g C·m-2; NP添加, 880.3 g C·m-2。N、P添加对生物量碳库的效应与其对生物量的效应相同。
表3 不同处理下的地上(AGB)、地下(BGB)和总生物量(TB)碳库(平均值±标准误差)
Table 3
| 对照 CK | 氮添加 N addition | 磷添加 P addition | 氮磷添加 N, P addition | |
|---|---|---|---|---|
| AGB (g C·m-2) | 146.5 ± 14.0a | 204.8 ± 7.8b | 250.9 ± 12.7c | 281.5 ± 7.9d |
| BGB (g C·m-2) | 558.0 ± 68.4a | 510.5 ± 36.8a | 709.1 ± 49.5b | 598.8 ± 101.3a |
| TB (g C·m-2) | 704.6 ± 64.3a | 715.3 ± 40.9a | 960.0 ± 41.8b | 880.3 ± 95.8ab |
不同小写字母表示处理间差异显著(p < 0.05)。
Different lowercase letters indicate significant difference among treatments (p < 0.05).
2.4 氮磷效应的比较
如2.2节中所述, 本研究中, N、P添加均促进了地上生物量的增加, P的促进作用(52%)高于N的促进作用(24%); 与此同时, P添加有增加地下生物量的趋势(27%), 而N添加对地下生物量没有显著作用; 对于总生物量, P的促进作用(30%)明显, 而N无显著影响。
3 讨论
青藏高原高寒草甸土壤中总N、总P含量均十分丰富, 然而由于低温限制了微生物对土壤有机质的分解作用, 能为植物吸收利用的有效N和有效P含量却很贫乏, 大约是总量的1% (周兴民, 2001)。本研究结果显示, N、P添加均显著促进了高寒草甸生态系统的地上生物量, 说明N、P均是限制青藏高原高寒草甸植物生长的重要营养元素。
3.1 青藏高原高寒草甸不同功能型植物对N、P添加的响应
本研究中, N、P添加均显著增加了禾草的绝对生物量及其在群落总生物量中所占的比例, 而对杂类草绝对生物量没有显著影响, 但降低了其在总生物量中所占的比例。禾草植物, 如垂穗披碱草、 异针茅等, 其植株较高, 处于群落的上层, 根系主要为须根系, 对水分、营养元素和光辐射的竞争在群落中处于优势地位。N、P的添加缓解了其对禾草植物生长的限制, 同时因禾草植物处于群落上方, 可获得更丰富的光资源(Hautier et al., 2009), 从而其生长受到明显的促进, 生物量大幅度增加。此外, 在青藏高原高寒草甸, 由于寒冷气候和多风的生态条件的综合影响, 杂类草植物多呈莲座状、半莲座状或垫状, 如美丽风毛菊(Saussurea pulchra)、麻花艽等, 一直处于群落的下层(周兴民, 2001)。尽管N、P添加可缓解矿质营养元素对杂类草植物生长的限制, 但由于上层生长迅速的禾草植物的遮阴作用增强, 会在一定程度上抵消N、P添加的促进作用, 最终其生物量无显著变化, 且由于禾草植物生物量增加明显, 使得杂类草在群落总生物量中所占的比例降低。莎草植物的绝对生物量及其在群落总生物量中所占的比例在N添加后无显著变化, 而在P添加后显著降低, 同一区域其他的研究中, N、P添加对莎草植物生物量没有显著影响(沈振西等, 2002; 辛小娟, 2011), 可能是由于与禾草植物相比, 莎草植物对养分资源的波动适应性较强(沈振西等, 2002)。在本研究中, 尽管N、P均促进了禾草植物生长, 但是P添加的促进作用(89.8%)大于N添加的作用(50.5%), 因此P添加处理下, 禾草植物的遮阴作用更强, 故而处于群落下层的莎草植物对光资源的获取受到较强的抑制, 导致生物量降低。
3.2 青藏高原高寒草甸生物量对N、P添加的响应
本研究结果显示, N、P添加均显著增加了青藏高原高寒草甸的地上生物量, 与前人的研究结果一致。地上生物量的增加主要是由于N、P添加后禾草生物量的大幅度增加, 与Zavaleta等(2003)和Henry等(2006)的研究结果一致。
与地上生物量对N、P添加的一致性正响应(LeBauer & Treseder, 2008)不同, 很多研究表明根系生物量对N、P添加的响应在不同的生态系统中差异较大, 如Majdi和Anderson (2005)发现N添加后云杉(Picea abies)的根系生物量增加, 而Henry等(2006)发现根系生物量对N添加没有响应, Stöcklin等(1998)发现根系生物量对P添加没有响应, 也有研究发现根系生物量在P添加后降低(Keith et al., 1997)或者增加(Shaver et al., 1998)。可能的原因是: (1)根据Liebig最小因子定律, 植物生长受限于生态系统中最受限制的因子, 因此根系生物量对N、P添加后的响应取决于在N、P限制减缓或消除后, 是否又受限于其他的生态因子, 如水分等(Chapin & Matson, 2011); (2) N、P对地下生物量的效应取决于其对总生物量的影响和对R/S的影响的平衡。本研究结果显示, 施肥4年后, N添加对R/S值的降低作用接近显著, 而P添加对R/S值没有影响, 同时N添加对总生物量没有显著影响, 而P添加显著增加了总生物量, 因此N倾向于降低地下生物量, P倾向于增加地下生物量。
3.3 N、P效应的比较
根据Liebig最小因子定律“植物的生长取决于那些处于最少量状态的营养元素”, 青藏高原高寒草甸植物生长极有可能更受土壤中可利用P含量的限制。自然状态下, 由于固氮菌的生物固氮作用非常有限, 同时生态系统中的可利用N很容易通过淋溶、挥发、硝化及反硝化等过程从生态系统中损失, 因此, N是限制植物生长的最重要的营养元素(Mooney et al., 1987), 很多生态系统表现为N限制。然而, 尽管研究者们普遍认为热带雨林等古老土壤中更易受P的限制, 但由于P的许多无机结构和有机结构在土壤中保持着不溶形式, 其中磷酸盐(PO43-)是土壤中可利用P的主要形式, 并且磷酸盐在高pH条件下, 易与钙结合形成磷酸钙(沉淀), 在低pH条件下易与氧化铁、铝结合成不溶的沉淀物, 只在pH为6.5附近的狭窄范围内为可溶形式, 此外, P与铁、铝和钙形成的大部分化合物沉淀不能被植物吸收利用(Chapin & Matson, 2011), 因此即使是在P含量丰富的土壤中P的可利用性也受到了极大的限制。本研究中, P添加使得地上生物量增加了约50%, 同时有促进地下生物量的趋势, 因而显著地增加了总生物量(30%), 比较而言, N添加仅使得地上生物量增加24%。有研究表明, 在青藏高原高寒草甸, 植物对N的重吸收效率平均为65.2%, 而对P的重吸收效率平均为67.4% (Jiang et al., 2012)。因此, 尽管N、P添加均可促进高寒草甸植物的生长, 但P的促进作用高于N的促进作用, 土壤中可利用P对植物生长的限制作用极有可能大于N的限制作用。
4 结论
青藏高原高寒草甸是青藏高原地区极为重要的生态系统, 其土壤有机质含量高, 然而由于低温限制了微生物活动, 土壤中可利用养分含量极低, 限制了植物的生长。本研究基于在青藏高原高寒草甸连续4年N、P添加后对群落地上生物量和地下生物量的测定, 探讨了青藏高原高寒草甸生态系统碳输入对N、P添加的响应, 得到以下结论: (1) N、P添加均显著促进了青藏高原高寒草甸的地上生物量, 分别增加了24%和52%; 不同功能群地上生物量的响应不同, 其中N、P添加均增加了禾草的绝对生物量及其在群落总生物量中所占的比例, 仅P添加降低了莎草生物量及其在群落总生物量中的比例, 而杂类草生物量在群落中的比例在N、P添加后均降低。因此, 养分添加后禾草类植物生物量(50.5%, 89.8%)的显著增加是N、P提高地上生物量的主要来源。(2) N添加对高寒草甸地下生物量无显著影响, 而P添加有促进地下生物量的趋势。(3) N添加对高寒草甸植物总生物量无显著影响, 而P添加显著促进了植物总生物量。(4)高寒草甸植物地上生物量碳库为147 g C·m-2, 地下生物量碳库为558 g C·m-2, 总生物量碳库约为705 g C·m-2, N、P添加均增加了植物地上生物量碳库, 但只有P添加增加了总生物量碳库。
本研究结果表明, N、P添加可缓解青藏高原高寒草甸植物生长的营养限制, 促进植物地上部分的生长, 然而高寒草甸植物的生长极有可能更受土壤中可利用P含量的限制。
参考文献
Principles of Terrestrial Ecosystem Ecology
2nd edn.
Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems

The cycles of the key nutrient elements nitrogen (N) and phosphorus (P) have been massively altered by anthropogenic activities. Thus, it is essential to understand how photosynthetic production across diverse ecosystems is, or is not, limited by N and P. Via a large-scale meta-analysis of experimental enrichments, we show that P limitation is equally strong across these major habitats and that N and P limitation are equivalent within both terrestrial and freshwater systems. Furthermore, simultaneous N and P enrichment produces strongly positive synergistic responses in all three environments. Thus, contrary to some prevailing paradigms, freshwater, marine and terrestrial ecosystems are surprisingly similar in terms of N and P limitation.
Competition for light causes plant biodiversity loss after eutrophication
Human activities have increased the availability of nutrients in terrestrial and aquatic ecosystems. In grasslands, this eutrophication causes loss of plant species diversity, but the mechanism of this loss has been difficult to determine. Using experimental grassland plant communities, we found that addition of light to the grassland understory prevented the loss of biodiversity caused by eutrophication. There was no detectable role for competition for soil resources in diversity loss. Thus, competition for light is a major mechanism of plant diversity loss after eutrophication and explains the particular threat of eutrophication to plant diversity. Our conclusions have implications for grassland management and conservation policy and underscore the need to control nutrient enrichment if plant diversity is to be preserved.
Interactive effects of fire, elevated carbon dioxide, nitrogen deposition, and precipitation on a California annual grassland
Although it is widely accepted that elevated atmospheric carbon dioxide (CO2), nitrogen (N) deposition, and climate change will alter ecosystem productivity and function in the coming decades, the combined effects of these environmental changes may be nonadditive, and their interactions may be altered by disturbances, such as fire. We examined the influence of a summer wildfire on the interactive effects of elevated CO2, N deposition, and increased precipitation in a full-factorial experiment conducted in a California annual grassland. In unburned plots, primary production was suppressed under elevated CO2. Burning alone did not significantly affect production, but it increased total production in combination with nitrate additions and removed the suppressive effect of elevated CO2. Increased production in response to nitrate in burned plots occurred as a result of the enhanced aboveground production of annual grasses and forbs, whereas the removal of the suppressive effect of elevated CO2 occurred as a result of increased aboveground forb production in burned, CO2-treated plots and decreased root production in burned plots under ambient CO2.The tissue nitrogen–phosphorus ratio, which was assessed for annual grass shoots, decreased with burning and increased with nitrate addition. Burning removed surface litter from plots, resulting in an increase in maximum daily soil temperatures and a decrease in soil moisture both early and late in the growing season. Measures of vegetation greenness, based on canopy spectral reflectance, showed that plants in burned plots grew rapidly early in the season but senesced early. Overall, these results indicate that fire can alter the effects of elevated CO2 and N addition on productivity in the short term, possibly by promoting increased phosphorus availability.]]>
Disturbance and element interactions
In: Melillo JM, Fieldd CB, Moldan B eds. Interactions of the Major Biogeochemical Cycles.
Climate Change 2007: Synthesis Report
Nutrient resorption of coexistence species in alpine meadow of the Qinghai-Tibetan Plateau explains plant adaptation to nutrient-poor environment
Controls of grass and shrub aboveground production in the Patagonian steppe
Allocation of carbon in a mature eucalypt forest and some effects of soil phosphorus availability
Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed
Our meta-analysis of 126 nitrogen addition experiments evaluated nitrogen (N) limitation of net primary production (NPP) in terrestrial ecosystems. We tested the hypothesis that N limitation is widespread among biomes and influenced by geography and climate. We used the response ratio (R approximately equal ANPP(N)/ANPP(ctrl)) of aboveground plant growth in fertilized to control plots and found that most ecosystems are nitrogen limited with an average 29% growth response to nitrogen (i.e., R = 1.29). The response ratio was significant within temperate forests (R = 1.19), tropical forests (R = 1.60), temperate grasslands (R = 1.53), tropical grasslands (R = 1.26), wetlands (R = 1.16), and tundra (R = 1.35), but not deserts. Eight tropical forest studies had been conducted on very young volcanic soils in Hawaii, and this subgroup was strongly N limited (R = 2.13), which resulted in a negative correlation between forest R and latitude. The degree of N limitation in the remainder of the tropical forest studies (R = 1.20) was comparable to that of temperate forests, and when the young Hawaiian subgroup was excluded, forest R did not vary with latitude. Grassland response increased with latitude, but was independent of temperature and precipitation. These results suggest that the global N and C cycles interact strongly and that geography can mediate ecosystem response to N within certain biome types.
A global comparison of grassland biomass responses to CO2 and nitrogen enrichment
Climatic warming in the Tibetan Plateau during recent decades
Spatial and temporal patterns of nitrogen deposition in China: synthesis of observational data
On 5 May 2017, MMS observed a crater-type flux rope on the dawnside tailward magnetopause with fluctuations. The boundary-normal analysis shows that the fluctuations can be attributed to nonlinear Kelvin-Helmholtz (KH) waves. Reconnection signatures such as flow reversals and Joule dissipation were identified at the leading and trailing edges of the flux rope. In particular, strong northward electron jets observed at the trailing edge indicated midlatitude reconnection associated with the 3-D structure of the KH vortex. The scale size of the flux rope, together with reconnection signatures, strongly supports the interpretation that the flux rope was generated locally by KH vortex-induced reconnection. The center of the flux rope also displayed signatures of guide-field reconnection (out-of-plane electron jets, parallel electron heating, and Joule dissipation). These signatures indicate that an interface between two interlinked flux tubes was undergoing interaction, causing a local magnetic depression, resulting in an M-shaped crater flux rope, as supported by reconstruction.
Effect of warming and grazing on litter mass loss and temperature sensitivity of litter and dung mass loss on the Tibetan Plateau
Fine root production and turnover in a Norway spruce stand in northern Sweden: effects of nitrogen and water manipulation
Fine root length production, biomass production, and turnover in forest floor and mineral soil (0–30 cm) layers were studied in relation to irrigated (I) and irrigated-fertilized (IL) treatments in a Norway spruce stand in northern Sweden over a 2-year period. Fine roots (<1 mm) of both spruce and understory vegetation were studied. Minirhizotrons were used to estimate fine root length production and turnover, and soil cores were used to estimate standing biomass. Turnover was estimated as both the inverse of root longevity (RTL) and the ratio of annual root length production to observed root length (RTR). RTR values of spruce roots in the forest floor in I and IL plots were 0.6 and 0.5 y−1, respectively, whereas the corresponding values for RTL were 0.8 and 0.9 y−1. In mineral soil, corresponding values for I, IL, and control (C) plots were 1.2, 1.2, and 0.9 y−1 (RTR) and 0.9, 1.1, and 1 y−1 (RTL). RTR and RTL values of understory vegetation roots were 1 and 1.1 y−1, respectively. Spruce root length production in both the forest floor and the mineral soil in I plots was higher than in IL plots. The IL-treated plots gave the highest estimates of spruce fine root biomass production in the forest floor, but, for the mineral soil, the estimates obtained for the I plots were the highest. The understory vegetation fine root production in the I and IL plots was similar for both the forest floor and the mineral soil and higher (for both layers) than in C plots. Nitrogen (N) turnover in the forest floor and mineral soil layers (summed) via spruce roots in IL, I, and C plots amounted to 2.4, 2.1, and 1.3 g N m−2 y−1, and the corresponding values for field vegetation roots were 0.6, 0.5, and 0.3 g N m−2 y−1. It was concluded that fertilization increases standing root biomass, root production, and N turnover of spruce roots in both the forest floor and mineral soil. Data on understory vegetation roots are required for estimating carbon budgets in model studies.]]>
Exchange of materials between terrestrial ecosystems and the atmosphere
Many biogenic trace gases are increasing in concentration or flux or both in the atmosphere as a consequence of human activities. Most of these gases have demonstrated or potential effects on atmospheric chemistry, climate, and the functioning of terrestrial ecosystems. Focused studies of the interactions between the atmosphere and the biosphere that regulate trace gases can improve both our understanding of terrestrial ecosystems and our ability to predict regional-and global-scale canges in atmospheric chemistry.
Fundamentals of Ecology
5th edn.
Changes in biomass carbon stocks in China’s grasslands between 1982 and 1999
Modification of the soil environment by vegetation fires, with particular reference to nitrogen transformations: a review
Effects of resource additions on species richness and ANPP in an alpine meadow community
Aims Theories based on resource additions indicate that plant species richness is mainly determined by the number of limiting resources. However, the individual effects of various limiting resources on species richness and aboveground net primary productivity (ANPP) are less well understood. Here, we analyzed potential linkages between additions of limiting resources, species loss and ANPP increase and further explored the underlying mechanisms.Methods Resources (N, P, K and water) were added in a completely randomized block design to alpine meadow plots in the Qinghai-Tibetan Plateau. Plant aboveground biomass, species composition, mean plant height and light availability were measured in each plot. Regression and analysis of variance were used to analyze the responses of these measures to the different resource-addition treatments.Important findings Species richness decreased with increasing number of added limiting resources, suggesting that plant diversity was apparently determined by the number of limiting resources. Nitrogen was the most important limiting resource affecting species richness, whereas P and K alone had negligible effects. The largest reduction in species richness occurred when all three elements were added in combination. Water played a different role compared with the other limiting resources. Species richness increased when water was added to the treatments with N and P or with N, P and K. The decreases in species richness after resource additions were paralleled by increases in ANPP and decreases in light penetration into the plant canopy, suggesting that increased light competition was responsible for the negative effects of resource additions on plant species richness.]]>
Warming and grazing increase mineralization of organic P in an alpine meadow ecosystem of Qinghai-Tibet Plateau, China
Estimating net primary productivity from grassland biomass dynamics measurements
Biomass and CO2 flux in wet sedge tundras: responses to nutrients, temperature, and light
Response of plant groups to simulated rainfall and nitrogen supply in alpine Kobresia humilis meadow
高寒矮嵩草草甸植物类群对模拟降水和施氮的响应
Different inter-annual responses to avail- ability and form of nitrogen explain species coexistence in an alpine meadow community after release from grazing
Plant species and functional groups in nitrogen (N) limited communities may coexist through strong eco-physiological niche differentiation, leading to idiosyncratic responses to multiple nutrition and disturbance regimes. Very little is known about how such responses depend on the availability of N in different chemical forms. Here we hypothesize that idiosyncratic year-to-year responses of plant functional groups to availability and form of nitrogen explain species coexistence in an alpine meadow community after release from grazing. We conducted a 6 year N addition experiment in an alpine meadow on the Tibetan Plateau released from grazing by livestock. The experimental design featured three N forms (ammonium, nitrate, and ammonium nitrate), crossed with three levels of N supply rates (0.375, 1.500 and 7.500 g N m(-2) yr(-1) ), with unfertilized treatments without and with light grazing as controls. All treatments showed increasing productivity and decreasing species richness after cessation of grazing and these responses were stronger at higher N rates. Although N forms did not affect aboveground biomass at community level, different functional groups did show different responses to N chemical form and supply rate and these responses varied from year to year. In support of our hypothesis, these idiosyncratic responses seemed to enable a substantial diversity and biomass of sedges, forbs, and legumes to still coexist with the increasingly productive grasses in the absence of grazing, at least at low and intermediate N availability regimes. This study provides direct field-based evidence in support of the hypothesis that idiosyncratic and annually varying responses to both N quantity and quality may be a key driver of community structure and species coexistence. This finding has important implications for the diversity and functioning of other ecosystems with spatial and temporal variation in available N quantity and quality as related to changing atmospheric N deposition, land-use, and climate-induced soil warming.
Effects of elevated CO2 and phosphorus addition on productivity and community composition of intact monoliths from calcareous grassland
/=P > 0.05). Belowground biomass and stubble after two seasons were not different in elevated CO2 and when P was added. The small initial increase in aboveground community biomass under elevated CO2 is explained by the fact that some species, in particular Carex flacca, responded very positively right from the beginning, while others, especially the dominant Bromus erectus, responded negatively to CO2 enrichment. Shifts in community composition towards more responsive species explain the much larger CO2 response in the second year. These shifts, i.e., a decline in xerophytic elements (B. erectus) and an increase in mesophytic grasses and legumes occurred independently of treatments in all monoliths but were accelerated significantly by elevated CO2. The difference in average biomass production at elevated compared to ambient CO2 was higher when P was supplied (at the community level the CO2 response was enhanced from 20% to 33% when P was added, in graminoids from 17% to 27%, in legumes from 4% to 60%, and in C. flacca from 120% to 298% by year two). Based on observations in this and similar studies, we suggest that interactions between CO2 concentration, species presence, and nutrient availability will govern community responses to elevated CO2.]]>
Phosphorus acquisition and use: critical adaptations by plants for securing a nonrenewable resource
Nitrogen- or phosphorus-limited growth in herbaceous, wet vegetation: relations with atmospheric inputs and management regimes
Terrestrial phosphorus limitation: mechanisms, implica- tions, and nitrogen-phosphorus interactions
Effects of N, P Addition on Above/Below- Ground Biomass Allocation and Plant Functional Types’ Composition in a Sub-Alpine Meadow
氮、磷添加对亚高山草甸地上/地下生物量分配及植物功能群组成的影响
Grassland responses to three years of elevated temperature, CO2, precipitation, and N deposition
Ecological basis of alpine meadow ecosystem management in Tibet: Haibei alpine meadow ecosystem research station

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}